
Inhibition of Fungi and Gram-Negative Bacteria by Bacteriocin BacTN635 Produced by Lactobacillus...
16
This is an author-deposited version published in: http://oatao.univ-toulouse.fr/ Eprints ID : 5699 To link to this article: DOI:10.1007/s12010-009-8821-7 URL: http://dx.doi.org/10.1007/s12010-009-8821-7 To cite this version: Smaoui, Slim and Elleuch, Lobna and Bejar, Wacim and Karrai-Rebai, Ines and Ayadi, Imen and Jaouadi, Bassem and Chouayekh, Hichem and Bejar, Samir and Mellouli, Lofti and Mathieu, Florence (2010) Inhibition of Fungi and Gram-Negative Bacteria by Bacteriocin BacTN635 Produced by Lactobacillus plantarum sp. TN635. Applied Chemistry and Biotechnology pp. 1132-1146. ISSN 0273-2289 Open Archive Toulouse Archive Ouverte (OATAO) OATAO is an open access repository that collects the work of Toulouse researchers and makes it freely available over the web where possible. Any correspondence concerning this service should be sent to the repository administrator: [email protected] hal-00725453, version 1 - 27 Aug 2012 Author manuscript, published in "Applied Chemistry and Biotechnology vol. 162 (2010) pp. 1132-1146" DOI : 10.1007/s12010-009-8821-7
Transcript of Inhibition of Fungi and Gram-Negative Bacteria by Bacteriocin BacTN635 Produced by Lactobacillus...

This is an author-deposited version published in: http://oatao.univ-toulouse.fr/ Eprints ID: 5699
To link to this article: DOI:10.1007/s12010-009-8821-7 URL: http://dx.doi.org/10.1007/s12010-009-8821-7
To cite this version: Smaoui, Slim and Elleuch, Lobna and Bejar, Wacim and Karrai-Rebai, Ines and Ayadi, Imen and Jaouadi, Bassem and Chouayekh, Hichem and Bejar, Samir and Mellouli, Lofti and Mathieu, Florence (2010) Inhibition of Fungi and Gram-Negative Bacteria by Bacteriocin BacTN635 Produced by Lactobacillus plantarum sp. TN635. Applied Chemistry and Biotechnology pp. 1132-1146. ISSN 0273-2289
Open Archive Toulouse Archive Ouverte (OATAO) OATAO is an open access repository that collects the work of Toulouse researchers and makes it freely available over the web where possible.
Any correspondence concerning this service should be sent to the repository administrator: [email protected]
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012
Author manuscript, published in "Applied Chemistry and Biotechnology vol. 162 (2010) pp. 1132-1146" DOI : 10.1007/s12010-009-8821-7

Inhibition of Fungi and Gram-Negative Bacteria
by Bacteriocin BacTN635 Produced by Lactobacillus
plantarum sp. TN635
Slim Smaoui & Lobna Elleuch & Wacim Bejar & Ines Karray-Rebai & Imen Ayadi &
Bassem Jaouadi & Florence Mathieu & Hichem Chouayekh & Samir Bejar &
Lotfi Mellouli
Abstract The aim of this study was to evaluate 54 lactic acid bacteria (LAB) strains
isolated from meat, fermented vegetables and dairy products for their capacity to produce
antimicrobial activities against several bacteria and fungi. The strain designed TN635 has
been selected for advanced studies. The supernatant culture of this strain inhibits the growth
of all tested pathogenic including the four Gram-negative bacteria (Salmonella enterica
ATCC43972, Pseudomonas aeruginosa ATCC 49189, Hafnia sp. and Serratia sp.) and
the pathogenic fungus Candida tropicalis R2 CIP203. Based on the nucleotide sequence of
the 16S rRNA gene of the strain TN635 (1,540 pb accession no FN252881) and the
phylogenetic analysis, we propose the assignment of our new isolate bacterium as
Lactobacillus plantarum sp. TN635 strain. Its antimicrobial compound was determined as a
proteinaceous substance, stable to heat and to treatment with surfactants and organic
solvents. Highest antimicrobial activity was found between pH 3 and 11 with an optimum
at pH=7. The BacTN635 was purified to homogeneity by a four-step protocol involving
ammonium sulfate precipitation, centrifugal microconcentrators with a 10-kDa membrane
cutoff, gel filtration Sephadex G-25, and C18 reverse-phase HPLC. SDS-PAGE analysis of
the purified BacTN635, revealed a single band with an estimated molecular mass of
approximately 4 kDa. The maximum bacteriocin production (5,000 AU/ml) was recorded
after a 16-h incubation in Man, Rogosa, and Sharpe (MRS) medium at 30°C. The mode of
S. Smaoui : L. Elleuch :W. Bejar : I. Karray-Rebai : I. Ayadi : B. Jaouadi : H. Chouayekh : S. Bejar :
L. Mellouli
Laboratoire d’Enzymes et de Métabolites des Procaryotes (LEMP), Centre de Biotechnologie de Sfax
(CBS), Route de Sidi Mansour Km 6, B.P. “1177”, 3018 Sfax, Tunisia
F. Mathieu
Département « Bioprocédés & Systèmes Microbiens » INP-ENSAT, Université de Toulouse, Laboratoire
de Génie Chimique UMR 5503 (CNRS/INPT/UPS), 1 Av, de l’Agrobiopôle, BP 32607, 31326 Castanet-
Tolosan, France
L. Mellouli (*)
Laboratoire d’Enzymes et de Métabolites des Procaryotes, Centre de Biotechnologie de Sfax, B.P
“1177”, Route Sidi Mansour Km 6, 3018 Sfax, Tunisia
e-mail: [email protected]
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

action of the partial purified BacTN635 was identified as bactericidal against Listeria
ivanovii BUG 496 and as fungistatic against C. tropicalis R2 CIP203.
Keywords Lactobacillus plantarum sp. TN635 . Identification . BacTN635 . Purification .
Broad inhibitory spectrum
Introduction
During the last two decades, with the misuse use of existing antibiotics in human and
veterinary medicine and in agriculture, the effectiveness of currently available antibiotics is
decreasing due to the increasing number of resistant pathogenic strains. This is presently an
urgent focus of research and it becomes inevitable to discover new antimicrobial agents for
combating such a problem. Numerous antibacterial agents are now being considered, such
as bacteriophage [1], probiotic bacteria [2], antimicrobial peptides [3], and bacteriocins [4].
Bacteriocins are ribosomally synthesized peptides with a narrower spectrum of antimicro-
bial activity than most antibiotics [5]. Their antimicrobial activity has been found more
effective against closely related strains [6]. Actually, attention is turning to the potential
application of bacteriocins in the protection of human health [7], in agriculture [8], and in
food preservation [9]. Although bacteriocins are produced by many Gram-positive and
Gram-negative bacteria, those produced by lactic acid bacteria (LAB) are of particular
interest because they are known as “good bacteria” and can be used as natural preservatives
in food industry [10]. During the last few years, a large number of new LAB bacteriocins
have been identified and characterized. However, only few have been described to possess
activities against Gram-negative bacteria, e.g., plantaricin 35d produced by Lactobacillus
plantarum [11], bacteriocin ST34BR produced by L. lactis subsp. Lactis [12], bacteriocins
ST26MS and ST28MS produced by L. plantarum [13] and bacteriocin AMA-K produced
by L. plantarum AMA-K [14].
During our search program for bioactive compounds from LAB, a new bacterium called
TN635 was selected. This paper describes the selection, identification, and phylogenetic
determination of this bacterium. The purification and the characterization of a bacteriocin-
like substance called BacTN635, from the supernatant culture of this new strain are also
reported.
Materials and Methods
Bacterial Strains and Plasmids
The TN635 strain was selected as a producer of potent antimicrobial activities and was used
as the source of chromosomal DNA to amplify the 16S rRNA gene. E. coli TOP10
(Invitrogen), F-mcrA Δ(mrr-hsdRMS-mcrBC) φ 80lacZΔM15 ΔlacX74 deoR recA1
araD139 Δ(ara-leu)7697 galU galK rpsL endA1 nupG, and E. coli DH5α [15] were used
as host strains. Bacterial strains: Gram-positive bacteria (Micrococcus luteus LB 14110,
Staphylococcus aureus ATCC 6538, Listeria ivanovii BUG 496, Enterococcus faecalis JH
2-2 and Enterococcus faecium ENSAIA 631), Gram-negative bacteria (Escherichia coli
ATCC 8739, Salmonella enterica ATCC43972, Pseudomonas aeruginosa ATCC 49189,
Hafnia sp. and Serratia sp.), and lactic acid bacteria: (Lactobacillus casei DSM 20011,
Lactobacillus sakei 2525, Lactobacillus delbrueckii DSM 20081, Lactococcus lactis ATCC
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

11454 and Lactococcus lactis subsp. cremoris DSM 11603) were used as indicator
microorganisms for the antibacterial activity assays. Antifungal activity was determined
against the Fusarium sp. and Candida tropicalis R2 CIP203.
pIJ2925 [16] derivative of pUC18 and pCR-Blunt vector (Invitrogen) Col E1 origin
(pUC-derived) KnR were used as the cloning vectors. pSS1, derivative of pCR-Blunt vector
carrying a 1,540-bp DNA fragment corresponding to the whole 16S rRNA gene of the
TN635 strain (this work).
Culture Conditions
E. coli DH5α was grown on Luria Bertani (LB) plates supplemented with ampicillin
(50 μg/ml) and 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (40 μg/ml) when
appropriate [17]. Transformation of E. coli DH5α with pIJ2925 derivatives was carried
out according to Hanahan [15]. Growth and transformation of TOP10 E. coli strain with the
pCR-Blunt vector derivative were carried out according to the manufacturer’s instructions
(Invitrogen). For determination of antibacterial activities, indicator microorganisms were
grown overnight in LB medium in aerobic conditions at 30°C for Lb. casei DSM 20011,
Lb. sakei 2525, Lc. lactis ATCC 11454, Lc. lactis subsp. cremoris DSM 11603, L. ivanovii
BUG 496, S. enterica ATCC43972, P. aeruginosa ATCC 49189, and M. luteus LB 14110,
in aerobic conditions at 37°C for E. coli ATCC 8739 and S. aureus ATCC 6538, in
anaerobic conditions at 30°C for E. faecalis JH 2-2, E. faecium ENSAIA 631, Hafnia sp
and Serratia sp, and in anaerobic conditions at 37°C for Lb. delbrueckii DSM 20081. For
antifungal activities determination, Fusarium sp. was grown in potato dextrose agar (PDA)
for 7 days at 30°C. Spores were collected in sterile distilled water and then concentrated to
produce a suspension with approximately 104spores/ml. C. tropicalis R2 CIP203 was
grown in YP10 medium (10 g/l yeast extract, 10 g/l peptone, 100 g/l glucose, 15 ml of 2 g/
l adenine solution) at 30°C for 24 h in an orbital incubator with shaking at 200 rpm. The
TN635 strain was maintained as frozen stocks at −80°C in Man, Rogosa, and Sharpe
(MRS) broth containing 20% (v/v) glycerol. Before experimental use, the cultures were
propagated twice in MRS medium at 30°C for 12 h and the transfer inoculum was 1% (v/v).
DNA Isolation and Manipulation
Total DNA preparation from TN635 strain was carried out according to the method of
Leenhouts et al. [18] with slight modifications as described by Lee et al. [19]. Small-scale
plasmid preparations from E. coli were performed as described by Sambrook et al. [17].
Digestion with restriction endonucleases, separation of DNA fragments by agarose gels
electrophoresis, dephosphorylation with alkaline calf intestinal phosphatase, ligation of DNA
fragments, and transformation of E. coli were performed according to Sambrook et al. [17].
PCR Amplification of the 16S rRNA Gene of TN635 Strain
PCR amplification of the 16S rRNA gene of TN635 strain was performed using two specific
primers P1: 5′-AGAGTTTGATCCTGGCTCAG-3′ and P2: 5′-ATGGCTCAGGAC
GAACGCT-3′. Approximately 200 ng genomic template DNA was used with 150 pmol of
each primer per 50 μl reaction volume. Amplification was performed in an automated
thermocycler (Perkin Elmer) using 1 U Pfu DNA polymerase (Stratagene) according to the
following amplification profile: 94°C (3 min) followed by 40 cycles of denaturation at 94°C
(30 s), annealing at 60°C (1 min), and extension at 72°C (3 min).
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

DNA Sequencing and Analysis
Nucleotide sequence of the whole 16S rRNA gene of TN635 strain was determined on both
strands by an automated 3100 Genetic Analyser (Applied Biosystems) using specific
primers. Homology search was performed using Blast Search algorithm. The nucleotide
sequence of the whole 16S rRNA gene (1,540 pb) of TN635 strain has been assigned
GenBank (EMBL) under accession number FN252881. Multiple sequence alignment was
carried out using CLUSTAL W [20] at the European Bioinformatics Institute website
(http://www.ebi.ac.uk/clustalw/). Phylogenetic analyses were performed using programs
from the PHYLIP package [21] and phylogenetic tree was constructed by the neighbor-
joining (NJ) algorithm [22] using Kimura ten-parameter distance. The robustness of the
inferred tree was evaluated by bootstrap (100 replications).
Antimicrobial Activities Determination
The ability of TN635 strain to produce diffusible metabolites was tested by the agar well
diffusion assay [23]. The diameters of the inhibition zones were measured. The bacteriocin
samples to be spotted were serially diluted twofold, and the reciprocal of the highest
inhibitory dilution was used to calculate the arbitrary activity units (AU) per milliliter. The
un-inoculated media were also tested for inhibitory zones as a control.
Effect of Different Treatments on the Antimicrobial Activity of the TN635 Strain
The indicator strains used in these experiments were: L. ivanovii BUG 496, S. enterica
ATCC43972, S. aureus ATCC 6538, E. faecalis JH 2-2 and E. faecium ENSAIA 631, and
the pathogenic fungus C. tropicalis R2 CIP203. All determinations were carried out
simultaneously three times under the same conditions. For each determination, control tests
were realized without the corresponding treatment.
Effect of Temperature and Stability at Different pH
Cultures of the TN635 strain in MRS broth under anaerobic condition were maintained at
different temperatures (25, 30, 37, and 45°C). The growth was followed by measuring the
optical density (OD) at 600 nm. Sensitivity to heat of antimicrobial compounds was
investigated by treating the 16-h culture supernatant of the TN635 strain growing at 30°C
in water bath at 45, 70, 85, and 100°C for 90 min and 2 h and by autoclaving (20 min at
121°C). In order to determine the sensitivity of the bacteriocin to pH, the supernatant of the
TN635 strain culture (30°C, 16 h incubation) was adjusted to pH levels ranging from 1 to
14 (intervals of 1.0) with sterile 1 N HCl or 1 N NaOH and kept at 4°C for 6 h. The treated
supernatants were adjusted to pH 7.0 and tested for their antimicrobial activities as
previously described.
Sensitivity to Different Enzymes and Surfactants
In order to determine the biological nature of the antimicrobial activity produced by the
strain TN635, 1 ml of the cell-free supernatant was incubated for 3 h at 37°C in the
presence of 1 mg/ml catalase (Boehringer). The untreated bacteriocin-containing cell-free
supernatant served as control. Sensitivity to proteolytic, lipolytic and α-amylase enzymes of
antimicrobial compounds was investigated by the addition of Trypsin, pronase E (Sigma),
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

Proteinase K (Boehringer), lipase A (Sigma) and α-amylase (Sigma) at final concentration
of 1 mg/ml to the culture supernatants of the TN635 strain (30°C, 16 h). The samples were
incubated for 2 h at 37°C and immediately after, the residual activity was determined as
described above. The surfactants used were Tween 20, Tween 80, urea, Triton X-100,
sodium dodecyl sulfate (SDS), ethylenediaminetetraacetic acid (EDTA), and phenyl-
methanesulfonylfluoride (PMSF) which were added to the culture supernatants of the
TN635 strain (30°C, 16 h) at a final concentration of 1% (v/v) and incubated at 30°C for
2 h. Surfactants at 1% in MRS broth were used as controls. All surfactants were prepared at
10% in water and filter-sterilized before use.
Effect of Organic Solvents
The culture supernatants of the TN635 strain (30°C, 16 h) were mixed with various organic
solvents (ethyl acetate, isopropanol and methanol) at a final concentration of 50% (v/v).
After incubation for 1 h at room temperature, the organic solvent was evaporated in a
vacuum concentrator and the residual antimicrobial activity was determined.
Bacteriocin Purification, Molecular Weight Determination, and Zymogram Analysis
Strain TN635 was inoculated (1% v/v) into 900 ml (Duran bottles, containing 900 ml of MRS
broth), and incubated without agitation at 30°C until early stationary phase (16 h)
corresponding to maximum bacteriocin production. For bacteriocin purification, proteins of
the crude supernatant fluid (CSF) were precipitated with 40, 60, and 80% saturated
ammonium sulfate, gradually added by slow stirring during 4 h at 4°C. After centrifugation at
9,000×g for 30 min at 4°C, the precipitate was suspended in 10 ml of 20 mmol/l sodium
phosphate buffer (pH 7.0) and desalted in centrifugal microconcentrators (Amicon, Inc.) with
a 10-kDa membrane cutoff. The resulting solution (3.5 ml) was loaded on a column (70×
1.5 cm) of gel filtration Sephadex G-25 equilibrated with 20 mmol/l sodium phosphate buffer
(pH 7). Elution of proteins was performed with the same buffer at 30 ml/h; 140 fractions (F1–
F140) with 5 ml each were eluted from the Sephadex G-25 column and collected
automatically. These 140 fractions detected using absorption measurements at 280 nm were
fractioned into six samples (S1–S6): S1 (F1–F40), S2 (F41–F70), S3 (F71–F76), S4 (F77–
F81), S5 (F82–F93), and S6 (F94–F140). The antimicrobial activity of the different collected
fractions was tested using L. ivanovii BUG 496, S. enterica ATCC43972, and C. tropicalis
R2 CIP203 as an indicator strains. The active fraction was then applied to a second round of
purification by a reverse-phase HPLC method using a C18 column (300×4.6 mm) at a flow
rate of 0.5 ml/min. Bacteriocin was eluted from the column with two mobile phases: A
(99.9% water, 0.1% trifluoroacetic acid “TFA”) and B (99.9% acetonitrile, 0.1% TFA); from
0 to 5 min (90% A, 10% B), from 5 to 30 min (50% A, 50% B), from 30 to 35 min (20% A,
80% B), from 35 to 50 min (10% A, 90% B) and from 50 to 60 min (90% A, 10% B).
Proteins were monitored at 280 nm. The pooled biological active fraction obtained from
HPLC elution was concentrated and stored at −20°C. The estimated molecular weight of the
purified bacteriocin was determined by SDS-PAGE according to Laemmli [24] with 20%
acrylamide gel. Protein concentration was measured using BSA as reference as described by
Bradford [25]. The used protein marker is from BioLabs with a broad range of 2–212 kDa.
To determine the apparent molecular mass of the bacteriocin, the gel was cut into two vertical
parts after SDS-PAGE. The part of the gel containing the sample and the protein marker was
stained with 0.25% Coomassie brilliant blue R-250 (BioRad), while the remaining part,
containing only the sample, was washed in 10 mmol/l phosphate buffer (pH 7) for 4 h, then
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

fixed and used for direct detection of antimicrobial activity by overlaying with LB soft agar
containing 100 μl of a 5 h culture of L. ivanovii BUG 496.
Mode of Action of BacTN635
For this realization, we have used two indicator microorganisms: L. ivanovii BUG 496 and
C. tropicalis R2 CIP203. Partially-purified BacTN635 (the fraction obtained from gel
filtration chromatography and freeze-dried) at 400 AU/ml, was added to 200 ml LB culture
of L. ivanovii BUG 496 and 200 ml YP10 of C. tropicalis R2 CIP203 at in early
exponential phase (107colony-forming units (CFU)/ml). The two indicator microorganisms
growing in LB (L. ivanovii BUG 496) and YP10 (C. tropicalis R2 CIP203) in the absence
of bacteriocin were used as controls. Changes in the turbidity of the cultures were recorded
at an O.D. of the 600 nm and the number of CFU was determined by plaiting the samples
on LB or YP10 agar.
Results and Discussion
Screening of LAB Strains Producing Bacteriocin
During previous works (data not shown), a collection of 54 LAB strains were isolated from
several isolates (meat, fermented vegetables, and dairy products). We have tested these
bacteria for their capacity to produce antimicrobial activities against the following indicator
microorganisms: Lb. casei DSM 20011, L. ivanovii BUG 496, S. enterica ATCC43972,
Fusarium sp., and C. tropicalis R2 CIP203. As shown in Table 1, different profiles of
inhibition were observed. Nine strains (TN600, TN606, TN615, TN618, TN623, TN627,
TN635, TN644, and TN653) were characterized by the broad-spectrum inhibition. These
nine isolates were tested for their capacity to inhibit the growth of other microorganisms
(Table 2). The largest spectrum of inhibition was showed by TN635 strain which inhibited
all used indicator microorganisms. Moreover, the active molecule(s) of the TN635 strain
presents the highest inhibitory effect against all tested microorganisms including the
pathogenic Gram-positive bacteria (L. ivanovii BUG 496, S. aureus ATCC 6538, E. faecalis
JH 2-2 and E. faecium ENSAIA 631), the four pathogenic Gram-negative bacteria (S.
enterica ATCC43972, P. aeruginosa ATCC 49189, Hafnia sp. and Serratia sp.), and the
pathogenic fungus C. tropicalis R2 CIP203. Since bacteriocins often have a narrow killing
spectrum and inhibit only bacteria closely related to those from which they have been
formed, bacteriocins from Gram-positive bacteria are generally not effective against Gram-
negative bacteria and have no effect with yeasts and fungi [26, 27]. The search for new
bacteriocins with a wider spectrum of activity which can be used in human health,
agriculture, and the food industry is being studied by several research groups. According to
its large inhibitory spectrum, our finding, BacTN635 of the new isolated TN635 strain, may
be of applied interest as food preservation and/or in the protection of human health.
Identification and Phylogenetic Analysis of TN635 Strain
Using the two specific primers: forward P1: 5′-AGAGTTTGATCCTGGCTCAG-3′ and
reverse P2: 5′-ATGGCTCAGGACGAACGCT-3′ and the TN635 strain genomic DNA as
template, we have amplified by PCR a DNA fragment having an expected size of
approximately 1,500 pb. After purification, this DNA fragment was cloned in the pGEM-T
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

Table 1 Inhibitory spectrum (zone of inhibition/mm) of the isolated 54 LAB (TN600- TN653) against Lb.
casei DSM 20011, L. ivanovii BUG 496, S. enterica ATCC43972, Fusarium.sp, and C. tropicalis R2
CIP203: L (listeria); Lb (Lactobacillus); C (Candida).
Isolates Indicator microorganisms
Lb. casei L. ivanovii S. enterica Fusarium sp. C. tropicalis
DSM 20011 BUG 496 ATCC43972 R2 CIP203
TN600 6 12 22 14 8
TN601 – 7 – – –
TN602 – – – – –
TN603 8 – – – 6
TN604 – – – 8 8
TN605 10 16 7 7 –
TN606 7 18 16 16 9
TN607 6 5 – – –
TN608 8 8 8 – –
TN609 9 – 7 8 –
TN610 – 13 15 – –
TN611 – – – – –
TN612 – – – – –
TN613 9 17 10 – 8
TN614 – 6 7 8 15
TN615 7 6 18 19 5
TN616 8 – – 16 –
TN617 – – – 18 –
TN618 8 14 16 18 9
TN619 8 7 – 10 –
TN620 7 16 – – –
TN621 6 – – 20 –
TN622 – 7 5 – 6
TN623 7 20 18 17 19
TN624 8 8 7 7 –
TN625 7 6 17 – –
TN626 – – – – –
TN627 8 9 12 14 10
TN628 – – 6 16 –
TN629 7 – – 6 –
TN630 9 9 – 6 6
TN631 – 18 17 9 –
TN632 – 8 8 – –
TN633 9 – – – 7
TN634 6 – 7 7 –
TN635 9 22 21 23 21
TN636 9 6 – 7 –
TN637 8 7 – 15 –
TN638 7 – 7 16 –
TN639 – 7 7 18 7
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

Table 1 (continued).
Isolates Indicator microorganisms
Lb. casei L. ivanovii S. enterica Fusarium sp. C. tropicalis
DSM 20011 BUG 496 ATCC43972 R2 CIP203
TN640 7 7 8 6 –
TN641 – 16 17 – 15
TN642 6 – – 16 –
TN643 – 6 7 17 6
TN644 7 16 6 22 6
TN645 6 7 8 – –
TN646 – – – 18 7
TN647 7 – 6 8 –
TN648 8 8 – 18 –
TN649 – – 8 16 6
TN650 – 16 – 17 17
TN651 6 – 7 7 6
TN652 – 8 – 12 7
TN653 7 16 17 18 8
Table 2 Inhibitory spectrum (zone of inhibition/mm) of TN600, TN606, TN615, TN618, TN623, TN627,
TN635, TN 644 and TN653 strains against Gram-positive and Gram-negative bacteria, LAB and fungi: L
(listeria); Lb (Lactobacillus); Lc (Lactococcus); C (Candida).
Indicator strains TN600 TN606 TN615 TN618 TN623 TN627 TN635 TN644 TN653
Gram-positive bacteria
M. luteus LB 14110 23 14 6 14 14 6 23 14 11
S. aureus ATCC 6538 12 15 16 14 9 16 22 21 8
L. ivanovii BUG 496 12 18 6 16 20 9 22 16 16
E. faecalis JH 2-2 6 18 8 – 15 16 22 7 8
E. faecium ENSAIA 631 6 14 7 – 14 14 21 8 6
Gram-negative bacteria
E. coli ATCC 8739 5 22 13 15 15 23 22 13 22
S. enterica ATCC43972 22 16 18 16 18 12 21 6 17
P. aeruginosa ATCC 49189 22 16 6 9 14 9 22 8 9
Hafnia sp. 5 15 6 13 24 6 24 5 7
Serratia sp. 12 6 12 15 21 7 23 8 9
LAB
Lb. Casei DSM 20011 6 7 7 8 7 8 9 7 7
Lb. delbrueckii DSM 20081 – 6 8 – – 9 9 6 –
Lb. Sakei 2525 – 6 7 – 7 6 9 – –
Lc. lactis ATCC 11454 – – 6 5 6 7 9 – 9
Lc. lactis cremoris DSM 11603 – – 5 7 7 9 8 – 8
Fungi
Fusarium sp. 14 16 19 18 17 14 23 22 18
C. tropicalis R2 CIP203 8 9 5 9 19 10 21 6 8
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

Easy vector, yielding the pSS1 plasmid. Total nucleotide sequence of 1,540 pb (accession
number FN252881) of the whole 16S rRNA gene of strain TN635 was determined in both
strands. The alignment of this sequence through matching with reported 16S rRNA gene
sequences in gene bank shows high similarity (97–99%) to the Lactobacillus 16S rRNA
genes. The organism most similar to the new isolate TN635 strain was L. plantarum NRIC
0383 (Fig. 1). Based on the nucleotide sequence of the 16S rRNA gene of the TN635 strain
and the phylogenetic analysis, we propose the assignment of our new isolate bacterium as
L. plantarum sp. TN635 strain.
Growth and Bacteriocin Production
To test the effect of incubation temperature on cell growth and antimicrobial activity
production, cultures of the L. plantarum sp. TN635 strain in MRS broth under anaerobic
conditions were maintained at different temperatures (25, 30, 37, and 45°C). The growth
was followed by measuring the OD at 600 nm and the antimicrobial activity was
determined against the three indicator microorganisms: L. ivanovii BUG 496, S. enterica
ATCC43972, and C. tropicalis R2 CIP203. Aliquots were withdrawn at 3 h time intervals.
Obtained results show that the secretion of biological activity is correlated with biomass
production. Highest growth and antimicrobial activity were obtained at 30°C (Fig. 2). We
can deduce that the extracellularly antimicrobial activity was produced in the logarithmic
growth phase and the production profile was a typical growth associated patter. The
maximum antimicrobial activity was detected 16 h after incubation. This activity remains
stable between 13 and 20 h and then gradually decreased.
2
88
6
18
24
40
4
9
10
1
10
92
66 76
94
95
88
51
53
Fig. 1 Phylogenetic trees derived from 16S rDNA sequence of L. plantarum sp. TN635 strain. All the
sequences used here were from LAB type strains
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

Effects of Enzymes, Heat, pH, Surfactants and Organic Solvents
Enzymatic tests showed that the antimicrobial activity against the three tested indicator
microorganisms, L. ivanovii BUG 496, S. enterica ATCC43972, and C. tropicalis R2
CIP203, of the supernatant culture of the L. plantarum sp. TN635 strain was not affected by
the addition of catalase, indicating that the observed growth inhibition was not due to
hydrogen peroxide production. In contrast, treatment with the proteolytic enzymes (Trypsin,
pronase E, and Proteinase K) caused complete inactivation of the antimicrobial compounds,
which therefore identified them as proteinaceous substances. The sensitivity of the
inhibitory compounds to heat treatment showed that inhibitory activity was not significantly
altered by boiling for 90 min showing that the antagonistic activity of the culture
supernatant is heat-resistant. The activity of the sample was insensitive to lipase and α-
amylase, which eliminates the possibility that synthesized active compounds are included in
the group of complex biologically active substances containing a lipid or carbohydrate
component [28]. Concerning the sensitivity to pH, the supernatant culture of the Lb.
plantarum sp. TN635 (30°C, 16 h incubation) was adjusted to pH levels ranging from 1 to
14 (intervals of 1.0). The obtained results show that bacteriocin activity was highest when
the pH of the supernatant was between 3 and 11, with an optimum at pH 7. About 50%
activity was maintained at pH 12 and was completely inactivated above pH 12. Exposure to
surfactants caused about 15% inactivation of the antimicrobial activity for the SDS agent
and about 40% for the other tested surfactants (Tween 20, Tween 80, Triton X-100, Urea,
EDTA, and PMSF). Concerning the treatment with organic solvents (ethyl acetate,
methanol, and isopropanol), no significant alteration of the BacTN635 activity was
observed.
0
5
10
15
20
25
30
35
0 6 12 18 24 30 36 42 48
Incubation time (h)
Inh
ibit
ory
Zo
ne D
iam
ete
r (m
m)
0
1
2
3
4
5
6
7
8
OD
600 n
m
Fig. 2 BacTN635 production during growth of L. plantarum TN635 in MRS broth at 30°C. (filled square)
Optical density at 600 nm; activities of BacTN635 against C. tropicalis R2 CIP203 (filled upright triangle);
against S. enterica ATCC43972 (filled circle), and against L. ivanovii BUG 496 (filled diamond) were
measured by diameter (mm) of zone of inhibition
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

Bacteriocin Purification and Molecular Weight
BacTN635 was purified to homogeneity from a cell-free culture supernatant of Lb.
plantarum sp. TN635 strain by a four-step protocol. Maximum antimicrobial activity was
obtained when the cell-free culture supernatant was precipitated with 80% ammonium
sulfate. One hundred forty fractions (F1–F140) with 5 ml each were eluted from a
Sephadex G-25 column. These 140 fractions detected using absorption measurements at
280 nm were fractioned into six samples (S1–S6). After concentration, the antagonistic
activity of each sample was essayed using L. ivanovii BUG 496, S. enterica ATCC43972,
and C. tropicalis R2 CIP203 as indicator microorganisms. Only sample S5 (F82–F93)
possesses inhibitory activity against the three tested cells (Fig. 3). Sample S5 was then
applied to another round of purification using the HPLC technique. Three well-defined
0
0,5
1
1,5
2
2,5
3
3,5
0 10 9080706050403020 100 110 120 130 140 150
Fraction number
OD
28
0n
m
S5
Fig. 3 Purification of BacTN635 by cation-exchange chromatography using Sephadex G-25 monitoring by
absorbance at 280 nm (filled square)
0 10 20 30 40 50 60 70
0.5
1.0
18
.50
0
31.1
33
34
.55
0
38
.3 8
3
39
.31
7
0
P1
P2
P3
Ab
sorb
an
ce (
AU
) at
280 n
m
Retention time (min)
Fig. 4 Elution profile of bacteriocin using HPLC reverse-phase chromatography on C18 column monitoring
by absorbance at 280 nm
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

peaks (P1–P3) were obtained having, respectively, the following retention times: P1:
34.55 min, P2: 38.38 min, and P3: 39.31 min (Fig. 4). Only the fraction of the peak P3
possesses an antimicrobial activity against the three tested microorganisms mentioned
above and no activity has been observed for the two other fractions. The yield, activity, and
purification fold of the BacTN635 of the four various purification steps were summarized in
Table 3. SDS-PAGE analysis of the purified BacTN635, revealed a single band with an
estimated molecular mass of approximately 4 kDa indicating that this bacteriocin had been
purified to homogeneity (Fig. 5a). Antibacterial activity determination against L. ivanovii
BUG 496 strain, reveals a growth inhibitory zone at the same position than that visualized
in the stained gel (Fig. 5b). During the last few years, a large number of new bacteriocins
produced by LAB bacteria have been identified and characterized. However, a few
bacteriocins produced by this species have been reported to be active against Gram-
negative bacteria and particularly against fungi. Most of the described bacteriocins as
antifungal agents from Lactobacillus are organic compounds or diketopiperazine
derivatives with low molecular mass, lower than 1 kDa [29]. Too rare are the bacteriocins
described from LAB bacteria with a molecular mass higher than 1 kDa and having
antifungal activities [30].
Table 3 Purification of BacTN635 produced by Lactobacillus plantarum sp. TN635 strain.
Step Volume
(ml)
Total activity
(AU)aTotal
protein (mg)
Specific activity
(AU/mg)
Purification
fold
Yield
%
Culture supernatant 900 45×105 2,160 2,083 1 100
Ammonium sulfate precipitation 50 5.2×105 52.5 9,904 4.75 11.55
Microconcentrators 32 3.32×105 23.2 14,310 6.87 7.38
Gel filtration 3.5 2.25×105 1.54 146,104 70.14 5
HPLC 0.7 1.125×105 0.57 197,368 94.75 2.5
aAntibacterial activity (in arbitrary units [AU]) was assayed by agar well diffusion assay using L. ivanovii
BUG 496 as an indicator strain
6.5
14.3
1 12
a b
3.4
20.0
27.0
34.6
42.7
55.6
M M
(kDa)
Fig. 5 Electrophoretic and zy-
mogram analyses of the purified
BacTN635. Coomassie brilliant
blue R-250 stained SDS-PAGE
gel. Lane 1 molecular mass
markers; lane 2 purified
BacTN635 (a). Portion of the
renaturated SDS-PAGE, overlaid
with LB soft agar containing L.
ivanovii BUG 496 (b)
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012
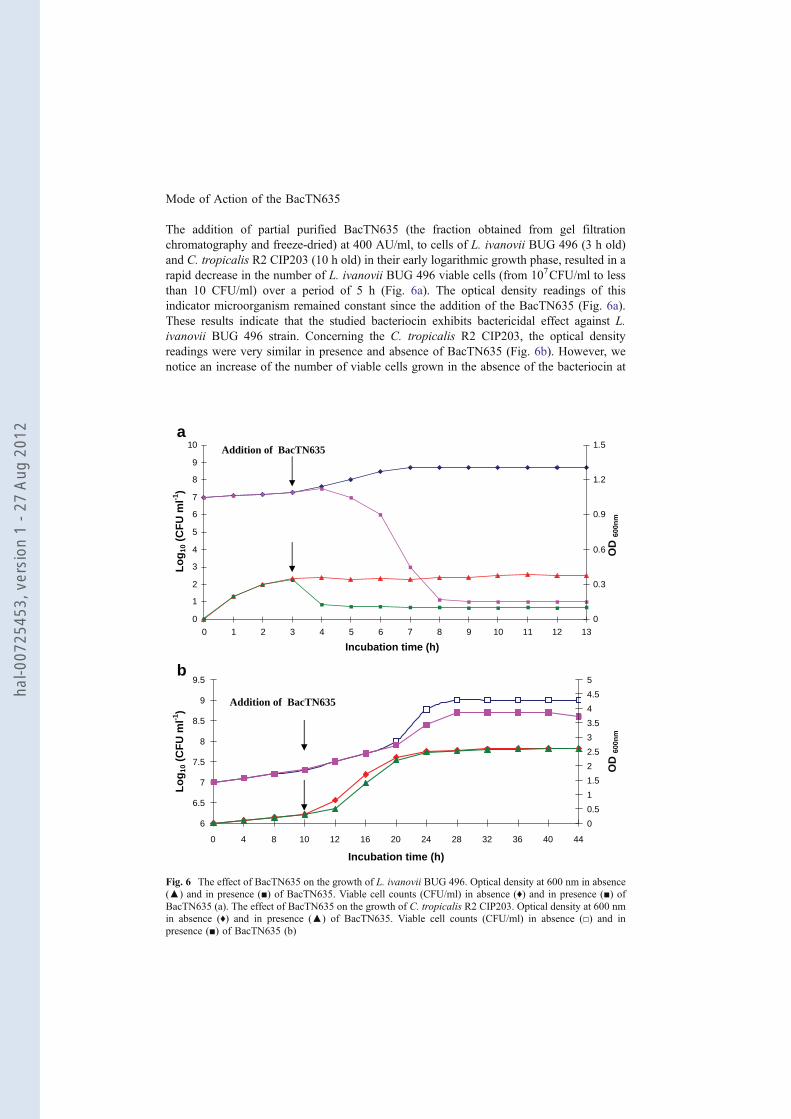
Mode of Action of the BacTN635
The addition of partial purified BacTN635 (the fraction obtained from gel filtration
chromatography and freeze-dried) at 400 AU/ml, to cells of L. ivanovii BUG 496 (3 h old)
and C. tropicalis R2 CIP203 (10 h old) in their early logarithmic growth phase, resulted in a
rapid decrease in the number of L. ivanovii BUG 496 viable cells (from 107CFU/ml to less
than 10 CFU/ml) over a period of 5 h (Fig. 6a). The optical density readings of this
indicator microorganism remained constant since the addition of the BacTN635 (Fig. 6a).
These results indicate that the studied bacteriocin exhibits bactericidal effect against L.
ivanovii BUG 496 strain. Concerning the C. tropicalis R2 CIP203, the optical density
readings were very similar in presence and absence of BacTN635 (Fig. 6b). However, we
notice an increase of the number of viable cells grown in the absence of the bacteriocin at
0
1
2
3
4
5
6
7
8
9
10
0 3 71 4 82 5 6 9 10 11 12 13
Incubation time (h)
0
0.3
0.6
0.9
1.2
1.5
6
6.5
7
7.5
8
8.5
9
9.5
0 4 8 10 12 16 20 24 28 32 36 40 44
Incubation time (h)
Lo
g1
0 (
CF
U m
l-1)
Lo
g1
0 (
CF
U m
l-1)
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
OD
60
0n
mO
D 6
00
nm
Addition of BacTN635
Addition of BacTN635
a
b
Fig. 6 The effect of BacTN635 on the growth of L. ivanovii BUG 496. Optical density at 600 nm in absence
(▲) and in presence (■) of BacTN635. Viable cell counts (CFU/ml) in absence (♦) and in presence (■) of
BacTN635 (a). The effect of BacTN635 on the growth of C. tropicalis R2 CIP203. Optical density at 600 nm
in absence (♦) and in presence (▲) of BacTN635. Viable cell counts (CFU/ml) in absence (□) and in
presence (■) of BacTN635 (b)
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

40 h of growth (Fig. 6b). These data indicate that BacTN635 acts with a fungistatic effect
against C. tropicalis R2 CIP203.
Conclusions
A new LAB strain has been selected for its antimicrobial activity against pathogenic Gram-
negative bacteria and fungi. Based on the analysis of the nucleotide sequence (1,540 pb) of
the whole 16S rRNA gene (accession no: FN252881) and the phylogenetic study, we
propose the assignment of our new isolate bacterium as Lb. plantarum sp. TN635 strain.
The antimicrobial compound of this strain was determined as a proteinaceous substance.
The maximum bacteriocin-like substance (BacTN635) production (5,000 AU/ml) was
recorded after a 16-h incubation in MRS medium at 30°C. BacTN635 was stable to heat
and to treatment with surfactants and organic solvents. Highest antimicrobial activity was
found between pH 3 and 11 with an optimum at pH=7. The purified BacTN635, revealed a
single band with an estimated molecular mass of approximately 4 kDa. BacTN635
produced by Lb. plantarum sp. TN635 strain, may possess potential practical applications,
since it was able to inhibit important Gram-negative pathogenic bacteria such as (S.
enterica ATCC43972, P. aeruginosa ATCC 49189, Hafnia sp., and Serratia sp.) and
pathogenic fungi such as C. tropicalis R2 CIP203. S. enterica is an important cause of
human salmonellosis and food-poisoning associated with consumption of contaminated
chicken eggs and poultry products [31]. P. aeruginosa is an opportunistic pathogen of
humans. It causes urinary tract infections, respiratory system infections, gastrointestinal
infections, and a variety of systemic infections, particularly in patients with severe burns
and in cancer and AIDS patients who are immunosuppressed [32]. Serratia and Hafnia sp.
are usually found in the intestinal tract. These pathogenic bacteria cause s wide variety of
infections primarily pneumonia, wound, and urinary tract infections. Moreover, it should be
noted that the number of patients with fungal infections has been rising in recent years. In
addition to the species Candida albicans and Candida stellatoidea, C. tropicalis is one of
the significant pathogenic yeasts in humans belonging to the genus Candida [33].
Acknowledgments This work was supported by the Tunisian government (Contract Program CBS-LEMP)
and the CMCU project (2006-2008) No 06/S 0901 “MELLOULI/AIGLE”. We are grateful to Dr. M.
Ferchichi for providing the indicator bacteria.
References
1. Alisky, J., Iczkowski, K., Rapoport, A., & Troitsky, N. (1998). Journal of Infection, 36, 5–15.
2. Macfarlane, G. T., & Cummings, J. H. (2002). Current Opinion in Infectious Diseases, 15, 501–506.
3. Papagianni, M. (2003). Biotechnology Advances, 21, 465–499.
4. Twomey, D., Ross, R. P., Ryan, M., Meaney, B., & Hill, C. (2002). Antonie Van Leeuwenhoek, 82,
165–18.
5. Brooks, G. F., Butel, J. S., & Morse, S. A. (1998). Jawetz, Melnick, and Adelberg’s medical
microbiology (p. 269). Upper Saddle River, New Jersey, USA: Appleton & Lange.
6. Settanni, L., & Corsetti, A. (2008). International Journal of Food Microbiology, 121, 123–138.
7. Nawaz, S. K., Riaz, S., Riaz, S., & Hasnain, S. (2009). African. J. Biotechnol., 8, 365–368.
8. Hammami, I., Rhouma, A., Jaouadi, B., Rebai, A., & Nesme, X. (2009). Letters in Applied
Microbiology, 48, 253–260.
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012

9. Diop, M. B., Dubois-Dauphin, R., Tine, E., Ngom, A., Destain, J., & Thonart, P. (2007). Biotechnology,
Agronomy, Society and Environment, 11, 275–281.
10. Paul, R. R., Morgan, S., & Hill, C. (2002). International Journal of Food Microbiology, 79, 3–16.
11. Messi, P., Bondi, M., Sabia, C., Battini, R., & Manicardi, G. (2001). International Journal of Food
Microbiology, 64, 193–198.
12. Todorov, S. D., & Dicks, L. M. (2004). Journal of Basic Microbiology, 44, 305–316.
13. Todorov, S. D., & Dicks, L. M. (2005). Enzyme and Microbial Technology, 36, 318–326.
14. Todorov, S. D., Nyati, H., Meincken, M., & Dicks, L. M. T. (2007). Food Control, 18, 656–664.
15. Hanahan, D. (1983). Journal of Molecular Biology, 16, 557–580.
16. Janssen, G. R., & Bibb, M. J. (1993). Gene, 124, 133–134.
17. Sambrook, J., Fritsch, E. F., & Maniatis, T. (1989). Molecular cloning: a laboratory manual (2nd ed.).
New York: Cold Spring Harbor Laboratory Press.
18. Leenhouts, K. J., Kok, J., & Venema, G. (1990). Applied and Environmental Microbiology, 56, 2726–
2735.
19. Lee, C. M., Sieo, C. C., Abdullah, N., & Ho, Y. W. (2008). FEMS Microbiology Letters, 287, 136–141.
20. Thompson, J. D., Higgins, D. G., & Gibson, T. J. (1994). Nucleic Acids Research, 22, 4673–4680.
21. Felsenstein, J. (1989). PHYLIP-Phylogeny Inference Package, version 3.2. Cladistics, 5, 164–166.
22. Saitou, N., & Nei, M. (1987). Molecular Biology and Evolution, 4, 406–425.
23. Tagg, J. R., & McGiven, A. R. (1971). Applied Microbiology, 21, 943.
24. Laemmli, U. K. (1970). Nature, 227, 680–685.
25. Bradford, M. M. (1976). Analytical Biochemistry, 72, 248–254.
26. Karunasagar, I., Krohne, G., & Goebel, W. (1993). Infection and Immunity, 61, 162–169.
27. Atanassova, V., Meindl, A., & Ring, C. (2001). International Journal of Food Microbiology, 68, 105–
113.
28. Aktypis, A., Kalantzopoulos, G., Huis IN’T Veld, J. H. J., & Ten Brink, B. (1998). Journal of Applied
Microbiology, 84, 568–576.
29. Lavermicocca, P., Valerio, F., Evidente, A., Lazzaroni, S., Corsetti, A., & Gobbetti, M. (2000). Applied
and Environmental Microbiology, 66, 4084–4090.
30. Atanassova, M., Choiset, Y., Dalgalarrond, D., Chobert, J. M., Dousset, X., Ivanova, I., et al. (2003).
International Journal of Food Microbiology, 8, 63–73.
31. Erich, L., & Martin, J. (1997). Antimicrobial food additives, characteristics, uses, effects (2nd Ed.).
Translated by S.F. Laichena Berlin: Springer.
32. Wiblin, R. T. (1997). Nosocomial pneumonia. In R. P. Wenzel (Ed.), Prevention and control of
nosocomial infections (3rd ed., pp. 807–819). Baltimore: Williams and Wilkins.
33. Kobayashi, H., Matsuda, K., Ikeda, T., Suzuki, M., Takahashi, S., Suzuki, A., et al. (1994). Infection and
Immunity, 62, 615–622.
hal-0
0725
453,
ver
sion
1 -
27 A
ug 2
012




















