
Effects of gonadotropin-releasing hormone (GnRH) on the cytodifferentiation of gonadotropes in rat...
9
Cell Tissue Res (1994) 276:35-43 Cell Tissue Research 9 Springer-Verlag 1994 Effects of gonadotropin-releasing hormone (GnRH) on the cytodifferentiation of gonadotropes in rat adenohypophysial primordia in organ culture Akihiko Kudo, Min Kyun Park, Seiichiro Kawashima Zoological Institute, School of Science, Universityof Tokyo, Hongo, Tokyo, 113 Japan Received: 2 June 1993 / Accepted: 3 August 1993 Abstract. The effects of gonadotropin-releasing hormone (GnRH) on the development of gonadotropes were inves- tigated by the use of organ culture and by means of im- munocytochemistry and radioimmunoassay. Pituitary primordia from rat fetuses were cultured in a medium with or without 104 M GnRH during the first 24 h of culture. The ratio of the number of immunoreactive LH cells to the total number of cells in the explants derived from 13.5-day fetuses was increased by the GnRH treat- ment after 6 or 8 days of culture, while the total number of cells was not altered. LH released into the medium and LH content of explants were not affected by the GnRH treatment. Subsequent treatment with 104 M GnRH for 4 h after 7 days of culture resulted in a marked release of LH, accompanying a significant decline in LH content, in both explants exposed or unexposed to the first GnRH treatment. However, the former explants contained a lower amount of LH than the latter explants. The present results indicate that pituitary primordia at 13.5 days of gestation are capable to respond to GnRH, and that Gn- RH is effective in stimulating the responsiveness of go- nadotropes to GnRH during early pituitary cytodifferen- tiation. Key words: Organ culture - Pituitary primordium - Go- nadotropes - LH - GnRH - Rat (Wistar) Introduction It is well established that hypothalamic neurosecretory substances regulate the synthesis and release of pituitary hormones. However, it is not yet clear whether these hy- pothalamic factors affect the cytodifferentiation of the pituitary. Several investigators have concentrated their efforts to clarify the effects of hypothalamic factors on the cytodifferentiation of the pituitary gland. Organ culture Correspondence to: S. Kawashima studies using pituitary primordium of rats isolated before the appearance of immunoreactive(ir)-LH gonadotropes have revealed a variety of results, some of them contra- dictory (Nemesk6ry et al. 1976; Begeot et al. 1981, 1983 a, 1984; Gash et al. 1982; Watanabe 1982, 1985, 1987). There are conflicting reports concerning the temporal de- velopment of ir-LH gonadotropes in isolated pituitary cultures, possibly due to the difference in the culture con- ditions. Watanabe (1987) described that ir-LH cells ap- pear spontaneously in explants derived from rat fetuses at 11.5 days of gestation, the earliest fetal age described to date. The pituitary primordium at this stage may have a self-differentiation potency to develop LH gonadotropes and/or has had already determined primordial LH go- nadotropes. On the other hand, co-culture with brain and/or mesenchyme affects the size of explants and the number ofir-ACTH cells (Daikoku et al. 1982; Watanabe 1982) and LH cells (Watanabe 1985), indicating that fac- tors derived from these tissues may modify cytogenesis. The effect of gonadotropin-releasing hormone (Gn- RH) on the cytodifferentiation of gonadotropes remains open to discussion. Some investigators have postulated that GnRH acts as an inductive factor of gonadotropes during the early developmental stage of Rathke's pouch (Begeot et al. 1983 b, 1984; Horacek et al. 1989). Howev- er, see a contradictory report by Watanabe (1987). GnRH-binding sites in the pituitary primordia were determined using a radioreceptor assay by Aubert et al. (1985). These authors found significant binding of a ra- dioiodinated-GnRH analog to pituitary homogenates of fetuses at 12.5 and later days of gestation. Similarly, Jen- nes (1990) reported that GnRH analog-binding sites ap- peared in the pituitary primordia at day 13.5. Therefore, GnRH receptors may exist in the pituitary primordium at earlier stages than the appearance of ir-LH or FSH gonadotropes. These facts prompted us to study the effects of GnRH on the pituitary primordia in organ culture by means of radioimmunoassay and immunocytochemistry in order to clarify the secretory profile of gonadotropes and their responsiveness to GnRH.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Effects of gonadotropin-releasing hormone (GnRH) on the cytodifferentiation of gonadotropes in rat...

Cell Tissue Res (1994) 276:35-43 Cell Tissue
Research �9 Springer-Verlag 1994
Effects of gonadotropin-releasing hormone (GnRH) on the cytodifferentiation of gonadotropes in rat adenohypophysial primordia in organ culture Akihiko Kudo, Min Kyun Park, Seiichiro Kawashima
Zoological Institute, School of Science, University of Tokyo, Hongo, Tokyo, 113 Japan
Received: 2 June 1993 / Accepted: 3 August 1993
Abstract. The effects of gonadotropin-releasing hormone (GnRH) on the development of gonadotropes were inves- tigated by the use of organ culture and by means of im- munocytochemistry and radioimmunoassay. Pituitary primordia from rat fetuses were cultured in a medium with or without 104 M GnRH during the first 24 h of culture. The ratio of the number of immunoreactive LH cells to the total number of cells in the explants derived from 13.5-day fetuses was increased by the GnRH treat- ment after 6 or 8 days of culture, while the total number of cells was not altered. LH released into the medium and LH content of explants were not affected by the GnRH treatment. Subsequent treatment with 104 M GnRH for 4 h after 7 days of culture resulted in a marked release of LH, accompanying a significant decline in LH content, in both explants exposed or unexposed to the first GnRH treatment. However, the former explants contained a lower amount of LH than the latter explants. The present results indicate that pituitary primordia at 13.5 days of gestation are capable to respond to GnRH, and that Gn- RH is effective in stimulating the responsiveness of go- nadotropes to GnRH during early pituitary cytodifferen- tiation.
Key words: Organ culture - Pituitary primordium - Go- nadotropes - LH - GnRH - Rat (Wistar)
Introduction
It is well established that hypothalamic neurosecretory substances regulate the synthesis and release of pituitary hormones. However, it is not yet clear whether these hy- pothalamic factors affect the cytodifferentiation of the pituitary. Several investigators have concentrated their efforts to clarify the effects of hypothalamic factors on the cytodifferentiation of the pituitary gland. Organ culture
Correspondence to: S. Kawashima
studies using pituitary primordium of rats isolated before the appearance of immunoreactive(ir)-LH gonadotropes have revealed a variety of results, some of them contra- dictory (Nemesk6ry et al. 1976; Begeot et al. 1981, 1983 a, 1984; Gash et al. 1982; Watanabe 1982, 1985, 1987). There are conflicting reports concerning the temporal de- velopment of ir-LH gonadotropes in isolated pituitary cultures, possibly due to the difference in the culture con- ditions. Watanabe (1987) described that ir-LH cells ap- pear spontaneously in explants derived from rat fetuses at 11.5 days of gestation, the earliest fetal age described to date. The pituitary primordium at this stage may have a self-differentiation potency to develop LH gonadotropes and/or has had already determined primordial LH go- nadotropes. On the other hand, co-culture with brain and/or mesenchyme affects the size of explants and the number ofir-ACTH cells (Daikoku et al. 1982; Watanabe 1982) and LH cells (Watanabe 1985), indicating that fac- tors derived from these tissues may modify cytogenesis.
The effect of gonadotropin-releasing hormone (Gn- RH) on the cytodifferentiation of gonadotropes remains open to discussion. Some investigators have postulated that GnRH acts as an inductive factor of gonadotropes during the early developmental stage of Rathke's pouch (Begeot et al. 1983 b, 1984; Horacek et al. 1989). Howev- er, see a contradictory report by Watanabe (1987).
GnRH-binding sites in the pituitary primordia were determined using a radioreceptor assay by Aubert et al. (1985). These authors found significant binding of a ra- dioiodinated-GnRH analog to pituitary homogenates of fetuses at 12.5 and later days of gestation. Similarly, Jen- nes (1990) reported that GnRH analog-binding sites ap- peared in the pituitary primordia at day 13.5. Therefore, GnRH receptors may exist in the pituitary primordium at earlier stages than the appearance of ir-LH or FSH gonadotropes.
These facts prompted us to study the effects of GnRH on the pituitary primordia in organ culture by means of radioimmunoassay and immunocytochemistry in order to clarify the secretory profile of gonadotropes and their responsiveness to GnRH.

36
Materials and methods
Animals and organ culture
Rats of the Wistar strain maintained in this laboratory were used in the present study. They were housed in a temperature-controlled room at 12 h light (06:00-18:00) and 12 h dark cycle with free access to laboratory chow and tap water. Sexually mature female rats were placed in a cage with males at the evening of proestrus. Vaginal smears were examined for successful pregnancy the follow- ing morning, and 12:00 noon was designated as day 0.5 of gesta- tion. This designation of fetal age was adopted by Watanabe et al. (1973).
Fetuses were taken out from pregnant females at 12.5, 13.5 or 14.5 days of gestation. Disregarding the sex of fetuses, pituitary primordia with surrounding tissue were dissected out from fetuses according to the method of Watanabe et al. (1973), with minor modifications, and incubated with 1000 PU/ml Dispase (Godo Shu- sei, Tokyo, Japan; the main activity of Dispase is that of trypsin) in culture medium at room temperature for 15 min. After immersion in Dispase solution, the adenohypophysial primordium (designated as pituitary primordium for brevity) was completely separated from surrounding tissue, which consisted of the diencephalic floor, neuro- hypophysial primordium and mesenchymal tissue, using watch- maker's forceps under a dissecting microscope.
The explants were maintained in Trowell chambers (Trowell 1958). Each isolated primordium was placed on a 2 x 2 mm 2 piece of Millipore filter (type HA; pore size 0.45 gm, Millipore, USA). These membranes were placed on a mesh support (#~ 40) of stainless steal in 0.8 ml culture medium in Falcon organ culture dishes (No.3037, Becton Dickinson, USA). In this system explants were immediately under the surface of the medium. The explants were incubated at 37~ in an atmosphere of 5% CO2 and 95% air and maintained in vitro usually until the day equivalent to 21.5 days of gestation, with one of exception (Experiment 2). Explants on Mil- lipore filters were fixed in Bouin's solution for 12 h for cytological observations. Other explants were homogenized in 0.01 M phos- phate-buffered saline (pH 7.5) containing 1% bovine serum albumin (1% BSA-PBS), centrifuged at 10000 x g for 20 min at 4~ and the supernatants were stored at - 3 0 ~ for radioimmunoassay. Ho- mogenates of the explants were made separately or from two ex- plants together.
Basal medium for culture consisted of a mixture of Dulbecco's modified Eagle's medium (DME) and Ham's nutrient mixture F-12 (DME/F12 mixture, Sigma, USA) containing 15 mM HEPES, 3% fetal bo vine serum (F B S; GI B CO, USA) and antibiotics ( 100 IU/ml penicillin and 100 mg/ml streptomycin). The solution used for Gn- RH treatment contained 10 9 M GnRH (Peptide Institute, Osaka, Japan) dissolved in acetic acid at a final concentration of 0.4 mM in the basal medium. For control experiments, the basal medium with acetic acid at the same concentration was used (control medium). GnRH treatment was carried out during the first 24 h of culture. After treatment, the media were renewed after washing culture chambers twice. In subsequent cultures, the medium was changed to fresh basal medium every other day. The media after culture were centrifuged at 10000 x g for 20 min at 4~ and the supernatants were stored at - 3 0 ~ until radioimmunoassay.
Radioimmunoassay (RIA)
NIDDK-ra t LH (rLH)-I-7 was radioiodinated with ~25I (Na125I, Amersham, UK) by the chloramine-T method (Greenwood et al. 1963) with minor modifications, and the radioiodinated LH was separated by gel filtration on a Sephadex G-50 (Pharmacia, Sweden) column with 0.05 M phosphate buffer (pH 7.5) as elutant. The frozen media and the supernatants of tissue homogenates were as- sayed by RIA using the kit provided by the National Hormone and Pituitary Program, NIDDK. The assay system consisted of 200 gl
1% BSA-PBS, 100 gl standard hormone (NIDDK-rLH-RP-3) or samples, 100 gl primary antiserum (rabbit; NIDDK-anti-rLH-S-10) diluted with 1% normal rabbit serum in 0.05 M EDTA-PBS, and 100 gl [125I] iodo-LH in 1% BSA-PBS. Sample (or standard hor- mone), antiserum and buffer were incubated for 24 h, and iodinated hormone was added and incubated for an additional 12 h. Then 250 gl second antiserum (goat anti-rabbit IgG serum supplied by Dr. K. Wakabayashi, Gunma University) diluted with 3.5% polyethylene glycol 6000 in 0.05 M EDTA-PBS were added, and incubated for 3 h. Bound [125I] iodo-LH was precipitated by centrifugation at 1500 x g for 15 min, and the radioactivity was counted in an au- towell gamma counter. All incubation procedures were performed at room temperature. The minimal detectable level of LH was 31.25 + 0.05 pg/ml. Values are expressed in terms of pg NIDDK-rLH- RP-3/explant.
Immunoc yt ochemistr y
Fixed explants adhering to the filters were dehydrated and embed- ded in Paraplast Plus (Sherwood Medical, USA). Serial sections were cut at 3 gm thickness in a transverse plane and mounted on glass slides coated with gelatin. Sections were divided into two series of successive 5 sections, and each series was used for immuno- cytochemistry of LH or FSH.
The sections were immunostained by using Vectastain elite ABC kit (Vector Laboratories, USA), following the accompanying proto- col. Rabbit anti-rat LH and anti-rat FSH antisera (NIDDK-anti- rLH-S-10 and NIDDK-anti-rFSH-S-11, respectively) were provid- ed by NIDDK. These primary antisera were used at the final dilu- tion of x 22500 for LH and x 15000 for FSH, which were diluted with 0.1% BSA-PBS, and sections were incubated with these antis- era at 4~ overnight. Sections were counterstained with Mayer's hematoxylin, dehydrated and mounted in Entellan (E. Merck, Ger- many).
Preabsorption tests were performed on sections of explants derived from 13.5-day fetuses. The addition of an excess amount of cold rat LH (NIDDK-rLH-I-7, 0.5 gg) or cold rat FSH (NIDDK- rFSH-I-7, 2 gg) to diluted primary antisera (100 gl anti LH-S-10 x 22500, or 100 gl anti FSH-S-I1 x 15000) completely inhibited specific binding of these antisera to tissue sections.
The total number of nuclei of immunoreactive (ir) cells in the sections was counted in every 5th section (30 gm apart) of each section series for LH or FSH immunocytochemistry. At the same time, the total number of nuclei in the sections was counted in order to calculate the ratio of the number of ir cells to that of total cells. The ratio was used as an index of the LH or FSH cell population in the explant.
Experimental protocol
Experiment 1. Pituitary primordia from fetuses at 12.5, 13.5 or 14.5 days of gestation were cultured until the day equivalent to 21.5 days of gestation with or without GnRH treatment on the first day of culture. Culture periods for the 3 age groups were 9, 8 and 7 days, respectively. Cultured explants were processed for immunocyto- chemistry for LH and FSH. As the initial controls for the total numbers of cells in explants, some cultures were terminated 3 h after the explantation to facilitate adhesion to Millipore filters and the total number of cells in explants was counted on sections stained with hematoxylin.
Experiment 2. Pituitary primordia from fetuses at 13.5 days of gesta- tion were cultured for 0, 2, 4, 6, 8 and 10 days with or without GnRH treatment on the first day of culture. The ratio (%) of the
number of ir-LH cells to that of the total number of cells was compared between the two groups. The explants incubated for 3 h in the same way as Experiment 1 served as 0-day cultures.
Experiment 3. Gonadotropin concentrations in the media and su- pernatants of tissue homogenates were measured by RIA to evalu- ate the responsiveness of pituitary explants to GnRH. In this exper- iment, GnRH treatment was performed twice with an interval of 7 days. The first GnRH treatment on the first day of culture lasted for 24 h and the second one on the 8th day after 7 days of culture lasted for 4 h. The total period of culture of the pituitary primordia from fetuses at 13.5 days of gestation was 8 days, the terminal day being equivalent to 21.5 days of gestation. Media were changed every other day before the 8th day and every 4 h after the second GnRH treatment. At the end of 8 days of culture, most cultures were termi- nated and homogenized, and some cultures were used for LH im- munocytochemistry.
Statistics
Statistical analysis was performed using the Student's t-test for the data with homologous variance or Cochran-Cox test for the data without homologous variance, depending on the results of an anal- ysis of variance.
Results
Effects of GnRH on the population of gonadotropes in pituitary explants derived from fetuses of different gestational ages (Experiment 1)
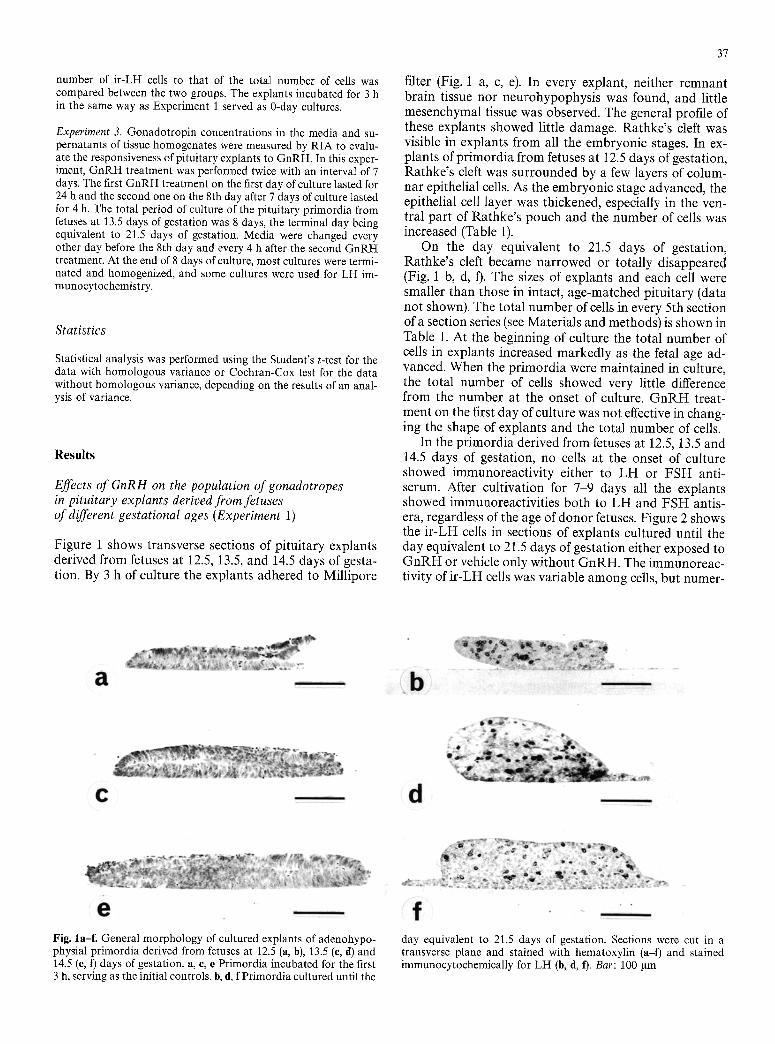
Figure 1 shows transverse sections of pituitary explants derived from fetuses at 12.5, 13.5, and 14.5 days of gesta- tion. By 3 h of culture the explants adhered to Millipore
37
filter (Fig. 1 a, c, e). In every explant, neither remnant brain tissue nor neurohypophysis was found, and little mesenchymal tissue was observed. The general profile of these explants showed little damage. Rathke's cleft was visible in explants from all the embryonic stages. In ex- plants of pr imordia from fetuses at 12.5 days of gestation, Rathke's cleft was surrounded by a few layers of colum- nar epithelial cells. As the embryonic stage advanced, the epithelial cell layer was thickened, especially in the ven- tral part of Rathke's pouch and the number of cells was increased (Table 1).
On the day equivalent to 21.5 days of gestation, Rathke's cleft became narrowed or totally disappeared (Fig. 1 b, d, ft. The sizes of explants and each cell were smaller than those in intact, age-matched pituitary (data not shown). The total number of cells in every 5th section of a section series (see Materials and methods) is shown in Table 1. At the beginning of culture the total number of cells in explants increased markedly as the fetal age ad- vanced. When the pr imordia were maintained in culture, the total number of cells showed very little difference from the number at the onset of culture. G n R H treat- ment on the first day of culture was not effective in chang- ing the shape of explants and the total number of cells.
In the pr imordia derived from fetuses at 12.5, 13.5 and 14.5 days of gestation, no cells at the onset of culture showed immunoreactivi ty either to LH or FSH anti- serum. After cultivation for 7-9 days all the explants showed immunoreactivities both to LH and FSH antis- era, regardless of the age of donor fetuses. Figure 2 shows the i r -LH cells in sections of explants cultured until the day equivalent to 21.5 days of gestation either exposed to G n R H or vehicle only without GnRH. The immunoreac- tivity of i r -LH cells was variable among cells, but numer-
Fig. la-f. General morphology of cultured explants of adenohypo- physial primordia derived from fetuses at 12.5 (a, b), 13.5 (e, d) and 14.5 (e, t) days of gestation, a, e, e Primordia incubated for the first 3 h, serving as the initial controls, b, d, f Primordia cultured until the
day equivalent to 21.5 days of gestation. Sections were cut in a transverse plane and stained with hematoxylin (a-f) and stained immunocytochemically for LH (b, d, t). Bar: 100 gm

38
Table 1. Changes in the total number of cells in explants derived from fetuses of dif- ferent gestational ages. Number of explants examined is indicated in parentheses. Sig- nificant difference from the value at the be- ginning of culture. * P < 0.05
Groups Gestational ages of donor fetuses
12.5 13.5 14.5
At the beginning of culture
At the termination of culture
Control GnRH
920 • 89 (6) a 2688 + 260 (6) 5238 • 136 (3)
887_+ 95 (5) 2729_+437 (8) 6404-t-765 (7) 924+191(6) 3204+207(8) 7929_+656(6)*
a Mean_+ SEM
Fig. 2a-f. Effects of GnRH treatment on ir-LH cells in the central part of cultured pituitary primordia. The primordia were cultured until the day equivalent to 21.5 days of gestation. Donor age and treatment: a 12.5 days, control; b 12.5 days, GnRH-treated; e 13.5
days, control; d 13.5 days, GnRH-treated; e 14.5 days, control; f 14.5 days, GnRH-treated. GnRH was added to the media on the first day of culture for 24 h. Bar: 20 I-tm
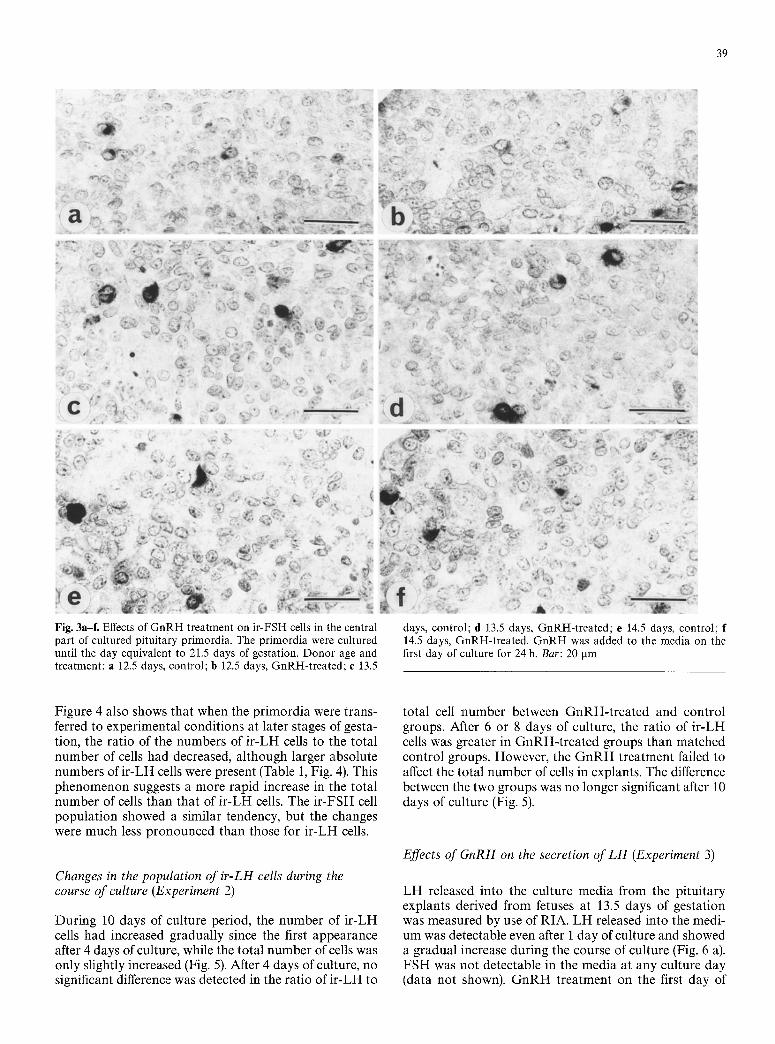
ous cells were s t rong ly s tained. Sect ions of G n R H - t r e a t - ed exp lan t s show s o m e w h a t m o r e i r - L H cells t han those of n o n - t r e a t e d explants . On the con t ra ry , on ly a smal l n u m b e r of cells showed s t rong i m m u n o r e a c t i v i t y to F S H and mos t of t hem were weak ly s ta ined (Fig. 3).
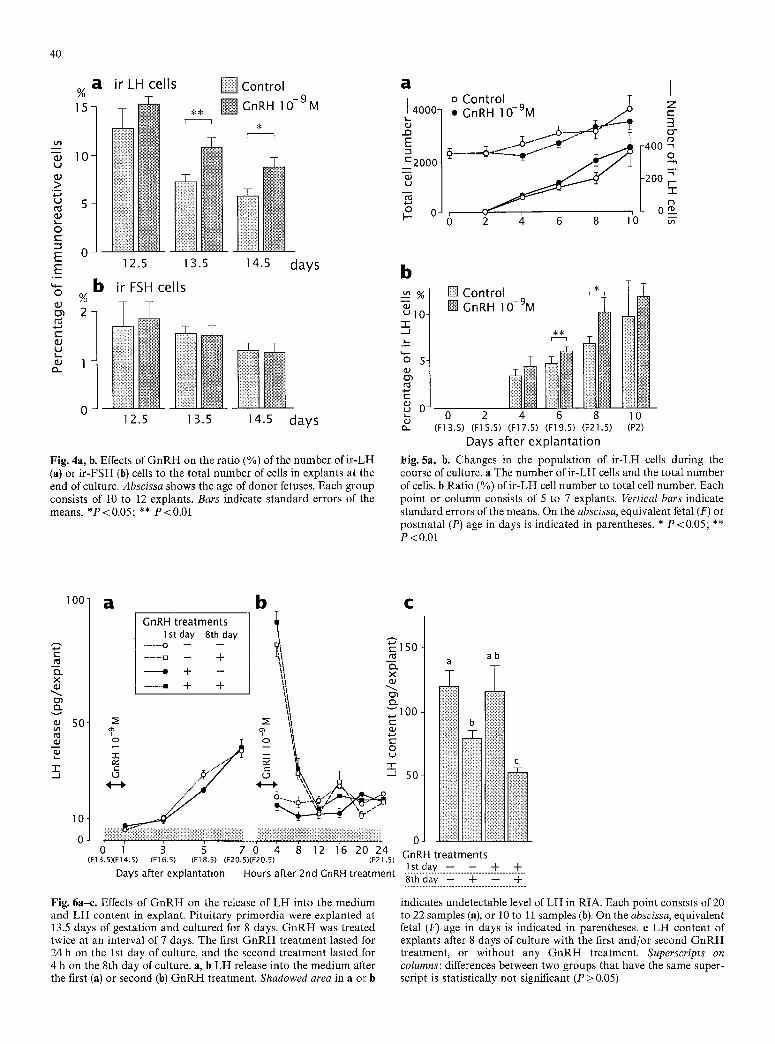
The ra t io of the n u m b e r of i r - L H and i r - F S H cells to tha t of to ta l cells is shown in Fig. 4. G n R H t r e a t m e n t was effective in increas ing the ra t io of i r - L H cells in cul tures der ived f rom 13.5 a n d 14.5 days of gestat ion. However , the t r e a t m e n t d id no t affect p o p u l a t i o n of i r - F S H cells.
39
Fig. 3a-f. Effects of GnRH treatment on ir-FSH cells in the central part of cultured pituitary primordia. The primordia were cultured until the day equivalent to 21.5 days of gestation. Donor age and treatment: a 12.5 days, control; b 12.5 days, GnRH-treated; e 13.5
days, control; d 13.5 days, GnRH-treated; e 14.5 days, control; f 14.5 days, GnRH-treated. GnRH was added to the media on the first day of culture for 24 h. Bar: 20 gm
Figure 4 also shows that when the pr imordia were trans- ferred to experimental conditions at later stages of gesta- tion, the ratio of the numbers of i r -LH cells to the total number of cells had decreased, although larger absolute numbers o f i r -LH cells were present (Table 1, Fig. 4). This phenomenon suggests a more rapid increase in the total number of cells than that of i r -LH cells. The i r -FSH cell populat ion showed a similar tendency, but the changes were much less pronounced than those for i r -LH cells.
Changes in the population of ir-LH cells during the course of culture (Experiment 2)
During 10 days of culture period, the number of i r -LH cells had increased gradually since the first appearance after 4 days of culture, while the total number of cells was only slightly increased (Fig. 5). After 4 days of culture, no significant difference was detected in the ratio of i r -LH to
total cell number between GnRH- t rea ted and control groups. After 6 or 8 days of culture, the ratio of i r -LH cells was greater in GnRH- t rea ted groups than matched control groups. However, the G n R H treatment failed to affect the total number of cells in explants. The difference between the two groups was no longer significant after 10 days of culture (Fig. 5).
Effects of GnRH on the secretion of LH (Experiment 3)
LH released into the culture media from the pituitary explants derived from fetuses at 13.5 days of gestation was measured by use of RIA. L H released into the medi- um was detectable even after 1 day of culture and showed a gradual increase during the course of culture (Fig. 6 a). FSH was not detectable in the media at any culture day (data not shown). G n R H treatment on the first day of
40
% a ir LH cells ~ Control
15- I ** ~ G n R H 10-9M
I i I ~ I
0t 10
1 2.5 1 3.5 14.5 d a y s
o o b ir FSH cells
g
0 12.5 13 .5 14 .5 d a y s
Fig. 4a, b. Effects of G n R H on the ratio (%) of the number of i r -LH (a) or ir-FSH (b) cells to the total number of cells in explants at the end of culture. Abscissa shows the age of donor fetuses. Each group consists of 10 to 12 explants. Bars indicate standard errors of the means. *P<0.05; ** P<0 .01
a
i 14000-
_Q
c2000-
o
I o Control 9 ~ z �9 GnRH 10- M <~
[ -
200 "-r"
L Ore_.. 0 2 4 6 8 10
b ~ %
w l O - "r- ._J
r ~
~ 0 -
[ ] C o n t r o l [ ] CnRH 1~
0 2 4 6 8 10 ~J ~- (F13.5) (F15.5) (F17.5) (F19.5) (F21.5) (P2)
Days after explantation Fig. 5a, b. Changes in the population of ir-LH cells during the course of culture, a The number of ir-LH cells and the total number of cells, b Ratio (%) of i r -LH cell number to total cell number. Each point or column consists of 5 to 7 explants. Vertical bars indicate standard errors of the means. On the abscissa, equivalent fetal (F) or postnatal (P) age in days is indicated in parentheses. * P < 0.05; ** P<0 .01
4.a r
n~
X
v
aJ u~ n~
-i- ._l
a b
50
10
-0
z; T O
"l- n,"
ID
GnRH treatments
. . . . . . . . . 0
- - - - - - Q
6 I' (F13.5)(F14.S) (F16.5)
Days after explantation
lstday 8thday _~
�9 + - ;i x ~, + + ~ - - - - - - i ~, r
i! r
z; ~i e-
~ �9 - - - k 9
_.1
/' 6 .......... ]); -.4 -~ ..o
i 7""6 4 8 I'2 1'6 2'0 2'4 (F18.5) (F20.S)(F20.5) (F21.5)
Hours after 2nd GnRH treatment
C
7 15o
~ 1 0 0
ab
~!i!!!i!!!i!!i~!i!~!
ililililililililililil b ii iiiiiiiiiiiiiiiiii, !i!i!i!i!i!i!i!iii!ill !i%ii!iii!i
iiii!~!ii~ii~ill ill
..................... iiiiiiiiii!i!i!iiiiiiiil ~ 50 ii!~ii~iiiii~i~iii~ �84 ii:iii~ii;~il iii~iil iii:,iiiiii'iiiiill ~ ii~!ii~ili~ii~ii~ii !iiiiiiiiii!i!iiiiiiiiil
!i!:!i!i!i!:!i!!i!!: i,~i~i~i~iiii~ii iiiiiiiiiiiiiiiiiiiiiil ~i!:!i!i!i!i!i!i~i!i~ iiiiiiiiiiiiiiiiiiiii iii~i;ii~i;!iil ..... !i~!i~i!i~i!i~i!i!. i~i!i?i!:i!.!i! .!i~i!i!i'i!i~? ~
0 !!ili!ili~ii!~ili~ili~ );iiiiiiiiiii!ill ~I~:~IS:!~:L,!
GnRH treatments 1st day -- -- + + -~-ff ~ ~-----------u165
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
indicates undetectable level of LH in RIA. Each point consists of 20 to 22 samples (a), or 10 to 11 samples (b). On the abscissa, equivalent fetal (F) age in days is indicated in parentheses, e LH content of explants after 8 days of culture with the first and/or second G n R H treatment, or without any G n R H treatment. Superscripts on columns: differences between two groups that have the same super- script is statistically not significant (P > 0.05)
Fig. 6a-e. Effects of G n R H on the release of LH into the medium and LH content in explant. Pituitary primordia were explanted at 13.5 days of gestation and cultured for 8 days. G n R H was treated twice at an interval of 7 days. The first G n R H treatment lasted for 24 h on the 1st day of culture, and the second treatment lasted for 4 h on the 8th day of culture, a, b LH release into the medium after the first (a) or second (b) G n R H treatment. Shadowed area in a or b
41
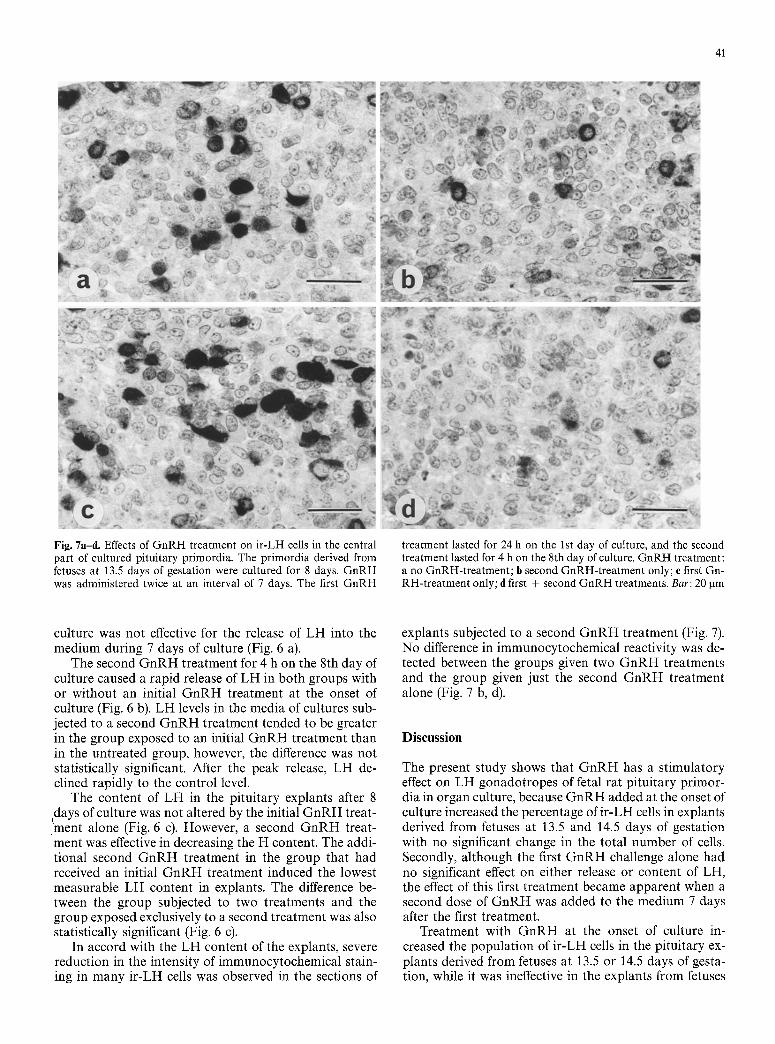
Fig. 7a-d. Effects of GnRH treatment on ir-LH cells in the central part of cultured pituitary primordia. The primordia derived from fetuses at 13.5 days of gestation were cultured for 8 days. GnRH was administered twice at an interval of 7 days. The first GnRH
treatment lasted for 24 h on the 1st day of culture, and the second treatment lasted for 4 h on the 8th day of culture. GnRH treatment: a no GnRH-treatment; b second GnRH-treatment only; e first Gn- RH-treatment only; d first + second GnRH treatments. Bar: 20 gm
culture was not effective for the release of LH into the medium during 7 days of culture (Fig. 6 a).
The second G n R H treatment for 4 h on the 8th day of culture caused a rapid release of L H in both groups with or without an initial G n R H treatment at the onset of culture (Fig. 6 b). LH levels in the media of cultures sub- jected to a second G n R H treatment tended to be greater in the group exposed to an initial G n R H treatment than in the untreated group, however, the difference was not statistically significant. After the peak release, LH de- clined rapidly to the control level.
The content of L H in the pituitary explants after 8 days of culture was not altered by the initial G n R H treat- 'ment alone (Fig. 6 c). However, a second G n R H treat- ment was effective in decreasing the H content. The addi- tional second G n R H treatment in the group that had received an initial G n R H treatment induced the lowest measurable L H content in explants. The difference be- tween the group subjected to two treatments and the group exposed exclusively to a second treatment was also statistically significant (Fig. 6 c).
In accord with the L H content of the explants, severe reduction in the intensity of immunocytochemical stain- ing in many i r -LH cells was observed in the sections of
explants subjected to a second G n R H treatment (Fig. 7). No difference in immunocytochemical reactivity was de- tected between the groups given two G n R H treatments and the group given just the second G n R H treatment alone (Fig. 7 b, d).
Discuss ion
The present study shows that G n R H has a stimulatory effect on L H gonadotropes of fetal rat pituitary primor- dia in organ culture, because G n R H added at the onset of culture increased the percentage of i r -LH cells in explants derived from fetuses at 13.5 and 14.5 days of gestation with no significant change in the total number of cells. Secondly, al though the first G n R H challenge alone had no significant effect on either release or content of LH, the effect of this first t reatment became apparent when a second dose of G n R H was added to the medium 7 days after the first treatment.
Treatment with G n R H at the onset of culture in- creased the populat ion of i r -LH cells in the pituitary ex- plants derived from fetuses at 13.5 or 14.5 days of gesta- tion, while it was ineffective in the explants from fetuses

42
at 12.5 days of gestation. Begeot et al. (1984) showed that GnRH at a concentration of 10 12 or 10 9 M in serum-free media stimulated the appearance of ir-LH cells in pitu- itary explants derived from 11.5- or 12.5-day fetuses. Ir- LH cells were missing in the pituitary explants in which GnRH was absent in the medium. On the other hand, Watanabe (1987) failed to detect any significant change in the population of ir-LH cells using GnRH at a higher concentration of 10 ng/ml (10 _8 M) in serum-free media. In his culture system ir-LH cells spontaneously appeared in explants derived from 11.5-day fetuses even when Gn- RH was not added to the media. These contradictory results with regard to the effect of GnRH may be caused by the difference in culture conditions.
Begeot et al. (1983 a) showed that medium supple- mented with insulin and transferrin was more effective for the differentiation of ir-LH gonadotropes than serum- supplemented medium. In their experiments, ir-LH go- nadotropes in explants from 13.5 day-fetuses appeared spontaneously in Medium 199 supplemented with insulin and transferrin, but not in serum-supplemented (20% FBS) medium. On the other hand, Watanabe (1987) found that the pituitary primordia of day 11.5- or 12.5-fe- tuses maintained in Medium 199 or ~ MEM developed ir-LH cells regardless of supplementation with insulin, transferrin, or thyroxine to serum-free medium or serum- supplemented medium. In our culture system, both ir-LH and ir-FSH cells appeared even in explants isolated from 12.5-day fetuses and cultured without GnRH stimulus. Concerning ir-LH gonadotropes, this result is in accord with the findings of Watanabe (1987). Therefore, it is sug- gested that the effects of GnRH found in the present study indicate its more important role for the modifica- tion of developmental processes than a role for the deter- mination of LH cells. Watanabe (1987) further showed that ir-LH cells in explants cultured in serum-supple- mented (5% horse and 2.5% calf serum) medium were more numerous and showed a stronger immunoreaction to anti LH serum. The sera used contain various agents for growth, differentiation and function, which are not thoroughly studied or identified and the precise role of which is not completely understood (Waymouth 1984). Although supplementation of serum to culture media is inferior to defined serum-free media because of consider- able variation in the composition of sera (Honn et al. 1975; Boone et al. 1972), culture conditions with supple- mentation of serum are favorable with respect to a greater potency of differentiation and/or proliferation of gonadotropes. Therefore, in the present study, the medi- um supplemented with 3% FBS was used throughout for the examination of GnRH effects. At this concentration of FBS, a maximal number of total cells was observed in the pituitary explants derived from days 12.5 and 13.5 fetuses in a preliminary experiment, where effects of 0 to 10% FBS were examined (data not shown). In the present study the ratio of ir-LH cells was clearly dependent on the addition of GnRH. It could not be decided in the present study whether any one factor in the serum played an important role in the alteration of the ir-LH cell pop- ulation or in its responsiveness to GnRH.
The present results suggest that the pituitary primor-
dia of 13.5- and 14.5-day fetuses are already responsive to GnRH and therefore possibly contain GnRH receptors, in corroboration of previous investigations on GnRH- binding sites in the pituitary primordia (Aubert et al. 1985; Jennes 1990). Because no ir-LH gonadotropes could be detected at the onset of culture, the GnRH re- ceptor-positive cells might not yet be 'functional' (hor- mone-secreting) gonadotropes. It is likely that 'primor- dial' gonadotropes, which are programmed to differenti- ate into functional gonadotropes, already possess the re- sponsiveness to GnRH. The stimulatory effect of GnRH on the ir-LH cell population was observed in the pitu- itary explants in culture with a long lag time of 6-8 days after withdrawal of GnRH. GnRH treatment at the onset of culture was without effect on the first appearance of ir-LH cells in explants. Moreover, the day of first appear- ance, i.e., day 5 of culture, is equivalent to 17.5 days of gestation, when ir-LH cells appear in fetal pituitary in vivo (Tougard et al. 1977; Watanabe and Daikoku 1979; Begeot et al. 1981; Nemesk6ri et al. 1988). These facts indicate that the onset of manifestation of ir-LH cells is programmed before 13.5 days of gestation; it is not al- tered by organ culture or GnRH treatment. At any rate, the increase in the ir-LH cell population may be the con- sequence of long-lasting GnRH effects on cytodifferentia- tion and/or proliferation of previously existing deter- mined cells.
Horacek et al. (1989) reported that the populations of ir-LH cells and ir-FSH cells were increased by GnRH treatment in the transplants of developing pituitary of the hamster. In the present study, a difference was noted be- tween ir-LH and ir-FSH cells. The results of the present study show that the population of FSH cells was not significantly affected by GnRH treatment, and suggest that the population of ir-LH and ir-FSH cells is differen- tially regulated by GnRH or that the culture conditions were not suitable to detect changes in the ir-FSH cell population.
A second GnRH challenge increased the release of LH into the medium, decreased LH content in the explants, and reduced immunoreactivity to anti-LH serum in sec- tions, regardless of the first GnRH treatment. Similar depletion of ir-LH by GnRH administration has been previously reported in the perinatal rat pituitary (Schafer and McShan 1974; Watanabe 1981). The present study indicates that gonadotropes respond to GnRH even in the pituitary primordium isolated from the fetuses at 13.5 days of gestation and cultured until the day equivalent to 21.5 days of gestation. Therefore, this responsiveness is self-differentiational, or previously determined.
The development of GnRH neurons in the hypothala- mus was investigated immunocytochemically by Daikoku et al. (1984). Ir-GnRH neurons were first detect- ed at 16.5 days of gestation in the dorso-medial portion of the hypothalamus and ventro-medial portion of the forebrain vesicles. At 17.5 days of gestation, GnRH neu- rons were further found in the septo-preoptic area (Daikoku et al. 1984). On the other hand, hypothalamic GnRH content was detectable by RIA as early as 12.5 days of fetal life (Aubert et al. 1985). Begeot et al. (1984) suggested that GnRH may be released from the brain

43
dur ing ear ly fetal ages. The G n R H - l i k e immunoreac t i v i - ty in the fetal p i t u i t a ry at 14.5 days of ges ta t ion (Auber t et al. 1985) indica tes tha t at this s tage the p i tu i t a ry con- ta ins G n R H . Therefore , any effect of G n R H t r ea tmen t on p i tu i t a ry p r i m o r d i u m in cul ture du r ing these fetal ages, as shown in the presen t s tudy, cer ta in ly reflects the phys- io logica l process of g o n a d o t r o p e different iat ion.
Acknowledgements. We would like to express our cordial thanks to Dr. S. Raiti, the National Hormone and Pituitary Program, Univer- sity of Maryland School of Medicine, NIDDK, and to Dr. A. F. Parlow, Pituitary Hormone/Antisera Center, Harbor-University of California-Los Angeles Medical Center, for supplying pituitary hor- mones and antisera. We also thank Dr. K. Wakabayashi, Institute of Endocrinology, Gunma University, for supplying anti-gamma glob- ulin serum. This work was supported in part by a Grant-in-Aid from the Ministry of Education, Science and Culture and a Re- search Grant from Zenyaku Kogyo, Japan to S. K.
References
Aubert ML, Begeot M, Winiger BP, Morel G, Sizonenko PO, Dubois PM (1985) Ontogeny of hypothalamic luteinizing hor- mone-releasing hormone (GnRH) and pituitary GnRH recep- tors in fetal and neonatal rats. Endocrinology 116:1565-1576
Begeot M, Dupouy JP, Dubois MP, Dubois PM (1981) Immunocy- tological determination of gonadotropic and thyrotropic cells in fetal rat anterior pituitary during normal development and un- der experimental conditions. Neuroendocrinology 32:285-294
Begeot M, Dubois MP, Dubois PM (1983 a) Comparative study in vivo and in vitro of the differentiation of immunoreactive go- nadotropic cells in fetal rat anterior pituitary. Neuroendocrinol- ogy 37:52-58
Begeot M, Hemming F J, Martinat N, Dubois MP, Dubois PM (1983 b) Gonadotropin releasing hormone (GnRH) stimulates immunoreactive lactotroph differentiation. Endocrinology 112:2224-2226
Begeot M, Morel G, Rivest RW, Aubert ML, Dubois MP, Dubois PM (1984) Influence of gonadoliberin on the differentiation of rat gonadotrophs: an in vivo and in vitro study. Neuroen- docrinology 38:217-225
Boone CW, Mantel N, Caruso TD, Kazam E, Stevenson RE (1972) Quality control studies of fetal bovine serum used in tissue cul- ture. In Vitro Cell Dev Biol 7:174-189
Daikoku S, Chikamori M, Adachi T, Maki Y (1982) Effects of the basal diencephalon on the development of Rathke's pouch in rats: a study in combined organ cultures. Dev Biol 90:198 202
Daikoku S, Maki Y, Okamura Y, Tsuruo Y, Hisano S (1984) Devel- opment of immunoreactive LHRH neurons in the fetal rat hy- pothalamus. Int J Dev Neurosci 2:113 120
Gash D, Ahmid N, Schechter J (1982) Comparison of gonadotroph, thyrotroph and mammotroph development in situ, in transplant and in organ culture. Neuroendocrinology 34:222-228
Greenwood FC, Hunter WM, Glover JS (1963) The preparation of 131I-labelled human growth hormone of high specific activity. Biochem J 89:114-123
Honn KV, Singley JA, Chavin W (1975) Fetal bovine serum: a muttivarience standard. Proc Soc Exp Biol Med 149:344-347
Horacek MJ, Campbell GT, Blake CA (1989) Luteinizing hormone (LH)-releasing hormone: effects on induction of LH, follicle- stimulating hormone, and prolactin cell differentiation. En- docrinology 124:1800-1806
Jennes L (1990) Prenatal development of gonadotropin-releasing hormone receptors in the rat anterior pituitary. Endocrinology 126: 942-947
Nemesk~ry A, N~meth A, S~tfil6 G, Vigh S, Hal~sz B (1976) Cell differentiation of the fetal rat anterior pituitary in vitro. Cell Tissue Res 170:263-273
Nemesk6ri A_, S6tfil6 G, Halfisz B (1988) Ontogenesis of the three parts of the fetal rat adenohypophysis: detailed immunohisto- chemical analysis Neuroendocrinology 48:534-543
Schafer S J, McShan WH (1974) Gonadotropic hormone release from fetal and adult pituitary glands after in vitro exposure to synthetic LH-FSH-RH. Neuroendocrinology 16:332-341
Tougard C, Picart R, Tixier-Vidal A (1977) Cytogenesis of im- munoreactive gonadotropic cells in the fetal rat pituitary at light and electron microscopic levels. Dev Biol 58:148-163
Trowell OA (1958) The culture of mature organs in a synthetic medium. Exp Cell Res 16:118-147
Watanabe YG (1981) Immunohistochemical study of the respon- siveness of the LH cells of fetal rats to synthetic LHRH in vitro. Cell Tissue Res 221:59-66
Watanabe YG (1982) Effects of brain and mesenchyme upon the cytogenesis of rat adenohypophysis in vitro. I. Differentiation of ACTH cells. Cell Tissue Res 227:257-266
Watanabe YG (1985) Effects of brain and mesenchyme upon the cytogenesis of rat adenohypophysis in vitro. II. Differentiation of LH cells. Cell Tissue Res 242:49-55
Watanabe YG (1987) Failure of luteinizing hormone-releasing hor- mone (LHRH) to affect the differentiation of LH cells in the rat hypophysial primordium in serum-free culture. Cell Tissue Res 250:35~42
Watanabe YG, Daikoku S (1979) An immunohistochemical study on the cytogenesis of adenohypophysial cells in fetal rats. Dev Biol 68:557-567
Watanabe YG, Matsumura H, Daikoku S (1973) Electron micro- scopic study of rat pituitary primordium in organ culture. Z Zellforsch 146:453461
Waymouth C (1984) Preparation and use of serum-free culture me- dia. In: Barnes DW, Sirbasku DA, Sato G (eds) Methods for preparation of media, supplements, and substrata for serum-free animal cell culture. Liss, New York, pp 23 68




















