
Leuprolide acetate reduces both in vivo and in vitro ovarian steroidogenesis in infertile women...
10
steroids 73 ( 2 0 0 8 ) 1475–1484 available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/steroids Leuprolide acetate reduces both in vivo and in vitro ovarian steroidogenesis in infertile women undergoing assisted reproduction Alcione L.A. Silva, Lauriane G.A. Abreu, Ana Carolina J.S. Rosa-e-Silva, Rui A. Ferriani, Marcos F. Silva-de-Sá ∗,1 Department of Gynecology and Obstetrics, Faculty of Medicine of Ribeirao Preto- University of Sao Paulo, Brazil article info Article history: Received 21 November 2007 Received in revised form 30 May 2008 Accepted 11 August 2008 Published on line 26 August 2008 Keywords: GnRH analogues Estradiol Progesterone Testosterone Androstenedione Luteinized granulosa cells abstract Despite the probable inhibitory effects of GnRH analogues on ovarian steroidogenesis in vitro, their association with assisted reproduction protocols shows favorable results. This suggests that there are important differences in the behaviors of these drugs when admin- istered in vivo versus in vitro. To clarify these differences, this study was designed to analyze the effect of leuprolide acetate (LA) on ovarian steroidogenesis in women undergoing In Vitro Fertilization (IVF). A prospective, randomized open label study was conducted on 14 women (26–35 years): seven receiving only gonadotrophins (Group 1) and seven receiv- ing gonadotrophin plus LA at 1mg/day (Group 2). The LA in vivo effect was determined with serum and follicular fluid (FF) samples and via luteinized granulosa cell cultivation (GCC), where cells were obtained during oocyte retrieval after ovarian hyperstimulation. In vitro analysis was performed via addition of LA to GCC only for Group 1 (without LA) at progressively higher concentrations (0, 10 −12 , 10 −9 and 10 −6 M). In vivo, the main observation was a reduction in androgen production in Group 2, represented by lower androstenedione production in FF (G1 = 6479 ± 3458; G2 = 3021 ± 1119 ng/ml; p = 0.04) and a lower testosterone peak in GC at 96 h (G1 = 0.64 ± 0.12 ng/ml; G2 = 0.50 ± 0.19ng/ml; P = 0.02), but a higher fertilization rate (G1 = 67%; G2 = 83%; p = 0.009). In vitro, testosterone, estradiol and progesterone were also reduced by LA, even though this reduction occurred for proges- terone only at the highest LA dosage (10 −6 M; 606.0 ± 114.3 ng/ml versus 1524.0 ± 246.5 ng/ml; p = 0.02). Results show that LA reduces ovarian steroidogenesis in vivo by essentially inhibiting androgen synthesis; whereas, in vitro, ovarian steroidogenesis is reduced over- all. © 2008 Elsevier Inc. All rights reserved. 1. Introduction The inclusion of gonadotropin-releasing hormone analogues (GnRHa) in controlled ovarian hyperstimulation schemes for assisted reproduction techniques has been used for more ∗ Corresponding author at: Av Bandeirantes, n ◦ 3900 – 8 ◦ andar – Departamento de Ginecologia e Obstetrícia, Bairro Monte Alegre, Ribeirão Preto – SP, CEP14049-900, Brazil. E-mail address: [email protected] (M.F. Silva-de-Sá). 1 Researcher of Nacional Council of Research-CNPq. than a decade. This is mainly because their combination with exogenous gonadotrophins results in a larger number of stimulated follicles, better oocyte quality, and higher fertil- ization and implantation rates [1–4]. They act by suppressing endogenous gonadotrophins via prevention of the premature 0039-128X/$ – see front matter © 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.steroids.2008.08.003
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Leuprolide acetate reduces both in vivo and in vitro ovarian steroidogenesis in infertile women...

Lsr
ARD
a
A
R
R
3
A
P
K
G
E
P
T
A
L
1
T(a
P
0d
s t e r o i d s 7 3 ( 2 0 0 8 ) 1475–1484
avai lab le at www.sc iencedi rec t .com
journa l homepage: www.e lsev ier .com/ locate /s tero ids
euprolide acetate reduces both in vivo and in vitro ovarianteroidogenesis in infertile women undergoing assistedeproduction
lcione L.A. Silva, Lauriane G.A. Abreu, Ana Carolina J.S. Rosa-e-Silva,ui A. Ferriani, Marcos F. Silva-de-Sá ∗,1
epartment of Gynecology and Obstetrics, Faculty of Medicine of Ribeirao Preto- University of Sao Paulo, Brazil
r t i c l e i n f o
rticle history:
eceived 21 November 2007
eceived in revised form
0 May 2008
ccepted 11 August 2008
ublished on line 26 August 2008
eywords:
nRH analogues
stradiol
rogesterone
estosterone
ndrostenedione
uteinized granulosa cells
a b s t r a c t
Despite the probable inhibitory effects of GnRH analogues on ovarian steroidogenesis in
vitro, their association with assisted reproduction protocols shows favorable results. This
suggests that there are important differences in the behaviors of these drugs when admin-
istered in vivo versus in vitro. To clarify these differences, this study was designed to analyze
the effect of leuprolide acetate (LA) on ovarian steroidogenesis in women undergoing In
Vitro Fertilization (IVF). A prospective, randomized open label study was conducted on 14
women (26–35 years): seven receiving only gonadotrophins (Group 1) and seven receiv-
ing gonadotrophin plus LA at 1mg/day (Group 2). The LA in vivo effect was determined
with serum and follicular fluid (FF) samples and via luteinized granulosa cell cultivation
(GCC), where cells were obtained during oocyte retrieval after ovarian hyperstimulation.
In vitro analysis was performed via addition of LA to GCC only for Group 1 (without
LA) at progressively higher concentrations (0, 10−12, 10−9 and 10−6 M). In vivo, the main
observation was a reduction in androgen production in Group 2, represented by lower
androstenedione production in FF (G1 = 6479 ± 3458; G2 = 3021 ± 1119 ng/ml; p = 0.04) and a
lower testosterone peak in GC at 96 h (G1 = 0.64 ± 0.12 ng/ml; G2 = 0.50 ± 0.19ng/ml; P = 0.02),
but a higher fertilization rate (G1 = 67%; G2 = 83%; p = 0.009). In vitro, testosterone, estradiol
and progesterone were also reduced by LA, even though this reduction occurred for proges-
terone only at the highest LA dosage (10−6 M; 606.0 ± 114.3 ng/ml versus 1524.0 ± 246.5 ng/ml;
p = 0.02). Results show that LA reduces ovarian steroidogenesis in vivo by essentially
inhibiting androgen synthesis; whereas, in vitro, ovarian steroidogenesis is reduced over-
with exogenous gonadotrophins results in a larger number of
all.
. Introduction
he inclusion of gonadotropin-releasing hormone analoguesGnRHa) in controlled ovarian hyperstimulation schemes forssisted reproduction techniques has been used for more
∗ Corresponding author at: Av Bandeirantes, n◦3900 – 8◦andar – Departareto – SP, CEP14049-900, Brazil.
E-mail address: [email protected] (M.F. Silva-de-Sá).1 Researcher of Nacional Council of Research-CNPq.039-128X/$ – see front matter © 2008 Elsevier Inc. All rights reserved.oi:10.1016/j.steroids.2008.08.003
© 2008 Elsevier Inc. All rights reserved.
than a decade. This is mainly because their combination
mento de Ginecologia e Obstetrícia, Bairro Monte Alegre, Ribeirão
stimulated follicles, better oocyte quality, and higher fertil-ization and implantation rates [1–4]. They act by suppressingendogenous gonadotrophins via prevention of the premature

( 2 0 0 8 ) 1475–1484
Fig. 1 – Study protocol. E1, estrone; E2, estradiol; P,progesterone; T, testosterone; A, androstenedione. Leu,
1476 s t e r o i d s 7 3
luteineized hormone (LH) peak and premature ovulation, aswell as the deleterious effects of excessive luteinization onthe oocyte [5].
The presence of specific GnRH receptors on the humanovary surface and specifically in granulosa cells has beendemonstrated, leading researchers to study the effects ofthese drugs, particularly on the ovaries [6–10]. GnRH carriesout paracrine and autocrine actions by influencing steroido-genesis, reducing proliferation and inducing ovarian cellsapoptosis [11]. In some animal species, the presence of aGnRH-binding inhibitor in the ovaries has been described[12,13]. In humans, the potential effects of GnRH analogueson ovarian cells and on the biology of the GnRH/GnRHreceptor are complex and several aspects must be eluci-dated.
The effects of GnRH agonists on steroidogenesis are contro-versial, although results tend to indicate the inhibitory effectsof GnRH agonists in vitro [3,14]. Modifications introduced inthe molecular structure of different agonists are responsi-ble for the pharmacological properties of these agents andinterfere with their ability to penetrate follicular fluid [15,16].Thus, contradictory results presented in various reports can beattributed to differences among the agonists used and espe-cially in the methodologies utilized, whether in vitro or in vivo,as different agonists seem to have different effects on thesteroidogenic function of granulosa cells [17].
Upon analyzing the association between steroid levels infollicular fluid and the oocyte maturation process, we noticeda paradox regarding agonist interference in steroidogenesisand their universally accepted use in assisted reproductionprograms, where they produce good results [18–20]. Thus,there might be differences in the behavior of these agonistswhen administered directly to patients or when applied togranulosa cell cultures in vitro. Behavioral differences are alsoimportant when considering their distinctive molecular formsand the different concentrations achieved in follicular fluid orthe culture medium.
Several studies specifically conducted on leuprolide acetateare available [3,14,21–24], demonstrating diverse resultsamong in vivo and in vitro studies. However, there are no reportsin the literature that compare the action of leuprolide on gran-ulosa cells using in vivo and in vitro models for the same groupof patients receiving gonadotrophins for in vitro fertilization(IVF)-related controlled ovarian hyperstimulation.
Thus, the aim of this study was to comparatively evalu-ate the effects of leuprolide acetate on steroidogenesis in vivoand in vitro by measuring steroids in serum, follicular fluidand cultured granulosa cells of women submitted to ovarianhyperstimulation for IVF.
2. Experimental
This study was conducted on 14 infertile women aged 26–35years who were submitted to IVF owing to a tuboperitonealor male factor. Work was followed at the Infertility Outpa-
tient Clinic of the University Hospital, Faculty of Medicine ofRibeirão Preto, University of São Paulo, Brazil. The study wasapproved by the Hospital Ethics Committee and all patientsgave written, informed consent to participate.leuprolide acetate.
In a prospective, simple and randomized open label study,patients were divided into two groups: Group 1 (seven patients)was submitted to controlled ovarian stimulation with recom-binant FSH (rFSH; Puregon®, Organon, São Paulo, SP, Brazil;Gonal®, Serono, São Paulo, SP, Brazil); Group 2 (seven patients)was submitted to ovarian stimulation with rFSH plus leupro-lide acetate (Lupron® - Abbott, Chicago, IL, USA) at 1 mg/day forpituitary desensitization starting on the first menstrual cycleday (short protocol) (Fig. 1). Randomization was performed bydrawing lots for the first participant, with each patient thenbeing introduced consecutively and alternately in a differ-ent group (1 or 2). All patients carried out the study to itscompletion and no significant adverse effects of leuprolidewere observed. Patients with endocrine diseases, includ-ing polycystic ovarian syndrome, were excluded from thestudy.
Blood samples were obtained on the first day of the cycleduring controlled ovarian stimulation, on the day of hCGadministration, and on the day of oocyte retrieval. Deter-mination of estradiol, estrone, progesterone, testosterone,and androstenedione levels in these samples was carriedout. Follicular fluid was also collected and granulosa cells,obtained from mature follicles punctured on the day ofretrieval, were cultured for evaluation of the same steroids.Plasma was centrifuged at 2500 rpm and stored at −20 ◦C for
later analysis. Follicular fluid was manipulated as describedbelow.Granulosa cells were cultured using a modified techniqueof the Instituto Valenciano de Infertilidad (Valencia, Spain) to

0 0 8
e[
2
2Tbcbcidtfw3datb
2aAgpmLstpw
2
Tscsifdcat
2
EworppoaiwH
s t e r o i d s 7 3 ( 2
valuate the in vivo and in vitro effects of LA on steroidogenesis25,26].
.1. Protocols for controlled ovarian stimulation
.1.1. Group 1: Gonadotrophins alonehe menstrual cycle was initially programmed using com-ined oral contraceptives that were initiated in the precedingycle. Transvaginal ultrasonography (USG) was performedetween the first and third days of the ovarian stimulationycle to exclude the presence of ovarian cysts. Controlled ovar-an stimulation with gonadotrophins was started with rFSH onay 3 of the cycle. Consecutive doses were adjusted accordingo ovarian response during US monitoring. When at least twoollicles reached 17 mm, 10.000 IU of hCG (Profasi®, Serono)as administered intramuscularly and oocytes were retrieved
4–36 h later. All follicles measuring more than 14 mm iniameter were aspirated. Embryo transfer was performed 72 hfter oocyte retrieval. Luteal supplementation was started onhe day of oocyte retrieval and maintained until menstrualleeding.
.1.2. Group 2: Gonadotrophins associated with leuprolidecetates described for Group 1, the menstrual cycle was pro-rammed using combined oral contraceptives during thereceding cycle. The GnRH agonist used for this experi-ent was leuprolide acetate (LA; [d-Leu6, Pro9-NHEt] GnRH,
upron®- Abbott) at a dose of 1 mg/day. This was administeredubcutaneously starting on the first day of the cycle and con-inuing until the day preceding administration of hCG (shortrotocol). Gonadotrophin administration and oocyte retrievalere performed as described for Group 1.
.2. In vivo versus in vitro analysis
he in vivo effect of LA was determined on the basis of sexteroid levels in serum, follicular fluid and granulosa cellultures obtained from patients submitted to ovarian hyper-timulation for IVF. Conversely, the in vitro effect was evaluatedn the culture medium of luteineized granulosa cells obtainedrom patients who did not receive leuprolide acetate in vivouring ovarian stimulation (Group 1). LA was added to theulture medium in progressively higher concentrations. Thenalyzed parameters were estradiol, progesterone and testos-erone.
.3. Cell culture
ach cell culture was performed using granulosa cells thatere harvested via aspiration of follicles containing matureocytes. This included 17 in Group 1 and 13 in Group 2. Mate-ial was collected in Falcon tubes (Costar) and poured intoreviously warmed (37 ◦C) Petri dishes. Fluid was left for aeriod of time to allow blood cells to fall out as sedimentn the dish bottom. Granulosa cells were then aspirated with
Pasteur pipette with simultaneously visualization using annverted photomicroscope (Olympus; IMT 2). The cellular clotas transferred to another dish for medium washing witham F-10 + bovine fetal serum (BFS, FCS Sigma St Louis, MO,
) 1475–1484 1477
USA) and HAM F10 medium (Cultilab, Campinas, SP, Brazil).Again, the blood cells were allowed to fall out as sediment onthe dish bottom. Dish content was subsequently transferredthrough a Pasteur pipette into a Falcon tube and centrifugedfor 10 min at 1500 rpm. The supernatant was removed and2 ml of medium was added to the pellet. This was vortexed toinduce cell separation. All of this material was transferred toa Percoll gradient column (Pharmacia, Uppsala, Sweden) thatwas previously prepared in Falcon tubes, and then centrifugedat 500 rpm for 30 min. Granulosa cells were then aspirated atthe interface of the Percoll column (clotted blood cells stayedat the bottom) and transferred to another sterile Falcon tube.Four milliliters of medium was added and the solution wasmixed to separate cells. This mixture was subsequently cen-trifuged at 1500 rpm for 10 min. Cells were then diluted withM199® culture medium (Adolfo Lutz, São Paulo, SP, Brazil),with FCS and glutamine (Sigma), then cultured at a concen-tration of 50,000 viable cells/wheel on 24-wheel cell cultureplates (Costar, Corning, NY, USA) at a volume of 1 ml perwheel.
A volume containing the desired number of cells for culturewas added to each wheel with culture medium. Cell viabil-ity was determined with Trypan Blue® (Merck, WhitehouseStation, NJ, USA). Cell cultures were incubated in a 5% CO2
atmosphere at 37 ◦C for 144 h, with changing of the mediumoccurring every 48 h. Cultures were then stored at −20 ◦Cfor later determination of estrone, estradiol, progesterone,androstenedione, and testosterone levels. All experimentswere carried out in triplicate.
After the first 144 h, cultures obtained from Group 1patients (i.e., patients who had not received leuprolide in vivo)were grouped into pools of four and reseeded. This producednew granulosa cell cultures (a total of four new cultures)to which progressively higher concentrations of leuprolideacetate (0, 10−12, 10−9 and 10−6 M) were added. These newcultures were maintained under identical conditions for morethan 144 h, with medium changes at 48 h intervals. Sampleswere then stored as described above.
2.4. Follicular fluid
After granulosa cell aspiration in the Petri dish, the follicu-lar fluid was poured into haemolysis tubes and centrifuged at2500 rpm. The supernatant was separated and submitted tovisual inspection and hematocrit, prior to storage at −20 ◦C,for hormonal dosage. Only the follicular fluid that was gradedas “clean”, according to criteria suggested by Levay et al. [27]was stored. Hormonal dosages were made only in follicularfluids in which the granulosa cells were successfully cultured(i.e., 17 in Group 1 and 13 in Group 2).
2.5. Hormone measurements
Estradiol, progesterone and testosterone were determined induplicate in plasma, follicular fluid and granulosa cell culturemedia using a radioimmunoassay with a double antibody kit
and a solid phase (DPC Medlab, Los Angeles, CA, USA). All sam-ples from the same patient were processed within the sameassay for each hormone. Minimum levels detected and intra-/inter-assay errors were: 8 pg/ml, 3.6% and 3% for estradiol;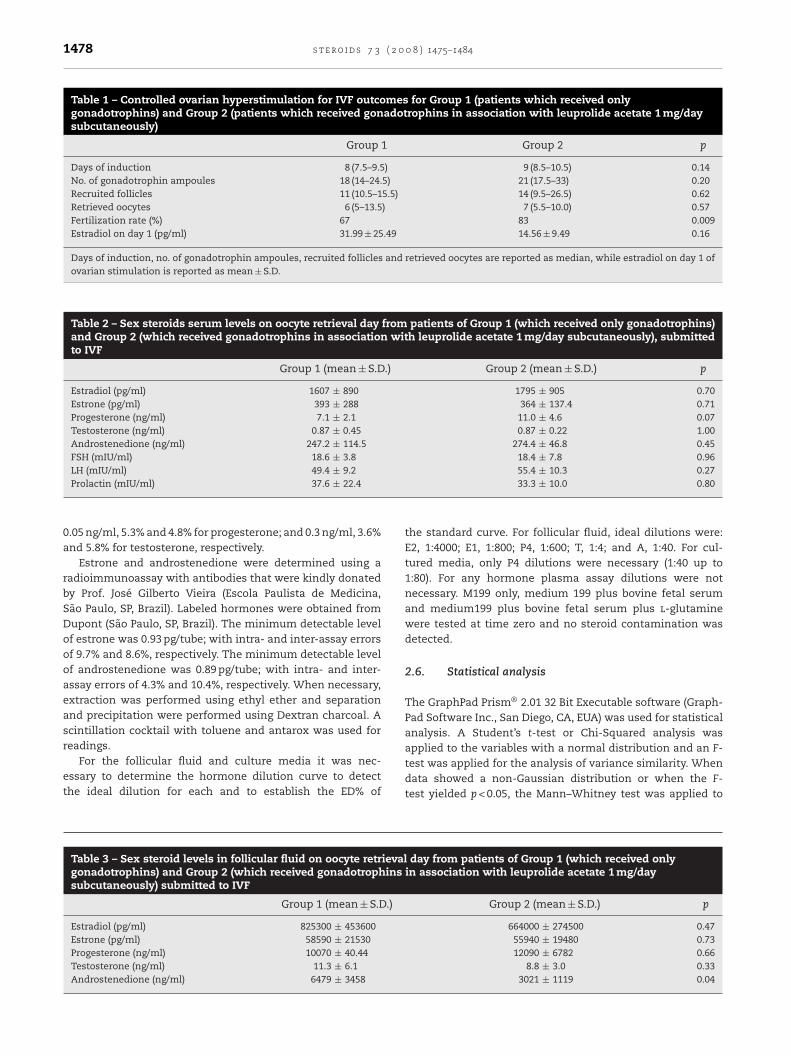
1478 s t e r o i d s 7 3 ( 2 0 0 8 ) 1475–1484
Table 1 – Controlled ovarian hyperstimulation for IVF outcomes for Group 1 (patients which received onlygonadotrophins) and Group 2 (patients which received gonadotrophins in association with leuprolide acetate 1 mg/daysubcutaneously)
Group 1 Group 2 p
Days of induction 8 (7.5–9.5) 9 (8.5–10.5) 0.14No. of gonadotrophin ampoules 18 (14–24.5) 21 (17.5–33) 0.20Recruited follicles 11 (10.5–15.5) 14 (9.5–26.5) 0.62Retrieved oocytes 6 (5–13.5) 7 (5.5–10.0) 0.57Fertilization rate (%) 67 83 0.009Estradiol on day 1 (pg/ml) 31.99 ± 25.49 14.56 ± 9.49 0.16
Days of induction, no. of gonadotrophin ampoules, recruited follicles and retrieved oocytes are reported as median, while estradiol on day 1 ofovarian stimulation is reported as mean ± S.D.
Table 2 – Sex steroids serum levels on oocyte retrieval day from patients of Group 1 (which received only gonadotrophins)and Group 2 (which received gonadotrophins in association with leuprolide acetate 1 mg/day subcutaneously), submittedto IVF
Group 1 (mean ± S.D.) Group 2 (mean ± S.D.) p
Estradiol (pg/ml) 1607 ± 890 1795 ± 905 0.70Estrone (pg/ml) 393 ± 288 364 ± 137.4 0.71Progesterone (ng/ml) 7.1 ± 2.1 11.0 ± 4.6 0.07Testosterone (ng/ml) 0.87 ± 0.45 0.87 ± 0.22 1.00Androstenedione (ng/ml) 247.2 ± 114.5 274.4 ± 46.8 0.45FSH (mIU/ml) 18.6 ± 3.8 18.4 ± 7.8 0.96
LH (mIU/ml) 49.4 ± 9.2Prolactin (mIU/ml) 37.6 ± 22.40.05 ng/ml, 5.3% and 4.8% for progesterone; and 0.3 ng/ml, 3.6%and 5.8% for testosterone, respectively.
Estrone and androstenedione were determined using aradioimmunoassay with antibodies that were kindly donatedby Prof. José Gilberto Vieira (Escola Paulista de Medicina,São Paulo, SP, Brazil). Labeled hormones were obtained fromDupont (São Paulo, SP, Brazil). The minimum detectable levelof estrone was 0.93 pg/tube; with intra- and inter-assay errorsof 9.7% and 8.6%, respectively. The minimum detectable levelof androstenedione was 0.89 pg/tube; with intra- and inter-assay errors of 4.3% and 10.4%, respectively. When necessary,extraction was performed using ethyl ether and separationand precipitation were performed using Dextran charcoal. Ascintillation cocktail with toluene and antarox was used for
readings.For the follicular fluid and culture media it was nec-essary to determine the hormone dilution curve to detectthe ideal dilution for each and to establish the ED% of
Table 3 – Sex steroid levels in follicular fluid on oocyte retrievalgonadotrophins) and Group 2 (which received gonadotrophinssubcutaneously) submitted to IVF
Group 1 (mean ± S.D.)
Estradiol (pg/ml) 825300 ± 453600Estrone (pg/ml) 58590 ± 21530Progesterone (ng/ml) 10070 ± 40.44Testosterone (ng/ml) 11.3 ± 6.1Androstenedione (ng/ml) 6479 ± 3458
55.4 ± 10.3 0.2733.3 ± 10.0 0.80
the standard curve. For follicular fluid, ideal dilutions were:E2, 1:4000; E1, 1:800; P4, 1:600; T, 1:4; and A, 1:40. For cul-tured media, only P4 dilutions were necessary (1:40 up to1:80). For any hormone plasma assay dilutions were notnecessary. M199 only, medium 199 plus bovine fetal serumand medium199 plus bovine fetal serum plus l-glutaminewere tested at time zero and no steroid contamination wasdetected.
2.6. Statistical analysis
The GraphPad Prism® 2.01 32 Bit Executable software (Graph-Pad Software Inc., San Diego, CA, EUA) was used for statisticalanalysis. A Student’s t-test or Chi-Squared analysis was
applied to the variables with a normal distribution and an F-test was applied for the analysis of variance similarity. Whendata showed a non-Gaussian distribution or when the F-test yielded p < 0.05, the Mann–Whitney test was applied today from patients of Group 1 (which received onlyin association with leuprolide acetate 1 mg/day
Group 2 (mean ± S.D.) p
664000 ± 274500 0.4755940 ± 19480 0.7312090 ± 6782 0.66
8.8 ± 3.0 0.333021 ± 1119 0.04
s t e r o i d s 7 3 ( 2 0 0 8 ) 1475–1484 1479
Table 4 – Hormone levels in the culture medium after 48, 96 and 144 h of granulosal luteinized cells cultivation, obtainedfrom patients submitted to IVF divided into two Groups: 1(which received only gonadotrophins) and 2 (which receivedgonadotrophins in association with leuprolide acetate 1 mg/day subcutaneously)
Time of culture Hormones Group 1 Group 2 p
48 h E2 762.5 ± 271.5 739.6 ± 194.3 0.79P4 1070.0 ± 323.1 964.5 ± 355.2 0.40T 0.57 ± 160.0 0.45 ± 0.18 0.10A 28.2 ± 12.4 33.3 ± 14.8 0.30
96 h E2 209.6 ± 69.0 190.9 ± 58.6 0.43P4 1750.3 ± 461.3 1693.2 ± 391.8 0.72T 0.64 ± 0.12 0.5 ± 0.19 0.02A 29.8 ± 16.9 33.4 ± 19.2 0.58
144 h E2 115.4 ± 35.5 111.4 ± 39.7 0.76P4 1058.1 ± 412.0 928.6 ± 311.7 0.35T 0.55 ± 0.15 0.45 ± 0.16 0.10
23
A, an
uvwmnp
FG(
A
E2, estradiol(pg/ml); P4, progesterone(ng/ml); T, testosterone(ng/ml);
npaired variables and the Wilcoxon test was applied to pairedariables. Simultaneous comparison of three or four groups
as performed with ANOVA when distributions were nor-al and by the Kruskal–Wallis test when distributions wereot normal. In all cases, the level of significance was set at< 0.05.
ig. 2 – Hormonal levels in granulosal luteinized cells culture meroup 1 (no leuprolide administration in vivo) and Group 2 (leupr
pg/ml); P, progesterone (ng/ml); T, testosterone (pg/ml); A, andro
.7 ± 11.9 27.9 ± 16.2 0.42
drostenedione (ng/ml).
3. Results
3.1. Ovarian stimulation cycle
There was no difference in patient mean age between Groups1 and 2 (G1 = 32.6 ± 2.6 years and G2 = 30.7 ± 3.3 years). Groups
dium after 48, 96 and 144 h obtained from patients fromolide administration in vivo) submitted to IVF. E2, estradiolstenedione (ng/ml). a = 0.02; b = 0.04.

( 2 0 0 8 ) 1475–1484
Fig. 3 – Peak of each hormone during granulosal luteinizedcells culture obtained only from patients from Group 1 (noleuprolide administration during ovarian stimulation forIVF) with the in vitro addition of progressively higher
1480 s t e r o i d s 7 3
also did not differ regarding the number of ovarian inductiondays, the number of gonadotrophin ampoules administered,the number of follicles recruited, the number of oocytesretrieved and the serum estradiol levels on the ovarian stimu-lation cycle starting day (Table 1). Serum estradiol levels weresimilar for both groups on the day of hCG administration(G1 = 2500 ± 922 and G2 = 3279 ± 1359 pg/ml; p = 0.23) as well ason the day of oocyte retrieval (Table 2). A reduction in serumestradiol levels, from the day of hCG administration to theday of oocyte retrieval, was observed in both groups. Withlevels going from 2500 ± 922 to 1607 ± 890 pg/ml in Group 1(p = 0.02) and from 3279 ± 1359 to 1795 ± 905 pg/ml in Group 2(p = 0.01). Estrone, progesterone, testosterone and androstene-dione were also determined on the day of oocyte retrieval, withno difference in their levels between groups (Table 2).
3.2. Assisted reproduction procedures
The recruited follicles and retrieved oocytes were analyzedin terms of their microscopic morphological characteristics[28,29]. Table 1 presents the number of follicles recruited andoocytes retrieved as well as the fertilization rates for eachgroup. No statistical differences were observed regarding therecruited follicles and retrieved oocytes (p = 0.62 and p = 0.57respectively; Student’s t-test). Oocyte fertilization rates were67% for Group 1 and 83% for Group 2 (p = 0.009; chi-squared).
3.3. Follicular fluid
Follicles and their contents were evaluated on the day ofoocyte retrieval. Mean follicular diameter was similar forboth groups (i.e., 18.7 ± 2.9 mm for Group 1 and 20.3 ± 2.8 mmfor Group 2; p = 0.15). Estrone, estradiol, progesterone,androstenediona and testosterone levels were measured inthe follicular fluid and were equivalent for both groups.Androstenedione, however, was an exception, with levels thatwere lower in Group 2 (6479 ± 3458 and 3021 ± 1119 ng/ml forGroups 1 and 2, respectively; p = 0.04) (Table 3).
3.4. Luteinized granulosa cell cultures
Estrone, estradiol, progesterone, testosterone and androstene-dione levels in the culture medium were determined after 48,96 and 144 h. Estrone and estradiol production were highestafter 48 h of culture, whereas progesterone and testosteroneproduction were highest after 96 h. Androstenedione levelswere constant during the 144 h of culture. A significant dif-ference between groups was observed only for testosteroneafter 96 h of culture, with Group 2 presenting significantlylower production than Group 1 (G2 = 500.0 ± 190.0 pg/ml versusG1 = 640.0 ± 120.0 pg/ml; p = 0.02) (Table 4 and Fig. 2).
When leuprolide acetate was added to these pooled cul-tures of Group 1 patients, curves of estradiol, progesteroneand testosterone production were found to follow the samepatterns for all LA concentrations used. The hormone levelsof these curves were compared at the peak point for each
hormone. For estradiol this point was at 48 h and for proges-terone and testosterone it was at 96 h (Fig. 3). The additionof LA resulted in a reduction of hormone levels for estradioland progesterone at the peak moment. However, this was notconcentrations of leuprolide acetate. E2, estradiol (pg/ml); P,progesterone (ng/ml); T, testosterone (pg/ml). (a, b) = p < 0.05.
observed for testosterone, even though there was a tendencyfor this reduction to occur (Fig. 3).
4. Discussion
Despite discrepancies in the results, most papers point toan inhibitory effect of GnRH agonists on ovarian steroido-
s t e r o i d s 7 3 ( 2 0 0 8 ) 1475–1484 1481
Table 5 – Mean hormone levels after the addition of progressively higher leuprolide acetate concentrations to the culturedgranulosal luteinized cells obtained from Group 1 patients (which received only gonadotrophins during ovarianhyperstimulation) after 48, 96 and 144 h of culture
Leuprolide concentration inthe culture medium
48 h 96 h 114 h
Estradiol (pg/ml) 0 881.6 ± 143.4a 331.6 ± 73.5 162.8 ± 45.410−12 M 769.2 ± 82.3 357.2 ± 49.2 202.2 ± 45.410−9 M 609.5 ± 98.8b 300.0 ± 69.5 191.2 ± 75.210−6 M 604.0 ± 129.9 265.0 ± 86.3 125.2 ± 50.6
Progesterone (ng/ml) 0 744.0 ± 216.4 1524.0 ± 246.5c 372.0 ± 128.010−12 M 696.0 ± 234.8 1440.0 ± 356.8 384.0 ± 108.810−9 M 558.0 ± 128.5 1062.0 ± 157.6 222.0 ± 72.510−6 M 450.0 ± 134.3 606.0 ± 114.3d 534.0 ± 123.4
Testosterone (pg/ml) 0 400.0 ± 80.0 600.0 ± 180.0 400.0 ± 110.010−12 M 400.0 ± 80.0 500.0 ± 140.0 400.0 ± 80.010−9 M 200.0 ± 80.0 400.0 ± 80.0 200.0 ± 80.010−6 M 300.0 ± 140.0 400.0 ± 80.0 200.0 ± 80.0
gfatg
ofirtL
ttamifwo
dppgctrloplmdTlff
M = Molar concentration. a > b, p = 0.02; c > d, p = 0.02.
enesis in vivo or in vitro. They may act at different steps,rom cholesterol transportation, inhibition on progesteronend pregnenolone production, reduction in conversion of C21o C19 compounds and in androgens aromatization to estro-ens [2,3,30–36].
Regarding the in vivo effect of leuprolide acetate, webserved a significant reduction of androstenedione levels inollicular fluid from patients who received leuprolide admin-stration (Group 2). In addition, testosterone production waseduced in luteinized granulosa cell cultures obtained fromhese same women in comparison with Group 1 patients (noA administration) (Table 5).
We believe that there is a relationship between thesewo results and that discovered differences can be attributedo differences among utilized methodologies. Androgensre intermediate compounds in steroidogenesis and theiretabolism is very complex. The interconversion from A to T
s regulated by 17 � HSD, which is present in tissues in multipleorms, having specific roles for synthesis and catabolism, asell as acting with different intensities according to the organr tissue under consideration [37].
Granulosal cells that are cultured in vitro have completelyifferent conditions for steroidogenesis compared to thoseresent in follicular fluid, where the flow of steroidogenesisrecursors is very dynamic. Here there is integration betweenranulosal and stromal cells and the oocyte, including theonversion of A to T. This occurs according to the necessi-ies of the oocyte-cumulus unit, which has its own complexegulation mechanism. On the other hand, in cultured granu-osa cells, the production of A and T is limited by the amountf enzymes and endogenous steroid precursors or the com-ounds offered in the culture medium, which is obviously
imited by time exposure. With dependency on the experi-ental model utilized, GnRH agonists may or may not have
emonstrable effects on androstenedione production [2,22].
able 3 shows that, in the follicular fluid, androstenedioneevels (ng/ml) were 6479 ± 3458 and 3021 ± 1119, respectively,or Groups 1 and 2. Testosterone levels (ng/ml) were 11.3 ± 6.1or Group 1 and 8.8 ± 3.0 for Group 2. So, the rate A:Twas 573.3 for Group 1 and 382.9 for Group 2 (Group 1 >Group 2).
There was a significant reduction in androgens producedin the culture medium. Androstenedione was 217 and testos-terone was 17.6 times less abundant than in follicular fluid.Certainly the time and reserve of culture cell endogenous pre-cursors were important factors that may have an influenceon these results. However, there is a clear reduction in theratio A:T in the culture medium. At the 96 h cell culture period(Table 4), the ratio A:T was 46.5 for Group 1 and 66.8 for Group2 (Group 1 < Group 2), which is significantly lower than forfollicular fluid. By considering the A:T ratio as an indirect mea-surement of 17 � HSD activity, results suggest that there aresignificant differences in the interconversion of A to T in twoexperimental situations. In one, the follicular fluid drives theconversion preferably to A, while in the other, the granulosacultured cells drive it to T. Therefore, the effect of LA would bebetter observed on A production, as is the case for follicularfluid, and on T in cultured granulosa cells.
For in vitro results, we observed that the addition of dif-ferent leuprolide concentrations to the luteinized granulosacell culture medium had an inhibitory effect on the threesteroids analyzed. The progesterone reduction was significantonly when the highest leuprolide concentration was addedto the culture medium. However, for estradiol this effect wasobserved even at lower leuprolide concentrations, suggestingvaried degrees of sensitivity for enzymes involved in the invitro steroidogenesis cascade. The reduction of progesteroneand estradiol levels observed only in vitro may also mean that,in addition to the doses, the experimental model itself mayinfluence results.
The in vitro inhibitory effects of leuprolide acetate ontestosterone production showed strong similarity to the otherhormones, having a dose-effect pattern. However, this inhi-bition was not statistically significant. There may be two
explanations for this fact. The first is that the small numberof experiments and resulting data did not reach statistical sig-nificance. The second, which seems quite attractive, is thattestosterone reduction was compensated by the blockade of
( 2 0
r
ovarian granulosa-luteal cells. Mol Endocrinol
1482 s t e r o i d s 7 3
its conversion to estradiol (i.e., the “consumption” of testos-terone did not occur with the same rate as its production).Although it was inhibited, testosterone was not converted toestradiol because of the aromatase blockade by the GnRH ago-nist. This was already demonstrated in a previous study withgranulosa cells from immature follicles or in luteinized gran-ulose cells [3,38] “masking” the true extent of inhibition fortestosterone production. It must be considered that, in thispaper, cultured granulosa cells, which were obtained fromwell-developed and luteinized follicles, were utilized. Resultscannot automatically be extended to all granulosa cells. How-ever, we may not rule out some similarities. It has been shownthat both immature and mature luteinized follicles are simi-larly inhibited in vivo and in vitro by GnRH agonists in differentanimal species, with marked intensity differences resultingfrom dependencies on follicle size and the steroid under con-sideration [38–40].
We recognize that in vitro studies have their limitationsand do not reflect a clinical situation; however, in thispaper they not only confirmed the inhibitory effect of LAon steroidogenesis, but also permitted the establishment ofthe dose-dependence of this phenomenon. Although differ-ent methodological approaches were utilized, results lead usto believe that the data are complementary.
Results from this study are quite coherent with findingsobserved during the clinical practice of assisted reproduction[16,41,42]. The randomization model utilized was adequateand did not influence results. Both groups were homogenousin age, number of stimulation days, number of gonadotrophinampoules, number of recruited follicles and estradiol levelsprior to ovarian stimulation (as was also observed for estrone,progesterone, androstenedione and testosterone). No differ-ence was observed for the cultured cells. These facts lead usto believe that differences in results were due to the LA effect.
The influence of follicular fluid composition on oocytematuration is well known [18,20,43–46] and higher andro-gen concentrations in the follicular microenvironment mayimpair follicular development [47]. The maturity and fertiliza-tion potential of the oocyte are inversely related to levels oftestosterone in follicular fluid and there is a clear relation-ship between androgen levels in follicular fluid and oocytequality [48,49]. The reduction of androstenedione in follicu-lar fluid and of testosterone in cell cultures from patients whohad received leuprolide indicates that this may be one of themechanisms by which agonists improve the clinical resultsof assisted reproduction. This is in terms of oocyte quality,retrieval and fertilization, and embryo implantation reportedin the literature [44–46].
Leuprolide, by reducing androgen levels, may have a bene-ficial effect on oocyte maturation, with an improved potentialfor mature oocyte fertilization. This fact, reported by others[1,16,48] was also observed in the present study, with a sig-nificant increase in the oocyte fertilization rate for the grouptreated with leuprolide. Thus, the GnRH agonist may act ongranulosa cells both in vivo and in vitro, interfering in a differentand dose-dependent manner on C21, C19 and C18 produc-
tion. Its action on androgen production is considered to be themost clinically significant effect. Inhibition of progesteroneand estradiol production, however, required higher leuprolideconcentrations in granulosa cells, which was achieved only0 8 ) 1475–1484
with direct in vitro applications.In conclusion, leuprolide acetate reduces granulosal
steroidogenesis in vivo by inhibition of androgen synthesis.This finding may explain the better fertilization rates observedin patients treated with this drug. On the other hand, in vitro,in addition to the inhibitory effects of LA on progesterone andtestosterone, we also observed a more intense reduction ofestradiol, suggesting a different mechanism of drug actionwhen applied in vivo or in vitro. The main difference seems tobe related to aromatase activity. At first this occurs by impair-ing its action in vitro, but not in vivo. Secondly, it shows greatersensitivity to the LA effect than other enzymes of the steroido-genic cascade.
Thus, it appears that there are still unknown mechanismsin vivo, but not in vitro, which perpetuate aromatase activity.This is an enzyme of primordial importance for the productionof estradiol and for the establishment of a favorable follicularmicroenvironment, which is necessary to reach good oocytequality.
Acknowledgement
CAPES - Coordenacão de Aperfeicoamento de Pessoal doEnsino Superior for providing financial support and to Mrs.MAV Bortolieiro and Mrs. MAM Ruzzeno for technical assis-tance.
e f e r e n c e s
[1] Meldrum DR, Wisot A, Hamilton F, Gutlay AL, Kempton WF,Huynh D. Routine pituitary suppression with leuprolidebefore ovarian stimulation for oocyte retrieval. Fertil Steril1989;51:455–9.
[2] Akaboshi K, Oda T, Yoshida J, Kohriyama S, Miyasaki T,Yoshimura Y. The effects of gonadotropin-releasinghormone agonist on androstenedione production andfollicular development during controlled ovarianhyperstimulation. J Assist Reprod Genet 1998;15:478–84.
[3] Dor J, Bider D, Shulman A, Levron JL, Shine S, Mashiach S, etal. Effects of gonadotrophin-releasing hormone agonists onhuman ovarian steroid secretion in vivo and in vitro -results of a prospective, randomized in vitro fertilizationstudy. Hum Reprod 2000;15:1225–30.
[4] Macklon NS, Stouffer RL, Giudice LC, Fauser BC. The sciencebehind 25 years of ovarian stimulation for in vitrofertilization. Endocr Rev 2006;27:170–207.
[5] Dodson WC. Gonadotropin-releasing hormone analogtherapy in ovulation induction. Semin Reprod Endocrinol1990;8:198–207.
[6] Peng C, Fan NC, Ligier M, Vaananen J, Leung PC. Expressionand regulation of gonadotropin-releasing hormone (GnRH)and GnRH receptor messenger ribonucleic acids in humangranulosa-luteal cells. Endocrinology 1994;135:1740–6.
[7] Cheng CK, Yeung CM, Chow BK, Leung PC. Characterizationof a new upstream GnRH receptor promoter in human
2002;16:1552–64.[8] Tarlatzis BC, Kolibianakis EM. Direct ovarian effects and
safety aspects of GnRH agonists and antagonists. ReprodBiomed Online 2002;5(Suppl 1):8–13, 3.

0 0 8
s t e r o i d s 7 3 ( 2[9] Khosravi S, Leung PC. Differential regulation ofgonadotropin-releasing hormone (GnRH) I and GnRH IImessenger ribonucleic acid by gonadal steroids in humangranulosa luteal cells. J Clin Endocrinol Metab2003;88:663–72.
[10] Ramakrishnappa N, Rajamahendran R, Lin YM, Leung PC.GnRH in nonhypothalamic reproductive tissues. AnimReprod Sci 2005;88:95–113.
[11] Metallinou C, Asimakopoulos B, Schröer A, Nikolettos N.Gonadotropin-releasing hormone in the ovary. Reprod Sci2007;14:737–49.
[12] Aten RF, Behrman HR. A gonadotropin-releasinghormone-binding inhibitor from bovine ovaries. Purificationand identification as histone H2A. J Biol Chem1989;264:11065–71.
[13] Aten RF, Behrman HR. Antigonadotropic effects of thebovine ovarian gonadotropin-releasing hormone-bindinginhibitor/histone H2A in rat luteal and granulosal cells. J BiolChem 1989;264:11072–5.
[14] Tureck RW, Mastroianni Jr L, Blasco L, Strauss 3rd JF.Inhibition of human granulosa cell progesterone secretionby a gonadotropin-releasing hormone agonist. J ClinEndocrinol Metab 1982;54:1078–80.
[15] Huirne JA, Lambalk CB.Gonadotropin-releasing-hormone-receptor antagonists.Lancet 2001;358:1793–803.
[16] Shapiro DB. An overview of GnRH antagonists in infertilitytreatments. Introduction. Fertil Steril 2003;80(suppl 1):S1–7.
[17] Balasch J, Jove IC, Moreno V, Civico S, Puerto B, Vanrell JA.The comparision of two gonadotropin-releasing hormoneagonists in an in vitro fertilization program. Fertil Steril1992;58:991–4.
[18] Fishel SB, Edwards RG, Walters DE. Follicular steroid as aprognosticator of successful fertilization of human oocytesin vitro. J Endocrinol 1983;99:335–44.
[19] Pellicer A, Gaitan P, Neuspiller F, Ardiles G, Albert C, RemohiJ, et al. Ovarian follicular dynamics: from basic science toclinical practice. J Reprod Immunol 1998;39:29–61.
[20] Costa LO, Mendes MC, Ferriani RA, Moura MD, Reis RM, Silvade Sa MF. Estradiol and testosterone concentrations infollicular fluid as criteria to discriminate between matureand immature oocytes. Braz J Med Biol Res 2004;37:1747–55.
[21] Dodson WC, Myers T, Morton PC, Conn PM. Leuprolideacetate: serum and follicular fluid concentrations andeffects on human fertilization, embryo growth, andgranulosa-lutein cell progesterone accumulation in vitro.Fertil Steril 1988;50:612–7.
[22] Frederick JL, Hickey MJ, Francis MM, Sauer MV, Paulson RJ.The effect of leuprolide acetate on steroidogenesis bygranulosa and theca cells in vitro. J In Vitro Fert EmbryoTransf 1991;8:230–4.
[23] Miro F, Sampaio MC, Tarin JJ, Pellicer A. Steroidogenesis invitro of human granulosa-luteal cells pretreated in vivo withtwo gonadotropin releasing hormone analogs employingdifferent protocols. Gynecol Endocrinol 1992;6:77–84.
[24] Fabbri R, Porcu E, Pession A, Sereni E, Marsella T, SeracchioliR, et al. The effect of leuprorelin on steroidogenesis ofhuman preovulatory granulosa cells in vitro. J Assist ReprodGenet 1996;13:287–92.
[25] Pellicer A, Miro F. Steroidogenesis in vitro of humangranulosa-luteal cells pretreated in vivo withgonadotropin-releasing hormone analogs. Fertil Steril1990;54:590–6.
[26] Garrido N, Albert C, Krussel JS, O’Connor JE, Remohi J, Simon
C, et al. Expression, production, and secretion of vascularendothelial growth factor and interleukin-6 by granulosacells is comparable in women with and withoutendometriosis. Fertil Steril 2001;76:568–75.) 1475–1484 1483
[27] Levay PF, Huyser C, Fourie F, Lê R, Rossowd J. The detectionof blood contamination in human follicular fluid. J AssistReprod Genet 1997;14:212–7.
[28] Testart J, Frydman R, Mouzon J, Lassalle B, Belaishc JC. Astudy of factors affecting the success of human fertilizationin vitro. I. Influence of ovarian stimulation upon the numberand condition of oocytes collected. Biol Reprod1983;28:415–24.
[29] Veeck LL, Wortham Jr JW, Witmyer J, Sandow BA, Acosta AA,Garcia JE, et al. Maturation and fertilization ofmorphologically immature human oocytes in a program ofin vitro fertilization. Fertil Steril 1983;39:594–602.
[30] Uemura T, Namiki T, Kimura A, Yanagisawa T, Minaguchi H.Direct effects of gonadotropin-releasing hormone on theovary in rats and humans. Horm Res 1994;41(Suppl-1):7–13.
[31] Srivastava RK, Luu-The V, Marrone BL, Sridaran R.Suppression of luteal steroidogenesis by an LHRHantagonist (Nal-Lys antagonist: antide) in vitro during earlypregnancy in the rat. J Mol Endocrinol 1994;13:87–94.
[32] Srivastava RK, Luu-THE, Marrone BL, Harris-Hooker S,Sridaran R. Inhibition of steridogenesis by luteal cells ofearly pregnancy in the rat in response to in vitroadministration of a gonadotropi-releasing hormone agonist.J Steroid Biochem Mol Biol 1994;49:73–9.
[33] Guerrero HE, Stein P, Asch RH, De Fried EP, Tesone M. Effectof a gonadotropin-releasing hormone agonist on luteinizinghormone receptors and steroidogenesis in ovarian cells.Fertil Steril 1993;59:803–8.
[34] Kowalik A, Barmat L, Damario M, Liu HC, Davis O,Rosenwakx Z. Ovarian estradiol production in vivo.Inhibitory effect of leuprolide acetate. J Reprod Med1998;43:413–7.
[35] Sridaran R, Lee MA, Haynes L, Srivastava RK, Ghose M,Sridaran G, et al. GnRH action on the luteal setroidogenesisduring pregnancy. Steroids 1999;64:618–23.
[36] Srivastava RK, Krishna A, Sridaran R. Follicular development,steroidogenesis and gondotrophin secretion in response tolong-term treatment with gonadotrophin-releasinghormone agonist in the rat. J Endocrinol 1995;146:349–57.
[37] Strauss JF. The synthesis and metabolism of steroidshormones. In: Yen, Jaffe’s, editors. ReproductiveEndocrinology. 5th ed. Philadelphia: Elsevier SaundersPhiladelphia; 2004. p. 125–54.
[38] Wickings EJ, Eidne KA, Dixson AF, Hillier SG.Gonadotropin-releasing hormone analogs inhibit primategranulosa cell steroidogenesis via mechanism distinct fromthat in the rat. Biol Reprod 1990;43:305–11.
[39] MaruoT, Otani T, Mochizuki M. Anti-gonadotropic actions ofGnRH agonist on avarian cells in vivo and in vitro. J SteroidBiochem 1985;23:765–70.
[40] Harlow CR, Shaw HJ, Hillier SG, Hodges JK. Factorsinfluencing follicle-stimulating hormone-responsivesteroidogenesis in marmoset granulosa cells: effects ofandrogens and the stages of follicular maturity.Endocrinolgy 1988;122:2780–7.
[41] Filicori M, Flamigni C, Cognigni GE, Falbo A, Arnone R,Capelli M, et al. Different gonadotropin and leuprorelinovulation induction regimens markedly affect follicular fluidhormone levels and folliculogenesis. Fertil Steril1996;65:387–93.
[42] Daya S, Gunby J. Recombinant versus urinary folliclestimulating hormone for ovarian stimulation in assistedreproduction cycles. Cochrane Database Syst Rev 2000;4.
CD002810.[43] McNatty KP, Smith DM, Makris A, Osathanondh R, Ryan KJ.The microenvironment of the human antral follicle:interrelationships among the steroid levels in antral fluid,

( 2 0
1484 s t e r o i d s 7 3the population of granulosa cells and the status of the oocytein vivo and in vitro. J Clin Endocrinol Metab 1979;49:851–60.
[44] Moor RM, Polge C, Willadsen SM. Effect of follicular steroidson the maturation and fertilization of mammalian oocytes. JEmbryol Exp Morphol 1980;56:319–35.
[45] Murdoch WJ, Dunn TG. Alterations in follicular steroidhormones during the preovulatory period in the ewe. BiolReprod 1982;27:300–7.
[46] Bottero-Ruiz W, Laufer N, De Cherney AH, Polan ML,Haseltine FP, Behrman HR. The relationship betweenfollicular fluid steroid concentration and successfulfertilization of human oocytes in vitro. Fertil Steril1984;41:820–6.
0 8 ) 1475–1484
[47] Ireland JJ, Roche JF. Development of antral follicles in cattleafter prostaglandin induced luteolysis: changes in serumhormones, steroids in follicular fluid, and gonadotropinreceptors. Endocrinology 1982;111:2077–86.
[48] Readhead C, Lobo LA, Kletzky OA. The activity of3�hidroxysteroid dehydrogenase and delta 4-5 isomerase inhuman follicular tissue. Am J Obstet Gynecol 1983;145:491–5.
[49] Franchimont P, Hazeé-Hagelstein MT, Hazout A, Frydman R,Schatz B, Demerle F. Correlation between follicular fluidcontent and the results of in vitro fertilization and embryotransfer. I. Sex steroids. Fertil Steril 1989;52:1006–11.








![Astrocytic tracer dynamics estimated from [1-11C]-acetate PET measurements](https://static.fdokumen.com/doc/165x107/6334cca03e69168eaf070c95/astrocytic-tracer-dynamics-estimated-from-1-11c-acetate-pet-measurements.jpg)











