
IQGAP Proteins Reveal an Atypical Phosphoinositide (aPI) Binding Domain with a Pseudo C2 Domain Fold
Upload
retroscreenCategory
view
2download
0
ACI International, 12/3 (2000)
Research Trends126
© 2000 Hogrefe & Huber Publishers
The Role of Bruton’sTyrosine Kinase (Btk) inPhosphoinositide-Dependent Signaling
by Beston F. Nore, Abdalla J. Mohamed, Leonardo Vargas, Lars J. Brandén, Carl-Magnus Bäckesjö, Mauno Vihinen,
Birger Christensson, & C.I. Edvard Smith
Mutations in the cytoplasmic tyrosine kinase, Bruton’s tyro-sine kinase (Btk), were identified in humans diagnosed as X-linked agammaglobulinemia (XLA) and as X-linkedimmunodeficiency (Xid) in mice. The functional alteration ofthe Btk mutation results in a block in B-cell development anddefects in the cellular signaling. Btk is a member of the Tecfamily of protein tyrosine kinases (Tec kinases) whichincludes Itk, Tec, Bmx, Txk, Drosophila Tec (dTec29), Rajaeglanteria Tec (sTec), and zebrafish Tec (zTec). Apart fromTxk and dTec29, these kinases possess an N-terminal pleck-strin homology domain (PH) which is essential for membranetargeting and also for biological activity. The PH domainsare capable of binding phosphoinositides in a specific man-ner and function mainly as membrane localization signals.In vitro, Btk binds strongly to PIP3 and IP4. Moreover,lipid specificity for other Tec kinase members is not known.We propose a model in which PH domains of Tec familykinases exhibit a switching mechanism regulating activationand deactivation states (ON–OFF states). This process isdetermined by exchange of lipid specificity where themembrane anchoring signal is an activation state of thatmechanism.
B-cell development and differentiation elicited by B-cellantigen receptor (BcR) engagement is driven by a complex net-work of signals (see review [1]). The BcR is composed of mem-brane immunoglobulins (mIg) noncovalently associated with themembrane proteins Igα and Igβ (CD79a and b). The mIg bindsantigen whereas Igα and Igβ each contain a single immunore-ceptor tyrosine-based activation motif (ITAM) in their cytoplas-mic domains and are responsible for intracellular signaling (fora review see [2]). Stimulation of the BcR results in the elevationof protein tyrosine phosphorylation [3]. These initial signalingevents triggered by BcR cross-linking involve the activation of
several nonreceptor tyrosine kinase families, including Src-fam-ily kinases (Src kinases), Tec family kinases, and Jak/Syk pro-tein kinases [4].
The members of the Src-family kinases phosphorylate twotyrosines in a conserved sequence of the ITAM motif (in single-letter amino acid code, D/Ex7D/Ex2Yx2L/Ix7Yx2L/I, where x isany amino acid) in the cytoplasmic domain of Igα and Igβ. Thedoubly phosphorylated ITAM recruits Syk, which is subse-quently activated by Src family kinase-mediated transphospho-rylation, leading to the activation of Syk [4]. This sequential ac-tivation mechanism has been proposed by Weiss and Littman[5]. In addition to Syk activation, the Tec family kinases are alsoactivated by Src family kinases, as indicated for Btk, but Btk isnot associated with known antigen receptor complex compo-nents [6, 7].
The aim of this review is to discuss recent advances of thesignal transduction pathways of Tec kinases and involvement ofphosphoinositide lipids in the activation event(s).
Tec Family Kinases
Members
Tec kinases belong to a subfamily of Src-like kinases, ofwhich 5 members have been identified in mammalians: Tec [8],Btk [9, 10], Itk [11, 12], Bmx [13, 14], and Txk [15]; three re-lated members have been identified in Drosophila (dTec29)[16], zebrafish kidney (zTec) [17], and Raja elganteria spleen(clearnose skate, sTec) [18].

ACI International, 12/3 (2000)
Research Trends 127
Expression
Each Tec kinase has its distinct expression pattern. Itk is ex-pressed in T cells and mast cells, whereas Btk is expressed inmost hematopoietic cells other than T lymphocytes. The pres-ence of Txk is restricted to mast and T cells, whereas Tec ismore widely expressed and not restricted to hematopoietic cells[19]. The expression of Bmx is detected in myeloid cells, thegranulomonocytic lineage within the hematopoietic system, andcertain endothelial cells [13, 14, 20].
Domains
The Tec kinases, excluding Txk and dTec29, contain thesame subdomain architecture: pleckstrin domain (PH) at theamino terminus, followed by the BTK motif and a proline-rich(PR) region that together have been termed as the “Tec homolo-gy” (TH) domain [21], and three Src homology domains, SH3,SH2, and kinase (SH1) in tandem (see Figure 1) [22].
Sequence Homologies of Tec Kinases
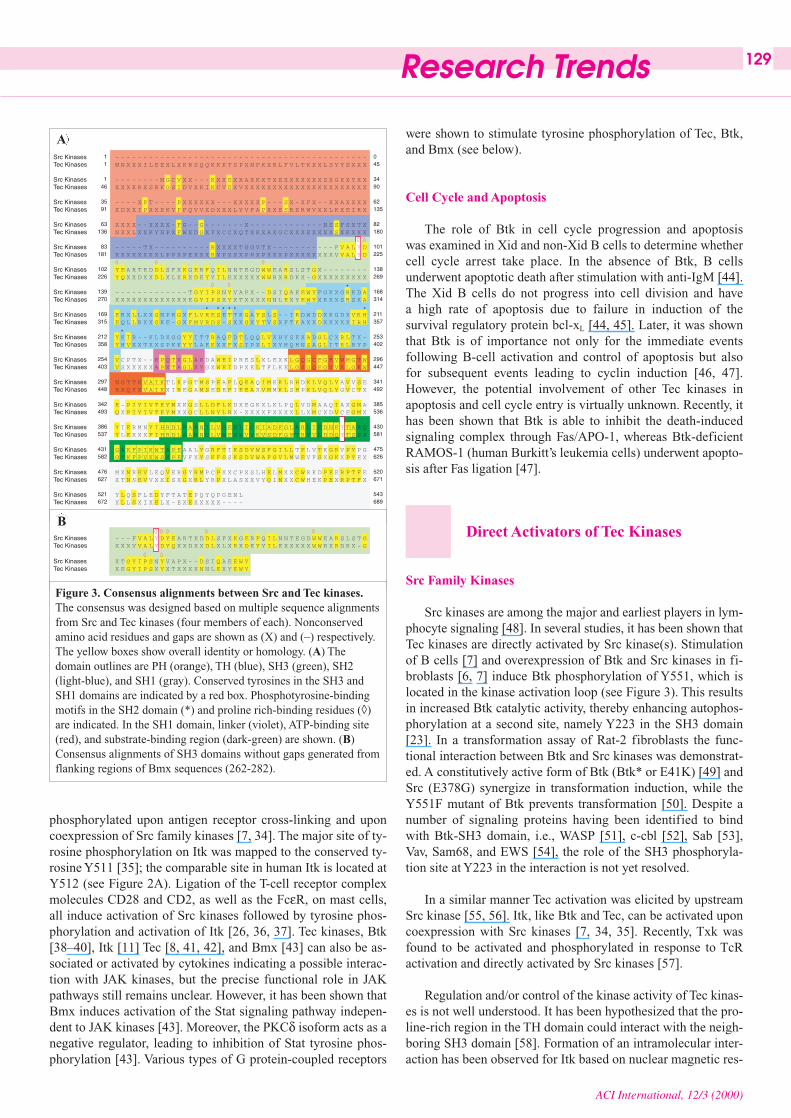
Sequence alignments between all Tec kinases predicted thelocation of conserved regions in the kinase domain, including acommon Btk phosphorylation site found in all cytoplasmic pro-tein tyrosine kinases (PTKs) (tyrosine (Y) 551 in Btk) whichsubsequently was found to be active [6, 7]. Corresponding sitesare located at hBmx (Y566), mBmx (Y542), hItk (Y512), mItk(Y511), hTec (Y519), mTec (Y518), m/hTxk (Y420), sTec(Y514), zTec (Y509), and dTec29 (Y479) (see Figure 2A).Phosphorylation at this site increases the kinase activity and isimportant for mediation of BcR-signaling downstream of Btk[7]. The transphosphorylation at Y551 of activated Btk is fol-lowed by phosphorylation at Y223 in the SH3 domain [23]. Thetyrosine 223 position in Btk is also conserved in all SH3 do-mains of Tec kinases, excluding zTec, whose tyrosine is substi-tuted by an arginine (see Figure 2B). This conserved tyrosinesite is located at hBmx (Y194), mBmx (Y193), hItk (Y180),mItk (Y180), hTec (Y188), mTec (Y187), h/mTxk (Y91), sTec(Y187), zTec (Y175), and dTec29 (Y153).
Phylogenetic Trees
Homology and identity of primary sequences applied to de-termine phylogenetic relationship between all Tec kinases arepresented in Figure 2C. The phylogenetic tree shows the great-est identity between human and mouse genes of Btk, thereafterTec, Itk, Bmx, and Txk, respectively (see Figure 2C). On theother hand, this comparison shows that sTec is more closely re-lated to Btk, while zTec has closer relations to Itk. On the basisof amino acid sequence similarity and expression of sTec in Blymphocytes, one may speculate that sTec is essential for B-cellresponses in skate, being a counterpart of Btk [18]. The mostdistant members of the Tec kinases are Txk and dTec29 whichare characterized by the absence of PH domains, and the overallsequence similarity with other Tec family members is low. The
conserved tyrosines (phosphorylated sites in Btk) in the SH3domain and in the kinase domain are, however, also present inTxk and dTec29, (see Figure 2a). Moreover, the genomic orga-nization of Txk is related to Btk [15] and, furthermore, the N-terminal region of Txk may also have a membrane localizingfunction.
Consensus Alignments Between Src andTec Family Kinases
Further multiple sequence alignments of Src and Tec kinasesfrom both human and mouse were analyzed. Four Src kinases(c-Src, Yes, Fgr, and Fyn) and Tec kinases (Btk, Itk, Tec, andBmx) were selected as the corresponding sequences wereavailable from databases. Sequence alignments for eachgroup of kinases were performed separately using the samemethod as described in Figure 2. One consensus sequencewas deduced from multiple sequence alignments. Consensusdetermination was chosen based on 62.5% identity and/orhomology. In the case of Tec kinases, the flanking regions(sequence repeats in the SH3 domain [13, 14]) in Bmx werenot included into the analysis, because they do not sharesimilarity with other Tec kinase members. These two consensussequences were subjected to further multiple sequence align-ments (see Figure 3). As shown in Figure 3, the overall domainhomologies of SH3, SH2, and SH1 are conserved. Excludingthe PH domain, there are very few insertions and deletions inthe alignment. Gaps are located in the N-terminus, where thesetwo families of kinases are divergent. It is worth mentioningthat the Src kinases lack a PH domain, which is replaced by aunique region (SH4) with a membrane-anchoring function viamyristylation. Alignment between the PH domain of Tec kinasesand the unique region of the Src kinases shows only singlehomologies and/or identities spread out in the region which re-flect the alignment of unrelated sequences with massive gapinsertions (see Figure 3). As mentioned earlier, tyrosines in thekinase and SH3 domains are conserved. High conservation atpolyproline binding motifs in the SH3 domain, the phosphoty-rosine-binding motif in the SH2 domain, and the linker motif
pY pY223 551
PH BTK PR
THSH3 SH2 Kinase (SH1)
PIP3 Zn+2
PI3-kinase
PKCβ Ι/ΙΙ
G proteins
Src kinasesAutophosphorylation
PR Interaction
p120cblSabVavWASPSam68EWS
BAP-135PLCγGqα, Gα12, Gβγ
Figure 1. Schematic model of Btk showing different domains andinteracting molecules in both upstream and downstream signal-ing. PH = Pleckstrin homology domain; TH = Tec homology domain(Btk motif and prolin-rich (PR) region); SH3 = Src homology 3domain; SH2 = Src homology 2 domain; SH1 = kinase domain.

ACI International, 12/3 (2000)
Research Trends128
of the N-terminal end of the SH1 domain are observed, reflect-ing the importance of these elements for the protein kinasefunction.
Defects
The role of Tec kinases in signal transduction has been bestcharacterized for Btk in the context of the BcR signaling. Btkwas shown to be activated following BcR stimulation [23–26].The naturally occurring mutations in the Btk gene result in X-linked agammaglobulinemia (XLA) in humans [27], eithercaused by loss of Btk expression or severe functional changesin any domain or motifs (for a recent review see [28]). Asevere block in B-cell development at the pre-B stage is themost characteristic feature of this disease in humans, whereas
mutated PH domains (R28C) in X-linked immunodeficiency(Xid) as well as knockout mice exhibit a milder phenotype[29–31].
Signaling
Studies on Itk, the second best characterized Tec kinase afterBtk, have shown that Itk plays an important role in T-cell signal-ing. Itk knockout mice have slightly reduced numbers of maturethymocytes and show impeded T-cell receptor (TcR) signalingresponses [32]. This suggests a role for Itk in T-cell develop-ment and function. The reciprocal pattern of expression sug-gests that Btk and Itk play analogous roles in antigen receptorsignaling pathways in B and T cells, respectively, as is also thecase with ZAP-70 and Syk [33]. Itk, like Btk, becomes tyrosine
BTK-HUMAN 519 HRDLAARNCL VNDQGVVKVS DFGLSRYVLD DEYTSSVGSK FPVRW 563BTK-MOUSE 519 HRDLAARNCL VNDQGVVKVS DFGLSRYVLD DEYTSSVGSK FPVRW 563BMX-HUMAN 534 HRDLAARNCL VDRDLCVKVS DFGMTRYVLD DQYVSSVGTK FPVKW 578BMX-MOUSE 510 HRDLAARNCL VDSDLSVKVS DFGMTRYVLD DQYVSSVGTK FPVKW 554TEC-HUMAN 487 HRDLAARNCL VSEAGVVKVS DFGMARYFLD DQYTSSSGAK FPVKW 531TEC-MOUSE 486 HRDLAARNCL VNEAGVVKVS DFGMARYVLD DQYTSSSGAK FPVKW 530ITK-HUMAN 480 HRDLAARNCL VGENQVIKVS DFGMTRFVLD DQYTSSTGTK FPVKW 524ITK-MOUSE 479 HRDLAARNCL VGENQVIKVS DFGMTRFVLD DQYTSSTGTK FPVKW 523TEC-SKATE 482 HRDLAARNCL VDDAATVKVS DFGLSRYVLD DEYTSSQGSK FPVRW 526TEC-ZEBRA 477 HRDLAARNCL VSENSVVKIA DFGMTRFVLD DQYTSSHGSK FPVRW 521TXK-HUMAN 388 HRDLAARNCL VSSTCIVKIS DFGMTRYVLD DEYVSSFGAK FPIKW 432TXK-MOUSE 388 HRDLAARNCL VSSACVVKIS DFGMARYVLD DEYISSSGAK FPVKW 432TEC29-DRm 447 HRDLAARNCL VGSENVVKVA DFGLARYVLD DQYTSSGGTK FPIKW 491
Figure 2. Sequence alignments of Tec kinases showing conserved tyrosines, allocated asphosphorylated tyrosines Y551 (A) and Y223 (B) (in red) in Btk. Alignment of the activationloop in the SH1 (kinase) domain (A) and of the conserved tyrosine in the SH3 domain (B), wasperformed by using a Macintosh version of the ClustalX program [94]. The multiple sequencealignments were refined using SeqVu (The Garvan Institute of Medical Research, DalingurstNSW, 2010 Sydney, Australia). Segments of the sequence alignments were chosen and viewedusing FreeHand (MACROMEDIA). (C) Phylogenetic tree of all Tec kinases as determined bymaximum proximity analysis from multiple sequence alignments on ClustalX software. Themethod applied is adapted from PHYLIP calculations (Joe Felsenstein). The graphical outputadjustment of the tree was made by TreeMap (v1.0) for Macintosh (Roderic D.M., Oxford, UK).
BTK-HUMAN 217 KKVVALYDYM PMNANDLQLR K 237BTK-MOUSE 217 KKVVALYDYM PMNANDLQLR K 237BMX-HUMAN 188 STTLAQYDNE SKKNYGSQPP S 208BMX-MOUSE 187 GAILPQYDSY SKKSCGSQP- T 207TEC-HUMAN 182 EIVVAMYDFQ AAEGHDLRLE R 202TEC-MOUSE 181 EIVVAMYDFQ ATEAHDLRLE R 201ITK-HUMAN 174 TVVIALYDYQ TNDPQELALR R 194ITK-MOUSE 174 TLVIALYDYQ TNDPQELALR C 194TEC-SKATE 181 KVVVALYDYD IMNPDDLPLR K 201TEC-ZEBRA 169 RIVVALRNFI PKEHTDLPLQ K 189TXK-HUMAN 85 IQVKALYDFL PREPCNLALR R 105TXK-MOUSE 85 IQVKALYDFL PREPGNLALK R 105TEC29-DRm 147 KLVVALYPFK AIEGGDLSLE K 167
0.1
TEC29 DRm
TXK HUMAN
TXK MOUSE
TEC SKATE
BTK HUMAN
BTK MOUSE
BMX HUMAN
BMX MOUSE
TEC HUMAN
TEC MOUSE
TEC ZEBRA
ITK HUMAN
ITK MOUSE
A
B C
ACI International, 12/3 (2000)
Research Trends 129
phosphorylated upon antigen receptor cross-linking and uponcoexpression of Src family kinases [7, 34]. The major site of ty-rosine phosphorylation on Itk was mapped to the conserved ty-rosine Y511 [35]; the comparable site in human Itk is located atY512 (see Figure 2A). Ligation of the T-cell receptor complexmolecules CD28 and CD2, as well as the FcεR, on mast cells,all induce activation of Src kinases followed by tyrosine phos-phorylation and activation of Itk [26, 36, 37]. Tec kinases, Btk[38–40], Itk [11] Tec [8, 41, 42], and Bmx [43] can also be as-sociated or activated by cytokines indicating a possible interac-tion with JAK kinases, but the precise functional role in JAKpathways still remains unclear. However, it has been shown thatBmx induces activation of the Stat signaling pathway indepen-dent to JAK kinases [43]. Moreover, the PKCδ isoform acts as anegative regulator, leading to inhibition of Stat tyrosine phos-phorylation [43]. Various types of G protein-coupled receptors
were shown to stimulate tyrosine phosphorylation of Tec, Btk,and Bmx (see below).
Cell Cycle and Apoptosis
The role of Btk in cell cycle progression and apoptosiswas examined in Xid and non-Xid B cells to determine whethercell cycle arrest take place. In the absence of Btk, B cellsunderwent apoptotic death after stimulation with anti-IgM [44].The Xid B cells do not progress into cell division and havea high rate of apoptosis due to failure in induction of thesurvival regulatory protein bcl-xL [44, 45]. Later, it was shownthat Btk is of importance not only for the immediate eventsfollowing B-cell activation and control of apoptosis but alsofor subsequent events leading to cyclin induction [46, 47].However, the potential involvement of other Tec kinases inapoptosis and cell cycle entry is virtually unknown. Recently, ithas been shown that Btk is able to inhibit the death-inducedsignaling complex through Fas/APO-1, whereas Btk-deficientRAMOS-1 (human Burkitt’s leukemia cells) underwent apopto-sis after Fas ligation [47].
Direct Activators of Tec Kinases
Src Family Kinases
Src kinases are among the major and earliest players in lym-phocyte signaling [48]. In several studies, it has been shown thatTec kinases are directly activated by Src kinase(s). Stimulationof B cells [7] and overexpression of Btk and Src kinases in fi-broblasts [6, 7] induce Btk phosphorylation of Y551, which islocated in the kinase activation loop (see Figure 3). This resultsin increased Btk catalytic activity, thereby enhancing autophos-phorylation at a second site, namely Y223 in the SH3 domain[23]. In a transformation assay of Rat-2 fibroblasts the func-tional interaction between Btk and Src kinases was demonstrat-ed. A constitutively active form of Btk (Btk* or E41K) [49] andSrc (E378G) synergize in transformation induction, while theY551F mutant of Btk prevents transformation [50]. Despite anumber of signaling proteins having been identified to bindwith Btk-SH3 domain, i.e., WASP [51], c-cbl [52], Sab [53],Vav, Sam68, and EWS [54], the role of the SH3 phosphoryla-tion site at Y223 in the interaction is not yet resolved.
In a similar manner Tec activation was elicited by upstreamSrc kinase [55, 56]. Itk, like Btk and Tec, can be activated uponcoexpression with Src kinases [7, 34, 35]. Recently, Txk wasfound to be activated and phosphorylated in response to TcRactivation and directly activated by Src kinases [57].
Regulation and/or control of the kinase activity of Tec kinas-es is not well understood. It has been hypothesized that the pro-line-rich region in the TH domain could interact with the neigh-boring SH3 domain [58]. Formation of an intramolecular inter-action has been observed for Itk based on nuclear magnetic res-
R
*
* ** * * *
*
◊ ◊
◊ ◊ ◊
◊ ◊
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
11
146
3591
63136
169315
212358
254403
297448
342493
386537
431582
476627
521672
045
3490
62135
82180
211357
253402
296447
341492
385536
430581
475626
520671
543689
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -M N X X X I L E E X L X K R S Q Q K K K T S P X N F K X R L F V L T K X X L S Y Y E X X X
- - - - - - - - M G C V X X - - - K X X E X X A X K X T X E X X X X X X X S X G X X Y X XX X X K R X S R K G S I D V X K I K C V E X V X X X X X X X X X X X X X X X X X X X X X X
- - - - X P T - - - - P X X X X X X - - - X X X X X P - - - S X - X F X - - X X A X X X XX D X X I P X X X K Y P F Q V V X D X X X L Y V F A P X X E S R X R W V X X L K X E I R X
X X X X - - X X X X - F G - - G - - - - - - - X - - - - - - - - - - - - - N S S F S X T XN X X L X X K Y H P X F W X D G X Y X C C X Q T X K X A X G C X X X E X X X X S X K X X X
E R X L L X X G N P R G X F L V R E S E T T K G A Y S L S - - I R D W D D X K G D X V K HE Q L L R X X G K E - G X F M V R D S - S X X G X Y T V S X F T K A X X D X X X X X I H
Y K I R - - K L D X G G Y Y I T T R A Q F D T L Q Q L V X H Y S E X A D G L C X R L T X -Y H V X X T X X S P K K Y Y L A E K X X F X S I P X L I X Y H Q H N S A G L I T R L R Y P
V C P T X - - K P Q T X G L A K D A W E I P R E S L X L E X K L G Q G C F G E V W G T WV S X X X X X A P X T A G L X Y G X W E I D P X E L T F L K E L G S G Q F G V V X G K W
N G T T K V A I K T L K P G T M S P E A F L Q E A Q I M K K L R H D K L V Q L Y A V V S ER X Q Y X V A I K X I R E G A M S E D E F I E E A X V M M K L S H P K L V Q L Y G V C T X
E - P I Y I V T E Y M X K G S L L D F L K D X E G K X L K L P Q L V D M A A Q I A X G M AQ X P I Y I V T E Y M X X G C L L N Y L R X - X X X X F X X X X L L X M C X D V C E G M X
Y I E R M N Y I H R D L R A A N I L V G E X L I C K I A D F G L A R L I E D N E Y T A R QY L E X X X F I H R D L A A R N C L V N E X X V V K V S D F G M T R Y V L D D Q Y T S X
G A K F P I K W T A P E A A L Y G R F T I K S D V W S F G I L L T E L V T K G R V P Y P GG T K F P V K W S X P E V F X Y S K F S S K S D V W A F G V L M W E V F S X G K X P Y E X
M X N R E V L E Q V E R G Y R M P C P X X C P X S L H E L M X X C W R K D P E E R P T F EX T N S E V V X X I S X G X R L Y R P X L A S X X V Y Q I M X X C W H E K P E X R P T F X
Src KinasesTec Kinases
Src KinasesTec Kinases
Src KinasesTec Kinases
83181
102226
139270
101225
138269
168314
- - - - - T X - - - - - - - - - - R X X X X T G G V T X - - - - - - - - V A L Y DX X X X X X K X L P P X P E X X X R X P X X X P P X P X X X P X X X X X V A L Y D
Y E A R T E D D L S F X K G E R F Q I L N N T E G D W W E A R S L S T G X - - - - - - - -Y Q X X D X X D L X L X R X D E Y Y I L E X X X X X W W R X R D K X - G X X X X X X X X X
- - - - - - - - - - - - - T G Y I P S N Y V A P X - - D S I Q A E E W Y F G K X G R K D AX X X X X X X X X X X X X E G Y I P S X Y X T X X X X N N L E X Y E W Y X K X X S R S X A
- - - FX X X V
Y L Q S F L E D Y F T A T E P Q Y Q P G E N LX L L S X I X E L X - E X E X X X X X - - - -
S
R
ML
Src KinasesTec Kinases
Src KinasesTec Kinases
I P S N Y V A P X - - D S I Q A E E W YI P S X Y X T X X X X N N L E X Y E W Y
T GE G
YY
XX
V A L Y DV A L Y D
Y E A R T E D D L S F X K G E R F Q I L N N T E G D W W E A R S L S T GY Q X X D X X D L X L X R X D E Y Y I L E X X X X X W W R X R D K X - G
- - - FX X X V
◊ ◊ ◊ ◊ ◊
◊ ◊
Figure 3. Consensus alignments between Src and Tec kinases.The consensus was designed based on multiple sequence alignmentsfrom Src and Tec kinases (four members of each). Nonconservedamino acid residues and gaps are shown as (X) and (–) respectively.The yellow boxes show overall identity or homology. (A) Thedomain outlines are PH (orange), TH (blue), SH3 (green), SH2(light-blue), and SH1 (gray). Conserved tyrosines in the SH3 andSH1 domains are indicated by a red box. Phosphotyrosine-bindingmotifs in the SH2 domain (*) and proline rich-binding residues (◊)are indicated. In the SH1 domain, linker (violet), ATP-binding site(red), and substrate-binding region (dark-green) are shown. (B)Consensus alignments of SH3 domains without gaps generated fromflanking regions of Bmx sequences (262-282).
A
B
ACI International, 12/3 (2000)
Research Trends130
onance (NMR) studies of a polypeptide consisting of the pro-line-rich (PR) subdomain with the adjacent SH3 domain. Thismay sequester these sites preventing an interaction with their“natural” ligands [59]. However, it remains to be shown thatsuch interactions take place in the full-length kinases. In Btk,deletion of the SH3 domain or a Y223F mutation result in en-hanced fibroblast transformation, implying that the SH3 domainmay play a negative regulatory role [23].
Despite the presence of several Src kinases in B cells, whensingle members were studied, only mutations in Lyn have beenreported to affect B-cell development [60–62]. Btk activationstill functions in Lyn-deficient cells, probably due to the redun-dant activation by other Src kinases, such as Fyn and Blk.
G-Coupled Proteins
The sequential activation of Tec family kinases is not onlymediated by Src kinases. Other pathways may also be involved.Involvement of heterotrimeric G proteins is one of the earliersuggestions for an interaction with Btk [63, 64], probablythrough binding to the PH [65] and/or TH domains. An interac-tion between Gβγ subunits and the PH domain of Btk [66], aswell as the PH domain of Itk [67], has been reported, thereby in-ducing kinase activation. Recently, it has been suggested thatGi-coupled receptor and Btk activation lie in the same pathway[68]. However, the same group reported later that only Gq-cou-pled receptors or purified Gqα subunits directly stimulate Btk[69]. The binding sites of the Gα subunit on Btk have beenmapped at the PH-TH and TH-SH3 regions with Gqα andGα12, respectively [70, 71].
Tec Kinases Are Lipid-RegulatedProteins
Function of the PH Domain
Mutations in the PH domain of Btk, including the Xid muta-tion (R28C), provided the first evidence for the functional im-portance of any PH domain in vivo [29, 72]. These mutationsclearly interfere with Btk’s role in B-cell development, despitethe intrinsic autophosphorylation activity remain intact. ThePH-containing proteins are recruited to the plasma membranethrough binding to phosphoinositides, end products generatedby the phosphatidylinositol 3-kinase (PI3-K) protein family [73](for reviews see [74–76]). It has been reported that the Btk PHdomain binds strongly to both PI(3,4,5)P3 (PIP3) [77, 78] andI(1,3,4,5)P4 (IP4) [79]. An intact PH domain is required for thefunction of wild type Btk in vivo [29, 72, 80] and is also re-quired for membrane localization, demonstrating the impor-tance of an intact PH domain in the early steps of Btk activation[81]. On the basis of structural studies on the Btk PH domainand the Btk motif, lipid interaction sites have been identified.Naturally occurring point mutations in XLA and Xid interferewith the lipid binding site(s) [82, 83].
Phosphatidylinositol 3-Kinase (PI3-K) Activates Tec Kinases
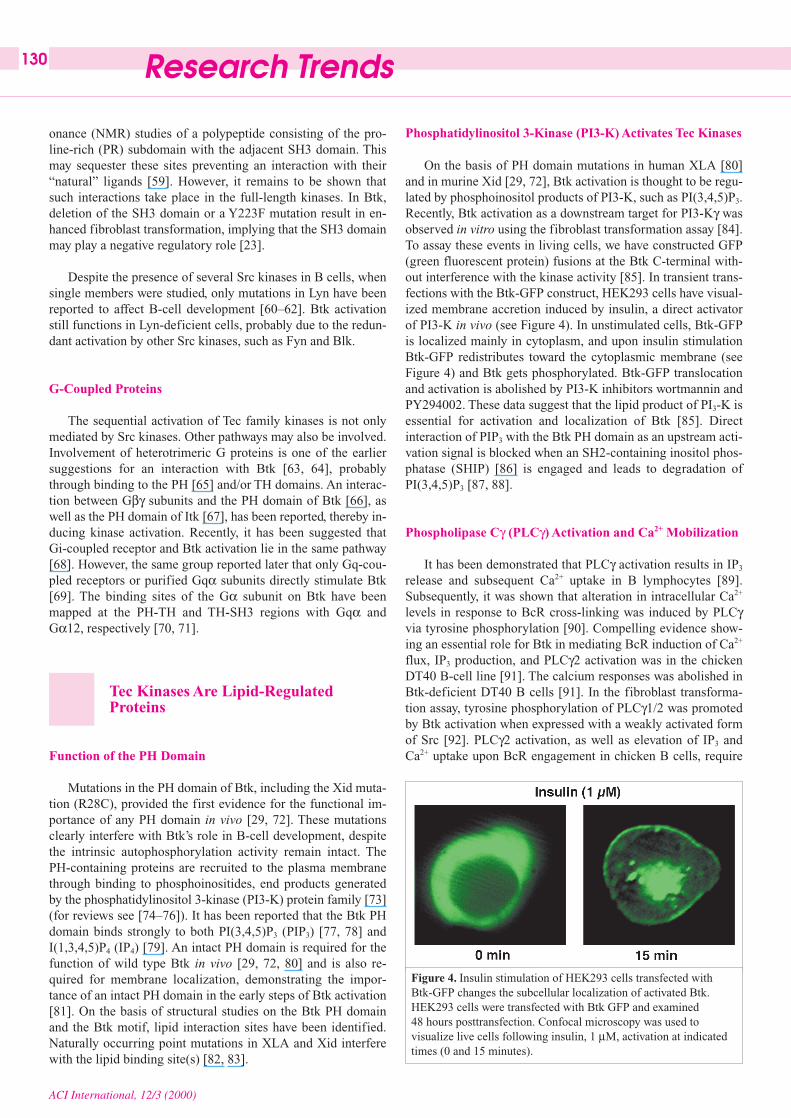
On the basis of PH domain mutations in human XLA [80]and in murine Xid [29, 72], Btk activation is thought to be regu-lated by phosphoinositol products of PI3-K, such as PI(3,4,5)P3.Recently, Btk activation as a downstream target for PI3-Kγ wasobserved in vitro using the fibroblast transformation assay [84].To assay these events in living cells, we have constructed GFP(green fluorescent protein) fusions at the Btk C-terminal with-out interference with the kinase activity [85]. In transient trans-fections with the Btk-GFP construct, HEK293 cells have visual-ized membrane accretion induced by insulin, a direct activatorof PI3-K in vivo (see Figure 4). In unstimulated cells, Btk-GFPis localized mainly in cytoplasm, and upon insulin stimulationBtk-GFP redistributes toward the cytoplasmic membrane (seeFigure 4) and Btk gets phosphorylated. Btk-GFP translocationand activation is abolished by PI3-K inhibitors wortmannin andPY294002. These data suggest that the lipid product of PI3-K isessential for activation and localization of Btk [85]. Directinteraction of PIP3 with the Btk PH domain as an upstream acti-vation signal is blocked when an SH2-containing inositol phos-phatase (SHIP) [86] is engaged and leads to degradation ofPI(3,4,5)P3 [87, 88].
Phospholipase Cγ (PLCγ) Activation and Ca2+ Mobilization
It has been demonstrated that PLCγ activation results in IP3
release and subsequent Ca2+ uptake in B lymphocytes [89].Subsequently, it was shown that alteration in intracellular Ca2+
levels in response to BcR cross-linking was induced by PLCγvia tyrosine phosphorylation [90]. Compelling evidence show-ing an essential role for Btk in mediating BcR induction of Ca2+
flux, IP3 production, and PLCγ2 activation was in the chickenDT40 B-cell line [91]. The calcium responses was abolished inBtk-deficient DT40 B cells [91]. In the fibroblast transforma-tion assay, tyrosine phosphorylation of PLCγ1/2 was promotedby Btk activation when expressed with a weakly activated formof Src [92]. PLCγ2 activation, as well as elevation of IP3 andCa2+ uptake upon BcR engagement in chicken B cells, require
Figure 4. Insulin stimulation of HEK293 cells transfected withBtk-GFP changes the subcellular localization of activated Btk.HEK293 cells were transfected with Btk GFP and examined 48 hours posttransfection. Confocal microscopy was used tovisualize live cells following insulin, 1 µM, activation at indicatedtimes (0 and 15 minutes).

ACI International, 12/3 (2000)
Research Trends 131
both PH and SH2 domains of Btk [91]. In human B cells defi-cient in Btk, as in Btk-deficient DT40 cells, induction of PLCγactivation, Ca2+ store depletion, and IP3 generation are impaired[92]. These events are restored when either Btk or Itk or Tec isexpressed in XLA B cells [92].
Lipid Specificity Determines Kinase Activity
As described earlier, PH domain mutations in human XLAand murine Xid are both interfering with phosphoinositide bind-ing sites as determined by three-dimensional structure analyses[82, 83]. Then, one could ask why is the phenotype of these mu-tations so severe? One reason is that the lipid-binding mutationsof the PH domain block membrane translocation. Along withthese observations, we propose a hypothesis for the regulationand activation of lipid-binding kinases (see Figure 5). Recently,it was shown that Btk gets activated entirely at the cytoplasmicmembrane [81]. Stimulatory signals modulate Btk not only atthe activation (phosphorylation) step but also in localization. Incontrast, expression of a constitutively active form of Btk carry-ing the E41K gain-of-function mutation results in a B-cell de-fect that is more severe than Xid [93]. This implies that continu-ous stimulatory signals which target Btk to the cell membraneare toxic. Therefore, inhibitory signals affecting PIP3 degrada-tion are needed, as shown for SHIP, which blocks upstream acti-vation of Btk [87, 88]. Thus, the inactivation step of Btk re-quires tyrosine dephosphorylation and PIP3 degradation (seeFigure 5). As a result of this, dephosphorylated Btk is releasedto the cytoplasm. In the cytoplasm, all Btk molecules are proba-bly in an “inactive” form (see Figure 5). Binding of IP4 to theinactive form of Btk might protect the protein stability againstproteolytic cleavage. Recent biochemical [78, 79] experimentsshow that the IP4 binding affinity for Btk is higher than that of
PIP3. On the other hand, the structural data reveal that both IP4
and PIP3 have identical positions in the Btk-PH domain [82,83]. In resting cells, signaling molecules and kinases must beintact in order to respond stimulatory signals rapidly (see Figure5). To support our model, experiments are underway to deter-mine if Btk bound IP4 is more stable compared to unbound Btk.This model might be generally applicable to rapid ON–OFFregulation of activity of proteins containing PH and PH-likedomains.
References
[1] Kurosaki T. Molecular mechanisms in B-cell antigen receptor signaling.Curr Opin Immunol 1997; 9:309–318
[2] Isakov N. ITIMs and ITAMs. The Yin and Yang of antigen and Fc re-ceptor-linked signaling machinery. Immunol Res 1997; 16:85–100
[3] Gold MR, Law DA, DeFranco AL. Stimulation of protein tyrosinephosphorylation by the B-lymphocyte antigen receptor. Nature 1990;345:810–813
[4] Bolen JB. Protein tyrosine kinases in the initiation of antigen-receptorsignaling. Curr Opin Immunol 1995; 7:306–311
[5] Weiss A, Littman DR. Signal transduction by lymphocyte antigen re-ceptors. Cell 1994; 76:263–274
[6] Mahajan S, Fargnoli J, Burkhardt AL, Kut SA, Saouaf SJ, Bolen JB. Srcfamily protein tyrosine kinases induce autoactivation of Bruton’s tyrosine ki-nase. Mol Cell Biol 1995; 15:5304–5311
[7] Rawlings DJ, Scharenberg AM, Park H, Wahl MI, Lin S, Kato RM,Fluckiger AC, Witte ON, Kinet JP. Activation of BTK by a phosphorylationmechanism initiated by SRC family kinases. Science 1996; 271:822–825
[8] Mano H, Yamashita Y, Sato K, Yazaki Y, Hirai H. Tec protein-tyrosinekinase is involved in interleukin-3 signaling pathway. Blood 1995; 85:343–350
[9] Vetrie D, Vorechovsky I, Sideras P, Holland J, Davies A, Flinter F,Hammarstrom L, Kinnon C, Levinsky R, Bobrow M et al. The gene involved inX-linked agammaglobulinaemia is a member of the src family of protein-tyro-sine kinases [published erratum appears in Nature 1993; 364(6435):362] [seecomments]. Nature 1993; 361:226–233
[10] Tsukada S, Saffran SD, Rawlings DJ, Parolini O, Allen RC, Klisak I,Sparkes RS, Kubagawa H, Mohandas T, Quan S et al. Deficient expression of aB-cell cytoplasmic tyrosine kinase in human X-linked agammaglobulinemia.Cell 1993; 72:279–290
[11] Siliciano JD, Morrow TA, Desiderio SV. Itk, a T-cell-specific tyrosinekinase gene inducible by interleukin 2. Proc Natl Acad Sci USA 1992;89:11194–11198
[12] Heyeck SD, Berg LJ. Developmental regulation of a murine T-cell-spe-cific tyrosine kinase gene, Tsk. Proc Natl Acad Sci USA 1993; 90:669–673
[13] Tamagnone L, Lahtinen I, Mustonen T, Virtaneva K, Francis F,Muscatelli F, Alitalo R, Smith CI, Larsson C, Alitalo K. BMX, a novel nonre-ceptor tyrosine kinase gene of the BTK/ITK/TEC/TXK family located in chro-mosome Xp22.2. Oncogene 1994; 9:3683–3688
[14] Kaukonen J, Lahtinen I, Laine S, Alitalo K, Palotie A. BMX tyrosinekinase gene is expressed in granulocytes and myeloid leukaemias. Br JHaematol 1996; 94:455–460
[15] Haire RN, Ohta Y, Lewis JE, Fu SM, Kroisel P, Litman GW. TXK, anovel human tyrosine kinase expressed in T cells shares sequence identity withTec family kinases and maps to 4p12. Hum Mol Genet 1994; 3:897–901
[16] Gregory RJ, Kammermeyer KL, Vincent WSD, Wadsworth SG.Primary sequence and developmental expression of a novel Drosophilamelanogaster src gene. Mol Cell Biol 1987; 7:2119–2127
[17] Haire RN, Strong SJ, Litman GW. Tec-family non-receptor tyrosinekinase expressed in zebrafish kidney. Immunogenetics 1998; 47:336-7
[18] Haire RN, Strong SJ, Litman GW. Identification and characterizationof a homologue of Bruton’s tyrosine kinase, a Tec kinase involved in B-cell de-velopment, in a modern representative of a phylogenetically ancient vertebrate.Immunogenetics 1997; 46:349–351
[19] Rawlings DJ, Witte ON. The Btk subfamily of cytoplasmic tyrosine ki-nases: Structure, regulation and function. Semin Immunol 1995; 7:237–246
[20] Weil D, Power MA, Smith SI, Li CL. Predominant expression ofmurine Bmx tyrosine kinase in the granulo- monocytic lineage. Blood 1997;90:4332–4340
B CA
Inactive
Src activeSHIP inactive
PTP inactive
IP4
SHIP activeSrc inactivePTP active
IP4
ActivePartially Active
pY551
Y223
IP4
membrane
PIP3
membrane membrane
pY551
pY223
PIP3
IP4
Y551Y223
SHIP inactiveSrc activePTP inactive
PIP3
Figure 5. Schematic model showing main steps in phospholipid-binding specificity in Btk function. The regulation in Btk kinaseactivity due to interchange of lipid specificity of inositol 1,3,4,5-tetrakis-phosphate (IP4) and phosphatidylinositol 3,4,5-triphosphate(PIP3). Exchange of PIP3 for IP4 turns the active form of Btk to aninactivate form, i.e., ON–OFF switcher of tyrosine kinase activitysignal. SHIP = SH2 domain-containing inositol-polyphosphate 5’-phosphate; PTP = protein tyrosine phosphatase.

ACI International, 12/3 (2000)
Research Trends132
[21] Vihinen M, Nilsson L, Smith CI. Tec homology (TH) adjacent to thePH domain. FEBS Lett 1994; 350:263–265
[22] Mattsson PT, Vihinen M, Smith CI. X-linked agammaglobulinemia(XLA): A genetic tyrosine kinase (Btk) disease. Bioessays 1996; 18:825–834
[23] Park H, Wahl MI, Afar DE, Turck CW, Rawlings DJ, Tam C,Scharenberg AM, Kinet JP, Witte ON. Regulation of Btk function by a major au-tophosphorylation site within the SH3 domain. Immunity 1996; 4:515–525
[24] Aoki Y, Isselbacher KJ, Pillai S. Bruton tyrosine kinase is tyrosinephosphorylated and activated in pre-B lymphocytes and receptor-ligated B cells.Proc Natl Acad Sci SA 1994; 91:10606–10609
[25] de Weers M, Brouns GS, Hinshelwood S, Kinnon C, Schuurman RK,Hendriks RW, Borst J. B-cell antigen receptor stimulation activates the humanBruton’s tyrosine kinase, which is deficient in X-linked agammaglobulinemia. JBiol Chem 1994; 269:23857–23860
[26] Gibson S, August A, Kawakami Y, Kawakami T, Dupont B, Mills GB.The EMT/ITK/TSK (EMT) tyrosine kinase is activated during TcR signaling:LCK is required for optimal activation of EMT. J Immunol 1996; 156:2716–2722
[27] Smith CI, Notarangelo LD. Molecular basis for X-linked immunodefi-ciencies. Adv Genet 1997; 35:57–115
[28] Vihinen M, Brandau O, Branden LJ, Kwan SP, Lappalainen I, Lester T,Noordzij JG, Ochs HD, Ollila J, Pienaar SM, Riikonen P, Saha BK, Smith CIE.BTKbase, mutation database for X-linked agammaglobulinemia (XLA). NuclAcids Res 1998; 26:242–247
[29] Thomas JD, Sideras P, Smith CI, Vorechovsky I, Chapman V, Paul WE.Colocalization of X-linked agammaglobulinemia and X-linked immunodefi-ciency genes. Science 1993; 261:355–358
[30] Kerner JD, Appleby MW, Mohr RN, Chien S, Rawlings DJ,Maliszewski CR, Witte ON, Perlmutter RM. Impaired expansion of mouse Bcell progenitors lacking Btk. Immunity 1995; 3:301–312
[31] Khan WN, Alt FW, Gerstein RM, Malynn BA, Larsson I, Rathbun G,Davidson L, Muller S, Kantor AB, Herzenberg LA et al. Defective B cell devel-opment and function in Btk-deficient mice. Immunity 1995; 3:283–299
[32] Liu KQ, Bunnell SC, Gurniak CB, Berg LJ. T cell receptor-initiatedcalcium release is uncoupled from capacitative calcium entry in Itk-deficient Tcells. J Exp Med 1998; 187:1721–177
[33] Kong GH, Bu JY, Kurosaki T, Shaw AS, Chan AC. Reconstitution ofSyk function by the ZAP-70 protein tyrosine kinase. Immunity 1995; 2:485–492
[34] Heyeck SD, Wilcox HM, Bunnell SC, Berg LJ. Lck phosphorylates theactivation loop tyrosine of the Itk kinase domain and activates Itk kinase activi-ty. J Biol Chem 1997; 272:25401–25408
[35] August A, Gibson S, Kawakami Y, Kawakami T, Mills GB, Dupont B.CD28 is associated with and induces the immediate tyrosine phosphorylationand activation of the Tec family kinase ITK/EMT in the human Jurkat leukemicT-cell line. Proc Natl Acad Sci USA 1994; 91:9347–9351
[36] King PD, Sadra A, Teng JM, Xiao-Rong L, Han A, Selvakumar A,August A, Dupont B. Analysis of CD28 cytoplasmic tail tyrosine residues asregulators and substrates for the protein tyrosine kinases, EMT and LCK. JImmunol 1997; 158:580-90
[37] Sato S, Katagiri T, Takaki S, Kikuchi Y, Hitoshi Y, Yonehara S, TsukadaS, Kitamura D, Watanabe T, Witte O et al. IL-5 receptor-mediated tyrosine phos-phorylation of SH2/SH3-containing proteins and activation of Bruton’s tyrosineand Janus 2 kinases. J Exp Med 1994; 180:2101–2111
[38] Hitoshi Y, Sonoda E, Kikuchi Y, Yonehara S, Nakauchi H, Takatsu K.IL-5 receptor positive B cells, but not eosinophils, are functionally and numeri-cally influenced in mice carrying the X-linked immune defect. Int Immunol1993; 5:1183–1190
[39] Matsuda T, Takahashi-Tezuka M, Fukada T, Okuyama Y, Fujitani Y,Tsukada S, Mano H, Hirai H, Witte ON, Hirano T. Association and activation ofBtk and Tec tyrosine kinases by gp130, a signal transducer of the interleukin-6family of cytokines. Blood 1995; 85:627–633
[40] Machide M, Mano H, Todokoro K. Interleukin 3 and erythropoietin in-duce association of Vav with Tec kinase through Tec homology domain.Oncogene 1995; 11:619–625
[41] Tang B, Mano H, Yi T, Ihle JN. Tec kinase associates with c-kit and istyrosine phosphorylated and activated following stem cell factor binding. MolCell Biol 1994; 14:8432–8437
[42] Qiu Y, Robinson D, Pretlow TG, Kung HJ. Etk/Bmx, a tyrosine kinasewith a pleckstrin-homology domain, is an effector of phosphatidylinositol 3’-ki-nase and is involved in interleukin 6-induced neuroendocrine differentiation ofprostate cancer cells. Proc Natl Acad Sci USA 1998; 95:3644–3649
[43] Saharinen P, Ekman N, Sarvas K, Parker P, Alitalo K, Silvennoinen O.The Bmx tyrosine kinase induces activation of the Stat signaling pathway, which is specifically inhibited by protein kinase Cδ. Blood 1997; 90:4341–4353
[44] Anderson JS, Teutsch M, Dong Z, Wortis HH. An essential role forBruton’s tyrosine kinase in the regulation of B-cell apoptosis. Proc Natl AcadSci U S A 1996; 93:10966–10971
[45] Solvason N, Wu WW, Kabra N, Lund-Johansen F, Roncarolo MG,Behrens TW, Grillot DA, Nunez G, Lees E, Howard M. Transgene expression ofbcl-xL permits anti-immunoglobulin (Ig)-induced proliferation in xid B cells. JExp Med 1998; 187:1081–1091
[46] Brorson K, Brunswick M, Ezhevsky S, Wei DG, Berg R, Scott D, SteinKE. xid affects events leading to B-cell cycle entry. J Immunol 1997; 159:135–143
[47] Vassilev A, Ozer Z, Navara C, Mahajan S, Uckun FM. Bruton’s tyro-sine kinase as an inhibitor of the Fas/CD95 death-inducing signaling complex. J Biol Chem 1999; 274:1646–1656
[48] Saouaf SJ, Mahajan S, Rowley RB, Kut SA, Fargnoli J, Burkhardt AL,Tsukada S, Witte ON, Bolen JB. Temporal differences in the activation of threeclasses of non-transmembrane protein tyrosine kinases following B-cell antigenreceptor surface engagement. Proc Natl Acad Sci USA 1994; 91:9524–9528
[49] Li T, Tsukada S, Satterthwaite A, Havlik MH, Park H, Takatsu K, WitteON. Activation of Bruton’s tyrosine kinase (BTK) by a point mutation in itspleckstrin homology (PH) domain. Immunity 1995; 2:451–460
[50] Afar DE, Park H, Howell BW, Rawlings DJ, Cooper J, Witte ON.Regulation of Btk by Src family tyrosine kinases. Mol Cell Biol 1996;16:3465–3471
[51] Cory GO, MacCarthy-Morrogh L, Banin S, Gout I, Brickell PM,Levinsky RJ, Kinnon C, Lovering RC. Evidence that the Wiskott-Aldrich syn-drome protein may be involved in lymphoid cell signaling pathways. J Immunol1996; 157:3791–3795
[52] Cory GO, Lovering RC, Hinshelwood S, MacCarthy-Morrogh L,Levinsky RJ, Kinnon C. The protein product of the c-cbl protooncogene is phos-phorylated after B cell receptor stimulation and binds the SH3 domain ofBruton’s tyrosine kinase. J Exp Med 1995; 182:611–615
[53] Matsushita M, Yamadori T, Kato S, Takemoto Y, Inazawa J, Baba Y,Hashimoto S, Sekine S, Arai S, Kunikata T, Kurimoto M, Kishimoto T, TsukadaS. Identification and characterization of a novel SH3-domain binding protein,Sab, which preferentially associates with Bruton’s tyrosine kinase (BtK).Biochem Biophys Res Commun 1998; 245:337–343
[54] Guinamard R, Fougereau M, Seckinger P. The SH3 domain of Bruton’styrosine kinase interacts with Vav, Sam68 and EWS. Scand J Immunol 1997;45:587–595
[55] Mano H, Yamashita Y, Miyazato A, Miura Y, Ozawa K. Tec protein-ty-rosine kinase is an effector molecule of Lyn protein-tyrosine kinase. FASEB J1996; 10:637–642
[56] August A, Sadra A, Dupont B, Hanafusa H. Src-induced activation ofinducible T cell kinase (ITK) requires phosphatidylinositol 3-kinase activity andthe Pleckstrin homology domain of inducible T cell kinase. Proc Natl Acad SciUSA 1997; 94:11227–11232
[57] Debnath J, Chamorro M, Czar MJ, Schaeffer EM, Lenardo MJ, VarmusHE, Schwartzberg PL. rlk/TXK encodes two forms of a novel cysteine string ty-rosine kinase activated by src family kinases. Mol Cell Biol 1999;19:1498–1507
[58] Smith CI, Islam KB, Vorechovsky I, Olerup O, Wallin E, Rabbani H,Baskin B, Hammarstrom L. X-linked agammaglobulinemia and other im-munoglobulin deficiencies. Immunol Rev 1994; 138:159–183
[59] Andreotti AH, Bunnell SC, Feng S, Berg LJ, Schreiber SL. Regulatoryintramolecular association in a tyrosine kinase of the Tec family. Nature 1997;385:93-7
[60] Chan VW, Meng F, Soriano P, DeFranco AL, Lowell CA.Characterization of the B lymphocyte populations in Lyn-deficient mice andthe role of Lyn in signal initiation and downregulation. Immunity 1997; 7:69–81
[61] Hibbs ML, Tarlinton DM, Armes J, Grail D, Hodgson G, Maglitto R,Stacker SA, Dunn AR. Multiple defects in the immune system of Lyn-deficientmice, culminating in autoimmune disease. Cell 1995; 83:301–311
[62] Nishizumi H, Taniuchi I, Yamanashi Y, Kitamura D, Ilic D, Mori S,Watanabe T, Yamamoto T. Impaired proliferation of peripheral B cells and indi-cation of autoimmune disease in lyn-deficient mice. Immunity 1995; 3:549–560
[63] Clapham DE, Neer EJ. New roles for G-protein βγ-dimers in trans-membrane signalling. Nature 1993; 365:403–406
[64] Clapham DE, Neer EJ. G protein βγ subunits. Annu Rev PharmacolToxicol 1997; 37:167–203
[65] Touhara K, Inglese J, Pitcher JA, Shaw G, Lefkowitz RJ. Binding of Gprotein βγ-subunits to pleckstrin homology domains. J Biol Chem 1994;269:10217–10220
[66] Tsukada S, Simon MI, Witte ON, Katz A. Binding of βγ subunits ofheterotrimeric G proteins to the PH domain of Bruton tyrosine kinase. ProcNatl Acad Sci USA 1994; 91:11256–11260

ACI International, 12/3 (2000)
Research Trends 133
[67] Langhans-Rajasekaran SA, Wan Y, Huang XY. Activation of Tsk andBtk tyrosine kinases by G protein βγ subunits. Proc Natl Acad Sci USA 1995;92:8601–8605
[68] Wan Y, Bence K, Hata A, Kurosaki T, Veillette A, Huang XY. Geneticevidence for a tyrosine kinase cascade preceding the mitogen-activated proteinkinase cascade in vertebrate G protein signaling. J Biol Chem 1997;272:17209–17215
[69] Bence K, Ma W, Kozasa T, Huang XY. Direct stimulation of Bruton’styrosine kinase by G(q)-protein α subunit. Nature 1997; 389:296–299
[70] Jiang Y, Ma W, Wan Y, Kozasa T, Hattori S, Huang XY. The G proteinG α12 stimulates Bruton’s tyrosine kinase and a rasGAP through a conservedPH/BM domain. Nature 1998; 395:808–813
[71] Ma YC, Huang XY. Identification of the binding site for Gqα on its ef-fector Bruton’s tyrosine kinase. Proc Natl Acad Sci USA 1998; 95:12197–12201
[72] Rawlings DJ, Saffran DC, Tsukada S, Largaespada DA, Grimaldi JC,Cohen L, Mohr RN, Bazan JF, Howard M, Copeland NG, et al. Mutation ofunique region of Bruton’s tyrosine kinase in immunodeficient XID mice.Science 1993; 261:358–361
[73] Harlan JE, Hajduk PJ, Yoon HS, Fesik SW. Pleckstrin homologydomains bind to phosphatidylinositol-4,5-bisphosphate. Nature 1994;371:168–170
[74] Wymann MP, Pirola L. Structure and function of phosphoinositide 3-kinases. Biochim Biophys Acta 1998; 1436:1270–150
[75] Bottomley MJ, Salim K, Panayotou G. Phospholipid-binding proteindomains. Biochim Biophys Acta 1998; 1436:165–183
[76] Hemmings BA. PtdIns(3,4,5)P3 gets its message across [comment].Science 1997; 277:534
[77] Salim K, Bottomley MJ, Querfurth E, Zvelebil MJ, Gout I, Scaife R,Margolis RL, Gigg R, Smith CI, Driscoll PC, Waterfield MD, Panayotou G.Distinct specificity in the recognition of phosphoinositides by the pleckstrin ho-mology domains of dynamin and Bruton’s tyrosine kinase. EMBO J 1996;15:6241–6250
[78] Rameh LE, Arvidsson A, Carraway, KL, 3rd, Couvillon AD, RathbunG, Crompton A, VanRenterghem B, Czech MP, Ravichandran KS, Burakoff SJ,Wang DS, Chen CS, Cantley LC. A comparative analysis of the phosphoinosi-tide binding specificity of pleckstrin homology domains. J Biol Chem 1997;272:22059–22066
[79] Fukuda M, Kojima T, Kabayama H, Mikoshiba K. Mutation of thepleckstrin homology domain of Bruton’s tyrosine kinase in immunodeficiencyimpaired inositol 1,3,4,5-tetrakisphosphate binding capacity. J Biol Chem 1996;271:30303–30306
[80] Vihinen M, Zvelebil MJ, Zhu Q, Brooimans RA, Ochs HD, Zegers BJ,Nilsson L, Waterfield MD, Smith CI. Structural basis for pleckstrin homologydomain mutations in X-linked agammaglobulinemia. Biochemistry 1995;34:1475–1481
[81] Li T, Rawlings DJ, Park H, Kato RM, Witte ON, Satterthwaite AB.Constitutive membrane association potentiates activation of Bruton tyrosine ki-nase. Oncogene 1997; 15:1375–1383
[82] Hyvonen M, Saraste M. Structure of the PH domain and Btk motiffrom Bruton’s tyrosine kinase: Molecular explanations for X-linked agamma-globulinaemia. EMBO J 1997; 16:3396–3404
[83] Baraldi E, Carugo KD, Hyvonen M, Surdo PL, Riley AM, Potter BV,O’Brien R, Ladbury JE, Saraste M. Structure of the PH domain from Bruton’s
tyrosine kinase in complex with inositol 1,3,4,5-tetrakisphosphate. Structure1999; 7:449–460
[84] Li Z, Wahl MI, Eguinoa A, Stephens LR, Hawkins PT, Witte ON.Phosphatidylinositol 3-kinase-γ activates Bruton’s tyrosine kinase in concertwith Src family kinases. Proc Natl Acad Sci USA 1997; 94:13820–13825
[85] Nore BF, Vargas L, Mohamed AJ, Branden LJ, Backesjo CM, IslamTC, Mattsson PT, Hultenby K, Christensson B, Smith CI. Redistribution ofBruton’s tyrosine kinase by activation of phosphatidylinositol 3-kinase and Rho-family GTPases. Eur J Immunol 2000; 30:145–154
[86] Chacko GW, Tridandapani S, Damen JE, Liu L, Krystal G, CoggeshallKM. Negative signaling in B lymphocytes induces tyrosine phosphorylation ofthe 145-kDa inositol polyphosphate 5-phosphatase, SHIP. J Immunol 1996;157:2234–2238
[87] Bolland S, Pearse RN, Kurosaki T, Ravetch JV. SHIP modulates im-mune receptor responses by regulating membrane association of Btk. Immunity1998; 8:509–516
[88] Scharenberg AM, El-Hillal O, Fruman DA, Beitz LO, Li Z, Lin S,Gout I, Cantley LC, Rawlings DJ, Kinet JP. Phosphatidylinositol-3,4,5-trisphos-phate (PtdIns-3,4,5-P3)/Tec kinase-dependent calcium signaling pathway: a tar-get for SHIP-mediated inhibitory signals. EMBO J 1998; 17:1961–1972
[89] Ransom JT, Harris LK, Cambier JC. Anti-Ig induces release of inositol1,4,5-trisphosphate, which mediates mobilization of intracellular Ca++ stores inB lymphocytes. J Immunol 1986; 137:708–714
[90] Padeh S, Cohen A, Roifman CM. ATP-induced activation of human Blymphocytes via P2-purinoceptors. J Immunol 1991; 146:1626–1632
[91] Takata M, Kurosaki T. A role for Bruton’s tyrosine kinase in B-cellantigen receptor-mediated activation of phospholipase C-γ 2. J Exp Med 1996;184:31–40
[92] Fluckiger AC, Li Z, Kato RM, Wahl MI, Ochs HD, Longnecker R,Kinet JP, Witte ON, Scharenberg AM, Rawlings DJ. Btk/Tec kinases regulatesustained increases in intracellular Ca2+ following B-cell receptor activation.EMBO J 1998; 17:1973–1985
[93] Dingjan GM, Maas A, Nawijn MC, Smit L, Voerman JS, Grosveld F,Hendriks RW. Severe B cell deficiency and disrupted splenic architecture intransgenic mice expressing the E41K mutated form of Bruton’s tyrosine kinase.EMBO J 1998; 17:5309–5320
[94] Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. TheCLUSTAL_X windows interface: Flexible strategies for multiple sequencealignment aided by quality analysis tools. Nucl Acid Res 1997; 25:4876–4882
C.I. Edvard Smith (to whom correspondence should beaddressed), Beston F. Nore, Abdalla J. Mohamed, LeonardoVargas, Lars J. Brandén, and Carl-Magnus Bäckesjö are withthe Center for BioTechnology, Department of Bioscience atNovum, Karolinska Institute, SE-141 57 Huddinge, Sweden(tel. +46 8 608-9114, fax +46 8 774-5538, [email protected]). Mauno Vihinen is with theDepartment of Medical Technology, University of Tampere,P.O. Box 607, FIN-33101 Tampere, Finland.
… continued from page 94
2nd European Conference on PediatricAsthmaNovember 23–24, 2000: London, UK
Contact: Castle House Medical,Conferences, 3 Linden Close, TunbridgeWells, Kent TN4 8HH, UK, Tel. +441892
539606, Fax +441892517773,E-mail [email protected]
Annual Meeting of the German Societyfor Immunology and of the Neder-landse Vereniging voor ImmunologieNovember 29–December 2, 2000:Düsseldorf, Germany
Contact: Prof. Dr. Ernst Gleichmann,Sekretariat Frau Steil, MedizinischesInstitut für Umwelthygiene, Auf’mHennekamp 50, D–40225 Düsseldorf,Germany, Tel. +49 211 3389212, Fax +49211 3190910, E-mail dgfi2000@uni-dues-
seldorf.de, Internet www.miu.uni-duessel-dorf.de/dgfi2000
XXth Congress of the EuropeanAcademy of Allergology & ClinicalImmunologyMay 9–13, 2001: Berlin, Germany
Dept. of Pediatric Pneumology & Immuno-logy, Charité, Augustenburger Platz 1,D–13353 Berlin, Tel. +49 30 4506-6131,Fax +49 30 4506-6931, E-mail [email protected], Internet www.eaaci.org
… continued on page 138
Meeting Calendar
Copyright © 2022 FDOKUMEN




















