Phosphoinositide binding and phosphorylation act sequentially in the activation mechanism of ezrin
Upload
independentCategory
view
2download
0
Phosphoinositide 3-kinase/Akt inhibition increases arsenictrioxide-induced apoptosis of acute promyelocytic and T-cellleukaemias
Acute promyelocytic leukaemia (APL) is a rare disease
representing 10–15% of all adult acute myeloid leukaemia
(AML) patients. APL is characterised by developmental arrest
of granulopoiesis at the promyelocytic stage and is generally
associated with a specific t(15;17) chromosomal translocation,
which fuses the PML protein and retinoic acid receptor (RAR)
genes to yield a PML–RAR fusion protein (Puccetti &
Ruthardt, 2004). Arsenic trioxide (As2O3) is a clinically
effective agent in the treatment of APL. At clinically achievable
concentrations (1–5 lmol/l in the plasma, see Evens et al,
2004; Douer & Tallman, 2005), As2O3 provokes the degrada-
tion of the PML-RAR oncogenetic fusion protein, expressed in
the vast majority of APLs, thus forcing the malignant
promyelocytes to terminal differentiation and/or apoptosis
(Zhu et al, 2002). Currently, As2O3 is considered the treatment
of choice for patients with relapsed disease, particularly in
patients exposed to retinoids within the previous 12 months
(Soignet et al, 2001). Furthermore, albeit with lower efficacy,
As2O3 also induces apoptosis in tumour cells lacking the
PML-RAR, including other types of leukaemia and multiple
Giovanna Tabellini,1 Pier Luigi Tazzari,2
Roberta Bortul,3 Camilla Evangelisti,4
Anna Maria Billi,4 Tiziana Grafone,5
Giovanni Martinelli,5 Michele Baccarani5
and Alberto M. Martelli4,6
1Dipartimento di Scienze Biomediche e
Biotecnologie, Sezione di Citologia e Istologia,
Universita di Brescia, Brescia, 2Servizio di
Immunoematologia e Trasfusionale, Policlinico
S.Orsola-Malpighi, Bologna, 3Dipartimento di
Morfologia Umana Normale, Universita di
Trieste, Trieste, 4Dipartimento di Scienze
Anatomiche Umane e Fisiopatologia
dell‘Apparato Locomotore, Sezione di Anatomia,
Cell Signalling Laboratory, Universita di Bologna,
Bologna, 5Istituto di Ematologia ed Oncologia
Medica Seragnoli, Universita di Bologna, Bologna,
and 6Istituto per i Trapianti d’Organo e
l’Immunocitologia del C.N.R., Sezione di Bologna
c/o I.O.R., Bologna, Italy
Received 01 May 2005; accepted for publication
21 June 2005
Correspondence: Alberto M. Martelli, MD,
Dipartimento di Scienze Anatomiche Umane e
Fisiopatologia dell’Apparato Locomotore,
Universita di Bologna, via Irnerio 48, 40126
Bologna, Italy.
E-mail: [email protected]
Summary
Recent studies suggest that the prosurvival signal transduction pathway
involving phosphoinositide 3-kinase (PI3K)/Akt can confer an aggressive,
apoptosis-resistant phenotype to acute leukaemia cells. We have investigated
the effect of modulating this signalling pathway on the sensitivity of
leukaemic cell lines (NB-4, CEM, Jurkat, MOLT-4) and acute promyelocytic
primary blasts to apoptosis induced by 1 lmol/l As2O3. Whereas parental
NB-4 cells did not display any phosphorylated (active) Akt, CEM, Jurkat and
MOLT-4 cells exhibited high levels of Akt activation. Consistently, treatment
of NB-4 cells with pharmacological inhibitors of the PI3K/Akt pathway
(LY294002, wortmannin) did not increase sensitivity of these cells to arsenic
trioxide (As2O3), whereas siRNA knock-down of Akt enhanced As2O3-
induced apoptosis of CEM, Jurkat and MOLT-4 cells. Overexpression of a
constitutively active Akt cDNA rendered NB-4 cells less susceptible to As2O3.
Upon prolonged exposure to As2O3, we isolated a NB-4 cell clone that was
resistant to As2O3 and displayed high levels of active Akt. LY294002
treatment of acute promyelocytic primary blasts with elevated Akt
phosphorylation levels resulted in an increased sensitivity to As2O3. These
results may provide a rationale for the development of combined or
sequential treatment with PI3K/Akt inhibitors to improve the efficacy of
As2O3 on acute leukaemias and also to overcome As2O3 resistance.
Keywords: apoptosis, leukaemia, protein phoshorylation, signal transduc-
tion.
research paper
doi:10.1111/j.1365-2141.2005.05679.x ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 130, 716–725

myeloma, which opens the possibility for broader clinical
application of this compound (Miller et al, 2002). For
example, As2O3 has been recently tested for the treatment of
acute T-cell leukaemia in combination with interferon
(Bazarbachi et al, 2004). Moreover, murine T-cell prolympho-
cytic leukaemia responds to As2O3 treatment in vitro and in
vivo (Recher et al, 2002). Nevertheless, it would be advisable to
elucidate strategies to increase the apoptotic action of As2O3
and reduce drug dosage. An important aspect of apoptosis
regulation is the signalling by serine/threonine kinases, a broad
category of kinases that includes, among others, the extracel-
lular-regulated protein kinases 1/2 (Erk 1/2) and Akt (Cross
et al, 2000). To be activated, Akt requires the 3¢ phosphory-
lated lipid products of phosphoinositide 3-kinase (PI3K)
(Hanada et al, 2004). Indeed, 3¢-phosphoinositides attract to
the plasma membrane phosphoinositide-dependent kinase 1
(PDK1) and Akt. PDK1 activates Akt by phosphorylation of
Thr 308 residue in the activation loop, although full activation
requires phosphorylation of Ser 473 in the C-terminal
hydrophobic motif of Akt. The latter phosphorylation step is
effected by a kinase that has not been conclusively identified so
far (Brazil et al, 2004; Hanada et al, 2004). The signalling
pathways that emanate from Erk 1/2 and PI3K/Akt are
considered very critical for cell survival to apoptotic stimuli
(Hanada et al, 2004; Wada & Penninger, 2004). For this
reason, these two signalling pathways represent promissory
targets of therapeutic intervention in haematological malig-
nancies (Platanias, 2003; Martelli et al, 2005). Indeed, recent
studies have highlighted that both Erk 1/2 and PI3K/Akt
pathways can be constitutively activated in AML blasts, and
their down-modulation by means of selective pharmacological
inhibitors induces apoptosis and/or increases blast sensitivity
to chemotherapeutic agents (Milella et al, 2001; Lunghi et al,
2003; Xu et al, 2003; Bortul et al, 2005). It has been shown that
inhibition of the Erk 1/2 pathway synergises with As2O3 to
induce apoptosis in APL cells (Lunghi et al, 2005). Moreover,
we have recently demonstrated that down-modulation of the
PI3K/Akt pathway increased sensitivity to As2O3 of a HL60 cell
clone with a constitutively activated PI3K/Akt axis (Tabellini
et al, 2005). Although HL60 cells are sensitive to retinoids,
which promote their granulocytic differentiation, they are not
representative of a typical APL, as they lack the t(15;17)
chromosomal translocation that is a distinctive feature of APL.
In this study, we investigated whether or not down-modula-
tion of PI3K/Akt pathway could potentiate the antileukaemic
action of As2O3 in cells from APL and acute T-leukaemia. We
found that pharmacological inhibitors of the PI3K/Ak signal-
ling network restored As2O3 sensitivity of a NB-4 cell clone
with an upregulated PI3K/Akt axis that was resistant to As2O3.
Down-modulation of Akt phoshorylation also significantly
enhanced As2O3-induced apoptosis of primary APL blasts.
Finally, blocking PI3K/Akt function with specific double-
stranded RNA oligonucleotides (siRNA) increased As2O3
sensitivity of T-leukaemia cell lines, which have constitutively
active Akt signalling.
Materials and methods
Chemicals and antibodies
As2O3, normal goat serum (NGS), bovine serum albumin (BSA,
Fraction V), monoclonal antibody to tubulin and peroxidase-
conjugated secondary antibodies were from Sigma (St Louis,
MO, USA). The COMPLETE Protease Inhibitor Cocktail,
Annexin V-fluorescein isothiocyanate (FITC) staining kit, and
the Lumi-LightPlus enhanced chemiluminescence (ECL) detec-
tion kit were from Roche Applied Science (Milan, Italy).
Wortmannin and LY294002 were from Calbiochem (La Jolla,
CA, USA). The In situ Cell Death Detection kit for TUNEL (TdT-
mediated dUTP Nick End Labelling) was from Roche Applied
Science. The Protein Assay kit (detergent compatible) was
from Bio-Rad (Hercules, CA, USA). Constitutively active and
dominant negative Akt cDNAs (both cloned in pUSEamp and
c-Myc tagged) were from Upstate (Charlottesville, VA, USA).
The constitutively active Akt is a Gag fusion protein, with a src
myristoylation signal sequence at its amino terminus. In the case
of dominant negative Akt, a mutation (K179M) results in loss of
affinity for ATP, and consequently a protein that is inactive as a
kinase. SMARTpool Akt siRNA was also from Upstate. Rabbit
polyclonals to total Akt and Ser 473 phosphorylated Akt (p-Akt,
Catalogue no. 9271, specific for Western blotting, and Catalogue
no. 9277, specific for immunocytochemistry) were from Cell
Signalling Technology (Beverly, MA, USA). Anti-c-Myc mono-
clonal antibody (clone 9E10) was from Santa Cruz Biotechno-
logy (Santa Cruz, CA, USA).
Cell culture
NB-4 [an APL cell line carrying the t(15;17) chromosomal
translocation], CEM, Jurkat, and MOLT-4 cells (all T-cell
leukaemia-derived cell lines) were routinely maintained in
Roswell Park Memorial Institute (RPMI) 1640 medium
supplemented with 10% fetal calf serum (complete medium)
at an optimal cell density of 3–8 · 105 cells/ml.
Selection of NB-4 As2O3-resistant cell line(NB-4-ArsRES/AKT)
An As2O3-resistant cell line, hereafter referred to as NB-4-
ArsRES/AKT, was derived from the parental NB-4 cell line
essentially as described by Gianni et al (1998). Briefly, cells
were continuously cultured in the presence of 1 lmol/l As2O3.
While most of the cell population died, rare As2O3-resistant
cells became evident after about 4 months of selection. To
separate the living from the dead cells, the method described
by Thøger-Andersen and Junker (1994) was employed. This
method exploits the specificity of the plant lectin concanavalin
A to bind glucosyl and mannosyl residues that are almost
universally present on the outer surface of mammalian cell
membranes. Before plating in tissue culture plates, the cell
suspension was maintained at 37�C and supplemented with
PI3K/Akt Inhibition and As2O3 in Leukaemia Cells
ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 130, 716–725 717

sterile agarose at the final concentration of 0Æ18%. After
2 weeks, clones were picked with a Pasteur pipette, and
expanded in complete medium. Individual clones were
screened for Ser 473 p-Akt levels by flow cytometry, as
reported elsewhere (Tazzari et al, 2004).
Primary APL blast culture
The APL patient samples were obtained at diagnosis from
peripheral blood after informed consent was given according
to the ethical standards of the institutional guidelines. All
samples had >80% blasts after Ficoll–Hypaque density-gradi-
ent centrifugation, as assessed by flow cytometric analysis with
anti-HLA-DR, CD13, CD33, CD34 antibodies (alone or in
combination). APL blasts (1 · 106/ml) were cultured in
methylcellulose medium (Methocult; Stem Cell Technologies,
Vancouver, Canada) supplemented with human recombinant
growth factors: interleukin (IL)-3 (20 ng/ml), IL-6 (20 ng/ml),
stem cell factor (50 ng/ml). They were treated with for 12 h
with LY294002 (30 lmol/l) and then incubated with As2O3
(1 lmol/l) for 48 h.
cDNA transfection
Transfection of parental NB-4 cells was performed by electro-
poration with a Bio-Rad Gene Pulser apparatus (Neri et al,
2003). Twenty microgram of plasmid DNA (either constitu-
tively active Akt or dominant negative Akt) were mixed with
107 cells in 0Æ5 ml of phosphate-buffered sucrose (272 mmol/l
sucrose and 7 mmol/l Na2HPO4, pH 7Æ4). Cells were electro-
porated with a pulse of 250 V for 18–20 ms. Control cells were
mock-transfected in the same conditions with the empty
vector. Following electroporation, cells were allowed to recover
in 20 ml of culture medium for 48 h before selection with
600 lg/ml of G418. G418-resistant transfected clones were
obtained by limited dilution.
siRNA transfection
Prior to electroporation, CEM, Jurkat and MOLT-4 cells were
washed twice with serum-free Opti-MEM (Gibco BRL, Paisley,
UK) and resusupended to a final concentration of
8 · 106 cells/ml in Opti-MEM. Subsequently, 0Æ5 ml of cell
suspension was mixed with either 0Æ5 nmol of siRNA against
Akt or non-specific control siRNA (as supplied by the
manufacturer) and electroporated in a 0Æ4-cm cuvette using
the Gene Pulser apparatus with a pulse of 260 V and a
capacitance of 1050 lF. Cells were immediately plated in
complete medium and treated with As2O3. After 48 h cells
were analysed by Western blotting for the levels of Akt.
Detection of apoptosis
For detection of apoptosis, cells were stained with Annexin
V-FITC (FITC) (Zhao et al, 2004). Briefly, cells were washed in
phosphate-buffered saline (pH 7Æ4, PBS) and resuspended in
100 ll of binding buffer containing Annexin V-FITC. Cells
were analysed by flow cytometry after addition of propidium
iodide (PI). Annexin V-FITC binds to those cells that express
phosphatidylserine on the outer layer of the cell membrane,
and PI stains the cellular DNA of those cells with a
compromised cell membrane. This enabled live cells (un-
stained with either fluorochrome) to be discriminated from
apoptotic cells (stained only with Annexin V-FITC) and
necrotic cells (stained with both Annexin V and PI) (Vermes
et al, 1995). In some cases, apoptotic cells were detected by the
TUNEL/PI technique, as reported elsewhere (Li et al, 1995;
Gao et al, 2004). Briefly, cells were first fixed at room
temperature with 4% paraformaldeheyde for 30 min and then
permeabilised for 2 min on ice in the presence of 0Æ1% Triton
X-100. Samples were then incubated (60 min at 37�C in the
dark) in the TUNEL reaction mixture containing TdT
(terminal deoxynucleotidyl transferase) and FITC-dUTP,
washed twice in PBS, treated with 100 U/ml RNase A, stained
with PI (50 g/ml), and finally analysed by flow cytometry. TdT
catalyses the polymerisation of the labelled nucleotide to free
3¢-OH DNA ends in a template-independent manner. The
TUNEL reaction preferentially labels DNA strand breaks
generated during apoptosis and enables discrimination
between apoptotic and necrotic cells. Cell cycle analysis was
performed on PI-stained samples as reported elsewhere
(Cappellini et al, 2003).
Intracellular immunostaining for Ser 473 p-Akt and flowcytometric analysis
This was performed essentially as described previously
(Tazzari et al, 2004). Cells (approximately 1 · 105/sample)
were fixed with Reagent 1 of the Intraprep kit, according to
the manufacturer’s instructions (Beckman Coulter, Miami,
FL, USA). Cells were then permeabilised with saponin-
based Reagent 2 and incubated at 4�C for 12 h with a 1:10
final dilution of antibody to Ser 473 p-Akt (Cell Signaling
Technology, catalogue no. 9277, specific for immunocyto-
chemistry). Samples were washed twice with PBS and
pellets were incubated with 5 ll of a FITC-conjugate swine
anti-rabbit IgG (DakoCytomation, Glostrup, Denmark).
Negative controls were run with FITC-conjugated swine
anti-rabbit IgG alone and with FITC-conjugated swine anti-
rabbit IgG plus normal rabbit IgG (from Sigma). All the
samples were analysed by EPICS XL and Cytomics FC 500
flow cytometers (Beckman Coulter) equipped with dedica-
ted software. Histograms were then analysed with EXPO
software.
Protein concentration assay
This was performed according to the instruction of the
manufacturer using the detergent-compatible Bio-Rad Protein
Assay.
G. Tabellini et al
718 ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 130, 716–725

Preparation of cell homogenates and Western blot analysis
Cells were washed twice in PBS, then lysed (107/ml) in
10 mmol/l Tris-HCl, pH 7Æ4, 1 mmol/l MgCl2, 1 mmol/l
EGTA, 1% Triton X-100, 0Æ25 mol/l sucrose, containing the
COMPLETE Protease Inhibitor Cocktail supplemented with
50 mmol/l NaF, 1 mmol/l 2-glycerophosphate, 25 mmol/l
NaPPi. After 15 min on ice, they were homogenised by 10
passages through a 25-gauge needle, and centrifuged in a
microfuge (9300 g) for 10 min at 4�C. An aliquot of the
supernatant was saved for protein assay. Supernatants were
mixed with 4x electrophoresis sample buffer, and then the
protein (80 lg/lane), separated by sodium dodecylsulphate-
polyacrylamide gel electrophoresis (SDS-PAGE), was trans-
ferred to nitrocellulose sheets using a semi-dry blotting
apparatus. Sheets were saturated in PBS containing 5% NGS,
4% BSA, 0Æ25 non-fat dry milk (blocking buffer) for 60 min
at 37�C, then incubated overnight at 4�C in blocking buffer
containing the primary antibody. After four washes in PBS
containing 0Æ1% Tween-20, they were incubated for 30 min
at room temperature with the appropriate peroxidase-conju-
gated secondary antibody, diluted 1:5000 in blocking buffer,
and washed as above. Bands were visualised by the ECL
method.
Statistical evaluation
Data are shown as mean ± SD. For statistical analyses, the
Student’s t-test for unpaired samples at the level of significance
of 0Æ01 was used.
Results
Ser 473 p-Akt levels and As2O3 sensitivity in human acuteleukaemia cell lines
We first analysed by Western blot the expression of total Akt
and the levels of Ser 473 p-Akt in four human acute
leukaemia cell lines. As shown in Fig 1A, NB-4, CEM, Jurkat
and MOLT-4 cells expressed comparable amount of total
Akt. In contrast, a marked Akt phosphorylation on Ser 473
was detected only in CEM, Jurkat and MOLT-4 cells, in
agreement with others (e.g. Xu et al, 2002; Uddin et al,
2004). NB-4 cells were completely negative for Ser 473
p-Akt. The sensitivity of these cells lines to incubation for
48 h with 1 lmol/l As2O3 was analysed next. NB-4, CEM
and MOLT-4 cells were highly sensitive to As2O3 as
indicated by the percentage of Annexin V-positive cells
detected by flow cytometry. On the contrary, Jurkat cells
were resistant to this concentration of As2O3. In some cases,
apoptotic cells were detected by means of the TUNEL/PI
technique with results similar to those obtained with
Annexin V staining (Fig 1B). Overall, our results were in
good agreement with the findings reported elsewhere
(Rojewski et al, 2002; Hu et al, 2003).
Cell cycle analysis
The effect of As2O3 treatment on cell cycle was evaluated by
flow cytometric analysis of PI-stained samples. As2O3
increased the percentage of NB-4 cells in the G2/M phase of
the cell cycle along with an increase in sub-G1 (apoptotic
cells), in agreement with others (e.g. Gao et al, 2004).
Consistent with the lack of Akt phosphorylation, PI3K
A
B
Fig 1. Akt expression, phosphorylation levels, and As2O3 sensitivity of
acute human leukaemia cell lines. (A) Western blot analysis for total
Akt and Ser 473 p-Akt. Protein (80 lg/lane) was separated by SDS-
PAGE and blotted to nitrocellulose sheets. Immunoreactive bands were
visualised by the ECL technique. For detection of p-Akt, antibody no.
9271 was employed. Immunostaining with a monoclonal antibody to
b-tubulin confirmed equal loading. Blots are representative of three
separate experiments. (B) Flow cytometric analysis of As2O3-induced
apoptosis. Cells were treated with 1 lmol/l As2O3 for 48 h. The per-
centage of apoptotic cells was evaluated by Annexin V-FITC staining or
TUNEL/PI technique. Untreated cells were negative for TUNEL reac-
tion (not shown). A representative of three separate experiments is
shown.
PI3K/Akt Inhibition and As2O3 in Leukaemia Cells
ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 130, 716–725 719
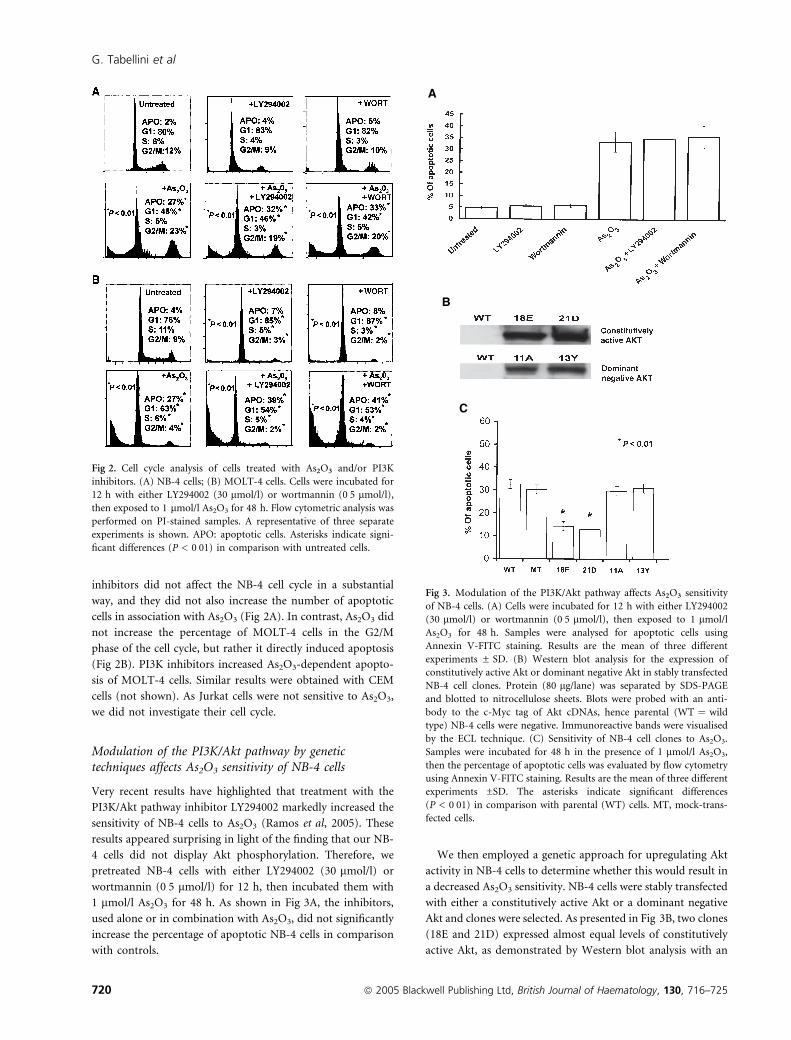
inhibitors did not affect the NB-4 cell cycle in a substantial
way, and they did not also increase the number of apoptotic
cells in association with As2O3 (Fig 2A). In contrast, As2O3 did
not increase the percentage of MOLT-4 cells in the G2/M
phase of the cell cycle, but rather it directly induced apoptosis
(Fig 2B). PI3K inhibitors increased As2O3-dependent apopto-
sis of MOLT-4 cells. Similar results were obtained with CEM
cells (not shown). As Jurkat cells were not sensitive to As2O3,
we did not investigate their cell cycle.
Modulation of the PI3K/Akt pathway by genetictechniques affects As2O3 sensitivity of NB-4 cells
Very recent results have highlighted that treatment with the
PI3K/Akt pathway inhibitor LY294002 markedly increased the
sensitivity of NB-4 cells to As2O3 (Ramos et al, 2005). These
results appeared surprising in light of the finding that our NB-
4 cells did not display Akt phosphorylation. Therefore, we
pretreated NB-4 cells with either LY294002 (30 lmol/l) or
wortmannin (0Æ5 lmol/l) for 12 h, then incubated them with
1 lmol/l As2O3 for 48 h. As shown in Fig 3A, the inhibitors,
used alone or in combination with As2O3, did not significantly
increase the percentage of apoptotic NB-4 cells in comparison
with controls.
We then employed a genetic approach for upregulating Akt
activity in NB-4 cells to determine whether this would result in
a decreased As2O3 sensitivity. NB-4 cells were stably transfected
with either a constitutively active Akt or a dominant negative
Akt and clones were selected. As presented in Fig 3B, two clones
(18E and 21D) expressed almost equal levels of constitutively
active Akt, as demonstrated by Western blot analysis with an
A
B
C
Fig 3. Modulation of the PI3K/Akt pathway affects As2O3 sensitivity
of NB-4 cells. (A) Cells were incubated for 12 h with either LY294002
(30 lmol/l) or wortmannin (0Æ5 lmol/l), then exposed to 1 lmol/l
As2O3 for 48 h. Samples were analysed for apoptotic cells using
Annexin V-FITC staining. Results are the mean of three different
experiments ± SD. (B) Western blot analysis for the expression of
constitutively active Akt or dominant negative Akt in stably transfected
NB-4 cell clones. Protein (80 lg/lane) was separated by SDS-PAGE
and blotted to nitrocellulose sheets. Blots were probed with an anti-
body to the c-Myc tag of Akt cDNAs, hence parental (WT ¼ wild
type) NB-4 cells were negative. Immunoreactive bands were visualised
by the ECL technique. (C) Sensitivity of NB-4 cell clones to As2O3.
Samples were incubated for 48 h in the presence of 1 lmol/l As2O3,
then the percentage of apoptotic cells was evaluated by flow cytometry
using Annexin V-FITC staining. Results are the mean of three different
experiments ±SD. The asterisks indicate significant differences
(P < 0Æ01) in comparison with parental (WT) cells. MT, mock-trans-
fected cells.
Fig 2. Cell cycle analysis of cells treated with As2O3 and/or PI3K
inhibitors. (A) NB-4 cells; (B) MOLT-4 cells. Cells were incubated for
12 h with either LY294002 (30 lmol/l) or wortmannin (0Æ5 lmol/l),
then exposed to 1 lmol/l As2O3 for 48 h. Flow cytometric analysis was
performed on PI-stained samples. A representative of three separate
experiments is shown. APO: apoptotic cells. Asterisks indicate signi-
ficant differences (P < 0Æ01) in comparison with untreated cells.
G. Tabellini et al
720 ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 130, 716–725

antibody to the c-Myc tag. Clones 11A and 13Y expressed a
comparable amount of dominant negative Akt.
When the As2O3 sensitivity of these clones was analysed, it
became evident that the two clones that overexpressed
constitutively active Akt were significantly less sensitive to
1 lmol/l As2O3 than wild type NB-4 cells or mock transfected
(i.e. transfected with the empty vector) cells. In contrast, cell
clones overexpressing a dominant negative form of Akt were as
sensitive to As2O3 as wild type cells (Fig 3C).
Selection of an As2O3 resistant NB-4 cell subline with highlevels of Ser 473 p-Akt
Previous results have indicated that it is possible to select NB-4
cell sublines that are resistant to As2O3 (e.g. Gianni et al,
1998). Moreover, recent findings have highlighted that an
As2O3-resistant NB-4 cell subline exhibited higher levels of
ERK1/2 activity and that downregulation of this survival
pathway markedly decreased As2O3 resistance (Lunghi et al,
2005). Taking also into account that exposure to apoptotic
stimuli could result in the selection of a HL60 cell subline with
an active PI3K/Akt signalling pathway (Neri et al, 2003), we
decided to investigate whether As2O3 resistance of NB-4 cell
sublines might somehow be related to upregulation of Akt
signalling. As2O3-resistant NB-4 cells were selected from
parental NB-4 cells treated with 1 lmol/l As2O3 over a
4-month period. Cell clones were then selected and screened
by flow cytometry (Tazzari et al, 2004) for high levels of Ser
473 p-Akt.
A cell subline with high levels of Ser 473 p-Akt was identified
(NB-4-ArsRES/AKT) and further analysed by Western blotting
(Fig 4A). The amount of total Akt detected in NB-4-ArsRES/
AKT was similar to that of parental NB-4 cells. However, also by
Western blotting, NB-4-ArsRES/AKT cells displayed a very high
level of Ser 473 p-Akt. Ser 473 p-Akt levels could be effectively
downregulated by incubation with two pharmacological
inhibitors of the PI3K/Akt pathway, LY294002 and wortman-
nin. As a control, we also analysed an As2O3-resistant clone,
referred to as NB-4-ArsRES, which did not display high levels of
Ser 473 p-Akt (Fig 4A) by Western blot.
The NB-4-ArsRES/AKT cells were much less sensitive to
1 lmol/l As2O3-induced apoptosis than parental cells (Fig 4B).
Treatment with either LY294002 or wortmannin significantly
increased the number of NB-4-ArsRES/AKT cells that underwent
apoptosis in response to As2O3 incubation, whereas LY294002
or wortmannin per se did not significantly increase the
percentage of apoptotic cells. In contrast, As2O3-induced
apoptosis of NB-4-ArsRES was not influenced at all by the
presence of LY294002 (Fig 4C).
PI3K/Akt inhibition enhances As2O3 sensitivity in APLprimary blasts
We next studied the effect of LY294002 on As2O3-dependent
apoptosis of primary APL blasts. Due to the limitations caused
by the amount of the samples, we decided to analyse the levels
of Ser 473 p-Akt of APL blasts prior to and after PI3K/Akt
inhibition by flow cytometry (Tazzari et al, 2004). Indeed, this
technique requires much fewer cells than Western blotting and
gives the same results. As controls we employed CEM cells,
CEM cells treated with LY294002, and NB-4 cells. LY294002
caused a dramatic fall in the levels of Ser 473 p-Akt, as revealed
by Western blotting (data not shown, but see Uddin et al,
2004). Flow cytometric analysis confirmed the positivity of
A
B
Fig 4. An NB-4 cell clone with high levels of Ser 473 –Akt (NB-4-
ARSRES/AKT) is resistant to As2O3-induced apoptosis. (A) Western blot
analysis for total Akt and Ser 473 p-Akt. Cells were incubated for 12 h
with either LY294002 (30 lmol/l) or wortmannin (0Æ5 lmol/l), then
lysed for SDS-PAGE and Western blot analysis. Immunostaining with
a monoclonal antibody to b-tubulin confirmed equal loading. Blots are
representative of three separate experiments. (B and C) Flow cyto-
metric analysis. NB-4-ARSRES/AKT (B) or NB-4-ARSRES (C) cells were
first incubated for 12 h with solvent carrier (dimethylsulphoxide) or
PI3K/Akt inhibitors (LY294002, 30 lmol/l; wortmannin, 0Æ5 lmol/
l ¼ WORT), then exposed to 1 lmol/l As2O3 for 48 h and analysed
by flow cytometry. The percentage of apoptotic cells was evaluated
by Annexin V-FITC staining (B) or TUNEL/PI technique (B, C).
Untreated cells were negative for TUNEL reaction (not shown).
A representative of three separate experiments is shown.
PI3K/Akt Inhibition and As2O3 in Leukaemia Cells
ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 130, 716–725 721

CEM cells for Ser 473 p-Akt and its decrease in LY294002-
treated samples (Fig 5A). NB-4 cells were negative for Ser 473
p-Akt, in agreement with Western blot data (see Fig 1). Of the
four APL patients, two were positive and two negative for Ser
473 p-Akt. In the two positive cases, incubation with LY294002
(12 h at 30 lmol/l) resulted in downregulation of the quantity
of Ser 473 p-Akt.
We next studied the apoptotic effect of the combined
treatment (LY294002 + As2O3) in APL blasts (Fig 5B). When
employed alone, LY294002 did not increase apoptosis in a
significant manner. The two samples with activated Akt were
less sensitive to As2O3 (either 1 or 2Æ5 lmol/l) than those
without activation. In p-Akt positive samples, even if the
As2O3 concentration was raised to 2Æ5 lmol/l, the percentage
of apoptotic cells was inferior to that detected in p-Akt
negative samples incubated with 1 lmol/l As2O3. The com-
bined treatment (1 lmol/l As2O3 + LY294002) resulted in a
dramatic increase of apoptotic cells only in samples with high
levels of activated Akt.
Abrogation of Akt expression by siRNA sensitises acuteT-leukaemia cell lines to As2O3
Recently, siRNAs have also been shown to achieve a high
degree of specificity with low toxicity in mammalian cells
acting through a degradative chain reaction catalysed by the
activation of a cellular RNA-dependent RNA polymerase
(Caplen et al, 2001; Elbashir et al, 2001). While much work
remains to optimise delivery and maintain specificity, the
therapeutic advantages of siRNAs for treatment of cancers, in
which genes are upregulated or misappropriately expressed,
show great promise (Ryther et al, 2005).
We therefore, decided to selectively knock-down Akt in
CEM, Jurkat and MOLT-4 cells by means of siRNA for
increasing sensitivity to As2O3. Transfection of Akt siRNA, but
not of the non-specific control siRNA, led to the decrease of
Akt in these cell lines without affecting the unrelated protein,
tubulin (Fig 6A). A significant increase in the number of
apoptotic cells in response to 1 lmol/l As2O3 treatment for
48 h was seen in samples in which Akt expression was
abrogated by siRNA (Fig 6B).
Discussion
The results presented in this work indicate that the generation
of apoptosis by 1 lmol/l As2O3 is potentiated by down-
modulation of the PI3K/Akt pathway in APL and T-leukaemia
cells. As2O3 also affected the cell cycle of NB-4 cells in that it
increased the number of G2/M phase cells. Therefore, it could
be suggested that apoptotic NB-4 cells derive from cells
blocked in G2/M, in agreement with the findings of others
(Gao et al, 2004). In contrast, As2O3 did not alter the cell cycle
of MOLT-4 or CEM cells but rather it directly induced
apoptosis. Conceivably, these differences are dependent on the
specific cell type.
As to NB-4 cells, a recent report showed that treatment with
LY294002 resulted in increased sensitivity to 1 lmol/l As2O3
(Ramos et al, 2005). However, these authors did not check, by
Western blot, whether their NB-4 cells had elevated levels of
Ser 473 p-Akt and whether or not there was a decrease in Akt
phosphorylation in response to LY294002. The NB-4 cells we
used for this work completely lacked Ser 473 p-Akt and,
consistently, were not further sensitised to 1 lmol/l As2O3 by
treatment with either LY294002 or wortmannin. Nevertheless,
they could be induced to become resistant to As2O3 by
overexpression of a constitutively active Akt cDNA, whereas a
dominant negative Akt cDNA was without effect. Conceivably,
Fig 5. Inhibition of PI3K/Akt sensitises APL blasts to As2O3. (A) flow
cytometric analysis of Ser 473 p-Akt in CEM and NB-4 cells, and APL
blasts. Cells were incubated for 12 h in the presence of LY294002
(30 lmol/l), then immunostained with antibody to Ser 473-Akt spe-
cific for immunocytochemistry. Controls were run with FITC-conju-
gated swine anti-rabbit IgG alone and with FITC-conjugated swine
anti-rabbit IgG plus normal rabbit IgG. (B) Flow cytometric analysis of
As2O3-induced apoptosis. Cells were incubated for 12 h with either
LY294002 (30 lmol/l) or an equivalent volume of solvent carrier
(dimethylsulphoxide), then exposed for 48 h to As2O3. The percentage
of apoptotic cells was then evaluated by Annexin V-FITC staining.
Results are the mean ± SD of a single experiment performed in trip-
licate.
G. Tabellini et al
722 ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 130, 716–725

the apparent discrepancy between their findings and ours
could be explained by the fact the NB-4 cells employed by
Ramos et al (2005) had elevated levels of active Akt.
Our results also indicate that, upon prolonged exposure to
As2O3, it was possible to isolate a cell subline, referred to as
NB-4-ArsRES/AKT, which displayed elevated levels of Ser 473
p-Akt and was resistant to 1 lmol/l As2O3. Previous reports
have indicated that As2O3 down-modulated Ser 473 p-Akt
levels of U937 cells (Choi et al, 2002; Ramos et al, 2005).
However, our results indicate that it is also possible that, in
response to As2O3, there is an upregulation of the PI3K/Akt
survival pathway that causes resistance to As2O3. In this
connection, it should be emphasised here that Akt can be
activated by many forms of cellular stress, including heat
shock, ultraviolet light, ischaemia, hypoxia and oxidative stress
(reviewed in West et al, 2002, and references therein). Stress-
induced Akt upregulation is likely to be viewed as a compen-
satory protective mechanism that cells activate to escape death.
Interestingly, the production of reactive oxygen species may be
stimulated by As2O3 in human myeloid leukaemia cells
(Perkins et al, 2000), and this might represent a sufficient
stimulus for the activation of Akt. We feel that the results
obtained with the NB-4-ArsRES/AKT cell subline might have
important consequences at the clinical level, as they might help
explaining the occurrence of resistance to As2O3. Indeed, early
relapses from As2O3 treatment within a few months were not
infrequently seen in APL patients (Huan et al, 2000). There-
fore, it would be interesting to investigate whether or not Akt
activation might be implicated in As2O3 also in APL patients.
In this connection, it is worthwhile to note that APL blasts
with activated Akt were less sensitive to As2O3 than those with
no Akt activation. Although the number of investigated
patients was low, we feel that these findings are suggestive
for a role of PI3K/Akt also in the resistance of APL patients to
As2O3 treatment in vivo. It might be hypothesised, for example,
that patients who are resistant to As2O3 have extremely
elevated levels of Ser 473 p-Akt. Indeed, the phosphorylation
levels of this residue have been demonstrated to display a great
variability among patients with AML (Xu et al, 2003).
However, it is important to emphasise that our findings seem
to indicate that activation of the PI3K/Akt pathway cannot be
considered per se the only mechanism that leads to As2O3
resistance in NB-4 cells and T-leukaemia cell lines. Indeed, an
As2O3-resistant clone (NB-4-ArsRES) was isolated that did not
display elevated levels of Akt. Moreover, CEM, Jurkat and
MOLT-4 cells have elevated basal levels of Ser 473 p-Akt, due to
the lack of the lipid phosphatase PTEN which downregulates
the levels of 3¢ phosphorylated inositol lipids (Uddin et al,
2004). While CEM and MOLT-4 were sensitive to 1 lmol/l
As2O3, Jurkat cells were resistant. However, Akt knock-down by
siRNA increased 1 lmol/l As2O3 sensitivity of all of these three
cell lines. Therefore, it is conceivable that other signalling
networks also mediate As2O3 resistance of APL cells, for
example the Erk 1/2 survival pathway (Lunghi et al, 2005).
In conclusion, our results strengthen the hypothesis that the
PI3K/Akt pathway may be one of the factors important for
resistance to As2O3-induced apoptosis in a variety of human
acute leukaemia cell lines. Moreover, our findings indicated
that down-modulation of this survival pathway could increase
APL blast sensitivity to As2O3. Therefore, it might be that
future innovative therapeutic strategies for acute leukaemias
A
B
Fig 6. Abrogation of Akt expression sensitises acute T-leukaemia cell
lines to As2O3. (A) Western blot analysis. Cells were transfected with
the indicated siRNA for 48 h. Subsequently, cells were harvested for
Western blot analysis, which was performed as indicated in legend to
Fig 1. (B) Samples in which Akt had been knocked-down by siRNA
transfection, were incubated for 48 h in the presence of 1 lmol/l As2O3
for 48 h, then the percentage of apoptotic cells was evaluated by flow
cytometry using Annexin V-FITC staining. Results are the mean of
three different experiments ±SD. Asterisks indicate significant differ-
ences (P < 0Æ01) in comparison with untreated cells.
PI3K/Akt Inhibition and As2O3 in Leukaemia Cells
ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 130, 716–725 723

will consist of inhibition of the PI3K/Akt pathway in addition
to As2O3. PI3K/Akt inhibition might be achieved not only
through the use of selective pharmacological inhibitors, but
also by means of siRNA, which is rapidly establishing itself as a
very promising technology for specific gene silencing also in
haematological oncology (Woessmann et al, 2003).
In this regard, it is worth emphasising that the first phase I
clinical trial involving a PI3K/Akt inhibitor, perifosine (Kon-
dapaka et al, 2003), was recently performed on patients with
incurable solid malignancies. These results suggested perifosine
activity in sarcoma and perhaps renal cell carcinoma, thus
justifying additional investigation of this agent in a phase II
sarcoma trial (Van Ummersen et al, 2005). Thus, perifosine
might also be employed for the treatment of patients with
haematological malignancies in the future.
Acknowledgements
This work was supported by grants from: AIRC, Italian MIUR
Cofin 2003 and FIRB 2001, Selected Topics Research Fund
from Bologna University, ‘Hairshow A.I.L.’, Fondazione del
Monte di Bologna e Ravenna.
References
Bazarbachi, A., Ghez, D., Lepelletier, Y., Nasr, R., de The, H.,
El-Sabban, M.E. & Hermine, O. (2004) New therapeutic approaches
for adult T-cell leukaemia. Lancet Oncology, 5, 664–672.
Bortul, R., Tazzari, P.L., Billi, A.M., Tabellini, G., Mantovani, I.,
Cappellini, A., Grafone, T., Martinelli, G., Conte, R. & Martelli,
A.M. (2005) Deguelin, a PI3K/Akt inhibitor, enhances chemo-
sensitivity of leukemia cells with an active PI3K/Akt pathway. British
Journal of Haematology, 129, 677–686.
Brazil, D.P., Yang, Z.Z. & Hemmings, B.A. (2004) Advances in protein
kinase B signalling: AKTion on multiple fronts. Trends in Bio-
chemical Sciences, 29, 233–242.
Caplen, N.J., Parrish, S., Imani, F., Fire, A. & Morgan, R.A. (2001)
Specific inhibition of gene expression by small double-stranded
RNAs in invertebrate and vertebrate systems. Proceedings of the
National Academy of Sciences of the United States of America, 98,
9742–9747.
Cappellini, A., Tabellini, G., Zweyer, M., Bortul, R., Tazzari, P.L., Billi,
A.M., Fala, F., Cocco, L. & Martelli, A.M. (2003) The phospho-
inositide 3-kinase/Akt pathway regulates cell cycle progression
of HL60 human leukemia cells through cytoplasmic relocalization of
the cyclin-dependent kinase inhibitor p27(Kip1) and control of
cyclin D1 expression. Leukemia, 17, 2157–2167.
Choi, Y.J., Park, J.W., Suh, S.I., Mun, K.C., Bae, J.H., Song, D.K., Kim,
S.P. & Kwon, T.K. (2002) Arsenic trioxide-induced apoptosis in
U937 cells involves generation of reactive oxygen species and
inhibition of Akt. International Journal of Oncology, 21, 603–610.
Cross, T.G., Scheel-Toellner, D., Henriquez, N.V., Deacon, E., Salmon,
M. & Lord, J.M. (2000) Serine/threonine protein kinases and
apoptosis. Experimental Cell Research, 256, 34–41.
Douer, D. & Tallman, M.S. (2005) Arsenic trioxide: new clinical ex-
perience with an old medication in hematologic malignancies.
Journal of Clinical Oncology, 23, 2396–2410.
Elbashir, S.M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K. &
Tuschl, T. (2001) Duplexes of 21-nucleotide RNAs mediate RNA
interference in cultured mammalian cells. Nature, 411, 494–498.
Evens, A.M., Tallman, M.S. & Gartenhaus, R.B. (2004) The potential of
arsenic trioxide in the treatment of malignant disease: past, present,
and future. Leukemia Research, 28, 891–900.
Gao, F., Yi, J., Yuan, J.Q., Shi, G.Y. & Tang, X.M. (2004) The cell cycle
related apoptotic susceptibility to arsenic trioxide is associated with
the level of reactive oxygen species. Cell Research, 14, 81–85.
Gianni, M., Koken, M.H., Chelbi-Alix, M.K., Benoit, G., Lanotte, M.,
Chen, Z. & de The, H. (1998) Combined arsenic and retinoic acid
treatment enhances differentiation and apoptosis in arsenic-resistant
NB4 cells. Blood, 91, 4300–4310.
Hanada, M., Feng, J. & Hemmings, B.A. (2004) Structure, regulation
and function of PKB/AKT – a major therapeutic target. Biochimica et
Biophysica Acta, 1697, 3–16.
Hu, X.M., Hirano, T. & Oka, K. (2003) Arsenic trioxide induces
apoptosis equally in T lymphoblastoid leukemia MOLT-4 cells and
P-gp-expressing daunorubicin-resistant MOLT-4 cells. Cancer Che-
motherapy and Pharmacology, 51, 119–126.
Huan, S.Y., Yang, C.H. & Chen, Y.C. (2000) Arsenic trioxide therapy
for relapsed acute promyelocytic leukemia: an useful salvage therapy.
Leukemia and Lymphoma, 38, 283–293.
Kondapaka, S.B., Singh, S.S., Dasmahapatra, G.P., Sausville, E.A. &
Roy, K.K. (2003) Perifosine, a novel alkylphospholipid, inhibits
protein kinase B activation. Molecular Cancer Therapeutics, 2, 1093–
1103.
Li, X., Traganos, F., Melamed, M.R. & Darzynkiewicz, Z. (1995) Single-
step procedure for labeling DNA strand breaks with fluorescein- or
BODIPY-conjugated deoxynucleotides: detection of apoptosis and
bromodeoxyuridine incorporation. Cytometry, 20, 172–180.
Lunghi, P., Tabilio, A., Dall’Aglio, P.P., Ridolo, E., Carlo-Stella, C.,
Pelicci, P.G. & Bonati, A. (2003) Downmodulation of ERK activity
inhibits the proliferation and induces the apoptosis of primary acute
myelogenous leukemia blasts. Leukemia, 17, 1783–1793.
Lunghi, P., Tabilio, A., Lo-Coco, F., Pelicci, P.G. & Bonati, A. (2005)
Arsenic trioxide (ATO) and MEK1 inhibition synergize to induce
apoptosis in acute promyelocytic leukemia cells. Leukemia, 19, 234–
244.
Martelli, A.M., Tabellini, G., Bortul, R., Tazzari, P.L., Cappellini, A.,
Billi, A.M. & Cocco, L. (2005) Involvement of the phosphoinositide
3-kinase/Akt signaling pathway in the resistance to therapeutic
treatments of human leukemias. Histology and Histopathology, 20,
239–252.
Milella, M., Kornblau, S.M., Estrov, Z., Carter, B.Z., Lapillonne, H.,
Harris, D., Konopleva, M., Zhao, S., Estey, E. & Andreeff, M. (2001)
Therapeutic targeting of the MEK/MAPK signal transduction
module in acute myeloid leukemia. Journal of Clinical Investigation,
108, 851–859.
Miller, Jr, W.H. Schipper, H.M., Lee, J.S., Singer, J. & Waxman, S.
(2002) Mechanisms of action of arsenic trioxide. Cancer Research,
62, 3893–3903.
Neri, L.M., Borgatti, P., Tazzari, P.L., Bortul, R., Cappellini, A.,
Tabellini, G., Bellacosa, A., Capitani, S. & Martelli, A.M. (2003) The
phosphoinositide 3-kinase/AKT1 pathway involvement in drug and
all-trans-retinoic acid resistance of leukemia cells. Molecular Cancer
Research, 1, 234–246.
Perkins, C., Kim, C.N., Fang, G. & Bhalla, K.N. (2000) Arsenic induces
apoptosis of multidrug-resistant human myeloid leukemia cells that
G. Tabellini et al
724 ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 130, 716–725

express Bcr-Abl or overexpress MDR, MRP, Bcl-2, or Bcl-x(L).
Blood, 95, 1014–1022.
Platanias, L.C. (2003) Map kinase signaling pathways and hematologic
malignancies. Blood, 101, 4667–4679.
Puccetti, E. & Ruthardt, M. (2004) Acute promyelocytic leukemia:
PML/RAR and the leukemic stem cell. Leukemia, 18, 1169–
1175.
Ramos, A.M., Fernandez, C., Amran, D., Sancho, P., de Blas, E. & Aller,
P. (2005) Pharmacological inhibitors of PI3K/Akt potentiate the
apoptotic action of the antileukemic drug arsenic trioxide via
glutathione depletion and increased peroxide accumulation in
myeloid leukemia cells. Blood, 105, 4013–4020.
Recher, C., Chopin, M., Raffoux, E., Pierron, G., Poupon, J., Sigaux, F.,
Dombret, H. & Stern, M.H. (2002) In vitro and in vivo effectiveness
of arsenic trioxide against murine T-cell prolymphocytic leukaemia.
British Journal of Haematology, 117, 343–350.
Rojewski, M.T., Baldus, C., Knauf, W., Thiel, E. & Schrezenmeier, H.
(2002) Dual effects of arsenic trioxide (As2O3) on non-acute pro-
myelocytic leukaemia myeloid cell lines: induction of apoptosis and
inhibition of proliferation. British Journal of Haematology, 116, 555–
563.
Ryther, R.C., Flynt, A.S., Phillips, III, J.A. & Patton, J.G. (2005) siRNA
therapeutics: big potential from small RNAs. Gene Therapy, 12,
5–11.
Soignet, S.L., Frankel, S.R., Douer, D., Tallman, M.S., Kantarjian, H.,
Calleja, E., Stone, R.M., Kalaycio, M., Scheinberg, D.A., Steinherz,
P., Sievers, E.L., Coutre, S., Dahlberg, S., Ellison, R. & Warrell, Jr,
R.P. (2001) United States multicenter study of arsenic trioxide in
relapsed acute promyelocytic leukemia. Journal of Clinical Oncology,
19, 3852–3860.
Tabellini, G., Cappellini, A., Tazzari, P.L., Fala, F., Billi, A.M., Manzoli,
L., Cocco, L. & Martelli, A.M. (2005) Phosphoinositide 3-kinase/Akt
involvement in arsenic trioxide resistance of human leukemia cells.
Journal of Cellular Physiology, 202, 623–634.
Tazzari, P.L., Cappellini, A., Grafone, T., Mantovani, I., Ricci, F., Billi,
A.M., Ottaviani, E., Conte, R., Martinelli, G. & Martelli, A.M. (2004)
Detection of serine 473 phosphorylated Akt in acute myeloid leu-
kaemia blasts by flow cytometry. British Journal of Haematology, 126,
675–681.
Thøger-Andersen, A.-S. & Junker, S. (1994) Simple and efficient
recovery of rare living lymphoid cells from a vast majority of dead
cells. Nucleic Acids Research, 22, 5769–5770.
Uddin, S., Hussain, A., Al-Hussein, K., Platanias, L.C. & Bhatia, K.G.
(2004) Inhibition of phosphatidylinositol 3¢-kinase induces prefer-
entially killing of PTEN-null T leukemias through AKT pathway.
Biochemical and Biophysical Research Communications, 320, 932–938.
Van Ummersen, L., Binger, K., Volkman, J., Marnocha, R., Tutsch, K.,
Kolesar, J., Arzoomanian, R., Alberti, D. & Wilding G. (2005) A
phase I trial of perifosine (NSC 639966) on a loading dose/main-
tenance dose schedule in patients with advanced cancer. Clinical
Cancer Research, 10, 7450–7456.
Vermes, I., Haanen, C., Steffens-Nakken, H. & Reutelingsperger, C.
(1995) A novel assay for apoptosis. Flow cytometric detection of
phosphatidylserine expression on early apoptotic cells using fluor-
escein labelled Annexin V. Journal of Immunological Methods, 184,
39–51.
Wada, T. & Penninger, J.M. (2004) Mitogen-activated protein kinases
in apoptosis regulation. Oncogene, 23, 2838–2849.
West, K.A., Castillo, S.S. & Dennis, P.A. (2002) Activation of the PI3K/
Akt pathway and chemotherapeutic resistance. Drug Resistance
Updates, 5, 234–248.
Woessmann, W., Damm-Welk, C., Fuchs, U. & Borkhardt, A. (2003)
RNA interference: new mechanisms for targeted treatment? Reviews
in Clinical and Experimental Hematology, 7, 270–291.
Xu, Z., Stokoe, D., Kane, L.P. & Weiss, A. (2002) The inducible
expression of the tumor suppressor gene PTEN promotes apoptosis
and decreases cell size by inhibiting the PI3K/Akt pathway in Jurkat
T cells. Cell Growth and Differentiation, 13, 285–296.
Xu, Q., Simpson, S.E., Scialla, T.J., Bagg, A. & Carroll, M. (2003)
Survival of acute myeloid leukemia cells requires PI3 kinase acti-
vation. Blood, 102, 972–980.
Zhao, S., Konopleva, M., Cabreira-Hansen, M., Xie, Z., Hu, W.,
Milella, M., Estrov, Z., Mills, G.B. & Andreeff, M. (2004) Inhibition
of phosphatidylinositol 3-kinase dephosphorylates BAD and pro-
motes apoptosis in myeloid leukemias. Leukemia, 18, 267–275.
Zhu, J., Chen, Z., Lallemand-Breitenbach, V. & de The, H. (2002) How
acute promyelocytic leukaemia revived arsenic. Nature Reviews on
Cancer, 2, 705–713.
PI3K/Akt Inhibition and As2O3 in Leukaemia Cells
ª 2005 Blackwell Publishing Ltd, British Journal of Haematology, 130, 716–725 725
Copyright © 2022 FDOKUMEN