
Contribution of C. beijerinckii and C. sporogenes in association with C. tyrobutyricum to the...
10
Contribution of C. beijerinckii and C. sporogenes in association with C. tyrobutyricum to the butyric fermentation in Emmental type cheese Anne-Gaëlle Le Bourhis a , Joël Doré b , Jean-Philippe Carlier c , Jean-François Chamba a, ⁎ , Michel-Robert Popoff c , Jean-Luc Tholozan d a Institut Technique Français des Fromages, 74801 La Roche-sur-Foron Cedex, France b Institut National de la Recherche Agronomique, Unité d'Ecologie et de Physiologie du Système Digestif, Domaine de Vilvert, 78352 Jouy-en-Josas Cedex, France c Institut Pasteur, Centre National de Référence des Bactéries Anaérobies et du Botulisme, 75724 Paris Cedex 15, France d Université de Provence, Laboratoire de Microbiologie, Institut de recherche pour le Développement, 13288 Marseille Cedex 9, France Received 27 June 2005; received in revised form 8 February 2006; accepted 6 June 2006 Abstract The relationship between C. tyrobutyricum, C. sporogenes and C. beijerinckii in experimental cheese conditions, and their influences on late- blowing and butyric fermentation, have been investigated. A molecular approach using a PCR-TTGE method in combination with conventional methods, such as microbiological and physico-chemical analysis, was performed to monitor the evolution of these clostridial species, simultaneously with the occurrence of cheese defects. Sixteen Emmental type cheeses were produced from milk inoculated with different clostridial spore associations. In all cheeses inoculated with C. tyrobutyricum, obvious signs of late blowing were detected. In cheeses inoculated with C. beijerinckii or C. sporogenes, a formation of holes in cheese body was observed, with a concomitant slight amount of butyric acid production. Even though C. beijerinckii and C. sporogenes were less metabolically active and less numerically important than C. tyrobutyricum in cheese as shown by TTGE profiles, the association of these species to C. tyrobutyricum enhanced the butyric fermentation and the cheese defects. The level of butyric content in ripened cheese increased to 268 mg 100 g - 1 in presence of C. tyrobutyricum, and reached a maximum of 414 mg 100 g - 1 in presence of the C. beijerinckii–C. tyrobutyricum (1:10) association. The propionic fermentation was also higher in cheese inoculated with C. tyrobutyricum, and was slowed down in presence of C. beijerinckii and C. sporogenes. From 30 days of ripening, a strong correlation between the chemical contents and the intensity of cheese defects was demonstrated. A chemical analysis of cheese associated with a molecular method for microbial spoilage investigation allows the prediction of the level of late blowing at early stages of ripening, and the understanding of the origin of the defect. © 2006 Published by Elsevier B.V. Keywords: Clostridium association; Butyric fermentation; Emmental type cheese 1. Introduction The defects caused by the growth of spore-forming bacteria during the ripening process of hard-cooked-cheeses such as Emmental, Comté, Beaufort and similar semi-hard cheeses, result in economic loss for the cheese industry. These spor- ulating bacteria contaminate milk mainly via silage and induce a defect called late blowing during the ripening of cheese (Das Gupta and Hull, 1989). Growth of these organisms is controlled to some extent by the pH, moisture content and by the presence of other bacteria in cheese (Bergère and Hermier, 1970). Owing to its physico-chemical characteristics and its ripening temper- ature, Emmental type cheese is highly susceptible to late blowing. Clostridia able to grow in cheese cause a subsequent spoilage either through gas formation by lactate-fermenting Clostridia such as C. beijerinckii and C. tyrobutyricum, or additional proteolysis in Clostridia such as C. sporogenes (Goudkov and Sharpe, 1965). C. tyrobutyricum is usually con- sidered to be the primary cause of late blowing, C. sporogenes, International Journal of Food Microbiology 113 (2007) 154 – 163 www.elsevier.com/locate/ijfoodmicro ⁎ Corresponding author. Mailing address: 419 route des champs laitiers, BP 30, 74801 La Roche-sur-Foron Cedex, France. Tel.: +33 4 50 03 33 03; fax: +33 4 50 25 82 26. E-mail address: [email protected] (J.-F. Chamba). 0168-1605/$ - see front matter © 2006 Published by Elsevier B.V. doi:10.1016/j.ijfoodmicro.2006.06.027
Transcript of Contribution of C. beijerinckii and C. sporogenes in association with C. tyrobutyricum to the...

biology 113 (2007) 154–163www.elsevier.com/locate/ijfoodmicro
International Journal of Food Micro
Contribution of C. beijerinckii and C. sporogenes in association withC. tyrobutyricum to the butyric fermentation in Emmental type cheese
Anne-Gaëlle Le Bourhis a, Joël Doré b, Jean-Philippe Carlier c, Jean-François Chamba a,⁎,Michel-Robert Popoff c, Jean-Luc Tholozan d
a Institut Technique Français des Fromages, 74801 La Roche-sur-Foron Cedex, Franceb Institut National de la Recherche Agronomique, Unité d'Ecologie et de Physiologie du Système Digestif, Domaine de Vilvert, 78352 Jouy-en-Josas Cedex, France
c Institut Pasteur, Centre National de Référence des Bactéries Anaérobies et du Botulisme, 75724 Paris Cedex 15, Franced Université de Provence, Laboratoire de Microbiologie, Institut de recherche pour le Développement, 13288 Marseille Cedex 9, France
Received 27 June 2005; received in revised form 8 February 2006; accepted 6 June 2006
Abstract
The relationship between C. tyrobutyricum, C. sporogenes and C. beijerinckii in experimental cheese conditions, and their influences on late-blowing and butyric fermentation, have been investigated. A molecular approach using a PCR-TTGE method in combination with conventionalmethods, such as microbiological and physico-chemical analysis, was performed to monitor the evolution of these clostridial species,simultaneously with the occurrence of cheese defects. Sixteen Emmental type cheeses were produced from milk inoculated with differentclostridial spore associations. In all cheeses inoculated with C. tyrobutyricum, obvious signs of late blowing were detected. In cheeses inoculatedwith C. beijerinckii or C. sporogenes, a formation of holes in cheese body was observed, with a concomitant slight amount of butyric acidproduction. Even though C. beijerinckii and C. sporogenes were less metabolically active and less numerically important than C. tyrobutyricum incheese as shown by TTGE profiles, the association of these species to C. tyrobutyricum enhanced the butyric fermentation and the cheese defects.The level of butyric content in ripened cheese increased to 268 mg 100 g−1 in presence of C. tyrobutyricum, and reached a maximum of414 mg 100 g−1 in presence of the C. beijerinckii–C. tyrobutyricum (1:10) association. The propionic fermentation was also higher in cheeseinoculated with C. tyrobutyricum, and was slowed down in presence of C. beijerinckii and C. sporogenes. From 30 days of ripening, a strongcorrelation between the chemical contents and the intensity of cheese defects was demonstrated. A chemical analysis of cheese associated with amolecular method for microbial spoilage investigation allows the prediction of the level of late blowing at early stages of ripening, and theunderstanding of the origin of the defect.© 2006 Published by Elsevier B.V.
Keywords: Clostridium association; Butyric fermentation; Emmental type cheese
1. Introduction
The defects caused by the growth of spore-forming bacteriaduring the ripening process of hard-cooked-cheeses such asEmmental, Comté, Beaufort and similar semi-hard cheeses,result in economic loss for the cheese industry. These spor-ulating bacteria contaminate milk mainly via silage and induce a
⁎ Corresponding author. Mailing address: 419 route des champs laitiers, BP30, 74801 La Roche-sur-Foron Cedex, France. Tel.: +33 4 50 03 33 03; fax: +334 50 25 82 26.
E-mail address: [email protected] (J.-F. Chamba).
0168-1605/$ - see front matter © 2006 Published by Elsevier B.V.doi:10.1016/j.ijfoodmicro.2006.06.027
defect called late blowing during the ripening of cheese (DasGupta and Hull, 1989). Growth of these organisms is controlledto some extent by the pH, moisture content and by the presenceof other bacteria in cheese (Bergère and Hermier, 1970). Owingto its physico-chemical characteristics and its ripening temper-ature, Emmental type cheese is highly susceptible to lateblowing. Clostridia able to grow in cheese cause a subsequentspoilage either through gas formation by lactate-fermentingClostridia such as C. beijerinckii and C. tyrobutyricum, oradditional proteolysis in Clostridia such as C. sporogenes(Goudkov and Sharpe, 1965). C. tyrobutyricum is usually con-sidered to be the primary cause of late blowing, C. sporogenes,

Table 1Experimental combinations used for the addition of clostridial suspensions incheese milk, and cheese identification
Bacterialcontaminationof cheese milka
Ratio between theclostridial species
Cheese no.
Essay no. 1 Essay no. 2
Control / T /Ctb / A1-1 /Cs / C1-1 /Cb / D1-1 /Ct–Cs 10:1 A2-1 A2-2
Ct–Cs 1:1 A3-1 A3-2
Ct–Cs 1:10 C2-1 C2-2
Ct–Cb 10:1 B1-1 B1-2
Ct–Cb 1:1 B2-1 B2-2
Ct–Cb 1:10 E1-1 E1-2
Cs–Cb 1:1 D3-1 /a Final concentration, 4 ·103 spores per l of milk.b Cb, C. beijerinckii (CIP 104308); Cs, C. sporogenes (ITFF 35CL13) and Ct,
C. tyrobutyricum (CNRZ 608). CIP; Collection de l'Institut Pasteur; CNRZ,Centre National de la Recherche Zootechnique; ITFF, Institut TechniqueFrançais des Fromages.
155A.-G. Le Bourhis et al. / International Journal of Food Microbiology 113 (2007) 154–163
C. beijerinckii, and C. butyricum are also significant secondarycauses of gas production in cheese (Hood and Bowen, 1950;Bergère et al., 1968; Matteuzzi et al., 1972; Toyoda et al., 1990;Klijn et al., 1995). In a recent work, C. tyrobutyricum and C.beijerinckii were reported to be the main organisms present indefectuous commercial cheeses (Le Bourhis et al., 2005). Thisstudy pointed out the ability of C. beijerinckii to grow in cheeseconditions and suggested the implication of this species in splitdefects. In order to confirm this finding and to investigate therelationship between C. tyrobutyricum, C. beijerinckii and C.sporogenes this work has been continued. The main purpose ofthis present study was to obtain fundamental information aboutthe influence of different types of clostridial associations on lateblowing under conditions occurring in Emmental cheese. Inorder to follow the evolution of the clostridial species simul-taneously with the occurrence of cheese defects during theripening, a molecular approach using PCR-TTGE in combina-tion with conventional methods implicating microbiologicaland physico-chemical analysis was performed.
2. Materials and methods
2.1. Production of spores
Spores from C. tyrobutyricum (CNRZ 608), C. beijerinckii(CIP 104308) and C. sporogenes (ITFF 35CL13) represented asCt, Cb, and Cs, respectively, were produced by growing culturesfor 15 days at 30 °C under anaerobic conditions in BBmB brothat pH 5.3 (Bergère's modification of the lactate medium ofBryant and Burkey). The spores were concentrated by centri-fugation at 12,000 g for 30 min, washed two times in a sodiumcitrate solution (trisodium citrate dihydrate 2% (wt/v)), thensuspended in 10 ml of sterile H2O, and stored at 4 °C (Cerf et al.,1967). The spore concentration was determined after pasteur-ization at 75 °C for 10 min of serial dilutions and spreading onRCM medium, and CFU were counted from an incubation timeof 3 days until 7 days at 37 °C under anaerobic conditions.
2.2. Cheese manufacture
Seventeen cheeses were made in 3 vats containing 120 l ofmicrofiltered and thermized milk (63 °C, 30 s). Beforemicrofiltration, the fat content of skimmed milk was standard-ized to 3% fat content by addition of skim (Chamba andPerreard, 2002). The milk was inoculated with a starter cultureconsisting of Streptococcus thermophilus starter PAL 82–87and Lactobacillus delbrueckki subsp. lactis; Lactobacillushelveticus starter PAL LH56-LL57 (Standa Industrie, Caen,France) added at levels of 1.4‰ and 0.5‰, respectively. Sporesuspensions were then added to the milk as described in Table. 1,with no addition for cheese control. The experimental cheeseswere made using a typical hard-cooked-cheese making schemeaccording to the procedures described previously (Chamba andPerreard, 2002). Cheeses were ripened in a cold room for 10 daysat 12 °C, followed by incubation for 50 days at 23 °C in a warmroom. This is an adaptation of the usual process where theripening in the cold room is followed by 15 days in temperate
temperature room at 15 °C, and then in a warm room at 23 °C.Samples of cheese were aseptically taken every fifteen days ofripening time for physico-chemical, microbiological andmolecular analysis as described below.
2.3. Microbiological analysis
Ten grams of cheese sample was homogenized in 90 ml ofsodium citrate solution with a stomacher. The samples wereheat-shocked at 75 °C for 10 min. Using 0.1% tryptone solutionas diluent, the multiple dilution procedure was used for deter-mination of spore concentration by plating method on RCMmedium as described above. Enumerations were performed induplicate, and average counts of colony forming units werereported.
2.4. Physico-chemical analysis
Samples of cheese were treated or stored at 4 °C untilprocessing. Acidification, moisture, fat, dry matter and lactatecontents were determined as previously described by Boutonand Grappin (1994). Butyric acid fermentation in cheeses wasdetected by gas–liquid chromatography (GLC) analysis ofvolatile fatty acids according to the method previously used(Berdagué, 1986; Le Bourhis et al., 2005). The gas productionwas monitored during ripening by visual inspections of cheese.
2.5. DNA and RNA extraction from cheese
Aliquots of 250 mg were stored in 2-ml screw-cap tubes at−80 °C until further use. Total DNAwas extracted according tothe method previously described (Godon et al., 1997). Thesamples were treated by bead-beating, in order to mechanicallydisrupt spores, by guanidine thiocyanate and N-lauroyl-sarcosinefor DNA extraction, and by ethanol for DNA precipitation. RNAisolation was carried out by phenol–chloroform extraction

Table 2Evolution of the spore concentration (spores g−1) in cheese during the ripening
Cheeseidentifications
Day
1 15 30 45 60
Min Max Min Max Min Max Min Max Min Max
Control NDa ND ND ND NDCtb 1.5 ·101 7.8 ·102 4.7 ·105 7.5 ·105 1.3 ·106
Cs 1.5 ·101 2.5 ·101 3.0 ·101 1.5 ·102 8.0 ·103
Cb 1.5 ·101 3.5 ·101 1.1 ·102 8.5 ·102 1.0 ·104
Ct–Cs (10:1) ND ND ND 1.0 ·102 2.0 ·105 3.1 ·105 1.1 ·106 1.1 ·106 1.1 ·106 1.2 ·106
Ct–Cs (1:1) 1.5 ·101 2.5 ·101 1.3 ·104 2.0 ·104 3.1 ·105 8.8 ·105 1.5 ·106 4.0 ·106 3.6 ·106 4.4 ·106
Ct–Cs (1:10) 1.0 ·101 1.5 ·101 6.9 ·101 2.5 ·102 2.7 ·104 7.3 ·104 1.9 ·104 3.3 ·105 6.1 ·104 5.0 ·105
Ct–Cb (10:1) 5.0 ·100 1.0 ·101 9.0 ·101 1.8 ·102 2.9 ·105 4.1 ·105 5.4 ·105 2.5 ·106 1.6 ·106 3.4 ·106
Ct–Cb (1:1) ND 2.0 ·101 5.5 ·101 4.4 ·104 3.4 ·104 1.3 ·105 1.8 ·106 2.7 ·106 3.7 ·106 4.8 ·106
Ct–Cb (1:10) 5.0 ·100 1.0 ·101 2.5 ·101 1.2 ·102 2.0 ·102 2.4 ·103 1.4 ·104 3.3 ·104 1.6 ·105 7.0 ·105
Cs–Cb (1:1) ND 8.0 ·101 1.1 ·103 4.0 ·103 6.0 ·103
Data reported are minimum and maximum values determined in experimental cheeses.Cheeses were ripened in cold room for 10 days at 12 °C, and then in a warm room for 50 days at 23 °C. The spore concentrations were determined every fifteen days byplating method on RCM medium.a Not detected.b Cb, C. beijerinckii (CIP 104308); Cs, C. sporogenes (ITFF 35CL13); and Ct, C. tyrobutyricum (CNRZ 608).
156 A.-G. Le Bourhis et al. / International Journal of Food Microbiology 113 (2007) 154–163
following upon bead-beating treatment according to the methodpreviously described (Doré et al., 1998). After ethanol precipi-tation, nucleic acids were resuspended in 100 μl of 10 mM Tris–HCl (pH 8.0). The concentration and integrity of the nucleic acidswere visually determined by electrophoresis on 1% agarose gelcontaining ethidium bromide.
2.6. RT-PCR and PCR amplification
TTGE samples were prepared by performing two successivePCRs (nested-PCR) as previously described (Le Bourhis et al.,2005). Primers S-*-Cbot-0691-a-S-23 (5′-GTG AAA TGCGTA GAG ATT AGG AA-3′) (P930), specific of cluster I, andS-*-Clit-1338-a-A-22 (5′-GAT YYG CGA TTA CTA GYAACT C-3′) (P932) were used to amplify the V5–V8 region ofthe 16S rRNA gene of the phylogenetic cluster I of the genusClostridium (Collins et al., 1994; Le Bourhis et al., 2005).
RT-PCR was performed with the rTth DNA polymerase W/Buffers Kit (Applied-Biosystems, Courtaboeuf, France). Re-verse transcriptase reaction templates of 20 μl contained 10 mMTris–HCl (pH 8.3), 90 mM KCl, 1 mM MnCl2, 200 mM eachdNTP, 5 U rTth DNA polymerase, 0.5 μM of primer P932, and2 μl of RNA. The mixtures were incubated at 70 °C for 15 min.After this incubation, 80 μl of the PCR additive was added. Theadditive consisted of 4% glycerol, 8 mM Tris–HCl (pH 8.3),80mMKCl, 0.04 Tween 20, 0.6 mMEGTA, 1.5 mMMgCl2 and0.5 μM of primer P930. The samples were subjected to theamplification in a Mastercycler (Eppendorf, Hamburg, Ger-many) by using the following program: 2min at 95 °C; 40 cyclesof 94 °C for 30 s, 58 °C for 30 s, and 72 °C for 45 s; and finally,72 °C for 5 min. To check their size (665 pb) and estimate theirconcentration, PCR products were analysed by electrophoresison a 1% agarose gel containing ethidium bromide. Negativecontrol without DNAwas run in each amplification.
PCR reactions were carried out in 25 μl of the followingbuffer: 12 μl of premix (0.5 μM of forward and reverse primer,
DMSO 4% final concentration), 12 μl of HotStar Taq MasterMix (Qiagen, Courtaboeuf, France), and 1 μl of genomic DNA.Amplification and analysis by agarose gel electrophoresis wereperformed as described above, with the following specific PCRprogram: 15 min at 95 °C to allow HotStar Taq activationand 40 cycles of 94 °C for 20 s, 58 °C for 30 s, and 72 °C for30 s.
2.7. Nested-PCR
Serial dilutions of PCR products of DNA and cDNA weretested to determine the optimal DNA concentrations for nested-PCR which amplified 290 bp of the V5–V6 region of the 16SrRNA gene. From these dilutions 1 μl was used as template. ThePCR was performed with primers S-D-Bact-1060-a-A-21 (5′-TCA CGA CAC GAG CTG ACG ACA-3′) and S-D-Bact-0786-a-S-20 (5′-CGC CCG CCG CGC CCC GCG CCC GGCCCG CCG CCC CCG CCC C GAT TAG ATA CCC TGG TAGTC-3′) following the complete procedure described above, butrunning 25 cycles, and with annealing temperature set at 64 °C(Woese, 1987). Amplification and analysis by agarose gelelectrophoresis were performed as described above.
2.8. TTGE analysis
DCode™ Universal Mutation Detection System (Biorad)was used for sequence specific separation of PCR and RT-PCRproducts according to the procedures described previously withthe following modifications (Seksik et al., 2003). Theelectrophoresis was performed in a 8% (w/v) polyacrylamide–bisacrylamide gel, and 7 M urea with 1.25× TAE buffer (Tris-acetic acid-EDTA; Biorad Laboratories), at a constant voltage of70 V during 16 h. A temperature gradient from 63 °C to 70 °Cand a temperature increase rate of 0.4 °C/h were applied. Five-microliter samples of PCR products were loaded in wells. Afterelectrophoresis, the gels were stained for 30 min with SYBR

Table 3Mean composition of experimental cheeses
Day Dry matter (%) F/DM (%)a M/NFS (%) S/M (%) pH
1 62.5±0.6 48.2±1.4 1.16±0.03 NDb 5.49±0.0660 65.4±0.9 48.1±1.3 1.07±0.23 1.3±0.1 5.90±0.16
Data reported are means±standard deviations of the 17 cheeses.a F/DM: fat in dry matter; M/NFS: moisture in the non-fat solid; S/M: salt in
moisture.b Not determined.
Fig. 1. TTGE profiles of amplicons generated by PCR and RT-PCR of the V5–V6 regions of 16S rRNA genes for the identification of Clostridial species present incheese during the ripening period. Cheese inoculated with Ct–Cs (1:1), Ct–Cb (1:1) and Cs–Cb (1:1) associations. ID, identification ladder. Numbers correspond tothe nucleic acids extracted after 15, 30, 45 and 60 days of ripening. Abbreviations: Cb, C. beijerinckii (CIP 104308); Cs, C. sporogenes (ITFF 35CL13); and Ct, C.tyrobutyricum (CNRZ 608).
157A.-G. Le Bourhis et al. / International Journal of Food Microbiology 113 (2007) 154–163
Green I Nucleic Acid Gel Stain (Roche Diagnostics, Manheim,Germany), and their image digitized using a Storm system(Molecular Dynamics, Sunnyvale, CA, USA).
2.9. Statistical analysis
Statistical analysis was performed using Statbox version 6.Pearson correlation coefficients were obtained to investigatepotential linear correlations between the evolution of pH, lactateand fatty acid contents during the ripening of cheese. PrincipalComponent Analysis (PCA) was used in order to study theinfluence of the different clostridial strains and their associa-tions on the characteristics of the experimental cheeses.
3. Results
3.1. Evolution of spore concentrations during the ripening ofcheese
The clostridial strains combinations used in cheese makingexperiments are described in Table 1. Microfiltered milk wascharacterized by a good microbial quality (<4.0 ·102 spores l−1
and 4.0 ·103 CFU l−1). The ability of Clostridia to grow inhard-cooked-cheese was assessed by the enumeration of spores.Table 2 shows the evolution of spore concentration throughoutthe ripening period. At early stages of ripening, from day 1 today 15, notable differences in spore population were observedbetween cheeses: spores detected in cheese inoculated with C.tyrobutyricum (Ct) after 15 days of ripening was approximately26 times higher than mean value of spore counts in cheeseinoculated with C. beijerinckii or C. sporogenes. An increase ofmore than 2 log units between day 15 and day 30 was observedfor the cheese Ct. In cheeses inoculated with C. beijerinckii andwith C. sporogenes, the development of clostridial spore beganslightly at 30 days of ripening, and increased progressively up tothe end of the ripening period. At the end of ripening, growth ofClostridia was particularly strong in cheese Ct, with sporecounts reaching 1.3 ·106 spores g−1 while cheese inoculated
with C. beijerinckii and C. sporogenes, reached 1.0 ·104
spores g−1 and 8.0 ·103 spores g−1, respectively. No clostridialspores were detected in control cheese up to 60 days of ripening.
Cheese inoculated with Ct–Cb mix and with Ct–Cs mixsignificantly differed in their spore contents from cheeseinoculated with a single clostridial species. At the end of ripeningperiod, cheese inoculated with Ct–Cb mix or Ct–Cs mix (1:1)displayed a three times higher level of spores than cheese inocu-lated with C. tyrobutyricum alone. Spore counts in cheeseinoculated with Cb–Cs mix (1:1), increased slowly duringripening, and reached 6.0 ·103 spores g−1 in the ripened cheese.The amount of spores in cheese inoculatedwithCs–Ct andCb–Ctmix 1:1 each, were approximately 2 times higher than in cheeseinoculatedwith a ratio 1:10 between these species. Spore numberswere significantly lower when large amounts ofC. sporogenes orC. beijerinckii were added to C. tyrobutyricum.
3.2. Application of PCR-TTGE to monitor changes ofclostridial population
Predominance of each clostridial species was estimated onthe basis of TTGE fingerprints of cheese, in parallel to countsperformed by the classical plating method. Preliminary worksdemonstrated that DNA and RNA extract from cheesefollowing the method proposed by Godon et al. and Doré etal. was successfully used as a template in PCR (Godon et al.,1997; Doré et al., 1998). Since the metabolically active cellscontain more rRNA than inactive or dormant cells, the profiles

Table 4pH values measured at 1, 15, 30, 45 and 60 days of the ripening time, and content of lactate in 60 days old cheeses
Cheeseidentifications
pH values Lactate (mg 100g−1)
Day
1 15 30 45 60 60
Min Max Min Max Min Max Min Max Min Max Min Max
Control 5.55 5.63 5.46 5.61 5.62 1084.2Cta 5.63 5.56 5.56 5.91 6.14 51.5Cs 5.51 5.63 5.41 5.64 5.82 1023.2Cb 5.54 5.67 5.42 5.60 5.82 945.7Ct–Cs (10:1) 5.42 5.45 5.58 5.67 5.53 5.55 5.77 5.80 6.09 6.13 64.3 106.2Ct–Cs (1:1) 5.45 5.52 5.42 5.43 5.51 5.57 5.82 5.90 5.81 5.83 61.2 70.7Ct–Cs (1:10) 5.42 5.47 5.31 5.38 5.50 5.51 5.52 5.60 5.74 5.84 676.1 837.1Ct–Cb (10:1) 5.44 5.58 5.57 5.63 5.52 5.61 5.69 5.84 5.98 6.11 51.5 73.9Ct–Cb (1:1) 5.43 5.43 5.39 5.59 5.53 5.55 5.78 5.79 5.96 6.11 22.4 70.8Ct–Cb (1:10) 5.48 5.53 5.42 5.42 5.45 5.48 5.60 5.66 5.70 5.77 637.5 808.0Cs–Cb (1:1) 5.43 5.54 5.54 5.63 5.76 1172.1
Data reported are minimum and maximum values determined in experimental cheeses.a Cb, C. beijerinckii (CIP 104308); Cs, C. sporogenes (ITFF 35CL13) and Ct, C. tyrobutyricum (CNRZ 608).
158 A.-G. Le Bourhis et al. / International Journal of Food Microbiology 113 (2007) 154–163
of cDNA conceptually correspond to the metabolically activepopulation (Rosset et al., 1966; Wagner, 1994). A too low levelof RNA from Clostridia did not allow an amplification with theCluster I-specific primer at day 15. With an inoculation withCt–Cs mix, C. tyrobutyricum was consistently predominantin cheese from 15 days to 60 days of ripening. Banding pat-terns for the V5–V6 rDNA amplicons corresponding to Ct–Cs,Ct–Cb and Cs–Cb (1:1) associations are presented in Fig. 1. Inaddition, with a Ct–Cb mix (1:1) inoculation, C. beijerinckiiwas detected at 15 days of ripening by PCR-TTGE, and was stilldetected at 30 days in cheese inoculated with C. beijerinckii 10times higher than C. tyrobutyricum. PCR-TTGE profiles ofcheeses containing a Cb–Cs mix (1:1) showed a predominance
Fig. 2. Cross section of cheese after 60 days of ripening. Control cheese (A), Cheese iCs (1:1) mix (E), Ct–Cb (1:1) mix (F) and Cs–Cb (1:1) mix (G). Abbreviations: Ctyrobutyricum (CNRZ 608).
of C. beijerinckii until 15 days of ripening, then C. sporogenesbecame predominant. After 30 days of ripening, cDNA profilesshowed the presence of only one species, C. sporogenes,underlining its higher metabolic activity than C. beijerinckii.
3.3. Physico-chemical composition
Values of the chemical composition of experimental cheeseswere not significantly different between cheeses at one day and60 days of ripening (Table. 3). These characteristics are usual forFrench Emmental cheese except for the pH value at one day ofripening which was higher: 5.49±0.06 instead of 5.2 (Accolaset al., 1980). Control cheese showed a slight decrease of pH until
noculated with C. tyrobutyricum (B), C. sporogenes (C), C. beijerinckii (D), Ct–b, C. beijerinckii (CIP 104308); Cs, C. sporogenes (ITFF 35CL13); and Ct, C.
159A.-G. Le Bourhis et al. / International Journal of Food Microbiology 113 (2007) 154–163
day 30 and increasing thereafter, as reported in Table. 4. Its valuethen remained rather constant at the end of ripening. In allcheeses inoculated with Clostridia, the pH value increased afterday 30, reaching a maximum of 6.14 at the end of ripeningfor the cheese inoculated with C. tyrobutyricum. Compared toC. tyrobutyricum alone, deacidification rate was lower in cheeseinoculated with C. sporogenes and C. beijerinckii. In addition,cheeses inoculated with C. beijerinckii and with C. sporogenespresented a similar pH evolution throughout the ripening period,and reached a pH final value of 5.82.
Regression analysis confirmed the negative correlationbetween the increase of pH and the decrease of lactate contentat 60 days of ripening time (P<0.001; r=−0.78). At the endof ripening, the lactate content of cheese control reached
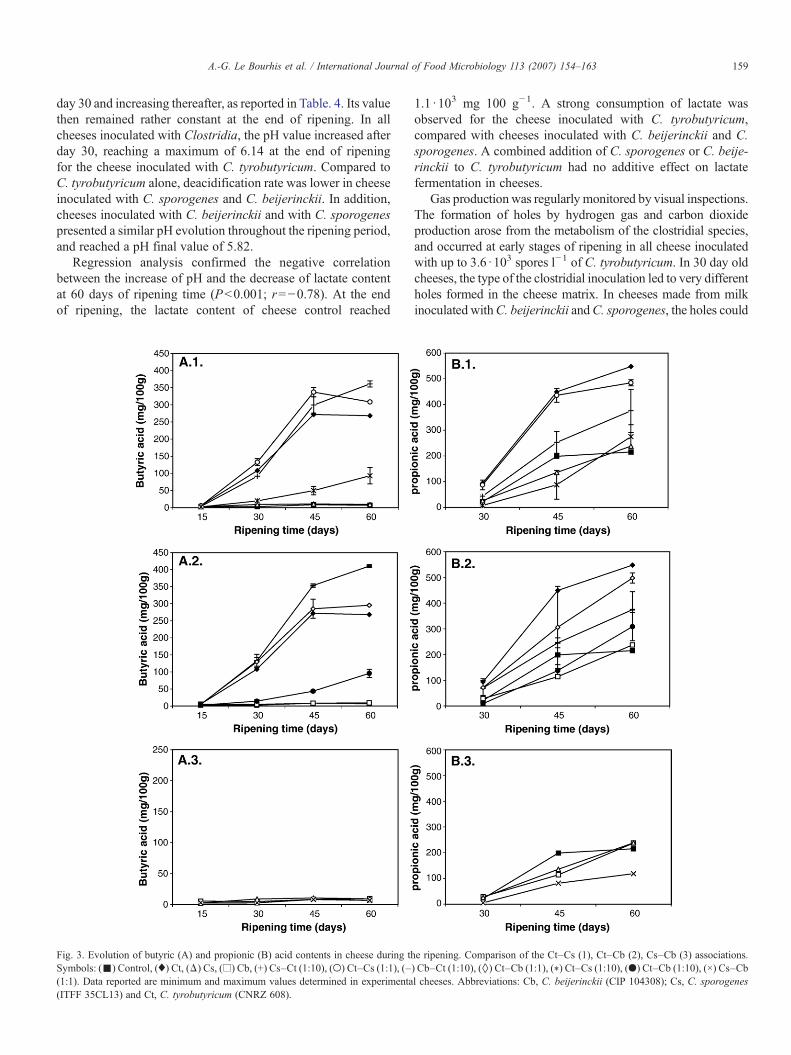
Fig. 3. Evolution of butyric (A) and propionic (B) acid contents in cheese during thSymbols: (▪) Control, (♦) Ct, (Δ) Cs, (□) Cb, (+) Cs–Ct (1:10), (○) Ct–Cs (1:1), (–(1:1). Data reported are minimum and maximum values determined in experimenta(ITFF 35CL13) and Ct, C. tyrobutyricum (CNRZ 608).
1.1 ·103 mg 100 g−1. A strong consumption of lactate wasobserved for the cheese inoculated with C. tyrobutyricum,compared with cheeses inoculated with C. beijerinckii and C.sporogenes. A combined addition of C. sporogenes or C. beije-rinckii to C. tyrobutyricum had no additive effect on lactatefermentation in cheeses.
Gas productionwas regularly monitored by visual inspections.The formation of holes by hydrogen gas and carbon dioxideproduction arose from the metabolism of the clostridial species,and occurred at early stages of ripening in all cheese inoculatedwith up to 3.6 ·103 spores l−1 of C. tyrobutyricum. In 30 day oldcheeses, the type of the clostridial inoculation led to very differentholes formed in the cheese matrix. In cheeses made from milkinoculated withC. beijerinckii andC. sporogenes, the holes could
e ripening. Comparison of the Ct–Cs (1), Ct–Cb (2), Cs–Cb (3) associations.) Cb–Ct (1:10), (◊) Ct–Cb (1:1), (⁎) Ct–Cs (1:10), (●) Ct–Cb (1:10), (×) Cs–Cbl cheeses. Abbreviations: Cb, C. beijerinckii (CIP 104308); Cs, C. sporogenes

160 A.-G. Le Bourhis et al. / International Journal of Food Microbiology 113 (2007) 154–163
be as small as 0.2–0.3 cm in diameter whereas in cheesesinoculated with C. tyrobutyricum their size could reach 1.0 cm.These strong differences of size remained rather constant until60 days of ripening, as reported in Fig. 2. Nevertheless, the holeswere larger for the cheese inoculated with C. sporogenes then forC. beijerinckii at the end of ripening. The holes also exhibiteddifferent shapes according to the type of contamination. Theywere quite clear and smooth for cheeses inoculated with C.sporogenes and withC. beijerinckii,whereas in cheese inoculatedwith C. tyrobutyricum the holes had the shape of a nut shell.
3.4. Volatile fatty acids analysis
Volatile fatty acids production in cheese were determinedthroughout the ripening period. Fig. 3A and B showed theevolution of the butyric (C4) and propionic (C3) acid contents.After 15 days of ripening, control cheese contained2.7mg 100 g−1
of butyric acid, and reached 6.7 mg 100 g−1 at the end of ripeningperiod. This low level of this volatile acid and low ratio of 2.3between butyric and caproïc acids (C6), showed that C4 originatedonly from lipolysis and was not produced by butyric fermentation(data not shown) (Mayenobe et al., 1983; Chamba and Perreard,2002).
3.4.1. Butyrate productionDifferences in the butyric pattern of cheese were clearly
detected after 15 days of ripening, and strongly depended on theclostridial inoculation. In cheese inoculated with C. tyrobutyr-icum, butyric acid was actively produced between day 15 andday 30, with an increase of 19 times, which resulted in thedevelopment of an intense rancid flavour and sweaty odor(Goudkov and Sharpe, 1966). The increase was still notableuntil 45 days of ripening, then an average constant value of270 mg 100 g−1 was determined till the end of the ripeningperiod. The time needed to reach 100 mg 100 g−1 of butyricacid was shorter for cheeses inoculated with 4 ·103 spores l−1 of
Fig. 4. Principal component analysis of the “late blowing” variables (pH, C4,C3) of the seventeen cheeses at 30 days of ripening (PC1 versus PC2). Thecheese numbers correspond to those in Table. 1.
C. tyrobutyricum than those inoculated with 4 ·102 spores l−1 ofthis species (30 days versus 60 days). Cheeses inoculated withC. sporogenes or with C. beijerinckii, demonstrated a con-tinuous butyric (C4) production, leading to a low final contentof approximately 9.4 mg 100 g−1 (Fig. 3.A.3). In cheeseinoculated with Ct–Cs or Ct–Cb mix (1:1), evolution of butyriclevel showed similar features than cheese inoculated withC. tyrobutyricum alone, with a higher amount of butyrate accu-mulated after 30 days of ripening. Surprisingly, butyric acid wasstill actively produced after 45 days of ripening with an initialspore count of C. sporogenes and C. beijerinckii 10 fold lowerthan C. tyrobutyricum. Cheeses inoculated with C. sporogenes–C. beijerinckii association and the control cheese displayed asimilar evolution of the C4 content. The C4/C6 ratio of allcheeses inoculated with C. tyrobutyricum measured throughoutthe ripening period indicated that the main part of butyric acidcontent came from butyric fermentation (data not shown). Anegative correlation (P<0.001; r=−0.96) was demonstrated bya statistical analysis of the decrease in lactic acid content in60 day old cheeses and the increase of butyrate accumulation.
3.4.2. Propionate productionThe type of contamination led to a strongly different pattern
of propionic acid production. At the end of ripening, the contentof C3 was increased from 1.3 to 2.5 times compared to controlcheese in presence of C. tyrobutyricum. A high correlationbetween propionic acid fermentation and butyric fermentationwas found from day 30 (P<0.001; r=0.86) until day 60(P=<0.001; r=0.74). In contrast, propionic fermentation incheeses inoculated with C. beijerinckii or with C. sporogenes,were lower than in the control cheese until 45 days, but reachedapproximately the same value at the end of the ripening (230±13 mg 100 g−1). With C. beijerinckii and C. sporogenes asso-ciated, the antagonistic effect on the propionic fermentation wasobserved, leading to an approximately 2 times lower C3 contentthan in the reference cheese control.
As reported in Fig. 4, Principal Component Analysis (PCA)provides a synthetic description of these data sets at 30 days ofthe ripening period. The variables pH, C4 and C3 were includedin the PCA. The map defined by principal components 1 and 2(81% and 16% of the variations at 30 days), allowed a gooddiscrimination between cheeses (P<0.05). PC1 is defined onthe one hand by C4 and C3 contents and on the other hand bypH values. PC2 is defined by C3 contents and by pH values.Control cheese which presented no sign of microbial spoilage orother undesirable changes in texture and odor, showed anegative value according to PC1, and a value close to zeroaccording to PC2. All cheeses characterized by a low butyricand propionic fermentation exhibited negative values accordingto PC1, and positive values according to PC2. All cheeses madefrom milk inoculated with up to 3.6 ·103 spores l−1 of C.tyrobutyricum, and characterized by a strong simultaneousbutyric and propionic fermentation showed positive valuesaccording to PC1. The negative or positive values according toPC2 represented slight differences concerning the intensities ofpropionic fermentation and deacidification between the cheeses.The slight differences between PCA at 30 and 60 days (data not

161A.-G. Le Bourhis et al. / International Journal of Food Microbiology 113 (2007) 154–163
shown), highlighted the early occurrence of the fermentationpattern of cheese.
4. Discussion
The present study investigated the behaviour of threedifferent species of Clostridia, C. tyrobutyricum, C. beijerinckiiand C. sporogenes, during the ripening of a hard-cooked-cheeseby a combination of classical and culture independent-basedmethods. The results showed no significant differences betweenany of the compositional parameters of the seventeen experi-mental cheeses before the ripening process, indicating similargrowth conditions between all cheeses. The relatively high levelof pH obtained at the start of ripening should have favoured theoutgrowth of clostridial spores. In all cheeses made withC. tyrobutyricum, the outgrowth of spores were particularlystrong whereas in cheeses inoculated with C. beijerinckii and/orC. sporogenes, the increase was lower. As the optimum pH rangefor growth of these two latter species is between 6.5 and 7.0(Cato et al., 1986), the relatively low pH of cheese may haveimpaired the outgrowth of spores in the beginning of ripening. Incontrast, the pH value of cheese was favourable to thedevelopment of C. tyrobutyricum, which displays an optimumgrowth pH at 5.8. This shows the variable levels of suitability ofcheese as “a culture medium” for the growth of these clostri-dial species. Surprisingly, an addition of C. beijerinckii orC. sporogenes to C. tyrobutyricum increased the level of viablespores in cheese. The development of C. tyrobutyricum at earlystages of ripening may have resulted in conditions stimulatinggermination, increasing the multiplication of C. beijerinckii andC. sporogenes, as suggested by the evolution of sporeconcentration and the lower deacidification rate of cheesecompared to cheese inoculated with C. tyrobutyricum alone. Inaddition, some compounds such as acetate and amino acidsissued from the metabolism of C. sporogenes and C. beijerinckiicould have favoured the germination of C. tyrobutyricum(Bergère and Hermier, 1970). In experiments to estimate thepredominance of each clostridial species, and to monitor theirevolution during the ripening period by using PCR-TTGEanalysis of the V5–V6 16S rDNA and cDNA amplicons,reproducibility of TTGE pattern from DNA amplicons ofClostridia was determined in previous work (Le Bourhis et al.,2005). Since the minor clostridial species cannot be detected ifthey account for less than 10% of the dominant species, the 16SrDNA- and 16S cDNA-derived profiles suggested thatC. beijerinckii and C. sporogenes in cheese were apparentlynumerically less important and metabolically less active thanC. tyrobutyricum (Le Bourhis et al., 2005).
During the ripening process, the increase of butyric acidconcentration in cheese without late blowing results fromlipolysis by secondary microflora (Kamaly and Marth, 1989). Anotable risk of defect due to an excessive butyric fermentationappears if the butyrate concentration in cheese is higher than20 mg 100 g−1, and if the C4/C6 ratio reaches values above 3(Mayenobe et al., 1983; Chamba and Perreard, 2002). At theend of the ripening period, the low level of C4 and the C4/C6ratio measured in control cheese, confirmed the usual ripening
of the reference cheese with the slight contribution of lipolysisto the production of C4. Results obtained for cheeses inoculatedwith C. beijerinckii and C. sporogenes demonstrated the lowinvolvement of these two clostridial species when compared tothe natural milk contaminants as previously observed (Klijn etal., 1995; Le Bourhis et al., 2005). These results suggested thatcollection strains C. beijerinckii and C. sporogenes may be notwell adapted any more to the cheese environment, and possiblydisplay a metabolism less active than wild strains. In all cheesesinoculated with C. tyrobutyricum, the rapid production ofbutyrate, simultaneously with the growth of this species, and thelow lactate content at the end of the ripening, demonstrated thestrong contribution of C. tyrobutyricum to butyric fermen-tation The addition of C. sporogenes or C. beijerinckii toC. tyrobutyricum enhanced the butyric production compared tothe cheese inoculated with C. tyrobutyricum alone. Further-more, the effect of the clostridial association in cheese was moreintense for C. beijerinckii–C. tyrobutyricum mix rather thanC. sporogenes–C. tyrobutyricum, and for a ratio between thesespecies of 1/10. As previously described in culture media, theformation of end-products related to the utilization of particularsubstrates, is clearly different in these three species, andinvolves a specific type of fermentation (Cato et al., 1986).C. beijerinckii and C. tyrobutyricym ferment carbohydrates viathe Embden–Meyerhof–Parnas pathway and produce butyrate,acetate, CO2 and H2. C. beijerinckii can also switch fromacidogenic phase to solventogenic phase, depending on the pHvalue, and solvents like acetone, butanol and ethanol areproduced as the final products (Yan et al., 1988; Rogers andGottschalk, 1993). C. sporogenes also uses the Embden–Meyerhof–Parnas pathway, converting lactate to butyrate, andpyruvate to acetate and butyrate, though its preferred substratesare amino acids, which are fermented by Stickland reactions(Cato et al., 1986; Allison and Macfarlane, 1990). Some end-products issued from the metabolism of C. beijerinckiiand C. sporogenes in cheese probably stimulate growthand metabolic activities of C. tyrobutyricum. In additionC. tyrobutyricum is not in competition with C. beijerinckiiand C. sporogenes for the utilization of lactate, the carbohydratesubstrate available in large amount in cheese.
Our results also demonstrated the implication ofC. beijerinckiiand C. sporogenes in the splitting of cheese, even though theywere numerically at a lower level than C. tyrobutyricum after30 days of ripening.Matteuzzi et al. observed a similar split defectin experimental cheese inoculated with C. sporogenes at 105
spores l−1 of milk (Matteuzzi et al., 1972). In a previous study,Klijn et al. demonstrated the relationship between C. tyrobutyr-icum and late blowing in experimental cheeses, but not theimplication of C. sporogenes and C. beijerinckii in the observeddefects (Klijn et al., 1995). The failure of these two latter speciesto induce a defect in experimental cheeses made with 105 to 106
spores l−1, suggested that the conditions of Gouda type cheese donot support a good growth of both, possibly due to the low pH.Even when they did grow, these two species apparently did notproduce hydrogen gas and butyric acid in this study. Kerjean et al.had also demonstrated that the reconstitution of butyricfermentation under controlled conditions was only possible in

162 A.-G. Le Bourhis et al. / International Journal of Food Microbiology 113 (2007) 154–163
cheese when the spores of Clostridia were prepared at low pH(Kerjean, 1995). In our study, the higher propionic productiondetected in 45 day old cheeses inoculated with C. tyrobutyricumcould be partly attributed to a greater deacidification resultingfrom the butyric fermentation. This is in good agreement withprevious works, where the development of C. tyrobutyricum wasfavoured in the presence of propionibacteria in cheese (Hunterand Frazier, 1961; Goudkov and Sharpe, 1966). All theseobservations suggested that C. tyrobutyricum and Propionibac-teria reciprocally stimulate their growth in cheese conditions. Incontrast, the correlation between the pH level and the C3 contentin cheese inoculated withC. beijerinckii and C. sporogenes couldexplain the weak propionic fermentation level. Even though, thelow pH is not a favourable factor for growth, other compoundsderived from the metabolism of these clostridial species couldhave contributed to this inhibition. No assumption can be madeconcerning the type of interaction between the propionicmicroflora and these Clostridia.
In agreement with previous works, the present studysupports the hypothesis that the presence of C. tyrobutyricumis a prerequisite for the occurrence of late blowing, but alsopoints out the implication of others clostridial species incheese defects (Bergère et al., 1968; Klijn et al., 1995; Inghamet al., 1998; Le Bourhis et al., 2005). Our results stronglysuggest that the metabolic activity of C. sporogenes and C.beijerinckii linked to the deacidification of cheese by C.tyrobutyricum, results in an increase in butyric acid fermen-tation. Chemical parameters such as cheese pH, propionic andbutyric acid contents had a strong correlation with theprevalence of late blowing. In association with molecularanalysis, they could be used to predict the intensity of cheesedefect at early stages of ripening and to understand the originof the defect.
A metabolic engineering approach should be useful toelucidate the physiological basis of these observations, and toquantify the relationship between the spoilage species undercheese conditions.
Acknowledgments
This project was financially supported by the SyndicatInterprofessionnel du Gruyère Français (Bourg en Bresse,France) and by the Association National de la RechercheTechnique (Paris, France).
The authors thank S. Zimmerman, M.O. Rimboud and O.Sgard for their technical assistance.
References
Accolas, J.P., Hemme, D., Desmazeaud, M.J., Vassal, L., Veaux, M., 1980. Leslevains lactiques thermophiles: propriétés et comportement en technologielaitière. Le Lait 60, 487–524.
Allison, C., Macfarlane, G.T., 1990. Regulation of protease production inClostridium sporogenes. Appl. Environ. Microbiol. 56, 3485–3490.
Berdagué, J.L., 1986. Méthode d'extraction rapide des acides gras volatils libresdes fromages. Le Lait 66, 233–246.
Bergère, J.L., Hermier, J., 1970. Spore properties of Clostridia occurring incheese. J. Appl. Bacteriol. 33, 167–179.
Bergère, J.L., Gouet, P., Hermier, J., Mocquot, G., 1968. Les Clostridium du groupebutyrique dans les produits laitiers. Ann. Inst. Pasteur, Lille 19, 41–54.
Bouton, Y., Grappin, R., 1994. Measurement of proteolysis in cheese: relationshipbetween phosphotungstic acid-soluble N fraction by Kjeldahl and 2,4,6trinitrobenzene–sulphonic acid reactive groups in water-soluble N. J. DairyRes. 61, 437–440.
Cato, E.P., George, W.L., Finegold, S.M., 1986. Genus Clostridium. In: Sneath,P.H.A., Mair, N.S., Sharpe, M.E., Holt, J.G. (Eds.), Bergey's Manu-al of Systematic Bacteriology, vol. 2. Williams and Wilkins, Baltimore,pp. 1141–1200.
Cerf, O., Bergère, J.L., Hermier, J., 1967. Thermorésistance des spores deClostridium tyrobutyricum et Clostridium butyricum. J. Dairy Res. 34,221–230.
Chamba, J.F., Perreard, E., 2002. Contribution of propionic acid bacteria tolipolysis of Emmental cheese. Le Lait 82, 33–44.
Collins, M.D., Lawson, P.A., Willems, A., Cordoba, J.J., Fernandez-Garayzabal,J., Garcia, P., Cai, J., Hippe, H., Farrow, J.A., 1994. The phylogeny of thegenus Clostridium: proposal of five new genera and eleven new speciescombinations. Int. J. Syst. Bacteriol. 44, 812–826.
Das Gupta, A.P., Hull, R.R., 1989. Late blowing of Swiss cheese: incidence ofClostridium tyrobutyricum in manufacturing milk. Aust. J. Dairy Technol.44, 82–87.
Doré, J., Sghir, A., Hannequart-Gramet, G., Corthier, G., Pochart, P., 1998.Design and evaluation of a 16S rRNA-targeted oligonucleotide probe forspecific detection and quantification of human faecal Bacteroides popula-tions. Syst. Appl. Microbiol. 21, 65–71.
Godon, J.J., Zumstein, E., Dabert, P., Habouzit, F., Moletta, R., 1997. Molecularmicrobial diversity of an anaerobic digestor as determined by small-subunitrDNA sequence analysis. Appl. Environ. Microbiol. 63, 2802–2813.
Goudkov, A.V., Sharpe, M.E., 1965. Clostridia in dairying. J. Appl. Bacteriol.28, 63–73.
Goudkov, A.V., Sharpe, M.E., 1966. A preliminary investigation of theimportance of Clostridia in the production of rancid flavour in Cheddarcheese. J. Dairy Res. 33, 139–149.
Hood, E.G., Bowen, J.F., 1950. A new type of bacterial spoilage in processcheese. Sci. Agric. 30, 38–42.
Hunter, J.E., Frazier, W.C., 1961. Gas production by associated Swiss cheesebacteria. J. Dairy Sci. 44, 2176–2186.
Ingham, S.C.,Hassler, J.R., Tsai, Y.W., Ingham,B.H., 1998.Differentiation of lactate-fermenting, gas-producing Clostridium spp. isolated from milk. Int. J. FoodMicrobiol. 43, 173–183.
Kamaly, K.M., Marth, E.H., 1989. Enzyme activities of lactic streptococci andtheir role in maturation of cheese: a review. J. Dairy Sci. 72, 1945–1966.
Kerjean, J.R. 1995. Study of antagonistic lactic acid bacteria against butyric acidfermentation in hard cooked cheeses. Flair Project 89057. Technical report.:12 pp.
Klijn, N.,Nieuwenhof, F.F., Hoolwerf, J.D., van derWaals, C.B.,Weerkamp, A.H.,1995. Identification ofClostridium tyrobutyricum as the causative agent of lateblowing in cheese by species-specific PCR amplification. Appl. Environ.Microbiol. 61, 2919–2924.
Le Bourhis, A.G., Saunier, K., Doré, J., Carlier, J.P., Chamba, J.F., Popoff, M.R.,Tholozan, J.L., 2005. Development and validation of PCR primers to assessthe diversity of Clostridium spp. in cheese by temporal temperature gradientgel electrophoresis. Appl. Environ. Microbiol. 71, 29–38.
Matteuzzi, D., Annibaldi, S., Sabatini, P., 1972. Importanza di Clostridiumsporogenes come agente di gonfiore del formaggio Grana. Ann. Microbiol.Enzimol. 22, 145–154.
Mayenobe, D., Didienne, R., Pradel, G., 1983. Caractérisation des gonflementstardifs dans les fromages de St-Nectaire et certaines pâtes pressées. Le Lait63, 15–24.
Rogers, P., Gottschalk, G., 1993. Biochemistry and regulation of acid andsolvent production in Clostridia. In: Woods, D.R. (Ed.), The Clostridia andBiotechnology. Butterworths-Heinemann, Stoneham, Mass, pp. 25–50.
Rosset, R., Julien, J., Monier, R., 1966. Ribonucleic acid composition ofbacteria as a function of growth rate. J. Mol. Biol. 18, 308–320.
Seksik, P., Rigottier-Gois, L., Gramet, G., Sutren, M., Pochart, P., Marteau, P.,Jian, R., Dore, J., 2003. Alterations of the dominant faecal bacterial groupsin patients with Crohn's disease of the colon. Gut 52, 237–242.

163A.-G. Le Bourhis et al. / International Journal of Food Microbiology 113 (2007) 154–163
Toyoda, S., Kobayashi, Y., Ahiko, K., 1990. Isolation and characterization ofClostridia from Gouda cheese with late-blowing. Jpn. J. Zootech. Sci. 61,591–598.
Yan, R.T., Zhu, C.X., Golemboski, C., et Chen, J.S., 1988. Expression ofsolvent-forming enzymes and onset of solvent production in batch cultures
of Clostridium beijerinckii (“Clostridium butylicum”). Appl. Environ.Microbiol. 54, 642–648.
Wagner, R., 1994. The regulation of ribosomal RNA synthesis and bacterial cellgrowth. Arch. Microbiol. 161, 100–109.
Woese, C.R., 1987. Bacterial evolution. Microbiol. Rev. 51, 221–271.















![Organic memory using [6,6] - phenyl-C 61 butyric acid methyl ester: morphology, thickness and concentration dependence studies](https://static.fdokumen.com/doc/165x107/634487eadf19c083b107a26c/organic-memory-using-66-phenyl-c-61-butyric-acid-methyl-ester-morphology.jpg)


![Intracellular reactions affecting 2-amino-4-([11C]methylthio)butyric acid ([11C]methionine) response to carbon ion radiotherapy in C10 glioma cells](https://static.fdokumen.com/doc/165x107/6343c86b88adeae9b9061aee/intracellular-reactions-affecting-2-amino-4-11cmethylthiobutyric-acid-11cmethionine.jpg)

