
Complete Sequence of the Enterocin Q-Encoding Plasmid pCIZ2 from the Multiple Bacteriocin Producer...
15
10.1128/AEM.00859-06. 2006, 72(10):6653. DOI: Appl. Environ. Microbiol. Ingolf F. Nes, Pablo E. Hernández and Luis M. Cintas Raquel Criado, Dzung B. Diep, Ågot Aakra, Jorge Gutiérrez, and Immunity Characterization of Enterocin Q Production L50 and Genetic Enterococcus faecium Multiple Bacteriocin Producer Q-Encoding Plasmid pCIZ2 from the Complete Sequence of the Enterocin http://aem.asm.org/content/72/10/6653 Updated information and services can be found at: These include: REFERENCES http://aem.asm.org/content/72/10/6653#ref-list-1 at: This article cites 53 articles, 30 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on January 3, 2014 by guest http://aem.asm.org/ Downloaded from on January 3, 2014 by guest http://aem.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Complete Sequence of the Enterocin Q-Encoding Plasmid pCIZ2 from the Multiple Bacteriocin Producer...

10.1128/AEM.00859-06.
2006, 72(10):6653. DOI:Appl. Environ. Microbiol. Ingolf F. Nes, Pablo E. Hernández and Luis M. CintasRaquel Criado, Dzung B. Diep, Ågot Aakra, Jorge Gutiérrez, and ImmunityCharacterization of Enterocin Q Production
L50 and GeneticEnterococcus faeciumMultiple Bacteriocin Producer Q-Encoding Plasmid pCIZ2 from the Complete Sequence of the Enterocin
http://aem.asm.org/content/72/10/6653Updated information and services can be found at:
These include:
REFERENCEShttp://aem.asm.org/content/72/10/6653#ref-list-1at:
This article cites 53 articles, 30 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Oct. 2006, p. 6653–6666 Vol. 72, No. 100099-2240/06/$08.00�0 doi:10.1128/AEM.00859-06Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Complete Sequence of the Enterocin Q-Encoding Plasmid pCIZ2 fromthe Multiple Bacteriocin Producer Enterococcus faecium L50 and
Genetic Characterization of Enterocin Q Production and ImmunityRaquel Criado,1 Dzung B. Diep,2 Agot Aakra,2 Jorge Gutierrez,1 Ingolf F. Nes,2
Pablo E. Hernandez,1 and Luis M. Cintas1*Departamento de Nutricion, Bromatologıa y Tecnologıa de los Alimentos, Facultad de Veterinaria, Universidad Complutense de Madrid,
28040 Madrid, Spain,1 and Laboratory of Microbial Gene Technology, Department of Chemistry, Biotechnology andFood Science, Norwegian University of Life Sciences, P.O. Box 5003, N-1432 As, Norway2
Received 12 April 2006/Accepted 25 July 2006
The locations of the genetic determinants for enterocin L50 (EntL50A and EntL50B), enterocin Q (EntQ),and enterocin P (EntP) in the multiple bacteriocin producer Enterococcus faecium strain L50 were determined.These bacteriocin genes occur at different locations; entL50AB (encoding EntL50A and EntL50B) are on the50-kb plasmid pCIZ1, entqA (encoding EntQ) is on the 7.4-kb plasmid pCIZ2, and entP (encoding EntP) is onthe chromosome. The complete nucleotide sequence of pCIZ2 was determined to be 7,383 bp long and contains10 putative open reading frames (ORFs) organized in three distinct regions. The first region contains threeORFs: entqA preceded by two divergently oriented genes, entqB and entqC. EntqB shows high levels of similarityto bacterial ATP-binding cassette (ABC) transporters, while EntqC displays no significant similarity to anyknown protein. The second region encompasses four ORFs (orf4 to orf7), and ORF4 and ORF5 display highlevels of similarity to mobilization proteins from E. faecium and Enterococcus faecalis. In addition, featuresresembling a transfer origin region (oriT) were found in the promoter area of orf4. The third region containsthree ORFs (orf8 to orf10), and ORF8 and ORF9 exhibit similarity to the replication initiator protein RepEfrom E. faecalis and to RepB proteins, respectively. To clarify the minimum requirement for EntQ synthesis,we subcloned and heterologously expressed a 2,371-bp fragment from pCIZ2 that encompasses only the entqA,entqB, and entqC genes in Lactobacillus sakei, and we demonstrated that this fragment is sufficient for EntQproduction. Moreover, we also obtained experimental results indicating that EntqB is involved in ABCtransporter-mediated EntQ secretion, while EntqC confers immunity to this bacteriocin.
Bacteriocins are a heterogeneous group of ribosomally syn-thesized antimicrobial peptides produced by both gram-nega-tive and gram-positive bacteria (14, 33, 52). They often have anarrow inhibitory spectrum; i.e., they are most active againstclosely related bacteria likely to occur in the same ecologicalniche as the producer. However, many lactic acid bacteria(LAB) seem to compensate for the narrow inhibitory spectraby producing several bacteriocins (19). In many cases, bacte-riocin production is a plasmid-encoded trait; however, bacte-riocin genes have been also found on the chromosome andmobile genetic elements, such as transposon-like elements orphage DNA (17).
LAB bacteriocins belonging to class II are nonmodifiedsmall heat-stable peptides (19, 39, 40, 41) that are grouped intothree subclasses; (i) subclass IIa comprises pediocin-like bac-teriocins with a conserved N-terminal motif (YGNGVXC), (ii)subclass IIb comprises bacteriocins whose full activity is de-pendent on the presence of two different peptides, and (iii)subclass IIc includes other peptide bacteriocins. Class II bac-teriocins are commonly synthesized as biologically inactive pre-cursors containing an N-terminal extension (the so-called dou-ble-glycine-type leader sequence or the sec-dependent leader
peptide), which is cleaved off concomitant with externalizationof the active bacteriocin (30, 33, 39). Interestingly, a few bac-teriocins described to date are synthesized without an N-ter-minal extension, including enterocin L50 (11), enterocin Q(13), enterocin EJ97 (47), aureocin A70 (42), aureocin A53(43), and LsbB (28). Production of most subclass IIa bacterio-cins relies on a well-conserved genetic organization includingat least the following four genes that are often organized in oneor two operon-like structures in gene clusters: (i) the structuralgene encoding the prebacteriocin; (ii) a gene encoding theimmunity protein, which confers producer self-protection againstthe toxicity of the bacteriocin; (iii) a gene encoding a dedicatedATP-binding cassette (ABC) transporter required for process-ing and transport of the bacteriocin; and (iv) a gene encodingan accessory protein required for proper bacteriocin external-ization (30, 39, 52).
Enterococcus faecium L50, a strain isolated from a Spanishdry-fermented sausage (8, 9, 11), harbors two plasmids, pCIZ1(ca. 50 kb) and pCIZ2 (ca. 7.4 kb), and produces three differ-ent bacteriocins; (i) enterocin P (EntP) is a pediocin-like bac-teriocin (subclass IIa), (ii) enterocin L50 (EntL50) is a subclassIIb bacteriocin whose full activity is dependent on the peptidesEntL50A and EntL50B, and (iii) enterocin Q (EntQ) is asubclass IIc bacteriocin. While EntL50 and EntQ are synthe-sized as leaderless bacteriocins and are secreted by a hithertounknown mechanism, EntP is synthesized with a sec-depen-dent leader peptide (11, 13) and is secreted by the Sec trans-
* Corresponding author. Mailing address: Departamento de Nutri-cion, Bromatologıa y Tecnologıa de los Alimentos, Facultad de Vet-erinaria, Universidad Complutense de Madrid, 28040 Madrid, Spain.Phone: 34-913943751. Fax: 34-913943743. E-mail: [email protected].
6653
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

locase (31). Moreover, the production of multiple bacteriocinsby E. faecium L50 is a temperature-regulated process; EntL50is produced at 16 to 32°C but production is negligible when thegrowth temperature is above 37°C, whereas EntP and EntQare synthesized at temperatures ranging from 16 to 47°C (13;R. Criado, J. Gutierrez, M. Martın, C. Herranz, P. E. Hernan-dez, and L. M. Cintas, submitted for publication). The produc-tion of several bacteriocins confers to E. faecium L50 a broadspectrum of antimicrobial activity, including activity againstfood-borne pathogens, such as Listeria monocytogenes, Clos-tridium perfringens, and Clostridium botulinum, and activityagainst human and animal clinical pathogens, such as Strep-tococcus pneumoniae, Streptococcus mitis, Streptococcus oralis,Streptococcus parasanguis, and Streptococcus agalactiae (11, 13).
In this work, the genetic locations of all three bacteriocins inthis multiple bacteriocin producer were established by plasmidcuring and Southern analysis, and both plasmid pCIZ1 andplasmid pCIZ2 were found to be bacteriocinogenic. The com-plete nucleotide sequence of plasmid pCIZ2, which is respon-
sible for EntQ production and immunity, was determined.Furthermore, the minimum genetic requirements for EntQproduction and immunity were established by heterologousexpression experiments.
MATERIALS AND METHODS
Bacterial strains, plasmids, media, and culture conditions. Bacterial strainsand plasmids used in this work are listed in Table 1. All LAB strains were grownin MRS broth (Oxoid Ltd., Basingstoke, United Kingdom) at 32°C, unless other-wise stated. Escherichia coli cells were grown in LB broth (Sigma-Aldrich Inc.,St. Louis, Mo.) at 37°C with shaking. The following antibiotics (Sigma) wereadded to the media as selective agents when required: ampicillin (100 �g/ml forE. coli) and erythromycin (300 �g/ml for E. coli, 20 �g/ml for E. faecium, and 5�g/ml for Lactococcus lactis and Lactobacillus sakei). Pure enterocin Q waschemically synthesized at the Molecular Biology Unit, University of NewcastleUpon Tyne, Newcastle Upon Tyne, United Kingdom.
Bacteriocin and immunity assays. The antimicrobial activity of individualcolonies was screened by a stab-on-agar test as previously described by Cintas(8), using as the indicator microorganisms Pediococcus acidilactici 347 (EntL50sensitive [EntL50s], EntP resistant [EntPr], and EntQ resistant [EntQr]) (12), E.faecium T136 (EntL50s EntPs EntQr) (7), and E. faecium P13 (EntL50s EntPr
TABLE 1. Bacterial strains and plasmids used in this study
Strain or plasmid Relevant characteristics Source orreference
Bacterial strainsEscherichia coli TOP10 Host strain, F� mcrA �(mrr-hsdRMS-mcrBC) �8lacZ�M15 �lacW74 deoR recA1 araD139
�(ara-leu)7697 galU galK rpsL endA1 nupGInvitrogen
Enterococcus faecium strainsL50 Wild-type strain (pCIZ1�, pCIZ2�); EntL50, EntP, and EntQ producer; Emr 12LA5 E. faecium L50 cured derivative (pCIZ1�, pCIZ2�); EntL50 and EntP producer; EntQs 13L50/30-2 E. faecium L50 cured derivative (pCIZ1�, pCIZ2�); EntL50 and EntP producer; EntQs Emr This studyL50/14-2 E. faecium L50 cured derivative (pCIZ1�, pCIZ2�); EntP producer; EntL50s EntQs Ems This studyL50/14-2(pMG36e) E. faecium L50/14-2 derivative carrying pMG36e; EntP producer; EntL50s EntQs Emr This studyL50/14-2(pRCG04-1) E. faecium L50/14-2 derivative carrying pRCG04-1; EntP producer; EntL50s EntQr Emr This studyL50/14-2(pRCG04-2) E. faecium L50/14-2 derivative carrying pRCG04-2; EntP producer; EntL50s EntQr Emr This studyP13 EntP producer; indicator microorganism; EntL50s EntPr EntQs 10T136 EntA and EntB producer; indicator microorganism; EntL50s EntPs EntQr 7
Enterococcus faecalis JH2-2 Plasmid-free recipient strain, non-bacteriocin producer (Bac�); Rifr Fusr Erys 34
Pediococcus acidilactici 347 Pediocin PA-1 producer; indicator microorganism; EntL50s EntPr EntQr 12
Lactobacillus sakei strainsLb790 Plasmid-free host strain, non-bacteriocin producer (Bac�); EntQr Ems 49Lb790(pELS200) L. sakei Lb790 derivative carrying pELS200 This studyLb790(pRCG01) L. sakei Lb790 derivative carrying pRCG01; entqA This studyLb790(pRCG02) L. sakei Lb790 derivative carrying pRCG02; entqB entqC This studyLb790(pRCG03) L. sakei Lb790 derivative carrying pRCG03; entqA entqB entqC; EntQ producer; Emr This study
Lactococcus lactis DPC5598 Plasmid-free host strain; Ems 55
PlasmidspCIZ1 50-kb plasmid from E. faecium L50; EntL50� Imm� Emr 13pCIZ2 7.4-kb plasmid from E. faecium L50; EntQ� Imm� 13pGEM-3Zf(�) 3-kb cloning vector; Ampr PromegapELS200 7.8-kb E. coli-Lactobacillus shuttle vector; Ampr Emr 54pMG36e 3.6-kb gene expression vector carrying P32 promoter; Emr 56pGEM-CIZ2 pGEM-3Zf(�) derivative containing a 7.4-kb HindIII pCIZ2 fragment This studypRCG01 pELS200 derivative carrying entqA This studypRCG02 pELS200 derivative carrying entqB-entqC This studypRCG03 pELS200 derivative carrying enqA-entqB-entqC This studypRCG04-1 pMG36e derivative carrying truncated entqB (due to nucleotide substitution at coordinate
1519) and entqCThis study
pRCG04-2 pMG36e derivative carrying truncated entqB (due to nucleotide deletion at coordinate 1961)and entqC
This study
6654 CRIADO ET AL. APPL. ENVIRON. MICROBIOL.
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

EntQs) (10) (Table 1). The antimicrobial activities of cell-free culture superna-tants and chemically synthesized EntQ were detected by an agar diffusion test(ADT) and/or a microtiter plate assay (MPA) as previously described (13). Theimmunity tests were performed by using ADT and MPA in which the targetstrains were challenged with either chemically synthesized EntQ, supernatantsfrom a culture of E. faecium P13 grown at 32°C containing EntP (10), orsupernatants from a culture of E. faecium mutant strain LA5 grown at 32°Ccontaining EntP and EntL50 (13) (Table 1). The MICs of chemically synthesizedEntQ were determined by an MPA.
DNA isolation and manipulations. DNA for Southern hybridization wasisolated from E. faecium L50 by the alkaline lysis method, modified as pre-viously described (8). Small-scale plasmid DNA isolation from E. coli wascarried out using a QIAgen Spin Miniprep kit (QIAGEN GmbH, Hilden,Germany). Large-scale plasmid DNA isolation from E. coli, E. faecium, andL. lactis was carried out using a QIAgen Plasmid Midi kit (QIAGEN) after aninitial treatment with lysozyme (final concentration, 5 mg/ml) at 37°C for 15min when required. Restriction enzymes (New England Biolabs Inc., Beverly,Mass.), calf alkaline phosphatase (Promega Corporation, Madison, Wis.), T4DNA ligase (Promega), and Platinum Taq DNA polymerase (Invitrogen LifeTechnologies, Carlsbad, Calif.) were used according to the suppliers’ instruc-tions. PCR products were purified by using a QIAquick PCR purification kit(QIAGEN). Most DNA manipulations were carried out using standard pro-cedures described elsewhere (46).
Plasmid curing. Bacteriocin-deficient variants of E. faecium wild-type strainL50 were obtained by plasmid curing using novobiocin. E. faecium L50 wasinoculated into MRS broth (about 1 � 105 CFU/ml) with increasing concentra-tions (0 to 10 �g/ml) of novobiocin (Sigma) and incubated at 32°C for 72 h. Theculture that grew at the highest novobiocin concentration was serially diluted andplated onto MRS agar (1.5%, wt/vol) plates to obtain individual colonies. Ran-domly selected colonies were replica plated onto MRS agar plates and screenedfor bacteriocin production by the stab-on-agar test and ADT. The presence ofbacteriocin structural genes for EntL50 (entL50A and entL50B), EntP (entP),and EntQ (entqA) was investigated by PCR using the specific EntL50S1/EntL50S2, EntPS1/EntPS2, and EntQS1/EntQS2 primer pairs (Table 2) de-signed on the basis of the previously published EntL50, EntQ, and EntP operonsequences (11, 13). Plasmid profiles of the wild-type strain and the derivedmutants were determined and analyzed by agarose (0.8%, wt/vol) gel electro-phoresis.
Southern hybridization. DNA from E. faecium L50 was isolated, separated byagarose (0.8%, wt/vol) gel electrophoresis, and subsequently transferred onto apositively charged nylon membrane (Hybond-N�; Amersham BiosciencesGmbH, Freiburg, Germany) by vacuum blotting (VacuGene XL; PharmaciaLKB, Uppsala, Sweden) as described by Sambrook and Russell (46). Specific
DNA probes for the EntL50 operon (877 bp), EntQ operon (678 bp), and EntPoperon (682 bp) were prepared by PCR amplification from E. faecium L50 DNAusing the specific EntL50S1/EntL50S2, EntQS1/EntQS2, and EntPS1/EntPS2primer pairs (Table 2) and were labeled with horseradish peroxidase (ECLDirect nucleic acid labeling kit; Amersham Biosciences) according to the man-ufacturer’s instructions. Prehybridization (42°C, 15 min), hybridization (42°C,overnight), and posthybridization washes (0.5� SSC–0.4% sodium dodecyl sul-fate [SDS] at 42°C for 10 min twice; 2� SSC at 25°C for 5 min twice [1� SSC is0.15 M NaCl plus 0.015 M sodium citrate]) were carried out with a MiniovenMKII (Hybaid, Middlesex, United Kingdom). Hybridization detection was per-formed as suggested in the ECL Direct nucleic acid detection system user’s guidefor Southern hybridization (Amersham Biosciences). The chemiluminescencesignals were detected following exposure of blue-light-sensitive autoradiographyfilms (Hyperfilm-ECL; Amersham Biosciences).
DNA sequencing of pCIZ2 and sequence analysis. Plasmid pCIZ2 was linear-ized with HindIII, and the purified 7.4-kb fragment obtained was ligated into theHindIII site of the pGEM7Z vector, resulting in pGEM-CIZ2, which was used asthe starting point for sequencing by primer walking. The DNA on both strandswas sequenced by the dideoxy chain termination method (48), using an ABIPrism BigDye cycle sequencing Ready Reaction kit with AmpliTaq DNA poly-merase and dye-labeled terminators and an ABI Prism 377 DNA sequencer(Perkin-Elmer, Applied Biosystems Division, Foster City, Calif.). Sequenceswere assembled using the Autoassembler software package (Perkin-Elmer) andthe GCG software package of the Genetics Computer Group (University ofWisconsin, Madison, Wis.). To predict the location of putative open readingframes (ORFs) in pCIZ2, the ORF-finder software (National Center for Bio-technology Information [NCBI], U.S. National Library of Medicine) (http://www.ncbi.nlm.nih.gov/gorf/gorf.html) was used. The BLAST software (NCBI) (http://www.ncbi.nlm.nih.gov) was used to conduct similarity searches with GenBankand EMBL sequence databases. Multiple-sequence alignments of related aminoacid sequences were constructed using the ClustalW software (Baylor College ofMedicine Human Genome Sequencing Center) (http://searchlauncher.bcm.tmc.edu) and the Boxshade software (Swiss Node of EMBnet, Swiss Institute forExperimental Cancer Research) (http://www.ch.embnet.org). Transmembranehelices in putative proteins were predicted by using the TMHMM software(Center for Biological Sequence Analysis, Technical University of Denmark)(http://www.cbs.dtu.dk). The presence of protein motifs was examined by usingthe CDD software (NCBI) (http://www.ncbi.nlm.nih.gov) and InterProScan soft-ware (Swiss Institute of Bioinformatics) (http://www.expasy.org). The presenceof nucleotide and amino acid conserved sequences was examined by using theFuzznuc software and the Fuzztran software, respectively (The German NationalGenome Research Network) (http://ngfnblast.gbf.de). The presence or absenceof coiled-coil regions in putative proteins was predicted by using the PepCoil
TABLE 2. PCR primer pairs used in this study
Oligonucleotides PCR productsize (bp)Designation Sequence (5�–3�)a
EntL50S1 TCCCTACAGTCTCCCTTCC 877EntL50S2 TCTAGCGTTAAGCCGAATG
EntPS1 TTATGCGCGTTATTGTGTG 682EntPS2 CATAACTCAAAGTCCCGACC
EntQS1 AACAAGAAAATTGCGGCTG 678EntQS2 AGGGCTACTTGGATAGTACAC
EntQC1XbaI CGCGTAACTCTAGAGAAATATTGCATAGCAACCTAGTG 2,207EntQC2XhoI GTACTACCTCGAGATCCATTTTGCGATACCATT
EntQC1KpnI GGTGGTGGTACCGAAATATTGCATAGCAACCTAG 2,207EntQC2SacI GGTGGTGAGCTCATCCATTTTGCGATACCATT
EntQC1XbaI CGCGTAACTCTAGAGAAATATTGCATAGCAACCTAGTG 2,371EntQC4XhoI TAATTAATCTCGAGCTAATCGTATAAATAACGAAGGAAATA
EntQC3XbaI CGCGTAACTCTAGATACTGATATGAGACTAAAAAGATTGG 381EntQC4XhoI TAATTAATCTCGAGCTAATCGTATAAATAACGAAGGAAATA
a Restriction sites are underlined. Boldface type indicates nucleotides in the nucleotide tail added to the specific sequences to ensure proper functioning of therestriction enzymes.
VOL. 72, 2006 pCIZ2-ENCODED ENTEROCIN Q PRODUCTION AND IMMUNITY 6655
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from
software (The German National Genome Research Network) (http://ngfnblast.gbf.de).
Plasmid construction, DNA transformation, and mating experiments. To in-vestigate the biological functions of entqB and entqC, several expression cloneswere constructed in E. coli TOP10 or L. lactis DPC5598 (55) and subsequentlytransferred to L. sakei Lb790 (49) or E. faecium L50/14-2 (Table 1). Inserts wereobtained by PCR amplification of entqA, entqBC, and entqABC using E. faeciumL50 plasmid DNA as the template and the specific primers carrying terminalrestriction sites listed in Table 2. Recombinant plasmids (Table 1) were con-structed by cloning entqA and/or entqBC into the cloning vectors pMG36e (56)and pELS200 (54). The entqA gene was PCR amplified from pCIZ2 with primers
EntQC3XbaI and EntQC4XhoI as a 381-bp fragment, digested with XbaI andXhoI, and ligated into the corresponding sites in dephosphorylated pELS200,resulting in pRCG01. DNA fragments containing entqB and entqC were ampli-fied from pCIZ2 with the EntQC1XbaI/EntQC2XhoI and EntQC1KpnI/EntQC2SacI primer pairs and digested with the appropriate restriction enzymesprior to ligation into the corresponding sites in dephosphorylated pELS200 andpMG36e, resulting in pRCG02 and in pRCG04-1 and pRCG04-2, respectively.The entqBC insert in pRCG04-1 and pRCG04-2 contains a single mutation(substitution of C by T at coordinate 1,519 in pRCG04-1 and deletion of T atcoordinate 1,961 in pRCG04-2); the result of this is that both plasmids encodeintact EntqC but truncated variants of EntqB. These mutations were spontane-
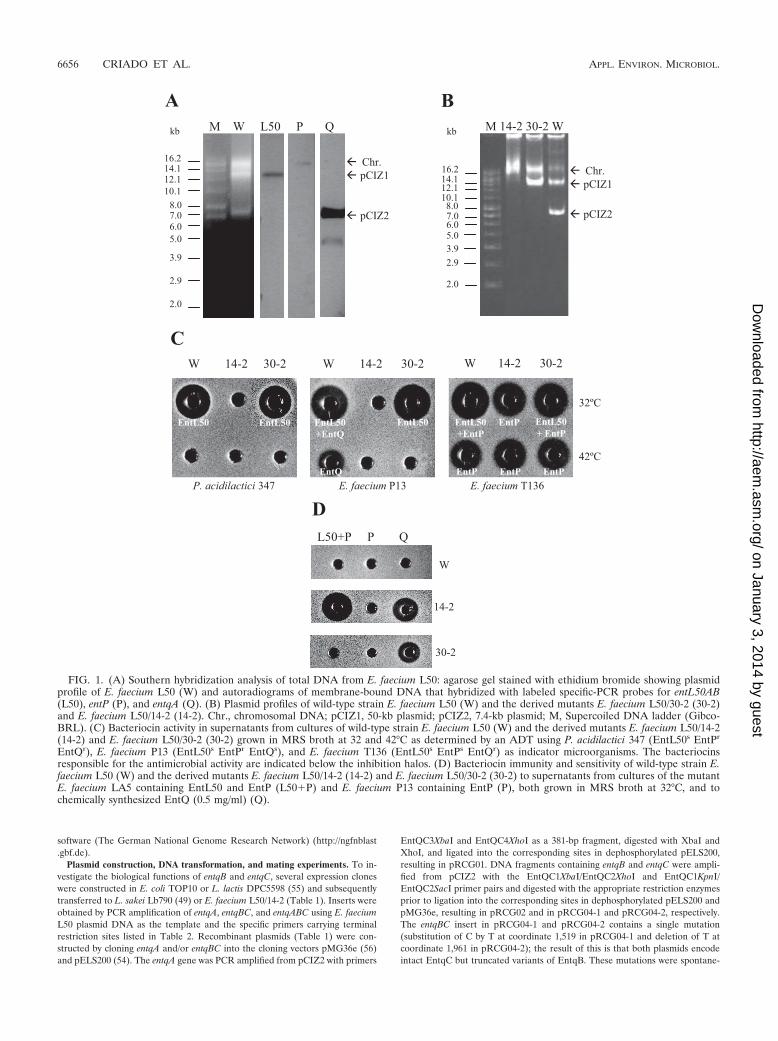
FIG. 1. (A) Southern hybridization analysis of total DNA from E. faecium L50: agarose gel stained with ethidium bromide showing plasmidprofile of E. faecium L50 (W) and autoradiograms of membrane-bound DNA that hybridized with labeled specific-PCR probes for entL50AB(L50), entP (P), and entqA (Q). (B) Plasmid profiles of wild-type strain E. faecium L50 (W) and the derived mutants E. faecium L50/30-2 (30-2)and E. faecium L50/14-2 (14-2). Chr., chromosomal DNA; pCIZ1, 50-kb plasmid; pCIZ2, 7.4-kb plasmid; M, Supercoiled DNA ladder (Gibco-BRL). (C) Bacteriocin activity in supernatants from cultures of wild-type strain E. faecium L50 (W) and the derived mutants E. faecium L50/14-2(14-2) and E. faecium L50/30-2 (30-2) grown in MRS broth at 32 and 42°C as determined by an ADT using P. acidilactici 347 (EntL50s EntPr
EntQr), E. faecium P13 (EntL50s EntPr EntQs), and E. faecium T136 (EntL50s EntPs EntQr) as indicator microorganisms. The bacteriocinsresponsible for the antimicrobial activity are indicated below the inhibition halos. (D) Bacteriocin immunity and sensitivity of wild-type strain E.faecium L50 (W) and the derived mutants E. faecium L50/14-2 (14-2) and E. faecium L50/30-2 (30-2) to supernatants from cultures of the mutantE. faecium LA5 containing EntL50 and EntP (L50�P) and E. faecium P13 containing EntP (P), both grown in MRS broth at 32°C, and tochemically synthesized EntQ (0.5 mg/ml) (Q).
6656 CRIADO ET AL. APPL. ENVIRON. MICROBIOL.
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

ously introduced during PCRs. A 2,371-bp fragment encompassing entqABC wasamplified from pCIZ2 using primers EntQC1XbaI and EntQC4XhoI, digestedwith the appropriate restriction enzymes, and ligated into the corresponding sitesin dephosphorylated pELS200, resulting in pRCG03. The integrity of all PCR-generated fragments was confirmed by DNA sequencing.
E. coli TOP10 was used as an intermediate host for amplification of plasmidspRCG01, pRCG02 and pRCG03, whereas L. lactis DPC5598 was used foramplification of pRCG04-1 and pRCG04-2. Recombinant plasmids were reiso-lated and then transferred to L. sakei Lb790 or E. faecium L50/14-2 by electro-poration as previously described (2, 32).
The transferability of bacteriocin production, as well as erythromycin resis-tance (Eryr) determinants, was tested by conjugation using a filter method (18),with plasmid-free Enterococcus faecalis JH2-2 as the recipient strain (rifampinresistant [Rifr], fusidic acid resistant [Fusr], erythromycin susceptible [Erys], andnonbacteriocin producer [Bac�]) and E. faecium L50 as the donor strain (Rifs
Fuss Eryr Bac�).NCI-ELISA. The presence of EntQ in cell-free supernatants and intracel-
lular extracts (5) obtained from growing (log-phase) or overnight cultures ofL. sakei Lb790 and the recombinant strains L. sakei Lb790(pRCG01), L. sakeiLb790(pRCG02), and L. sakei Lb790(pRCG03) was determined and quantifiedby a noncompetitive indirect enzyme-linked immunosorbent assay (NCI-ELISA)essentially as previously described (29), using rabbit polyclonal antibodies withspecificity for EntQ (anti-EntQ-KLH) (Criado et al., submitted for publication).Briefly, wells of flat-bottom polysterene microtiter plates (Maxisorp; Nunc,Roskilde, Denmark) were coated overnight (4°C) with supernatants or intracel-lular extracts. After coating, wells were blocked and then washed. Next, dilutedanti-EntQ-KLH serum was added to each well, unbound antibodies were re-moved by washing, and goat anti-rabbit immunoglobulin G-peroxidase conjugate(Cappel Laboratories, West Chester, PA) was added. The amount of boundperoxidase was determined using ABTS [2,2�-azinobis(3-ethylbenzthiazoline-6-sulfonic acid)]) (Sigma) as the substrate by measuring the absorbance at 405 nmof the wells with an iEMS reader (Labsystems, Helsinki, Finland). The platesincluded control wells coated with (i) coating buffer (0.1 M sodium carbonate-bicarbonate buffer, pH 9.6)-MRS broth to determine the background level of theplate and (ii) six twofold dilutions of samples containing known concentrations ofpure EntQ (determined from the A280 using the molar extinction coefficient) incoating buffer-MRS broth to determine a standard curve.
Bacteriocin purification and mass spectrometry. The EntQ heterologouslyproduced by L. sakei Lb790(pRCG03) was purified from a 1-liter culture grownin MRS broth at 32°C for 14 h, as previously described by Cintas et al. (13). Thefraction corresponding to the peak containing the purified bacteriocin was sub-jected to mass spectrum analysis by using a matrix-assisted laser desorptionionization—time of flight Voyager-DE STR mass spectrometer (PerSeptive Bio-systems, Foster City, Calif.).
Tricine-SDS-PAGE analysis, Western blotting, and overlay assay. The purityof EntQ at the final reversed-phase fast protein liquid chromatography purifi-cation step was also analyzed by sodium dodecyl sulfate-polyacrylamide gelelectrophoresis on 16% Tricine gels (Tricine-SDS-PAGE) (51) after silver stain-ing. Western blotting using anti-EntQ-KLH antibodies was performed essen-tially as previously described (29). To determine the antimicrobial activity ofthe purified heterologously produced EntQ, an overlay assay (6) was per-formed using E. faecium P13 (about 1 � 105 CFU/ml) as the indicatormicroorganism.
Nucleotide sequence accession number. The nucleotide sequence reported inthis paper has been deposited in the GenBank database under accession no.DQ832184.
RESULTS
Localization of the bacteriocin structural genes for EntL50,EntQ, and EntP. E. faecium L50 harbors two plasmids, pCIZ1(about 50 kb) and pCIZ2 (about 7.4 kb) (11, 13). To examinewhether the genetic determinants responsible for bacteriocinproduction are plasmid encoded in E. faecium L50, plasmidcuring was performed using novobiocin (3 �g/ml) as the curingagent. Two plasmid profiles were obtained from the plasmid-cured mutants; the type I profile had only the larger plasmid,pCIZ1, while the type II profile had no plasmid, as shown forone isolate (designated 30-2) with the type I profile and oneisolate (designated 14-2) with the type II profile in Fig. 1B andTable 3. Isolate 30-2 still produces EntL50 and EntP but is notable to produce EntQ (Fig. 1C), indicating that one or more ofthe genetic determinants required for EntQ production arelocated on pCIZ2. Likewise, only EntP activity and not EntL50or EntQ activity were detected in isolate 14-2, indicating thatEntP determinants are chromosomally located. Together, theresults also imply that the genetic determinants for EntL50 arelocated on pCIZ1. These results were confirmed by Southernanalysis using DNA probes specific for the different bacteriocingenes. Thus, we found that the entP-specific probe hybridizedto the chromosome, while the probes specific to entL50AB andentqA hybridized to pCIZ1 and pCIZ2, respectively (Fig. 1A).
To examine whether the genes conferring immunity are ge-netically linked with their cognate bacteriocin structural genes,as commonly found in other bacteriocin systems (17, 40), weexamined their immunity potentials by exposing the isolates tovarious bacteriocins on agar plates. As expected, isolate 30-2,which did not contain the EntQ-encoding plasmid (pCIZ2),was sensitive only to EntQ and not to EntP or EntL50, whereasisolate 14-2, which did not contain either plasmid, was sensitiveto both EntL50 and EntQ but not to EntP, which is chromo-somally encoded (Fig. 1D).
DNA sequence of pCIZ2. The complete nucleotide sequenceof pCIZ2 (Fig. 2) was determined by primer walking. It is 7,383bp long, and its G�C content is 32.3%, which is lower than thevalue reported for the E. faecium genomic DNA (38.3 to 39%)(38). Computer analysis of the pCIZ2 sequence revealed thepresence of 10 putative ORFs (orf1 to orf10) (Table 4), eachpreceded by a putative ribosome binding site (Fig. 2). All ofthese ORFs except orf1 have the same direction on the plasmid(Fig. 2). The ORFs on pCIZ2 seem to be organized into threedistinct functional regions (Fig. 3).
Region I is comprised of three ORFs (orf1 to orf3). orf1corresponds to the enterocin Q structural gene entqA (previ-ously reported as entQ) (13), which is preceded by two putative
TABLE 3. Presence of EntL50 (entL50A and entL50B), EntP (entP), and EntQ (entqA) structural genes, phenotypes, and plasmid profilesfor wild-type strain E. faecium L50 and the derived mutants E. faecium L50/30-2 and E. faecium L50/14-2
StrainStructural genes
Phenotypes Plasmid profileentL50ABa entP entqA
E. faecium L50 � � � EntL50� Imm�/EntP� Imm�/EntQ� Imm� pCIZ1�/pCIZ2�
E. faecium L50/30-2 � � � EntL50� Imm�/EntP� Imm�/EntQ� Imm� pCIZ1�/pCIZ2�
E. faecium L50/14-2 � � � EntL50� Imm�/EntP� Imm�/EntQ� Imm� pCIZ1�/pCIZ2�
a entL50AB corresponds to a PCR fragment containing entL50A and entL50B amplified using primers EntL50S1 and EntL50S2.
VOL. 72, 2006 pCIZ2-ENCODED ENTEROCIN Q PRODUCTION AND IMMUNITY 6657
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

FIG. 2. Nucleotide sequence of the 7,383-bp plasmid pCIZ2. The deduced amino acid sequences are shown below the DNA sequence. Putativeribosome binding sites (RBS) are indicated by boldface type. The putative �35 and �10 promoter sequences are underlined, and start codons areindicated by boldface italics. Stop codons are indicated by asterisks at the ends of protein sequences. The horizontal arrows under the nucleotidesequence indicate inverted repeats. Direct repeats are enclosed in boxes and overlined. Primers used in this study are indicated by dashedunderlining. Transmembrane segments are underlined in the amino acid sequence. Nucleotide and amino acid motifs and conserved sequences areenclosed in boxes. The vertical solid and open arrows indicate the nucleotides substituted and deleted in the truncated entqB encoded by pRCG04-1and pRCG04-2, respectively. The 1,020-bp sequence from coordinates 40 to 1060 was previously published by Cintas et al. (13).
6658
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

FIG. 2—Continued.
VOL. 72, 2006 pCIZ2-ENCODED ENTEROCIN Q PRODUCTION AND IMMUNITY 6659
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

promoters (Fig. 2). Downstream of entqA are two invertedrepeats (IR1L and IR1R) that could serve as a rho-indepen-dent transcriptional terminator. Together, these findings sug-gest that the structural bacteriocin gene can be transcribed asa monocistronic unit (Fig. 2).
orf2 (designated entqB), which is divergently located 79 bpupstream of entqA, is preceded by a putative consensus pro-moter (Fig. 2). entqB encodes a 572-amino-acid protein withhigh levels of sequence homology to many prokaryotic ABCtransporters, including LmrB and AurT, which are involved inexport of the leaderless bacteriocins LsbB and aureocin A70,respectively (28, 42) (Table 4). Sequence analyses of EntqBrevealed the presence of the following features typical of ABCtransporters: (i) the Walker A and B motifs; (ii) a highlyconserved C motif (linker peptide) preceding the Walker Bmotif; and (iii) the so-called “switch region,” located after theC motif (15, 21, 30, 50) (Fig. 4A). Moreover, analysis of theprimary sequence of this polypeptide revealed extensive hydro-phobic stretches (residues 18 to 40, 53 to 75, 124 to 146, 153 to175, 244 to 266, and 273 to 295) that are likely to form sixputative transmembrane segments.
orf3 (designated entqC), located immediately downstream of
FIG. 3. Physical map showing the genetic organization of the7,383-bp plasmid pCIZ2. ORFs are represented by arrows indicatingthe direction of transcription. Inverted repeats are indicated by stem-loop symbols. Direct repeats are indicated by boxes.
TABLE 4. ORFs in 7,383-bp plasmid pCIZ2 from E. faecium L50 and the closest relatives of the deduced proteins
ORF
Position innucleotidesequence
Gene Protein
5� 3� Designation G�Ccontent (%)
Length(amino acids)
Molecularmass
(kDa)pI
Closest relative (length; e value, level ofamino acid identity/level of amino
acid similarity; microorganism)Accession no.
ORF1 259 155 entqA 33.33 34 3.95 9.39 Structural gene of enterocin Q A4318955
ORF2 340 2058 entqB 25.48 572 64.66 9.51 Ej97B, ABC transporter (583 amino acids;e�123, 41%/65%; E. faecalis)
CAD35294
LmrB, MDR transporter (567 aminoacids; e�119, 41%/63%; L. lactis)
NP_861550
ORF3 2069 2272 entqC 20.59 67 7.69 9.63 No significant similarity found
ORF4 2686 3012 orf4 44.95 108 12.11 9.89 Mobilization protein (129 amino acids;5e�46, 78%/78%; E. faecium)
EAN08864
MobC, mobilization protein (127 aminoacids; 3e�16, 61%/77%; E. faecalis)
NP_863267
ORF5 2994 3908 orf5 39.23 304 35.77 9.36 Relaxase/mobilization nuclease domain(304 amino acids; e�167, 94%/97%;E. faecium)
EAN08860
MobA, mobilization protein (346 aminoacids; 3e�64, 44%/58%; E. faecalis)
NP_863268
ORF6 3700 4146 orf6 40.94 148 17.70 5.98 Hypothetical protein (538 amino acids;1e�4, 34%/55%; Mycoplasma synoviae)
YP_278611
ATP synthase F0, subunit b (171 aminoacids; 0.001, 26%/49%; Helicobacterpylori)
NP_223781
ORF7 4148 4837 orf7 33.33 229 27.09 8.74 Hypothetical protein (229 amino acids;2e�120, 94%/98%; E. faecium)
EAN08861
ORF8 5690 6427 orf8 26.00 245 29.17 9.46 RepE, replication protein (240 aminoacids; 2e�44, 40%/62%; E. faecalis)
NP_863355
ORF9 6455 6991 orf9 33.89 178 21.10 5.40 Protein with unknown function (179amino acids; 1e�91, 96%/98%;E. faecium)
EAN08855
RepB, replication protein (168 aminoacids; 1e�11, 28%/52%; L. casei)
NP_542222
ORF10 7007 7177 orf10 30.41 56 6.33 9.16 No significant similarity found
6660 CRIADO ET AL. APPL. ENVIRON. MICROBIOL.
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

FIG. 4. (A) Alignment of the predicted amino acid sequence of EntqB encoded by pCIZ2 (EntqB-pCIZ2) with the sequences of severalbacterial ABC transporters, including those from Bacillus halodurans (ABC-Bh) (accession no. NP_241579), Lactobacillus johnsonii (ABC-Lj)(accession no. NP_965013), Oceanobacillus iheyensis (ABC-Oi) (accession no. NP_691347), S. aureus (AurT [accession no. AAK73550] andABC-A-Sa [accession no. CAA62898]), Staphylococcus epidermidis (PepT [accession no. CAA90021] and ABC-Se [accession no. YP_187899]), andE. faecalis (Ej97B) (accession no. CAD35294), and with the sequences of ABC-type multidrug transporters (MDR) from Bacillus thuringiensis(MDR-Bt) (accession no. YP_036121), L. lactis (LmrB [accession no. AAC14278] and MDR-Ll [accession no. NP_266867]), and Lactobacillusbrevis (HorA) (accession no. BAD80897). (B) Alignment of the predicted amino acid sequence of ORF5 encoded by pCIZ2 with the predictedamino acid sequences of several plasmid relaxases and mobilization proteins, including those from E. faecium (RLX-Ef [pRUM; accession no.NP_863170] and VirD2-Ef [accession no. ZP_00287779]), E. faecalis (MobA-Efs [pEF1071; accession no. AAN76342], RLX-Efs [pAD1; accessionno. AAF72355], and orf-Efs [accession no. NP_815959]), Pediococcus pentosaceus (Mob-Pp [pMD136; accession no. AAD39619]), S. aureus(RLX-Sa [pC221; accession no. CAA26106]), and S. epidermidis (RLX-Se [accession no. NP_863255]). Identical residues and conservativesubstitutions are indicated by black and gray backgrounds, respectively. The conserved motifs are indicated above the sequences. Asterisks and dotsin the consensus sequence indicate highly conserved and 100% conserved residues, respectively. Solid and open arrows indicate the last amino acidsof the truncated EntqB proteins encoded by pRCG04-1 and pRCG04-2, respectively.
VOL. 72, 2006 pCIZ2-ENCODED ENTEROCIN Q PRODUCTION AND IMMUNITY 6661
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

entqB (Fig. 2), encodes a putative 67-amino-acid peptide con-taining two possible transmembrane segments. However, EntqCshows no significant sequence similarity to any known protein indata banks.
Region II is comprised of four ORFs (orf4 to orf7). orf4 ispreceded by a putative promoter (Fig. 2) and encodes a puta-tive 108-amino-acid protein which most closely resembles amobilization protein from E. faecium and the mobilizationprotein MobC from E. faecalis (3, 4) (Table 4). Features typicalof a transfer origin (oriT) (24, 35, 53, 58) were found in theregion containing the putative promoter of orf4; these featuresinclude (Fig. 2) (i) two inverted repeats (IR3 and IR4) and (ii)a sequence (GAGCTTGC) with homology to the core se-quence of the cis-acting nic site of the IncP/MobP family ofgram-positive plasmids (RYGCTTGC) (53).
orf5, which overlaps the end of orf4, encodes a 304-amino-acid protein with high levels of sequence similarity to a relax-ase/mobilization nuclease domain from E. faecium and to themobilization protein MobA from E. faecalis (4) (Table 4).Typical features for this protein family include the presence ofthree highly conserved motifs (motifs I, II, and III) at theN-terminal relaxase domain (24, 44) (Fig. 4B). The presence oftwo mob genes and a possible oriT sequence suggested thatpCIZ2 could be mobilizable. However, numerous mating ex-periment attempts, carried out with the plasmidless recipientstrain E. faecalis JH2-2 (Rifr Fusr Erys) (34) and the donorstrain E. faecium L50 (Rifs Fuss Eryr), were unsuccessful.
orf6, which overlaps the end of orf5, encodes a 148-amino-acid protein which shows similarity to a hypothetical proteinfrom Mycoplasma synoviae and to subunit b of ATP synthaseF0 from Helicobacter pylori (1) (Table 4).
orf7, which is immediately downstream of orf6 (Fig. 2),encodes a putative 229-amino-acid protein exhibiting a highlevel of similarity to a hypothetical protein from E. faecium(Table 4).
Finally, region III is comprised of three ORFs (orf8 toorf10). orf8 is preceded by a putative promoter (Fig. 2) andis predicted to encode a 245-amino-acid protein with simi-larity to the replication initiator protein RepE from E. fae-calis (23) (Table 4). The region containing the putativepromoter of orf8 includes two sets of direct repeats (12 bpand 22 bp), which is a feature typical of a putative plasmidreplication origin (ori) (20).
orf9, which is located 26 bp downstream of orf8, is predictedto encode a 178-amino-acid protein with a putative DNA-binding domain, exhibiting sequence similarity to a proteinhaving an unknown function from E. faecium and to RepBproteins in data banks (Table 4).
orf10, found 13 bp downstream of orf9, encodes a putative65-amino-acid peptide showing no significant sequence simi-larity to any known protein in data banks.
EntqBC is involved in secretion and immunity. Genes in-volved in transport and immunity are often closely associatedwith bacteriocin structural genes (17, 41, 52). We are thereforeinterested in determining whether entqBC are involved in sucha relationship. To examine their possible roles in transport,different DNA fragments from entqABC were cloned into theexpression vector pELS200. The resulting recombinant plas-mids, including pRCG01 containing only entqA, pRCG02containing entqBC but not entqA, and pRCG03 containing
entqABC (Table 1), in addition to an empty plasmid, weretransferred into the heterologous host L. sakei Lb790, which isa naturally EntQ-resistant strain. Bacteriocin production inculture supernatants from the resulting Lb790 clones wasassessed by an ADT. As shown in Fig. 5A, only cloneLb790(pRCG03), expressing all three genes (entqABC), wasable to produce antimicrobial activity in supernatant againstgrowth of E. faecium P13, while the other two clones,Lb790(pRCG01) and Lb790(pRCG02), were not able to dothis. The presence of EntQ in the supernatant of Lb790(pRCG03) was confirmed, and the EntQ quantified by anNCI-ELISA using anti-EntQ-KLH-specific antibodies (datanot shown). The maximum heterologous production of EntQ(390 ng/ml) by Lb790(pRCG03) was obtained after 14 h ofgrowth at 32°C in MRS broth; this corresponds to 22% of themaximum amount of EntQ produced by cultures of E. faeciumL50 grown under similar conditions (Criado et al., submittedfor publication). While no EntQ was found in the culturesupernatant of Lb790(pRCG01), the bacteriocin was detectedin the intracellular extract from an overnight culture by anNCI-ELISA (data not shown), demonstrating that the regionencompassing entqBC is involved in ABC transporter-medi-ated secretion of EntQ.
During cloning of the fragment containing entqBC into lac-tococcal plasmid pMG36e, we obtained two plasmid clones,each of which had a mutation in entqB. The mutations causedtruncation of EntqB, while EntqC was still intact in these
FIG. 5. (A) Bacteriocin activity in supernatants from cultures of L.sakei Lb790(pELS200) (HpELS), E. faecium L50 (W), L. sakeiLb790(pRCG01) (Hp01), L. sakei Lb790(pRCG02) (Hp02), and L.sakei Lb790(pRCG03) (Hp03) grown in MRS broth at 32°C and MRSbroth containing erythromycin (5 �g/ml) (MRS-E) as determinedby an ADT using E. faecium P13 (P13) (EntL50s EntPr EntQs) asthe indicator microorganism. (B) Bacteriocin immunity or sensitiv-ity of E. faecium L50/14-2(pRCG04-1) (Hp04-1), E. faecium L50/14-2(pRCG04-2) (Hp04-2), and E. faecium L50/14-2(pMG36e) (HpMG)to supernatants from cultures of the mutant E. faecium LA5 containingEntL50 and EntP (L50�P) and E. faecium P13 containing EntP (P),both grown in MRS broth at 32°C, and to chemically synthesized EntQ(0.5 mg/ml) (Q).
6662 CRIADO ET AL. APPL. ENVIRON. MICROBIOL.
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

clones. The truncated EntqB (393 amino acids) encoded by thepRCG04-1 clone lacked most of the C-terminal half, includingthe C motif (involved in ATP hydrolysis and mediating con-formational changes induced by ATP hydrolysis between theATPase and the transmembrane domains), the Walker B motif(involved in nucleotide binding and ATPase activity), and theswitch region (involved in signal transduction to the ABC do-main by sensing conformational changes in the membrane-spanning domains upon substrate binding) (15, 50). The sec-ond clone (pRCG04-2) with a truncated version of EntqB (540amino acids) lacked the last 32 amino acids that encompass theswitch region (Fig. 4A). Therefore, we assumed that the func-tionality of entqB, which encodes an ABC transporter and isinvolved in secretion of EntQ, was destroyed in the two clones,thereby providing the possibility of examining whether theremaining entqC gene in these two clones can confer immunity.These two plasmid clones were therefore transformed into theEntQ-sensitive plasmidless mutant E. faecium L50/14-2 (Table1), and the resulting transformants were examined for immu-nity to EntQ. The ADT immunity analyses were performedwith the transformants E. faecium L50/14-2(pRCG04-1) and E.faecium L50/14-2(pRCG04-2), and the results conclusivelyshowed that both strains were immune to EntQ, while thecontrol clone E. faecium L50/14-2(pMG36e) was inhibited bythis bacteriocin under the same conditions (Fig. 5B). Interest-ingly, the acquired immunity was specific to EntQ as these twoclones were sensitive to EntL50 (Fig. 5B). Besides, by using anMPA, it was shown that the EntQ immunity level of E. faeciumL50/14-2(pRCG04-1) and E. faecium L50/14-2(pRCG04-2)was the same as that exhibited by the wild-type strain E. fae-cium L50; i.e., their growth was not affected by chemicallysynthesized EntQ at a concentration of 125 �g/ml. In compar-
ison, the growth of the control clones E. faecium L50/14-2(pMG36e) and E. faecium L50/14-2 was inhibited by a muchlower concentration of EntQ (2.0 �g/ml). Taken together, allthese results established that EntqC is involved in EntQ-ded-icated immunity, conferring full protection against the toxicityof this bacteriocin.
Purification and characterization of the heterologously pro-duced EntQ. The EntQ heterologously produced by L. sakeiLb790(pRCG03) was purified to homogeneity. The last re-versed-phase fast protein liquid chromatography resulted in asingle well-separated absorbance peak that coincided with theantimicrobial activity peak (results not shown). The purity andmolecular mass of EntQ were confirmed by matrix-assistedlaser desorption ionization—time of flight mass spectrometry,which revealed a single peptide with the expected molecularmass (3,952 Da) (13). The purified EntQ produced a band ofthe expected size similar to the band produced by chemicallysynthesized EntQ when it was analyzed on a silver-stainedTricine-SDS-PAGE gel (Fig. 6A). The purified bacteriocin andbacteriocin aggregates, likely due to hydrophobic interactions,were detected by Western blotting using the anti-EntQ-KLHantibodies specific to EntQ (Fig. 6B), and they were shown tobe biologically active by an overlay assay (Fig. 6C). Based onthese results, we concluded that the heterologously producedEntQ is identical to the EntQ produced by the wild-type strainE. faecium L50 (13).
DISCUSSION
Genetic determinants responsible for multiple bacteriocinproduction have been described for a number of systems. Inthe case of the plantaricin (16) and sakacin (37) systems, thegenes involved are clustered in a locus-like unit and exposed toa common regulatory system. In this work, we demonstratedthat the genetic determinants involved in the multiple bacte-riocin production by E. faecium L50 are organized differentlyand occur at different genetic locations, as follows: entL50AB,entqA, and entP are located on the 50-kb plasmid pCIZ1, the7.4-kb plasmid pCIZ2, and the chromosome, respectively (Fig.1A). Likewise, Carnobacterium piscicola LV17 produces threecarnobacteriocins, carnobacteriocins A, BM1, and B2, whosestructural genes are located on the chromosome and 72-kb and61-kb plasmids, respectively (45, 57). The genetic organizationfound in the enterocin system might allow production of theindividual bacteriocins to be subjected to different regulatoryregimens, each of which is dependent on specific environmen-tal signals and growth conditions. In fact, the level of produc-tion of EntL50 is high at 16 to 25°C but negligible when thegrowth temperature is above 37°C, whereas EntP and EntQare synthesized in a wider temperature window, from 16 to47°C (13; Criado et al., submitted for publication).
Nucleotide sequence analysis of pCIZ2 from E. faecium L50revealed the presence of the EntQ structural gene (entqA) andtwo divergently oriented genes, entqB and entqC, which wereshown to be involved in ATP-mediated bacteriocin transportand immunity, respectively. These notions are based on theresults of expression of these genes in L. sakei Lb790 and E.faecium L50/14-2. L. sakei Lb790(pRCG01) expressing onlyentqA was found to be unable to secrete the bacteriocin intothe growth medium. Only when entqA was coexpressed with
FIG. 6. (A) Tricine-SDS-PAGE of EntQ heterologously producedby L. sakei Lb790(pRCG03) after silver staining. (B) Western blottingusing specific anti-EntQ-KLH antibodies. (C) Antimicrobial activityafter overlay with the indicator strain E. faecium P13. Lane 1, 1 �g ofpure chemically synthesized EntQ; lane 2, pure EntQ heterologouslyproduced by L. sakei Lb790(pRCG03). The positions of ultra-low-range molecular mass markers (Sigma) are indicated on the left.
VOL. 72, 2006 pCIZ2-ENCODED ENTEROCIN Q PRODUCTION AND IMMUNITY 6663
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

entqBC in L. sakei Lb790(pRCG03) was the bacteriocin se-creted into the medium (Fig. 5A). Similarly, the cloned entqC(together with truncated and apparently nonfunctional entqBmutants) was found to confer immunity to E. faecium L50/14-2(pRCG04-1/2) (Fig. 5B).
The role of ABC transporters in processing and secretion ofleader-containing class II bacteriocins is well documented (14,33, 36, 39, 41, 52). Furthermore, it has been reported thatsecretion of the leaderless bacteriocin LsbB from L. lactisBGM-1 (28) and secretion of aureocin A70 from Staphylococ-cus aureus A70 (42) are mediated by the ABC-type multidrugresistance transporter LmrB and the ABC transporter AurT,respectively; also, it has been suggested that secretion of theleaderless enterocin EJ97 from E. faecalis EJ97 is mediated bythe ABC transporter Ej97B (47). Interestingly, EntqB exhibitsextensive homology to these ABC-type transporters (Table 4and Fig. 4A). There are two important features that distinguishthe transporters directing leaderless bacteriocin secretion fromtheir counterparts directing leader-containing bacteriocin se-cretion. First, the former proteins, including EntqB, lack thetypical N-terminal proteolytic domain that is found in andrequired by the latter proteins to process and direct transportof leader-containing bacteriocins (30). Second, in the transportof leader-containing bacteriocins, a so-called accessory proteinfunctions together with the cognate ABC transporter to medi-ate bacteriocin secretion, and its gene is commonly located justdownstream of the ABC transporter gene (17, 41); such a geneis not required for transporters directing leaderless bacteriocinsecretion. Based on these findings, EntqB is therefore believedto be involved in transport of the leaderless bacteriocin EntQ.This notion is further supported by the coexpression of thebacteriocin structural gene (entqA) with entqB, which allowsthe bacterium to secrete active EntQ into the growth medium(Fig. 5A).
The immunity gene of most nonlantibiotics is located imme-diately downstream of, and in the same operon as, the bacte-riocin structural gene (17, 41, 52). However, there are someexamples of atypical locations, such as the immunity genes ofbacteriocins LsbA and LsbB, enterocin B, and carnobacterio-cin A, which are located next to the structural genes but havethe opposite orientation (25, 26, 28). With regard to this, entqAis neither followed nor preceded by a typical immunity gene.However, entqC, which is located immediately downstream ofentqB, encodes a putative 64-amino-acid protein (EntqC) withsome physical properties typical of immunity proteins (14, 16,22, 52), such as (i) a high pI value (pI 9.63), (ii) a high hydro-phobic residue content (50%), and (iii) the presence of twopredicted transmembrane segments, previously reported to benecessary for the integration of immunity proteins into themembrane of the bacteriocin producers (27). It is noteworthythat the genetic content of the EntQ gene cluster found inpCIZ2 is similar to that of the mundticin KS gene cluster inEnterococcus mundtii NFRI 7393 (36) and that of the enterocinEJ97 gene cluster in E. faecalis EJ97 (47). In these systems, thebacteriocin structural gene (munA or ej97A) is followed by anABC transporter gene (munB or ej97B) and a gene (munC orej97C) encoding a small protein (71 to 98 amino acids). WhileEJ97C is a hypothetical protein having an unknown function,MunC exhibits homology to other bacteriocin immunity pro-teins and functions as the mundticin KS immunity protein. In
the present study, a 2,207-bp fragment encompassing an intactentqC (and a truncated entqB) was shown to confer full immu-nity to EntQ, thus suggesting strongly that EntqC plays a rolein host self-protection against EntQ. However, one might ar-gue that the remaining part of the truncated forms of EntqB inpRCG04-1 and pRCG04-2 might have an immunity function.It is noteworthy that these EntqB variants (in particular, theone encoded by pRCG04-1, which is only 393 amino acidslong) are truncated from the C terminus, where importantconserved motifs required for functional ABC transporters(15, 50) are missing (Fig. 4). Nevertheless, a possible role ofEntqB in immunity can be clarified only by further investiga-tion.
The genetic determinants involved in multiple bacteriocinproduction frequently occur in clusters; each bacteriocin has itsown dedicated immunity protein, while the bacteriocins sharethe transport system (39, 41, 52). In this sense, it seems thatsecretion of the leaderless bacteriocins EntQ and EntL50 doesnot require a common ABC transporter, which was deducedfrom the following observations: (i) the loss of pCIZ2 in themutant E. faecium L50/30-2 did not affect the production ofEntL50 (Fig. 1C, left panel) and (ii) the presence of the re-combinant plasmid pRCG03 in L. sakei Lb790 was sufficientfor EntQ production and secretion (Fig. 5A). Likewise, it alsoseems that EntQ and EntL50 do not have a common immunityprotein, since (i) the loss of pCIZ2 in the mutant E. faeciumL50/30-2 did not alter the immunity to EntL50 (Fig. 1D, upperand lower panels) and (ii) the presence of recombinant plas-mid pRCG04-1 or pRCG04-2 in the host E. faecium L50/14-2conferred immunity to EntQ but not immunity to EntL50 (Fig.5B, upper and middle panels).
The biochemical, genetic, and immunological data pre-sented in this paper demonstrate that the 2,371-bp fragmentfrom the 7.4-kb pCIZ2 plasmid encompassing entqA, entqB,and entqC contains the genetic determinants required forEntQ production and immunity. Although the presence of theABC transporter EntqB alone is probably sufficient for export-ing the leaderless EntQ, the mechanism by which EntqB acts inthis process and what feature(s) on the leaderless bacteriocinpeptide serves as a signal for export are still unknown. Unrav-eling this process is therefore a challenging task that has greatscientific interest.
ACKNOWLEDGMENTS
This research was partially supported by grants 07G/0026/2000 andS-0505/AGR/0265 from the Comunidad de Madrid, Spain, and bygrants AGL2000-0706 and AGL2003-01508 from the Ministerio deEducacion, Cultura y Deporte (MECD), Spain. R.C. was the recipientof a fellowship from the MECD. J.G. holds a fellowship from theMinisterio de Ciencia y Tecnologıa, Spain.
We are indebted to Morten Skaugen and Lars Axelsson for provid-ing the expression vector pELS200, to Rosa del Campo for supplyingthe recipient strain E. faecalis JH2-2, and to Maria Victoria Francia forher help in searching for an oriT. We also thank Carmen Herranz,Marıa Victoria Francia, Morten Skaugen, and Rosa del Campo forhelpful discussions.
REFERENCES
1. Alm, R. A., L. S. L. Ling, D. T. Moir, B. L. King, E. D. Brown, P. C. Doig,D. R. Smith, B. Noonan, G. C. Guild, B. L. de Jonge, G. Carmel, P. J.Tummino, A. Caruso, M. Uria-Nickelsen, D. M. Mills, C. Ives, R. Gibson, D.Merberg, S. D. Mills, Q. Jiang, D. E. Taylor, G. F. Vovis, and T. J. Trust.1999. Genomic-sequence comparison of two unrelated isolates of the humangastric pathogen Helicobacter pylori. Nature 397:176–180.
6664 CRIADO ET AL. APPL. ENVIRON. MICROBIOL.
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

2. Aukrust, T. W., M. B. Brurberg, and I. F. Nes. 1995. Transformation oflactobacilli by electroporation. Methods Mol. Biol. 47:201–208.
3. Balla, E., L. M. T. Dicks, M. Du Toit, M. J. Van der Merwe, and W. H.Holzapfel. 2000. Characterization and cloning of the genes encodingenterocin 1071A and enterocin 1071B, two antimicrobial peptides pro-duced by Entetococcus faecalis BFE 1071. Appl. Environ. Microbiol.66:1298–1304.
4. Balla, E., and L. M. T. Dicks. 2005. Molecular analysis of the gene clusterinvolved in the production and secretion of enterocins 1071A and 1071B andof the genes responsible for the replication and transfer of plasmid pEF1071.Int. J. Food Microbiol. 99:33–45.
5. Bhowmik, T., and J. L. Steele. 1994. Cloning, characterization and inser-tional inactivaton of the Lactobacillus helveticus D(�) lactate dehydrogenasegene. Appl. Microbiol. Biotechnol. 41:432–439.
6. Bhunia, A. K., M. G. Johnson, and B. Ray. 1987. Direct detection of anantimicrobial peptide of Pediococcus acidilactici in sodium dodecyl sulphate-polyacrylamide gel electrophoresis. J. Ind. Microbiol. 2:319–322.
7. Casaus, P., T. Nilsen, L. M. Cintas, I. F. Nes, P. E. Hernandez, and H. Holo.1997. Enterocin B, a new bacteriocin from Enterococcus faecium T136 whichcan act synergistically with enterocin A. Microbiology 143:2287–2294.
8. Cintas, L. M. 1995. Ph.D. thesis. Universidad Autonoma de Madrid, Madrid,Spain.
9. Cintas, L. M., J. M. Rodrıguez, M. F. Fernandez, K. Sletten, I. F. Nes, P. E.Hernandez, and H. Holo. 1995. Isolation and characterization of pediocinL50, a new bacteriocin from Pediococcus acidilactici with a broad inhibitoryspectrum. Appl. Environ. Microbiol. 61:2643–2648.
10. Cintas, L. M., P. Casaus, L. S. Havarstein, P. E. Hernandez, and I. F. Nes.1997. Biochemical and genetic characterization of enterocin P, a novel sec-dependent bacteriocin from Enterococcus faecium P13 with a broad antimi-crobial spectrum. Appl. Environ. Microbiol. 63:4321–4330.
11. Cintas, L. M., P. Casaus, H. Holo, P. E. Hernandez, I. F. Nes, and L. S.Havarstein. 1998. Enterocins L50A and L50B, two novel bacteriocins fromEnterococcus faecium L50, are related to staphylococcal hemolysins. J. Bac-teriol. 180:1988–1994.
12. Cintas, L. M., P. Casaus, M. F. Fernandez, and P. E. Hernandez. 1998.Comparative antimicrobial activity of enterocin L50, pediocin PA-1, nisin Aand lactocin S against spoilage and foodborne pathogenic bacteria. FoodMicrobiol. 15:289–298.
13. Cintas, L. M., P. Casaus, C. Herranz, L. S. Havarstein, H. Holo, P. E.Hernandez, and I. F. Nes. 2000. Biochemical and genetic evidence thatEnterococcus faecium L50 produces enterocins L50A and L50B, the sec-dependent enterocin P, and a novel bacteriocin secreted without an N-terminal extension termed enterocin Q. J. Bacteriol. 182:6806–6814.
14. Cintas, L. M., P. Casaus, C. Herranz, I. F. Nes, and P. E. Hernandez. 2001.Bacteriocins of lactic acid bacteria. Food Sci. Technol. Int. 7:281–305.
15. Dassa, E. 2000. ABC transport, p. 1–12. In J. Lederberg (ed.), Encyclopediaof microbiology, vol. 1. Academic Press, New York, N.Y.
16. Diep, D. B., L. S. Havarstein, and I. F. Nes. 1996. Characterization of thelocus responsible for the bacteriocin production in Lactobacillus plantarumC11. J. Bacteriol. 178:4472–4483.
17. Diep, D. B., and I. F. Nes. 2002. Ribosomally synthesized antibacterialpeptides in Gram positive bacteria. Curr. Drug Targets 3:107–122.
18. Dunny, G. M., R. A. Craig, R. Carron, and D. B. Clewell. 1979. Plasmidtransfer in Streptococcus faecalis. Production of multiple sex pheromones byrecipients. Plasmid 2:454–465.
19. Eijsink, V. G. H., L. Axelsson, D. B. Diep, L. S. Havarstein, H. Holo, and I. F.Nes. 2002. Production of class II bacteriocins by lactic acid bacteria, anexample of biological warfare and communication. Antonie Leeuwenhoek81:639–645.
20. Espinosa, M., S. Cohen, M. Couturier, G. del Solar, R. Dıaz-Orejas, R.Giraldo, L. Janniere, C. Miller, M. Osborn, and C. M. Thomas. 2000.Plasmid replication and copy number control, p. 1–47. In C. M. Thomas(ed.), The horizontal gene pool: bacterial plasmids and gene spread. Har-wood Academic Publishers, London, United Kingdom.
21. Fath, M. J., and R. Kolter. 1993. ABC transporters: bacterial exporters.Microbiol. Rev. 57:995–1017.
22. Fimland, G., V. G. H. Eijsink, and J. Nissen-Meyer. 2002. Comparativestudies of immunity proteins of pediocin-like bacteriocins. Microbiology148:3661–3670.
23. Francia, M. V., and D. B. Clewell. 2002. Amplification of the tetracyclineresistance determinant of pAM�1 in Enterococcus faecalis requires asite-specific recombination event involving relaxase. J. Bacteriol. 184:5187–5193.
24. Francia, M. V., A. Varsaki, M. P. Garcillan-Barcia, A. Latorre, C. Drainas,and F. de la Cruz. 2004. A classification scheme for mobilization regions ofbacterial plasmids. FEMS Microbiol. Rev. 28:79–100.
25. Franz, C. M. A. P., R. W. Worobo, L. E. Quadri, U. Schillinger, W. H.Holzapfel, J. C. Vederas, and M. E. Stiles. 1999. Atypical genetic locusassociated with constitutive production of enterocin B by Enterococcus fae-cium BFE 900. Appl. Environ. Microbiol. 65:2170–2178.
26. Franz, C. M. A. P., M. J. van Belkum, R. W. Worobo, J. C. Vederas, andM. E. Stiles. 2000. Characterization of the genetic locus responsible for
production and immunity of carnobacteriocin A: the immunity gene conferscross-protection to enterocin B. Microbiology 146:621–631.
27. Fremaux, C., C. Ahn, and T. R. Klaenhammer. 1993. Molecular analysis ofthe lactacin F operon. Appl. Environ. Microbiol. 59:3906–3915.
28. Gajic, O., G. Buist, M. Kojic, L. Topisirovic, O. P. Kuipers, and J. Kok. 2003.Novel mechanism of bacteriocin secretion and immunity carried out bylactococcal multidrug resistance proteins. J. Biol. Chem. 278:34291–34298.
29. Gutierrez, J., R. Criado, R. Citti, M. Martın, C. Herranz, M. F. Fernandez,L. M. Cintas, and P. E. Hernandez. 2004. Performance and applications ofpolyclonal antipeptide antibodies specific for the enterococcal bacteriocinenterocin P. J. Agric. Food Chem. 52:2247–2255.
30. Havarstein, L. S., D. B. Diep, and I. F. Nes. 1995. A family of bacteriocinABC transporters that carry out proteolytic processing of their substratesconcomitant with export. Mol. Microbiol. 16:229–240.
31. Herranz, C., and A. J. M. Driessen. 2005. sec-mediated secretion of bacte-riocin enterocin P by Lactococcus lactis. Appl. Environ. Microbiol. 71:1959–1963.
32. Holo, H., and I. F. Nes. 1989. High-frequency transformation, by electropo-ration, of Lactococcus lactis subsp. cremoris grown with glycine in osmoticallystabilized media. Appl. Environ. Microbiol. 55:3119–3123.
33. Jack, R. W., J. R. Tagg, and B. Ray. 1995. Bacteriocins of gram-positivebacteria. Microbiol. Rev. 59:171–200.
34. Jacob, A. E., and S. J. Hobbs. 1974. Conjugal transfer of plasmid-bornemultiple antibiotic resistance in Streptococcus faecalis var. zymogenes. J. Bac-teriol. 117:360–372.
35. Jaworski, D., and D. B. Clewell. 1995. A functional origin of transfer (oriT)on the conjugative transposon Tn916. J. Bacteriol. 177:6644–6651.
36. Kawamoto, S., J. Shima, R. Sato, T. Eguchi, S. Ohmomo, J. Shibato, N.Horikoshi, K. Takeshita, and T. Sameshima. 2002. Biochemical andgenetic characterization of mundticin KS, an antilisterial peptide pro-duced by Enterococcus mundtii NFRI 7393. Appl. Environ. Microbiol.68:3830–3840.
37. Mathiesen, G., K. Kuehne, L. Kroeckel, L. Axelsson, and V. G. Eijsink. 2005.Characterization of a new bacteriocin operon in sakacin P-producing Lacto-bacillus sakei, showing strong translational coupling between the bacteriocinand immunity genes. Appl. Environ. Microbiol. 71:3565–3574.
38. Mundt, J. O. 1986. Genus Enterococcus, p. 1063–1065. In P. H. A. Sneath,N. S. Mair, M. E. Sharpe, and J. G. Holt (ed.), Bergey’s manual of systematicbacteriology, vol. 2. Williams and Wilkins Co., Baltimore, Md.
39. Nes, I. F., D. B. Diep, L. S. Havarstein, and M. B. Brurberg. 1996. Biosyn-thesis of bacteriocins in lactic acid bacteria. Antonie Leeuwenhoek 70:113–128.
40. Nes, I. F., and H. Holo. 2000. Class II antimicrobial peptides from lactic acidbacteria. Biopolym. Pept. Sci. 55:50–61.
41. Nes, I. F., H. Holo, G. Fimland, H. H. Hauge, and J. Nissen-Meyer. 2002.Unmodified peptide-bacteriocins (class II) produced by lactic acid bacteria,p. 81–115. In C. J. Dutton, M. A. Haxell, H. A. I. McArthur, and R. G. Wax(ed.), Peptide antibiotics: discovery, modes of action and application. MarcelDekker, New York, N.Y.
42. Netz, D. J. A., H. G. Sahl, R. Marcolino, J. Dos Santos Nascimiento, S.Soares de Oliveira, M. Bento Soares, and M. C. F. Bastos. 2001. Molecularcharacterisation of aureocin A70, a multipeptide bacteriocin isolated fromStaphylococcus aureus. J. Mol. Biol. 311:939–949.
43. Netz, D. J. A., R. Pohl, A. G. Beck-Sickinger, T. Selmer, A. J. Pierik, M. C. F.Bastos, and H. G. Sahl. 2002. Biochemical characterisation and geneticanalysis of aureocin A53, a new, atypical bacteriocin from Staphylococcusaureus. J. Mol. Biol. 319:745–756.
44. Pansegrau, W., W. Schroder, and E. Lanka. 1994. Concerted action of threedistinct domains in the DNA cleaving-joining reaction catalyzed by relaxase(TraI) of conjugative plasmid RP4. J. Biol. Chem. 269:2782–2789.
45. Quadri, L. E. N., M. Sailer, K. L. Roy, J. C. Vederas, and M. E. Stiles. 1994.Chemical and genetic characterization of bacteriocins produced by Carno-bacterium piscicola LV17B. J. Biol. Chem. 269:12204–12211.
46. Sambrook, J., and D. Russell. 2001. Molecular cloning: a laboratory manual,3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
47. Sanchez-Hidalgo, M., M. Maqueda, A. Galvez, H. Abriouel, E. Valdivia, andM. Martınez-Bueno. 2003. The genes coding for enterocin EJ97 productionby Enterococcus faecalis EJ97 are located on a conjugative plasmid. Appl.Environ. Microbiol. 69:1633–1641.
48. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequencing with chainterminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463–5467.
49. Schillinger, U., and F. K. Lucke. 1989. Antibacterial activity of Lactobacillussake isolated from meat. Appl. Environ. Microbiol. 55:1901–1906.
50. Schneider, E., and S. Hunke. 1998. ATP-binding-cassette (ABC) transportsystems: functional and structural aspects of the ATP-hydrolysing subunits/domains. FEMS Microbiol. Rev. 22:1–20.
51. Shagger, H., and G. von Jagow. 1987. Tricine-sodium dodecyl sulphate-polyacrylamide gel electrophoresis for the separation of proteins in the range1 to 100 kDa. Anal. Biochem. 166:368–379.
52. Skaugen, M., L. M. Cintas, and I. F. Nes. 2003. Genetics of bacteriocinproduction in lactic acid bacteria, p. 225–260. In B. J. B. Wood and P. J.
VOL. 72, 2006 pCIZ2-ENCODED ENTEROCIN Q PRODUCTION AND IMMUNITY 6665
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

Warner (ed.), The lactic acid bacteria, vol. 3. Genetics of lactic acid bacteria.Kluwer Academic/Plenum Publisher, New York, N.Y.
53. Smith, M. C. A, and C. D. Thomas. 2004. An accessory protein is required forrelaxosome formation by small staphylococcal plasmids. J. Bacteriol. 186:3363–3373.
54. Sorvig, E., M. Skaugen, K. Naterstad, V. G. Eijsink, and L. Axelsson. 2005.Plasmid p256 from Lactobacillus plantarum represents a new type of repliconin lactic acid bacteria, and contains a toxin-antitoxin-like plasmid mainte-nance system. Microbiology 151:421–431.
55. Trotter, M., R. P. Ross, G. F. Fitzgerald, and A. Coffey. 2002. Lactococcuslactis DPC5598, a plasmid-free derivative of a commercial starter, provides avaluable alternative host for culture improvement studies. J. Appl. Micro-biol. 93:134–143.
56. van de Guchte, M., J. M. van der Vossen, J. Kok, and G. Venema. 1989.Construction of a lactococcal expression vector: expression of hen egg whitelysozyme in Lactococcus lactis subsp. lactis. Appl. Environ. Microbiol. 55:224–228.
57. Worobo, R. W., T. Henkel, M. Sailer, K. L. Roy, J. C. Vederas, and M. E.Stiles. 1994. Characteristics and genetic determinants of a hydrophobic pep-tide bacteriocin, carnobacteriocin A, produced by Carnobacterium piscicolaLV17A. Microbiology 140:517–526.
58. Zechner, E. L., F. de la Cruz, R. Eisenbrandt, A. M. Grahn, G. Koraimann,E. Lanka, G. Muth, W. Pansegrau, C. M. Thomas, B. M. Wilkins, and M.Zaryka. 2000. Conjugative-DNA transfer processes, p. 87–174. In C. M.Thomas (ed.), The horizontal gene pool: bacterial plasmids and gene spread.Harwood Academic Publishers, London, United Kingdom.
6666 CRIADO ET AL. APPL. ENVIRON. MICROBIOL.
on January 3, 2014 by guesthttp://aem
.asm.org/
Dow
nloaded from

![=^f EPiTSa^_b³Tgc^acX^]´ Q^\Q^]Tg 7< BT]P](https://static.fdokumen.com/doc/165x107/631b1814c51d6b41aa0517ad/f-epitsabtgcacx-qqtg-7-btp.jpg)










![\ ] q § n ' § b](https://static.fdokumen.com/doc/165x107/633204227f0d9c38da013cb8/-q-n-b.jpg)







