
Increased Apoptosis in Acute Puromycin Aminonucleoside Nephrosis
10
Original Paper Exp Nephrol 2001;9:99–108 Increased Apoptosis in Acute Puromycin Aminonucleoside Nephrosis Lucas Fernandez b Maritza Romero a Haydee Soto a Jesu ´ s Mosquera a a Instituto de Investigaciones Clı ´nicas ‘Dr Américo Negrette’ y b Centro de Cirugı ´a Experimental, Facultad de Medicina, Universidad del Zulia, Maracaibo, Venezuela Received: August 9, 1999 Accepted: February 18, 2000 Jesu ´ s A. Mosquera, MD Instituto de Investigaciones Clinicas, Facultad de Medicina Universidad del Zulia, Apartado Postal 1151 Maracaibo 4001-A, Zulia (Venezuela) Tel./Fax +58 61 597248, E-Mail [email protected] ABC Fax + 41 61 306 12 34 E-Mail [email protected] www.karger.com © 2001 S. Karger AG, Basel 1018–7782/01/0092–0099$17.50/0 Accessible online at: www.karger.com/journals/exn Key Words Apoptosis W Puromycin W Nephrosis W Proliferation W Infiltration Abstract Acute puromycin aminonucleoside nephrosis (PAN) in rats is characterized by heavy proteinuria associated with renal hypercellularity. The role of apoptosis in the resolution of renal hypercellularity was investigated in PAN. To study the participation of apoptosis in PAN, renal tissues were collected from nephrotic and control rats on weeks 1, 2 and 7 after a single puromycin amino- nucleoside injection. Apoptosis was evaluated by light and electron microscopy. Apoptotic DNA fragmentation was detected by TUNEL staining. Renal tissues were also evaluated by the presence of leukocyte common antigen (LCA), ED1 (macrophages) and proliferating cell nuclear antigen (PCNA) with corresponding monoclonal anti- bodies. An increased number of apoptotic (TUNEL+) cells was observed in the glomerulus at week 1. Elec- tron microscopy analysis showed glomerular apoptosis mainly in endothelial cells. In the interstitium and tu- bules, increased apoptosis was observed at weeks 1 and 2. Increased apoptosis was accompanied with increased LCA+, ED1+ and PCNA+ cells in the interstitium and with increased PCNA+ cells in tubules. There was a high sig- nificant correlation between the number of apoptotic cells and the number of interstitial LCA+, ED1+ and PCNA+ cells. Tubular PCNA expression was correlated with tubular apoptosis. We also observed significant cor- relation between glomerular, interstitial and tubular apoptosis with proteinuria during the nephrosis. Double staining analysis showed that about 13% of interstitial or tubular apoptotic cells were positive for PCNA. All these values returned to normal by week 7. These results indi- cate that apoptosis is involved in the repairing process of this disease model. Copyright © 2001 S. Karger AG, Basel Introduction The administration of puromycin aminonucleoside (PA) to rats produces well-defined glomerular lesions, and increased numbers of glomerular and interstitial infiltrat- ing cells. In the acute phase of puromycin aminonucleo- side nephrosis (PAN), leukocyte infiltration reaches a peak on day 14 and declines thereafter [1–5]. Little is known about the nature of the mechanisms of healing in
Transcript of Increased Apoptosis in Acute Puromycin Aminonucleoside Nephrosis

Original Paper
Exp Nephrol 2001;9:99–108
Increased Apoptosis in AcutePuromycin Aminonucleoside Nephrosis
Lucas Fernandezb Maritza Romeroa Haydee Sotoa Jesus Mosqueraa
aInstituto de Investigaciones Clınicas ‘Dr Américo Negrette’ y bCentro de Cirugıa Experimental,Facultad de Medicina, Universidad del Zulia, Maracaibo, Venezuela
Received: August 9, 1999Accepted: February 18, 2000
Jesus A. Mosquera, MDInstituto de Investigaciones Clinicas, Facultad de MedicinaUniversidad del Zulia, Apartado Postal 1151Maracaibo 4001-A, Zulia (Venezuela)Tel./Fax +58 61 597248, E-Mail [email protected]
ABCFax + 41 61 306 12 34E-Mail [email protected]
© 2001 S. Karger AG, Basel1018–7782/01/0092–0099$17.50/0
Accessible online at:www.karger.com/journals/exn
Key WordsApoptosis W Puromycin W Nephrosis W Proliferation W
Infiltration
AbstractAcute puromycin aminonucleoside nephrosis (PAN) inrats is characterized by heavy proteinuria associatedwith renal hypercellularity. The role of apoptosis in theresolution of renal hypercellularity was investigated inPAN. To study the participation of apoptosis in PAN,renal tissues were collected from nephrotic and controlrats on weeks 1, 2 and 7 after a single puromycin amino-nucleoside injection. Apoptosis was evaluated by lightand electron microscopy. Apoptotic DNA fragmentationwas detected by TUNEL staining. Renal tissues were alsoevaluated by the presence of leukocyte common antigen(LCA), ED1 (macrophages) and proliferating cell nuclearantigen (PCNA) with corresponding monoclonal anti-bodies. An increased number of apoptotic (TUNEL+)cells was observed in the glomerulus at week 1. Elec-tron microscopy analysis showed glomerular apoptosismainly in endothelial cells. In the interstitium and tu-bules, increased apoptosis was observed at weeks 1 and2. Increased apoptosis was accompanied with increased
LCA+, ED1+ and PCNA+ cells in the interstitium and withincreased PCNA+ cells in tubules. There was a high sig-nificant correlation between the number of apoptoticcells and the number of interstitial LCA+, ED1+ andPCNA+ cells. Tubular PCNA expression was correlatedwith tubular apoptosis. We also observed significant cor-relation between glomerular, interstitial and tubularapoptosis with proteinuria during the nephrosis. Doublestaining analysis showed that about 13% of interstitial ortubular apoptotic cells were positive for PCNA. All thesevalues returned to normal by week 7. These results indi-cate that apoptosis is involved in the repairing process ofthis disease model.
Copyright © 2001 S. Karger AG, Basel
Introduction
The administration of puromycin aminonucleoside(PA) to rats produces well-defined glomerular lesions, andincreased numbers of glomerular and interstitial infiltrat-ing cells. In the acute phase of puromycin aminonucleo-side nephrosis (PAN), leukocyte infiltration reaches apeak on day 14 and declines thereafter [1–5]. Little isknown about the nature of the mechanisms of healing in

100 Exp Nephrol 2001;9:99–108 Fernandez/Romero/Soto/Mosquera
the disease, which involve the resolution of hypercellu-larity.
Apoptosis has been widely recognized as a manner ofcell death with important biological roles [6–8]. One ofthe roles of apoptosis is the regulation of cell number intissues by deletion of ‘unwanted’ cells [9–13]. Recentreports indicate that apoptosis is present in several hu-man [14, 15] and experimental proliferative nephritis [16,17], suggesting that apoptosis may participate in therepair process of some renal diseases. Therefore, in thepresent study we have used a well-established rat model ofnephrosis to study if apoptosis is present in the kidneyduring the acute phase of PAN, and to analyze its relation-ship with cellular infiltration and proliferation.
Materials and Methods
AnimalsMale Sprague-Dawley rats, weighing 250–300 g and obtained
from the Instituto Venezolano de Investigaciones Cientıficas (Vene-zuela), were used for this study. They had free access to food andwater throughout the experiments.
PAN ModelExperimental rats were made nephrotic by a single intraperitone-
al injection of 15 mg/100 g body weight of PA (Sigma Chemical Co.,St. Louis, Mo., USA). Rats injected with 0.9% saline were used ascontrols. Experimental and control rat groups (n = 5–6 per group)were sacrificed on weeks 1, 2 and 7 after PA injection. In order todetermine proteinuria, 24-hour urine samples were collected from allrats before the sacrifice and proteinuria was determined by the sulfo-salicylic acid method. At the time of sacrifice, kidneys were removedafter perfusion with 20 ml of 0.9% saline solution by abdominal aor-ta. For immunofluorescence and immunohistochemical studies,pieces of perfused kidneys were included in OCT compound (TissueTek, Miles Inc. Diagnostics Division, Kankakee, Ill., USA), frozen indry ice and acetone and stored at –70°C until use. For light andelectron microscopy studies, isolated glomeruli or pieces of renal cor-tex from control and experimental rats were fixed in 2.5% of glutaral-dehyde.
Antibodies and Immunofluorescence StudiesTo identify leukocyte surface markers and proliferative activity
in renal frozen sections, a panel of mouse monoclonal antibodiesagainst rat antigens were used: antileukocyte common antigen (LCA;clone OX1), and antimonocyte/macrophage cells (ED1) from Accu-rate Chemical & Scientific Corp., Westbury, N.Y., USA. Mouseimmunoglobulins were detected in the tissues by a rat FITC-labelled(Fab))2 antibody against mouse IgG (Boehringer Mannheim, Germa-ny) for immunofluorescence studies. Renal proliferative activity wasdetected by a monoclonal antibody antiproliferating cell nuclearantigen (PCNA) (Zymed Laboratories, San Francisco, Calif., USA)followed by a biotinylated F(ab))2 goat anti-mouse IgG antibody (Or-ganon Teknika Corp., Cappel, Durham, N.C., USA) for immunohis-tochemical studies (see below). To examine the expression of cell
markers in the kidney specimens, 4-Ìm thick cryostat sections werereacted with each mAb listed above for 30 min. After washing, tis-sues were reacted with a rat FITC-labelled (Fab))2 antibody againstmouse IgG in a solution of PBS containing 10% of normal rat serumas a second antibody. Sections were mounted in a solution of p-phenylenediamine in PBS-glycerol and observed in a fluorescencemicroscopy (Axioskop; Zeiss, Germany).
Identification of ApoptosisTo determine the cellular apoptosis during PAN, we utilized a
histologic assay that allows for selective labelling of cells withdegraded DNA in tissue sections [18, 19]. This assay takes advantageof the ability of Klenow DNA polymerase to fill gaps of degradedDNA in apoptotic cell nuclei with digoxigenin-modified nucleotides.Nuclear DNA repaired with digoxigenin-labelled nucleotides canthen be easily detected by immunofluorescence staining proceduresutilizing a fluorescein isothiocyanate (FITC)-labelled antibody thatrecognizes digoxigenin. The frozen sections (4 Ìm) from normal andnephrotic animals were fixed in 10% neutral-buffered formalin andpostfixed in ethanol:acetic acid for 5 min at –20°C. Tissular frag-mented DNA was nick end-labelled using the ApopTag™. In situApoptosis Detection Kit-Fluorescein (ONCOR, Gaithersburg, Md.,USA). Embryonic rat forelimb bud to detect developmental cell dele-tion was used as positive control. Negative controls were representedby renal and embryonic rat forelimb bud tissues incubated with dis-tilled water instead of TdT enzyme in the reaction buffer. Slides weremounted with SlowFadeTM Antifade Kit (Molecular Probes, Inc.,USA) to delay fluorescence quenching and examined with an epi-fluorescence microscope. Apoptotic morphological features were alsostudied by light and electron microscopy procedures. In order todetermine the number of apoptotic glomerular cells, isolated glomer-uli from control animals (n = 5) and animals at 1 week of PAN (n = 5)were studied using the electron microscopy (see below). At least 50areas of isolated glomeruli were randomly studied per animal at low-er magnification (!5,000). Data were expressed as percentage ofapoptotic cells in endothelium, epithelium or mesangium.
Light and Electron Microscopy StudiesFor electron microscopy, glomeruli were obtained by pressing
minced renal cortexes from normal and nephrotic rats through sievesof graded sizes. Tissue samples were also taken from renal cortexesand cut into 1-mm cubes. Glomeruli or renal pieces were fixed for 5 hat room temperature in 2.5% of glutaraldehyde (Merck, Darmstadt,Germany) in 0.1 M cacodylate buffer (pH 7.2), rinsed in cacodylatebuffer, postfixed for 1 h in 2% OsO4, dehydrated in a graded ethanolseries, and embedded in Epon 812 (Fluka Chemie, Buchs, Switzer-land). Thin sections were cut with a diamond knife, collected on cop-per grids, and double stained with uranyl acetate and lead citratebefore examination in a Philips 400 electron microscope. Thin sec-tions were also stained with toluidine blue for light microscopy stud-ies.
Identification of Proliferating CellsRenal frozen sections (4 Ìm thick) fixed in 10% neutral buffered
formalin were used in these studies for the detection of PCNA. Sec-tions were postfixed in methanol, washed with PBS and incubatedwith avidin (2 Ìg/ml; Sigma Chemical Co.) for 20 min. After washingwith PBS, sections were incubated with 0.001% biotin solution (Sig-ma Chemical Co.) for 20 min. Afterwards, sections were incubatedfor 2 h with a monoclonal antibody against PCNA diluted 1:20 in
Apoptosis in Aminonucleoside Nephrosis Exp Nephrol 2001;9:99–108 101
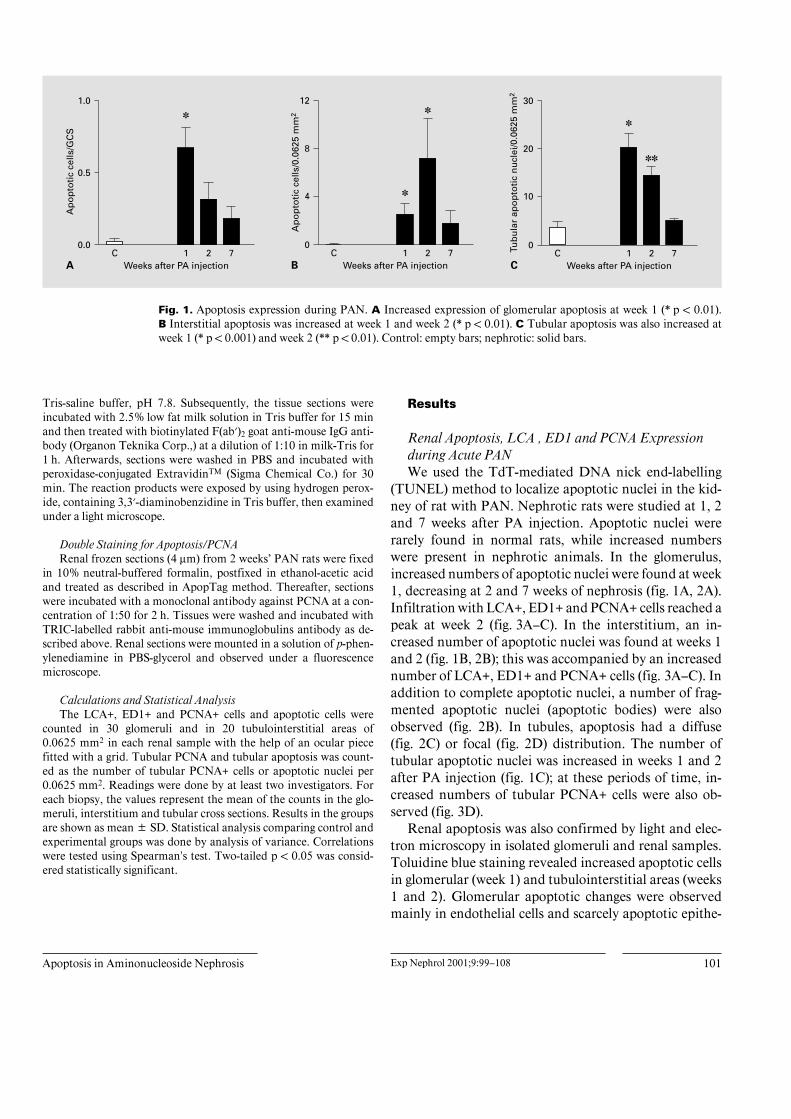
Fig. 1. Apoptosis expression during PAN. A Increased expression of glomerular apoptosis at week 1 (* p ! 0.01).B Interstitial apoptosis was increased at week 1 and week 2 (* p ! 0.01). C Tubular apoptosis was also increased atweek 1 (* p ! 0.001) and week 2 (** p ! 0.01). Control: empty bars; nephrotic: solid bars.
Tris-saline buffer, pH 7.8. Subsequently, the tissue sections wereincubated with 2.5% low fat milk solution in Tris buffer for 15 minand then treated with biotinylated F(ab))2 goat anti-mouse IgG anti-body (Organon Teknika Corp.,) at a dilution of 1:10 in milk-Tris for1 h. Afterwards, sections were washed in PBS and incubated withperoxidase-conjugated ExtravidinTM (Sigma Chemical Co.) for 30min. The reaction products were exposed by using hydrogen perox-ide, containing 3,3)-diaminobenzidine in Tris buffer, then examinedunder a light microscope.
Double Staining for Apoptosis/PCNARenal frozen sections (4 Ìm) from 2 weeks’ PAN rats were fixed
in 10% neutral-buffered formalin, postfixed in ethanol-acetic acidand treated as described in ApopTag method. Thereafter, sectionswere incubated with a monoclonal antibody against PCNA at a con-centration of 1:50 for 2 h. Tissues were washed and incubated withTRIC-labelled rabbit anti-mouse immunoglobulins antibody as de-scribed above. Renal sections were mounted in a solution of p-phen-ylenediamine in PBS-glycerol and observed under a fluorescencemicroscope.
Calculations and Statistical AnalysisThe LCA+, ED1+ and PCNA+ cells and apoptotic cells were
counted in 30 glomeruli and in 20 tubulointerstitial areas of0.0625 mm2 in each renal sample with the help of an ocular piecefitted with a grid. Tubular PCNA and tubular apoptosis was count-ed as the number of tubular PCNA+ cells or apoptotic nuclei per0.0625 mm2. Readings were done by at least two investigators. Foreach biopsy, the values represent the mean of the counts in the glo-meruli, interstitium and tubular cross sections. Results in the groupsare shown as mean B SD. Statistical analysis comparing control andexperimental groups was done by analysis of variance. Correlationswere tested using Spearman's test. Two-tailed p ! 0.05 was consid-ered statistically significant.
Results
Renal Apoptosis, LCA , ED1 and PCNA Expressionduring Acute PANWe used the TdT-mediated DNA nick end-labelling
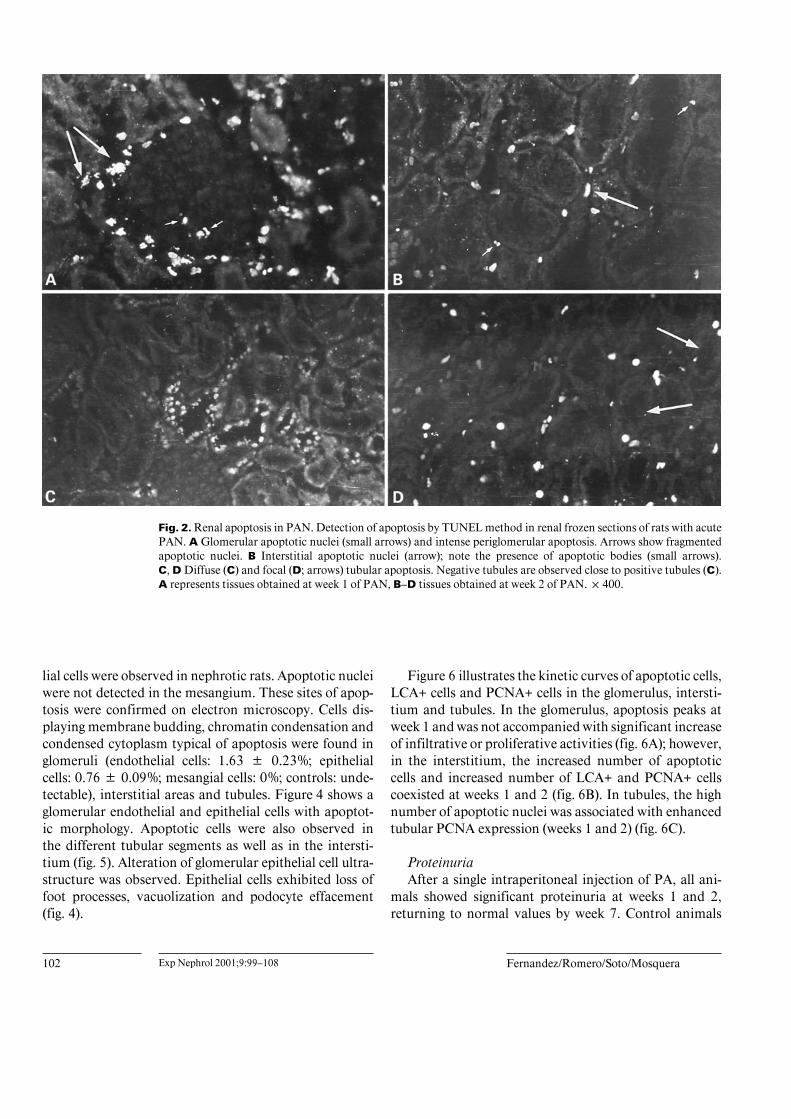
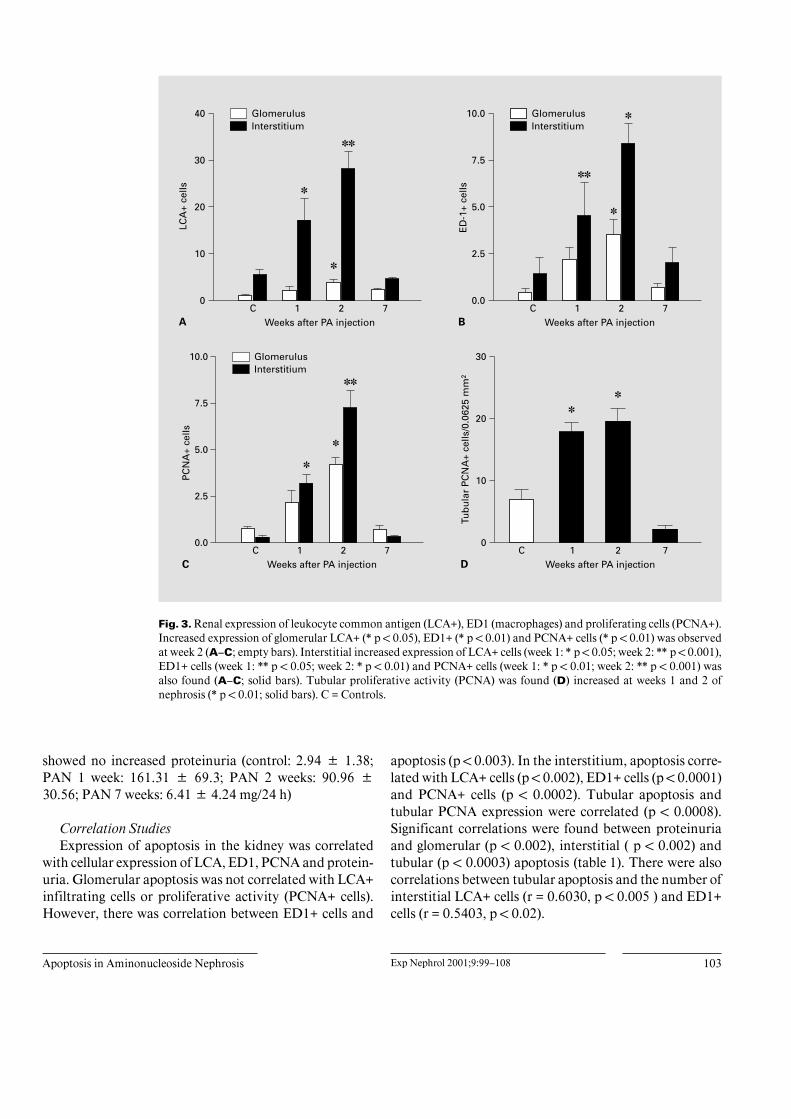
(TUNEL) method to localize apoptotic nuclei in the kid-ney of rat with PAN. Nephrotic rats were studied at 1, 2and 7 weeks after PA injection. Apoptotic nuclei wererarely found in normal rats, while increased numberswere present in nephrotic animals. In the glomerulus,increased numbers of apoptotic nuclei were found at week1, decreasing at 2 and 7 weeks of nephrosis (fig. 1A, 2A).Infiltration with LCA+, ED1+ and PCNA+ cells reached apeak at week 2 (fig. 3A–C). In the interstitium, an in-creased number of apoptotic nuclei was found at weeks 1and 2 (fig. 1B, 2B); this was accompanied by an increasednumber of LCA+, ED1+ and PCNA+ cells (fig. 3A–C). Inaddition to complete apoptotic nuclei, a number of frag-mented apoptotic nuclei (apoptotic bodies) were alsoobserved (fig. 2B). In tubules, apoptosis had a diffuse(fig. 2C) or focal (fig. 2D) distribution. The number oftubular apoptotic nuclei was increased in weeks 1 and 2after PA injection (fig. 1C); at these periods of time, in-creased numbers of tubular PCNA+ cells were also ob-served (fig. 3D).
Renal apoptosis was also confirmed by light and elec-tron microscopy in isolated glomeruli and renal samples.Toluidine blue staining revealed increased apoptotic cellsin glomerular (week 1) and tubulointerstitial areas (weeks1 and 2). Glomerular apoptotic changes were observedmainly in endothelial cells and scarcely apoptotic epithe-
102 Exp Nephrol 2001;9:99–108 Fernandez/Romero/Soto/Mosquera
Fig. 2. Renal apoptosis in PAN. Detection of apoptosis by TUNEL method in renal frozen sections of rats with acutePAN. A Glomerular apoptotic nuclei (small arrows) and intense periglomerular apoptosis. Arrows show fragmentedapoptotic nuclei. B Interstitial apoptotic nuclei (arrow); note the presence of apoptotic bodies (small arrows).C, D Diffuse (C) and focal (D; arrows) tubular apoptosis. Negative tubules are observed close to positive tubules (C).A represents tissues obtained at week 1 of PAN, B–D tissues obtained at week 2 of PAN. !400.
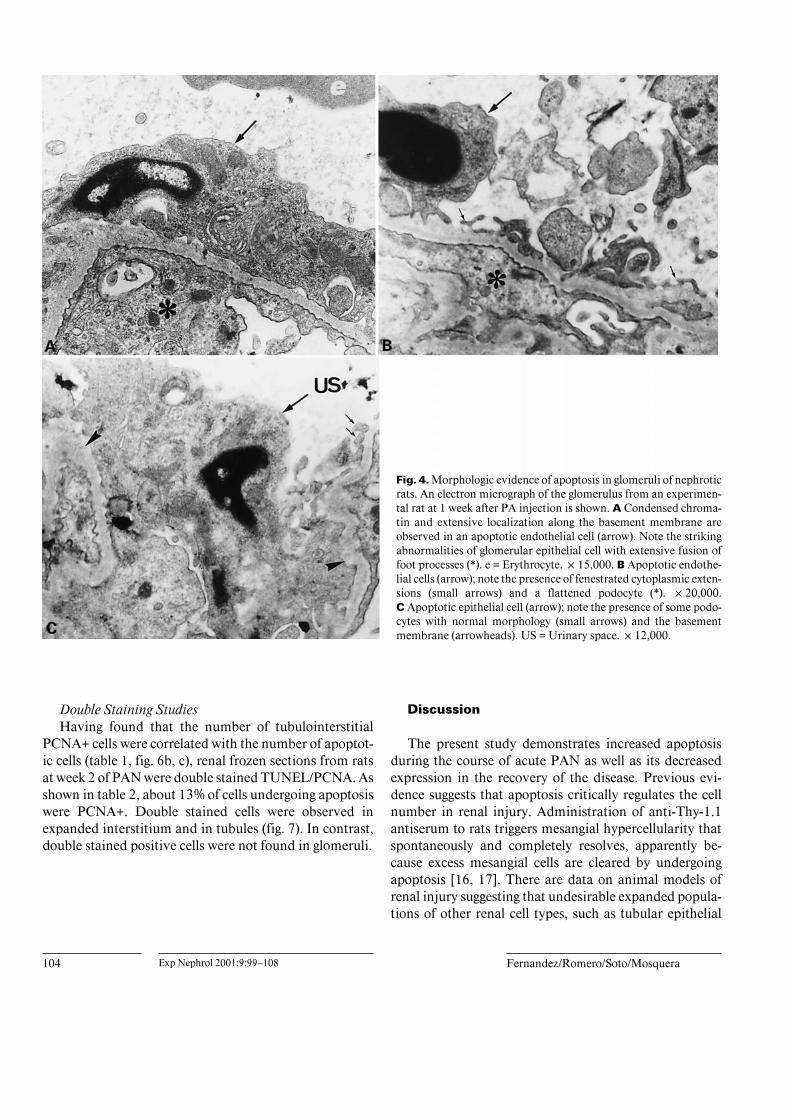
lial cells were observed in nephrotic rats. Apoptotic nucleiwere not detected in the mesangium. These sites of apop-tosis were confirmed on electron microscopy. Cells dis-playing membrane budding, chromatin condensation andcondensed cytoplasm typical of apoptosis were found inglomeruli (endothelial cells: 1.63 B 0.23%; epithelialcells: 0.76 B 0.09%; mesangial cells: 0%; controls: unde-tectable), interstitial areas and tubules. Figure 4 shows aglomerular endothelial and epithelial cells with apoptot-ic morphology. Apoptotic cells were also observed inthe different tubular segments as well as in the intersti-tium (fig. 5). Alteration of glomerular epithelial cell ultra-structure was observed. Epithelial cells exhibited loss offoot processes, vacuolization and podocyte effacement(fig. 4).
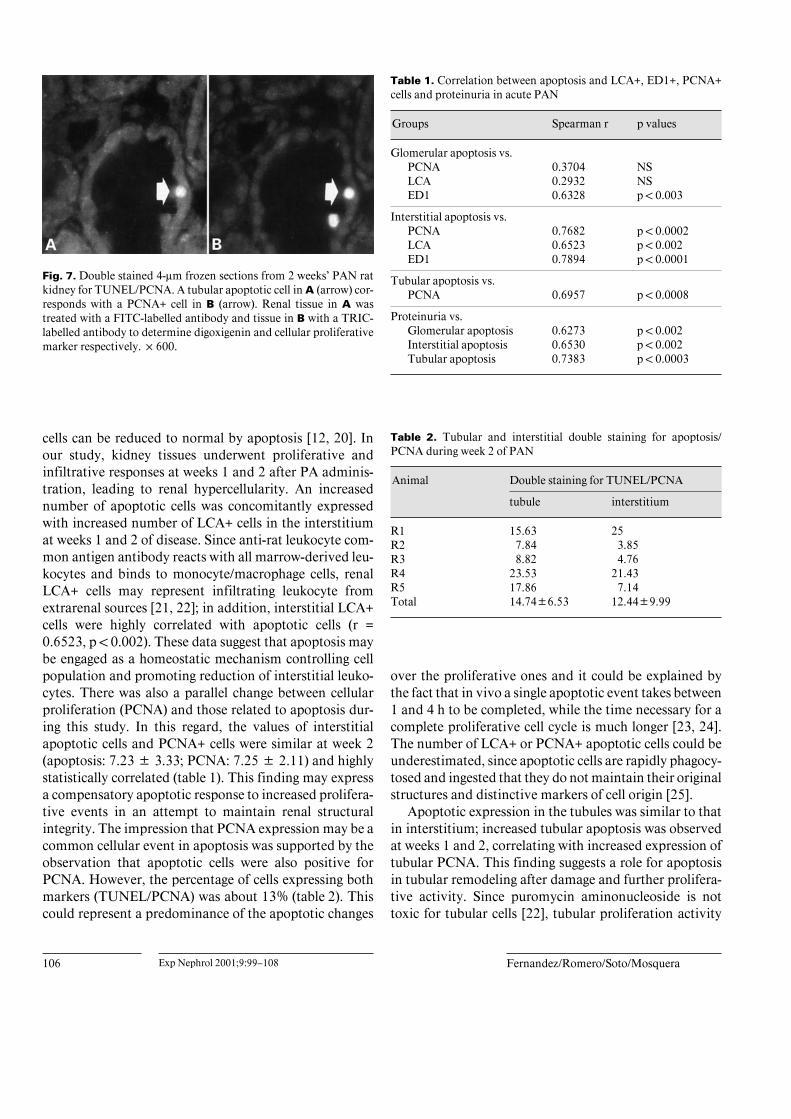
Figure 6 illustrates the kinetic curves of apoptotic cells,LCA+ cells and PCNA+ cells in the glomerulus, intersti-tium and tubules. In the glomerulus, apoptosis peaks atweek 1 and was not accompanied with significant increaseof infiltrative or proliferative activities (fig. 6A); however,in the interstitium, the increased number of apoptoticcells and increased number of LCA+ and PCNA+ cellscoexisted at weeks 1 and 2 (fig. 6B). In tubules, the highnumber of apoptotic nuclei was associated with enhancedtubular PCNA expression (weeks 1 and 2) (fig. 6C).
ProteinuriaAfter a single intraperitoneal injection of PA, all ani-
mals showed significant proteinuria at weeks 1 and 2,returning to normal values by week 7. Control animals
Apoptosis in Aminonucleoside Nephrosis Exp Nephrol 2001;9:99–108 103
Fig. 3. Renal expression of leukocyte common antigen (LCA+), ED1 (macrophages) and proliferating cells (PCNA+).Increased expression of glomerular LCA+ (* p ! 0.05), ED1+ (* p ! 0.01) and PCNA+ cells (* p ! 0.01) was observedat week 2 (A–C; empty bars). Interstitial increased expression of LCA+ cells (week 1: * p ! 0.05; week 2: ** p ! 0.001),ED1+ cells (week 1: ** p ! 0.05; week 2: * p ! 0.01) and PCNA+ cells (week 1: * p ! 0.01; week 2: ** p ! 0.001) wasalso found (A–C; solid bars). Tubular proliferative activity (PCNA) was found (D) increased at weeks 1 and 2 ofnephrosis (* p ! 0.01; solid bars). C = Controls.
showed no increased proteinuria (control: 2.94 B 1.38;PAN 1 week: 161.31 B 69.3; PAN 2 weeks: 90.96 B30.56; PAN 7 weeks: 6.41 B 4.24 mg/24 h)
Correlation StudiesExpression of apoptosis in the kidney was correlated
with cellular expression of LCA, ED1, PCNA and protein-uria. Glomerular apoptosis was not correlated with LCA+infiltrating cells or proliferative activity (PCNA+ cells).However, there was correlation between ED1+ cells and
apoptosis (p ! 0.003). In the interstitium, apoptosis corre-lated with LCA+ cells (p ! 0.002), ED1+ cells (p ! 0.0001)and PCNA+ cells (p ! 0.0002). Tubular apoptosis andtubular PCNA expression were correlated (p ! 0.0008).Significant correlations were found between proteinuriaand glomerular (p ! 0.002), interstitial ( p ! 0.002) andtubular (p ! 0.0003) apoptosis (table 1). There were alsocorrelations between tubular apoptosis and the number ofinterstitial LCA+ cells (r = 0.6030, p ! 0.005 ) and ED1+cells (r = 0.5403, p ! 0.02).
104 Exp Nephrol 2001;9:99–108 Fernandez/Romero/Soto/Mosquera
Fig. 4. Morphologic evidence of apoptosis in glomeruli of nephroticrats. An electron micrograph of the glomerulus from an experimen-tal rat at 1 week after PA injection is shown. A Condensed chroma-tin and extensive localization along the basement membrane areobserved in an apoptotic endothelial cell (arrow). Note the strikingabnormalities of glomerular epithelial cell with extensive fusion offoot processes (*). e = Erythrocyte. !15,000. B Apoptotic endothe-lial cells (arrow); note the presence of fenestrated cytoplasmic exten-sions (small arrows) and a flattened podocyte (*). !20,000.C Apoptotic epithelial cell (arrow); note the presence of some podo-cytes with normal morphology (small arrows) and the basementmembrane (arrowheads). US = Urinary space. !12,000.
Double Staining StudiesHaving found that the number of tubulointerstitial
PCNA+ cells were correlated with the number of apoptot-ic cells (table 1, fig. 6b, c), renal frozen sections from ratsat week 2 of PAN were double stained TUNEL/PCNA. Asshown in table 2, about 13% of cells undergoing apoptosiswere PCNA+. Double stained cells were observed inexpanded interstitium and in tubules (fig. 7). In contrast,double stained positive cells were not found in glomeruli.
Discussion
The present study demonstrates increased apoptosisduring the course of acute PAN as well as its decreasedexpression in the recovery of the disease. Previous evi-dence suggests that apoptosis critically regulates the cellnumber in renal injury. Administration of anti-Thy-1.1antiserum to rats triggers mesangial hypercellularity thatspontaneously and completely resolves, apparently be-cause excess mesangial cells are cleared by undergoingapoptosis [16, 17]. There are data on animal models ofrenal injury suggesting that undesirable expanded popula-tions of other renal cell types, such as tubular epithelial
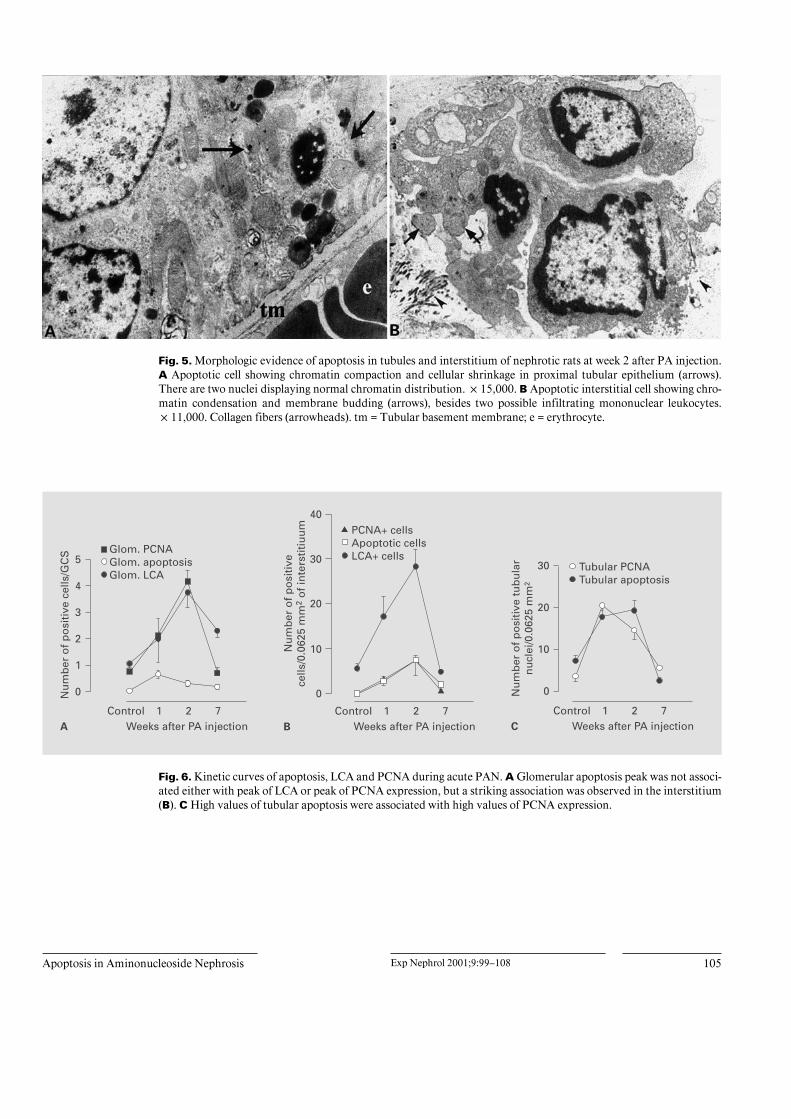
Apoptosis in Aminonucleoside Nephrosis Exp Nephrol 2001;9:99–108 105
Fig. 5. Morphologic evidence of apoptosis in tubules and interstitium of nephrotic rats at week 2 after PA injection.A Apoptotic cell showing chromatin compaction and cellular shrinkage in proximal tubular epithelium (arrows).There are two nuclei displaying normal chromatin distribution. !15,000. B Apoptotic interstitial cell showing chro-matin condensation and membrane budding (arrows), besides two possible infiltrating mononuclear leukocytes.!11,000. Collagen fibers (arrowheads). tm = Tubular basement membrane; e = erythrocyte.
Fig. 6. Kinetic curves of apoptosis, LCA and PCNA during acute PAN. A Glomerular apoptosis peak was not associ-ated either with peak of LCA or peak of PCNA expression, but a striking association was observed in the interstitium(B). C High values of tubular apoptosis were associated with high values of PCNA expression.
Glomerular apoptosis vs.
R1
106 Exp Nephrol 2001;9:99–108 Fernandez/Romero/Soto/Mosquera
Fig. 7. Double stained 4-Ìm frozen sections from 2 weeks’ PAN ratkidney for TUNEL/PCNA. A tubular apoptotic cell in A (arrow) cor-responds with a PCNA+ cell in B (arrow). Renal tissue in A wastreated with a FITC-labelled antibody and tissue in B with a TRIC-labelled antibody to determine digoxigenin and cellular proliferativemarker respectively. !600.
cells can be reduced to normal by apoptosis [12, 20]. Inour study, kidney tissues underwent proliferative andinfiltrative responses at weeks 1 and 2 after PA adminis-tration, leading to renal hypercellularity. An increasednumber of apoptotic cells was concomitantly expressedwith increased number of LCA+ cells in the interstitiumat weeks 1 and 2 of disease. Since anti-rat leukocyte com-mon antigen antibody reacts with all marrow-derived leu-kocytes and binds to monocyte/macrophage cells, renalLCA+ cells may represent infiltrating leukocyte fromextrarenal sources [21, 22]; in addition, interstitial LCA+cells were highly correlated with apoptotic cells (r =0.6523, p ! 0.002). These data suggest that apoptosis maybe engaged as a homeostatic mechanism controlling cellpopulation and promoting reduction of interstitial leuko-cytes. There was also a parallel change between cellularproliferation (PCNA) and those related to apoptosis dur-ing this study. In this regard, the values of interstitialapoptotic cells and PCNA+ cells were similar at week 2(apoptosis: 7.23 B 3.33; PCNA: 7.25 B 2.11) and highlystatistically correlated (table 1). This finding may expressa compensatory apoptotic response to increased prolifera-tive events in an attempt to maintain renal structuralintegrity. The impression that PCNA expression may be acommon cellular event in apoptosis was supported by theobservation that apoptotic cells were also positive forPCNA. However, the percentage of cells expressing bothmarkers (TUNEL/PCNA) was about 13% (table 2). Thiscould represent a predominance of the apoptotic changes
Table 1. Correlation between apoptosis and LCA+, ED1+, PCNA+cells and proteinuria in acute PAN
Groups Spearman r p values
PCNA 0.3704 NSLCA 0.2932 NSED1 0.6328 p ! 0.003
Interstitial apoptosis vs.PCNA 0.7682 p ! 0.0002LCA 0.6523 p ! 0.002ED1 0.7894 p ! 0.0001
Tubular apoptosis vs.PCNA 0.6957 p ! 0.0008
Proteinuria vs.Glomerular apoptosis 0.6273 p ! 0.002Interstitial apoptosis 0.6530 p ! 0.002Tubular apoptosis 0.7383 p ! 0.0003
Table 2. Tubular and interstitial double staining for apoptosis/PCNA during week 2 of PAN
Animal Double staining for TUNEL/PCNA
tubule interstitium
15.63 25R2 7.84 3.85R3 8.82 4.76R4 23.53 21.43R5 17.86 7.14Total 14.74B6.53 12.44B9.99
over the proliferative ones and it could be explained bythe fact that in vivo a single apoptotic event takes between1 and 4 h to be completed, while the time necessary for acomplete proliferative cell cycle is much longer [23, 24].The number of LCA+ or PCNA+ apoptotic cells could beunderestimated, since apoptotic cells are rapidly phagocy-tosed and ingested that they do not maintain their originalstructures and distinctive markers of cell origin [25].
Apoptotic expression in the tubules was similar to thatin interstitium; increased tubular apoptosis was observedat weeks 1 and 2, correlating with increased expression oftubular PCNA. This finding suggests a role for apoptosisin tubular remodeling after damage and further prolifera-tive activity. Since puromycin aminonucleoside is nottoxic for tubular cells [22], tubular proliferation activity

Apoptosis in Aminonucleoside Nephrosis Exp Nephrol 2001;9:99–108 107
could be the result of damage induced by interstitial cellu-lar infiltration [26], with further regeneration and apopto-sis. A correlation between the number of interstitial mac-rophages (ED1+ cells) and tubular apoptosis expression(r = 0.5403, p ! 0.02) and tubular apoptosis and PCNAtubular expression (r = 0.6957, p ! 0.0008; table 1) foundin our experiments could be related with that proposal. Inaddition, previous studies have shown an increased num-ber of T-cytotoxic/suppressor cells (OX8+) and naturalkiller cells (OX8+, OX19–) in the interstitium of rats withPA nephrosis [3, 27], cell types capable of inducing apop-tosis. Alternatively, tubular cell apoptosis and prolifera-tion may ocurr in response to proteinuria (table 1) [28,29]; in either case, apoptosis could represent a mechanismof resolution of tubular hypercellularity.
It is important to appreciate that although apoptosismay be a useful mechanism of cell deletion, it is not neces-sary a beneficial event; injurious stimuli can trigger apop-tosis and produce loss of desirable renal cells [30]. Thismay be the case in glomerular apoptosis in PAN. Glomer-ular apoptosis expression was found incremented in PANnephrosis at week 1 and decreased at weeks 2 and 7. Thisfinding was not associated with increment of LCA+ andPCNA+ glomerular cells which appears at week 2 whenapoptosis is subsiding. It is conceivable that the toxiceffect of PA to glomerular cells could induce apoptosis.Previous reports have shown in vitro and in vivo toxiceffect of PA on glomerular epithelial cells [31, 32], and arecent report [33] showed increased apoptosis in epithe-lial glomerular cells after multiple subcutaneous doses ofPA in rats. In the current study a toxic effect of PA onepithelial cells was observed. Ultrastructural analysisshowed vacuolization, loss of foot processes and in someareas podocyte effacement; however, a scanty number ofepithelial apoptotic cells was observed. Ultrastructuralfeatures of apoptosis were found mainly in endothelialcells. In this regard, previous studies have shown that PAinduces apoptosis in other cell types [34, 35]. We used amodel of a single intraperitoneal injection of PA and thesedifferent findings could be related to different susceptibil-ity of endothelial and epithelial cells to undergo apoptosisunder toxic effect of PA in vivo (multiple doses vs. singledose). Alternatively, cytotoxic lipid fraction produced inrats with PAN may be in part involved in glomerular cellapoptosis [36]. Apoptosis of endothelial cells could be rel-evant in the progression of focal and segmental glomeru-losclerosis found in PA nephrosis [37].
Significant positive correlations were found betweenglomerular, interstitial and tubular apoptosis with pro-teinuria (table 1). These correlations could be related to
puromycin-induced glomerular damage [31, 32, 38] andthe tubulointerstitial damage resulting from interactionbetween interstitial and tubular cells [3, 21, 27]. Alterna-tively, damage to tubular epithelial cells by filtered pro-teins may represent another example of an apoptoticstimulus. Native serum albumin with its lipidic compo-nents elicited apoptosis of tubular epithelial cells [28, 29].In this regard, increased activity of two cysteine proteasesin the proximal tubules in rats with PAN have beenreported [39].
Apoptosis expression, leukocyte infiltration, prolifera-tive activity and proteinuria decline to normal values byweek 7. This sequence of events is characteristic of othertypes of renal injury in man and animals: spontaneousresolution of poststreptococcal glomerulonephritis andexperimental mesangioproliferative glomerulonephritis,in which increased apoptosis has been reported [14–17],are good examples of this pattern.
In summary, in this study we report the developmentof apoptotic, infiltrative and proliferative events duringthe course of nephrotic syndrome in PA-treated rats. Theintensity of infiltration and proliferation correlates withapoptosis and apoptotic, infiltrative and proliferativeevents correlate with the severity of proteinuria. Detailedanalysis of cell markers suggests that proliferating intersti-tial and tubular cells are undergoing apoptosis. All numer-ic values returned to normal values at week 7 of nephrosis.In conclusion, during the resolution of infiltrative andproliferative events during acute PAN in rats, apoptosiscould play an important role for the restoration to the nor-mal cellularity.
Acknowledgments
The authors are grateful to Mr. Juan Pablo Hernandez for hiscooperation in the ultrastructural studies. This study was supportedby Consejo Cientifico y Humanistico de la Universidad del ZuliaCONDES (grant 1459-96).

108 Exp Nephrol 2001;9:99–108 Fernandez/Romero/Soto/Mosquera
References
1 Diamond JR, Pesek I: Glomerular tumor ne-crosis factor and interleukin-1 during acuteaminonucleoside nephrosis. Lab Invest 1991;64:21–28.
2 Diamond JR, Ding G, Frye J, Pesek-DiamondI: Glomerular macrophages and the mesangialproliferative response in the experimental ne-phrotic syndrome. Am J Pathol 1992;141:887–894.
3 Eddy AA, Michael AF: Acute tubulointerstitialnephritis associated with aminonucleoside ne-phrosis. Kidney Int 1988;33:14–23.
4 Jones CL, Buch S, Post M, McCulloch L, Liu E,Eddy AA: Renal extracellular matrix accumu-lation in acute puromycin aminonucleoside ne-phrosis in rats. Am J Pathol 1992;141:1381–1396.
5 Mampaso FM, Egido J, Martinez-Quintero JC,Bricio T, Gonzalez E, Cobo ME, Pirotzky E,Braquet P, Hernando L: Interstitial mononu-clear cell infiltrates in experimental nephrosis:Effect of PAF antagonist. Nephrol Dial Trans-plant 1989;4:1037–1044.
6 Kerr JFR: Shrinkage necrosis. A distinct modeof cellular death. J Pathol 1971;105:13–20.
7 Searle J, Kerr JFR, Bishop CJ: Necrosis andapoptosis: Distinct modes of cellular deathwith fundamentally different significance. Pa-thol Annu 1982;17:229–259.
8 Wyllie AH, Kerr JFR, Currie AR: Cell death:The significance of apoptosis. Int Rev Cytol1980;68:251–306.
9 Bhathal PS, Gall JAM: Deletion of hyperplasticbilliary epithelial cells by apoptosis followingremoval of the proliferative stimulus. Liver1985;5:311–325.
10 Bursch W, Lauer B, Timmermann-Trosiener I,Barthel G, Souppler J, Schulte-Hermann R:Controlled death (apoptosis) of normal and pu-tative preneoplastic cells in rat liver followingwithdrawal of tumor promoters. Carcinogene-sis 1984;5:453–458.
11 Columbano A, Ledda-Columbano GM, ConiPP, Faa G, Liguori C, Santacruz GS, Pani P:Occurrence of cell death (apoptosis) during theinvolution of liver hyperplasia. Lab Invest1985;52:670–675.
12 Ledda-Columbano GM, Columbano A, ConiP, Faa G, Pani P: Cell detection by apoptosisduring regression of renal hyperplasia. Am JPathol 1989;135:657–662.
13 Savill J: Apoptosis: A mechanism for regula-tion of the cell complement of inflamed glo-meruli. Kidney Int 1992;41:607–612.
14 Soto H, Mosquera J, Rodrıguez-Iturbe B, Hen-riquez C, Pinto A: Apoptosis in proliferativeglomerulonephritis: Decreased apoptosis ex-pression in lupus nephritis. Nephrol DialTransplant 1997;12:273–280.
15 Takemura T, Murakami K, Miyazato H, YagiK, Yoshioka K: Expression of Fas antigen andBcl-2 in human glomerulonephritis. Kidney Int1995;48:1886–1892.
16 Shimizu A, Kitamura H, Masuda Y, IshizakiM, Sugiga K, Yamanaka M: Apoptosis in therepair process of experimental proliferativeglomerulonephritis. Kidney Int 1995;47:114–121.
17 Baker AJ, Mooney A, Hughes J, Lombardi D,Johnson RJ, Savill J: Mesangial cells apoptosis:The major mechanism for resolution of glomer-ular hypercellularity in experimental mesangialproliferative nephritis. J Clin Invest 1994;94:2105–2116.
18 Wijsman JH, Jonker RR, Keuzer R, VeldeCJH, Cornelisse CJ, Dierendonck JH: Newmethod to detect apoptosis in paraffin sections:In situ end-labeling of fragmented DNA. J His-tochem Cytochem 1993;41:7–12.
19 Gravrieli Y, Sherman Y, Ben-Sasson SA: Iden-tification of programed cell death in situ viaspecific labeling of nuclear DNA fragmenta-tion. J Cell Biol 1992;119:493–501.
20 Nouwen EJ, Verstrepen WA, Buyssens N, ZhuMQ, De Broe ME: Hyperplasia, hypertrophyand phenotypic alterations in the distal neph-ron after acute proximal tubular injury in therat. Lab Invest 1994;70:479–493.
21 Eddy AA, McCulloch L, Liu E, Adams J: Arelationship between proteinuria and acute in-terstitial disease in rats with experimental ne-phrosis syndrome. Am J Pathol 1991;138:1111–1123.
22 Diamond JR, Anderson S: Irreversible tubu-lointerstitial damage associated with chronicaminonucleoside nephrosis. Amelioration byangiotensin I converting enzyme inhibition.Am J Pathol 1990;137:1323–1332.
23 Arends MJ, Wyllie AH: Apoptosis: Mecha-nisms and role in pathology. Int Rev Exp Pa-thol 1991;32:223–254.
24 Bravo R, MacDonald-Bravo H: Existence oftwo populations of cyclin/proliferating cell nu-clear antigen during the cell cycle: Associationwith DNA replication sites. J Cell Biol 1987;105:1549–1554.
25 Savill J, Fadok V, Herison P, Haslett C: Phago-cyte recognition of cells undergoing apoptosis.Immunol Today 1992;14:131–136.
26 Eddy AA: Experimental insights into the tubu-lointerstitial disease accompanying primaryglomerular lesions. J Am Soc Nephrol 1994;5:1273–1287.
27 Rossmann P, Bukovcky A, Matousovic K, Ho-lub M, Kral J: Puromycin aminonucleoside ne-phropathy: Ultrastructure, glomerular polyan-ion and cell surface markers. J Pathol 1986;148:337–348.
28 Savill J, Mooney A, Hughes J: Apoptosis andrenal scarring. Kidney Int 1996;49:S14–S17.
29 Amore A, Cirina P, Iavarone A, Fiorucci GC,Coppo R: Proteinuria induces apoptosis in cul-tured tubular cells. J Am Soc Nephrol 1996;7:1691.
30 Thomas GL, Yang B, Wagner BE, Savill J,Nahas ME: Cellular apoptosis and prolifera-tion in experimental renal fibrosis. NephrolDial Transplant 1998;13:2216–2226.
31 Coers W, Huitema S, van der Horst ML, Ween-ing JJ: Puromycin aminonucleoside and Adria-mycin disturb cytoskeletal and extracellularmatrix protein organization, but not proteinsynthesis of cultured glomerular epithelial cells.Exp Nephrol 1994;2:40–50.
32 Fishman JA, Karnovsky MJ: Effects of theaminonucleoside of puromycin on glomerularepithelial cells in vitro. Am J Pathol 1985;118:398–407.
33 Shiiki H, Sasaki Y, Nishino T, Kimura T,Kirioka H, Fujimoto S, Dohi K: Cell prolifera-tion and apoptosis of the glomerular epithelialcells in rats with puromycin aminonucleosidenephrosis. Pathology 1998;66:221–229.
34 Davidoff AN, Mendelow BV: Puromycin-elic-ited c-myc mRNA superproduction precedesapoptosis in HL-60 leukemic cells. AnticancerRes 1993;13:2257–2260.
35 Sugita M, Morita T, Yonesaki T: Puromycininduces apoptosis of developing chick sympa-thetic neurons in a similar manner to NGFdeprivation. Zool Sci 1995;12:419–425.
36 Natori Y, Igawa Y, Nakao N, Natori Y: Cyto-toxicity of sera from rats with puromycin ami-nonucleoside nephrosis. Nephron 1996;73:258–263.
37 Diamond JR, Karnovsky MJ: Focal and seg-mental glomerulosclerosis following a single in-travenous dose of puromycin aminonucleo-side. Am J Pathol 1986;122:481–487.
38 Sugiyama H, Kashihara N, Yamasaki Y, Seri-kawa T, Okamoto K, Kanao K, Maeshima Y,Makino H, Ota Z: Reactive oxygen speciesinduce apoptosis in cultured mesangial cells(abstract). J Am Soc Nephrol 1993;6:796.
39 Olbricht CJ, Cannon JK, Tisher CC: CathepsinB and L in nephron segment of the rats withpuromycin aminonucleoside nephrosis. Kid-ney Int 1987;32:354–361.




















