
The impact of hemochromatosis mutations and transferrin genotype on gonadotropin serum levels in...
8
The impact of hemochromatosis mutations and transferrin genotype on gonadotropin serum levels in infertile men Alena Bureti c-Tomljanovi c, Ph.D., a Ivan Vlasteli c, M.D., M.S., b AnCelka Radoj ci c Badovinac, M.D., Ph.D., a,b Nada Star cevi c- Cizmarevi c, M.S., a Sergej Nadalin, M.D., a and Smiljana Risti c, Ph.D. a a Department of Biology and Medical Genetics, School of Medicine, University of Rijeka; and b Department of Human Reproduction, Clinic of Obstetrics and Gynaecology, Clinical Medical Centre Rijeka, Rijeka, Croatia Objective: To address the possibility that HFE mutations and TF gene polymorphism cause dysfunction of spermatogenesis and/or the hypothalamic-pituitary-gonadal axis via contribution to long-term iron overload in the testes and brain. Design: Case-control and association study. Setting: Clinic of obstetrics and gynecology and university-based research laboratory. Patient(s): 127 infertile men (including 97 with idiopathic infertility) and 188 controls of proven fertility. Intervention(s): Polymerase chain reaction/restriction fragment length polymorphism (PCR-RFLP). Main Outcome Measure(s): HFE mutations and transferrin allelic polymorphism, and testosterone, prolactin, and gonadotropin serum levels. Result(s): The frequencies of the analyzed alleles and genotypes showed no statistically significant difference between infertile men and controls. Sperm count and progressive sperm motility did not correlate with HFE or TF genotype, or their combination. After excluding patients with clinical hypogonadism or varicocele from further analysis, a statistically significant correlation between serum follicle-stimulating hormone and luteinizing hor- mone levels and the combined HFE H63D/TFC2 genotype was found in 97 men with idiopathic infertility. Conclusion(s): The combined HFE H63D/TF-C2 genotype contributed to 4.1% and 10.6% of follicle-stimulating hormone and luteinizing hormone variation, respectively, in infertile men, raising mean hormonal values above the normal physiologic range. Therefore, HFE and TF genes together may influence the hypothalamic-pituitary- gonadal axis, functioning at the pituitary or testes level. (Fertil Steril Ò 2009;91:1793–800. Ó2009 by American Society for Reproductive Medicine.) Key Words: Male infertility, HFE C282Y mutation, HFE H63D mutation, transferrin gene polymorphism, gonad- otropic hormones Iron is an essential cofactor in many cellular processes. As free iron is toxic due to induction of hydroxyl radical forma- tion and lipid peroxidation producing dangerous free radi- cals, iron homeostasis is a tightly regulated and balanced process (1). Both the hemochromatosis (Hfe) and transferrin (Tf) proteins control iron release from plasma and its subse- quent uptake by cells. Mutations in HFE (H63D and C282Y) have been shown to be associated with hereditary hemochro- matosis (HH), an autosomal recessive inherited disorder that is characterized by slow rises in iron and accumulation in dif- ferent tissues, including the liver, heart, pancreas, lungs, and brain (2, 3). Iron deposition has been well documented in the pituitary of the patients with iron overload (4), leading to im- paired gonadotropin secretion and hypogonadism, the second most common endocrinopathy after diabetes mellitus in HH. Peillon and Racadot (5) and Bergeron and Kovacs (6) previ- ously demonstrated iron deposition mainly in the gonadotro- pin secretory cells compared with other secretory cells within the anterior pituitary. However, the prevalence of hypogona- dotrophic hypogonadism (low testosterone level in associa- tion with low follicle-stimulating hormone [FSH] and luteinizing hormone [LH] levels) is only 6.4% in HH (7). The C282Y HFE mutation affects the b 2 microglobulin chain of HFE, allowing a higher rate of transferrin binding to the TfR1 transferrin receptor and thus a higher rate of in- ternalization of the Tf–TfR1 complex (8). The frequency of this mutation is decreased in southeastern Europe (1% to 5%) compared with northwestern Europe (6% to 10%) (9). By comparison, the distribution of the HFE H63D mutation in Europe is more generally distributed, with frequencies higher than 10% (9). The HFE H63D mutation has a milder effect on iron uptake and more effectively competes against the C282Y HFE mutation to prevent Tf-TfR1 binding and uptake. Transferrin is a plasma glycoprotein that binds to its recep- tors tightly, but reversibly, and delivers iron ions to cells. Received September 20, 2007; revised and accepted February 15, 2008; published online April 8, 2008. A.B-T. has nothing to disclose. I.V. has nothing to disclose. A.R.B. has nothing to disclose. N.S-C. has nothing to disclose. S.N. has nothing to disclose. S.R. has nothing to disclose. Reprint requests: Alena Bureti c-Tomljanovi c, Ph.D., Department of Biol- ogy and Medical Genetics, School of Medicine, University of Rijeka, Bra ce Branchetta 22, 51 000 Rijeka, Croatia (FAX: þ385-51-678-896; E-mail: [email protected]). 0015-0282/09/$36.00 Fertility and Sterility â Vol. 91, No. 5, May 2009 1793 doi:10.1016/j.fertnstert.2008.02.129 Copyright ª2009 American Society for Reproductive Medicine, Published by Elsevier Inc.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The impact of hemochromatosis mutations and transferrin genotype on gonadotropin serum levels in...

0015-0doi:10.
The impact of hemochromatosis mutations andtransferrin genotype on gonadotropin serum levels ininfertile menAlena Bureti�c-Tomljanovi�c, Ph.D.,a Ivan Vlasteli�c, M.D., M.S.,b AnCelka Radoj�ci�c Badovinac, M.D.,Ph.D.,a,b Nada Star�cevi�c-�Cizmarevi�c, M.S.,a Sergej Nadalin, M.D.,a and Smiljana Risti�c, Ph.D.a
a Department of Biology and Medical Genetics, School of Medicine, University of Rijeka; and b Department of Human
Reproduction, Clinic of Obstetrics and Gynaecology, Clinical Medical Centre Rijeka, Rijeka, Croatia
Objective: To address the possibility that HFE mutations and TF gene polymorphism cause dysfunction ofspermatogenesis and/or the hypothalamic-pituitary-gonadal axis via contribution to long-term iron overload inthe testes and brain.Design: Case-control and association study.Setting: Clinic of obstetrics and gynecology and university-based research laboratory.Patient(s): 127 infertile men (including 97 with idiopathic infertility) and 188 controls of proven fertility.Intervention(s): Polymerase chain reaction/restriction fragment length polymorphism (PCR-RFLP).Main Outcome Measure(s): HFE mutations and transferrin allelic polymorphism, and testosterone, prolactin, andgonadotropin serum levels.Result(s): The frequencies of the analyzed alleles and genotypes showed no statistically significant differencebetween infertile men and controls. Sperm count and progressive sperm motility did not correlate with HFE orTF genotype, or their combination. After excluding patients with clinical hypogonadism or varicocele from furtheranalysis, a statistically significant correlation between serum follicle-stimulating hormone and luteinizing hor-mone levels and the combined HFE H63D/TFC2 genotype was found in 97 men with idiopathic infertility.Conclusion(s): The combined HFE H63D/TF-C2 genotype contributed to 4.1% and 10.6% of follicle-stimulatinghormone and luteinizing hormone variation, respectively, in infertile men, raising mean hormonal values above thenormal physiologic range. Therefore, HFE and TF genes together may influence the hypothalamic-pituitary-gonadal axis, functioning at the pituitary or testes level. (Fertil Steril� 2009;91:1793–800. �2009 by AmericanSociety for Reproductive Medicine.)
Key Words: Male infertility, HFE C282Y mutation, HFE H63D mutation, transferrin gene polymorphism, gonad-otropic hormones
Iron is an essential cofactor in many cellular processes. Asfree iron is toxic due to induction of hydroxyl radical forma-tion and lipid peroxidation producing dangerous free radi-cals, iron homeostasis is a tightly regulated and balancedprocess (1). Both the hemochromatosis (Hfe) and transferrin(Tf) proteins control iron release from plasma and its subse-quent uptake by cells. Mutations in HFE (H63D and C282Y)have been shown to be associated with hereditary hemochro-matosis (HH), an autosomal recessive inherited disorder thatis characterized by slow rises in iron and accumulation in dif-ferent tissues, including the liver, heart, pancreas, lungs, andbrain (2, 3). Iron deposition has been well documented in thepituitary of the patients with iron overload (4), leading to im-paired gonadotropin secretion and hypogonadism, the second
Received September 20, 2007; revised and accepted February 15, 2008;
published online April 8, 2008.
A.B-T. has nothing to disclose. I.V. has nothing to disclose. A.R.B. has
nothing to disclose. N.S-C. has nothing to disclose. S.N. has nothing
to disclose. S.R. has nothing to disclose.
Reprint requests: Alena Bureti�c-Tomljanovi�c, Ph.D., Department of Biol-
ogy and Medical Genetics, School of Medicine, University of Rijeka,
Bra�ce Branchetta 22, 51 000 Rijeka, Croatia (FAX: þ385-51-678-896;
E-mail: [email protected]).
282/09/$36.001016/j.fertnstert.2008.02.129 Copyright ª2009 American
most common endocrinopathy after diabetes mellitus in HH.Peillon and Racadot (5) and Bergeron and Kovacs (6) previ-ously demonstrated iron deposition mainly in the gonadotro-pin secretory cells compared with other secretory cells withinthe anterior pituitary. However, the prevalence of hypogona-dotrophic hypogonadism (low testosterone level in associa-tion with low follicle-stimulating hormone [FSH] andluteinizing hormone [LH] levels) is only 6.4% in HH (7).
The C282Y HFE mutation affects the b2 microglobulinchain of HFE, allowing a higher rate of transferrin bindingto the TfR1 transferrin receptor and thus a higher rate of in-ternalization of the Tf–TfR1 complex (8). The frequency ofthis mutation is decreased in southeastern Europe (1% to5%) compared with northwestern Europe (6% to 10%) (9).By comparison, the distribution of the HFE H63D mutationin Europe is more generally distributed, with frequencieshigher than 10% (9). The HFE H63D mutation has a mildereffect on iron uptake and more effectively competes againstthe C282Y HFE mutation to prevent Tf-TfR1 binding anduptake.
Transferrin is a plasma glycoprotein that binds to its recep-tors tightly, but reversibly, and delivers iron ions to cells.
Fertility and Sterility� Vol. 91, No. 5, May 2009 1793Society for Reproductive Medicine, Published by Elsevier Inc.

According to Wong and Saha (10), the transferrin gene vari-ant C2 (TF C2) may have a lower affinity for iron comparedwith the C1 variant. Based on this finding, Robson et al. (11)suggested that bi-carriers of the HFE C282Y mutation and TFC2 variant may internalize more Tf–TfR1 complexes (due toHFE C282Y) and release more iron to endosomes (due to TFC2), possibly leading to an iron overload. Although the effectof iron overload on iron metabolism is largely unknown in or-gans such as the brain, several studies have suggested ironmisregulation in brain may be a source of oxidative stressand neurodegeneration (11, 12).
The first report of the possible association of HFE muta-tions with idiopathic male infertility showed no associationwith clinical data (13). Our study broadens the analysis ofthe relationship of iron metabolism-related genes with malefertility by including the transferrin gene C1/C2 polymor-phism because of the close functional relationship of thetwo proteins. We further hypothesize that synergistic effectof HFE mutations and transferrin gene variants may influencehormonal parameters in men with idiopathic infertility, pos-sibly acting on the hypothalamic-pituitary-gonadal (HPG)axis or on the testis level.
MATERIALS AND METHODS
Patients
Our study included 127 male patients in the Family PlanningDepartment of Clinic for Obstetrics and Gynaecology, ClinicalMedical Centre in Rijeka, Croatia, who were receiving assis-ted reproduction treatment due to spermatogenic impairmentor decreased sperm motility. The patients were all previouslyscreened for chromosomal aberrations and microdeletions ofthe AZF region of chromosome Y and were confirmed asnegative for both tests. Patients with obstructive azoospermiaor undergoing cancer treatment were excluded from the study.Karyotyping analyses were performed on GTG-banded slides,and 30 metaphases were examined for each patient. Patientswho were found to carry structural or numerical chromosomalaberrations were not included in the study.
Microdeletion of chromosome Y analyses were performedaccording to EMQN guidelines (14). The clinical character-istics of our 127 patients are presented in Table 1. Groupsof patients with spermatogenic impairment were defined ac-cording to sperm count as azoospermic, oligoasthenozoo-spermic (<5 � 106/mL and R5 � 106/mL, respectively),or normozoospermic. Twenty-four normozoospermic males(sperm count R20� 106/mL) were included due to astheno-spermia (progressive sperm motility <50%). Thirty of the127 patients (23.62%) were clinically diagnosed with hypo-gonadism, varicocele, unilateral cryptorchidism, prostatitis,inguinal hernia, or a combination. Clinical hypogonadismwas present in six men (hypergonadotropic in five casesand hypogonadotropic in one case). In the remaining 97 pa-tients (76.38%), infertility was idiopathic. The control pa-tients were 188 men with proven fertility. All tested andcontrol men were Croatian citizens who met the objectivesof the study and had signed an informed consent form. The
1794 Bureti�c-Tomljanovi�c et al. Iron-metabolizing genes i
study was approved by the School of Medicine (Universityof Rijeka, Croatia) and Clinical Medical Centre (Rijeka, Cro-atia) ethics committees.
Semen was analyzed according to the World HealthOrganization guidelines (15) using the UltiMate Sperm Ana-lyzer (Hamilton Thorne Biosciences, Beverly, MA). Spermconcentration was considered normal at R20 � 106 sperma-tozoa/mL, and progressive sperm motility was considerednormal at R50%. Serum FSH, LH, and prolactin levelswere assessed by a chemoluminescent assay (ImmuliteFSH, DPC; Immulite/Immulite 1000 LH, DPC; Immulite/Im-mulite 1000 Prolactin, Siemens Medical Diagnostics Ltd.,Llanberis, Gwynedd, United Kingdom), while total serumtestosterone levels were measured by a solid-phase radioim-munoassay (Coat-A-Count Total Testosterone In-vitro Diag-nostic Test Kit, Diagnostic Products Corporation, LosAngeles, CA) (16). Results were considered normal withinthe following ranges: FSH, 1.1–13.5 IU/L; LH, 0.4–5.7 IU/L; testosterone, 10.0–49.0 nmol/L; and prolactin, 65.0–350.0 mIU/L.
Genomic DNA was extracted from whole blood usinga QIAmp Blood Kit (Qiagen, Hildren, Germany) as describedby the manufacturer. Genotyping for HFE C282Y, HFEH63D mutations, and the TF C1/C2 polymorphism was per-formed by the polymerase chain reaction/restriction fragmentlength polymorphism (PCR-RFLP) method as described pre-viously elsewhere (11, 17). The possible effects of differentgonadotoxins in the form of chemicals, medications, tobacco,or alcohol were not evaluated in this study.
Statistical Analyses
Allele frequencies were calculated by gene counting, anddeviations from those estimated by the Hardy-Weinbergequilibrium were tested using the chi-square test. The statis-tical significance of the differences in frequency of observedalleles and genotypes between patients and controls wasdetermined using the chi-square test or Fisher’s exact test,as appropriate.
Mean, standard deviation, and difference of clinical pa-rameters (age, progressive sperm motility, hormonal parame-ters) between subgroups of patients were tested using one-way analysis of variance (ANOVA). Multiple response tableswere used to assess the frequencies of hypogonadism, varico-cele, cryptorchidism, prostatitis, or hernia in the patients.
We further tested the possible dependence of several vari-ables: sperm count, progressive sperm motility, gonadotropinlevel, serum testosterone, and serum prolactin level to HFEand TF genotypes, and different clinical characteristics (inde-pendent variables) such as the presence of hypogonadism, var-icocele, cryptorchidism, hernia, or prostatitis in the patients.The possible dependence was assessed using multiple for-ward stepwise regression analysis that enabled the identifica-tion of the independent variable(s) that highly correlate to themeasured variables. We only included 97 men with idiopathicinfertility in the separate multiple forward stepwise
n infertile men Vol. 91, No. 5, May 2009
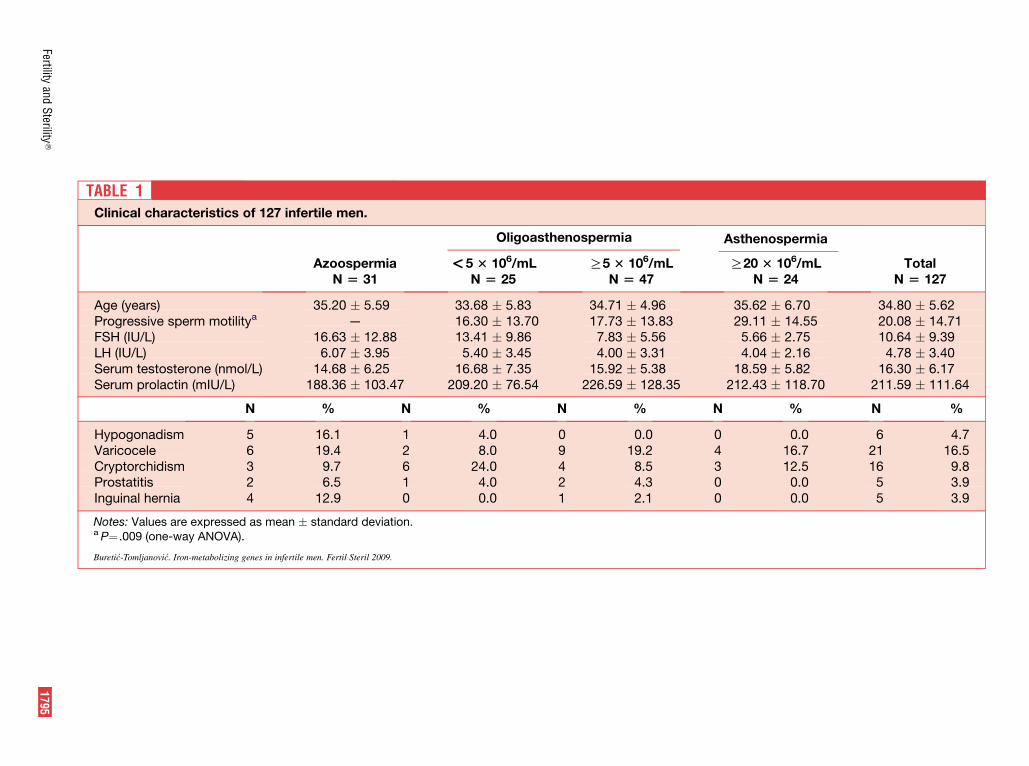
TABLE 1Clinical char
spermia Asthenospermia
R5 3 106/mL R20 3 106/mL TotalN [ 47 N [ 24 N [ 127
Age (years) 34.71 � 4.96 35.62 � 6.70 34.80 � 5.62Progressive s 17.73 � 13.83 29.11 � 14.55 20.08 � 14.71FSH (IU/L) 7.83 � 5.56 5.66 � 2.75 10.64 � 9.39LH (IU/L) 4.00 � 3.31 4.04 � 2.16 4.78 � 3.40Serum testos 15.92 � 5.38 18.59 � 5.82 16.30 � 6.17Serum prolac 226.59 � 128.35 212.43 � 118.70 211.59 � 111.64
% N % N %
Hypogonadis 0.0 0 0.0 6 4.7Varicocele 19.2 4 16.7 21 16.5Cryptorchidis 8.5 3 12.5 16 9.8Prostatitis 4.3 0 0.0 5 3.9Inguinal herni 2.1 0 0.0 5 3.9
Notes: Values aa P¼ .009 (one-
Bureti�c-Tomljanovi�c.
Fertilityand
Sterility
�1795
acteristics of 127 infertile men.
Oligoastheno
Azoospermia <5 3 106/mLN [ 31 N [ 25
35.20 � 5.59 33.68 � 5.83perm motilitya — 16.30 � 13.70
16.63 � 12.88 13.41 � 9.866.07 � 3.95 5.40 � 3.45
terone (nmol/L) 14.68 � 6.25 16.68 � 7.35tin (mIU/L) 188.36 � 103.47 209.20 � 76.54
N % N % N
m 5 16.1 1 4.0 06 19.4 2 8.0 9
m 3 9.7 6 24.0 42 6.5 1 4.0 2
a 4 12.9 0 0.0 1
re expressed as mean � standard deviation.way ANOVA).
Iron-metabolizing genes in infertile men. Fertil Steril 2009.
regression analysis to specifically assess the effects of HFEand/or TF genotype on sperm count, progressive sperm mo-tility, or serum hormonal concentrations.
For both multiple forward stepwise regression analyses,the statistical significance level was set at P<.05. Multipleforward stepwise regression analyses started with no predic-tor variables in the regression equation. Variables were addedin the subsequent steps according to defined criteria (F ¼ 4.0to enter; F¼ 1 to remove). In the first step, the predictor vari-able with the largest correlation with the dependent variableentered the regression equation. In the second or furthersteps, if possible, the predictor variable was selected basedon the highest partial correlation with the dependent variable.Partial coefficients of correlation were calculated to test thecorrelation between investigated genotypes and hormonalstatus controlling for age and sperm count.
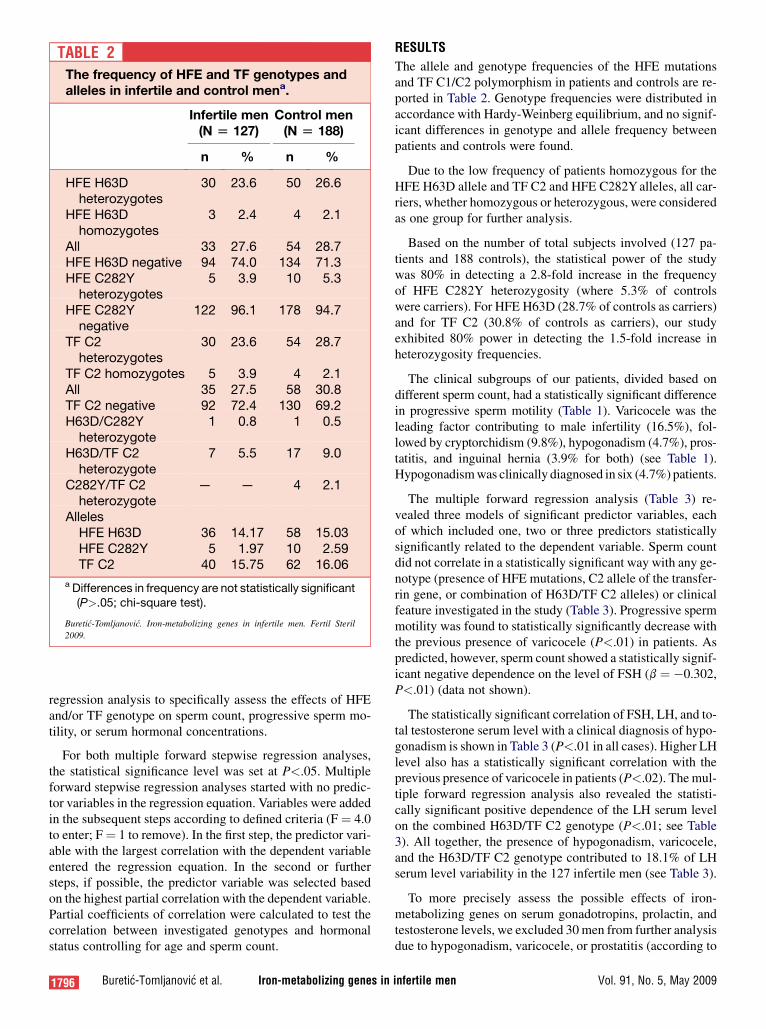
TABLE 2The frequency of HFE and TF genotypes andalleles in infertile and control mena.
Infertile men(N [ 127)
Control men(N [ 188)
n % n %
HFE H63Dheterozygotes
30 23.6 50 26.6
HFE H63Dhomozygotes
3 2.4 4 2.1
All 33 27.6 54 28.7HFE H63D negative 94 74.0 134 71.3HFE C282Y
heterozygotes5 3.9 10 5.3
HFE C282Ynegative
122 96.1 178 94.7
TF C2heterozygotes
30 23.6 54 28.7
TF C2 homozygotes 5 3.9 4 2.1All 35 27.5 58 30.8TF C2 negative 92 72.4 130 69.2H63D/C282Y
heterozygote1 0.8 1 0.5
H63D/TF C2heterozygote
7 5.5 17 9.0
C282Y/TF C2heterozygote
— — 4 2.1
AllelesHFE H63D 36 14.17 58 15.03HFE C282Y 5 1.97 10 2.59TF C2 40 15.75 62 16.06
a Differences in frequency are not statistically significant(P>.05; chi-square test).
Bureti�c-Tomljanovi�c. Iron-metabolizing genes in infertile men. Fertil Steril
2009.
1796 Bureti�c-Tomljanovi�c et al. Iron-metabolizing genes in
RESULTS
The allele and genotype frequencies of the HFE mutationsand TF C1/C2 polymorphism in patients and controls are re-ported in Table 2. Genotype frequencies were distributed inaccordance with Hardy-Weinberg equilibrium, and no signif-icant differences in genotype and allele frequency betweenpatients and controls were found.
Due to the low frequency of patients homozygous for theHFE H63D allele and TF C2 and HFE C282Y alleles, all car-riers, whether homozygous or heterozygous, were consideredas one group for further analysis.
Based on the number of total subjects involved (127 pa-tients and 188 controls), the statistical power of the studywas 80% in detecting a 2.8-fold increase in the frequencyof HFE C282Y heterozygosity (where 5.3% of controlswere carriers). For HFE H63D (28.7% of controls as carriers)and for TF C2 (30.8% of controls as carriers), our studyexhibited 80% power in detecting the 1.5-fold increase inheterozygosity frequencies.
The clinical subgroups of our patients, divided based ondifferent sperm count, had a statistically significant differencein progressive sperm motility (Table 1). Varicocele was theleading factor contributing to male infertility (16.5%), fol-lowed by cryptorchidism (9.8%), hypogonadism (4.7%), pros-tatitis, and inguinal hernia (3.9% for both) (see Table 1).Hypogonadism was clinically diagnosed in six (4.7%) patients.
The multiple forward regression analysis (Table 3) re-vealed three models of significant predictor variables, eachof which included one, two or three predictors statisticallysignificantly related to the dependent variable. Sperm countdid not correlate in a statistically significant way with any ge-notype (presence of HFE mutations, C2 allele of the transfer-rin gene, or combination of H63D/TF C2 alleles) or clinicalfeature investigated in the study (Table 3). Progressive spermmotility was found to statistically significantly decrease withthe previous presence of varicocele (P<.01) in patients. Aspredicted, however, sperm count showed a statistically signif-icant negative dependence on the level of FSH (b ¼ �0.302,P<.01) (data not shown).
The statistically significant correlation of FSH, LH, and to-tal testosterone serum level with a clinical diagnosis of hypo-gonadism is shown in Table 3 (P<.01 in all cases). Higher LHlevel also has a statistically significant correlation with theprevious presence of varicocele in patients (P<.02). The mul-tiple forward regression analysis also revealed the statisti-cally significant positive dependence of the LH serum levelon the combined H63D/TF C2 genotype (P<.01; see Table3). All together, the presence of hypogonadism, varicocele,and the H63D/TF C2 genotype contributed to 18.1% of LHserum level variability in the 127 infertile men (see Table 3).
To more precisely assess the possible effects of iron-metabolizing genes on serum gonadotropins, prolactin, andtestosterone levels, we excluded 30 men from further analysisdue to hypogonadism, varicocele, or prostatitis (according to
infertile men Vol. 91, No. 5, May 2009
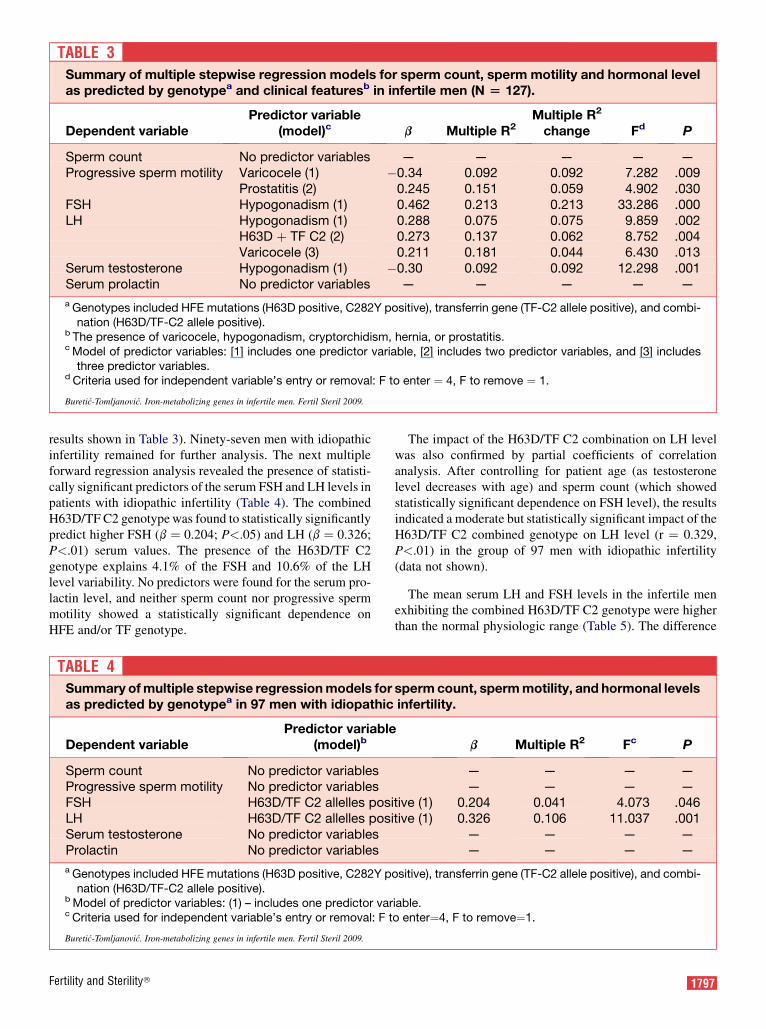
TABLE 3Summary of multiple stepwise regression models for sperm count, sperm motility and hormonal levelas predicted by genotypea and clinical featuresb in infertile men (N [ 127).
Dependent variablePredictor variable
(model)c b Multiple R2Multiple R2
change Fd P
Sperm count No predictor variables — — — — —Progressive sperm motility Varicocele (1) �0.34 0.092 0.092 7.282 .009
Prostatitis (2) 0.245 0.151 0.059 4.902 .030FSH Hypogonadism (1) 0.462 0.213 0.213 33.286 .000LH Hypogonadism (1) 0.288 0.075 0.075 9.859 .002
H63D þ TF C2 (2) 0.273 0.137 0.062 8.752 .004Varicocele (3) 0.211 0.181 0.044 6.430 .013
Serum testosterone Hypogonadism (1) �0.30 0.092 0.092 12.298 .001Serum prolactin No predictor variables — — — — —
a Genotypes included HFE mutations (H63D positive, C282Y positive), transferrin gene (TF-C2 allele positive), and combi-nation (H63D/TF-C2 allele positive).
b The presence of varicocele, hypogonadism, cryptorchidism, hernia, or prostatitis.c Model of predictor variables: [1] includes one predictor variable, [2] includes two predictor variables, and [3] includes
three predictor variables.d Criteria used for independent variable’s entry or removal: F to enter ¼ 4, F to remove ¼ 1.
Bureti�c-Tomljanovi�c. Iron-metabolizing genes in infertile men. Fertil Steril 2009.
results shown in Table 3). Ninety-seven men with idiopathicinfertility remained for further analysis. The next multipleforward regression analysis revealed the presence of statisti-cally significant predictors of the serum FSH and LH levels inpatients with idiopathic infertility (Table 4). The combinedH63D/TF C2 genotype was found to statistically significantlypredict higher FSH (b ¼ 0.204; P<.05) and LH (b ¼ 0.326;P<.01) serum values. The presence of the H63D/TF C2genotype explains 4.1% of the FSH and 10.6% of the LHlevel variability. No predictors were found for the serum pro-lactin level, and neither sperm count nor progressive spermmotility showed a statistically significant dependence onHFE and/or TF genotype.
Fertility and Sterility�
The impact of the H63D/TF C2 combination on LH levelwas also confirmed by partial coefficients of correlationanalysis. After controlling for patient age (as testosteronelevel decreases with age) and sperm count (which showedstatistically significant dependence on FSH level), the resultsindicated a moderate but statistically significant impact of theH63D/TF C2 combined genotype on LH level (r ¼ 0.329,P<.01) in the group of 97 men with idiopathic infertility(data not shown).
The mean serum LH and FSH levels in the infertile menexhibiting the combined H63D/TF C2 genotype were higherthan the normal physiologic range (Table 5). The difference
TABLE 4Summary of multiple stepwise regression models for sperm count, sperm motility, and hormonal levelsas predicted by genotypea in 97 men with idiopathic infertility.
Dependent variablePredictor variable
(model)b b Multiple R2 Fc P
Sperm count No predictor variables — — — —Progressive sperm motility No predictor variables — — — —FSH H63D/TF C2 allelles positive (1) 0.204 0.041 4.073 .046LH H63D/TF C2 allelles positive (1) 0.326 0.106 11.037 .001Serum testosterone No predictor variables — — — —Prolactin No predictor variables — — — —
a Genotypes included HFE mutations (H63D positive, C282Y positive), transferrin gene (TF-C2 allele positive), and combi-nation (H63D/TF-C2 allele positive).
b Model of predictor variables: (1) – includes one predictor variable.c Criteria used for independent variable’s entry or removal: F to enter¼4, F to remove¼1.
Bureti�c-Tomljanovi�c. Iron-metabolizing genes in infertile men. Fertil Steril 2009.
1797

TABLE 5LH and FSH serum levels according to HFE and TF genotype in 97 men with idiopathic infertilitya.
Genotype N LH (IU/L) FSH (IU/L)
H63D carriers 24 4.826 � 3.032 11.702 � 8.229H63D noncarriers 73 4.017 � 2.331 8.553 � 6.897TF C2 carriers 30 4.760 � 3.277 11.488 � 9.382TF C2 noncarriers 67 3.973 � 2.090 8.364 � 6.025H63D/TF C2 carriers 6 7.383 � 4.671b 15.100 � 10.038c
H63D/TF C2 noncarriers 91 4.008 � 2.212 8.956 � 7.029
a One-way ANOVA.b P¼ .001.c P¼ .046.
Bureti�c-Tomljanovi�c. Iron-metabolizing genes in infertile men. Fertil Steril 2009.
between biallelic carriers versus noncarriers was shown to bestatistically significant also using ANOVA (see Table 5).
The HFE mutation and TF C2 allelic frequencies were notstatistically significantly different in the group of 97 menwith idiopathic infertility compared with the 188 controlmen (data not shown).
DISCUSSION
We investigated the possible effects of variants in two genes af-fecting iron metabolism (HFE and transferrin gene) on malefertility. In a previous report, Peterlin et al. (13) did not findany implication of the HFE H63D and C282Y mutations inthe risk for male infertility in the Slovenian population. No sta-tistically significant differences were found in the frequenciesof HFE gene mutations between the Croatian and Sloveniangeneral populations (18). The HFE C282Y mutation is rarein both populations (3.3% and 4%, respectively), which is con-sistent with the northwest–southeast decline in frequency inEurope (18). The HFE H63D mutation is more prevalentwith an overall frequency of 14.5% in both populations (18).No statistically significant difference in the frequencies ofHFE mutations and TF alleles were observed between groupsof infertile and control fertile men. Therefore, our resultscorroborate the aforementioned findings and showed that ina Croatian population, neither the HFE mutation (H63D orC282Y) nor transferrin gene variants (C1/C2) directly affectmale fertility, with no statistically significant correlation tosperm count or progressive sperm motility variability. Further-more, the frequencies of the investigated transferrin variantsalso do not differ in a statistically significant manner betweeninfertile men and controls (see Table 2).
Multiple forward stepwise regression analysis that in-cluded the presence of HFE mutation, TF C2 allele, and com-bined H63D-TF C2 genotype as independent variables (seeTable 4) showed a statistically significant positive associationof the serum LH and FSH levels with the combined H63D/TFC2 genotype in the 97 men with idiopathic infertility. Partialcoefficients of correlation analysis further confirmed the in-terdependence of the combined HFE/TF genotype and LH
1798 Bureti�c-Tomljanovi�c et al. Iron-metabolizing genes in
level (r ¼ 0.329, P¼.002), but not FSH level. Therefore,our results showed the impact of iron-metabolizing genevariants on LH levels and, to a smaller extent, on FSH levels.These findings suggest that genes involved in iron metabo-lism may influence the function of the HPG axis.
The mechanisms of iron metabolism in brain are stillincompletely understood. Because iron metabolism was re-cently connected with signal transduction processes (19,20) and innate immunity (21), the synergistic action ofHFE mutations and the TF C2 gene variant might causechanges in downstream signalization processes and/or inter-fere with the inflammation response. A signaling pathwaythat includes iron sensing involves hepcidin synthesis in theliver. Hepcidin, a main regulator of whole-body iron homeo-stasis, is induced by HFE protein, which represents a part ofthe systemic iron sensor (20). Mutations in HFE interferewith hepcidin induction in cases of Tf saturation, whichmay contribute to iron misregulation, pro-oxidative stress,and anti-oxidative imbalance in the body. Pro-oxidative stressand anti-oxidative imbalance in the semen has also been re-lated to human male infertility (22).
The possible physiologic consequence of the variation inthe transferrin gene may affect few or many of the variousfunctions of the Tf protein described to date. Transferrin ispredominantly synthesized in the liver, but smaller amountsare also produced in the brain and Sertoli cells of the testes.After internalization of the Tf–TfR1 complex, transferrinlocalizes in the perinuclear position in the cytoplasm and sub-sequently releases iron. Tf is required for cell viability andproliferation, as shown in many cell culture systems, whileits deprivation from cells has been associated with elevatedindexes of apoptosis (23).
The involvement of the transferrin in many cellularprocesses underscores the importance of the iron metabolismregulation in different tissues and organs. In the brain, Tf func-tions in the delivery of iron through the blood–brain barrier,iron management (24), neuronal transport, and also neuromo-dulatory function (25). Transferrin was shown to be a majorproduct secreted by differentiated Sertoli cells in adult rat
infertile men Vol. 91, No. 5, May 2009

testes, while its secretion in fetal testis may serve as a markerfor Sertoli cell function and differentiation. Migrenne et al.(26) demonstrated that pituitary hormones differentially regu-late Sertoli cell function during fetal life and in the adult testis.In the fetus, pituitary hormones negatively regulate the activityof Sertoli cells, and this shifts to a positive regulation in theadult testis. That indicates that basal testicular transferrin se-cretion from the Sertoli cells decreases in the fetal rat testiswith the onset of pituitary hormone secretion. Sertoli cellsprovide the structural support and optimal environment forspermatogenesis, and their number in the testis is a key featureaffecting the number of mature spermatozoa formed in theadult testis (26). The role of FSH is well established in Sertolicell proliferation in the fetus and maintenance in the adult tes-tis (27). Further, FSH was shown to, through Sertoli cell func-tion, affect sperm motility and sperm shape characteristics.
Infertile males show impairment of both Leydig andSertoli cell function in parallel with their impaired spermato-genesis (28). Increased Tf secretion by the adult testis hasbeen shown to be associated with impaired spermatogenicfunction in men (29) and dogs (30, 31). Decreased Tf levelswere found in oligozoospermic men with varicocele (32,33). Levels of LH above the 97.5 percentile of the levels infertile men were found in 15% of infertile men (28). HigherLH content may stimulate Sertoli cell activity and highertransferrin release, as demonstrated in dogs with azoospermia(31). Sertoli cells are further known to control the testoster-one secretory function of the Leydig cells (34), probablythrough transferrin release. A positive feedback mechanismconnects the number of germ cells in the testis and transferrinsecretion from the Sertoli cells (35). Sertoli cells positivelycontrol testosterone secretion from Leydig cells, and a nega-tive feedback mechanism is well established between testos-terone released from the testis and LH synthesis and secretionfrom the anterior pituitary (36). The finding that Sertoli cellsof the adult rat testis did not increase FSH-stimulated trans-ferrin secretion argues in favor of the prediction of LH tobe an essential pituitary hormone required for transferrinsecretion by Sertoli cells in the adult testes. Abnormalitiesof LH pulsatile secretion may be associated with male infer-tility, and may cause disruption of the regulatory role of bothLeydig and Sertoli cells and disruption of spermatogenesis(37). Furthermore, seminal plasma levels of LH in oligozoo-spermic and normoasthenozoospermic men were several-fold higher than in serum.
The role of transferrin in the testes and iron deposits in thepituitary of patients with HH suggests an involvement of ironmetabolism in the maintenance of fertility status in men. Theeffect of the HFE and TF gene variants on serum LH and FSHconcentration in the group of infertile men as established inthis study supports this view. Our results showed the HFEH63D mutation and C2 allele of the transferrin gene to bethe interacting factors highly associated with increased serumLH level—that is, modulation of the hormone release in thepituitary or dysfunction of the testicular compartments dur-ing fetal/infant or adult life. In southern European popula-
Fertility and Sterility�
tions where the prevalence of the C282Y mutation is low,the H63D mutation may represent the genetic predisposingfactor acting in accordance with other genetic or environmen-tal factors in disrupting iron homeostasis. It is worth mention-ing that HFE and/or TF gene variants explain 10.6% and4.1% of LH and FSH variation in the group of men withidiopathic infertility (Table 4). This finding argues for theinvolvement of many other genes and/or factors influencinggonadotropic hormone regulation (38).
REFERENCES1. Andrews NC. Iron metabolism: iron deficiency and iron overload. Ann
Rev Genom Hum Genet 2000;1:75–98.
2. Parkkila S, Waheed A, Britton RS, Feder JN, Tsuchihashi Z, Schatzman RC,
et al. Immunohistochemistry of HLA-H, the protein defective in patients
with hereditary haemochromatosis, reveals unique pattern of expression
in gastrointestinal tract. Proc Natl Acad Sci USA 1997;94:2534.
3. Limdi JK, Crampton JR. Hereditary haemochromatosis. QJM 2004;97:
315–24.
4. O’Sullivan EP, Walsh CH. Endocrinopathy of HFE-related hemochroma-
tosis. Expert Rev Endocrinol Metab 2007;2:277–86.
5. Peillon F, Racadot J. Histopathological in the hypophysis in 6 cases of
hemochromatosis. Ann Endocrinol (Paris) 1969;30:800–7.
6. Bergeron C, Kovacs K. Pituitary siderosis a histologic, immunocyto-
logic, and ultrastructural study. Am J Pathol 1978;93:295–310.
7. McDermott JH, Walsh CH. Hypogonadism in hereditary hemochromato-
sis. J Clin Endocrinol Metab 2005;90:2451–5.
8. Okamoto CT. From genetics to cellular physiology. Focus on ‘‘regula-
tion of transferrin-induced endocytosis by wild-type and C282Y-mutant
HFE in transfected HeLa cells’’. Am J Physiol Cell Physiol 2002;282:
C971–2.
9. Merryweather-Clarke AT, Pointon JJ, Jouanolle AM, Rochette J,
Robson KJ. Geography of HFE C282Y and H63D mutations. Genet
Test 2000;4:183–98.
10. Wong CT, Saha N. Effects of transferrin genetic phenotypes on total iron-
binding capacity. Acta Haematol 1986;75:215–8.
11. Robson KJH, Lehmann DJ, Wimhurst VLC, Livesey KJ, Combrinck M,
Merryweather-Clarke AT, et al. Synergy between the C2 alelle of
transferrin and C282Y allele of the haemochromatosis gene (HFE) as
risk factors for developing Alzheimer’s disease. J Med Genet
2004;41:261–5.
12. Hayflick SJ. Neurodegeneration with brain iron accumulation: from
genes to pathogenesis. Semin Pediatr Neurol 2006;13:182–5.
13. Peterlin B, Kunej T, Hruskovicov�a H, Ferk P, Gersak K, Zorn B. Analysis
of the hemochromatosis mutations C282Y and H63D in infertile men.
Fertil Steril 2006;86:1796–8.
14. Simoni M, Bakker E, Eurlings MC, Matthijs G, Moro E, Muller CR, et al.
Laboratory guidelines for molecular diagnosis of Y-chromosomal micro-
deletions. European Molecular Genetics Quality Network (EMQN),
2001;1–7.
15. World Health Organisation. Laboratory manual for the examination of
human semen and semen-cervical mucus interaction. 4th ed. Cambridge
University Press, Cambridge, United Kingdom, 1999.
16. Abraham GE, ed. Handbook of radioimmunoassay. Marcel Dekker,
1977.
17. Merryweather-Clarke AT, Pointon JJ, Shearman JD, Robson KJ. Global
prevalence of putative haemochromatosis mutations. J Med Genet
1997;34:275–8.
18. Risti�c S, Makuc J, Star�cevi�c N, Logar N, Brajenovi�c-Mili�c B, Stepec S,
et al. Hemochromatosis gene mutations in the Croatian and Slovenian
populations. Clin Genet 2003;64:444–6.
19. Anderson GJ, Frazer DM. Iron metabolism meets signal transduction.
Nat Genet 2006;38:503–4.
20. Goswami T, Andrews NC. Hereditary hemochromatosis protein, HFE,
interaction with transferrin receptor 2 suggests a molecular mechanism
for mammalian iron sensing. Biol Chem 2006;281:28494–8.
1799

21. Roy CN, Custodio AO, de Graaf J, Schneider S, Akpan I, Montross LK,
et al. An HFE-dependent pathway mediates hyposideremia in response to
lipopolysaccharide-induced inflammation in mice. Nat Genet 2004;36:
481–5.
22. Garrido N, Meseguer M, Simon C, Pellicer A, Remoh J. Pro-oxidative
and anti-oxidative imbalance in human semen and its relation with
male fertility. Asian J Androl 2004;6:59–65.
23. Weinzimer SA, Gibson TB, Collett-Solberg PF, Khare A, Liu B, Cohen P.
Transferrin is an insulin-like growth factor-binding protein-3 binding
protein. J Clin Endocrinol Metab 2001;86:1806–13.
24. Takeda A. Significance of transferrin in iron delivery to the brain.
J Health Sci 2001;47:520–4.
25. Hill JM, Ruff MR, Weber RJ, Pert CB. Transferrin receptors in rat brain:
neuropeptide-like pattern and relationship to iron distribution. Proc Natl
Acad Sci USA 1985;82:4553–7.
26. Migrenne S, Chryst�ele Racine, Guillou F, Habert R. Pituitary hormones
inhibit the function and differentiation of fetal Sertoli cells. Endocrinol-
ogy 2003;144:2617–22.
27. Grover A, Smith CE, Gregory M, Cyr DG, Sairam MR, Hermo L. Effects
of FSH receptor deletion on epididymal tubules and sperm morphology,
numbers, and motility. Mol Reprod Dev 2005;72:135–44.
28. Andersson AM, Jørgensen N, Frydelund-Larsen L, Raypert-De Meyts E,
Skakkebæk NE. Impaired Leydig cell function in infertile men: a study
of 357 idiopathic infertile men and 318 proven fertile controls. J Clin
Endocrinol Metab 2004;89:3161–7.
29. Anaplitoou ML, Goulandris N, Douvara R. Seminal fibronectin-like
antigen and transferrin concentrations in infertile and fertile men. Andro-
logia 1995;27:137–42.
1800 Bureti�c-Tomljanovi�c et al. Iron-metabolizing genes i
30. Kawakami E, Hori T, Tsutsui T. Azoospermia of dogs with apoptotic
germ cells and Leydig cells. J Vet Med Sci 2000;62:529–31.
31. Kawakami E, Hori T, Tsutsui T. Relationship between testicular transfer-
rin and plasma estradiol-17beta concentrations of dogs with azoospermia
and dogs with Sertoli cell tumors. Vet Med Sci 2001;63:579–81.
32. Yoshida KI, Nakame Y, Uchijima Y. Seminal plasma transferrin concen-
tration in normozoospermic fertile men and oligozoospermic men asso-
ciated with varicocele. Int J Fertil 1988;33:432–6.
33. Kosar A, Sarica K, Ozdiler E. Effect of varicocelectomy on seminal
plasma transferrin values: a comparative clinical trial. Andrologia
2000;32:19–22.
34. Wu N, Murono EP. Temperature and germ cell regulation of Leydig cell
proliferation stimulated by Sertoli cell-secreted mitogenic factor: a possi-
ble role in cryptorchidism. Andrologia 1996;28:247–57.
35. Roberts KP, Santulli R, Seiden J, Zirkin BR. The effect of testosterone
and subsequent germ cell depletion on transferrin and sulfated glycopro-
tein-2 messenger ribonucleic acid levels in the adult rat testis. Biol
Reprod 1992;47:92–6.
36. Tang K, Bartke A, Gardiner CS, Wagner TE, Yun JS. Testosterone feed-
back on gonadotropin secretion and gene expression in transgenic mice
expressing human growth hormone gene. J Androl 1994;15:9–14.
37. Pryor JL, Krieg RJ Jr, Evans WS, Batson J, Turner TT. Luteinizing
hormone secretion by male rat pituitary cells perifused in vitro: effect
of experimental left varicocele and orchiectomy. J Androl 1989;10:
372–7.
38. Dohle GR, Jungwirth A, Colpi G, Givercman A, Diemer T,
Hargreave TB. Guidelines on male infertility. European Association of
Urology 2007;7–70.
n infertile men Vol. 91, No. 5, May 2009




















