
Crystal Structure of Mycobacterium tuberculosis Catalase-Peroxidase

Inhibition of leukotriene biosynthesis abrogates the host controlof Mycobacterium tuberculosis
Camila M. Peresa, Lúcia de Paulaa, Alexandra I. Medeirosa, Carlos A. Sorgia, Edson G.Soaresb, Daniela Carlosa, Marc Peters-Goldenc, Célio L. Silvad, and Lúcia H. Facciolia,*
a Departamento de Análises Clínicas, Toxicológicas e Bromatológicas, Faculdade de CiênciasFarmacêuticas de Ribeirão Preto, Universidade de Sao Paulo(USP), Av. do Cafe s/n(o), Ribeirao Preto, SP14040-903, Brazil
b Departamento de Patologia, Faculdade de Medicina de Ribeirão Preto (FMRP-USP), Av. Bandeirantes3.900, Ribeirao Preto, SP 14049-900, Brazil
c Division of Pulmonary and Critical Care Medicine, Department of Internal Medicine, Medical School,University of Michigan Health System, 6301 MSRB III, Box 0642, 1150 W. Medical Center Drive, Ann Arbor,MI 48109-0642, USA
d REDE-TB, Departamento de Bioquímica e Imunologia (FMRP – USP), Av. Bandeirantes 3.900, RibeiraoPreto, SP 14049-900, Brazil
AbstractLeukotrienes produced from arachidonic acid by the action of 5-lipoxygenase (5-LO) are classicalmediators of inflammatory responses. Recently, it has been demonstrated that leukotrienes also playan important role in host defense against microorganisms. In vitro studies have shown thatleukotrienes augmented the anti-mycobacterial activity of neutrophils. In this study, we examinedthe role of leukotrienes in regulating host response and cytokine generation in a murine model oftuberculosis. Administration of the 5-LO pathway inhibitor MK 886, which reduced lung levels ofboth leukotriene B4 and the anti-inflammatory substance lipoxin A4 by ~50%, increased 60-daymortality from 14% to ~57% in Mycobacterium tuberculosis-infected mice, and increased lungbacterial burden by ~15-fold. Although MK 886-treated animals exhibited no reduction in pulmonaryleukocyte accumulation, they did manifest reduced levels of nitric oxide generation and of theprotective type 1 cytokines interleukin-12 and gamma interferon. Together our results demonstratethat 5-LO pathway product(s) – presumably leukotrienes- positively regulate protective Th1responses against mycobacterial infection in vivo. Moreover, the immunosuppressive phenotype ininfected mice observed with MK 886 is most consistent with inhibition of an activator (LTB4) ratherthan a suppressor (LXA4) of antimicrobial defense, suggesting the major effect of leukotrienes.
KeywordsM. tuberculosis; 5-LO metabolites; Lung inflammation; Cytokines; Immunomodulation
*Corresponding author. Mailing address: Departamento de Análises Clínicas, Toxicológicas e Bromatológicas, Faculdade de CiênciasFarmacêuticas de Ribeirão Preto, Universidade de São Paulo - Av. do Café, s/n. Ribeirão Preto, São Paulo, Brazil. 14.040-903. Telephonenumber: 55-16-3602 4303, Fax number: 55-16-3633 1936; e-mail: [email protected]'s Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptMicrobes Infect. Author manuscript; available in PMC 2008 November 7.
Published in final edited form as:Microbes Infect. 2007 April ; 9(4): 483–489. doi:10.1016/j.micinf.2007.01.006.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

1. IntroductionTuberculosis (TB) is a classic example of an infection in which the disease process is causedby the immune response directed at the infectious agent. The death of the diseased individuallargely reflects the immunopathology of the cell-mediated immune response. Nevertheless, itis also the cell-mediated response that protects against the disease by arresting, killing, andremoving the multiplying bacteria. An important question that arises from this balance betweenthe protective and harmful effects of the immune response is whether the mediators or elementsof the immune response that protect can be distinguished from those that harm. The essentialrole of an intact Th1 response in host defense against TB is illustrated by reports of a markedlyenhanced susceptibility to TB of mice deficient for gamma interferon (IFN-γ) [1] orinterleukin-12 (IL-12) [2]. Besides the CD4+ T-cell, it has also been shown that the inductionof a population of antigen specific cytotoxic CD8+ T cells producing IFN-γ is important forthe protective response [3]. In contrast to cytokines, the class of potent lipid mediators ofinflammation known as leukotrienes has received far less attention for their role in host defensein this infection. Leukotrienes are derived via the 5-lipoxygenase (5-LO) pathway ofarachidonic acid metabolism. Many studies have demonstrated that leukotriene B4 (LTB4) isa powerful leukotropic, proinflammatory, and immunoregulatory mediator [4–6]. Moreover,it has been shown that these mediators are involved in antimicrobial host defense in a varietyof infectious diseases [7–9]. Elevated levels of LTB4 are found in the bronchoalveolar lavagefluid of patients with bacterial pneumonia [10] and the levels of LTB4 and leukotriene C4(LTC4) are similarly elevated in lung homogenates of animals with experimental bacterialpneumonia [7]. These findings provide evidence of 5-LO pathway activation during the courseof lower respiratory tract infections. Our group has previously demonstrated that leukotrienesare the main chemoattractants involved in the cell migration induced by H. capsulatum or thecell wall fraction thereof [11] and are involved in eliminating pulmonary histoplasmosis [8].Recently, in vitro studies have shown that endogenous synthesis of leukotrienes by neutrophilsfollowing phagocytosis of mycobacteria may play a role in the killing of M. bovis [12]. Lipoxinsrepresent another class of 5-LO- derived eicosanoids, but in contrast to leukotrienes, thesepossess anti-inflammatory properties. It has been shown that high levels of lipoxin A4(LXA4) are generated during infection with Toxoplasma gondii [13].
In the present study, we investigated the effects of 5-LO product inhibition during the immuneresponse in a murine model of pulmonary TB. The results reveal for the first time thatleukotrienes play a role in the protective host response to M. tuberculosis.
2. Materials and methods2.1. Animals
Male or female 5–8-week-old BALB/c mice were obtained from the animal facilities of theFaculdade de Ciências Farmacêuticas – Universidade de São Paulo and were kept under barrierconditions at a level III biohazard laboratory. All experiments were approved and conductedin accordance with guidelines of the Animal Care Committee of the University.
2.2. Preparation of M. tuberculosis, infection and treatment of miceM. tuberculosis H37Rv strain (American Type Culture Collection, Rockville, MD, USA) andinfection were performed as described previously [14]. Briefly, mice were anesthetized withtribromoethanol 2.5%, restrained on a small board and infected by intratracheal route. Animalsreceived either 100 μl of phosphate buffered saline (PBS) or 105 viable CFU of M.tuberculosis H37Rv in 100 μl of PBS. Animals were divided into three groups. Group 1 animalswere subjected to i.t. infection with M. tuberculosis and water (0.5 ml given perorally [p.o.]by gavage) was given 1 h before infection and then every 24 h for 60 days. Group 2 animals
Peres et al. Page 2
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

were treated with MK 886 (Merck Frosst Canada) p.o. (5 mg/kg/0.5 ml) 1 h prior to infectionwith M. tuberculosis and again every 24 h for 60 days. A dose of 5 mg/kg/0.5 ml has previouslybeen shown to inhibit the synthesis of leukotrienes in a model of pulmonary histoplasmosis[8]. Group 3 animals received i.t. injections of PBS and were given water p.o. for 60 days.
2.3. Bronchoalveolar lavage fluid (BALF)On days 2, 4, 7, 15, 30 and 60 following infection, animals were euthanized with sodiumpentobarbitone and lavage fluid was recovered as previously described [8]. Total cell countswere immediately performed in a Neubauer Chamber. Differential counts were obtained usingRosenfeld-stained cytospin preparations [15].
2.4. Quantitation of bacterial growthThe number of live bacteria recovered from the lungs was determined by plating 10-fold serialdilutions of homogenized tissue on Middlebrook 7H11 agar (Difco) and counting colonies after30 days at 37 °C [14].
2.5. Measurement of eicosanoids and cytokinesThe lungs were removed and homogenized (Mixer Homogenizer, Labortechnik, Germany) in2 ml of medium, centrifuged at 1500 g, filtered, and stored at −70 °C until assayed. Lipids werepurified from filtered lung homogenate supernatants with Sep-Pak C18 cartridges according tothe instructions of the manufacturer (Waters Corp., Milford, Mass.). Levels of eicosanoids andcytokines were measured using commercial ELISA kits obtained from Neogen Corp.(LXA4), Cayman Chemical Co. (LTB4) and PharMingen (cytokines).
2.6. Quantitation of nitric oxide (NO)NO production was assessed by measuring the amount of nitrite in lung homogenates, obtainedas described above, using the Greiss reagent [16]. Values were determined using a standardcurve with serial dilutions of NaNO2 (Sigma, St. Louis, MO).
2.7. Statistical analysisData are presented as mean ± SEM. Comparisons were performed with ANOVA followed bythe Bonferroni test using Prism 3.0 statistical program (GraphPad Software, San Diego, CA).Differences were considered significant if P < 0.05.
3. Results3.1. Lung leukotriene production is increased in M. tuberculosis-infected mice and isinhibited with MK 886
Throughout the entire experimental observation period, PBS-inoculated animals expresseddetectable but low levels of LTB4 (Fig. 1). M. tuberculosis-infected animals demonstratedincreased lung LTB4 levels as early as day 2. Overproduction of LTB4 persisted throughoutthe course of the experiment, peaking at day 30. Except on day 7, treatment of infected animalswith MK 886 significantly inhibited LTB4 synthesis compared to infected untreated mice, withthe degree of inhibition averaging ~50% (Fig. 1).
3.2. Inhibition of endogenous leukotriene synthesis is associated with impaired M.tuberculosis clearance and animal survival
The number of M. tuberculosis CFU in the lungs increased during the course of infection (Fig.2A). Recovery of CFU from lungs of infected animals treated with MK 886 increased by 1.73log10 and 1.5 log10 at 30 and 60 days after infection, respectively, in comparison to infected,
Peres et al. Page 3
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

untreated animals (Fig. 2A). In association with the increased mycobacterial burden observedwith MK 886, treated animals also exhibited an increase in 60-day mortality from 14% to 57%(Fig. 2B). In order to evaluate whether the increase in mortality in the treated animals was dueto MK 886 toxicity, i.t. injections of PBS were given to a group of animals who were thensubmitted to p.o. MK 886 treatment for 60 days. 100% of animals inoculated with PBS andtreated with MK 886 survived, as did the group receiving PBS and H2O (data not shown).
3.3. MK 886 has no effect on lung leukocyte recruitmentIntratracheal inoculation of M. tuberculosis induced significant neutrophil accumulation in thelungs at the second day post infection, as judged by BALF numbers. Neutrophil accumulationin this group peaked on day 30 post infection, but still exceeded the control value at 60 daysafter infection. No such accumulation occurred in the control group (Fig. 3A). MK 886abrogated the early BALF neutrophilia seen at day 2, but not the more pronouncedaccumulation at days 30 and 60 (Fig. 3A). Except for day 7, M. tuberculosis elicited significantincreases in BALF mononuclear cells (Fig. 3B). As with neutrophils, MK 886 significantlyinhibited mononuclear cell accumulation at day 2, but not at other time points (Fig. 3B).
3.4. MK 886 reduces levels of protective cytokines in infected miceWe determined whether MK 886 treatment altered cytokines that are involved in inflammatoryand immune responses to M. tuberculosis. In M. tuberculosis-infected animals, TNF-α andIL-6 levels in lungs increased progressively from the second day of infection, reaching theirpeak on day 30 (Fig. 4A and C). Treatment of infected animals with MK 886 decreased thelevels of TNF-α on days 2, 15 and 30 and of IL-6 on days 2 and 7 post infection (Fig. 4A andC). The levels of IL-1 in lungs of M. tuberculosis-infected animals were greater than in controlmice at all time points except day 7 (Fig. 4B). Treatment with MK 886 decreased the IL-1levels only at day 2 post infection (Fig. 4B). Levels of IL-12 in the lungs of M. tuberculosis-infected animals increased from the second day of infection (Fig. 4D), while IFN-γ was onlydetected after day 7 (Fig. 4E). Nevertheless, levels of these cytokines remained above thoseobserved in control animals until day 60. Treatment of infected animals with MK 886 inhibitedsynthesis of IL-12 from day 7 (Fig. 4D) and of IFN-γ from day 30 post-infection (Fig. 4E). Ondays 30 and 60 post infection, production of IL-10 in M. tuberculosis-infected animals washigher than in controls; this was unaffected by treatment with MK 886 (Fig. 4F). Althoughproduction of IL-4 during M. tuberculosis lung infection has been previously described [17],we were unable to detect this Th2 cytokine at any time point, perhaps because of the lowsensitivity of our ELISA assay (data not shown). In PBS-inoculated mice, MK 886 had noeffect on lung levels of IFN-γ, IL-12, TNF-α or IL-1 at day 60 (data not shown).
3.5. Production of NO in lung homogenatesIn comparison to those from uninfected control animals, lung homogenates from infected micecontained increased levels of nitrite at all time points analyzed (Fig. 5). At every time pointexcept day 15, treatment with MK 886 resulted in a significant reduction of NO production,which reached ~69% on day 60 (Fig. 5).
3.6. Production of LXA4 in lung homogenatesTo determine whether MK 886 treatment also affected lipoxin biosynthesis, we quantified thelevels of LXA4 on days 30 and 60 post infection. The infection by M. tuberculosis inducedpronounced synthesis of this anti-inflammatory mediator compared to PBS-inoculated animals(Fig. 6). However, the peak level observed at day 30 was ~50% less than this observed forLTB4 at the same time point (Fig. 1). At 30 days, treatment of infected mice with MK 886resulted in a 54% decrease in LXA4 production (Fig. 6).
Peres et al. Page 4
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

4. DiscussionThe present study for the first time demonstrates the important role that leukotrienes play inthe host immune response to pulmonary TB. Local generation of the proinflammatory lipid,LTB4, was increased ~5- to 15-fold above baseline levels in control infected mice. As expected,the leukotriene synthesis inhibitor MK 886 significantly (but incompletely) inhibited thisendogenous response during infection. Inhibition of LTB4 generation was associated withimpaired mycobacterial clearance from the lungs and increased animal mortality. These resultsare consistent with a pivotal role of 5-LO metabolites in the protective host response seen inthis model of TB, as previously demonstrated by others in various infections, includingbacterial and fungal infections of the lung [7,8].
A recent paper by Bafica et al. [18] showed that mice that were genetically deficient inleukotriene production had improved bacterial clearance and survival in a model of M.tuberculosis; these results were attributed to abrogation of synthesis of the alternative 5-LOmetabolite, lipoxin A4, which has immunosuppressive and anti-inflammatory effects. Wemeasured lung homogenate levels of lipoxin A4 at days 30 and 60 post infection. We foundthat lipoxin A4 production generally paralleled LTB4, with an increase in infected mice thatwas inhibited by ~50% when animals were treated with MK 886. Although MK 886 inhibitedboth LTB4 and LXA4 by only 50%, it is clear that in our model, the fact that it worsenedmycobacterial clearance and animal survival in our experiments can be explained by itsinhibition of immunoprotective 5-LO products (namely, leukotrienes), but not by its inhibitionof immunosuppressive lipoxin A4.
Lipid mediators have been shown to have opposing effect in host defense depending ondifferent experimental conditions [19]. The relative contributions of these two 5-LO productswith opposing actions may be model-specific, and the predominant contribution of leukotrienesin our hands vs. lipoxin A4 in the study of Bafica and colleagues may reflect differencesbetween the two models in mouse strain, infection severity, and genetic vs. pharmacologicapproaches. One important distinction is that LTB4 levels exceeded those of LXA4 in lunghomogenates in our study, while Bafica et al. observed much higher levels of LXA4 thanLTB4 measured in plasma, and did not measure those two lipids in the lungs. It should also bepointed out that these investigators only measured CFU and 5-LO products prior to the onsetof mortality.
Leukotrienes exert potent effects on phagocytosis and killing of microorganisms by bothalveolar macrophages [19] and neutrophils [20]. Indeed, leukotrienes have been reported to benecessary for optimal killing of Mycobacterium bovis by neutrophils [12]. In addition, previousstudies from our group have shown that in Histoplasma capsulatum infection, reductions inleukotriene synthesis caused by administration of MK 886 resulted in 100% 14-day mortalityin mice and increased lung CFU at 7 and 14 days post infection [8]. In light of these findings,we hypothesize that the absence of leukotrienes during infection decreases the antimicrobialaction of neutrophils and mononuclear cells, allowing persistence and growth of the bacilli inthe lung. This may reflect direct effects on killing mechanisms such as generation of NO [8,21], or indirect effects mediated by modulation of Th1 cytokines. Our data confirmed that MK886 interfered with the generation of NO, IL-12 and IFN-γ in the lungs of infected mice.
Several Th1-type cytokines, such as IL-12 and IFN-γ, have been shown to promote the immuneresponse against M. tuberculosis infection in both humans and mice. IFN-γ is essential for thedevelopment of protective immunity, and is probably the most important factor that activatesmacrophages for antimycobacterial action, at least in mice [1]. Other authors have shown thatLTB4 is involved in T-cell activation and upregulation of IL-2, IL-4, IFN-γ, IL-1 and TNF-α[5,8,22]. We therefore examined the effect of MK 886 treatment on M. tuberculosis-infected
Peres et al. Page 5
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

mice, with a focus on lung production of IL-12, IFN-γ, and IL-10 during infection. The resultsdemonstrated that decreased leukotriene biosynthesis was associated with impairment of IL-12and IFN-γ generation. In fact, lung levels of IL-12 and IFN-γ were markedly lower in infectedand MK 886-treated mice than in infected untreated mice. In studies using a neutralizingantibody against IL-12 [23] or IL-12 knockout mice (p35−/−, p40−/−, or double knockout) [2],the lack of IL-12 caused a decrease in the IFN-γ response and an increased susceptibility ofmice to M. tuberculosis infection.
Leukotrienes are directly chemotactic for neutrophils [11] and T lymphocytes [24] and canpromote neutrophil survival as well [25]. They can also contribute to leukocyte recruitmentindirectly by augmenting generation of chemotactic cytokines and chemokines [22,26].However, our BALF quantitation revealed no impairment in neutrophil or total mononuclearcell accumulation in the lungs of infected mice treated with MK 886. Similar findings wereobserved in a model of Klebsiella pneumoniae pneumonia [7]. Inflammatory cytokines suchas IL-1, IL-6, and TNF-α are known to be potent neutrophil chemotactic agents in mice [15].Moreover, TNF-α, IL-1, and IL-6 have been implicated in murine TB [27,28]. Therefore, wecompared the levels of these cytokines in lung homogenates from M. tuberculosis-infectedmice that were treated with MK 886 with those from M. tuberculosis-infected untreated mice.The results showed that M. tuberculosis infection induced release of IL-1, IL-6, and TNF-αinto the lungs in the early and the late phases of infection. However, except for TNF-α, theinhibition of endogenous leukotriene biosynthesis did not regulate production of theseinflammatory cytokines.
In summary, we have demonstrated that M. tuberculosis infection induces local production ofLTB4 in the lungs of mice which persists at least through day 60 post inoculation. Thisendogenous production of leukotrienes is important for development of a protective hostimmune response. Leukotrienes likely contribute to such protection by virtue of their effectson phagocyte ingestion and killing of microbes, processes which are themselves influenced bythe further ability of leukotrienes to promote Th1 cytokine synthesis. As leukotriene productioncan be down-regulated not only by drugs used in the treatment of asthma, but also by suchcircumstances as HIV infection [29] and malnutrition [30], recognition of the role ofleukotrienes in the immune response to TB may be clinically important and may define apotential target for immunomodulatory therapy.
AcknowledgementsThis study was supported by grants from the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP –01/12400-6; 02/12856-2; 00/09663-2), the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq),the Fundação de Amparo ao Ensino, Pesquisa e Assistência do Hospital das Clínicas da FMRP-USP (FAEPA), Brazil,and National Institutes of Health grant HL58897. We thank Ana Paula Masson for preparing of M. tuberculosisinoculum. We are grateful to Merck Frosst Canada Inc. for furnishing the MK 886.
References1. Cooper AM, Dalton DK, Stewart TA, Griffin JP, Russel DG, Orme IM. Disseminated tuberculosis in
interferon gamma gene-disrupted mice. J Exp Med 1993;178:2243–2247. [PubMed: 8245795]2. Cooper AM, Magram J, Ferrante J, Orme IM. Interleukin 12 (IL-12) is crucial to the development of
protective immunity in mice intravenously infected with Mycobacterium tuberculosis. J Exp Med1997;186:39–45. [PubMed: 9206995]
3. Lowrie DB, Silva CL. Enhancement of immunocompetence in tuberculosis by DNA vaccination.Vaccine 2000;18:1712–1716. [PubMed: 10689154]
4. Ford-Hutchinson AW, Bray MA, Doig MV, Shipley ME, Smith MJ. Leukotriene, a potentchemokinetic aggregating substance released from polymorphonuclear leukocytes. Nature1980;286:264–265. [PubMed: 6250050]
Peres et al. Page 6
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

5. Marcinkiewicz J, Grabowska A, Bryniarski K, Chain BM. Enhancement of CD4+ T-cell-dependentinterleukin-2 production in vitro by murine alveolar macrophages: the role of leukotriene B4.Immunology 1997;91:369–374. [PubMed: 9301525]
6. Smith MJH, Ford-Hutchinson AW, Bray MA. Leukotriene, a potential mediator of inflammation. JPharm Pharmacol 1980;32:517–518. [PubMed: 6105196]
7. Bailie MB, Standiford TJ, Laichalk LL, Coffey MJ, Strieter R, Peters-Golden M. Leukotriene-deficientmice manifest enhanced lethality from Klebsiella pneumoniae in association with decreased alveolarmacrophage phagocytic and bactericidal activities. J Immunol 1996;157:5221–5224. [PubMed:8955165]
8. Medeiros AI, Sá-Nunes A, Soares EG, Peres CM, Silva CL, Faccioli LH. Blockade of endogenousleukotrienes exacerbates pulmonary histoplasmosis. Infect Immun 2004;72:1637–1644. [PubMed:14977971]
9. Peters-Golden M, Canetti C, Mancuso P, Coffey MJ. Leukotrienes: underappreciated mediators ofinnate immune response. J Immunol 2005;173:589–594. [PubMed: 15634873]
10. Hopkins H, Stull T, Von Essen SG, Robbins RA, Rennard SI. Neutrophil chemotactic factors inbacterial pneumonia. Chest 1989;95:1021–1027. [PubMed: 2539955]
11. Medeiros AI, Silva CL, Malheiro A, Maffei CM, Faccioli LH. Leukotrienes are involved in leukocyterecruitment induced by live Histoplasma capsulatum or by the β-glucan present in their cell wall. BrJ Pharmacol 1999;128:1529–1537. [PubMed: 10602333]
12. Coffey MJ, Phare SM, Peters-Golden M. Role of leukotrienes in killing of Mycobacterium bovis byneutrophils. Prostaglandins Leukot Essent Fatty Acids 2004;71:185–190. [PubMed: 15253889]
13. Aliberti J, Serhan C, Sher A. Parasite-induced lipoxin A4 is an endogenous regulator of IL-12production and immunopathology in Toxoplasma gondii infection. J Exp Med 2002;196:1253–1262.[PubMed: 12417634]
14. Bonato VL, Gonçalves ED, Soares EG, Santos RR Junior, Sartori A, Coelho-Castelo AA, Silva CL.Immune regulatory effect of pHSP65 DNA therapy in pulmonary tuberculosis: activation of CD8+cells, interferon-gamma recovery and reduction of lung injury. Immunol 2004;113:130–138.
15. Faccioli LH, Souza GE, Cunha FQ, Poole S, Ferreira SH. Recombinant interleukin-1 and tumornecrosis factor induce neutrophil migration “in vivo” by indirect mechanisms. Agents Actions1990;30:344–349. [PubMed: 2201176]
16. Stuehr DJ, Nathan CF. Nitric oxide. A macrophage product responsible for cytostasis and respiratoryinhibition in tumor target cells. J Exp Med 1989;169:1543–1555. [PubMed: 2497225]
17. Lima VM, Bonato VL, Dos Santos SA, Dos Santos RR, Goncalves ED, Faccioli LH, Brandao IT,Rodrigues JM Junior, Silva CL. Role of trehalose dimycolate in recruitment of cells and modulationof production of production of cytokines and NO in tuberculosis. Infect Immun 2001;69:5305–5312.[PubMed: 11500399]
18. Bafica A, Scanga CA, Serhan C, Machado F, White S, Sher A, Aliberti J. Host control ofMycobacterium tuberculosis is regulated by 5-lipoxygenase-dependent lipoxin production. J ClinInvest 2005;115:1601–1606. [PubMed: 15931391]
19. Mancuso P, Standiford TJ, Marshall T, Peters-Golden M. 5-lipoxygenase reaction products modulatealveolar macrophage phagocytosis of klebsiella pneumoniae. Infect Immun 1998;66:5140–5146.[PubMed: 9784515]
20. Mancuso P, Nana-Sinkam P, Peters-Golden M. Leukotriene B4 augments neutrophil phagocytosisof Klebsiella pneumoniae. Infect Immun 2001;69:2011–2016. [PubMed: 11254552]
21. Larfars G, Lantoine F, Devynck MA, Palmblad J, Gyllenhammar H. Activation of nitric oxide releaseand oxidative metabolism by leukotrienes B4, C4, and D4 in human polymorphonuclear leukocytes.Blood 1999;93:1399–1405. [PubMed: 9949184]
22. Gagnon L, Filion LG, Dubois C, Rola-Pleszczinski M. Leukotrienes and macrophage activation:augmented cytotoxic activity and enhanced interleukin 1, tumor necrosis factor and hydrogenperoxide production. Agents Actions 1989;26:141–147. [PubMed: 2540636]
23. Cooper AM, Roberts AD, Rhoades E, Callahan JE, Getzy DM, Orme IM. The role of interleukin-12in acquired immunity to mycobacterium tuberculosis infection. Immunol 1995;84:423–432.
Peres et al. Page 7
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

24. Tager AM, Bromley SK, Medoff BD, Islam SA, Bercury SD, Friedrich EB, Carafone AD, GersztenRE, Luster AD. Leukotriene B4 receptor BLT1 mediates early effector T cell recruitment. NatureImmunol 2003;4:982–990. [PubMed: 12949531]
25. Hebert MJ, Takano T, Holthofer H, Brady HR. Sequential morphologic events during apoptosis ofhuman neutrophils: modulation by lipoxygenase-derived eicosanoids. J Immunol 1996;157:3105–3115. [PubMed: 8816421]
26. Huang L, Zhao A, Wong F, Ayala JM, Struthers M, Ujjainwalla F, Wright SD, Springer MS, EvansJ, Cui J. Leukotriene B4 strongly increases monocyte chemoattractant protein-1 in human monocytes.Arterioscler Thromb Vasc Biol 2004;24:1783–1788. [PubMed: 15271789]
27. Ladel CH, Blum C, Reifenberg A, Kopf M, Kaufmann SHE. Lethal tuberculosis in inteleukin-6-deficient mice. Infect Immun 1997;65:4843–4849. [PubMed: 9353074]
28. Law K, Weiden M, Harkin T, Tchou-Wong K, Chi C, Rom WN. Increased release of interleukin-1beta, interkeukin-6, and tumor necrosis factor-alpha by bronchoalveolar cells lavage from sites inpulmonary tuberculosis. Am J Respir Crit Care Med 1996;153:799–804. [PubMed: 8564135]
29. Coffey M, Phare SM, Cinti S, Peters-Golden M, Kazanjian PH. Granulocyte-macrophage colony-stimulating factor upregulates reduced 5-lipoxygenase metabolism in peripheral blood monocytesand neutrophils in AIDS. Blood 1999;94:3897–3905. [PubMed: 10572106]
30. Skerret S, Henderson W, Martin T. Alveolar macrophage function in rats with severe protein caloriemalnutrition: arachidonic acid metabolism, cytokine release, and antimicrobial activity. J Immunol1990;144:1052–1061. [PubMed: 2104909]
Peres et al. Page 8
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 1.Lung LTB4 production in M. tuberculosis-infected mice and its inhibition by MK 886. Enzymeimmunoassay quantification of LTB4 concentrations in lungs from mice submitted to eitheri.t. PBS injection and daily p.o. doses of water or i.t. infection with M. tuberculosis (Mtb) anddaily p.o. doses of water, or MK 886 (5 mg/kg/0.5 ml). Lungs removed on days 2, 4, 7, 15, 30and 60 and homogenized in medium are shown. Leukotriene concentrations in supernatantsare expressed as ng/ml. Data are presented as mean ± SEM from one representative result ofthree independent experiments (n = 6). * PBS + H2O vs Mtb + H2O; # Mtb + H2O vs Mtb +MK. P < 0.05.
Peres et al. Page 9
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 2.Endogenous leukotriene synthesis inhibition abrogates mycobacterial clearance and mousesurvival rate. (A) Bacterial load in lungs. Tissue samples were harvested at 30 and 60 days ofMtb-infection with daily administration of water or MK 886. Data are expressed as mean ±SEM from one representative result of three independent experiments (n = 8). * Mtb + H2Ovs Mtb + MK. P < 0.05. (B) Survival of mice after i.t. PBS injection and daily p.o.administration of water, after i.t. infection with M. tuberculosis (Mtb) and daily p.o.administration of water or MK 886 (5 mg/kg/0.5 ml). Mice were monitored for 60 days (n =7).
Peres et al. Page 10
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
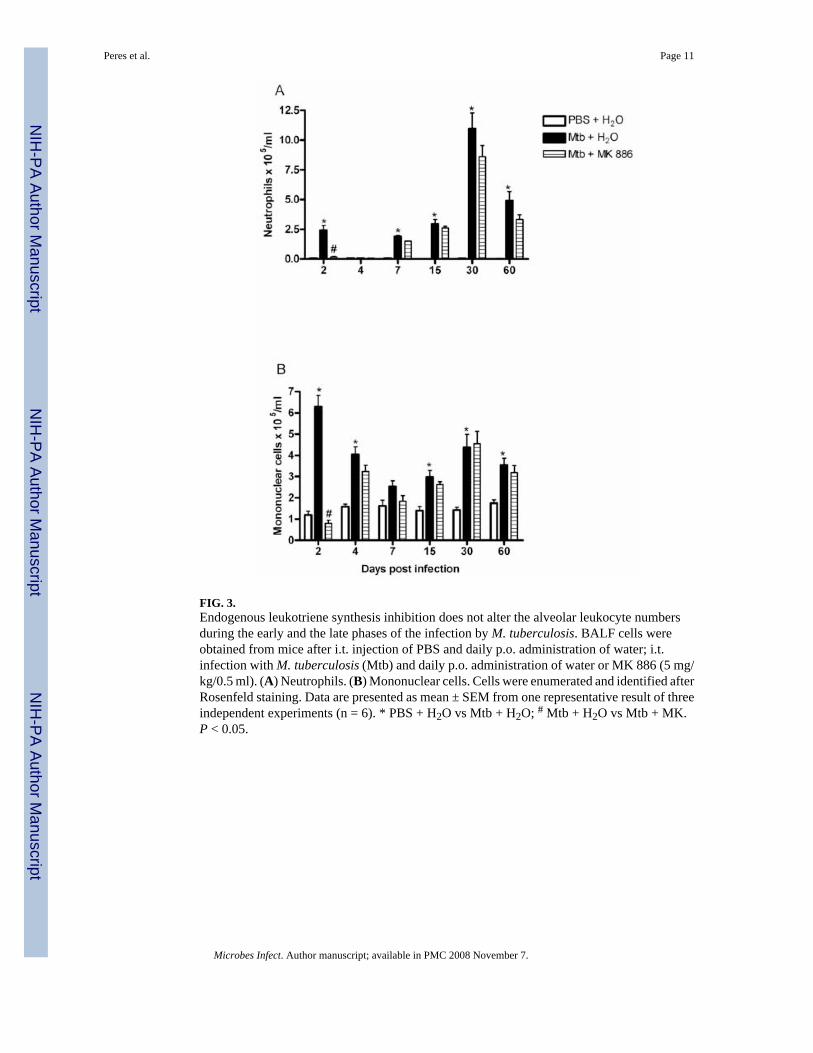
FIG. 3.Endogenous leukotriene synthesis inhibition does not alter the alveolar leukocyte numbersduring the early and the late phases of the infection by M. tuberculosis. BALF cells wereobtained from mice after i.t. injection of PBS and daily p.o. administration of water; i.t.infection with M. tuberculosis (Mtb) and daily p.o. administration of water or MK 886 (5 mg/kg/0.5 ml). (A) Neutrophils. (B) Mononuclear cells. Cells were enumerated and identified afterRosenfeld staining. Data are presented as mean ± SEM from one representative result of threeindependent experiments (n = 6). * PBS + H2O vs Mtb + H2O; # Mtb + H2O vs Mtb + MK.P < 0.05.
Peres et al. Page 11
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 4.Administration of MK 886 modulates levels of cytokines in lung tissue of infected mice. Lungsremoved on days 2, 4, 7, 15, 30 and 60 and homogenized in medium are shown. TNF-α (A),IL-1 (B), IL-6 (C), IL-12 (D), IFN-γ (E) and IL-10 (F) levels were determined by ELISA. Dataare expressed as mean ± SEM from one representative result of three independent experiments(n = 6). * PBS + H2O vs Mtb + H2O; # Mtb + H2O vs Mtb + MK. P < 0.05.
Peres et al. Page 12
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 5.Endogenous leukotriene synthesis inhibition impaired the production of NO2 in infected mice.Lung parenchyma homogenized in medium from mice subjected to either i.t. PBS injectionand daily p.o. doses of water or i.t. infection with M. tuberculosis (Mtb) and daily p.o. dosesof water or MK 886 (5 mg/kg/0.5 ml). NO2 production was quantified by Griess reaction inthe supernatants of cellular lysates from lung parenchyma recovered on days 2, 4, 7, 15, 30and 60. Data are expressed as mean ± SEM from one representative result of two independentexperiments (n = 6). * PBS + H2O vs Mtb + H2O; # Mtb + H2O vs Mtb + MK 886. P < 0.05.
Peres et al. Page 13
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FIG. 6.Effect of MK 886 on LXA4 synthesis in lung tissue. Enzyme immunoassay quantification ofLXA4 concentrations in lungs from mice submitted to either i.t. PBS injection and daily p.o.doses of water or i.t. infection with M. tuberculosis (Mtb) and daily p.o. doses of water, or 5mg/kg/0.5 ml of MK 886. Lungs removed on days 30 and 60 and homogenized in medium areshown. Lipoxin concentrations in supernatants were measured using a commercial kit(Neogen). Data are presented as mean ± SEM from one experiment (n = 6). * PBS + H2O vsMtb + H2O and Mtb + MK; # Mtb + H2O vs Mtb + MK. P < 0.05.
Peres et al. Page 14
Microbes Infect. Author manuscript; available in PMC 2008 November 7.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Copyright © 2022 FDOKUMEN




















