
Length of Androgen Receptor–CAG Repeats in Fertile and Infertile Egyptian Men
10
Length of Androgen Receptor–CAG Repeats in Fertile and Infertile Egyptian Men WAEL A. BADRAN,* IBRAHIM FAHMY,*{ WAEL M. ABDEL-MEGID,{ KAY ELDER,§ RAGAA MANSOUR,* AND MARIJO KENT-FIRST{ From *The Egyptian In Vitro Fertilization and Embryo Transfer Centre, Hadayk El-Maadi, Cairo, Egypt; ÀAndrology Department, Faculty of Medicine, Cairo University, Cairo, Egypt; `Obstetrics & Gynecology Unit, University of Wisconsin Medical School, Madison, Wisconsin; and §Bourn Hall Clinic, Cambridge, United Kingdom. ABSTRACT: Androgens play key roles in spermatogenesis, and they exert their effect via the androgen receptor (AR). The AR gene has a repetitive DNA sequence in exon 1 that encodes a polyglutamine tract. Instability in the glutamine (CAG) repeat unit length is polymorphic across ethnic groups. Previous studies of the relationship between the repeat unit length and male infertility have been contradictory. To establish the range of wild-type alleles in Egyptian men, we determined the range of repeat lengths in a population of normally fertile, ethnically selected Egyptian men. We also investigated the association between trinucleotide repeat length within the AR gene and male factor infertility in a population of ethnically selected Egyptian infertile men, who were compared with fertile, ethnic group–matched and age-matched controls. The study included 129 clinically selected infertile Egyptian men who were scheduled for intracytoplasmic sperm injection and 52 ethnically matched fertile controls. The experimental population was grouped according to sperm counts ranging from nonobstructive azoosper- mia to normozoospermia. The CAG repeat N-terminal domain region of the AR gene was amplified in peripheral blood DNA, and allele size was determined by fragment analysis. Allele size and single-nucleotide polymorphism and mutation rates were deter- mined by sequencing individual amplified alleles. The mean CAG repeat length in the azoospermia group was 18.55 6 2.0; in the severe oligozoospermia group it was 18.21 6 3.42; in the oligozoospermia group it was 18.27 6 2.93; and in the infertile with normal sperm count group it was 17.72 6 2.0. In the control group, the mean CAG repeat length was 18.18 6 3.63. No significant correlation was found between CAG repeat length and the risk of male factor infertility in an ethnically defined population of Egyptian men. However, a significant and positive correlation between CAG repeat length and serum testosterone concentration was demonstrated. This suggests the involvement of epigenetic regulation linked to this region. Key words: Male infertility, spermatogenesis, trinucleotide re- peat. J Androl 2009;30:416–425 A ndrogens control development of the male pheno- type, including spermatogenesis (Yong et al, 2003). The actions of androgens are mediated by the androgen receptor (AR), a member of the steroid hormone receptor subfamily of nuclear receptors (Molenda et al, 2003; Verrijdt et al, 2006). The AR gene mutations are linked to several pathologic conditions (Gottlieb, 2005; Gottlieb et al, 2005). Mutations that cause total disruptions of AR function result in complete androgen insensitivity syndrome (Gobinet et al, 2002). On the other hand, mutations that do not result in complete ablation of AR function can cause a wide spectrum of milder phenotypes ranging from ambiguous genitalia in the newborn to a normal male phenotype associated with infertility (Hiort and Holterhus, 2003). The AR gene is a single-copy gene located in the pericentric region of the long arm of the X chromosome, at locus Xq11-12 in the human (Brown et al, 1989). The AR gene contains 8 exons. Toward the 59 end of exon 1 there is a polymorphic glutamine (CAG) repeat that encodes a polyglutamine tract. This microsatellite locus is the most variable region in the AR gene. In a healthy individual, the number of CAG repeats ranges from 9 to 36, with an average of 21–22 repeats. The average number of repeat lengths varies significantly between different racial groups (Quigley et al, 1995). For example, individuals of African descent have much shorter average repeat lengths of CAG (18–20) com- pared with Hispanics (CAG, 23). Studies of CAG repeat lengths in whites and East Asians have reported the repeat lengths to be 21–22 and 23, respectively. Moreover, there are about 20 different allele lengths reported across ethnic groups, and 90% of women are heterozygous for the length of the repeat (Sartor et al, 1999; Bennett et al, 2002). Spinal and bulbar muscular atrophy (SBMA; Online Mendelian Inheritance in Man no. 313200), also known Correspondence to: Dr Marijo Kent-First, Department of Biological Sciences, Box GY, Mississippi State University, Mississippi State, MS 39672 (e-mail: [email protected]). Received for publication May 24, 2008; accepted for publication January 21, 2009. DOI: 10.2164/jandrol.108.005843 Journal of Andrology, Vol. 30, No. 4, July/August 2009 Copyright E American Society of Andrology 416
-
Upload
dsm-academy -
Category
Documents
-
view
5 -
download
0
Transcript of Length of Androgen Receptor–CAG Repeats in Fertile and Infertile Egyptian Men

Length of Androgen Receptor–CAG Repeats in Fertile andInfertile Egyptian Men
WAEL A. BADRAN,* IBRAHIM FAHMY,*{ WAEL M. ABDEL-MEGID,{ KAY ELDER,§
RAGAA MANSOUR,* AND MARIJO KENT-FIRST{
From *The Egyptian In Vitro Fertilization and Embryo Transfer Centre, Hadayk El-Maadi, Cairo, Egypt; �Andrology
Department, Faculty of Medicine, Cairo University, Cairo, Egypt; `Obstetrics & Gynecology Unit, University of
Wisconsin Medical School, Madison, Wisconsin; and §Bourn Hall Clinic, Cambridge, United Kingdom.
ABSTRACT: Androgens play key roles in spermatogenesis, and
they exert their effect via the androgen receptor (AR). The AR gene
has a repetitive DNA sequence in exon 1 that encodes a
polyglutamine tract. Instability in the glutamine (CAG) repeat unit
length is polymorphic across ethnic groups. Previous studies of the
relationship between the repeat unit length and male infertility have
been contradictory. To establish the range of wild-type alleles in
Egyptian men, we determined the range of repeat lengths in a
population of normally fertile, ethnically selected Egyptian men. We
also investigated the association between trinucleotide repeat length
within the AR gene and male factor infertility in a population of
ethnically selected Egyptian infertile men, who were compared with
fertile, ethnic group–matched and age-matched controls. The study
included 129 clinically selected infertile Egyptian men who were
scheduled for intracytoplasmic sperm injection and 52 ethnically
matched fertile controls. The experimental population was grouped
according to sperm counts ranging from nonobstructive azoosper-
mia to normozoospermia. The CAG repeat N-terminal domain
region of the AR gene was amplified in peripheral blood DNA, and
allele size was determined by fragment analysis. Allele size and
single-nucleotide polymorphism and mutation rates were deter-
mined by sequencing individual amplified alleles. The mean CAG
repeat length in the azoospermia group was 18.55 6 2.0; in the
severe oligozoospermia group it was 18.21 6 3.42; in the
oligozoospermia group it was 18.27 6 2.93; and in the infertile
with normal sperm count group it was 17.72 6 2.0. In the control
group, the mean CAG repeat length was 18.18 6 3.63. No
significant correlation was found between CAG repeat length and
the risk of male factor infertility in an ethnically defined population
of Egyptian men. However, a significant and positive correlation
between CAG repeat length and serum testosterone concentration
was demonstrated. This suggests the involvement of epigenetic
regulation linked to this region.
Key words: Male infertility, spermatogenesis, trinucleotide re-
peat.
J Androl 2009;30:416–425
Androgens control development of the male pheno-
type, including spermatogenesis (Yong et al, 2003).
The actions of androgens are mediated by the androgen
receptor (AR), a member of the steroid hormone
receptor subfamily of nuclear receptors (Molenda et
al, 2003; Verrijdt et al, 2006). The AR gene mutations
are linked to several pathologic conditions (Gottlieb,
2005; Gottlieb et al, 2005). Mutations that cause total
disruptions of AR function result in complete androgen
insensitivity syndrome (Gobinet et al, 2002). On the
other hand, mutations that do not result in complete
ablation of AR function can cause a wide spectrum of
milder phenotypes ranging from ambiguous genitalia in
the newborn to a normal male phenotype associated
with infertility (Hiort and Holterhus, 2003).
The AR gene is a single-copy gene located in the
pericentric region of the long arm of the X chromosome,
at locus Xq11-12 in the human (Brown et al, 1989). The
AR gene contains 8 exons. Toward the 59 end of exon 1
there is a polymorphic glutamine (CAG) repeat that
encodes a polyglutamine tract. This microsatellite locus
is the most variable region in the AR gene. In a healthy
individual, the number of CAG repeats ranges from 9 to
36, with an average of 21–22 repeats. The average
number of repeat lengths varies significantly between
different racial groups (Quigley et al, 1995). For
example, individuals of African descent have much
shorter average repeat lengths of CAG (18–20) com-
pared with Hispanics (CAG, 23). Studies of CAG repeat
lengths in whites and East Asians have reported the
repeat lengths to be 21–22 and 23, respectively.
Moreover, there are about 20 different allele lengths
reported across ethnic groups, and 90% of women are
heterozygous for the length of the repeat (Sartor et al,
1999; Bennett et al, 2002).
Spinal and bulbar muscular atrophy (SBMA; Online
Mendelian Inheritance in Man no. 313200), also known
Correspondence to: Dr Marijo Kent-First, Department of Biological
Sciences, Box GY, Mississippi State University, Mississippi State, MS
39672 (e-mail: [email protected]).
Received for publication May 24, 2008; accepted for publication
January 21, 2009.
DOI: 10.2164/jandrol.108.005843
Journal of Andrology, Vol. 30, No. 4, July/August 2009Copyright E American Society of Andrology
416

as Kennedy disease, is linked to an abnormal elongation
of more than 40 CAG repeats in exon 1 of the AR gene
(Quigley et al, 1995). SBMA is a neurodegenerativedisease characterized by progressive weakness and
atrophy of proximal muscles. Most affected men display
progressive androgen insensitivity in the form of
progressive testicular atrophy, gynecomastia, feminized
skin changes, and reduced fertility, which are either due
to azoospermia or severe oligozoospermia (Greenland
and Zajac, 2004).
Many studies have examined the possible correlationbetween the length of the polyglutamine repeat in the
AR gene and male factor infertility (Table 1). A number
of these reports suggested a link between male factor
infertility and expansion of the polymorphic trinucleo-
tide (CAG) repeat in the AR gene (Mifsud et al, 2001;
Patrizio et al, 2001; Madgar et al, 2002). Other reports
failed to demonstrate such an association (Giwercman et
al, 1998; Dadze et al, 2000; Tufan et al, 2005). The basisfor these investigations is the finding that the length of
the polyglutamine repeat is inversely proportional to the
degree of normal functionality of the AR (Kazemi-
Esfarjani et al, 1995). This observation led to the
hypothesis that significantly longer or significantly
contracted polyglutamine tracts should be considered
as a risk factor for male infertility. To test this
hypothesis in any population, it is critical that the rangeof alleles present in ethnically matched (relative to the
experimental population) fertile control men be deter-
mined (Zitzmann and Nieschlag, 2003). Unfortunately,
data interpretations in many studies are confusing
because of the general inherent instability of microsat-
ellite loci and the specific selective instability of loci
containing trinucleotide CAG repeats. Furthermore,
given fluid migration patterns in human populations,the wide variation in repeat lengths within an ethnically
mixed population(s) and the overlapping ranges between
defined ethnic groups make it difficult to assess data
interpretation and comparisons of data between ethnic
groups (Hiort and Holterhus, 2003).
The present study was designed to test the hypothesis
that significant deviation from the mean CAG repeat
length in the AR gene is correlated with male factor
infertility. To test this hypothesis, first the relativestability (including the range of allele sizes) of the AR-
CAG repeat length in an ethnically defined and
population of fertile Egyptian men with normal semen
parameters was determined. Second, the relationship of
the AR-CAG repeat length relative to male factor
infertility in an ethnically defined population of
Egyptian men who were intracytoplasmic sperm injec-
tion (ICSI) candidates was determined. An ethnicallyselected wild-type population of fertile Egyptian men
served as the control group in this study.
Materials and Methods
Subjects
A total of 129 infertile men presenting consecutively who were
scheduled for the ICSI program at the Egyptian In Vitro
Fertilization and Embryo Transfer Centre were prospectively
recruited for the study during the period from 2004 through
2005. Patients with obstructive azoospermia, hypogonadotro-
phic hypogonadism, genital tract infection, and genetic or
karyotype abnormalities were excluded from the study.
Infertile couples with definite female factor infertility were
also excluded. The control subjects were fertile volunteers and
patients attending urology clinics for complaints other than
infertility. Fifty-two men were recruited, all with proven
fertility, having fathered at least one child by natural
conception. These men had no previous history of infertility
or fertility treatment and had no identified genetic disorders.
All cases and controls were identified as Egyptians according
to self-categorization. The geographic regions of descent for all
subjects in the control and experimental populations were
derived from the Nile Delta and Upper Egypt, both along the
Nile River.
This study was conducted with institutional review board
approval in The Egyptian In Vitro Fertilization and Embryo
Transfer Centre and in the United States (Promega, Madison,
Wisconsin). Each participant (patients and controls) gave
written informed consent. The samples were made anonymous
throughout the study.
Clinical Evaluation
Each patient was subjected to history taking and clinical
examination. Determination of testicular volume was per-
formed by Orchidometer (Holtain Ltd, Crymych, United
Kingdom). Laboratory investigations included: semen analysis
according to World Health Organization (1999) guidelines,
measurement of serum levels of reproductive hormones
(follicle-stimulating hormone [FSH], luteinizing hormone
[LH], testosterone, and prolactin), and cytogenetic analysis.
The patient population was restricted according to the status
of the female partner’s age (younger than 40 years), normal
basal FSH level, no polycystic ovary syndrome, and normal
uterus with no hydrosalpinges detected by ultrasonography.
For each of the control subjects, at least one semen analysis
was performed, as well as clinical examination. For each
participant in the experimental and control groups, a
peripheral blood sample was collected for DNA analysis of
sequence of CAG repeat length within the AR gene. None of
the patients received infertility treatment during the 3 months
prior to these investigations
Molecular Analysis
Allele Sizing and Trinucleotide Repeat Allele Fragment
Analysis—DNA was purified from the peripheral blood of
patients and control subjects using a Wizard Genomic DNA
Purification System (Promega) and DNA IQ (Promega)
according to the manufacturer’s recommended protocol. The
CAG repeat within the N-terminal domain region was
Badran et al N AR-CAG in Fertile and Infertile Egyptians 417

Table
1.
Sum
mary
of
pre
vio
usly
publis
hed
stu
die
sin
vestigating
AR
-CA
Gre
peat
length
infe
rtile
and
infe
rtile
popula
tions
Sourc
e(L
ocatio
nof
Stu
dy)
Infe
rtile
Contr
ol
Sig
nific
ant
Diffe
rence
Betw
een
2G
roups
(P)
CA
GR
epeat
Cuto
ffP
oin
t
No.
of
Men,
n
Mean
6S
DR
ange
No.
of
Men,
nM
ean
6S
DR
ange
Azo
osperm
iaO
ligosperm
iaT
ota
l
Giw
erc
man,
1998
(Sw
eden)
617
33
21.9
16–27
294
23.2
8–30
No
n/a
Legiu
set
al,
1999
(Belg
ium
)22
201
223
21
(media
n)
15–30
181
21
(media
n)
14–29
Yes
$26
Dow
sin
get
al,
1999
(Austr
alia
)10
20
30
23.2
60.7
15–34
32
20.5
60.3
17–25
Yes
(.0001)
$26
Hio
rtet
al,
1999
(Germ
any)
13
167
180
23
13–30
53
24
17–39
No
n/a
Dadze
et
al,
2000
(Germ
any)
18
101
119
22
63.2
16–34
22
20.8
63.3
15–26
No
n/a
Patr
izio
et
al,
2001
(Unite
dS
tate
s)
16
53
69
23.5
63.4
18–39
45
22
62.8
12–30
Yes
(.03)
n/a
Walle
rand
et
al,
2001
(Fra
nce)
37
...
37
23.9
60.5
13–28
50
22.2
60.4
17–27
Yes
(.008)
n/a
Mifsud
et
al,
2001
(Unite
dS
tate
s)
23
72
95
21.9
56
0.3
114–31
55
20.7
26
0.5
28–27
Yes
(.034)
$26
Von
Eck
ard
ste
in,
2001
(Germ
any)
29
14
43
20.4
62.9
17–27
131
20
63.1
12–28
No
n/a
Rajp
ert
-De
Meyt
set
al,
2002
(Denm
ark
)30
89
119
21.5
62.9
15–29
110
21.8
63.4
14–33
No
n/a
Kukuvitis
et
al,
2002
(Gre
ece
)42
67
109
21
(media
n)
15–33
64
22
(media
n)
14–31
No
n/a
Van
Gold
eet
al,
2002
(The
Neth
erlands)
75
...
75
22.2
63.1
13–36
70
21.7
63.4
15–31
No
n/a
Era
sm
uson
et
al,
2003
(New
Zeala
nd)
...
105
105
21.4
66
0.3
12–30
93
20.9
96
0.2
89–26
No
n/a
Lund
et
al,
2003
(Fin
land)
...
192
192
22.1
66
2.8
16–30
149
22.4
62.8
15–29
No
n/a
Asa
tianiet
al,
2003
(Germ
any)
...
217
217
21.5
62.9
17–30
131
20
63.1
12–28
Yes
(.001)
n/a
Mengualet
al,
2003
(Spain
)102
...
102
23.2
56
2.7
18–32
96
22.4
622.8
15–34
Yes
(.033)
n/a
Ruhayelet
al,
2004
(Sw
eden)
36
49
85
...
14–29
223
...
12–30
No
n/a
Tufa
net
al,
2005
(Turk
ey)
25
22
47
22.2
86
0.3
718–29
32
22.4
16
0.5
416–29
No
n/a
Lavery
et
al,
2005
(Ire
land)
55
11
66
23.3
06
2.4
419–30
77
23.0
76
2.3
219–30
No
n/a
Kom
ori
et
al,
1999
(Japan)
...
59
59
21.2
64.2
14–32
36
21.4
63.5
16–21
Yes
(,.0
5)
,16
Yoshid
aet
al,
1999
(Japan)
41
...
41
26.5
63.5
20–34
48
23.9
62.9
17–30
Yes
(.0013)
31–40
Sasagaw
aet
al,
2001
(Japan)
30
...
30
23.4
62.9
19–30
51
23.7
63.2
17–28
No
n/a
Nakabaya
shiet
al,
2003
(Japan)
...
...
133
22.6
63
12–34
100
23.3
62.8
16–31
No
n/a
Tut
et
al,
1997
(Chin
a)
28
125
153
...
...
72
...
...
Yes
$27
Mifsud
et
al,
2001
(Chin
a)
33
87
120
23.8
26
0.4
14–33
87
22.3
86
0.3
211–29
Yes
(.043)
$26
Liet
al,
2005
(Chin
a)
31
52
...
22.1
6..
...
...
...
.Y
es
n/a
Thangara
jet
al,
2002
(India
)280
...
280
21.7
60.1
812–32
201
22.4
60.1
912–32
No
n/a
Dhill
on
et
al,
2003
(India
)103
80
183
22.2
61.5
13–33
59
21.4
61.4
12–33
No
n/a
Pan
et
al,
2002
(Taiw
an)
48
...
48
23
60.6
10–41
47
21
60.4
14–27
Yes
(.001)
.27
Tse
et
al,
2003
(Hong
Kong)
53
32
85
23
63.9
14–36
45
23
63.1
16–30
No
,16
to.
30
Kata
giriet
al,
2006
(United
Sta
tes)
757
64
22.2
63
17–32
13
19.3
65
10–24
Yes
(.02)
n/a
Madgar
et
al,
2002
(Isra
el)
46
15
61
18.6
63
13–24
50
16.6
62.6
11–22
Yes
(.003)
n/a
Hadjk
acem
et
al,
2004
(Tunis
ia)
27
38
129
20.8
60.2
712–28
98
21.1
36
0.3
114–29
No
n/a
West
erv
eld
et
al,
2008
(The
Neth
erlands)
...
...
700
21.5
63.1
...
...
...
...
No
n/a
Pre
sent
stu
dy
(Egypt)
44
74
118
+11
a18.2
96
36–26
52
18.1
86
3.6
38–26
No
n/a
Abbre
via
tions:
...,
an
absence
of
data
for
that
stu
dy;
n/a
,not
applic
able
.a
Ele
ven
infe
rtile
patients
with
norm
alsperm
count.
418 Journal of Andrology N July �August 2009

amplified using 2 flanking sets of primers: A(F) 59-TCC AGA
ATC TGT TCC AGA GCG TGC-39 and A1(R) 59-GCT GTG
AAG GTT GCT GTT CCT CAT-39. In set 1, the forward
primer, A(F), was fluorescently labeled and paired with the
unlabeled high-performance liquid chromatography (HPLC)–
purified reverse primer, A1(R). In set 2, the unlabeled HPLC–
purified forward primer was paired with a fluorescently labeled
reverse A1(R) primer. A total of 120 ng of DNA was amplified
in duplicate experiments designed to establish and to confirm
allele size. The PCR-amplified products were also sequenced as
needed to confirm CAG repeat length and polymorphism.
Conditions for large-pool PCR were as follows: primers were
diluted to 500 mM in a 40-mL total reaction volume consisting
of 4 mL of STAR buffer (Promega) and AmpliTaq Gold DNA
polymerase (0.5 mL/reaction; Applied Biosystems, Foster City,
California) PCR was performed using the ABI GeneAmp PCR
system 9600 (Perkin Elmer, Norwalk, Connecticut) according
to the Hot Start cycle protocol. The Hot Start cycle was 95uCfor 9 minutes, followed by 95u denaturation for 1 minute, 60uannealing for 1 minute, and 72u extension for 1 minute for 40
cycles, followed by a final extension at 72u for 5 minutes. The
products were separated by capillary electrophoresis on ABI
PRISM 3100 Genetic Analyzers with allele sizing using ILS-
600TM, 60–600 bp (Promega). PCR controls used in amplifica-
tion experiments included water, male DNA, and female DNA.
Fragment Size Confirmation and SNP Genotyping—PCR
products from a total of 75 individuals (40 samples from the
experimental group and 35 samples from the control group)
were gel purified according to standard methods. Both the
forward and reverse strands were sequenced to confirm the
length of the CAG repeat and to identify SNPs within the
region immediately flanking the CAG repeat.
Statistical Analysis
Statistical analysis was performed using the statistical package
SPSS for Windows (version 11.0; SPSS Inc, Chicago, Illinois).
The difference in the CAG repeat length between control,
combined infertile group, and various subgroups of infertile
men was evaluated by the Mann-Whitney U test. Spearman’s
correlation coefficient was used for bivariate regression
analysis to determine the correlation between CAG repeat
length, sperm count, bitesticular volume, and hormone data. A
probability of P , .05 was considered statistically significant.
Results
Clinical Findings
This study included 129 infertile, ethnically restricted
patients scheduled for an ICSI program. The group
included 44 patients with nonobstructive azoospermia
(absence of sperm in the ejaculate confirmed by
examination of sediment after centrifugation), 43
patients with severe oligozoospermia (sperm count
,5 million/mL), 31 patients with oligozoospermia
(sperm count range from 5 million/mL to less than
20 million/mL), and 11 patients with normal sperm
counts and varying degrees of asthenozoospermia/
teratozoospermia (sperm count $20 million/mL). A
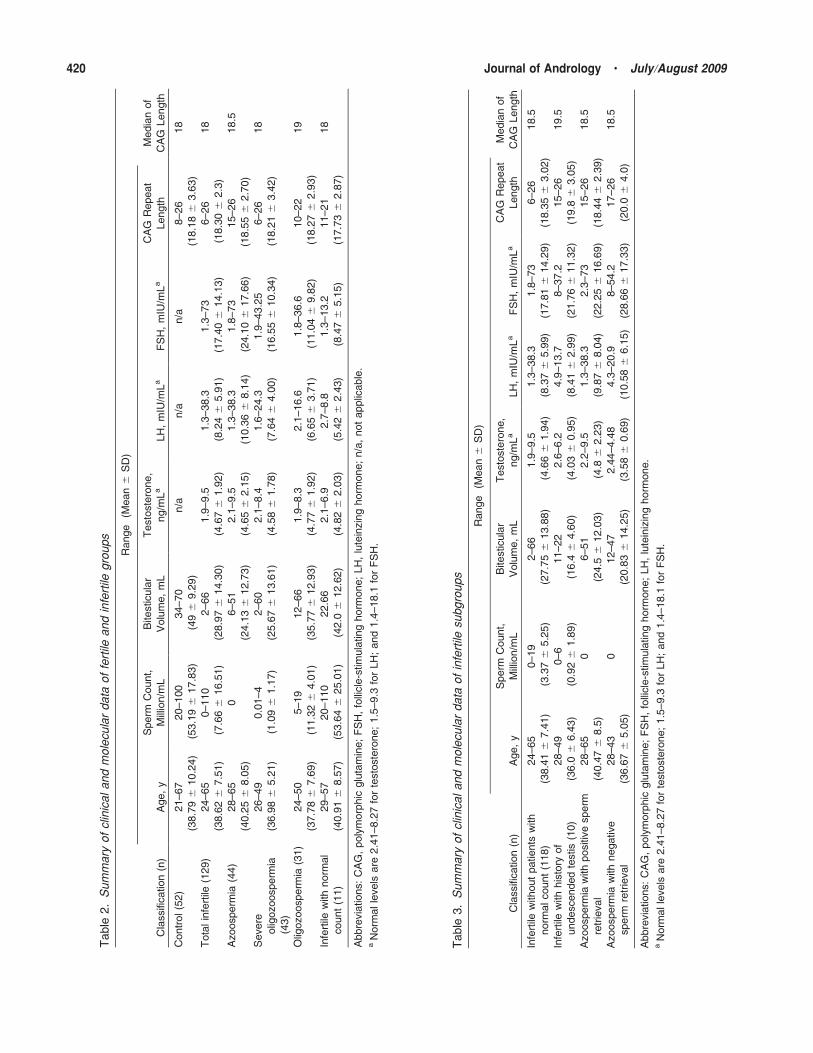
summary of the clinical and molecular data is shown
in Table 2. The control group consisted of 52 fertile men
with fertility demonstrated by live births and sperm
counts ranging from 20 million/mL to 100 million/mL
(mean, 53.19 6 17.83 million/mL). The ethnically
selected control population was matched with the
experimental population according to both ethnicity
and age. In the combined infertile group, excluding
infertile patients with normal sperm counts, the sperm
concentrations ranged between 0 and 19 million/mL
(mean, 3.37 6 5.25 million/mL; Table 3). Bitesticular
volume in the infertile group ranged between 2 and
66 mL (mean, 28.97 6 14.3 mL), whereas in the control
group the range was between 34 and 70 mL (mean, 49 6
9.29 mL), which was significantly different (P , .05).
The sperm count was significantly correlated with
bitesticular volume (P 5 .01) and serum levels of FSH
(P 5 .001), LH (P 5 .001), and testosterone (P 5 .04).
High serum FSH was detected in 40 patients from the
experimental population, and an isolated high serum
FSH was detected in 18 infertile patients. High serum
LH was detected in 28 patients. Combined high serum
LH and serum testosterone were detected in only 3
patients. These 3 patients had sperm counts of 0
(azoospermia), 0.5 million/mL, and 6 million/mL, re-
spectively, and the lengths of their mean CAG repeats
were 19, 19, and 21, respectively. A total of 38 patients
(29.5%) had undergone bilateral or left varicocelectomy.
A total of 10 patients (7.8%) had an undescended testis.
One patient had a nonmalignant testicular tumor with
subsequent left orchidectomy. Parents’ consanguinity
was reported by 26 patients (20.2%), whereas another 9
patients (7%) had a history of infertility in first- or
second-degree relatives.
Molecular Findings: Length of AR-CAG Repeats
The means and ranges of CAG repeat lengths in control
and different infertile groups are summarized in Table 2.
The mean CAG repeat length did not differ statistically
between the combined infertile group (18.30 6 2.3) and
the control group (18.18 6 3.63). When infertile men
with normal sperm count were excluded, the mean CAG
repeat length of the remaining group (18.35 6 3.02) did
not differ significantly from that of the control group
(Table 3). The mean CAG repeat length did not differ
statistically between the control group and each of the
infertile subgroups in the experimental population.
A total of 40 patients with azoospermia were
subjected to testicular sperm extraction (TESE) as a
part of their ICSI schedule (another 4 patients were
scheduled for ICSI/TESE and dropped out before the
Badran et al N AR-CAG in Fertile and Infertile Egyptians 419
Table
2.
Sum
mary
of
clin
icaland
mole
cula
rdata
of
fert
ileand
infe
rtile
gro
ups
Range
(Mean
6S
D)
Cla
ssifi
cation
(n)
Age,
y
Sperm
Count,
Mill
ion
/mL
Bitestic
ula
r
Volu
me,
mL
Testo
ste
rone,
ng
/mL
aLH
,m
IU/m
La
FS
H,
mIU
/mL
aC
AG
Repeat
Length
Media
nof
CA
GLength
Contr
ol(5
2)
21–67
(38.7
96
10.2
4)
20–100
(53.1
96
17.8
3)
34–70
(49
69.2
9)
n/a
n/a
n/a
8–26
(18.1
86
3.6
3)
18
Tota
lin
fert
ile(1
29)
24–65
(38.6
26
7.5
1)
0–110
(7.6
66
16.5
1)
2–66
(28.9
76
14.3
0)
1.9
–9.5
(4.6
76
1.9
2)
1.3
–38.3
(8.2
46
5.9
1)
1.3
–73
(17.4
06
14.1
3)
6–26
(18.3
06
2.3
)
18
Azo
osperm
ia(4
4)
28–65
(40.2
56
8.0
5)
06–51
(24.1
36
12.7
3)
2.1
–9.5
(4.6
56
2.1
5)
1.3
–38.3
(10.3
66
8.1
4)
1.8
–73
(24.1
06
17.6
6)
15–26
(18.5
56
2.7
0)
18.5
Severe
olig
ozoosperm
ia
(43)
26–49
(36.9
86
5.2
1)
0.0
1–4
(1.0
96
1.1
7)
2–60
(25.6
76
13.6
1)
2.1
–8.4
(4.5
86
1.7
8)
1.6
–24.3
(7.6
46
4.0
0)
1.9
–43.2
5
(16.5
56
10.3
4)
6–26
(18.2
16
3.4
2)
18
Olig
ozoosp
erm
ia(3
1)
24–50
(37.7
86
7.6
9)
5–19
(11.3
26
4.0
1)
12–66
(35.7
76
12.9
3)
1.9
–8.3
(4.7
76
1.9
2)
2.1
–16.6
(6.6
56
3.7
1)
1.8
–36.6
(11.0
46
9.8
2)
10–22
(18.2
76
2.9
3)
19
Infe
rtile
with
norm
al
count
(11)
29–57
(40.9
16
8.5
7)
20–110
(53.6
46
25.0
1)
22.6
6
(42.0
612.6
2)
2.1
–6.9
(4.8
26
2.0
3)
2.7
–8.8
(5.4
26
2.4
3)
1.3
–13.2
(8.4
76
5.1
5)
11–21
(17.7
36
2.8
7)
18
Abbre
via
tions:
CA
G,
poly
morp
hic
glu
tam
ine;
FS
H,
folli
cle
-stim
ula
ting
horm
one;
LH
,lu
tein
zin
ghorm
one;
n/a
,not
applic
able
.a
Norm
alle
vels
are
2.4
1–8.2
7fo
rte
sto
ste
rone;
1.5
–9.3
for
LH
;and
1.4
–18.1
for
FS
H.
Table
3.
Sum
mary
of
clin
icaland
mole
cula
rdata
of
infe
rtile
subgro
ups
Range
(Mean
6S
D)
Cla
ssifi
cation
(n)
Age,
y
Sperm
Count,
Mill
ion
/mL
Bite
sticula
r
Volu
me,
mL
Testo
ste
rone,
ng
/mL
aLH
,m
IU/m
La
FS
H,
mIU
/mL
aC
AG
Repeat
Length
Media
nof
CA
GLength
Infe
rtile
without
patients
with
norm
alcount
(118)
24–65
(38.4
16
7.4
1)
0–19
(3.3
76
5.2
5)
2–66
(27.7
56
13.8
8)
1.9
–9.5
(4.6
66
1.9
4)
1.3
–38.3
(8.3
76
5.9
9)
1.8
–73
(17.8
16
14.2
9)
6–26
(18.3
56
3.0
2)
18.5
Infe
rtile
with
his
tory
of
undesce
nded
testis
(10)
28–49
(36.0
66.4
3)
0–6
(0.9
26
1.8
9)
11–22
(16.4
64.6
0)
2.6
–6.2
(4.0
36
0.9
5)
4.9
–13.7
(8.4
16
2.9
9)
8–37.2
(21.7
66
11.3
2)
15–26
(19.8
63.0
5)
19.5
Azoosperm
iaw
ith
positi
ve
sperm
retr
ieval
28–65
(40.4
76
8.5
)
06–51
(24.5
612.0
3)
2.2
–9.5
(4.8
62.2
3)
1.3
–38.3
(9.8
76
8.0
4)
2.3
–73
(22.2
56
16.6
9)
15–26
(18.4
46
2.3
9)
18.5
Azoosperm
iaw
ith
negative
sperm
retr
ieval
28–43
(36.6
76
5.0
5)
012–47
(20.8
36
14.2
5)
2.4
4–4.4
8
(3.5
86
0.6
9)
4.3
–20.9
(10.5
86
6.1
5)
8–54.2
(28.6
66
17.3
3)
17–26
(20.0
64.0
)
18.5
Abbre
via
tions:
CA
G,
poly
morp
hic
glu
tam
ine;
FS
H,
folli
cle
-stim
ula
ting
horm
one;
LH
,lu
tein
izin
ghorm
one.
aN
orm
alle
vels
are
2.4
1–8.2
7fo
rte
sto
ste
rone;
1.5
–9.3
for
LH
;and
1.4
–18.1
for
FS
H.
420 Journal of Andrology N July �August 2009

procedure). Sperm retrieval was successful in 34
patients. The mean length of CAG repeat did not reveal
a significant difference between azoospermic patients
with positive sperm retrieval and those with negative
sperm retrieval (Table 3).
There was a mild positive correlation between length
of CAG repeat and serum testosterone level. This
correlation was detected in the combined infertile group(P 5 .014, r 5 0.330) and the combined infertile group
after exclusion of patients with normal sperm counts (P
5 .027, r 5 0.312), and the correlation was strongly
positive in infertile men with normal sperm counts (P 5
.005, r 5 0.975), but the correlation was not detected in
the other infertile subgroups (azoospermia, severe
oligozoospermia, and oligzoospermia). However, the
strong positive correlation detected in infertile men withnormal sperm count may be affected by the small
number of this subgroup (11 patients).
None of the semen parameters or the other hormone
parameters was significantly correlated with length of
CAG repeat. On the other hand, there was no
correlation between length of CAG repeat and bitestic-
ular volume or sperm count in the fertile control group.
The distribution of CAG repeat lengths in the control
group and experimental infertile group is shown in theFigure. The CAG repeat lengths in the control, total
infertile group, and infertile subgroups overlap consid-
erably. The longest allele (26 repeats) was detected in 3
individuals: 1 person from the control group, 1 patient
with azoospermia, and 1 patient with severe oligozoo-spermia. The shortest allele (6 repeats) was detected in 1
patient with severe oligozoospermia.
In the combined infertile group, the most common
alleles were 17 and 19 repeats. Each of these respective
alleles was detected in 20 patients (15.5%). In the control
group, the most common allele (18 repeats) was detected
in 9 individuals (17.3%). No sequence variation or
polymorphism was observed in alleles isolated from 75individuals.
Sequence analysis of PCR products confirmed repeat
length in all of the sampled alleles.
Discussion
The present study examines the association between
expanded CAG repeat length within the AR gene and
defective spermatogenesis in 129 infertile and 52 control
men of matched Egyptian origin. The first experiment
established the common alleles present in an ethnically
restricted population of normally fertile men. This
population then served as the control for the clinicallyselected experimental population. There was no corre-
lation between CAG repeat length and defective
Figure. Distribution of CAG repeat length in normal fertile control and infertile Egyptian men. (A) Combined cases and control (number of men ingroup, 181; mean, 18.26 6 3.18; median, 18). (B) Control (number of men in group, 52; mean, 18.18 6 3.63; median, 18). (C) Combinedinfertile (number of men in group, 129; mean, 18.3 6 2.3). (D) Infertile after excluding patients with normal sperm count (number of men ingroup, 118; mean, 18.35 6 3.02; median, 18.5). (E) Azoospermia (number of men in group, 44; mean, 18.55 6 2.7; median, 18.5). (F)Oligozoospermia (number of men in group, 31; mean, 18.27 6 2.93; median, 19). (G) Severe oligozoospermia (number of men in group, 43;mean, 18.21 6 3.42; median, 18). (H) Infertile with normal sperm count (number of men in group, 11; mean, 17.73 6 2.87; median, 18).
Badran et al N AR-CAG in Fertile and Infertile Egyptians 421

spermatogenesis. The mean numbers of repeats in the
infertile population and fertile controls were similar
(18.30 6 2.30 and 18.18 6 3.63, respectively).The current study is the first to examine this
correlation in a defined Egyptian population, and this
adds to the data available for different ethnic back-
grounds. The mean CAG repeat length for the
combined control and infertile groups was 18.24 6
2.96, with a range of 6–26 repeats. Although several
studies have investigated the proposed association, these
reports have yielded conflicting results (Table 1). Theobserved variations in the results from previous studies
stem from several factors: 1) ethnically diverse popula-
tions; 2) the studied infertile men may represent a
heterogeneous group with respect to the causes of
infertility and may be under the effect of different
genetic mutations or even epigenetic phenomena; and 3)
different inclusion criteria in each study. The previously
studied infertile populations included various categoriesof infertility (eg, patients with varicocele, genital tract
obstruction, and/or genital tract infection) and semen
parameters (azoospermia, oligozoospermia, severe oli-
gozoospermia, or infertile men with normal sperm count
associated with varying degrees of asthenozoospermia/
teratozoospermia; Kukuvitis et al, 2002; Asatiani et al,
2003; Hadjkacem et al, 2004). Most importantly, the
control groups in many of these previous studies werenot well matched in terms of ethnicity and age. The
control groups in these studies often included not only
individuals with proven fertility, but also individuals
with normal sperm count but not proven fertility and/or
individuals from unselected populations (Legius et al,
1999; Erasmuson et al, 2003; Lund et al, 2003).
Unfortunately, these different reports render the
comparison between various studies inconclusive. Be-cause of racial variation and the impact of ethnicity, it is
possible that the results for AR-CAG repeat length in
selectively restricted racial/ethnic populations outweigh
those for a mixed population (Asatiani et al, 2003).
Some biomedical scientists assume that racial/ethnic
categorizations are inadequate descriptors of the distri-
bution of genetic variation in humans (Schwartz, 2001;
Hega and Venter, 2003). However, studies of different
populations have documented genetic, and therefore,biological differences among races (Keita et al, 2004).
Genetic variation tends to be geographically structured
in accordance with the classic common racial classifica-
tion, which is based on continental ancestry: African,
white, Asian, Pacific Islander, and Native American
(Jorde and Wooding, 2004; Tishkoff and Kidd, 2004). It
could be argued that population clusters identified by
genotype analysis seem to be more informative thanthose identified by self-declaration of race (Wilson et al,
2001). However, Risch et al (2002) demonstrated that
from both an objective and scientific (genetic and
epidemiologic) perspective, there is great validity in
racial/ethnic self-categorizations.The present work studied only an ethnically defined
population; all subjects were Egyptian by self-declara-
tion. Egypt’s geographic location has prompted its
population’s genetic diversity, but despite this, the
Egyptian population seems to be fairly homogenous.
This is supported by the findings of genetic studies that
characterize the Egyptian population. Studies of mito-
chondrial DNA variation, the Y-chromosome genepool, and different allelic variants in the modern
Egyptian reflect a mixture of European, Middle Eastern,
and African characteristics (Manni et al, 2002; Hamdy
et al, 2003). In addition, there is still a homogenous
genetic tie to ancient Egyptians (Kring et al, 1999). On
the other hand, these studies distinguished genetic
characteristics of the Egyptian from those of sub-
Saharan populations (Luis et al, 2004). This supportsthe concept that human populations are seldom
demarcated by precise genetic boundaries, and substan-
tial overlap can occur (Risch et al, 2002). The Egyptian
population, although fairly homogenous, is no excep-
tion in this regard.
A recent comprehensive meta-analysis included a
summary of 33 reports. This study investigated the
source of CAG variation between published results
(Davis-Dao et al, 2007). The report indicated that caseand control definitions likely influenced study results as
an important determinant of differences in repeat length
between cases and controls. In the present work, in
accordance with the recommendation of that meta-
analysis, we considered stringency of case and control
definitions. On the other hand, the analysis of the full set
of 33 reports revealed statistically significant longer
CAG repeat length among cases compared with controlpopulations. In an attempt to avoid methodologic
concerns, particularly small sample size and unmatched
case-control, Westerveld et al (2008) investigated AR-
CAG repeat in 700 men who presented for fertility
workup with varying degrees of semen quality. The
study concluded that there was no correlation between
CAG repeat length expansion in the AR gene and semen
quality. However, the study included an experimentalpopulation of men from mixed ethnic origins with no
matched control population. Exon 1 of the AR gene
contains another island of polymorphic microsatellite
GGC repeats. It was proposed that certain combina-
tions of CAG and GGC repeats may confer a risk of
infertility to the carriers. However, previous studies
showed that there was no association between AR
CAG/GGN microsatellites and impaired spermatogen-esis (Ruhayel et al, 2004; Rajender et al, 2006; Saare et
al, 2007).
422 Journal of Andrology N July �August 2009

In the present study, there was a mild positive
correlation between CAG repeat length and the level
of serum testosterone in 3 of the subgroups of theinfertile patients studied (combined infertile, infertile
excluding patients with normal sperm count, and
patients with normal sperm count; but not in azoosper-
mia, severe oligospermia, or oligspermia groups).
Although some studies found no significant association
between AR-CAG and hormonal levels, particularly
testosterone (Alevizaki et al, 2003; Harkonen et al,
2003), one study reported strikingly higher level of LHand androgen sensitivity index (ASI) in azoospermic
patients with short CAG repeats (Tse et al, 2003). Other
reports are in agreement with our findings and show a
positive correlation between AR-CAG repeats and
serum levels of LH, free testosterone, and ASI (Giwerc-
man et al, 2004). This hormone profile is often reported
in men with a range of clinical phenotypes associated
with aberrant secondary sex characteristics. Lim et al(2000) reported longer AR-CAG repeats to be associ-
ated with moderate to severe undermasculinized genita-
lia in XY men. In addition, Canale et al (2005)
investigated CAG repeat length in 3 groups: control,
infertile, and hypoandrogenized subjects. They conclud-
ed that hypoandrogenic traits, such as hypoplasia of the
prostate and seminal vesicles, and reduced beard and
body hair, are associated with longer CAG repeats.However, they found no difference in the mean of CAG
repeats in infertile populations compared with fertile
control populations.
In the present work, the secondary sexual characteristics
of infertile groups were comparable to those of controls.
Because there was no difference between the CAG repeat
length in the 2 different groups (infertile vs control), the
proposed effect of CAG repeat length on testosteronefunctionality may be limited to a subclinical level.
Thus, it might be assumed, at least in our studied
population, that the AR modulates its transactivation
potential via its N-terminal polyglutamine tract without
being associated with any pathologically apparent
disease (pathologic phenotype; Vogt, 1999).
In conclusion, our study established the range of
alleles in an ethnically and geographically restricted
population of Egyptian men with normal fertility. Therewas no significant correlation between CAG repeat
length and risk of male infertility in our ethnically
restricted experimental population compared with the
matched control population. Therefore, polymorphism
detected in the polyglutamine-rich region of the AR
gene is not a useful genetic indication of male factor
infertility. A significant positive correlation between
CAG repeat length and serum testosterone concentra-tion was detected, and this suggests that epigenetic
regulation linked to this region is involved.
ReferencesAlevizaki M, Cimponeriu AT, Garofallaki M, Sarika HL, Alevizaki
CC, Papamichel C, Philippou G, Anastasiou EA, Lekakis JP,
Mavrikakis M. The androgen receptor gene CAG polymorphism is
associated with the severity of coronary artery disease in men. Clin
Endocrinol. 2003;59:749–755.
Asatiani K, Von Eckardstein S, Simoni M, Gromoli J, Nieschlag E.
CAG repeat length in the androgen receptor gene affects the risk of
male infetility. Int J Androl. 2003;26:255–261.
Bennett CI, Price DK, Kim S, Liu D, Jovanovice BD, Nathan D,
Johnson ME, Montgomery JS, Cude K, Brockbank JC, Sartor O,
Figg WD. Racial variation in CAG repeat length within the
androgen receptor gene among prostate cancer patients of lower
socioecomomic status. J Clin Oncol. 2002;20:3599–3604.
Brown CJ, Goss SJ, Lubahn DB, Joseph DR, Wilson EM, French FS,
Willard HF. Androgen receptor locus on the human X-chromo-
some: regional localization to Xp11-12 and description of a DNA
polymorphism. Am J Hum Genet. 1989;44:264–269.
Canale D, Caglieresi C, Moschini C, Liberati CD, Macchia E,
Pinchera A, Martino E. Androgen receptor polymorphism (CAG
repeats) and androgenicity. Clin Endocrinol. 2005;63:356–361.
Dadze S, Wieland C, Jakubiczka S, Funke K, Schroder E, Royer-
Pokora B, Willers R, Wieacker P. The size of the CAG repeat in
exon 1 of the androgen receptor gene shows no significant
relationship to impaired spermatogenesis in an infertile Caucasian
sample of German origin. Mol Hum Reprod. 2000;6:207–214.
Davis-Dao CA, Tuazon ED, Sokol RZ, Cortessis VK. Male infertility
and variation in CAG repeat length in the androgen receptor gene:
a meta-analysis. J Clin Endocrin Metab. 2007;92(11):4319–4326.
Dhillon VS, Husain SA. Cytogentic and molecular analysis of the Y
chromosome: absence of a significant relationship between CAG
repeat length in exon 1 of the androgen receptor gene and infertility
in Indian men. Int J Androl. 2003;26:286–295.
Dowsing AT, Yong EL, Clark M, Mclachlan RI, de Kretser DM,
Trounson A. Linkage between male infertility and trinucleotids
repeat expansion in the androgen receptor gene. Lancet.
1999;354:640–643.
Erasmuson T, Sin IL, Sin YT. Absence of association of androgen
receptor trinucleotide expansion and poor semen quality.
Int J Androl. 2003;26:46–51.
Giwercman Y, Richthoff J, Lilja H, Anderberg C, Abrahamsson P,
Giwercman A. Androgen receptor CAG repeat length correlates
with semen PSA level in adolesence. Prostate. 2004;59:227–233.
Giwercman YL, Xu C, Arver S, Pousette A, Reneland R. No
association between the androgen receptor gene CAG repeat and
impaired sperm production in Swedish men. Clin Genet.
1998;54:435–436.
Gobinet J, Poujol N, Sultan C. Molecular action of androgens. Mol
Cell Endocrinol. 2002;198:15–24.
Gottlieb B. The androgen receptor gene mutation database. Montreal,
QC, Canada: McGill University; April 4, 2005. http://www.
mcgill.ca/androgendb. Accessed April 27, 2008.
Gottlieb B, Lombroso R, Beitel LK, Trifiro MA. Molecular pathology
of the androgen receptor in male (in)fertility. Reprod Biomed
Online. 2005;10:42–48.
Greenland KJ, Zajac JD. Kennedy’s disease: pathogenesis and clinical
approaches. Intern Med J. 2004;34:279–286.
Hadjkacem L, Hadj-Kacem H, Boulila A, Bahloul A, Ayadi H,
Ammar-Keskes L. Androgen receptor gene CAG repeats length in
fertile and infertile Tunisian men. Ann Genet. 2004;47:217–224.
Hamdy S, Hiratsuka M, Narahara K, Endo N, El-Enany M, Moursi
N, Ahmed M, Mizugaki M. Gentotype and allele frequencies of
Badran et al N AR-CAG in Fertile and Infertile Egyptians 423

TPMT, NAT2, GST, SULTIAI and MDR-I in the Egyptian
population. Br J Clin Pharmacol. 2003;55(6):560–569.
Harkonen K, Hhtaniemi I, Makinen J, Hubler D, Irjala K, Koskenvo M,
Oettel M, Raitakari O, Saad F, Pollanen P. The polymorphic androgen
receptor gene CAG repeat, pituitary-testicular function and andro-
pausal symptomes in ageing men. Int J Androl. 2003;26:187–194.
Hega SB, Venter JC. Genetics. FDA races in wrong direction. Science.
2003;301:466.
Hiort O, Holterhus PM. Androgen insensitivity and male infertility.
Int J Androl. 2003;26:16–20.
Jorde LB, Wooding SP. Genetic variation, classification and race. Nat
Genet. 2004;36:S28–S33.
Katagiri Y, Neri QV, Takeuchi T, Moy F, Sills ES, Palermo GD.
Androgen receptor CAG polymorphism (Xq11-12) status and
human spermatogenesis: a prospective analysis of infertile males
and their offspring conceived by intracytoplasmic sperm injection.
Int J Mol Med. 2006;18(3):405–413.
Kazemi-Esfarjani P, Trifiro MA, Pinsky L. Evidence for a repressive
function of the long polyglutamine tract in the human androgen
receptor: possible pathogenetic relevance for the (CAG)n-expanded
neuronopathies. Hum Mol Genet. 1995;4:523–537.
Keita SOY, Kitles RA, Royal CD, Bonney GE, Furbert-Harris P,
Dunston GM, Rotimi CN. Conceptualizing human variation. Nat
Genet. 2004;36:S17–S20.
Komori S, Kasumi H, Kanazawa R, Sakata K, Nakata Y, Kato H,
Kovama K. CAG repeat length in the androgen receptor gene of
infertile Japanese males with oligozoospermia. Mol Hum Reprod.
1999;5:14–16.
Kring M, Salem A, Bauer K, Geisert H, Malek A, Chaix L, Simon C,
Welsby D, Di-Rienzo A, Utermann G. mtDNA analysis of Nile
River valley population: a genetic corridor or a barrier to
migration? Am J Hum Genet. 1999;64:1166–1176.
Kukuvitis M, Gerorgiou I, Bouba I, Tsirka A, Ciannouli C, Yapijakis
C, Traiatzis B, Bontis J, Lolis D, Sofikitis N, Papadimas J.
Association of oestrogen receptor polymorphisms and androgen
receptor CAG trinucleotide repeats with male infertility: a study in
109 Greek infertile men. Int J Androl. 2002;25:149–152.
Lavery R, Houghton JA, Nolan A, Glennon M, Egan D, Maher M.
CAG repeat length in an infertile male population of Irish origin.
Genetica. 2005;123:295–302.
Legius E, Vanderschueren D, Spiessens C, D’Hooghe T, Matthijs G.
Association between CAG repeat number in the androgen receptor
and male infertility in a Belgian study. Clin Genet. 1999;56:166–167.
Lim H, Chen H, McBride S, Dunning A, Nixon R, Hughes I, Hawkins
J. Longer polyglutamine tracts in the androgen receptor are
associated with moderate to severe undermasculinized genitalia in
XY males. Hum Mol Genet. 2000;9:829–834.
Luis J, Rowold D, Regueiro M, Caeiro B, Cinnioglu C, Roseman C,
Underhill P, Cavalli-Sforza L, Herrera R. The Levant versus the
Horn of Africa: evidence for biderictional corridor of human
migration. Am J Hum Genet. 2004;74:532–544.
Lund A, Tapanainen JS, Lahdetie J, Savontaus M, Attomaki K. Long
CAG repeats in the AR gene are not associated with infertility in
Finnish males. Acta Obstet Gynecol Scand. 2003;82:162–166.
Madgar I, Green L, Kent-First M, Weissenberg R, Gershoni-Baruch
R, Goldman B, Friedman E. Genotyping of Israeli infertile men
with idiopathic oligozoospermia. Clin Genet. 2002;62:203–207.
Manni F, Leonardi P, Barakat A, Rouba H, Heyer E, Klintschar M,
Mcelreavey K, Quintana-Murci L. Y-chromosome analysis in
Egypt suggests a genetic regional continuity in Northeastern
Africa. Hum Biol. 2002;74:645–658.
Mengual L, Oriola J, Ascaso C, Oliva R. An increase CAG repeat
length in the androgen receptor gene in azoospermic ICSI
candidates. J Androl. 2003;24:279–284.
Mifsud A, Sim KS, Boettger-Tong H, Moreira S, Lamb DJ, Lipshultz
I, Yong EL. Trinucleotide (CAG) repeat polymorphisms in the
androgen receptor gene: molecular markers of risk for male
infertility. Fertil Steril. 2001;75:275–281.
Molenda H, Kilts C, Allen R, Tetel M. Nuclear receptor coactivator
funuction in reproductive physiology and behavior. Biol Reprod.
2003;69:1449–1457.
Nakabayashi A, Sueoka K, Matsuda N, Asada H, Tanigaki R, Sato K,
Tajima H, Ogata T, Kuji N, Yoshimura Y. Incidental deviation of
short and long CAG repeats in the androgen receptor gene for
Japanese male inferility. Reprod Med Biol. 2003;2:145–150.
Pan H, Li YY, Li TC, Tsai WT, Li SY, Hsiao KM. Increased (CTG/
CAG)n lengths in myotonic dystrophy type 1 and Machado-Joseph
disease genes in idiopathic azoospermia patients. Hum Reprod.
2002;17:1578–1583.
Patrizio P, Leonard DB, Chen K, Hernandez-Ayup S, Trounson A.
Larger trinucleotide repeat size in the androgen receptor gene of
infertile men with extremely severe oligozoospermia. J Androl.
2001;22:444–448.
Quigley CA, De Bellis A, Marscheke KB, El-Awady MK, Wilson EM,
French FS. Androgen receptor defects: historical, clinical, and
molecular perspectives. Endocr Rev. 1995;16:271–317.
Rajender S, Rajani V, Gupta N, Chakravarty B, Singh L, Thangaraj K.
No association of androgen receptor GGN repeat length polymor-
phism with infertility in Indian men. J Androl. 2006;27:785–789.
Rajpert-De Meyts ER, Leffers H, Petersen JH, Anderson AG, Carlsen
E, Jorgensen N, Skakkebaek NE. CAG repeat length in androgen
receptor gene and reproductive variables in fertile and infertile
men. Lancet. 2002;359:44–46.
Risch N, Burchard E, Ziv E, Tang H. Categorization of humans in
biomedical research: genes, race and disease. Genome Biol.
2002;3:1–12.
Ruhayel Y, Lundin KB, Giwercman Y, Hallden C, Willen M,
Giwercman A. Androgen receptor gene GGN and CAG polymor-
phisms among severely oligozoospermic and azoospermic Swedish
men. Hum Reprod. 2004;19:2076–2083.
Saare M, Belousova A, Punab M, Peters M, Haller K, Ausmees K,
Poolamets O, Karro H, Metspalu A, Salumets A. Androgen
receptor gene haplotype is associated with male infertility.
Int J Androl. 2007;31:395–402.
Sartor O, Zheng Q, Eastham JA. Androgen receptor gene CAG repeat
length varies in a race-specific fashion in men without prostate
cancer. Urology. 1999;53:378–380.
Sasagawa I, Suzuki Y, Ashida J, Nakada T, Muroya K, Ogata T.
CAG repeat length analysis and mutation screening of the
androgen receptor gene in japanese man with idiopathic azoosper-
mia. J Androl. 2001;22:804–808.
Schwartz RS. Racial profiling in medical research. N Engl J Med.
2001;344:1392–1393.
Thangaraj K, Joshi MB, Reddy AG, Gupta NJ, Chakravarty B, Singh
L. CAG repeat expansion in the androgen receptor gene is not
associated with male infertility in Indian populations. J Androl.
2002;23:815–818.
Tishkoff SA, Kidd KK. Implications of biogeography of human
populations for ‘race’ and medicine. Nat Genet. 2004;36:521–527.
Tse JYM, Liu VWS, Yeung WSB, Lau EYL, Ng EHY, Ho PC.
Molecular analysis of the androgen receptor gene in Hong Kong
Chinese infertile men. J Assist Reprod Genet. 2003;20:227–233.
Tufan A, Stiroglu-Tufan NL, Aydinuraz B, Stiroglu MH, Aydos K, Bagci
H. No association of the CAG repeat length in exon 1 of the androgen
receptor gene with idiopathic infertility in Turkish men: implications
and literature review. Tohoku J Exp Med. 2005;206:105–115.
Tut TG, Ghadessy FJ, Trifiro MA, Pinsky L, Yong EL. Long
polyglutamine tracts in the androgen receptor are assoicated with
424 Journal of Andrology N July �August 2009

reduced trans activation, impaired sperm production, and male
infertility. J Clin Endocrinol Metab. 1997;82:3777–3782.
Van Golde R, Van Houwelingen K, Kiemeney L, Kremer J, Tuerlings
J, Schalken J, Meuleman E. Is increased CAG repeat length in the
androgen receptor gene a risk factor for male subfertility? J Urol.
2002;167:621–623.
Verrijdt G, Tanner T, Moehren U, Gallewaert L, Haelens A, Claessens
F. The androgen receptor DNA-binding domain determines
androgen selectivity of transcriptional response. Biochem Soc
Trans. 2006;34:1089–1094.
Vogt PH. Risk of neurodegenerative diseases in children conceived by
intracytoplasmic sperm injection. Lancet. 1999;354:61.
Von Eckardstein S, Syska A, Gromoll J, Kamischke A, Simoni M,
Nieschlag E. Inverse correlation between sperm concentration and
number of androgen receptor CAG repeats in normal men. J Clin
Endocrinol Metab. 2001;86:2585–2590.
Wallerand H, Remy-Martin A, Chabannes E, Bermont L, Adessi G,
Bittard H. Relationship between expansion of the CAG Repeat in
exon 1 of the androgne receptor gene and idiopathic male
infertility. Fertil Steril. 2001;76:769–774.
Westerveld H, Visser L, Tanck M, van deer Veen F. CAG repeat
length variation in the androgen receptor gene is not associated
with spermatogenic failure. Fertil Steril. 2008;89:253–259.
Wilson JF, Weale ME, Smith AC, Gratrix F, Fletcher B, Thomas MF,
Bradman N, Goldstein DB. Population genetic structure of
variable drug response. Nat Genet. 2001;29:265–269.
World Health Organization. WHO Laboratory Manual for the Exam-
ination of Human Semen and Sperm-Cervical Mucus Interaction. 4th
ed. Cambridge, United Kingdom: Cambridge University Press; 1999.
Yong EL, Loy CJ, Sim KS. Androgen receptor gene and male
infertility. Hum Reprod Update. 2003;9:1–7.
Yoshida KI, Yano M, Chiba K, Honda M, Kitahara S. CAG repeat
length in the androgen receptor gene is enhanced in patients with
idiopathic azoospermia. Urology. 1999;54:1078–1081.
Zitzmann M, Nieschlag E. The CAG repeat polymorphism within
androgen receptor gene and maleness. Int J Androl. 2003;26:73–83.
Badran et al N AR-CAG in Fertile and Infertile Egyptians 425




















