
Enhanced cytotoxicity of an anti-transferrin receptor IgG3-avidin fusion protein in combination with...
12
ORIGINAL ARTICLE Enhanced cytotoxicity of an anti-transferrin receptor IgG3-avidin fusion protein in combination with gambogic acid against human malignant hematopoietic cells: functional relevance of iron, the receptor, and reactive oxygen species E Ortiz-Sa ´nchez 1 , TR Daniels 1 , G Helguera 1 , O Martinez-Maza 2,3,4 , B Bonavida, 3,4 and ML Penichet 1,3,4 1 Division of Surgical Oncology, Department of Surgery, David Geffen School of Medicine, University of California, Los Angeles, CA, USA; 2 Department of Obstetrics and Gynecology, David Geffen School of Medicine, University of California, Los Angeles, CA, USA; 3 Jonsson Comprehensive Cancer Center, David Geffen School of Medicine, University of California, Los Angeles, CA, USA and 4 Department of Microbiology, Immunology, and Molecular Genetics, David Geffen School of Medicine, University of California, Los Angeles, CA, USA The human transferrin receptor (hTfR) is a target for cancer immunotherapy due to its overexpression on the surface of cancer cells. We previously developed an antibody-avidin fusion protein that targets hTfR (anti-hTfR IgG3-Av) and exhibits intrinsic cytotoxicity against certain malignant cells. Gambogic acid (GA), a drug that also binds hTfR, induces cytotoxicity in several malignant cell lines. We now report that anti-hTfR IgG3-Av and GA induce cytotoxicity in a new broader panel of hematopoietic malignant cell lines. Our results show that the effect of anti-hTfR IgG3-Av is iron-dependent whereas that of GA is iron-independent in all cells tested. In addition, we observed that GA exerts a TfR-independent cytotoxicity. We also found that GA increases the generation of reactive oxygen species that may play a role in the cytotoxicity induced by this drug. Additive cytotoxicity was observed by simultaneous combination treatment with these drugs and synergy by using anti-hTfR IgG3-Av as a chemosensitizing agent. In addition, we found a concentration of GA that is toxic to malignant hematopoietic cells but not to human hematopoietic progenitor cells. Our results suggest that these two compounds may be effective, alone or in combination, for the treatment of human hematopoietic malignancies. Leukemia (2009) 23, 59–70; doi:10.1038/leu.2008.270; published online 23 October 2008 Keywords: TfR; iron; antibody fusion protein; gambogic acid; hematopoietic malignances; ROS Introduction Iron is an important cofactor of enzymes that participate in cellular metabolism including those of the respiratory chain and DNA synthesis. 1 Iron associated with transferrin is internalized by cells through transferrin receptor (TfR)-mediated endocyto- sis. 2 The TfR family consists of TfR1 (CD71) and TfR2, which are type II transmembrane glycoproteins. TfR1 is expressed at different levels on the surface of most normal cells in which the majority show low TfR1 expression level. 2 However, dividing cells such as basal epidermis and intestinal epithelium cells express high levels of TfR1. 2 In contrast, although TfR2 mRNA is present in different tissues, 3 the TfR2 protein is limited to hepatocytes and enterocytes of the small intestine. 2,4 As cancer cells require large amounts of iron to maintain their high rate of cell proliferation, TfR1 is overexpressed on their surface. 2 The TfR1 overexpression in malignant cells, its ability to internalize and its central role in the cellular pathology of human cancer make this receptor an attractive target for the therapy of cancer. 2,5 TfR has been targeted by monoclonal antibodies that either induce cytotoxicity 2 or are used for drug delivery. 5 Our group developed earlier an antibody fusion protein that specifically recognizes hTfR1 and is composed of chicken avidin, geneti- cally fused to the C H 3 domain of a mouse/human chimeric IgG3 (anti-hTfR IgG3-Av, Figure 1a), to be used as a universal delivery system of biotinylated drugs into cancer cells. 6–8 Unexpectedly, we discovered that this fusion protein exhibits a significant intrinsic cytotoxicity against certain hematopoietic malignant cell lines compared with the antibody alone (anti-hTfR IgG3). 6,7,9 Anti-hTfR IgG3-Av disrupts TfR recycling and redirects it into the lysosomes where it is presumably degraded, promoting iron starvation and the induction of apoptosis that can be blocked by iron supplementation. 7,9 As avidin is a non-covalent tetrameric protein, the presence of two avidin moieties in anti-hTfR IgG3-Av favors the dimerization of this fusion protein. 6 Although the structural requirements responsible for the cytotoxicity of anti- hTfR IgG3-Av are still unclear, the activity of this fusion protein is considered to be, at least in part, due to its dimeric structure (Figure 1a). 7 We have recently shown that anti-hTfR IgG3-Av conjugated to biotinylated saporin (b-SO6), a plant toxin of Saponaria officinialis, increases its cytotoxicity in malignant cells and more importantly, overcomes the resistance of cells to the fusion protein alone. 9 In addition, as anti-hTfR IgG3-Av specifi- cally binds hTfR1 (not hTfR2), this antibody fusion protein delivers b-SO6 into cells that express hTfR1. 8 Therefore, the fusion of avidin to anti-hTfR IgG3 results in a relevant tool that can be used in cancer immunotherapy as it elicits cytotoxic effects by itself and can also deliver biotinylated cytotoxic compounds such as b-SO6 into cancer cells that overexpress hTfR1. We are also interested in the effect of the combination of anti- hTfR IgG3-Av with non-biotinylated cytotoxic drugs. We have focused on gambogic acid (GA) as it can also bind hTfR. 10 GA is a xanthone isolated from a resin obtained from the Garcinia hanburyi tree that is used in oriental traditional medicine (Figure 1b). 11 GA induces cytotoxicity in malignant epithelial cell lines and also in malignant hematopoietic cell lines such as JURKAT and HL-60 cells. 12–21 GA can also inhibit the growth of human lung carcinoma and hepatoma xenografts in nude mice. 21 This drug blocks hTfR internalization and induces Received 17 April 2008; revised 14 August 2008; accepted 27 August 2008; published online 23 October 2008 Correspondence: Dr ML Penichet, Division of Surgical Oncology, Department of Surgery, University of California, UCLA, 10833 Le Conte Avenue, CHS 54-140, Box 951782, Los Angeles, California, 90095-1782, USA. E-mail: [email protected] Leukemia (2009) 23, 59–70 & 2009 Macmillan Publishers Limited All rights reserved 0887-6924/09 $32.00 www.nature.com/leu
Transcript of Enhanced cytotoxicity of an anti-transferrin receptor IgG3-avidin fusion protein in combination with...

ORIGINAL ARTICLE
Enhanced cytotoxicity of an anti-transferrin receptor IgG3-avidin fusion protein incombination with gambogic acid against human malignant hematopoietic cells:functional relevance of iron, the receptor, and reactive oxygen species
E Ortiz-Sanchez1, TR Daniels1, G Helguera1, O Martinez-Maza2,3,4, B Bonavida,3,4 and ML Penichet1,3,4
1Division of Surgical Oncology, Department of Surgery, David Geffen School of Medicine, University of California, Los Angeles,CA, USA; 2Department of Obstetrics and Gynecology, David Geffen School of Medicine, University of California, Los Angeles,CA, USA; 3Jonsson Comprehensive Cancer Center, David Geffen School of Medicine, University of California, Los Angeles, CA,USA and 4Department of Microbiology, Immunology, and Molecular Genetics, David Geffen School of Medicine, University ofCalifornia, Los Angeles, CA, USA
The human transferrin receptor (hTfR) is a target for cancerimmunotherapy due to its overexpression on the surface ofcancer cells. We previously developed an antibody-avidinfusion protein that targets hTfR (anti-hTfR IgG3-Av) andexhibits intrinsic cytotoxicity against certain malignant cells.Gambogic acid (GA), a drug that also binds hTfR, inducescytotoxicity in several malignant cell lines. We now report thatanti-hTfR IgG3-Av and GA induce cytotoxicity in a new broaderpanel of hematopoietic malignant cell lines. Our results showthat the effect of anti-hTfR IgG3-Av is iron-dependent whereasthat of GA is iron-independent in all cells tested. In addition, weobserved that GA exerts a TfR-independent cytotoxicity. Wealso found that GA increases the generation of reactive oxygenspecies that may play a role in the cytotoxicity induced by thisdrug. Additive cytotoxicity was observed by simultaneouscombination treatment with these drugs and synergy by usinganti-hTfR IgG3-Av as a chemosensitizing agent. In addition,we found a concentration of GA that is toxic to malignanthematopoietic cells but not to human hematopoietic progenitorcells. Our results suggest that these two compounds may beeffective, alone or in combination, for the treatment of humanhematopoietic malignancies.Leukemia (2009) 23, 59–70; doi:10.1038/leu.2008.270;published online 23 October 2008Keywords: TfR; iron; antibody fusion protein; gambogic acid;hematopoietic malignances; ROS
Introduction
Iron is an important cofactor of enzymes that participate incellular metabolism including those of the respiratory chain andDNA synthesis.1 Iron associated with transferrin is internalizedby cells through transferrin receptor (TfR)-mediated endocyto-sis.2 The TfR family consists of TfR1 (CD71) and TfR2, which aretype II transmembrane glycoproteins. TfR1 is expressed atdifferent levels on the surface of most normal cells in whichthe majority show low TfR1 expression level.2 However,dividing cells such as basal epidermis and intestinal epitheliumcells express high levels of TfR1.2 In contrast, although TfR2mRNA is present in different tissues,3 the TfR2 protein is limitedto hepatocytes and enterocytes of the small intestine.2,4 As
cancer cells require large amounts of iron to maintain their highrate of cell proliferation, TfR1 is overexpressed on their surface.2
The TfR1 overexpression in malignant cells, its ability tointernalize and its central role in the cellular pathology ofhuman cancer make this receptor an attractive target for thetherapy of cancer.2,5
TfR has been targeted by monoclonal antibodies that eitherinduce cytotoxicity2 or are used for drug delivery.5 Our groupdeveloped earlier an antibody fusion protein that specificallyrecognizes hTfR1 and is composed of chicken avidin, geneti-cally fused to the CH3 domain of a mouse/human chimeric IgG3(anti-hTfR IgG3-Av, Figure 1a), to be used as a universal deliverysystem of biotinylated drugs into cancer cells.6–8 Unexpectedly,we discovered that this fusion protein exhibits a significantintrinsic cytotoxicity against certain hematopoietic malignantcell lines compared with the antibody alone (anti-hTfRIgG3).6,7,9
Anti-hTfR IgG3-Av disrupts TfR recycling and redirects it intothe lysosomes where it is presumably degraded, promoting ironstarvation and the induction of apoptosis that can be blocked byiron supplementation.7,9 As avidin is a non-covalent tetramericprotein, the presence of two avidin moieties in anti-hTfRIgG3-Av favors the dimerization of this fusion protein.6 Althoughthe structural requirements responsible for the cytotoxicity of anti-hTfR IgG3-Av are still unclear, the activity of this fusion proteinis considered to be, at least in part, due to its dimeric structure(Figure 1a).7 We have recently shown that anti-hTfR IgG3-Avconjugated to biotinylated saporin (b-SO6), a plant toxin ofSaponaria officinialis, increases its cytotoxicity in malignant cellsand more importantly, overcomes the resistance of cells to thefusion protein alone.9 In addition, as anti-hTfR IgG3-Av specifi-cally binds hTfR1 (not hTfR2), this antibody fusion protein deliversb-SO6 into cells that express hTfR1.8 Therefore, the fusion ofavidin to anti-hTfR IgG3 results in a relevant tool that can be usedin cancer immunotherapy as it elicits cytotoxic effects by itselfand can also deliver biotinylated cytotoxic compounds such asb-SO6 into cancer cells that overexpress hTfR1.
We are also interested in the effect of the combination of anti-hTfR IgG3-Av with non-biotinylated cytotoxic drugs. We havefocused on gambogic acid (GA) as it can also bind hTfR.10 GA isa xanthone isolated from a resin obtained from the Garciniahanburyi tree that is used in oriental traditional medicine(Figure 1b).11 GA induces cytotoxicity in malignant epithelialcell lines and also in malignant hematopoietic cell lines such asJURKAT and HL-60 cells.12–21 GA can also inhibit the growth ofhuman lung carcinoma and hepatoma xenografts in nudemice.21 This drug blocks hTfR internalization and induces
Received 17 April 2008; revised 14 August 2008; accepted 27 August2008; published online 23 October 2008
Correspondence: Dr ML Penichet, Division of Surgical Oncology,Department of Surgery, University of California, UCLA, 10833 LeConte Avenue, CHS 54-140, Box 951782, Los Angeles, California,90095-1782, USA.E-mail: [email protected]
Leukemia (2009) 23, 59–70& 2009 Macmillan Publishers Limited All rights reserved 0887-6924/09 $32.00
www.nature.com/leu

cytotoxicity in an iron-independent manner in JURKAT cells.10
When hTfR is downregulated by RNA interference, the effect ofGA is decreased suggesting that this drug induces cytotoxicitythrough hTfR.10,22
As anti-hTfR IgG3-Av and GA have shown cytotoxicity tocertain malignant cells, the first objective of this study was toevaluate the cytotoxicity of anti-hTfR IgG3-Av and GA in abroader panel of human hematopoietic malignant cell linesincluding leukemia, lymphoma and multiple myeloma cells.The second objective was to analyze the role of iron and hTfR inthe anti-hTfR IgG3-Av- and GA-mediated cytotoxicity. The thirdand last objective was to evaluate the effect of the combinationof both agents to determine if their potential therapeutic usecould be increased. Here, we report the cytotoxicity of anti-hTfRIgG3-Av and GA alone or in combination in a variety ofmalignant hematopoietic cell lines. The toxic effects of eitheragent alone or in combination on normal hematopoieticprogenitor cells were also evaluated.
Material and methods
Antibodies, antibody fusion protein and gambogic acidThe mouse/human chimeric anti-hTfR IgG3-Av was describedearlier6–9 and consists of chicken avidin fused to the CH3
domains of human IgG3 (Figure 1a). This fusion protein wasexpressed in murine myeloma cells, purified and characterizedas described earlier.6,7,23 Phycoerythrin-conjugated anti-humanCD71 (PE anti-hCD71) and its isotype control (phycoerythrin-conjugated mouse IgG2a k) were purchased from BD Pharmi-gen (BD Bioscience, Franklin Lakes, NJ, USA). GA waspurchased from BIOMOL International L.P (MW: 628.75;Plymouth meeting, PA, USA).
Cell linesThe human cell lines, JURKAT (acute T-cell leukemia), HL-60(acute promyelocytic leukemia), IM-9 (EBV-transformedB-lymphoblastoid), U266 (multiple myeloma), RAMOS(American Burkitt’s lymphoma), RAJI and HS-SULTAN (Burkitt’slymphoma) were purchased from ATCC (American Type CultureCollection, Manassas, VA, USA). The cell lines were maintainedin RPMI 1640 medium (Invitrogen Corporation, Carlsbad, CA,USA) supplemented with 10% heat-inactivated fetal bovineserum (Atlanta Biologicals, Lawrenceville, GA, USA) andcultured at 37 1C in 5% CO2. IM-9 and U266 cell lines wereused as controls of highly sensitive and resistant cells to anti-hTfR IgG3-Av-induced cytotoxicity, respectively.7,9 JURKATand HL-60 cell lines were used as positive controls10 tocompare the cytotoxicty of GA in cells not tested earlier.CHO-TRVb-hTfR1 cells (a cell line that only expresses hTfR1)and CHO-TRVb-neo cells (a cell line transfected with the emptyneomycin vector) are derived from CHO-TRVb cells, a TfR-deficient mutant Chinese hamster ovary (CHO) cell line,24 werekindly donated by Dr H Phillip Koeffler of Cedars Sinai MedicalCenter, Los Angeles, CA, USA. The CHO cell lines werecultured in F-12 (Hams’ medium; Invitrogen) medium supple-mented with 10% fetal bovine serum plus 1 mg/ml G418antibiotic as a selectable marker (Invitrogen). Cell images werecaptured using a Zeiss Axivovert 40 CFL PlasDIC InvertedMicroscope using a � 20 objective (Mikron Instruments Inc, SanMarcos, CA, USA) and a Canon PowerShot A620 digital camera(Mikron Instruments Inc.).
Detection of TfR1 by flow cytometryChinese hamster ovary (CHO) cells and hematopoietic cells(4� 105) were incubated with phycoerythrin-conjugated anti-human CD71 (anti-hTfR1) or with the isotype control antibodyphycoerythrin-conjugated mouse IgG2a k for 15 min on ice in0.5% bovine serum albumin and 2 mM ethylenediaminetetraa-cetic acid phosphate-buffered saline. Cells were washedand fixed with 4% paraformaldehyde in phosphate-bufferedsaline. Cells were analyzed on a FACScan Analytic FlowCytometer (BD-Bioscience). Data were analyzed using theWinMDI 2.8 software (The Scripps Research Institute, La Jolla,CA, USA).
Evaluation of cytotoxic effectsProliferation assay. The antiproliferative effects inducedby either anti-hTfR IgG3-Av or GA were measured by the[3H]-Thymidine incorporation assay as described earlier.9 Cellswere incubated with anti-hTfR IgG3-Av or GA for the indicatedtimes, then harvested and the radioactivity was counted asdescribed earlier.9 Negative controls for each agent were testedin each experiment and included their diluents, buffer (150 mM
NaCl, 50 mM Tris-HCl, pH 7.8) for anti-hTfR IgG3-Av ordimethylsulfoxide (DMSO) for GA. The amount of buffer orDMSO added in control treatments was consistent with the
Figure 1 Schematic representation of anti-hTfR IgG3-Av andchemical structure of GA. (a) In the anti-hTfR IgG3-Av, chickenavidin was fused to the carboxy terminus of the heavy chain of mouse/human chimeric anti-hTfR1 IgG3 containing human kappa (k) lightchain constant region. The antibody fusion protein forms homodimersin solution due to non-covalent tetrameric interaction of four avidinmoieties.6 (b) Chemical structure of GA (C38H44O8, MW.: 628.75).The a,b-unsaturated ketone is marked by the punctuated box.12
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
60
Leukemia

highest volume of each drug used in their respective assays. Foriron supplementation, cells were incubated with or without25mM ferric ammonium citrate (Sigma-Aldrich Co., St Louis, MI,USA) simultaneously with each treatment.
Apoptosis assay. Cells (105 per well for hematopoietic cellsand 2000 cells for CHO cells) were incubated in 24-well plates(total volume per well¼ 1 ml) with the different treatments for48 h. Cells were washed in cold phosphate-buffered saline andstained with Alexa Flour 488-labeled Annexin V and propidiumiodide (PI) contained in the Vybrant Apoptosis Assay Kit no.2(Invitrogen) following the procedure suggested by the manu-facturer. In total, 104 events were recorded for each flowcytometry measurement using FACScan Analytic Flow Cyt-ometer (BD-Bioscience). The cells Annexin Vþ /PI� and AnnexinVþ /PIþ were considered as apoptotic cells. The level ofapoptosis of cells treated with anti-hTfR IgG3-Av or GA iscalculated by the subtraction of the total percentage of apoptosisof cells treated with buffer or DMSO alone from the percentageof apoptosis of cells treated with the fusion protein and GA,respectively. The WinMDI 2.8 software (The Scripps ResearchInstitute) was used for data analysis.
Evaluation of the cytotoxic effect induced by thecombination of anti-hTfR IgG3-Av and GA. The effectof the simultaneous combination or the chemosensitizationtreatments of anti-hTfR IgG3-Av and GA was determined by
isobolographic analysis as described.25 For simultaneous treat-ments 0.31, 0.625, 1.25, 2.5, 5.0 and 10 nM anti-hTfR IgG3-Avplus 0.0375, 0.075, 0.15, 0.3 and 0.6mM GA were incubatedsimultaneously at 37 1C for 48 h. In chemosensitizationtreatments, cells were pre-incubated with the same concentra-tions mentioned above of anti-hTfR IgG3-Av at 37 1C for 24 h,followed by the treatment with the same concentrationsmentioned above of GA for additional 24 h. The additiveeffect is shown by the concentration of both agents representedby the points that lie on the straight line that connect theconcentration of both agents required to produce a definedsingle-agent effect (concentration at which 90% of cells showinhibition of proliferation [IC90]). The points that lie belowor above the straight line represent the concentrations ofboth agents that produce synergistic or antagonistic effects,respectively.
Detection of intracellular reactive oxygen speciesgenerationCells treated with 0.3 or 1 mM GA for 6 h were co-incubatedwith or without 3 mM dithiotheritol (Invitrogen) or with orwithout 2000 U/ml catalase from bovine liver (Sigma-Aldrich,St. Louis, MO, USA) to block reactive oxygen species (ROS)induction. As a positive control, a sample of untreated cells wasincubated with 100mM tert-butyl hydroperoxide (Sigma) for30 min at 37 1C. All of the cells from the different treatmentswere incubated with 5mM of the dye 5-(and-6)-carboxy-20,
Buffer
Anti-hTfRIgG3-Av
Alexa fluor 488 labeled annexin V
Pro
pidi
um lo
dide
U266 HL-60 RAJI JURKAT RAMOS HS-SULTAN IM-9
Figure 2 Anti-hTfR IgG3-Av induces cytotoxic effects in malignant hematopoietic cells. (a) Proliferation assay of hematopoietic malignant cellsincubated with 10 nM anti-hTfR IgG3-Av or buffer for 48 h. The punctuated line represents the percentage of [3H]-Thymidine incorporation in cellstreated with buffer alone. Data shown are the average of quadruplicates and are representative of three independent experiments. Error barsindicate the standard deviation. Statistically significant (Po0.001) growth inhibition compared with buffer alone treated cells is marked (*).(b) Induction of apoptosis in the panel of cells tested above incubated with 10 nM anti-hTfR IgG3-Av for 48 h. The percentage of cells in eachquadrant is localized in the corner. The cells in the right quadrants (Annexin Vþ /PI� and Annexin Vþ /PIþ ) are considered as apoptotic cells.Data are representative of three independent experiments.
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
61
Leukemia

70-dichlorodihydrofluorescein diacetate (CM-H2DCFDA:Invitrogen) in serum-free medium at 37 1C for 20 min. Cellswere washed and analyzed by flow cytometry using FACScanAnalytic Flow Cytometer (BD-Bioscience). Data were analyzedusing the WinMDI 2.8 software (The Scripps Research Institute).The oxidation of CM-H2DCFDA is detected by the increaseof fluorescence that is proportional to the amount of totalintracellular ROS generated. The value of ROS generation isrepresented as the mean of relative fluorescence units.
Human colony forming assayThe human colony forming assay was performed to determinethe effect of GA, anti-hTfR IgG3-Av or their combination onnormal human hematopoietic progenitor cells as describedearlier.9 Human bone marrow mononuclear cells werepurchased from StemCell Technologies (StemCell TechnologiesInc., Vancouver, BC, Canada) and were plated in quadruplicatewith diluent alone (buffer or DMSO: negative controls),GA (0.1, 0.3, 0.5 or 1 mM), anti-hTfR IgG3-Av (1 or 10 nM) ortheir combination. In total, 2� 104 cells were seeded per 35 mmdish in MethoCult GF H4434 (‘Complete’ MethylcelluloseMedium with Recombinant Cytokines and Erythropoietin;StemCell Technologies). Cells were incubated for 14 days at37 1C in 5% CO2 following the instructions of the manufacturer.
The number of CFU-E (colony forming unit-erythroid, matureerythroid progenitors), BFU-E (burst forming unit-erythroid,more primitive progenitor than CFU-E), CFU-GM (colonyforming unit-granulocyte/macrophage, more mature than CFU-GEMM) and CFU-GEMM (colony forming unit granulocyte/erythroid/macrophage/megakaryocyte) was determined using anOlympus CK2 inverted microscope (Olympus America Inc.,Center Valley, PA, USA) and the criteria defined by StemCellTechnologies for each colony type.
Statistical analysisThe student’s t-test was used to determine the significantdifferences in the data, using Microsoft Excel 2004 (MicrosoftCo., Redmond, WA, USA). Po0.05 were considered to besignificant.
Results
Cytotoxic effect of anti-hTfR IgG3-Av in hematopoieticmalignant cell linesWe have reported earlier the cytotoxic effect of anti-hTfRIgG3-Av against myeloma and B-lymphoblastoid cell lines.7,9
To explore the cytotoxic effect of anti-hTfR IgG3-Av in a new
U266
Alexa fluor 488 labeled annexin V
Pro
pidi
um lo
dide
HL-60 RAJI JURKAT RAMOS HS-SULTAN IM-9
DMSO
0.3 µM GA
0.39
2.89
Figure 3 GA induces cytotoxic effects in hematopoietic malignant cells. (a) Anti-proliferative effect of GA at the concentrations and timesindicated in IM-9 cells. Cell proliferation was determined by [3H]-Thymidine incorporation assay. (b) Proliferation assay of hematopoieticmalignant cells incubated with 0.3 mM GA for 48 h. The punctuated line represents the percentage of [3H]-Thymidine incorporation in cells treatedwith DMSO alone. Data represent the average of quadruplicates and are representative of three independent experiments. Statistically significant(Po0.001) growth inhibition compared with DMSO alone treated cells is marked (*). Error bars indicate the standard deviation. (c) Detection ofapoptosis in the panel of cells tested above incubated with 0.3 mM GA or DMSO alone for 48 h. The percentage of cells in each quadrant islocalized in the corner. The cells in the right quadrants (Annexin Vþ /PI� and Annexin Vþ /PIþ ) are considered as apoptotic cells. Data representsthe results of three independent experiments. DMSO, dimethylsulfoxide; GA, gambogic acid.
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
62
Leukemia
broader panel of human hematopoietic malignant cell linesincluding leukemia and lymphoma cells, we first evaluatedthe expression level of hTfR1 on the surface of the cells. Theanti-hTfR IgG-Av binds specifically to TfR1,8 which is expressedon the surface of all hematopoietic cell lines used in thisstudy (Supplementary Figure 1). This fusion protein elicited asignificant (P o 0.001) antiproliferative effect in all lymphomaand leukemia cell lines tested (Figure 2a). It is important to notethat although the IM-9 cell line has a lower TfR1-expressionlevel compared with RAJI and HL-60 (Supplementary Figure 1),IM-9 cells are highly sensitive to the fusion protein. Changes incell morphology suggest that the treatment with anti-hTfRIgG3-Av induces cell death (Supplementary Figure 2). Inductionof cell death by anti-hTfR IgG3-Av was confirmed by an increaseof apoptosis (Annexin Vþ /PI�, early apoptotic cells; andAnnexin Vþ /PIþ , late apoptotic cells) in the panel of the celllines tested with the exception of U266 cells that also expresshTfR1 (resistant to the fusion protein as reported earlier7,9)(Figure 2b). Different sensitivity levels of apoptosis ranging from10 to 40% compared with buffer alone was observed, however,it is important to note that although the level of apoptosisinduced by anti-hTfR IgG3-Av is low in some cancer cell lines,cells in the bottom left quadrant (Annexin V�/PI�) show a shift tothe right compared with buffer alone treated cells suggestingthat the apoptosis has been initiated in most cells.
Cytotoxic effect of GA in hematopoietic malignant celllinesThe cytotoxic effect of GA, a drug that also binds hTfR1,10 hasbeen tested in several cancer cell lines. We analyzed the toxiceffect of GA over a period of 72 h in the panel of malignanthematopoietic cell lines at concentrations below 1 mM used inprevious studies. Figure 3a shows the dose–response of GA overtime in IM-9 cells. Cells incubated with 0.3mM GA, concentra-tion chosen for subsequent experiments, for 48 h blocked80–95% of the proliferation in all the cell lines tested includingU266 cells (Figure 3b), which are resistant to anti-hTfR IgG3-Av(Figure 2). At this concentration, GA also induces high levels ofapoptosis ranging from 30 to 80% compared with the innocuouseffect of DMSO alone in all cell lines (Figure 3c), which wasconsistent with the morphological changes observed by thetreatment with this drug (Supplementary Figure 2).
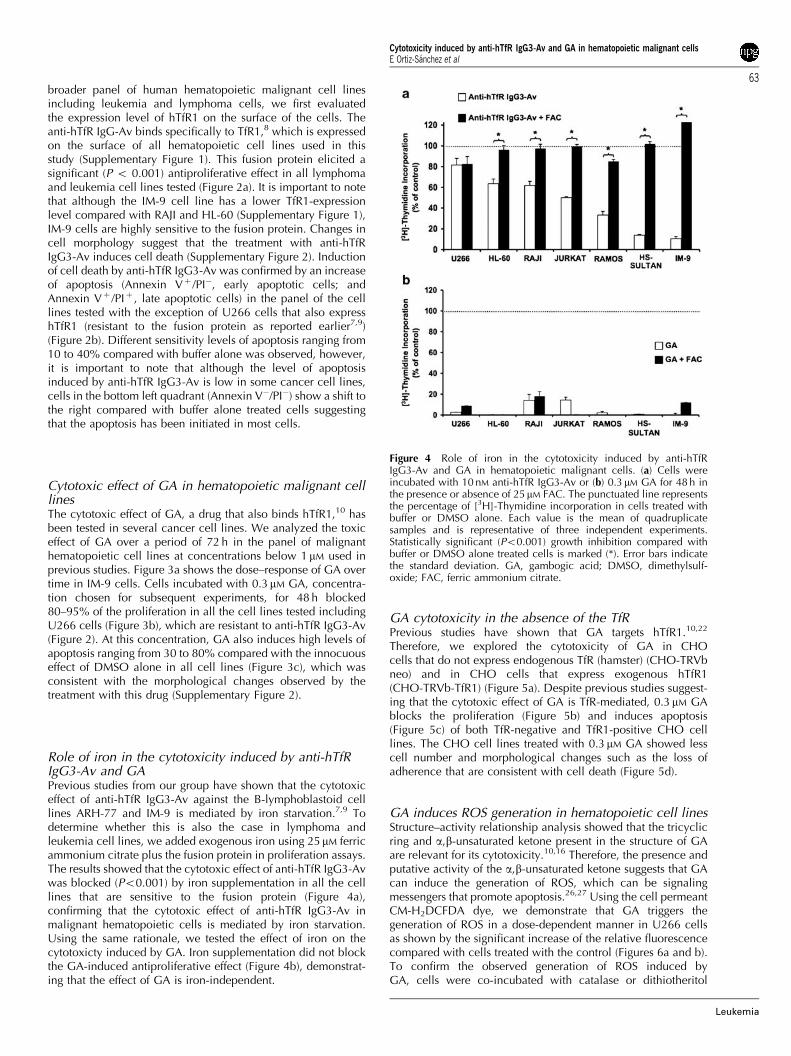
Role of iron in the cytotoxicity induced by anti-hTfRIgG3-Av and GAPrevious studies from our group have shown that the cytotoxiceffect of anti-hTfR IgG3-Av against the B-lymphoblastoid celllines ARH-77 and IM-9 is mediated by iron starvation.7,9 Todetermine whether this is also the case in lymphoma andleukemia cell lines, we added exogenous iron using 25mM ferricammonium citrate plus the fusion protein in proliferation assays.The results showed that the cytotoxic effect of anti-hTfR IgG3-Avwas blocked (Po0.001) by iron supplementation in all the celllines that are sensitive to the fusion protein (Figure 4a),confirming that the cytotoxic effect of anti-hTfR IgG3-Av inmalignant hematopoietic cells is mediated by iron starvation.Using the same rationale, we tested the effect of iron on thecytotoxicty induced by GA. Iron supplementation did not blockthe GA-induced antiproliferative effect (Figure 4b), demonstrat-ing that the effect of GA is iron-independent.
GA cytotoxicity in the absence of the TfRPrevious studies have shown that GA targets hTfR1.10,22
Therefore, we explored the cytotoxicity of GA in CHOcells that do not express endogenous TfR (hamster) (CHO-TRVbneo) and in CHO cells that express exogenous hTfR1(CHO-TRVb-TfR1) (Figure 5a). Despite previous studies suggest-ing that the cytotoxic effect of GA is TfR-mediated, 0.3mM GAblocks the proliferation (Figure 5b) and induces apoptosis(Figure 5c) of both TfR-negative and TfR1-positive CHO celllines. The CHO cell lines treated with 0.3mM GA showed lesscell number and morphological changes such as the loss ofadherence that are consistent with cell death (Figure 5d).
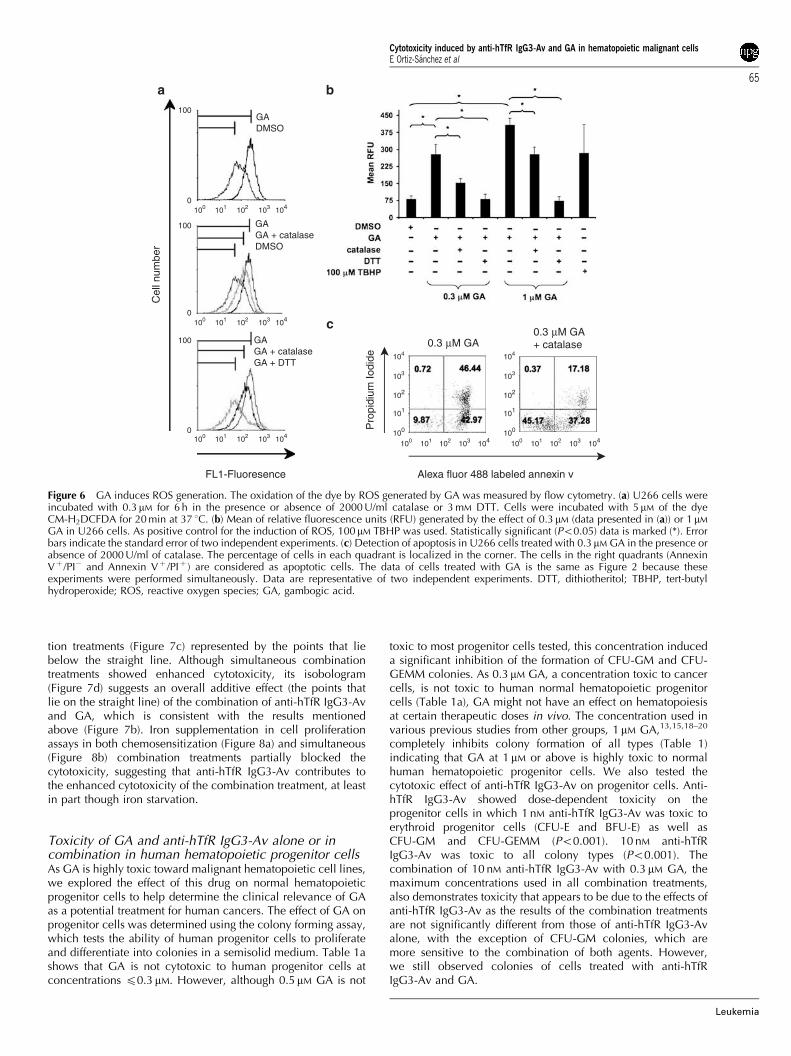
GA induces ROS generation in hematopoietic cell linesStructure–activity relationship analysis showed that the tricyclicring and a,b-unsaturated ketone present in the structure of GAare relevant for its cytotoxicity.10,16 Therefore, the presence andputative activity of the a,b-unsaturated ketone suggests that GAcan induce the generation of ROS, which can be signalingmessengers that promote apoptosis.26,27 Using the cell permeantCM-H2DCFDA dye, we demonstrate that GA triggers thegeneration of ROS in a dose-dependent manner in U266 cellsas shown by the significant increase of the relative fluorescencecompared with cells treated with the control (Figures 6a and b).To confirm the observed generation of ROS induced byGA, cells were co-incubated with catalase or dithiotheritol
Figure 4 Role of iron in the cytotoxicity induced by anti-hTfRIgG3-Av and GA in hematopoietic malignant cells. (a) Cells wereincubated with 10 nM anti-hTfR IgG3-Av or (b) 0.3 mM GA for 48 h inthe presence or absence of 25mM FAC. The punctuated line representsthe percentage of [3H]-Thymidine incorporation in cells treated withbuffer or DMSO alone. Each value is the mean of quadruplicatesamples and is representative of three independent experiments.Statistically significant (Po0.001) growth inhibition compared withbuffer or DMSO alone treated cells is marked (*). Error bars indicatethe standard deviation. GA, gambogic acid; DMSO, dimethylsulf-oxide; FAC, ferric ammonium citrate.
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
63
Leukemia

simultaneously. The co-incubation of GA with catalase, anenzyme that protects the cells from oxidative stress by breakingdown hydrogen peroxide (H2O2) into water and oxygen,significantly decreased the amount of ROS generated by GAmeasured directly by flow cytometry (Figure 6a and b). Inaddition, the co-incubation of GA with dithiotheritol, a reducingagent that inhibits oxidative reactions, decreased the amount ofROS to basal levels (Figures 6a and b). GA-induced apoptosiswas partially blocked in U266 cells (Figure 6c) as well as in IM-9, HL-60 and RAMOS cells (Supplementary Figure 3) byco-incubation with catalase. These findings suggest that thegeneration of ROS plays a significant role in the apoptosisinduced by GA in malignant hematopoietic cells.
Anti-hTfR IgG3-Av enhances the cytotoxic effect of GAon malignant hematopoietic cell linesAs we demonstrated that both anti-hTfR IgG3-Av and GA inducecytotoxic effects in human malignant hematopoietic cells, we
analyzed the effect of the combination of these drugs. Thecombination of both components showed an enhanced anti-proliferative effect compared with either compound alone(Figure 7 for IM-9 cells and Supplementary Table 1 for thepanel of cell lines). All cells pre-treated with 10 nM anti-hTfRIgG3-Av, but not with 2.5 nM of the fusion protein, were moresensitive to the effect of GA (chemosensitization treatments,Figure 7a for IM-9 cells and Supplementary Table 1a for thepanel of cell lines). These results were confirmed by theincreased apoptosis observed in IM-9 cells in the chemosensi-tization treatments of anti-hTfR IgG3-Av and GA (Supplemen-tary Figure 4). The simultaneous combination of 2.5 or 10 nM
anti-hTfR IgG3-Av and 0.1 mM GA enhances the antiproliferativeeffect observed in most of the malignant cells tested (Figure 7bfor IM-9 cells and Supplementary Table 1b for the panel of celllines), however, this enhanced effect was less compared withthat observed in chemosensitization treatments. Isobologramanalysis showed synergistic enhancement of cytotoxicity by thecombination of GA and anti-hTfR IgG3-Av in chemosensitiza-
CHO-TRVb-neo
PE CD71 fluorescence
Pro
pidi
um lo
dide
CHO-TRVb-hTfR1 CHO-TRVb-hTfR1
Untreated
DMSO
0.3 µM GA
Alexa fluor 488 labeled annexin V
CHO-TRVb-neo
Cel
l num
ber
Figure 5 GA induces cytotoxic effects in the absence of TfR. (a) CHO-TRVb-neo (no TfR expression) or CHO-TRVb-hTfR1 (exogenous hTfR1(hCD71) expression) cells were incubated with PE-conjugated anti-hCD71 (thick lines) or PE-conjugated isotype control antibody (thin lines).CD71 cell surface expression correlates with the increase of relative fluorescence that was detected by flow cytometry. (b) Proliferation assay inCHO-TRVb-neo and CHO-TRVb-hTfR1 cells incubated with 0.3 mM GA or DMSO alone for 48 h. Statistically significant (Po0.001) growthinhibition compared to DMSO alone treated cells is marked (*). Error bars indicate the standard deviation. (c) Detection of apoptosis in theindicated CHO cells incubated with 0.3 mM GA or DMSO alone for 48 h. The percentage of cells in each quadrant is localized in the corner. Thecells in the right quadrants (Annexin Vþ /PI� and Annexin Vþ /PIþ ) are considered as apoptotic cells. (d) Morphological changes of the cells treatedwith 0.3 mM GA for 48 h analyzed by differential interferential contrast microscopy using a � 20 objective. Data are representative of twoindependent experiments. GA, gambogic acid; DMSO, dimethylsulfoxide; TfR, transferrin receptor; CHO, Chinese hamster ovary;PE, phycoerythrin.
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
64
Leukemia
tion treatments (Figure 7c) represented by the points that liebelow the straight line. Although simultaneous combinationtreatments showed enhanced cytotoxicity, its isobologram(Figure 7d) suggests an overall additive effect (the points thatlie on the straight line) of the combination of anti-hTfR IgG3-Avand GA, which is consistent with the results mentionedabove (Figure 7b). Iron supplementation in cell proliferationassays in both chemosensitization (Figure 8a) and simultaneous(Figure 8b) combination treatments partially blocked thecytotoxicity, suggesting that anti-hTfR IgG3-Av contributes tothe enhanced cytotoxicity of the combination treatment, at leastin part though iron starvation.
Toxicity of GA and anti-hTfR IgG3-Av alone or incombination in human hematopoietic progenitor cellsAs GA is highly toxic toward malignant hematopoietic cell lines,we explored the effect of this drug on normal hematopoieticprogenitor cells to help determine the clinical relevance of GAas a potential treatment for human cancers. The effect of GA onprogenitor cells was determined using the colony forming assay,which tests the ability of human progenitor cells to proliferateand differentiate into colonies in a semisolid medium. Table 1ashows that GA is not cytotoxic to human progenitor cells atconcentrations p0.3 mM. However, although 0.5 mM GA is not
toxic to most progenitor cells tested, this concentration induceda significant inhibition of the formation of CFU-GM and CFU-GEMM colonies. As 0.3mM GA, a concentration toxic to cancercells, is not toxic to human normal hematopoietic progenitorcells (Table 1a), GA might not have an effect on hematopoiesisat certain therapeutic doses in vivo. The concentration used invarious previous studies from other groups, 1 mM GA,13,15,18–20
completely inhibits colony formation of all types (Table 1)indicating that GA at 1 mM or above is highly toxic to normalhuman hematopoietic progenitor cells. We also tested thecytotoxic effect of anti-hTfR IgG3-Av on progenitor cells. Anti-hTfR IgG3-Av showed dose-dependent toxicity on theprogenitor cells in which 1 nM anti-hTfR IgG3-Av was toxic toerythroid progenitor cells (CFU-E and BFU-E) as well asCFU-GM and CFU-GEMM (Po0.001). 10 nM anti-hTfRIgG3-Av was toxic to all colony types (Po0.001). Thecombination of 10 nM anti-hTfR IgG3-Av with 0.3 mM GA, themaximum concentrations used in all combination treatments,also demonstrates toxicity that appears to be due to the effects ofanti-hTfR IgG3-Av as the results of the combination treatmentsare not significantly different from those of anti-hTfR IgG3-Avalone, with the exception of CFU-GM colonies, which aremore sensitive to the combination of both agents. However,we still observed colonies of cells treated with anti-hTfRIgG3-Av and GA.
Cel
l num
ber
Pro
pidi
um Io
dide
0.3 µM GA
Alexa fluor 488 labeled annexin v
0.3 µM GA+ catalase
GADMSO
GAGA + catalaseDMSO
GAGA + catalaseGA + DTT
FL1-Fluoresence
100
100
0
0
100
0
Figure 6 GA induces ROS generation. The oxidation of the dye by ROS generated by GA was measured by flow cytometry. (a) U266 cells wereincubated with 0.3mM for 6 h in the presence or absence of 2000 U/ml catalase or 3 mM DTT. Cells were incubated with 5mM of the dyeCM-H2DCFDA for 20 min at 37 1C. (b) Mean of relative fluorescence units (RFU) generated by the effect of 0.3 mM (data presented in (a)) or 1 mM
GA in U266 cells. As positive control for the induction of ROS, 100 mM TBHP was used. Statistically significant (Po0.05) data is marked (*). Errorbars indicate the standard error of two independent experiments. (c) Detection of apoptosis in U266 cells treated with 0.3mM GA in the presence orabsence of 2000 U/ml of catalase. The percentage of cells in each quadrant is localized in the corner. The cells in the right quadrants (AnnexinVþ /PI� and Annexin Vþ /PIþ ) are considered as apoptotic cells. The data of cells treated with GA is the same as Figure 2 because theseexperiments were performed simultaneously. Data are representative of two independent experiments. DTT, dithiotheritol; TBHP, tert-butylhydroperoxide; ROS, reactive oxygen species; GA, gambogic acid.
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
65
Leukemia

Discussion
Initially, anti-hTfR IgG3-Av was developed by our group as auniversal drug delivery system.6,8,9 Our previous studies inmultiple myeloma (MM. 1S, S6B45, OCI-My5, 8226/S, 8226/DOX40), B-lymphoblastoid (IM-9 and ARH-77), and K562human erythroleukemia cell lines, demonstrated that anti-hTfRIgG3-Av elicits significant cytotoxic effects by itself.6,7,9 In the
present work, we demonstrate that anti-hTfR IgG3-Av is alsocytotoxic to a broader variety of human malignant hematopoie-tic cells including leukemia (HL-60 and JURKAT) and B-celllymphoma (RAMOS, HS-SULTAN and RAJI) cell lines. Wefound that this cytotoxicity is iron-dependent consistent with ourprevious reports using IM-9 and ARH-77 cells,7,9 suggesting thatiron starvation may be a general mechanism of cytotoxicityinduced by anti-hTfR IgG3-Av in human malignant hematopoietic
Figure 7 The combination of anti-hTfR IgG3-Av and GA enhances the cytotoxic effects in IM-9 cells. IM-9 cells were treated as follows:(a) Chemosensitization treatments. For cytotoxocity induced by GA alone, cells were incubated for 24 h in medium alone followed by theincubation with 0.1 or 0.3 mM GA for additional 24 h; for the cytotoxicity mediated by anti-hTfR IgG3-Av, cells were incubated with 2.5 or 10 nM ofthe fusion protein alone for 48 h; for chemosensitization treatment combinations, cells were pre-incubated with 2.5 or 10 nM of anti-hTfR IgG3-Avalone for 24 h followed by the incubation with 0.1 or 0.3mM GA for additional 24 h in the presence of anti-hTfR IgG3-Av. (b) Simultaneouscombination treatments. Cells were treated with 0.1 mM GA and 2.5 or 10 nM anti-hTfR IgG3-Av for 48 h as controls. For the combinationtreatments, cells were treated with 0.1 mM GA and 2.5 or 10 nM anti-hTfR IgG3-Av simultaneously for 48 h. The punctuated line represents thepercentage of [3H]-Thymidine incorporation in cells treated with buffer alone. Each value is the mean of quadruplicate samples and isrepresentative of three independent experiments. Error bars indicate the standard deviation. Statistically significant growth inhibition comparedwith buffer plus DMSO treated cells is marked (**Po0.001 or *Po0.05). (c) Isobologram curve obtained from the chemosensitization treatmentswith 0.0375, 0.075, 0.15, 0.3 or 0.6 mM GA with 1.25, 2.5, 5 or 10 nM anti-hTfR IgG3-Av, shows the synergistic effect of these drugs.(d) Isobologram curve obtained from the simultaneous combination of anti-hTfR IgG3-Av and GA with the same concentrations used in (c), showsa mostly an additive effect of these drugs. The concentrations of each drug in the combination that produce the specified effect is expressed as afractional inhibitory concentration (F.C.I.) that produce the same effect by the treatment with each drug alone. Points below and above to thestraight line mean synergy or antagonism respectively. The additive effect is represented by the points that lie on the straight line. GA, gambogicacid; DMSO, dimethylsulfoxide.
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
66
Leukemia

cells. Even though all cell lines tested express significant levelsof hTfR1, the variable sensitivity to anti-hTfR IgG3-Av did notcorrelate with the level of TfR1 cell surface expression. Studiesfrom our group are now in progress to analyze this variablesensitivity described in this report and earlier.7 Toxicity of anti-hTfR IgG3-Av on normal hematopoiesis (bone marrow mono-nuclear cells for a 14-day incubation) showed a cytotoxic effectas evidenced by a significant decrease in colony formation of allcolony types tested. Our previous results showed that 1 nM anti-hTfR IgG3-Av does not have cytotoxic effects on humanprogenitor cells if they are treated for 1 h in liquid culturefollowed by washing to remove unbound fusion protein prior tothe 14-day incubation in semi-solid medium required for colonyformation.9 The colony forming assay tests the toxicity of a
compound on late progenitor cells, many of which expresshTfR1 (CD71) at various stages of differentiation.2,28 Therefore,it is expected that anti-hTfR IgG3-Av would have an effect onthese proliferating cells during the extended 14-day incubationperiod. The hematopoietic pluripotent stem cells and the earlyprogenitor cells may be less or not sensitive to the fusion proteinbecause these cells express low to no TfR.28–31 In fact, earlyprogenitor cells are distinguished from late progenitor cells bysurface antigen expression, including the lack of hTfR1expression.28,29,31 The low number of pluripotent cells in thebone marrow makes it difficult to study their sensitivity to anti-hTfR IgG3-Av. In this case, although anti-hTfR IgG3-Av hadcytotoxic effects on committed progenitor cells, it is expectedthat they could be repopulated by their early counterparts.However, a transient myelosuppresion may be expected for theduration of the treatment in patients. Other anticancer drugssuch as homoharringtonine and the aurora kinase inhibitor(VX-680), used for the treatment of hematopoietic malignancesincluding leukemia and multiple myeloma, have shown a dose–response cytotoxicity on human progenitor cells in vitro.32–34
Immunogenicity of potential biological therapeutic agentssuch as antibodies and antibody fusion proteins is always aconcern. In our case, to avoid the immune response that is oftenobserved with the use of murine therapeutic monoclonalantibodies, anti-hTfR IgG3-Av was developed using humanIgG3 and human kappa (k) light chain constant regions. Inaddition, the human Fc can elicit effector functions such ascomplement-dependent cytotoxicity and antibody-dependentcell-mediated cytotoxicty in patients.35 The chicken avidinpresent in anti-hTfR IgG-Av is potentially immunogenic.However, avidin was used instead of the highly immunogenicbacterial protein streptavidin.6,35 The frequent exposure ofhumans to avidin by eating eggs, may decrease its immuno-genecity compared with streptavidin, as it is known that oralantigens induce tolerance.36 Consistent with this possibility,there is a report describing the use of avidin/111Indium-biotin asan imaging method for diagnosis of prosthetic vascular graftinfection. In this study, immunogenicity analysis showed thatnone of the 25 patients studied developed a human anti-avidinhumoral response.37 However, in case that anti-hTfR IgG3-Avinduces an antibody response in patients, this response can beprevented by the use of rituximab (Rituxan, Genentech Inc., SanFrancisco, CA, USA and Biogen IDEC, San Diego, CA, USA), atherapeutic antibody used to treat B-cell non-Hodgkin’slymphoma,38 which also depletes normal B cells and, therefore,the humoral immunity.39,40 Additionally, patients with hemato-poietic malignances such as multiple myeloma and lymphomashow an impaired immune response;41,42 therefore, theimmunogenicity of anti-hTfR IgG3-Av may not be an issue inthese patients.
Previous reports have shown that GA binds hTfR110,22 andblocks its internalization.10 In addition, it has been reported thatGA induces apoptosis in several types of cancer cells, includingJURKAT and HL-60 cell lines.12–22 In this study, we demonstratethe cytotoxicity of GA on malignant hematopoietic cell linesincluding multiple myeloma, lymphoma and leukemia. Ironsupplementation showed that the cytotoxic effect of GA wasiron-independent, as previously described in JURKAT cells.10 Itis important to note that we observed a strong cytotoxic effect ofGA using a lower concentration (0.3 mM GA) compared withthose of previous studies conducted by other groups that usedconcentrations of at least 1 mM GA. Moreover, using humancolony-forming assays, we found that 1 mM GA inhibits thegrowth of human hematopoietic progenitor cells. Thus, it ispossible that the use of GA at concentrations at or above 0.5 mM
Figure 8 Role of iron in the combination of anti-hTfR IgG3-Av andGA in IM-9 cells. IM-9 cells were treated in the presence or absence of25mM FAC in the following treatments: (a) Chemosensitizationtreatments. For cytotoxocity induced by GA alone, cells wereincubated for 24 h in medium alone followed by the incubation with0.1 or 0.3 mM GA for additional 24 h; for the cytotoxicity mediated byanti-hTfR IgG3-Av, cells were incubated with 2.5, or 10 nM of thefusion protein alone for 48 h; for chemosensitization treatmentcombinations, cells were pre-incubated with 2.5 or 10 nM ofanti-hTfR IgG3-Av alone for 24 h followed by the incubation with0.1 or 0.3 mM GA for additional 24 h in the presence of anti-hTfR IgG3-Av. (b) Simultaneous combination treatments. Cells were simulta-neously treated with 0.1 mM GA and 2.5 or 10 nM anti-hTfR IgG3-Avfor 48 h as controls. For the combination treatments, cells were treatedwith 0.1 mM GA and 2.5 or 10 nM anti-hTfR IgG3-Av simultaneously for48 h. The punctuated line represents the percentage of [3H]-Thymidineincorporation in cells treated with buffer alone. Error bars indicate thestandard deviation. Statistically significant growth inhibition comparedwith buffer plus DMSO treated cells is marked (**Po0.001 or*Po0.05). Each value is the mean of quadruplicate samples and isrepresentative of three independent experiments. GA, gambogic acid;DMSO, dimethylsulfoxide; FAC, ferric ammonium citrate.
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
67
Leukemia

would induce severe hematopoietic toxicity in patients.However, in this study we demonstrated that there is aneffective concentration range, where GA can specifically induceapoptosis in malignant hematopoietic cells without toxicity tonormal hematopoiesis. Toxicologic studies have demonstratedthe innocuous effect of GA in rats, mice and dogs,43–45
suggesting that GA can be a viable treatment option for multiplemyeloma, lymphoma and leukemia in humans.
It has been suggested that GA induces cytotoxic effectsthrough its ability to target hTfR1.10,22 In the present study, weshow that GA induces cytotoxicity independent of cell surfaceTfR expression as demonstrated using CHO cells expressinghTfR1or not. Our results are consistent with several mechanismsof cytotoxicity mediated by GA such as the induction of cellcycle arrest in G2/M phase19,20 by the deregulation of CDK7phosphorylation, a cell cycle kinase of the mitosis-promotingfactor.19 In addition, GA increases the level of Bax(a pro-apoptotic protein) and decreases the level of Bcl-2(anti-apoptotic protein).17,20 GA also blocks the activity of thetranscription factor NF-kB through inhibition of IkBa kinase22
resulting in the induction of apoptosis. Moreover, GA reducesthe expression of the human telomerase reverse transcriptaseresulting in decreased telomerase activity, which is required tomaintain malignant cell proliferation.12,18,21 Therefore, GA canbe considered to be a pleiotropic drug that induces cytotoxicitythrough multiple mechanisms.
The structure of GA shows the presence of an a,b-unsaturedketone (Figure 1b) that has been shown to play an important rolein the GA cytotoxicity.10 As a,b-unsatured ketone groups arepresent in some drugs that induce apoptosis through thegeneration of ROS,46,47 we explored the possibility that thecytotoxic effects of GA could be also mediated through thegeneration of ROS. We found that GA increases the ROSgeneration in malignant hematopoietic cell lines. In addition,the effect of GA can be partially blocked by its co-incubationwith catalase, an enzyme that decreases the presence of H2O2, apotent ROS. The partial protective effect of catalase is related tothe specific break down of H2O2, suggesting the presence ofother ROS, such as hydroxyl (.OH), alkoxyl (RO.), peroxyl(ROO.), superoxide (O2
. ) or nitroxyl radical (NO.),27 which maybe also generated by GA. As signaling messengers, ROS areinvolved in pathways that result in apoptosis.26,27,48 It has been
shown that ROS induce the oxidation of sulfydryl groups presentin NF-kB, inhibiting its DNA-binding activity in vitro.49–51 Aswe mentioned earlier, GA also inhibits the NF-kB activity byblocking its translocation to the nucleus.22 As we demonstratedthat GA induces ROS generation, we suggest that in additionto its ability to inhibit NF-kB translocation, GA can also blockNF-kB activity through abolishing its DNA-binding activityresulting in cooperative mechanisms of GA-mediated apoptosis.Additionally, it has been reported that GA-mediated apoptosisinvolves the mitochondrial pathway as evidenced by the releaseof cytochrome c,10 which can be related with the abilityof ROS to induce mitochondrial membrane permeability thatresults in the release of pro-apoptotic proteins such asapoptosis inducing factor, caspase activators (Smac/DIABLO)as well as cytochrome c.48 Taken together, our results suggestthat the generation of ROS by GA is an additional mechanisminvolved in the strong cytotoxic effect of this drug in cancercells.
Previous studies demonstrated that anti-hTfR IgG3-Av disruptsthe cycling of hTfR1 leading to its degradation,7 in contrast withGA, which has been reported to block the internalization ofhTfR1.10 Despite the potential antagonist effect of these agents, thesimultaneous combination of anti-hTfR IgG3-Av and GA enhancestheir cytotoxicity mainly by an additive effect in the panel of cellstested, including U266 that is resistant to anti-hTfR IgG3-Av alone,which can be explained by the multiple mechanisms of GA toinduce cytotoxicity including ROS generation. The incompleteblocking effect of iron supplementation showed the partial role ofthis fusion protein in the additive cytotoxic effect. The simulta-neous combination treatment shows an enhancement of thecytotoxicity on CFU-GM progenitor cells compared with thefusion protein alone. Even though the simultaneous combinationtreatment resulted in fewer CFU-E and BFU-E colonies than thoseof anti-hTfR IgG3-Av alone this variation was not significantlydifferent, suggesting that the cytotoxicity observed by thecombination of anti-hTfR IgG3-Av and GA is mostly due to theeffect of the antibody-avidin fusion protein alone. However,colonies are still formed with the treatment of the combination ofanti-hTfR IgG3-Av and GA.
Some reports have shown that therapeutic antibodies, suchas trastuzumab (Herceptin Genentech Inc.) and rituximab(Rituxan), induce chemosensitization of drug-resistant cancer
Table 1 Cytotoxicity of gambogic acid, anti-hTfR IgG3-Av and their combination on human hematopoietic progenitor cells
CFU-E BFU-E CFU-GM CFU-GEMM
(a) Cytotoxicity of GAUntreated 44±10.6 41±4.4 90±13.2 15±1.6DMSO alone 44±4.0 47±7.0 89±7.7 15±4.20.1 mM GA 40±3.7 35±2.1 94±10.0 19±3.10.3 mM GA 40±3.0 39±5.0 89±6.7 12±3.50.5 mM GA 30±4.5 37±6.5 70±2.1* 9±3.0*1.0 mM GA 0* 0** 0** 0**
(b) Cytotoxicity of anti-hTfR IgG3-Av alone or in combination with GAUntreated 37±1.9 26±2.5 91±7.9 9±1.0Buffer alone 42±6.2 28±4.2 92±7.1 8±1.51 nM anti-hTfR IgG3-Av 32±9.9 19±4.7* 56±9.6** 1±1.0**10 nM anti-hTfR IgG3-Av 15±5.3* 5±2.9** 39±6.8** 0**10 nM anti-hTfR IgG3-Av + 0.3 mM GA 11±4.5** 2±1.2** 24±2.9**,# 0**
anti-hTfR IgG3-Av, antibody-avidin fusion protein that targets hTfR; BFU-E: burst forming units-erythroid; CFU-E: colony forming units-erythroid;CFU-GM: colony forming units-granulocyte/macrophage; CFU-GEMM: colony-forming units-granulocyte/erythroid/macrophage/megakaryocyte;GA, gambogic acid.Each value represents the mean of the number of colonies±the s.d. of quadruplicate samples of two independent experiments.*Po0.05, **Po0.001 compared with buffer alone, #Po0.05 compared with anti-hTfR IgG3-Av.
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
68
Leukemia

cells and thus can be used in combination with other antitumoragents to overcome drug resistance.52–54 Although GA alone hasa cytotoxic effect, the pre-treatment with anti-hTfR IgG3-Av cansensitize cells to the effect of GA through a synergisticcytotoxicity. In addition, iron supplementation confirms therole of anti-hTfR IgG3-Av in chemosensitization treatments,because of this fusion protein induces cytotoxicity in aniron-dependent manner, suggesting for the first time that theiron stress induced by anti-hTfR IgG3-Av can sensitize cancercells to GA. These results suggest that in addition to the intrinsiccytotoxicity and delivery capabilities of anti-hTfR IgG3-Av, itmay also be used as a chemosensitizing agent. However, thischemosensitizing effect of anti-hTfR IgG3-Av needs to be furtherexplored using other chemotherapeutic agents to determine ifthis is a general phenomenon. In addition, owing to theuniversal delivery capability of anti-hTfR IgG3-Av, it can alsobe used to deliver drugs that are able to promote chemosensi-tization. In fact, a broad spectrum of therapeutics includingantisense oligonuleotides and toxins have been successfullydelivered into tumor cells by targeting TfR.5
In summary, we have demonstrated the cytotoxicity of anti-hTfR IgG3-Av in new human malignant hematopoietic cell linesincluding leukemia and lymphoma cells through an iron-dependent mechanism. We also demonstrated the cytotoxiceffect of GA in leukemia, lymphoma and multiple myelomacells through an iron-independent mechanism. We show thatGA can also induce cytotoxicity by a hTfR-independent mecha-nism. Moreover, we demonstrate that GA induces the genera-tion of ROS, suggesting that oxidative stress is part of themechanism of action of this pleiotropic drug. We also found aconcentration of GA that is cytotoxic to malignant hemato-poietic cells, but not to human hematopoietic progenitorcells. Importantly, additive cytotoxicity was observed with thesimultaneous combination of anti-hTfR IgG3-Av and GA,and synergistic cytotoxicty in chemosentitization treatments,suggesting these combination strategies may be effectivetreatments for a wide variety hematopoietic malignances.
Acknowledgements
We would like to thank Dr Sara Huerta-Yapez (Hospital Infantil deMexico ‘Federico Gomez’, Mexico City), Dr Mario I Vega(Hospital de Infectologıa ‘Dr Daniel Mendez Hernandez’ CentroMedico ‘La Raza’ del Instituto Mexicano del Seguro Social,Mexico City) and Jose A. Rodrıguez (University of California, LosAngeles) for assistance with isobolographic analysis. We alsowould like to thank Dr H Phillip Koeffler (Cedars Sinai MedicalCenter, Los Angeles, CA) for providing us with the CHO-TRVbcells. Our study was supported in part by the NIH/NCI GrantsCA86915, CA107023 and SPORE in Lymphoma P50CA096888;and also by the NIH/NCI research supplements CA107023-02S1,CA057152-13S1 and Fogarty AITRP AIDS Malignancies ProgramD43-TW000013-S1.
References
1 Andrews NC, Schmidt PJ. Iron homeostasis. Annu Rev Physiol2007; 69: 69–85.
2 Daniels TR, Delgado T, Rodriguez JA, Helguera G, Penichet ML.The transferrin receptor part I: Biology and targeting with cytotoxicantibodies for the treatment of cancer. Clin Immunol 2006; 121:144–158.
3 Kawabata H, Yang R, Hirama T, Vuong PT, Kawano S, Gombart AFet al. Molecular cloning of transferrin receptor 2. A new member
of the transferrin receptor-like family. J Biol Chem 1999; 274:20826–20832.
4 Deaglio S, Capobianco A, Cali A, Bellora F, Alberti F, Righi L et al.Structural, functional, and tissue distribution analysis of humantransferrin receptor-2 by murine monoclonal antibodies and apolyclonal antiserum. Blood 2002; 100: 3782–3789.
5 Daniels TR, Delgado T, Helguera G, Penichet ML. The transferrinreceptor part II: targeted delivery of therapeutic agents into cancercells. Clin Immunol 2006; 121: 159–176.
6 Ng PP, Dela Cruz JS, Sorour DN, Stinebaugh JM, Shin SU, Shin DSet al. An anti-transferrin receptor-avidin fusion protein exhibitsboth strong proapoptotic activity and the ability to deliver variousmolecules into cancer cells. Proc Natl Acad Sci USA 2002; 99:10706–10711.
7 Ng PP, Helguera G, Daniels TR, Lomas SZ, Rodriguez JA, Schiller Get al. Molecular events contributing to cell death in malignanthuman hematopoietic cells elicited by an IgG3-avidin fusion proteintargeting the transferrin receptor. Blood 2006; 108: 2745–2754.
8 Rodriguez JA, Helguera G, Daniels TR, Neacato II, Lopez-ValdesHE, Charles AC et al. Binding specificity and internalizationproperties of an antibody-avidin fusion protein targeting the humantransferrin receptor. J Control Release 2007; 124: 35–42.
9 Daniels TR, Ng PP, Delgado T, Lynch MR, Schiller G, Helguera Get al. Conjugation of an anti transferrin receptor IgG3-avidin fusionprotein with biotinylated saporin results in significant enhance-ment of its cytotoxicity against malignant hematopoietic cells.Mol Cancer Ther 2007; 6: 2995–3008.
10 Kasibhatla S, Jessen KA, Maliartchouk S, Wang JY, English NM,Drewe J et al. A role for transferrin receptor in triggering apoptosiswhen targeted with gambogic acid. Proc Natl Acad Sci USA 2005;102: 12095–12100.
11 Asano J, Chiba K, Tada M, Yoshii T. Cytotoxic xanthones fromGarcinia hanburyi. Phytochemistry 1996; 41: 815–820.
12 Guo QL, Lin SS, You QD, Gu HY, Yu J, Zhao L et al. Inhibition ofhuman telomerase reverse transcriptase gene expression bygambogic acid in human hepatoma SMMC-7721 cells. Life Sci2006; 78: 1238–1245.
13 Lu N, Yang Y, You QD, Ling Y, Gao Y, Gu HY et al. Gambogicacid inhibits angiogenesis through suppressing vascular endo-thelial growth factor-induced tyrosine phosphorylation of KDR/Flk-1. Cancer Lett 2007; 258: 80–89.
14 Qin Y, Meng L, Hu C, Duan W, Zuo Z, Lin L et al. Gambogic acidinhibits the catalytic activity of human topoisomerase IIalphaby binding to its ATPase domain. Mol Cancer Ther 2007; 6:2429–2440.
15 Yang Y, Yang L, You QD, Nie FF, Gu HY, Zhao L et al. Differentialapoptotic induction of gambogic acid, a novel anticancer naturalproduct, on hepatoma cells and normal hepatocytes. Cancer Lett2007; 256: 259–266.
16 Zhang HZ, Kasibhatla S, Wang Y, Herich J, Guastella J, Tseng Bet al. Discovery, characterization and SAR of gambogic acid as apotent apoptosis inducer by a HTS assay. Bioorg Med Chem 2004;12: 309–317.
17 Liu W, Guo QL, You QD, Zhao L, Gu HY, Yuan ST. Anticancereffect and apoptosis induction of gambogic acid in humangastric cancer line BGC-823. World J Gastroenterol 2005; 11:3655–3659.
18 Yu J, Guo QL, You QD, Lin SS, Li Z, Gu HY et al. Repressionof telomerase reverse transcriptase mRNA and hTERT promoterby gambogic acid in human gastric carcinoma cells. CancerChemother Pharmacol 2006; 58: 434–443.
19 Yu J, Guo QL, You QD, Zhao L, Gu HY, Yang Y et al. Gambogicacid-induced G2/M phase cell-cycle arrest via disturbingCDK7-mediated phosphorylation of CDC2/p34 in human gastriccarcinoma BGC-823 cells. Carcinogenesis 2007; 28: 632–638.
20 Zhao L, Guo QL, You QD, Wu ZQ, Gu HY. Gambogic acidinduces apoptosis and regulates expressions of Bax and Bcl-2protein in human gastric carcinoma MGC-803 cells. Biol PharmBull 2004; 27: 998–1003.
21 Wu ZQ, Guo QL, You QD, Zhao L, Gu HY. Gambogic acidinhibits proliferation of human lung carcinoma SPC-A1 cellsin vivo and in vitro and represses telomerase activity andtelomerase reverse transcriptase mRNA expression in the cells.Biol Pharm Bull 2004; 27: 1769–1774.
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
69
Leukemia

22 Pandey MK, Sung B, Ahn KS, Kunnumakkara AB, Chaturvedi MM,Aggarwal BB. Gambogic acid, a novel ligand for transferrinreceptor, potentiates TNF-induced apoptosis through modulationof the nuclear factor-\{kappa\}B signaling pathway. Blood 2007;110: 3517–3525.
23 Helguera G, Penichet ML. Antibody-Cytokine Fusion Proteins forthe Therapy of Cancer. Adoptive Immunotherapy. Methods MolMed 2005; 109: 347–3774.
24 McGraw TE, Greenfield L, Maxfield FR. Functional expression ofthe human transferrin receptor cDNA in Chinese hamster ovarycells deficient in endogenous transferrin receptor. J Cell Biol 1987;105: 207–214.
25 Berenbaum MC. A method for testing for synergy with any numberof agents. J Infect Dis 1978; 137: 122–130.
26 Jacobson MD. Reactive oxygen species and programmed celldeath. Trends Biochem Sci 1996; 21: 83–86.
27 Simon HU, Haj-Yehia A, Levi-Schaffer F. Role of reactive oxygenspecies (ROS) in apoptosis induction. Apoptosis 2000; 5: 415–418.
28 Lansdorp PM, Dragowska W. Long-term erythropoiesis fromconstant numbers of CD34+ cells in serum-free cultures initiatedwith highly purified progenitor cells from human bone marrow.J Exp Med 1992; 175: 1501–1509.
29 Andrews RG, Singer JW, Bernstein ID. Precursors of colony-forming cells in humans can be distinguished from colony-formingcells by expression of the CD33 and CD34 antigens and lightscatter properties. J Exp Med 1989; 169: 1721–1731.
30 Bender JG, Unverzagt K, Walker DE, Lee W, Smith S, Williams Set al. Phenotypic analysis and characterization of CD34+ cellsfrom normal human bone marrow, cord blood, peripheral blood,and mobilized peripheral blood from patients undergoing auto-logous stem cell transplantation. Clin Immunol Immunopathol1994; 70: 10–18.
31 Gross S, Helm K, Gruntmeir JJ, Stillman WS, Pyatt DW, Irons RD.Characterization and phenotypic analysis of differentiating CD34+human bone marrow cells in liquid culture. Eur J Haematol 1997;59: 318–326.
32 Mi Y, Xue Y, Yu W, Liu S, Zhao Y, Meng Q et al. Therapeuticexperience of adult acute myeloid leukemia in a single institutionof China and its relationship with chromosome karyotype. LeukLymphoma 2008; 49: 524–530.
33 Shi Y, Reiman T, Li W, Maxwell CA, Sen S, Pilarski L et al.Targeting aurora kinases as therapy in multiple myeloma. Blood2007; 109: 3915–3921.
34 Umbach GE, Hug V, Spitzer G, Thames H, Drewinko B. Responsesof human bone marrow progenitor cells to fluoro-ara-AMP,homoharringtonine, and elliptinium. Invest New Drugs 1984; 2:263–265.
35 Penichet ML, Morrison SL. Design and Engineering Human Formsof Monoclonal Antibodies. Drug Development Research 2004; 61:121–136.
36 Weiner HL. Oral tolerance. Proc Natl Acad Sci USA 1994; 91:10762–10765.
37 Samuel A, Paganelli G, Chiesa R, Sudati F, Calvitto M, MelissanoG et al. Detection of prosthetic vascular graft infectionusing avidin/indium-111-biotin scintigraphy. J Nucl Med 1996;37: 55–61.
38 McLaughlin P, Grillo-Lopez AJ, Link BK, Levy R, Czuczman MS,Williams ME et al. Rituximab chimeric anti-CD20 monoclonalantibody therapy for relapsed indolent lymphoma: half of patientsrespond to a four-dose treatment program. J Clin Oncol 1998; 16:2825–2833.
39 Frankel AE. Reducing the immune response to immunotoxin. ClinCancer Res 2004; 10 (1 Part 1): 13–15.
40 van der Kolk LE, Baars JW, Prins MH, van Oers MH. Rituximabtreatment results in impaired secondary humoral immune respon-siveness. Blood 2002; 100: 2257–2259.
41 Grulich AE, Vajdic CM, Cozen W. Altered immunity as a riskfactor for non-Hodgkin lymphoma. Cancer Epidemiol BiomarkersPrev 2007; 16: 405–408.
42 Pratt G, Goodyear O, Moss P. Immunodeficiency and immuno-therapy in multiple myeloma. Br J Haematol 2007; 138: 563–579.
43 Guo Q, Qi Q, You Q, Gu H, Zhao L, Wu Z. Toxicological studiesof gambogic acid and its potential targets in experimental animals.Basic Clin Pharmacol Toxicol 2006; 99: 178–184.
44 Hao K, Liu XQ, Wang GJ, Zhao XP. Pharmacokinetics, tissuedistribution and excretion of gambogic acid in rats. Eur J DrugMetab Pharmacokinet 2007; 32: 63–68.
45 Qi Q, You Q, Gu H, Zhao L, Liu W, Lu N et al. Studies on thetoxicity of gambogic acid in rats. J Ethnopharmacol 2008; 117:433–438.
46 Chen YC, Shen SC, Tsai SH. Prostaglandin D(2) and J(2) induceapoptosis in human leukemia cells via activation of the caspase 3cascade and production of reactive oxygen species. BiochimBiophys Acta 2005; 1743: 291–304.
47 Kondo M, Shibata T, Kumagai T, Osawa T, Shibata N, KobayashiM et al. 15-Deoxy-Delta(12,14)-prostaglandin J(2): the endo-genous electrophile that induces neuronal apoptosis. Proc NatlAcad Sci USA 2002; 99: 7367–7372.
48 Fleury C, Mignotte B, Vayssiere JL. Mitochondrial reactive oxygenspecies in cell death signaling. Biochimie 2002; 84: 131–141.
49 Meyer M, Schreck R, Baeuerle PA. H2O2 and antioxidants haveopposite effects on activation of NF-kappa B and AP-1 in intactcells: AP-1 as secondary antioxidant-responsive factor. Embo J1993; 12: 2005–2015.
50 Nakano H, Nakajima A, Sakon-Komazawa S, Piao JH, Xue X,Okumura K. Reactive oxygen species mediate crosstalk betweenNF-kappaB and JNK. Cell Death Differ 2006; 13: 730–737.
51 Toledano MB, Leonard WJ. Modulation of transcription factorNF-kappa B binding activity by oxidation-reduction in vitro.Proc Natl Acad Sci USA 1991; 88: 4328–4332.
52 Lee S, Yang W, Lan KH, Sellappan S, Klos K, Hortobagyi G et al.Enhanced sensitization to taxol-induced apoptosis by herceptinpretreatment in ErbB2-overexpressing breast cancer cells. CancerRes 2002; 62: 5703–5710.
53 Jazirehi AR, Huerta-Yepez S, Cheng G, Bonavida B. Rituximab(chimeric anti-CD20 monoclonal antibody) inhibits the constitu-tive nuclear factor-\{kappa\}B signaling pathway. Cancer Res2005; 65: 264–276.
54 Jazirehi AR, Vega MI, Chatterjee D, Goodglick L, Bonavida B.Inhibition of the Raf-MEK1/2-ERK1/2 signaling pathway, Bcl-xLdown-regulation, and chemosensitization of non-Hodgkin’s lym-phoma B cells by Rituximab. Cancer Res 2004; 64: 7117–7126.
Supplementary Information accompanies the paper on the Leukemia website (http://www.nature.com/leu)
Cytotoxicity induced by anti-hTfR IgG3-Av and GA in hematopoietic malignant cellsE Ortiz-Sanchez et al
70
Leukemia




















