
Regulation of glucose metabolism in T cells: new insight into the role of Phosphoinositide 3-kinases
7
MINI REVIEW ARTICLE published: 07 August 2012 doi: 10.3389/fimmu.2012.00247 Regulation of glucose metabolism in T cells: new insight into the role of phosphoinositide 3-kinases David K. Finlay 1,2 * 1 School of Biochemistry and Immunology,Trinity Biomedical Sciences Institute,Trinity College Dublin, Dublin, Ireland 2 School of Pharmacy and Pharmaceutical Sciences,Trinity Biomedical Sciences Institute,Trinity College Dublin, Dublin, Ireland Edited by: Klaus Okkenhaug, Babraham Institute, UK Reviewed by: Stephen Geoffrey Ward, University of Bath, UK Lawrence Kane, University of Pittsburgh School of Medicine, USA *Correspondence: David K. Finlay, School of Biochemistry and Immunology,Trinity Biomedical Sciences Institute,Trinity College, Dublin 2, Ireland. e-mail: fi[email protected] Naïve T cells are relatively quiescent cells that only require energy to prevent atrophy and for survival and migration. However, in response to developmental or extrinsic cues T cells can engage in rapid growth and robust proliferation, produce of a range of effec- tor molecules and migrate through peripheral tissues. To meet the significantly increased metabolic demands of these activities, T cells switch from primarily metabolizing glucose to carbon dioxide through oxidative phosphorylation to utilizing glycolysis to convert glucose to lactate (termed aerobic glycolysis). This metabolic switch allows glucose to be used as a source of carbon to generate biosynthetic precursors for the production of protein, DNA, and phospholipids, and is crucial for T cells to meet metabolic demands. Phosphoinositide 3-kinases (PI3K) are a family of inositol lipid kinases linked with a broad range of cellular functions in T lymphocytes that include cell growth, proliferation, metabolism, differenti- ation, survival, and migration. Initial research described a critical role for PI3K signaling through Akt (also called protein kinase B) for the increased glucose uptake and glycolysis that accompanies T cell activation. This review article relates this original research with more recent data and discusses the evidence for and against a role for PI3K in regulating the metabolic switch to aerobic glycolysis in T cells. Keywords: PI3K, Glucose metabolism, Akt,T lymphocyte, aerobic glycolysis, c-Myc, PDK1 PI3K IN T CELLS Class 1 phosphoinositide 3-kinases (PI3K), lipid kinases that phos- phorylate phosphatidylinositol-(4,5)-bisphosphate [PI(4,5)P 2 ] to generate the lipid signaling molecule phosphatidylinositol-(3,4,5)- trisphosphate [PI(3,4,5)P 3 ], play a crucial role in many aspects of T cell biology (Okkenhaug and Vanhaesebroeck, 2003). Class 1 PI3Ks consist of a catalytic subunit, responsible for the lipid kinase activity, and an adapter subunit which links the catalytic subunit to upstream activating signals. They are subdivided into Class 1A; p110α, β, or δ catalytic subunits coupled to p85 adapter subunits, and Class 1B; p110γ catalytic subunit coupled to the p101 adapter protein. The p85 subunit couples Class 1A PI3Ks to docking sites created by tyrosine-kinase signaling while the Class IB p101 links the p110γ kinase with G-protein-coupled recep- tors (Vanhaesebroeck et al., 2010). The levels of the lipid signaling molecule PI(3,4,5)P 3 are coordinately regulated by both class 1 PI3Ks and by the action of 3 and 5 phosphatases PTEN and SHIP1 that dephosphorylate PI(3,4,5)P 3 to generate PI(4,5)P 2 and PI(3,4)P 2 respectively (Figure 1A). PI(3,4,5)P 3 is an important lipid signaling molecule in T cells, present at low levels in naïve T cells and elevated in response to signaling initiated by the T cell receptor (TCR) and various cytokine and chemokine receptors (Koyasu, 2003). Following TCR engagement by antigen presenting cells (APC) levels of PI(3,4,5)P 3 accumulate in the plasma membrane and are maintained for prolonged periods, requiring continual TCR engagement and activation of PI3K (Costello et al., 2002; Har- riague and Bismuth, 2002; Huppa et al., 2003). Signals from co- stimulatory molecules such as CD28 are important for sustaining PI(3,4,5)P 3 levels following TCR engagement but are not in them- selves sufficient to stimulate PI3K as PI(3,4,5)P 3 levels are only induced when the TCR engages APC presenting cognate pep- tide antigen (Costello et al., 2002; Garcon et al., 2007). Other physiological stimuli that can stimulate PI(3,4,5)P 3 levels in T cells include cytokines and chemokines (Koyasu, 2003). While PI(3,4,5)P 3 levels are induced by a number of cytokines it should be noted that the potency of these different cytokines can vary, e.g., IL2 maintains high levels of PI(3,4,5)P 3 while IL15 maintains comparatively low levels (Cornish et al., 2006; Sinclair et al., 2008). In the thymus, PI3Kδ and PI3Kγ are the major isoforms required in developing thymocytes. While deletion of PI3Kδ or γ individually does not have a pronounced effect on thymopoiesis, deletion or inactivation of both isoforms in developing T cells results in a block early in thymocyte development at the CD4 CD8 double negative (DN) stage (Sasaki et al., 2000; Okkenhaug et al., 2002; Webb et al., 2005; Swat et al., 2006; Ji et al., 2007). Negative regulation of PI(3,4,5)P 3 signaling is also important in developing thymocytes. Thus, deletion of PTEN in early thymocyte progen- itors results in constitutive PI(3,4,5)P 3 signaling that leads to the development of T cell leukemia or lymphoma (Suzuki et al., 2001; Hagenbeek and Spits, 2008; Finlay et al., 2009). In contrast to developing thymocytes, in mature T cells PI3Kδ appears to be the major PI3K isoform responsible for promoting PI(3,4,5)P 3 signal- ing in activated T cells. Thus, a point mutation that makes p110δ catalytically inactive (D910A) abolishes PI(3,4,5)P 3 signaling in activated T cell subsets (Okkenhaug et al., 2002, 2006; Macintyre et al., 2011). www.frontiersin.org August 2012 | Volume 3 | Article 247 | 1
Transcript of Regulation of glucose metabolism in T cells: new insight into the role of Phosphoinositide 3-kinases

“fimmu-03-00247” — 2012/8/6 — 11:02 — page 1 — #1
MINI REVIEW ARTICLEpublished: 07 August 2012
doi: 10.3389/fimmu.2012.00247
Regulation of glucose metabolism in T cells: new insightinto the role of phosphoinositide 3-kinasesDavid K. Finlay1,2*
1 School of Biochemistry and Immunology, Trinity Biomedical Sciences Institute, Trinity College Dublin, Dublin, Ireland2 School of Pharmacy and Pharmaceutical Sciences, Trinity Biomedical Sciences Institute, Trinity College Dublin, Dublin, Ireland
Edited by:
Klaus Okkenhaug, BabrahamInstitute, UK
Reviewed by:
Stephen Geoffrey Ward, University ofBath, UKLawrence Kane, University ofPittsburgh School of Medicine, USA
*Correspondence:
David K. Finlay, School ofBiochemistry and Immunology, TrinityBiomedical Sciences Institute, TrinityCollege, Dublin 2, Ireland.e-mail: [email protected]
Naïve T cells are relatively quiescent cells that only require energy to prevent atrophyand for survival and migration. However, in response to developmental or extrinsic cuesT cells can engage in rapid growth and robust proliferation, produce of a range of effec-tor molecules and migrate through peripheral tissues. To meet the significantly increasedmetabolic demands of these activities,T cells switch from primarily metabolizing glucose tocarbon dioxide through oxidative phosphorylation to utilizing glycolysis to convert glucoseto lactate (termed aerobic glycolysis). This metabolic switch allows glucose to be used asa source of carbon to generate biosynthetic precursors for the production of protein, DNA,and phospholipids, and is crucial for T cells to meet metabolic demands. Phosphoinositide3-kinases (PI3K) are a family of inositol lipid kinases linked with a broad range of cellularfunctions in T lymphocytes that include cell growth, proliferation, metabolism, differenti-ation, survival, and migration. Initial research described a critical role for PI3K signalingthrough Akt (also called protein kinase B) for the increased glucose uptake and glycolysisthat accompanies T cell activation. This review article relates this original research withmore recent data and discusses the evidence for and against a role for PI3K in regulatingthe metabolic switch to aerobic glycolysis in T cells.
Keywords: PI3K, Glucose metabolism, Akt,T lymphocyte, aerobic glycolysis, c-Myc, PDK1
PI3K IN T CELLSClass 1 phosphoinositide 3-kinases (PI3K), lipid kinases that phos-phorylate phosphatidylinositol-(4,5)-bisphosphate [PI(4,5)P2] togenerate the lipid signaling molecule phosphatidylinositol-(3,4,5)-trisphosphate [PI(3,4,5)P3], play a crucial role in many aspectsof T cell biology (Okkenhaug and Vanhaesebroeck, 2003). Class1 PI3Ks consist of a catalytic subunit, responsible for the lipidkinase activity, and an adapter subunit which links the catalyticsubunit to upstream activating signals. They are subdivided intoClass 1A; p110α, β, or δ catalytic subunits coupled to p85 adaptersubunits, and Class 1B; p110γ catalytic subunit coupled to thep101 adapter protein. The p85 subunit couples Class 1A PI3Ks todocking sites created by tyrosine-kinase signaling while the ClassIB p101 links the p110γ kinase with G-protein-coupled recep-tors (Vanhaesebroeck et al., 2010). The levels of the lipid signalingmolecule PI(3,4,5)P3 are coordinately regulated by both class 1PI3Ks and by the action of 3′ and 5′ phosphatases PTEN andSHIP1 that dephosphorylate PI(3,4,5)P3 to generate PI(4,5)P2 andPI(3,4)P2 respectively (Figure 1A).
PI(3,4,5)P3 is an important lipid signaling molecule in Tcells, present at low levels in naïve T cells and elevated inresponse to signaling initiated by the T cell receptor (TCR)and various cytokine and chemokine receptors (Koyasu, 2003).Following TCR engagement by antigen presenting cells (APC)levels of PI(3,4,5)P3 accumulate in the plasma membrane andare maintained for prolonged periods, requiring continual TCRengagement and activation of PI3K (Costello et al., 2002; Har-riague and Bismuth, 2002; Huppa et al., 2003). Signals from co-stimulatory molecules such as CD28 are important for sustaining
PI(3,4,5)P3 levels following TCR engagement but are not in them-selves sufficient to stimulate PI3K as PI(3,4,5)P3 levels are onlyinduced when the TCR engages APC presenting cognate pep-tide antigen (Costello et al., 2002; Garcon et al., 2007). Otherphysiological stimuli that can stimulate PI(3,4,5)P3 levels in Tcells include cytokines and chemokines (Koyasu, 2003). WhilePI(3,4,5)P3 levels are induced by a number of cytokines itshould be noted that the potency of these different cytokinescan vary, e.g., IL2 maintains high levels of PI(3,4,5)P3 whileIL15 maintains comparatively low levels (Cornish et al., 2006;Sinclair et al., 2008).
In the thymus, PI3Kδ and PI3Kγ are the major isoformsrequired in developing thymocytes. While deletion of PI3Kδ or γ
individually does not have a pronounced effect on thymopoiesis,deletion or inactivation of both isoforms in developing T cellsresults in a block early in thymocyte development at the CD4 CD8double negative (DN) stage (Sasaki et al., 2000; Okkenhaug et al.,2002; Webb et al., 2005; Swat et al., 2006; Ji et al., 2007). Negativeregulation of PI(3,4,5)P3 signaling is also important in developingthymocytes. Thus, deletion of PTEN in early thymocyte progen-itors results in constitutive PI(3,4,5)P3 signaling that leads to thedevelopment of T cell leukemia or lymphoma (Suzuki et al., 2001;Hagenbeek and Spits, 2008; Finlay et al., 2009). In contrast todeveloping thymocytes, in mature T cells PI3Kδ appears to be themajor PI3K isoform responsible for promoting PI(3,4,5)P3 signal-ing in activated T cells. Thus, a point mutation that makes p110δ
catalytically inactive (D910A) abolishes PI(3,4,5)P3 signaling inactivated T cell subsets (Okkenhaug et al., 2002, 2006; Macintyreet al., 2011).
www.frontiersin.org August 2012 | Volume 3 | Article 247 | 1

“fimmu-03-00247” — 2012/8/6 — 11:02 — page 2 — #2
Finlay Glucose Metabolism in T cells
FIGURE 1 | PI3K and Akt signaling. (A) PI(4,5)P2 is phosphorylated byClass 1 PI3K isoforms generating the second messenger moleculePI(3,4,5)P3. Levels of PI(3,4,5)P3 are negatively regulated by the lipidphosphatases PTEN and SHIP. Binding of the PH domain of Akt to PI(3,4,5)P3in the membrane results in a conformational change that allows for thephosphorylation of Akt on key residues(T308 and S473) by PDK1 andmTORC2. The recruitment of PDK1, via its PH domain, to the site of
PI(3,4,5)P3 is required for efficient Akt activation. (B) Active Aktphosphorylates Foxo transcription factors on multiple sites resulting intheir translocation to the cytoplasm where they are retained in complexwith 14-3-3 proteins. mTORC1 can be activated downstream of Akt, thoughthis does not occur in all T cell types. (C) PDK1 also phosphorylates andactivates a number of other members of the AGC kinase family in a PI3Kindependent manner.
PI3K SIGNALINGPI(3,4,5)P3 acts as a signaling molecule through its interactionwith the pleckstrin homology (PH) domains of a diverse arrayof signal transduction proteins. These proteins include Akt (alsocalled PKB), Tec family kinases, and guanine-nucleotide-exchangeproteins for Rho family GTPases. This interaction primarily con-trols the subcellular localization of PH domain containing proteinsbut can also control protein conformation and enzyme activity.Consider, for example, the activation of Akt, which is the bestcharacterized PI(3,4,5)P3 effector in T cells (for reviews Alessiand Cohen, 1998; Hanada et al., 2004; Cameron et al., 2007). Theinteraction of PI(3,4,5)P3 with the PH domain of Akt stimu-lates its kinase activity by inducing a conformational change thatallows Akt to be phosphorylated on threonine 308 and serine 473by its upstream activating kinases phosphoinositide-dependentkinase 1 (PDK1) and mechanistic Target Of Rapamycin Complex2 (mTORC2) respectively (Figure 1A; Calleja et al., 2007). PDK1
also contains a PH domain and co-localization of Akt and PDK1 tosites of PI(3,4,5)P3 is required for efficient Akt activation (Bayascaset al., 2008; Waugh et al., 2009). Once activated, Akt phosphory-lates a number of important signaling molecules including theFoxo transcription factors (Manning and Cantley, 2007). Foxotranscription factors localize to the nucleus where they promotethe expression of target genes. Once phosphorylated by Akt,Foxos translocate into the cytoplasm where they are retainedthrough their interaction with 14-3-3 proteins (Figure 1B;Coffer and Burgering, 2004; Burgering, 2008). PI3K/Akt signalingalso activates the mTOR Complex 1 (mTORC1) in many cellu-lar systems through multiple mechanisms (Laplante and Sabatini,2009). mTORC1 is an important regulator of cellular metabolismthat senses environmental cues such as nutrient availability andenergy homeostasis (Delgoffe and Powell, 2009). However, it isnow becoming apparent that mTORC1 activity is not universallydependent upon PI3K/Akt signaling in T cells. Thus, in activated
Frontiers in Immunology | T Cell Biology August 2012 | Volume 3 | Article 247 | 2
“fimmu-03-00247” — 2012/8/6 — 11:02 — page 3 — #3
Finlay Glucose Metabolism in T cells
CD8 cells, mTORC1 activity is not blocked by the disruptionof PI3K/Akt signaling by various pharmacological and geneticstrategies (unpublished data; Macintyre et al., 2011).
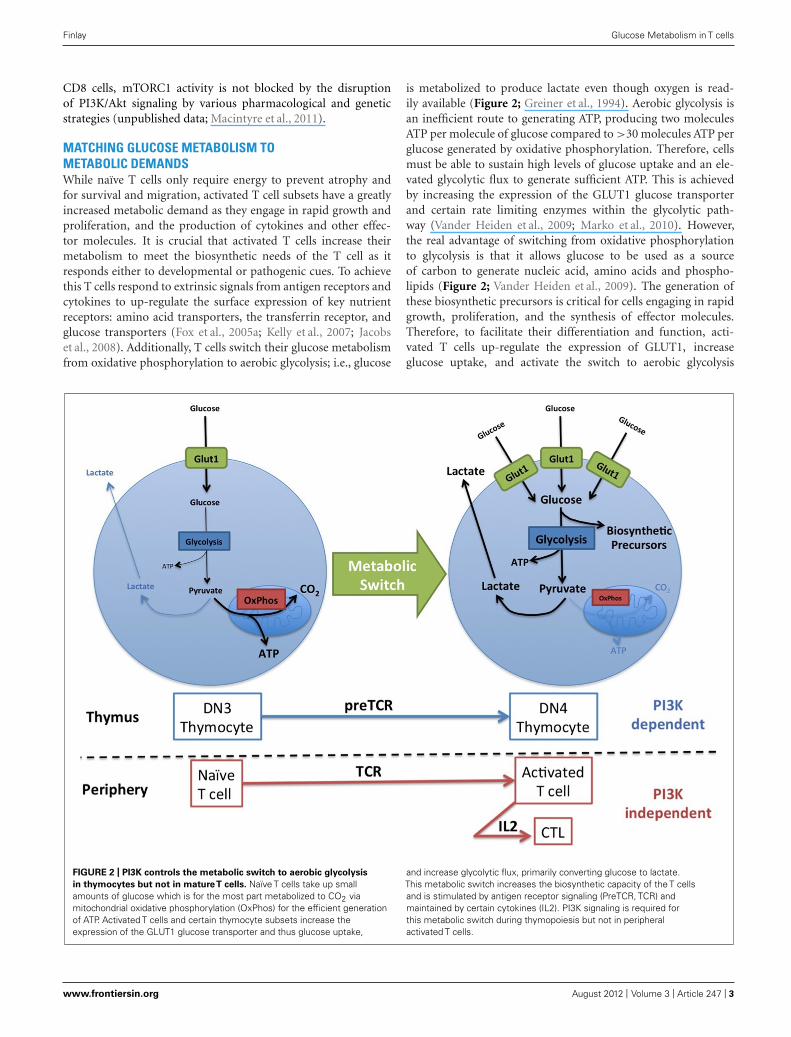
MATCHING GLUCOSE METABOLISM TOMETABOLIC DEMANDSWhile naïve T cells only require energy to prevent atrophy andfor survival and migration, activated T cell subsets have a greatlyincreased metabolic demand as they engage in rapid growth andproliferation, and the production of cytokines and other effec-tor molecules. It is crucial that activated T cells increase theirmetabolism to meet the biosynthetic needs of the T cell as itresponds either to developmental or pathogenic cues. To achievethis T cells respond to extrinsic signals from antigen receptors andcytokines to up-regulate the surface expression of key nutrientreceptors: amino acid transporters, the transferrin receptor, andglucose transporters (Fox et al., 2005a; Kelly et al., 2007; Jacobset al., 2008). Additionally, T cells switch their glucose metabolismfrom oxidative phosphorylation to aerobic glycolysis; i.e., glucose
is metabolized to produce lactate even though oxygen is read-ily available (Figure 2; Greiner et al., 1994). Aerobic glycolysis isan inefficient route to generating ATP, producing two moleculesATP per molecule of glucose compared to >30 molecules ATP perglucose generated by oxidative phosphorylation. Therefore, cellsmust be able to sustain high levels of glucose uptake and an ele-vated glycolytic flux to generate sufficient ATP. This is achievedby increasing the expression of the GLUT1 glucose transporterand certain rate limiting enzymes within the glycolytic path-way (Vander Heiden et al., 2009; Marko et al., 2010). However,the real advantage of switching from oxidative phosphorylationto glycolysis is that it allows glucose to be used as a sourceof carbon to generate nucleic acid, amino acids and phospho-lipids (Figure 2; Vander Heiden et al., 2009). The generation ofthese biosynthetic precursors is critical for cells engaging in rapidgrowth, proliferation, and the synthesis of effector molecules.Therefore, to facilitate their differentiation and function, acti-vated T cells up-regulate the expression of GLUT1, increaseglucose uptake, and activate the switch to aerobic glycolysis
FIGURE 2 | PI3K controls the metabolic switch to aerobic glycolysis
in thymocytes but not in matureT cells. Naïve T cells take up smallamounts of glucose which is for the most part metabolized to CO2 viamitochondrial oxidative phosphorylation (OxPhos) for the efficient generationof ATP. Activated T cells and certain thymocyte subsets increase theexpression of the GLUT1 glucose transporter and thus glucose uptake,
and increase glycolytic flux, primarily converting glucose to lactate.This metabolic switch increases the biosynthetic capacity of the T cellsand is stimulated by antigen receptor signaling (PreTCR, TCR) andmaintained by certain cytokines (IL2). PI3K signaling is required forthis metabolic switch during thymopoiesis but not in peripheralactivated T cells.
www.frontiersin.org August 2012 | Volume 3 | Article 247 | 3

“fimmu-03-00247” — 2012/8/6 — 11:02 — page 4 — #4
Finlay Glucose Metabolism in T cells
(Brand et al., 1984; Greiner et al., 1994; Frauwirth et al., 2002;Wofford et al., 2008).
PI3K AND GLUCOSE METABOLISM IN THE THYMUSIn the thymus discrete subpopulations, e.g., DN4 thymocytes,engage in rapid growth and robust proliferation. PI3K and Aktsignaling is crucial in allowing these thymocyte subsets to matchtheir metabolism with metabolic demands (Ciofani and Zuniga-Pflucker, 2005; Juntilla et al., 2007; Kelly et al., 2007; Finlay et al.,2010). Thus, mice lacking Akt or PDK1 or both PI3Kδ and γ iso-forms during early thymopoiesis have a profound developmentalblock at the DN3/DN4 stage of T cell development(Hinton et al.,2004; Webb et al., 2005; Swat et al., 2006; Fayard et al., 2007; Juntillaet al., 2007; Mao et al., 2007). In the absence of PI3K/PDK1/Aktsignaling DN4 thymocytes fail to up-regulate the expression ofthe glucose transporter, GLUT1, and also the expression of otherkey nutrient receptors for the uptake of amino acids (CD98,component of the L-amino acid transporter) and iron (transfer-rin receptor, CD71). Increased provision of these nutrients is akey requirement for these cells to meet the metabolic demandsof rapid growth and proliferation and PI3K/PDK1/Akt deficientDN4 thymocytes that fail to do so atrophy and fail to develop(Juntilla et al., 2007; Kelly et al., 2007). Therefore, in T cells devel-oping in the thymus, PI3K and Akt signaling is crucial to allowcertain thymocyte subsets to match glucose metabolism withmetabolic demands.
PI3K AND GLUCOSE METABOLISM IN ACTIVATED T CELLSThe metabolic switch to aerobic glycolysis is crucial during theactivation and differentiation of T cells in the periphery. Thus,limiting glucose availability in activating T cells compromisesTCR induced growth and proliferation and also the expressionof certain effector molecules such as interferon γ (IFNγ; Chamet al., 2008; Jacobs et al., 2008). The transcription factor c-Mycis crucial for the metabolic switch in glucose metabolism thataccompanies the activation of naïve T cells (Wang et al., 2011).Accordingly, deletion of c-Myc in naïve T cells prevents TCRinduced glucose uptake and glycolysis, and activated c-Myc-nullT cells completely fail to grow or proliferate (Trumpp et al., 2001;Iritani et al., 2002; Dose et al., 2006; Wang et al., 2011). Is PI3Kand Akt signaling also required for the increase in glucose uptakeand glycolysis in TCR activated T cells in the periphery? Cer-tainly, antigen receptor induced c-Myc expression and glucoseuptake have been attributed to PI3K signaling (Frauwirth et al.,2002; Grumont et al., 2002; Doughty et al., 2006; Jacobs et al.,2008; Wang et al., 2011). However, one criticism of these stud-ies is that they rely on experiments involving the overexpressionof Akt and the use of the PI3K inhibitor LY294002. Overexpres-sion studies can be difficult to interrupt and while LY294002 wasinitially believed to be a highly specific PI3K inhibitor, and as suchwas used in good faith, it has since emerged that this inhibitor israther non-specific. LY294002 potently inhibits a number kinasesother than PI3K, including those with described roles in regulat-ing T cell growth and proliferation, i.e., mTORC1 and Pim familykinases (Brunn et al., 1996; Davies et al., 2000; Fox et al., 2005b;Bain et al., 2007). The importance of these other LY204002 tar-gets for T cell metabolism can be appreciated by a comparison
of the cellular sizes of PI3K/Akt deficient CD8 cytotoxic T lym-phocytes (CTL) and wild-type CTL cultured in the presence ofLY294002. While PI3K/Akt deficient CTL are comparable in sizeto wild-type CTL, LY294002-treated CTL are substantially smaller(Cornish et al., 2006; Sinclair et al., 2008; Macintyre et al., 2011).Therefore, the question as to whether PI3K regulates TCR inducedc-Myc expression has not been satisfactorily investigated to date.Nevertheless, a comparison of PI3K/Akt and c-Myc deficient Tcells is extremely informative. In contrast to the failure of c-Myc-null T cells to engage in TCR stimulated growth and proliferation,PI3K deficient T cells show a relatively mild defect in growth andproliferation with activated T cells capable of completing numer-ous divisions, though at a reduced rate (Okkenhaug et al., 2002,2006). Furthermore, T cells expressing a PDK1 K465E mutantwhich have defective TCR stimulated Akt activity, undergo normalTCR induced growth and proliferation (Waugh et al., 2009). Theseobservations coupled to the fact that T cells activated in limitingconcentrations of glucose do show a marked defect in prolifera-tion argue that TCR induced c-Myc expression, glucose uptake andglycolysis is not compromised by disruption of PI3K and Akt sig-naling (Jacobs et al., 2008). More recently, PI3K/Akt independentglucose uptake and glycolysis in TCR activated T cells has been con-firmed using pharmacological inhibitors of PI3Kδ (IC87114) andAkt (Akti1/2) with substantially greater selectivity than LY294002(Macintyre et al., 2011). Akti1/2 is particularly selective toward Aktdue to its unique allosteric mechanism of inhibition, binding tothe PH domain of Akt and preventing the PH domain-PI(3,4,5)P3
interaction and the resultant conformational change that is a pre-requisite for Akt activation (Zhao et al., 2005; Bain et al., 2007).Both IC87114 and Akti1/2 prevent PI3K/Akt signaling in T cellswhile having no effect on TCR induced glucose uptake (Macintyreet al., 2011). Therefore, it seems clear that PI3K has differentialroles in regulating glucose metabolism in developing thymocytesin the thymus and mature T cells in the periphery.
Once activated, T cells differentiate into various different effec-tor T cell subsets depending on the local environment and cytokineavailability. Many of these effector T cell subsets maintain an ele-vated glycolytic rate in response to cytokine signaling (Macintyreet al., 2011; Shi et al., 2011). For example, activated CD8 T cellsundergo rapid growth and proliferation in response to interleukin2 (IL2) as they differentiate into functional CTL. In response toIL2 signaling CTL maintain high levels of glucose uptake and lac-tate production indicative of elevated glycolysis (Macintyre et al.,2011). IL2 also promotes glucose uptake and glycolysis indepen-dently of PI3K and Akt but a key role has been revealed for PDK1.Thus, while IC87114 and Akti1/2 have no effect on CTL glucoseuptake, a pronounced decrease is observed following the deletionof PDK1 using a Cre/loxP strategy (Macintyre et al., 2011). WhilePDK1 is responsible for the activation of Akt, it also activates anumber of other members of the AGC kinase family includingprotein kinase C (PKC), 70-kDa ribosomal S6 kinase (p70S6K),90-kDa ribosomal S6 kinase (p90RSK), and serum/glucocorticoidregulated kinase (SGK; Pearce et al., 2010). However, unlike theactivation of Akt, PDK1 mediated activation of these other AGCfamily members are independent of PI3K signaling (Figure 1C).As members of this kinase family have overlapping substratespecificity it is likely that PDK1 dependent, Akt independent
Frontiers in Immunology | T Cell Biology August 2012 | Volume 3 | Article 247 | 4

“fimmu-03-00247” — 2012/8/6 — 11:02 — page 5 — #5
Finlay Glucose Metabolism in T cells
regulation of glucose metabolism reflects functional redundancywithin the AGC family of protein kinases (Brunet et al., 2001;Zhang et al., 2006; Sapkota et al., 2007).
mTORC1 has described roles in regulating various aspects ofcellular metabolism and given that its activity is independent ofPI3K and Akt in some activated T cell subsets, it remains likely thatmTORC1 is involved in maintaining glucose uptake and glycoly-sis (unpublished data; Duvel et al., 2010; Macintyre et al., 2011).Indeed, inhibition of mTORC1 decreases glycolysis in T cells acti-vated under Th17 polarizing conditions (Shi et al., 2011). However,as mTORC1 inhibition also disrupts Th17 differentiation, it is dif-ficult to interpret whether the effect of rapamycin on glycolysisis direct or as a result of the differentiation of different T cells.Therefore, a role for mTORC1 in controlling glycolysis in T cellshas still to be formally demonstrated.
T CELL MIGRATION AND METABOLISMThe expression of adhesion molecules and chemokine receptorsorchestrate the peripheral trafficking of activated T cells. Thep110γ catalytic subunit is the major PI3K isoform in T cells thatpromotes PI(3,4,5)P3 signaling in response to chemokines. Thus,migration to a range of chemokines is deficient in p110γ−/− T cellsand these T cells fail to traffic normally to sites of inflammation(Reif et al., 2004; Smith et al., 2007; Martin et al., 2008; Thomaset al., 2008). T cell migration and motility are energy demandingprocesses and it is tempting to speculate that chemokine recep-tor signaling might promote T cell glucose metabolism to meetthese energy demands. However, while there is some tentativedata linking chemokine receptor signaling to cell growth andmetabolism in transformed T cells and developing thymocytesthe relationship between chemokine receptor signaling and T cellmetabolism has not been directly studied (Janas et al., 2010; Loet al., 2010). Nonetheless, it is worth mentioning that factors thatinfluence T cell migration and/or homing, and thus the peripheraltissue destination of T cells, will affect T cell metabolism, albeitindirectly, by determining the cytokine environment to which
they are exposed. A comparison of activated T cells respondingto related cytokines IL2 and IL15 illustrates the differential reg-ulation of T cell metabolism by distinct cytokine environments.IL2 promotes elevated glucose metabolism and glycolysis whileIL15 does not maintain this metabolic state and T cells respondingto IL15 are smaller with reduced nutrient uptake and glycolysis(Cornish et al., 2006; Macintyre et al., 2011; unpublished data).While PI3Kγ controls T cell migration in response to chemokines,PI3Kδ regulates the repertoire of adhesion and chemokine recep-tors expressed by activated T cells. PI3Kδ, signaling through Aktand the Foxo transcription factors, regulates the expression ofkey molecules required for T cell homing between the blood andthe lymphoid organs; the adhesion molecule CD62L (also calledL-selectin) and the chemokine receptors CC-chemokine receptor7 (CCR7) and sphingosine-1-phosphate receptor 1 (S1P1). Dis-ruption of PI3Kδ/Akt signaling in activated CD8 T cells preventsthe down-regulation of CD62L, CCR7, and S1P1 and these T cellsretain a lymph node trafficking pattern rather than migratingto non-lymphoid tissues and the sites of inflammation (Sinclairet al., 2008; Waugh et al., 2009; Finlay and Cantrell, 2010; Mac-intyre et al., 2011). Thus, PI3Kδ deficient T cells activated invivo or wild-type T cells activated in mice treated with a PI3Kδ
inhibitor fail to traffic into the periphery to antigenic sites (Jarminet al., 2008). Therefore, both PI3Kγ and δ isoforms coordinatelyregulate T cell peripheral tissue homing thereby dictating thecytokine environments encountered and indirectly impactingupon T cell metabolism.
FINAL REMARKIt has recently become clear that the PI3K/Akt signaling axis is notthe important regulator of glucose uptake and glycolysis in matureT cells, as initially described. However, disrupting PI3K may in factimpact upon T cell metabolism through indirect mechanisms, i.e.,through altering their in vivo trafficking pattern, which will dictatethe cytokines these T cells encounter.
REFERENCESAlessi, D. R., and Cohen, P. (1998).
Mechanism of activation and func-tion of protein kinase B. Curr. Opin.Genet. Dev. 8, 55–62.
Bain, J., Plater, L., Elliott, M., Shpiro,N., Hastie, C. J., McLauchlan, H.,Klevernic, I., Arthur, J. S., Alessi, D.R., and Cohen, P. (2007). The selec-tivity of protein kinase inhibitors:a further update. Biochem. J. 408,297–315.
Bayascas, J. R., Wullschleger, S.,Sakamoto, K., Garcia-Martinez, J. M.,Clacher, C., Komander, D., van Aal-ten, D. M. F., Boini, K. M., Lang,F., Lipina, C., Logie, L., Sutherland,C., Chudek, J. A., van Diepen, J.A., Voshol, P. J., Lucocq, J. M., andAlessi, D. R. (2008). Mutation of thePDK1 PH domain inhibits proteinkinase B/Akt, leading to small size andinsulin resistance. Mol. Cell. Biol. 28,3258–3272.
Brand, K., Williams, J. F., andWeidemann, M. J. (1984). Glu-cose and glutamine metabolism inrat thymocytes. Biochem. J. 221,471–475.
Brunet, A., Park, J., Tran, H., Hu, L. S.,Hemmings, B. A., and Greenberg, M.E. (2001). Protein kinase SGK medi-ates survival signals by phosphorylat-ing the forkhead transcription factorFKHRL1 (FOXO3a). Mol. Cell. Biol.21, 952–965.
Brunn, G. J., Williams, J., Sabers,C., Wiederrecht, G., Lawrence, J.C., and Abraham, R. T. (1996).Direct inhibition of the signalingfunctions of the mammalian targetof rapamycin by the phosphoinosi-tide 3-kinase inhibitors, wortmanninand LY294002. EMBO J. 15, 5256—5267.
Burgering, B. M. T. (2008). A briefintroduction to FOXOlogy. Oncogene27, 2258–2262.
Calleja, V., Alcor, D., Laguerre, M., Park,J., Vojnovic, B., Hemmings, B. A.,Downward, J., Parker, P. J., and Lar-ijani, B. (2007). Intramolecular andintermolecular interactions of pro-tein kinase B define its activationin vivo. PLoS Biol. 5, e95. doi:10.1371/journal.pbio.0050095
Cameron, A. J., De Rycker, M., Calleja,V., Alcor, D., Kjaer, S., Kostelecky,B., Saurin, A., Faisal, A., Laguerre,M., Hemmings, B. A., McDonald,N., Larijani, B., and Parker, P. J.(2007). Protein kinases, from B to C.Biochem. Soc. Trans. 35, 1013–1017.
Cham, C. M., Driessens, G., O’Keefe,J. P., and Gajewski, T. F. (2008). Glu-cose deprivation inhibits multiple keygene expression events and effectorfunctions in CD8 + T cells. Eur. J.Immunol. 38, 2438–2450.
Ciofani, M., and Zuniga-Pflucker, J.C. (2005). Notch promotes survivalof pre-T cells at the beta-selection
checkpoint by regulating cellularmetabolism. Nat. Immunol. 6, 881–888.
Coffer, P. J., and Burgering, B. M.(2004). Forkhead-box transcriptionfactors and their role in the immunesystem. Nat. Rev. Immunol. 4,889–899.
Cornish, G. H., Sinclair, L. V., andCantrell, D. A. (2006). Differ-ential regulation of T-cell growthby IL-2 and IL-15. Blood 108,600–608.
Costello, P. S., Gallagher, M., andCantrell, D. A. (2002). Sustained anddynamic inositol lipid metabolisminside and outside the immunologi-cal synapse. Nat. Immunol. 3, 1082–1089.
Davies, S. P., Reddy, H., Caivano, M.,and Cohen, P. (2000). Specificity andmechanism of action of some com-monly used protein kinase inhibitors.Biochem. J. 351, 95–105.
www.frontiersin.org August 2012 | Volume 3 | Article 247 | 5

“fimmu-03-00247” — 2012/8/6 — 11:02 — page 6 — #6
Finlay Glucose Metabolism in T cells
Delgoffe, G. M., and Powell, J.D. (2009). mTOR: taking cuesfrom the immune microenviron-ment. Immunology 127, 459–465.
Dose, M., Khan, I., Guo, Z., Koval-ovsky, D., Krueger, A., von Boehmer,H., Khazaie, K., and Gounari, F.(2006). c-Myc mediates pre-TCR-induced proliferation but not devel-opmental progression. Blood 108,2669–2677.
Doughty, C. A., Bleiman, B. F., Wag-ner, D. J., Dufort, F. J., Mataraza, J.M., Roberts, M. F., and Chiles, T. C.(2006). Antigen receptor-mediatedchanges in glucose metabolism in Blymphocytes: role of phosphatidyli-nositol 3-kinase signaling in the gly-colytic control of growth. Blood 107,4458–4465.
Duvel, K., Yecies, J. L., Menon, S.,Raman, P., Lipovsky, A. I., Souza, A.L., Triantafellow, E., Ma, Q., Gorski,R., Cleaver, S., Vander Heiden, M. G.,MacKeigan, J. P., Finan, P. M., Clish,C. B., Murphy, L. O., and Manning, B.D. (2010). Activation of a metabolicgene regulatory network downstreamof mTOR complex 1. Mol. Cell 39,171–183.
Fayard, E., Gill, J., Paolino, M., Hynx,D., Hollander, G. A., and Hem-mings, B. A. (2007). Deletion ofPKBalpha/Akt1 affects thymic devel-opment. PLoS ONE 2, e992. doi:10.1371/journal.pone.0000992
Finlay, D., and Cantrell, D. (2010).Phosphoinositide 3-kinase and themammalian target of rapamycinpathways control T cell migra-tion. Ann. N. Y. Acad. Sci. 1183,149–157.
Finlay, D. K., Kelly, A. P., Clarke, R., Sin-clair, L. V., Deak, M., Alessi, D. R.,and Cantrell, D. A. (2010). Tempo-ral differences in the dependency onphosphoinositide-dependent kinase1 distinguish the development ofinvariant Valpha14 NKT cells andconventional T cells. J. Immunol. 185,5973–5982.
Finlay, D. K., Sinclair, L. V., Feijoo,C., Waugh, C. M., Hagenbeek, T. J.,Spits, H., and Cantrell, D. A. (2009).Phosphoinositide-dependent kinase1 controls migration and malignanttransformation but not cell growthand proliferation in PTEN-null lym-phocytes. J. Exp. Med. 206, 2441–2454.
Fox, C. J., Hammerman, P. S., andThompson, C. B. (2005a). Fuel feedsfunction: energy metabolism and theT-cell response. Nat. Rev. Immunol.5, 844–852.
Fox, C. J., Hammerman, P. S., andThompson, C. B. (2005b). The Pimkinases control rapamycin-resistant
T cell survival and activation. J. Exp.Med. 201, 259–266.
Frauwirth, K. A., Riley, J. L., Harris,M. H., Parry, R. V., Rathmell, J. C.,Plas, D. R., Elstrom, R. L., June, C.H., and Thompson, C. B. (2002). TheCD28 signaling pathway regulatesglucose metabolism. Immunity 16,769–777.
Garcon, F., Patton, D. T., Emery, J. L.,Hirsch, E., Rottapel, R., Sasaki, T.,and Okkenhaug, K. (2007). CD28provides T-cell costimulation andenhances PI3K activity at the immunesynapse independently of its capac-ity to interact with the p85/p110heterodimer. Blood 111, 1464–1471.
Greiner, E. F., Guppy, M., and Brand, K.(1994). Glucose is essential for pro-liferation and the glycolytic enzymeinduction that provokes a transitionto glycolytic energy production. J.Biol. Chem. 269, 31484–31490.
Grumont, R. J., Strasser, A., andGerondakis, S. (2002). B cell growthis controlled by phosphatidylinoso-tol 3-kinase-dependent inductionof Rel/NF-kappaB regulated c-myctranscription. Mol. Cell 10, 1283–1294.
Hagenbeek, T. J., and Spits, H. (2008).T-cell lymphomas in T-cell-specificPten-deficient mice originate in thethymus. Leukemia 22, 608–619.
Hanada, M., Feng, J., and Hemmings, B.A. (2004). Structure, regulation andfunction of PKB/AKT – a major ther-apeutic target. Biochim. Biophys. Acta1697, 3–16.
Harriague, J., and Bismuth, G. (2002).Imaging antigen-induced PI3K acti-vation in T cells. Nat. Immunol. 3,1090–1096.
Hinton, H. J., Alessi, D. R., and Cantrell,D. A. (2004). The serine kinasephosphoinositide-dependent kinase1 (PDK1) regulates T cell develop-ment. Nat. Immunol. 5, 539–545.
Huppa, J. B., Gleimer, M., Sumen, C.,and Davis, M. M. (2003). ContinuousT cell receptor signaling required forsynapse maintenance and full effectorpotential. Nat. Immunol. 4, 749–755.
Iritani, B. M., Delrow, J., Grandori, C.,Gomez, I., Klacking, M., Carlos, L. S.,and Eisenman, R. N. (2002). Modu-lation of T-lymphocyte development,growth and cell size by the Myc antag-onist and transcriptional repressorMad1. EMBO J. 21, 4820–4830.
Jacobs, S. R., Herman, C. E., Maciver,N. J., Wofford, J. A., Wieman, H.L., Hammen, J. J., and Rathmell, J.C. (2008). Glucose uptake is limit-ing in T cell activation and requiresCD28-mediated Akt-dependent andindependent pathways. J. Immunol.180, 4476–4486.
Janas, M. L., Varano, G., Gudmunds-son, K., Noda, M., Nagasawa, T., andTurner, M. (2010). Thymic develop-ment beyond beta-selection requiresphosphatidylinositol 3-kinase activa-tion by CXCR4. J. Exp. Med. 207,247–261.
Jarmin, S. J., David, R., Ma, L., Chai,J. G., Dewchand, H., Takesono, A.,Ridley, A. J., Okkenhaug, K., andMarelli-Berg, F. M. (2008). T cellreceptor-induced phosphoinositide-3-kinase p110delta activity isrequired for T cell localization toantigenic tissue in mice. J. Clin.Invest. 118, 1154–1164.
Ji, H., Rintelen, F., Waltzinger, C.,Bertschy Meier, D., Bilancio, A.,Pearce, W., Hirsch, E., Wymann, M.P., Ruckle, T., Camps, M., Vanhae-sebroeck, B., Okkenhaug, K., andRommel, C. (2007). Inactivation ofPI3Kgamma and PI3Kdelta distortsT-cell development and causes mul-tiple organ inflammation. Blood 110,2940–2947.
Juntilla, M. M., Wofford, J. A., Birn-baum, M. J., Rathmell, J. C., andKoretzky, G. A. (2007). Akt1 andAkt2 are required for alphabeta thy-mocyte survival and differentiation.Proc. Natl. Acad. Sci. U.S.A. 104,12105–12110.
Kelly, A. P., Finlay, D. K., Hinton, H.J., Clarke, R. G., Fiorini, E., Radtke,F., and Cantrell, D. A. (2007). Notch-induced T cell development requiresphosphoinositide-dependent kinase1. EMBO J. 26, 3441–3450.
Koyasu, S. (2003). The role of PI3Kin immune cells. Nat. Immunol. 4,313–319.
Laplante, M., and Sabatini, D. M.(2009). mTOR signaling at a glance.J. Cell. Sci. 122, 3589–3594.
Lo, B. K., Yu, M., Zloty, D., Cowan,B., Shapiro, J., and McElwee, K. J.(2010). CXCR3/ligands are signifi-cantly involved in the tumorigene-sis of basal cell carcinomas. Am. J.Pathol. 176, 2435–2446.
Macintyre, A. N., Finlay, D., Preston, G.,Sinclair, L. V., Waugh, C. M., Tamas,P., Feijoo, C., Okkenhaug, K., andCantrell, D. A. (2011). Protein kinaseB controls transcriptional programsthat direct cytotoxic T cell fate butis dispensable for T cell metabolism.Immunity 34, 224–236.
Manning, B. D., and Cantley, L. C.(2007). AKT/PKB signaling: navi-gating downstream. Cell 129, 1261–1274.
Mao, C., Tili, E. G., Dose, M., Haks,M. C., Bear, S. E., Maroulakou, I.,Horie, K., Gaitanaris, G. A., Fidanza,V., Ludwig, T., Wiest, D. L., Gounari,F., and Tsichlis, P. N. (2007). Unequal
contribution of Akt isoforms in thedouble-negative to double-positivethymocyte transition. J. Immunol.178, 5443–5453.
Marko, A. J., Miller, R. A., Kelman, A.,and Frauwirth, K. A. (2010). Induc-tion of glucose metabolism in stim-ulated T lymphocytes is regulated bymitogen-activated protein kinase sig-naling. PLoS ONE 5, e15425. doi:10.1371/journal.pone.0015425
Martin, A. L., Schwartz, M. D., Jame-son, S. C., and Shimizu, Y. (2008).Selective regulation of CD8 effectorT cell migration by the p110 gammaisoform of phosphatidylinositol 3-kinase. J. Immunol. 180, 2081–2088.
Okkenhaug, K., Bilancio, A., Far-jot, G., Priddle, H., Sancho, S.,Peskett, E., Pearce, W., Meek, S.E., Salpekar, A., Waterfield, M. D.,Smith, A. J., and Vanhaesebroeck, B.(2002). Impaired B and T cell anti-gen receptor signaling in p110deltaPI 3-kinase mutant mice. Science 297,1031–1034.
Okkenhaug, K., Patton, D. T., Bilan-cio, A., Garcon, F., Rowan, W. C.,and Vanhaesebroeck, B. (2006). Thep110delta isoform of phosphoinosi-tide 3-kinase controls clonal expan-sion and differentiation of Th cells. J.Immunol. 177, 5122–5128.
Okkenhaug, K., and Vanhaesebroeck, B.(2003). PI3K in lymphocyte develop-ment, differentiation and activation.Nat. Rev. Immunol. 3, 317–330.
Pearce, L. R., Komander, D., and Alessi,D. R. (2010). The nuts and bolts ofAGC protein kinases. Nat. Rev. Mol.Cell Biol. 11, 9–22.
Reif, K., Okkenhaug, K., Sasaki, T., Pen-ninger, J. M., Vanhaesebroeck, B.,and Cyster, J. G. (2004). Cuttingedge: differential roles for phospho-inositide 3-kinases, p110gamma andp110delta, in lymphocyte chemo-taxis and homing. J. Immunol. 173,2236–2240.
Sapkota, G. P., Cummings, L., Newell, F.S., Armstrong, C., Bain, J., Frodin,M., Grauert, M., Hoffmann, M.,Schnapp, G., Steegmaier, M., Cohen,P., and Alessi, D. R. (2007). BI-D1870is a specific inhibitor of the p90 RSK(ribosomal S6 kinase) isoforms invitro and in vivo. Biochem. J. 401,29–38.
Sasaki, T., Irie-Sasaki, J., Jones, R. G.,Oliveira-dos-Santos, A. J., Stanford,W. L., Bolon, B., Wakeham, A., Itie, A.,Bouchard, D., Kozieradzki, I., Joza,N., Mak, T. W., Ohashi, P. S., Suzuki,A., and Penninger, J. M. (2000). Func-tion of PI3Kgamma in thymocytedevelopment, T cell activation, andneutrophil migration. Science 287,1040–1046.
Frontiers in Immunology | T Cell Biology August 2012 | Volume 3 | Article 247 | 6

“fimmu-03-00247” — 2012/8/6 — 11:02 — page 7 — #7
Finlay Glucose Metabolism in T cells
Shi, L. Z., Wang, R., Huang, G.,Vogel, P., Neale, G., Green, D. R.,and Chi, H. (2011). HIF1alpha-dependent glycolytic pathway orches-trates a metabolic checkpoint forthe differentiation of TH17 andTreg cells. J. Exp. Med. 208,1367–1376.
Sinclair, L. V., Finlay, D., Feijoo,C., Cornish, G. H., Gray, A.,Ager, A., Okkenhaug, K., Hagen-beek, T. J., Spits, H., and Cantrell,D. A. (2008). Phosphatidylinositol-3-OH kinase and nutrient-sensingmTOR pathways control T lympho-cyte trafficking. Nat. Immunol. 9,513–521.
Smith, L. D., Hickman, E. S., Parry,R. V., Westwick, J., and Ward, S.G. (2007). PI3Kgamma is the dom-inant isoform involved in migra-tory responses of human T lym-phocytes: effects of ex vivo mainte-nance and limitations of non-viraldelivery of siRNA. Cell. Signal. 19,2528–2539.
Suzuki, A., Yamaguchi, M. T., Ohteki,T., Sasaki, T., Kaisho, T., Kimura, Y.,Yoshida, R., Wakeham, A., Higuchi,T., Fukumoto, M., Tsubata, T.,Ohashi, P. S., Koyasu, S., Penninger,J. M., Nakano, T., and Mak, T.W. (2001). T cell-specific loss ofPten leads to defects in central andperipheral tolerance. Immunity 14,523–534.
Swat, W., Montgrain, V., Doggett, T. A.,Douangpanya, J., Puri, K., Vermi, W.,and Diacovo, T. G. (2006). Essentialrole of PI3Kdelta and PI3Kgammain thymocyte survival. Blood 107,2415–2422.
Thomas, M. S., Mitchell, J. S., DeNucci,C. C., Martin, A. L., and Shimizu,Y. (2008). The p110gamma isoformof phosphatidylinositol 3-kinase reg-ulates migration of effector CD4 Tlymphocytes into peripheral inflam-matory sites. J. Leukoc. Biol. 84,814–823.
Trumpp, A., Refaeli, Y., Oskarsson, T.,Gasser, S., Murphy, M., Martin, G.R., and Bishop, J. M. (2001). c-Mycregulates mammalian body size bycontrolling cell number but not cellsize. Nature 414, 768–773.
Vander Heiden, M. G., Cantley, L. C.,and Thompson, C. B. (2009). Under-standing the Warburg effect: themetabolic requirements of cell pro-liferation. Science 324, 1029–1033.
Vanhaesebroeck, B., Guillermet-Guibert, J., Graupera, M., andBilanges, B. (2010). The emergingmechanisms of isoform-specificPI3K signalling. Nat. Rev. Mol. CellBiol. 11, 329–341.
Wang, R., Dillon, C. P., Shi, L.Z., Milasta, S., Carter, R., Finkel-stein, D., McCormick, L. L., Fitzger-ald, P., Chi, H., Munger, J.,and Green, D. R. (2011). The
transcription factor Myc controlsmetabolic reprogramming upon Tlymphocyte activation. Immunity 35,871–882.
Waugh, C., Sinclair, L., Finlay, D.,Bayascas, J. R., and Cantrell,D. (2009). Phosphoinositide(3,4,5)-triphosphate binding tophosphoinositide-dependent kinase1 regulates a protein kinase B/Aktsignaling threshold that dictatesT-cell migration, not proliferation.Mol. Cell. Biol. 29, 5952–5962.
Webb, L. M., Vigorito, E., Wymann,M. P., Hirsch, E., and Turner, M.(2005). Cutting edge: T cell develop-ment requires the combined activitiesof the p110gamma and p110deltacatalytic isoforms of phosphatidyli-nositol 3-kinase. J. Immunol. 175,2783–2787.
Wofford, J. A., Wieman, H. L., Jacobs,S. R., Zhao, Y., and Rathmell, J. C.(2008). IL-7 promotes Glut1 traffick-ing and glucose uptake via STAT5-mediated activation of Akt to supportT-cell survival. Blood 111, 2101–2111.
Zhang, H. H., Lipovsky, A. I., Dib-ble, C. C., Sahin, M., and Man-ning, B. D. (2006). S6K1 regulatesGSK3 under conditions of mTOR-dependent feedback inhibition ofAkt. Mol. Cell 24, 185–197.
Zhao, Z., Leister, W. H., Robinson, R.G., Barnett, S. F., Defeo-Jones, D.,
Jones, R. E., Hartman, G. D., Huff, J.R., Huber, H. E., Duggan, M. E., andLindsley, C. W. (2005). Discovery of2,3,5-trisubstituted pyridine deriva-tives as potent Akt1 and Akt2 dualinhibitors. Bioorg. Med. Chem. Lett.15, 905–909.
Conflict of Interest Statement: Theauthor declares that the research wasconducted in the absence of any com-mercial or financial relationships thatcould be construed as a potential con-flict of interest.
Received: 01 June 2012; paper pendingpublished: 13 June 2012; accepted: 24July 2012; published online: 07 August2012.Citation: Finlay DK (2012) Regulationof glucose metabolism in T cells: newinsight into the role of phosphoinositide3-kinases. Front. Immun. 3:247. doi:10.3389/fimmu.2012.00247This article was submitted to Frontiers inT Cell Biology, a specialty of Frontiers inImmunology.Copyright © 2012 Finlay. This is an open-access article distributed under the termsof the Creative Commons AttributionLicense, which permits use, distributionand reproduction in other forums, pro-vided the original authors and sourceare credited and subject to any copy-right notices concerning any third-partygraphics etc.
www.frontiersin.org August 2012 | Volume 3 | Article 247 | 7




















