
Carbohydrate Starvation Causes a Metabolically Active but Nonculturable State in Lactococcus lactis
Upload
independentCategory
view
3download
0
Detection and characterization of a bacteriocin,garviecin L1-5, produced by Lactococcus garvieae isolatedfrom raw cow's milk
F. Villani, M. Aponte, G. Blaiotta, G. Mauriello, O. Pepe and G. MoschettiDipartimento di Scienza degli Alimenti, Sezione di Microbiologia Agraria, Alimentare ed Ambientale e di Igiene,
UniversitaÁ degli Studi di Napoli `Federico II', Portici, Italy
321/4/00: received 14 April 2000, revised 9 November 2000 and accepted 15 November 2000
F. V ILLANI , M. APONTE, G. BLAIOTTA, G. MAURIELLO, O. PEPE AND G. MOSCHETTI . 2001.
Aims: The identi®cation of a bacteriocin-producing lactococcal strain isolated from raw cow's
milk is reported, along with production conditions, physical and chemical properties, and mode
of action of the bacteriocin.
Methods and Results: On the basis of resistance to clindamycin, species-speci®c PCR and
ampli®cation of the 16S-23S rDNA spacer region, the strain was identi®ed as Lactococcus
garvieae. Its bacteriocin, designated garviecin L1-5, was bactericidal against closely related
species and strains of species from different genera, including Listeria monocytogenes and
Clostridium spp. Garviecin L1-5 was shown to be proteinaceous by protease inactivation and
was unaffected by heat treatments, also at low pH values. When amplifying known lactococcal
bacteriocin genes using DNA from strain L1-5 as template, no ampli®cation products were
observed on the agarose gel. The molecular weight of garviecin L1-5 was about 2á5 kDa. As far
as is known, no bacteriocins have been detected from Lactococcus garvieae.
Conclusions: The general properties of garviecin L1-5 are characteristic of the low-
molecular-weight bactericidal peptide group.
Signi®cance and Impact of the Study: The survey of micro-organisms for novel
antimicrobial substances provided valuable information on their physiology, ecology and
practical application.
INTRODUCTION
Antimicrobial proteinaceous compounds produced by bac-
teria that are active against other bacteria, despite varying
greatly in chemical nature, mode of action and host
speci®city, have traditionally been de®ned as bacteriocins
(Jack et al. 1995). Bacteriocin-producing bacteria may occur
over the whole range of Gram-negative and Gram-positive
species and have been isolated from a wide variety of
habitats (Dykes 1995).
Bacteriocins and lantibiotics produced by lactic acid
bacteria (LAB) are extremely important in preventing the
growth of spoilage and pathogenic bacteria, and have
been the subject of extensive studies in recent years
because of their potential use as novel, natural food
preservatives (Nettles and Barefoot 1993; Holzapfel et al.1995). The study of the biochemistry and genetics of
bacteriocins, and their use as phenotypic markers, has
also provided much valuable information on the genetics
and physiology of lactic acid bacteria (Klaenhammer
1993).
Bacteriocin-producing species have now been identi®ed
among all the genera that comprise the LAB, including
Lactococcus, Streptococcus, Lactobacillus, Leuconostoc, Pedio-coccus and Carnobacterium as well as several Enterococcus spp.
(Jack et al. 1995).
Production of bacteriocins by lactococci has been studied
extensively. Nisin, produced by certain strains of Lactococcuslactis subsp. lactis, is perhaps the best known lantibiotic
because of its importance and approved use as a food
preservative (Delves-Broughton 1990). It inhibits the
Correspondence to: Dr F. Villani, Dipartimento di Scienza degli Alimenti,
Sezione di Microbiologia Agraria, Alimentare ed Ambientale e di Igiene,
UniversitaÁ degli Studi di Napoli `Federico II', Via UniversitaÁ 100, I 80055
Portici, Italy (e-mail: [email protected]).
ã 2001 The Society for Applied Microbiology
Journal of Applied Microbiology 2001, 90, 430±439

growth of several Gram-positive micro-organisms, including
LAB, clostridia and pathogenic bacteria. Several other
lactococcal bacteriocins have been identi®ed from a variety
of Lactococcus spp. They can be distinguished by their
spectrum of activity, mode of action, biochemical and
genetic characteristics (for a review up to 1994, see De Vuyst
1994; Morgan et al. 1995; Villani et al. 1995; Martinez et al.1996; Ryan et al. 1996).
As far as is known, no bacteriocins have been detected
from Lactococcus garvieae. During the course of a survey of
LAB for novel antimicrobial substances, a bacteriocin was
found, termed garviecin L1-5, which was produced by a
strain of L. garvieae.
Lactococcus garvieae was originally isolated from cows with
mastitis (Garvie et al. 1981; Collins et al. 1984) and has been
reported, together with L. lactis, to be associated with a
variety of different types of infections in humans and
animals (Elliott et al. 1991), although its role as an infectious
agent remains unclear (Aguirre and Collins 1993). However,
strains belonging to the species L. lactis are commonly used
as starter cultures for a variety of fermented and cultured
dairy products and are generally-recognized-as-safe (GRAS)
micro-organisms.
Lactococcus lactis and L. garvieae are genealogically quite
distinct but phenotypically closely related, making their
differentiation on the basis of phenotypic criteria very
dif®cult (Rodrigues et al. 1991). Erroneous identi®cation of
the two species has been reported for clinical isolates (Elliott
et al. 1991). Unequivocal differentiation between L. garvieaeand L. lactis can be made, at present, only by DNA
homology (Garvie et al. 1981), rRNA sequence analysis
(Collins et al. 1989), rRNA gene restriction patterns
(Rodrigues et al. 1991) or by less time-consuming tests
such as resistance of L. garvieae to clindamycin (Elliott and
Facklam 1996), whole-cell protein analysis (Elliott et al.1991), or PCR assay speci®c to L. garvieae (Zlotkin et al.1998).
This paper reports the identi®cation of the bacteriocin-
producing strain isolated from raw cow's milk, and produc-
tion conditions, physical and chemical properties, and action
mode of the bacteriocin.
MATERIALS AND METHODS
Cultures and growth conditions
Bacterial strains used as indicator organisms and the
conditions of their incubation are listed in Table 1. Cultures
of lactic acid bacteria and non-lactic cultures were main-
tained in appropriate broths with 25% (v/v) sterile glycerol
and stored at ) 20°C. Plating and overlay media were
prepared by adding 1á5 and 0á75% agar, respectively, to the
broth media listed in Table 1.
Detection and isolation of bacteriocin-producingstrains
Fresh raw cows' milk samples from local farms were 10-fold
diluted in 0á1% Peptone water and plated in triplicate on
M17 agar (Oxoid). After incubation at 30°C for 24 h,
countable plates were overlaid with the appropriate soft
(0á75% agar) medium seeded with about 106 cfu ml)1 of
logarithmic phase cultures of a given indicator strain. The
indicator organisms tested singly were Lactococcus lactissubsp. lactis DSM 20481T and Listeria monocytogenes Scott
A. After further incubation at 30 or 37°C for 18 h, colonies
exhibiting inhibition were removed from their sandwiched
position using a snipped pipette tip and streaked onto M17
agar plates to obtain a pure culture. Antagonistic activity of
the isolates was con®rmed by the agar-spot deferred method
as previously described (Villani et al. 1993, 1995). Inhibition
due to proteinaceous substances was assessed by spotting,
near the spotted culture, 5 ll proteolytic enzymes
(10 mg ml)1; trypsin, pronase E, a-chymotrypsin and
pepsin, all from Sigma, were tested); the enzymes were
dissolved in 50 mmol l)1 Tris-HCl (pH 7á0), which resulted
in the disappearance of the inhibition zone near the enzyme
spot.
One of the isolates exhibiting inhibitory activity against
L. lactis DSM 20481T and Listeria monocytogenes Scott A
was identi®ed as Lactococcus garvieae L1-5 (see below) and
studied in detail for its antimicrobial substances.
Identi®cation of bacteriocin-producing strains
Phenotypic characterization. Conventional morphologi-
cal and physiological tests were determined as described by
Smibert and Krieg (1994). They included: cell morphology
and Gram-reaction; catalase and oxidase activity; hydrolysis
of arginine, esculine and Hippurate; growth in broth
containing 4 and 6á5% NaCl; growth at 10 and 45°C.
Production of pyrrolidonyl arylamidase was determined using
the commercial kit Pyr test (Oxoid); gas production from
glucose was determined by the hot loop test (Sperber and
Swan 1976); serogroup D was assessed by the Dryspot
Streptococcal Grouping Kit (Oxoid); susceptibility to clynda-
mycin (2 lg disc, Oxoid) was determined as described by
Elliott and Facklam (1996); acid production from carbohy-
drates was performed using API-50 CH fermentation panels
(API, Biomerieux Italia SpA, Ponte a Ema, Florence, Italy).
PCR assay. DNA extraction was carried out as previously
reported (Moschetti et al. 1996), and 25 ng DNA were used
for PCR ampli®cations, as described below.
Species-speci®c PCR ampli®cation. Species-speci®c oli-
gonucleotide primers described by Zlotkin et al. (1998)
LACTOCOCCUS GARVIEAE BACTERIOCIN 431
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 430±439

(pLG-1 5¢-CATAACAATGAGAATCGC-3¢ and pLG-2
5¢-GCACCCTCGCGGGTTG-3¢), targeting part of the
16S rRNA of L. garvieae, were used. The PCR was carried
out in a programmable heating incubator (MJ Research Inc.,
Watertown, Madison, USA) for 35 cycles (1 min at 94°C,
1 min at 55°C and 1á5 min at 72°C per cycle) followed by
10 min at 72°C. DNA from L. garvieae DSM 20684T
(positive control) and L. lactis subsp. lactis DSM 20481T
(negative control) were included in the analysis. The
presence of speci®c PCR products of 1100 bp was revealed
by electrophoresis in 1á5% (w/v) agarose gel.
Ampli®cation of 16S-23S rDNA spacer region. Syn-
thetic oligonucleotide primers described by Jensen et al.(1993) (G1, 5¢GAAGTCGTAACAAGG3¢ and L1, 5¢CA-
AGGCATCCACCGT 3¢) were used to amplify the 16S-
23S rDNA intergenic spacer region. PCR conditions
consisted of 25 cycles (1 min at 94°C, 2 min at 55°C,
3 min at 72°C) plus one additional cycle with a ®nal 10 min
chain elongation. DNA from L. garvieae DSM 20684T and
L. lactis subsp. lactis DSM 20481T were included in the
analysis. The presence of PCR products was controlled by
agarose (2% w/v) gel electrophoresis at 7 V cm)1 for 2 h.
Table 1 Sensitivity of indicator strains to garviecin L1-5 as determined by the critical dilution assay
Indicator strains Substrate* Incubation temperature °C Source AU ml)1à
Lactococcus garvieae 20684T M17 30 DSM 40
L. garvieae§ M17 30 RM 0
L. garvieae± M17 30 RM 40
L. lactis subsp. lactis 20481T M17 30 DSM 40
L. lactis subsp. lactis 124 M17 30 NWC 40
L. lactis subsp. cremoris 125 M17 30 NWC 40
L. lactis subsp. diacetylactis 152 M17 30 NWC 20
L. lactis subsp. diacetylactis RD M17 30 NWC 40
Streptococcus thermophilus 304 M17 42 NWC 0
Enterococcus faecalis 19433 M17 37 ATCC 20
Ent. faecium 19434 M17 37 ATCC 20
Ent. durans 11576 M17 37 ATCC 20
Ent. casseli¯avus 14436 M17 37 ATCC 0
Lactobacillus delbrueckii subsp. lactis MRS 37 NWC 0
Lact. delbrueckii subsp. bulgaricus 414 MRS 42 NWC 0
Lact. salivarius 432 MRS 37 NWC 0
Lact. helveticus 421 MRS 42 NWC 0
Leuconostoc cremoris 592 MRS 30 NWC 0
Leuc. mesenteroides 530 MRS 30 NWC 0
Listeria monocytogenes OH BHI 32 Carminati 40
Listeria monocytogenes V7 BHI 32 Carminati 40
Listeria monocytogenes CAL BHI 32 Carminati 40
Listeria monocytogenes SA BHI 32 Carminati 40
Listeria innocua C6 BHI 32 Cheese 40
Staphylococcus aureus 5S4 BHI 37 Cheese 0
Brucella melitensis F3 BHI 37 Clinical 0
Brucella abortus F4 BHI 37 Cheese 0
Klebsiella spp. CL10 BHI 37 Clinical 0
Yersinia spp. CL12 BHI 37 Clinical 0
Escherichia coli O157:H7 BHI 37 Sausage 0
Salmonella typhi CL3 BHI 37 Clinical 0
Clostridium butyricum RCM 30 Bottazzi 20
Cl. tyrobutyricum 25755 RCM 30 Bottazzi 20
Cl. sporogenes 7955 RCM 37 Bottazzi 20
* M17 (Oxoid, CM785B); MRS (Oxoid, CM395B); BHI (Oxoid, CM225B); RCM (Oxoid, CM149B).
NWC: natural whey cultures; RM: Raw milk; DSM: Deutsche sammlung von Mikroorganismen und Zellkulturen, Gmbh, Braunschweig,
Germany; ATCC: American Type Culture Collection, Rockville, Maryland, USA; Prof. V. Bottazzi, UniversitaÁ Cattolica Sacro Cuore Piacenza, Italy;
Dr D. Carminati, Istituto Sperimentale Lattiero-caseario, Lodi, Italy.
à Titres of garviecin L1-5 using a given strain as indicator.
§ Producer strain.
± Nine other strains of Lactococcus garvieae from RCM were inhibited; the titre of the bacteriocin activity range from 20 and 40 AU ml)1.
432 F. VILLANI ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 430±439

Ampli®cation of lactococcal bacteriocin genes. Oligo-
nucleotides targeting several bacteriocin genes are reported
in Table 2. PCRs were carried out in a programmable
heating incubator (MJ Research Inc., Watertown, Madison,
USA) for 25 cycles (45 s at 94°C, 45 s at 56°C and 1 min at
72°C per cycle) followed by 3 min at 72°C. The presence of
PCR products was revealed by electrophoresis in 1á5%
agarose gel.
Bacteriocin production and measurementof bacteriocin activity
Lactococcus garvieae L1-5 was grown in M17 broth
(Oxoid) + 0á5% lactose, M17 broth (Oxoid) + 0á5% glu-
cose (GM17), MRS (Oxoid), TSB broth (Oxoid) and BHI
broth (Oxoid) for 16 h at 30°C. The cells were removed
by centrifugation (10 000 g for 10 min) and the superna-
tant ¯uids were adjusted to pH 7á0 with 2 mol l)1 NaOH
and treated at 80°C for 10 min. Aliquots (50 ll) of the
heat-treated supernatant ¯uids were assayed for bacterio-
cin activity using the agar-well diffusion method (see
below).
The kinetics of bacteriocin production in MRS broth was
determined by inoculating L. garvieae L1-5 (1% v/v) and
incubating at 30°C. Viable cell count of the culture on M17
agar, and bacteriocin activity, were measured at hourly
intervals. The samples were centrifuged at 10 000 g for
10 min and the supernatant ¯uids were adjusted to pH 7á0,
heat-treated, and assayed for bacteriocin activity as des-
cribed below.
Bacteriocin activity was measured by the agar-well
diffusion method and critical dilution assay as previously
described (Villani et al. 1993, 1995). Brie¯y, 50 ll of serial
twofold dilution of heat-treated supernatant ¯uids were
added to wells of 6 mm in diameter cut in M17 agar plates
previously inoculated with the indicator strain, L. lactisDSM 20481T, and allowed to diffuse. The plates were
incubated at 30°C for 20±24 h and examined for zones of
inhibition. One Arbitrary Unit (AU) per millilitre was
de®ned as the reciprocal of the highest dilution yielding a
zone of growth inhibition on the indicator lawn.
Agar plugs bored out of the area of inhibition were tested
for the presence of lytic bacteriophage, by the plaque assay
of Terzaghi and Sandine (1975), using soft-BHI agar and
L. lactis DSM 20481T and Listeria monocytogenes Scott A as
indicator strains.
Partial puri®cation of bacteriocin
The procedure described by Stoffels et al. (1993) was used
with some modi®cation. The producing strain was inocu-
lated at 1% (v/v) and incubated at 30°C in MRS broth for
15 h. Cells were removed by centrifugation at 10 000 g for
15 min at 4°C. The bacteriocin was precipitated from the
supernatant ¯uid by ammonium sulphate (40% saturation,
4 h at 4°C). The resulting precipitate was collected by
centrifugation at 10 000 g for 35 min, dissolved in 5 ml
50 mmol l)1 potassium phosphate buffer, pH 7á0, and
exhaustively dialysed (3500 MWCO, Spectra por 3; Spec-
trum Medical Industries Inc., Los Angeles, CA, USA)
against the same buffer for 24 h at 4°C. The dialysed
fractions was used as the crude sample.
The crude preparation was adsorbed on Amberlite XAD-
16 (Sigma). Adsorption was carried out in a batch process by
adding 2á5% resin, twice within 1 h, under stirring. The
resin was transferred into a chromatographic column and
washed with 50% ethanol until the eluate was colourless.
Antagonistic substance was eluted with 70% 2-propanol
complemented with 10 mmol l)1 acetic acid, and adjusted to
pH 2á0 with concentrated HCl. After evaporation on a BuÈchi
Table 2 PCR primers used in this study amplifying lactococcal bacteriocin genes
Primers Sequences (5¢-3¢) PCR product Bacteriocin References
NISL CGAGCATAATAAACGGC 320 bp Nisin A/Z Ward et al. (1994)
NISR GGATAGTATCCATGTCTGAAC
LCNA 1 CAATCAGTAGAGTTATTAACATTTG 870 bp Lactococcin A Martinez et al. (1998)
LCNA 2 GATTTAAAAAGACATTCGATAATTAT
LCNB 1 GCT TGC AGT ATG TTA TGA GTG 431 bp Lactococcin B van Belkum et al. (1992)
LCNB 2 CCT ACC ATC CAG GAT TTT CTT
513F GCT CCA AAA AGC GCT AGA TC 466 bp Lactococcin 513 GenBank n. AF056207
513R GCT GGC TAC GAT ATT GCT AG
RM 1 ATC CTA TCC GAT ACC GTC AG 644 bp Lacticin RM GenBank n. AF080265
RM 2 GTT TTC CCT GAA CCA TTG GG
972F GCG CTC TCT TGC ATA GTG AG 531 bp Lactococcin 972 Martinez et al. (1999)
972R ACT CCT CCA TTA GTA CCA GC
LCT1 (481) GCA TCT GCA CTC ACT TCA 254 bp Lacticin 481 Piard et al. (1993)
LCT2 (481) GCA AGT AAA TAC AAA TTG CC
LACTOCOCCUS GARVIEAE BACTERIOCIN 433
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 430±439

apparatus, the precipitate was resuspended in 50 mmol l)1
potassium phosphate buffer (pH 7á0) and applied to an
S-Sepharose Fast Flow Column (2á5 ´ 7 cm; Pharmacia
LKB Biotechnology, Uppsala, Sweden). The column was
washed with 50 mmol l)1 potassium phosphate buffer
(pH 7á0) and the bacteriocin was eluted with the same
buffer supplemented with 1á0 mol l)1 NaCl. The active
fractions obtained were used as the partially puri®ed sample.
Bacteriocin and protein concentrations of each fraction were
determined by critical dilution assay and by Bio-Rad protein
assay kit, respectively.
Detection of bacteriocin activity by SDS-PAGEanalysis
Crude and partially puri®ed bacteriocin samples were
examined by Tricine-SDS-PAGE (4% in the stacking gel,
10% in the spacer gel and 16á5% in the running gel) as
described by Schagger and Von Jagow (1987) using standard
peptides as molecular weight markers (Promega, Madison,
WI, USA; Low-Range Markers, 2á5±31 kDa). After elec-
trophoresis at 200 V for 45 min, the gel was cut in two parts.
The ®rst half was stained with Coomassie brilliant blue R
(Sigma) while the other was assayed for antimicrobial
activity by the direct method of Bhunia et al. (1987).
Spectrum of antimicrobial activity
Crude garviecin L1-5 was examined for inhibition of the
indicator organisms listed in the Table 1 by the agar-well
diffusion method. Bacteriocin titre, determined by critical
dilution assay using a given strain as indicator, was taken as a
measure of strain sensitivity.
Sensitivity of garviecin L1-5 to enzymes,pH and heat
Crude bacteriocin (640 AU ml)1) was treated with trypsin,
pronase E, a-chymotrypsin, pepsin, papain, proteinase K,
lipase, catalase and a-amylase, all from Sigma. The enzymes
were dissolved (1 mg ml)1) in 50 mmol l)1 Tris±HCl
(pH 7á0) and ®lter sterilized. Bacteriocin in buffers without
enzymes, enzyme solutions and buffers alone were used as
controls. Samples were incubated at 37°C for 1 h, and the
residual activity was determined by critical dilution assay
against L. lactis DSM 20481T.
The effect of heat treatments at various pH upon
bacteriocin activity was determined by adjusting the pH of
crude garviecin L1-5 (640 AU ml)1) in the range pH 2á0 to
pH 9á0. Samples were heated at 30, 60, 100 and 121°C for
various times (Table 4), readjusted to pH 7á0 and assayed
for residual bacteriocin activity.
Mode of action
The effect of crude garviecin L1-5 was determined on non-
growing indicator cells. Lactococcus lactis subsp. lactis DSM
20481T was grown at 30°C up to log-phase in LM17 broth.
The cells were harvested by centrifugation (8000 g for
10 min) and resuspended in sterile 50 mmol l)1 potassium
phosphate buffer, pH 7á0, containing 640, 320 and 0 AU
ml)1, to yield about 109 cfu ml)1. The cell suspensions were
incubated at 30°C. Samples were withdrawn at 0, 30, 60, 120
and 180 min and viable counts were determined on LM17
agar (30°C for 24 h). At each sampling time, optical density at
600 nm and 260 nm was also measured to check for cell lysis.
Cell suspension without bacteriocin was used as control.
RESULTS
Detection and identi®cationof bacteriocin-producing strains
A total of eight bacteriocin-producing isolates was detected
in raw cow's milk by deferred antagonism assay using
Lactococcus lactis DSM 20481T and Listeria monocytogenesScott A as indicators. One of these Bac+ isolates, called
L1-5, exhibiting inhibitory activity against both indicators
tested, was selected for further study. The L1-5 isolate was a
Gram-positive, catalase-negative coccus that did not pro-
duce gas from glucose. It was able to grow at 10°C and in
broth with 6á5% NaCl but failed to grow at 45°C. No group
D antigen was detected. It showed acid production from the
following carbohydrates: glucose, ribose, galactose, fructose,
mannose, mannitol, N-acetyglucosamine, amygdaline, arbu-
tine, salicine, cellobiose, esculine, maltose, lactose, trehalose,
gentibiose and tagatose. On the basis of fermentation
patterns and positive results due to pyrrolidonyl arylamidase
activity and resistance to clindamycin, isolate L1-5 was
tentatively identi®ed as Lactococcus garvieae. Molecular
techniques were used to further con®rm the identity of
the isolate. PCR was used to amplify a fragment from the
16S rRNA gene of L. garvieae. The L1-5 isolate and
L. garvieae DSM 20684T strain showed a band of 1100 bp
on the agarose gel. By contrast, no ampli®cation product was
obtained using DNA from L. lactis subsp. lactis DSM
20481T as template (results not shown).
To differentiate L. garvieae from L. lactis de®nitively,
ampli®cation of the 16S±23S rDNA spacer region was
performed for both species. Results are shown in Fig. 1.
The bacteriocin-producing strain L1-5, and L. garvieaeDSM 20684T, showed an ampli®cation band migrating at
approximately 410 bp (Fig. 1, lane 2 and 1, respectively).
For L. lactis subsp. lactis DSM 20481T, a different
ampli®cation band located at 380 bp (Fig. 1, lane 3) was
obtained.
434 F. VILLANI ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 430±439

Bacteriocin production
Lactococcus garvieae L1-5 was found to have antimicrobial
activity. This could be shown both on solid media (agar spot
deferred antagonism) and in the neutralized and heat-treated
(10 min at 80°C) culture supernatant ¯uid of all broths
tested after incubation for 24 h at 30°C (data not shown).
MRS broth was found to yield maximum activity by critical
dilution assay against L. lactis DSM 20481T and was used in
subsequent experiments. The kinetics of the production of
antibacterial substance was monitored during the growth of
L. garvieae L1-5 in MRS broth at 30°C (Fig. 2). Activity
was detected after 4 h of incubation, when the micro-
organisms were in the mid-log phase of growth (3á3 ´ 108
cfu ml)1). Maximum activity was reached at 6±8 h, in the
late logarithmic to early stationary phase of growth.
Thereafter, the concentration declined after further incuba-
tion. During bacterial growth, the pH of the medium ranged
between 6á21 and 4á67 (data not shown).
The antagonistic activity was not associated with phages
as the material extracted from the agar in the zones of
inhibition did not produce plaques when assayed by the
double-layer technique.
In order to verify whether the L1-5 strain of L. garvieaeproduced a bacteriocin different from those formerly
sequenced, PCR was performed with speci®c primers
amplifying lactococcal bacteriocin genes (Table 2). No
ampli®cation products were observed on the agarose gel
using DNA from strain L1-5 as template in PCR ampli®-
cation (results not shown).
Partial puri®cation and detectionof garviecin L1-5 by SDS-PAGE
Bacteriocin was partially puri®ed from spent culture ¯uids
by a combination of ammonium sulphate precipitation,
XAD-16 and cation exchange chromatography. From
1000 ml of culture supernatant ¯uid, garviecin L1-5 was
puri®ed 136-fold with a recovery yield of 4% (Table 3).
Tricine SDS-PAGE was used to estimate the molecular
mass of garviecin L1-5 (Fig. 3). Duplicate samples of both
crude and partially-puri®ed bacteriocin were run on the
same gel, one half being stained with Coomassie blue and
the other being assayed for antimicrobial activity, overlaying
with the sensitive indicator L. lactis DSM 20481T. The
resulting zone of inhibition corresponded to a protein of
about 2á5 kDa. No other protein bands showed any antimi-
crobial property.
Spectrum of activity
The activity of crude garviecin L1-5 was assayed by the
agar-well diffusion method on several Gram-positive and
Gram-negative bacteria (Table 1). The bacteriocin was
Fig. 1 Ethidium bromide stained 2% agarose gel of PCR products
using primers in the region of 16S±23S intergenic spacer region.
Lane 1: Lactococcus garvieae DSM 20684T; lane 2: L. garvieae L1-5;
lane 3: L. lactis subsp. lactis DSM 20481T; lane 4: negative control;
M: 1 Kb DNA ladder (Gibco BRL) used as molecular marker
Fig. 2 Growth of Lactococcus garvieae L1±5 (s) and bacteriocin
production (d) in MRS broth at 30 °C. AU (Arbitrary units) ml)1
of neutralized and heat-treated culture supernatant ¯uid as determined
by critical dilution assay against L. lactis subsp. lactis DSM 20481T
LACTOCOCCUS GARVIEAE BACTERIOCIN 435
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 430±439

active against species relatively closely related to the
producer and against all strains of Listeria monocytogenesand Clostridium spp. tested. The producer strain L. garvieaeL1-5 and representative strains of Gram-negative bacteria
were not inhibited.
Effect of enzymes, pH and heaton the antimicrobial substance
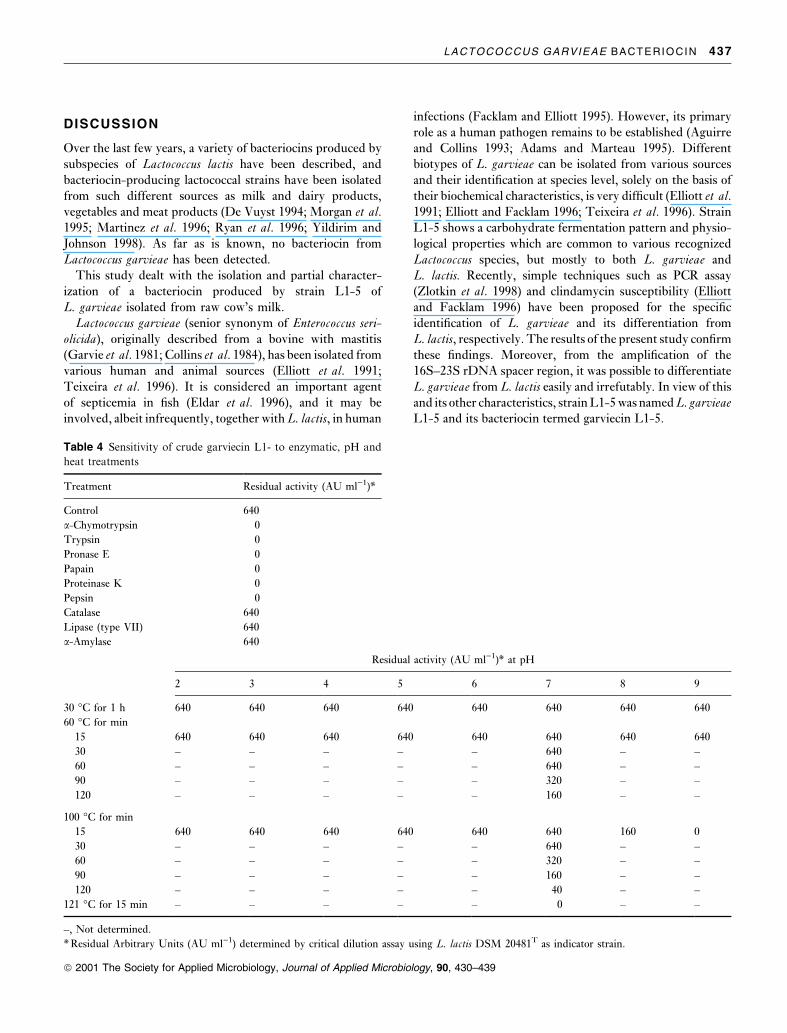
Table 4 reports the effects of enzymes, pH and temperature
on bacteriocin activity. Complete inactivation of the anti-
microbial activity of garviecin L1-5 resulted after treatment
with all proteolytic enzymes tested. Non-proteolytic
enzymes did not cause inactivation. Buffer and enzyme
solutions alone had no effect on the sensitive indicator.
Garviecin L1-5 fully retained its activity at pH levels
between 2 and 9 when treated at 30 and 60°C for 1 h. After
treatment at 100°C for 15 min, the bacteriocin lost 75 and
100% of its activity at pH 8 and 9, respectively. At pH 7á0,
activity decreased by 50, 75 and 93á75% after 60, 90 and
120 min, respectively. Treatment at 121°C for 15 min led to
complete loss of activity.
Mode of action
The effect of two concentrations of garviecin L1-5 on the
viability of non-growing cells of L. Lactis DSM 20481T,
suspended in potassium phosphate buffer at pH 7á0, is
shown in Fig. 4. Within 30 min, at 640 AU ml)1, the
population was reduced from 3á5 ´ 108 cfu ml)1 to 1á3 ´ 105
cfu ml)1, while addition of 320 AU ml)1 garviecin L1-5
resulted in a reduction in colony counts from 3á5 ´ 108 to
9á1 ´ 105 cfu ml)1. Viable counts of cell suspensions without
bacteriocin were unchanged. Incubation for more than
30 min did not signi®cantly change the number of viable
micro-organisms. Optical densities of the control and treated
cell suspensions remained stable throughout the experiment,
and no cell lysis was seen microscopically (data not shown).
Fig. 3 SDS-PAGE of garviecin L1±5. (a)
Stained half of the gel showing proteins
present in the crude (lane 1) and partially-
puri®ed samples (lane 2). Lane M contains
Low Molecular Marker (Promega). (b) Sec-
ond half of the gel overlaid with L. lactis
subsp. lactis DSM 20481T shows a zone of
inhibition corresponding to the active crude
(lane 3) and partially-puri®ed (lane 4) garvie-
cin L1-5
Puri®cation step
Total
activity*
(AU)
Total
protein
(mg)
Speci®c activity
(AU mg)1 protein)
Yield
(%)
Puri®cation
(fold)
Culture supernatant 40000 57á6 694 100 1
Ammonium sulphate
precipitation 6400 1á77 3615 16 5á2XAD-16
chromatography 3200 0á82 3902 8 5á6S-Sepharose
chromatography 600 0á017 94117 4 136
* Activity was expressed in Arbitrary Units and determined by critical dilution assay using
Lactococcus lactis subsp. lactis DSM 20481T as indicator strain.
Table 3 Partial puri®cation of garviecin
L1-5
436 F. VILLANI ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 430±439
DISCUSSION
Over the last few years, a variety of bacteriocins produced by
subspecies of Lactococcus lactis have been described, and
bacteriocin-producing lactococcal strains have been isolated
from such different sources as milk and dairy products,
vegetables and meat products (De Vuyst 1994; Morgan et al.1995; Martinez et al. 1996; Ryan et al. 1996; Yildirim and
Johnson 1998). As far as is known, no bacteriocin from
Lactococcus garvieae has been detected.This study dealt with the isolation and partial character-
ization of a bacteriocin produced by strain L1-5 of
L. garvieae isolated from raw cow's milk.
Lactococcus garvieae (senior synonym of Enterococcus seri-olicida), originally described from a bovine with mastitis
(Garvie et al. 1981; Collins et al. 1984), has been isolated from
various human and animal sources (Elliott et al. 1991;
Teixeira et al. 1996). It is considered an important agent
of septicemia in ®sh (Eldar et al. 1996), and it may be
involved, albeit infrequently, together with L. lactis, in human
infections (Facklam and Elliott 1995). However, its primary
role as a human pathogen remains to be established (Aguirre
and Collins 1993; Adams and Marteau 1995). Different
biotypes of L. garvieae can be isolated from various sources
and their identi®cation at species level, solely on the basis of
their biochemical characteristics, is very dif®cult (Elliott et al.1991; Elliott and Facklam 1996; Teixeira et al. 1996). Strain
L1-5 shows a carbohydrate fermentation pattern and physio-
logical properties which are common to various recognized
Lactococcus species, but mostly to both L. garvieae and
L. lactis. Recently, simple techniques such as PCR assay
(Zlotkin et al. 1998) and clindamycin susceptibility (Elliott
and Facklam 1996) have been proposed for the speci®c
identi®cation of L. garvieae and its differentiation from
L. lactis, respectively. The results of the present study con®rm
these ®ndings. Moreover, from the ampli®cation of the
16S±23S rDNA spacer region, it was possible to differentiate
L. garvieae from L. lactis easily and irrefutably. In view of this
and its other characteristics, strain L1-5 was named L. garvieaeL1-5 and its bacteriocin termed garviecin L1-5.
Residual activity (AU ml)1)* at pH
2 3 4 5 6 7 8 9
30 °C for 1 h 640 640 640 640 640 640 640 640
60 °C for min
15 640 640 640 640 640 640 640 640
30 ± ± ± ± ± 640 ± ±
60 ± ± ± ± ± 640 ± ±
90 ± ± ± ± ± 320 ± ±
120 ± ± ± ± ± 160 ± ±
100 °C for min
15 640 640 640 640 640 640 160 0
30 ± ± ± ± ± 640 ± ±
60 ± ± ± ± ± 320 ± ±
90 ± ± ± ± ± 160 ± ±
120 ± ± ± ± ± 40 ± ±
121 °C for 15 min ± ± ± ± ± 0 ± ±
±, Not determined.
* Residual Arbitrary Units (AU ml)1) determined by critical dilution assay using L. lactis DSM 20481T as indicator strain.
Table 4 Sensitivity of crude garviecin L1- to enzymatic, pH and
heat treatments
Treatment Residual activity (AU ml)1)*
Control 640
a-Chymotrypsin 0
Trypsin 0
Pronase E 0
Papain 0
Proteinase K 0
Pepsin 0
Catalase 640
Lipase (type VII) 640
a-Amylase 640
LACTOCOCCUS GARVIEAE BACTERIOCIN 437
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 430±439

Garviecin L1-5 was detected in cell-free supernatant ¯uids
of various broths, with maximal activity in MRS broth. This
medium proved to be better than others for bacteriocin
production (De Vuyst 1994). The bacteriocin reached a
maximum in the late logarithmic to early stationary phase of
growth, with a sharp decline at later stages. The reason for
this rapid loss of activity was not investigated, but similar
results have been reported for other bacteriocins and are
possibly due to extracellular proteases or adsorption to the
producer cells (De Vuyst 1994; Yildirim and Johnson 1998).
The experiments reported here rule out any possibility of
antagonistic action due to acids, hydrogen peroxide or
bacteriophage. Garviecin L1-5 was shown to be proteina-
ceous by protease inactivation (a-chymotrypsin, trypsin,
pronase E, papain and proteinase K), but was unaffected by
heat treatments, also at low pH values. However, bacteriocin
activity decreased or disappeared at alkaline pH.
Like other lactococcal bacteriocins, such as nisin, lacto-
coccin R (Yildirim and Johnson 1998) or lacticin 3147 (Ryan
et al. 1996), garviecin L1-5 exhibits a wide activity spectrum
towards closely related species and strains of species from
different genera, including Listeria monocytogenes and Clos-tridium spp. The ability to inhibit certain organisms outside
their genus is also typical of Gram-positive bacteriocins
(Jack et al. 1995). However, L. garvieae L1-5 was not a nisin
producer, or a lacticin and lactococcin producer, because no
ampli®ed products were observed when its genomic DNA
was used as template in PCR ampli®cation. Information on
amino acid or genetic determinant sequences will be
required to distinguish garviecin L1-5 from the many
others bacteriocins described.
Activity of garviecin L1-5 against sensitivity indicators
was bactericidal without concomitant cell lysis, and the
effectiveness of the inhibitor appears to depend upon the
inhibitor concentration. A similar mode of action has been
observed in many other bacteriocins from lactic acid bacteria
(Nettles and Barefoot 1993; De Vuyst 1994; Jack et al.1995). Direct detection of antimicrobial activity after
electrophoresis on SDS-PAGels overlaid with L. lactisDSM 20481T indicated that garviecin L1-5 has a molecular
mass of 2á5 kDa. In this respect, and in its heat stability, it
resembles lactococcin R produced by a strain of L. lactissubsp. cremoris (Yildirim and Johnson 1998). Unfortunately,
this bacteriocin is not yet sequenced.
In conclusion, the general properties of garviecin L1-5 are
characteristic of the low-molecular-weight bactericidal pep-
tide group (Klaenhammer 1993). Further work will be
necessary to extend the knowledge of this bacteriocin.
Puri®cation to homogeneity, as well as its characterization at
the molecular level, is now in progress.
ACKNOWLEDGEMENTS
The authors thank Prof. Salvatore Coppola for his encour-
agement to perform this study during research to develop the
microbial biodiversity occurring in traditional cheesemaking
processes supported by the MURST, Rome (PRIN 1998).
REFERENCES
Adams, M.R. and Marteau, P. (1995) On the safety of lactic acid bacteria
from food. International Journal of Food Microbiology 27, 263±264.
Aguirre, M. and Collins, M.D. (1993) Lactic acid bacteria and human
clinical infection. Journal of Applied Bacteriology 75, 95±107.
Bhunia, A.K., Johnson, M.C. and Ray, B. (1987) Direct detection of an
antimicrobial peptide of Pediococcus acidilactici in sodium dodecyl-
sulfate polyacrylamide gel electrophoresis. Journal of Industrial
Microbiology 2, 319±322.
Collins, M.D., Ash, C., Farrow, J.A.E., Wallbanks, S. and Williams,
A.M. (1989) 16S Ribosomal ribonucleic acid sequence analysis of
lactococci and related taxa. Description of Vagococcus ¯uvialis gen.
nov., sp. nov. Journal of Applied Bacteriology 67, 453±460.
Collins, M.D., Farrow, J.A.E., Phillips, B.A. and Kandler, O. (1984)
Streptococcus garvieae sp. nov. & Streptococcus plantarum sp. nov.
Journal of General Microbiology 129, 3427±3431.
De Vuyst, L. (1994) Bacteriocins produced by Lactococcus lactis strains.
In Bacteriocins of Lactic Acid Bacteria. Microbiology, Genetics and
Applications ed.De Vuyst, L. and Vandamme, E.J. pp. 143±149.
Blackie Academic & Professional. London: Chapman & Hall.
Delves-Broughton, J. (1990) Nisin and its uses as a food preservative.
Food Technology 44, 100±117.
Dykes, G.A. (1995) Bacteriocins: ecological and evolutionary signi®-
cance. Trends in Ecology and Evolution 10, 186±189.
Eldar, A., Ghittino, C., Assenta, L. et al. (1996) Enterococcus seriolicida
is a junior synonym of Lactococcus garvieae, a causative agent of
Fig. 4 Effect of garviecin L1-5 on non-growing cells of Lactococcus
lactis DSM 20481T suspended in 50 mmol l)1 potassium phosphate
buffer, pH 7. (h), Control, no bacteriocin added; (d) 640 AU ml)1;
(s) 320 AU ml)1
438 F. VILLANI ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 430±439

septicemia and meningoencephalitis in ®sh. Current Microbiology 32,
85±88.
Elliott, J.A., Collins, M.D., Pigott, N.E. and Facklam, R.R. (1991)
Differentiation of Lactococcus lactis and Lactococcus garvieae from
humans by comparison of whole-cell protein patterns. Journal of
Clinical Microbiology 29, 2731±2734.
Elliott, J.A. and Facklam, R.R. (1996) Antimicrobial susceptibilities of
Lactococcus lactis and Lactococcus garvieae and proposed method to
discriminate between them. Journal of Clinical Microbiology 34,
1296±1298.
Facklam, R.R. and Elliott, J.A. (1995) Identi®cation, classi®cation, and
clinical relevance of catalase-negative, Gram-positive cocci, exclu-
ding the streptococci and enterococci. Clinical Microbiology Review
8, 479±495.
Garvie, E.I., Farrow, J.A.E. and Phillips, B.A. (1981) A taxonomic
study of some strains of streptococci that grow at 10°C but not at
45°C, including Streptococcus lactis and Streptococcus cremoris.
Zentralblatt fuÈr Bakteriologie, Parasitenkunde, Infektionskrankheiten
und Hygiene, 1 Abteilung, Originale C 2, 151±165.
Holzapfel, W.H., Geisen, R. and Schillinger, U. (1995) Biological
preservation of foods with reference to protective cultures, bacte-
riocins and food-grade enzymes. International Journal of Food
Microbiology 24, 343±362.
Jack, R.W., Tagg, J.R. and Ray, B. (1995) Bacteriocins of Gram-
positive bacteria. Microbiological Review 59, 171±200.
Jensen, M.A., Webster, J.A. and Straus, N. (1993) Rapid identi®cation
of bacteria on the basis of Polymerase Chain Reaction-ampli®ed
ribosomal DNA spacer polymorphism. Applied and Environmental
Microbiology 59, 945±952.
Klaenhammer, T.R. (1993) Genetics of bacteriocins produced by lactic
acid bacteria. FEMS Microbiology Review 12, 39±86.
Martinez, B., Fernandez, M., Suarez, J.E. and Rodriguez, A. (1999)
Synthesis of lactococcin 972, a bacteriocin produced by Lactococcus
lactis IPLA 972, depends on the expression of a plasmid-encoded
bicistronic operon. Microbiology 145, 3155±3161.
Martinez, B., Rodriguez, A., Medina, M., Hernandez, P.E. and
Rodriguez, J.M. (1998) Detection of speci®c bacteriocin-producing
lactic acid bacteria by colony hybridization. Journal of Applied
Microbiology 84, 1099±1103.
Martinez, B., Suarez, J.E. and Rodriguez, A. (1996) Lactococcin 972:
a homodimeric lactococcal bacteriocin whose primary target is not
the plasma membrane. Microbiology 74, 5686±5692.
Morgan, S., Rossand, R.P. and Hill, C. (1995) Bacteriolytic activity
caused by the presence of a novel lactococcal plasmid encoding
lactococcin A, B and M. Applied and Environmental Microbiology 61,
2995±3001.
Moschetti, G., Villani, F., Blaiotta, G., Baldinelli, A. and Coppola, S.
(1996) Presence of non functional nisin genes in Lactococcus lactis
subsp. lactis isolated from natural starters. FEMS Microbiology
Letters 145, 27±32.
Nettles, C.G. and Barefoot, S.F. (1993) Biochemical and genetic
characteristics of bacteriocins of food-associated lactic acid bacteria.
Journal of Food Protection 56, 338±356.
Piard, J.C., Kuipers, O.P., Rollema, H.S., Desmazeaud, M.J. and De
Vos, W.M. (1993) Structure, organization and expression of the lct
gene for lacticin 481, a novel lantibiotic produced by Lactococcus
lactis. Journal of Biological Chemistry 268, 16361±16368.
Rodrigues, U.M., Aguirre, M., Facklam, R.R. and Collins, M.D.
(1991) Speci®c and intraspeci®c molecular typing of lactococci based
on polymorphism of DNA encoding rRNA. Journal of Applied
Bacteriology 71, 509±516.
Ryan, M.P., Rea, M.C., Hill, C. and Ross, P. (1996) An application in
cheddar cheese manufacture for a strain of Lactococcus lactis
producing a novel broad-spectrum bacteriocin, Lacticin 3147.
Applied and Environmental Microbiology 62, 612±619.
Schagger, H. and Von Jagow, G. (1987) Tricine-sodium dodecyl
sulfate-polyacrylamide gel electrophoresis for the separation of
proteins in the range from 1 to 100 KDa. Analytical Biochemistry
166, 369±379.
Smibert, R.M. and Krieg, N.R. (1994) Phenotypic characterization. In
Methods for General and Molecular Bacteriology ed. Gerhardt, P.,
Murray, R.G.E., Wood, W.A. and Kieg, N.R. pp. 607±654.
Washington, D.C.: American Society for Microbiology.
Sperber, W.H. and Swan, J. (1976) Hot loop test for the determination
of carbon dioxide production from glucose by lactic acid bacteria.
Applied and Environmental Microbiology 31, 990±991.
Stoffels, G., Sahl, H.-G. and GudmundsdoÁttir, A. (1993) Carnocin
UI49, a potential biopreservative produced by Carnobacterium
piscicola: large scale puri®cation and activity against gram-positive
bacteria including Listeria sp. International Journal of Food Micro-
biology 20, 199±210.
Teixeira, L.M., Merquior, V.L.C., Vianni, M.C.E. et al. (1996)
Phenotypic and genotypic characterization of atypical Lactococcus
garvieae strains isolated from water buffalos with subclinical mastitis
and con®rmation of L. garvieae as a senior subjective synonym of
Enterococcus seriolicida. International Journal of Systematic Bacterio-
logy 46, 664±668.
Terzaghi, B.E. and Sandine, W.E. (1975) Improved medium for lactic
streptococci and their bacteriophages. Applied Microbiology 29,
807±813.
van Belkum, M.J., Kok, J. and Venema, G. (1992) Cloning,
sequencing, and expression in Escherichia coli of lcnB, a third
bacteriocin determinant from the lactococcal bacteriocin plasmid
p9B4-6. Applied and Environmental Microbiology 58, 572±577.
Villani, F., Pepe, O., Moschetti, G., Salzano, G., Parente, E. and
Coppola, S. (1995) Antagonistic activity of lactic acid bacteria
isolated from natural whey starters for water-buffalo Mozzarella
cheese manufacture. Italian Journal of Food Science 3, 221±234.
Villani, F., Salzano, G., Sorrentino, E., Pepe, O., Marino, P. and
Coppola, S. (1993) Enterocin 226 NWC, a bacteriocin produced by
Enterococcus faecalis 226, active against Listeria monocytogenes.
Journal of Applied Bacteriology 74, 380±387.
Ward, L.J.H., Brown, J.C.S. and Davey, G.P. (1994) Application of
the ligase chain reaction to the detection of nisin A and nisin Z genes
in Lactococcus lactis ssp. lactis. FEMS Microbiology Letters 117,
29±34.
Yildirim, Z. and Johnson, M.G. (1998) Detection and characterization
of a bacteriocin produced by Lactococcus lactis subsp. cremoris R
isolated from radish. Letters in Applied Microbiology 26, 297±304.
Zlotkin, A., Eldar, A., Ghittino, C. and Bercovier, H. (1998)
Identi®cation of Lactococcus garvieae by PCR. Journal of Clinical
Microbiology 36, 983±985.
LACTOCOCCUS GARVIEAE BACTERIOCIN 439
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 430±439
Copyright © 2022 FDOKUMEN




















