
Increased alkaline phosphatase specific activity in reeler cerebella
Upload
independentCategory
view
2download
0
Molecular Cell, Vol. 17, 49–59, January 7, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.molcel.2004.11.043
Inactivation of the Cdc25 Phosphatase by theStress-Activated Srk1 Kinase in Fission Yeast
this, the activity of Cdc25 is tightly regulated, being lowthroughout interphase and activated only as cells entermitosis in part because of phosphorylation of Cdc25 by
Sandra Lopez-Aviles,1 Maribel Grande,1
Marta Gonzalez,1 Ase-Lill Helgesen,1
Vicenc Alemany,1 Maribel Sanchez-Piris,1,3
both Cdc2/cyclin B and polo-like kinases (Kumagai andOriol Bachs,1 Jonathan B.A. Millar,2
Dunphy, 1992; Hoffmann et al. 1993; Gabrielli et al.,and Rosa Aligue1,*1997; Qian et al., 2001). This phosphorylation has been1Departament de Biologia Cel·lularproposed to initiate an autoactivation loop that effi-Institut de Investigacions Biomediques Augustciently drives cells into mitosis (Hoffmann et al. 1993).Pi i Sunyer
Cdc25 is also subject to negative regulatory phos-Universitat de Barcelonaphorylation when the genome is either damaged or notE-08036 Barcelonacompletely replicated. In these situations, Cdc25 isSpainphosphorylated by the Chk1 and Cds1 (Chk2) kinases2 Division of Yeast Genetics(Sanchez et al., 1997; Zeng et al., 1998; Furnari et al.National Institute for Medical Research1999). In fission yeast, this occurs on at least nine phos-The Ridgeway, Mill Hillphorylation sites in the N-terminal regulatory domainLondon NW7 1AAof the protein, and phosphorylation of these sites isUnited Kingdomnecessary for checkpoint arrest (Furnari et al. 1999;Zeng and Piwnica-Worms, 1999). Notably, Cdc25 is ex-ported from the nucleus when the DNA damage or repli-Summarycation checkpoints are activated or when the Cds1 orChk1 kinases are overexpressed (Kumagai and Dunphy,The mechanisms by which environmental stress regu-1999; Lopez-Girona et al., 1999). Because Cdc2/cyclin Blates cell cycle progression are poorly understood. Inremains nuclear throughout the cell cycle, it was thoughtfission yeast, we show that Srk1 kinase, which associ-that this may provide a mechanism to keep Cdc25 fromates with the stress-activated p38/Sty1 MAP kinase,its substrate (Furnari et al. 1999; Zeng and Piwnica-regulates the onset of mitosis by inhibiting the Cdc25Worms, 1999). However, nuclear exclusion of Cdc25phosphatase. Srk1 is periodically active in G2, andis not sufficient to induce a checkpoint arrest (Lopez-its overexpression causes cell cycle arrest in late G2Girona et al., 2001). Instead, phosphorylation of Cdc25phase, whereas cells lacking srk1 enter mitosis pre-on these sites also inhibits its catalytic activity in vitromaturely. We find that Srk1 interacts with and phos-(Lopez-Girona et al., 2001). In addition, phosphorylationphorylates Cdc25 at the same sites phosphorylatedof Cdc25 by Cds1 and Chk1 kinases provides a bindingby the Chk1 and Cds1 (Chk2) kinases and that thissite for the 14-3-3 proteins Rad24 and Rad25, which arephosphorylation is necessary for Srk1 to delay mitoticthemselves required for checkpoint arrest (Peng et al.,entry. Phosphorylation by Srk1 causes Cdc25 to bind1997; Kumagai and Dunphy, 1999; Lopez-Girona et al.,to Rad24, a 14-3-3 protein family member, and accu-1999; Zeng and Piwnica-Worms, 1999; Graves et al., 2001).mulation of Cdc25 in the cytoplasm. However, Srk1Rad24 and Rad25 are thought to protect Cdc25 fromdoes not regulate Cdc25 in response to replicationdephosphorylation during checkpoint arrest and stabi-
arrest or DNA damage but, rather, during a normal celllize the protein in an inactive state in the cytoplasm.
cycle and in response to nongenotoxic environmen-However, Rad24 also interacts with Chk1, and this inter-
tal stress. action is both induced and required for checkpoint ar-rest, suggesting the role of Rad24 is complex (Chen et
Introduction al., 1999; Walworth, 2001). Indeed, interaction of Rad24with Cdc25 is observed in a normal interphase indicating
In eukaryotic cells, entry into mitosis requires the activity that as-yet-unidentified kinase(s), other than Chk1 orof the cyclin-dependent kinase Cdc2 (Cdk1). In fission Cds1, promote this association presumably to preventyeast, Schizosaccharomyces pombe, the tyrosine ki- ectopic activation of Cdc25 during interphase (Peng etnases Wee1 and Mik1 downregulate mitosis by phos- al., 1997; Chen et al., 1999).phorylating Cdc2 on Tyr15 (Russell and Nurse, 1987; In mammalian cells, the G2/M transition can be de-Lundgren et al., 1991; Aligue et al., 1997). Cdc25 is a layed by activation of an “antephase checkpoint” in re-universally conserved protein phosphatase that pro- sponse to a variety of environmental insults, includingmotes mitotic entry by dephosphorylating Cdc2 on hyperosmotic stress, microtubule disassembly, and in-Tyr15/Thr14, thereby activating Cdc2 (Russell and hibition of histone deacetylases (Matsusaka and Pines,Nurse, 1986; Gautier et al., 1991; Millar et al., 1991). In 2004; Mikhailov et al., 2004). This checkpoint is indepen-fission yeast, Cdc25 protein accumulates in the nucleus dent of the ATM and ATR checkpoint proteins but in-through the cell cycle and is most abundant in late G2 stead requires activation of the p38 MAP kinase pathway(Moreno et al., 1990; Lopez-Girona et al., 1999). Despite (Wang et al., 2000; Bulavin et al., 2001; Matsusaka and
Pines, 2004; Mikhailov et al., 2004). Intriguingly, the p38MAP kinase has been reported to bind and phosphory-*Correspondence: [email protected] Cdc25B at Ser309 and Ser361 and Cdc25C at3Present address: Instituto de Investigaciones Citologicas, 46010
Valencia, Spain. Ser216, which promotes binding to 14-3-3 proteins (Bu-
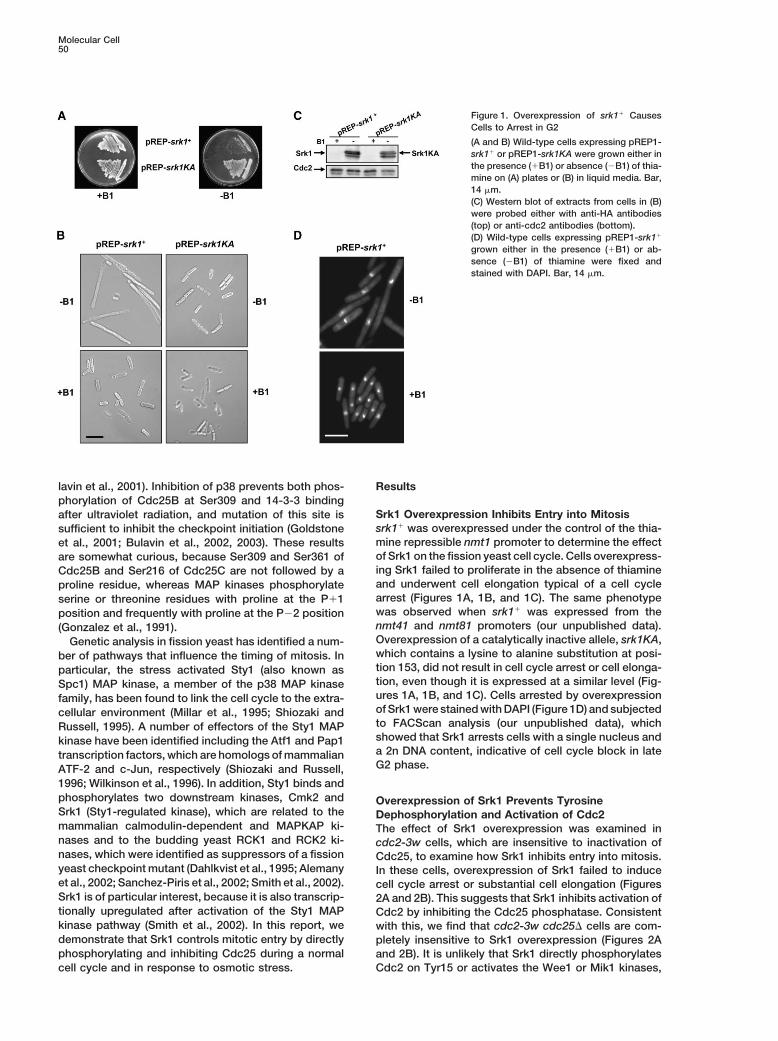
Molecular Cell50
Figure 1. Overexpression of srk1� CausesCells to Arrest in G2
(A and B) Wild-type cells expressing pREP1-srk1� or pREP1-srk1KA were grown either inthe presence (�B1) or absence (�B1) of thia-mine on (A) plates or (B) in liquid media. Bar,14 �m.(C) Western blot of extracts from cells in (B)were probed either with anti-HA antibodies(top) or anti-cdc2 antibodies (bottom).(D) Wild-type cells expressing pREP1-srk1�
grown either in the presence (�B1) or ab-sence (�B1) of thiamine were fixed andstained with DAPI. Bar, 14 �m.
lavin et al., 2001). Inhibition of p38 prevents both phos- Resultsphorylation of Cdc25B at Ser309 and 14-3-3 binding
Srk1 Overexpression Inhibits Entry into Mitosisafter ultraviolet radiation, and mutation of this site issrk1� was overexpressed under the control of the thia-sufficient to inhibit the checkpoint initiation (Goldstonemine repressible nmt1 promoter to determine the effectet al., 2001; Bulavin et al., 2002, 2003). These resultsof Srk1 on the fission yeast cell cycle. Cells overexpress-are somewhat curious, because Ser309 and Ser361 ofing Srk1 failed to proliferate in the absence of thiamineCdc25B and Ser216 of Cdc25C are not followed by aand underwent cell elongation typical of a cell cycleproline residue, whereas MAP kinases phosphorylatearrest (Figures 1A, 1B, and 1C). The same phenotypeserine or threonine residues with proline at the P�1was observed when srk1� was expressed from theposition and frequently with proline at the P�2 positionnmt41 and nmt81 promoters (our unpublished data).(Gonzalez et al., 1991).Overexpression of a catalytically inactive allele, srk1KA,Genetic analysis in fission yeast has identified a num-which contains a lysine to alanine substitution at posi-ber of pathways that influence the timing of mitosis. Intion 153, did not result in cell cycle arrest or cell elonga-particular, the stress activated Sty1 (also known astion, even though it is expressed at a similar level (Fig-Spc1) MAP kinase, a member of the p38 MAP kinaseures 1A, 1B, and 1C). Cells arrested by overexpressionfamily, has been found to link the cell cycle to the extra-of Srk1 were stained with DAPI (Figure 1D) and subjectedcellular environment (Millar et al., 1995; Shiozaki andto FACScan analysis (our unpublished data), whichRussell, 1995). A number of effectors of the Sty1 MAPshowed that Srk1 arrests cells with a single nucleus andkinase have been identified including the Atf1 and Pap1a 2n DNA content, indicative of cell cycle block in latetranscription factors, which are homologs of mammalianG2 phase.ATF-2 and c-Jun, respectively (Shiozaki and Russell,
1996; Wilkinson et al., 1996). In addition, Sty1 binds andphosphorylates two downstream kinases, Cmk2 and Overexpression of Srk1 Prevents TyrosineSrk1 (Sty1-regulated kinase), which are related to the Dephosphorylation and Activation of Cdc2mammalian calmodulin-dependent and MAPKAP ki- The effect of Srk1 overexpression was examined innases and to the budding yeast RCK1 and RCK2 ki- cdc2-3w cells, which are insensitive to inactivation ofnases, which were identified as suppressors of a fission Cdc25, to examine how Srk1 inhibits entry into mitosis.yeast checkpoint mutant (Dahlkvist et al., 1995; Alemany In these cells, overexpression of Srk1 failed to induceet al., 2002; Sanchez-Piris et al., 2002; Smith et al., 2002). cell cycle arrest or substantial cell elongation (FiguresSrk1 is of particular interest, because it is also transcrip- 2A and 2B). This suggests that Srk1 inhibits activation oftionally upregulated after activation of the Sty1 MAP Cdc2 by inhibiting the Cdc25 phosphatase. Consistentkinase pathway (Smith et al., 2002). In this report, we with this, we find that cdc2-3w cdc25� cells are com-demonstrate that Srk1 controls mitotic entry by directly pletely insensitive to Srk1 overexpression (Figures 2Aphosphorylating and inhibiting Cdc25 during a normal and 2B). It is unlikely that Srk1 directly phosphorylates
Cdc2 on Tyr15 or activates the Wee1 or Mik1 kinases,cell cycle and in response to osmotic stress.
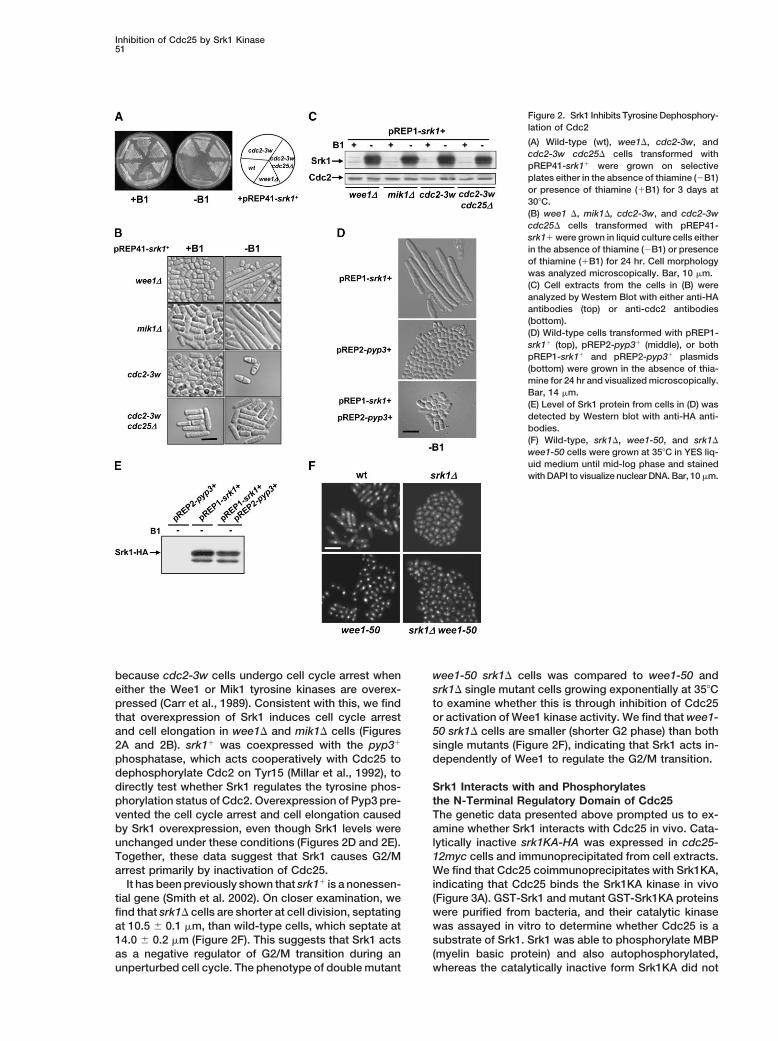
Inhibition of Cdc25 by Srk1 Kinase51
Figure 2. Srk1 Inhibits Tyrosine Dephosphory-lation of Cdc2
(A) Wild-type (wt), wee1�, cdc2-3w, andcdc2-3w cdc25� cells transformed withpREP41-srk1� were grown on selectiveplates either in the absence of thiamine (�B1)or presence of thiamine (�B1) for 3 days at30�C.(B) wee1 �, mik1�, cdc2-3w, and cdc2-3wcdc25� cells transformed with pREP41-srk1� were grown in liquid culture cells eitherin the absence of thiamine (�B1) or presenceof thiamine (�B1) for 24 hr. Cell morphologywas analyzed microscopically. Bar, 10 �m.(C) Cell extracts from the cells in (B) wereanalyzed by Western Blot with either anti-HAantibodies (top) or anti-cdc2 antibodies(bottom).(D) Wild-type cells transformed with pREP1-srk1� (top), pREP2-pyp3� (middle), or bothpREP1-srk1� and pREP2-pyp3� plasmids(bottom) were grown in the absence of thia-mine for 24 hr and visualized microscopically.Bar, 14 �m.(E) Level of Srk1 protein from cells in (D) wasdetected by Western blot with anti-HA anti-bodies.(F) Wild-type, srk1�, wee1-50, and srk1�
wee1-50 cells were grown at 35�C in YES liq-uid medium until mid-log phase and stainedwith DAPI to visualize nuclear DNA. Bar, 10 �m.
because cdc2-3w cells undergo cell cycle arrest when wee1-50 srk1� cells was compared to wee1-50 andsrk1� single mutant cells growing exponentially at 35�Ceither the Wee1 or Mik1 tyrosine kinases are overex-
pressed (Carr et al., 1989). Consistent with this, we find to examine whether this is through inhibition of Cdc25or activation of Wee1 kinase activity. We find that wee1-that overexpression of Srk1 induces cell cycle arrest
and cell elongation in wee1� and mik1� cells (Figures 50 srk1� cells are smaller (shorter G2 phase) than bothsingle mutants (Figure 2F), indicating that Srk1 acts in-2A and 2B). srk1� was coexpressed with the pyp3�
phosphatase, which acts cooperatively with Cdc25 to dependently of Wee1 to regulate the G2/M transition.dephosphorylate Cdc2 on Tyr15 (Millar et al., 1992), todirectly test whether Srk1 regulates the tyrosine phos- Srk1 Interacts with and Phosphorylates
the N-Terminal Regulatory Domain of Cdc25phorylation status of Cdc2. Overexpression of Pyp3 pre-vented the cell cycle arrest and cell elongation caused The genetic data presented above prompted us to ex-
amine whether Srk1 interacts with Cdc25 in vivo. Cata-by Srk1 overexpression, even though Srk1 levels wereunchanged under these conditions (Figures 2D and 2E). lytically inactive srk1KA-HA was expressed in cdc25-
12myc cells and immunoprecipitated from cell extracts.Together, these data suggest that Srk1 causes G2/Marrest primarily by inactivation of Cdc25. We find that Cdc25 coimmunoprecipitates with Srk1KA,
indicating that Cdc25 binds the Srk1KA kinase in vivoIt has been previously shown that srk1� is a nonessen-tial gene (Smith et al. 2002). On closer examination, we (Figure 3A). GST-Srk1 and mutant GST-Srk1KA proteins
were purified from bacteria, and their catalytic kinasefind that srk1� cells are shorter at cell division, septatingat 10.5 � 0.1 �m, than wild-type cells, which septate at was assayed in vitro to determine whether Cdc25 is a
substrate of Srk1. Srk1 was able to phosphorylate MBP14.0 � 0.2 �m (Figure 2F). This suggests that Srk1 actsas a negative regulator of G2/M transition during an (myelin basic protein) and also autophosphorylated,
whereas the catalytically inactive form Srk1KA did notunperturbed cell cycle. The phenotype of double mutant
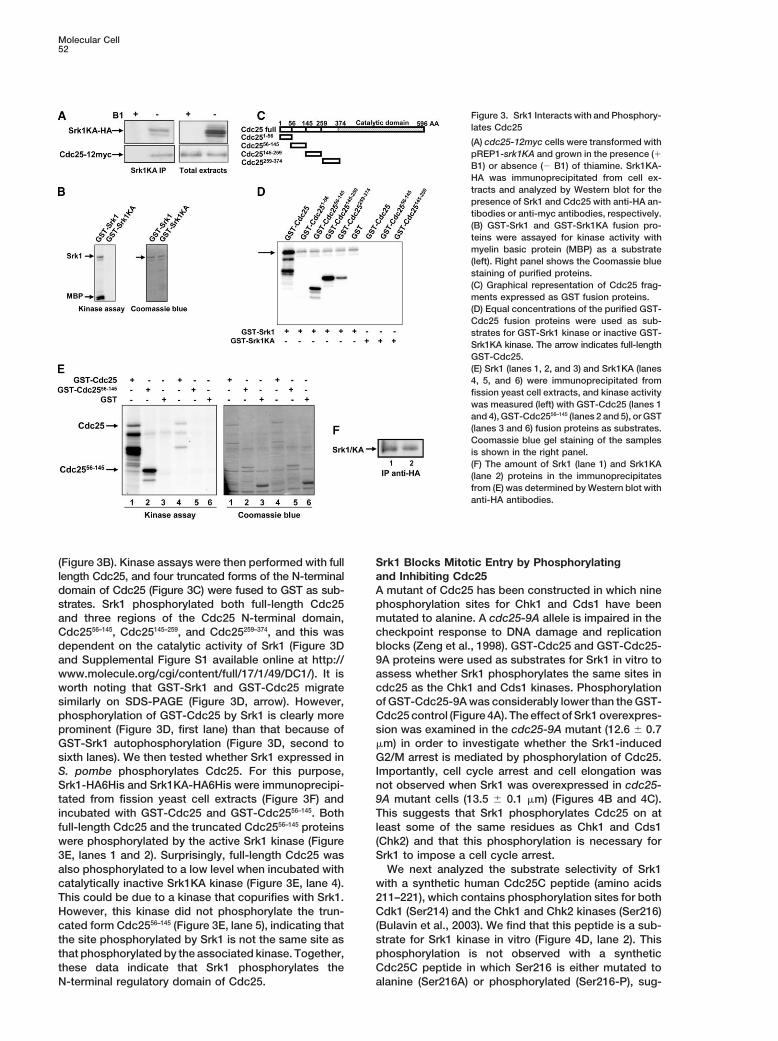
Molecular Cell52
Figure 3. Srk1 Interacts with and Phosphory-lates Cdc25
(A) cdc25-12myc cells were transformed withpREP1-srk1KA and grown in the presence (�B1) or absence (� B1) of thiamine. Srk1KA-HA was immunoprecipitated from cell ex-tracts and analyzed by Western blot for thepresence of Srk1 and Cdc25 with anti-HA an-tibodies or anti-myc antibodies, respectively.(B) GST-Srk1 and GST-Srk1KA fusion pro-teins were assayed for kinase activity withmyelin basic protein (MBP) as a substrate(left). Right panel shows the Coomassie bluestaining of purified proteins.(C) Graphical representation of Cdc25 frag-ments expressed as GST fusion proteins.(D) Equal concentrations of the purified GST-Cdc25 fusion proteins were used as sub-strates for GST-Srk1 kinase or inactive GST-Srk1KA kinase. The arrow indicates full-lengthGST-Cdc25.(E) Srk1 (lanes 1, 2, and 3) and Srk1KA (lanes4, 5, and 6) were immunoprecipitated fromfission yeast cell extracts, and kinase activitywas measured (left) with GST-Cdc25 (lanes 1and 4), GST-Cdc2556–145 (lanes 2 and 5), or GST(lanes 3 and 6) fusion proteins as substrates.Coomassie blue gel staining of the samplesis shown in the right panel.(F) The amount of Srk1 (lane 1) and Srk1KA(lane 2) proteins in the immunoprecipitatesfrom (E) was determined by Western blot withanti-HA antibodies.
(Figure 3B). Kinase assays were then performed with full Srk1 Blocks Mitotic Entry by Phosphorylatingand Inhibiting Cdc25length Cdc25, and four truncated forms of the N-terminal
domain of Cdc25 (Figure 3C) were fused to GST as sub- A mutant of Cdc25 has been constructed in which ninephosphorylation sites for Chk1 and Cds1 have beenstrates. Srk1 phosphorylated both full-length Cdc25
and three regions of the Cdc25 N-terminal domain, mutated to alanine. A cdc25-9A allele is impaired in thecheckpoint response to DNA damage and replicationCdc2556–145, Cdc25145–259, and Cdc25259–374, and this was
dependent on the catalytic activity of Srk1 (Figure 3D blocks (Zeng et al., 1998). GST-Cdc25 and GST-Cdc25-9A proteins were used as substrates for Srk1 in vitro toand Supplemental Figure S1 available online at http://
www.molecule.org/cgi/content/full/17/1/49/DC1/). It is assess whether Srk1 phosphorylates the same sites incdc25 as the Chk1 and Cds1 kinases. Phosphorylationworth noting that GST-Srk1 and GST-Cdc25 migrate
similarly on SDS-PAGE (Figure 3D, arrow). However, of GST-Cdc25-9A was considerably lower than the GST-Cdc25 control (Figure 4A). The effect of Srk1 overexpres-phosphorylation of GST-Cdc25 by Srk1 is clearly more
prominent (Figure 3D, first lane) than that because of sion was examined in the cdc25-9A mutant (12.6 � 0.7�m) in order to investigate whether the Srk1-inducedGST-Srk1 autophosphorylation (Figure 3D, second to
sixth lanes). We then tested whether Srk1 expressed in G2/M arrest is mediated by phosphorylation of Cdc25.Importantly, cell cycle arrest and cell elongation wasS. pombe phosphorylates Cdc25. For this purpose,
Srk1-HA6His and Srk1KA-HA6His were immunoprecipi- not observed when Srk1 was overexpressed in cdc25-9A mutant cells (13.5 � 0.1 �m) (Figures 4B and 4C).tated from fission yeast cell extracts (Figure 3F) and
incubated with GST-Cdc25 and GST-Cdc2556–145. Both This suggests that Srk1 phosphorylates Cdc25 on atleast some of the same residues as Chk1 and Cds1full-length Cdc25 and the truncated Cdc2556–145 proteins
were phosphorylated by the active Srk1 kinase (Figure (Chk2) and that this phosphorylation is necessary forSrk1 to impose a cell cycle arrest.3E, lanes 1 and 2). Surprisingly, full-length Cdc25 was
also phosphorylated to a low level when incubated with We next analyzed the substrate selectivity of Srk1with a synthetic human Cdc25C peptide (amino acidscatalytically inactive Srk1KA kinase (Figure 3E, lane 4).
This could be due to a kinase that copurifies with Srk1. 211–221), which contains phosphorylation sites for bothCdk1 (Ser214) and the Chk1 and Chk2 kinases (Ser216)However, this kinase did not phosphorylate the trun-
cated form Cdc2556–145 (Figure 3E, lane 5), indicating that (Bulavin et al., 2003). We find that this peptide is a sub-strate for Srk1 kinase in vitro (Figure 4D, lane 2). Thisthe site phosphorylated by Srk1 is not the same site as
that phosphorylated by the associated kinase. Together, phosphorylation is not observed with a syntheticCdc25C peptide in which Ser216 is either mutated tothese data indicate that Srk1 phosphorylates the
N-terminal regulatory domain of Cdc25. alanine (Ser216A) or phosphorylated (Ser216-P), sug-
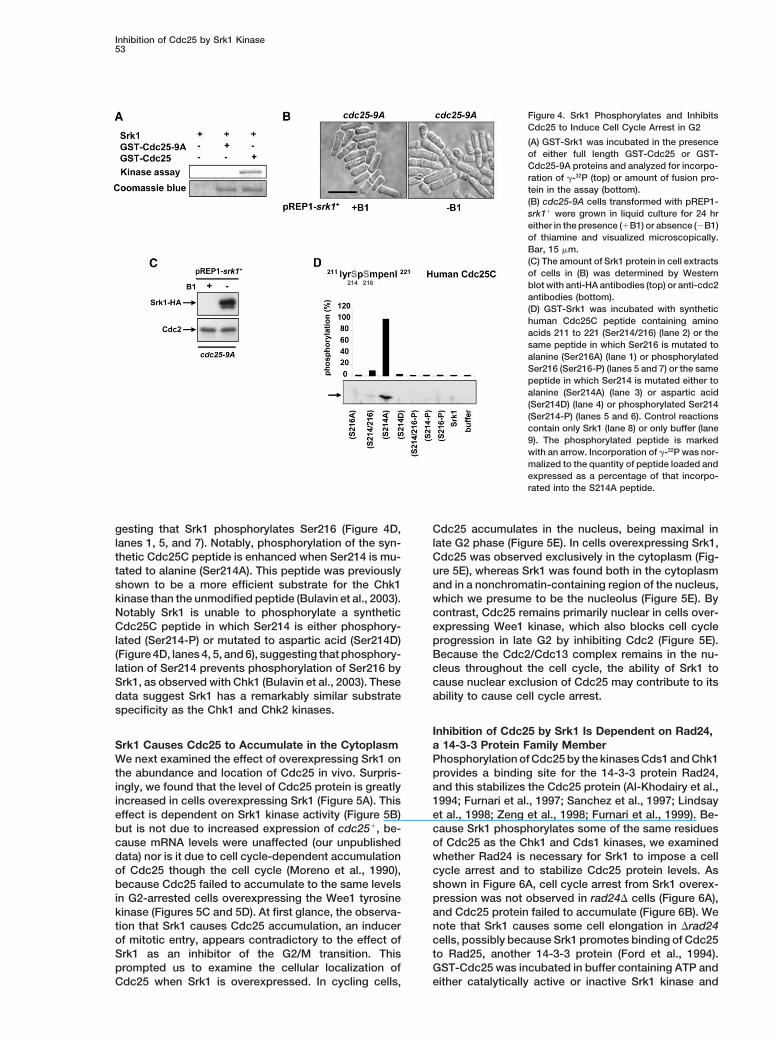
Inhibition of Cdc25 by Srk1 Kinase53
Figure 4. Srk1 Phosphorylates and InhibitsCdc25 to Induce Cell Cycle Arrest in G2
(A) GST-Srk1 was incubated in the presenceof either full length GST-Cdc25 or GST-Cdc25-9A proteins and analyzed for incorpo-ration of �-32P (top) or amount of fusion pro-tein in the assay (bottom).(B) cdc25-9A cells transformed with pREP1-srk1� were grown in liquid culture for 24 hreither in the presence (�B1) or absence (�B1)of thiamine and visualized microscopically.Bar, 15 �m.(C) The amount of Srk1 protein in cell extractsof cells in (B) was determined by Westernblot with anti-HA antibodies (top) or anti-cdc2antibodies (bottom).(D) GST-Srk1 was incubated with synthetichuman Cdc25C peptide containing aminoacids 211 to 221 (Ser214/216) (lane 2) or thesame peptide in which Ser216 is mutated toalanine (Ser216A) (lane 1) or phosphorylatedSer216 (Ser216-P) (lanes 5 and 7) or the samepeptide in which Ser214 is mutated either toalanine (Ser214A) (lane 3) or aspartic acid(Ser214D) (lane 4) or phosphorylated Ser214(Ser214-P) (lanes 5 and 6). Control reactionscontain only Srk1 (lane 8) or only buffer (lane9). The phosphorylated peptide is markedwith an arrow. Incorporation of �-32P was nor-malized to the quantity of peptide loaded andexpressed as a percentage of that incorpo-rated into the S214A peptide.
gesting that Srk1 phosphorylates Ser216 (Figure 4D, Cdc25 accumulates in the nucleus, being maximal inlate G2 phase (Figure 5E). In cells overexpressing Srk1,lanes 1, 5, and 7). Notably, phosphorylation of the syn-
thetic Cdc25C peptide is enhanced when Ser214 is mu- Cdc25 was observed exclusively in the cytoplasm (Fig-ure 5E), whereas Srk1 was found both in the cytoplasmtated to alanine (Ser214A). This peptide was previously
shown to be a more efficient substrate for the Chk1 and in a nonchromatin-containing region of the nucleus,which we presume to be the nucleolus (Figure 5E). Bykinase than the unmodified peptide (Bulavin et al., 2003).
Notably Srk1 is unable to phosphorylate a synthetic contrast, Cdc25 remains primarily nuclear in cells over-expressing Wee1 kinase, which also blocks cell cycleCdc25C peptide in which Ser214 is either phosphory-
lated (Ser214-P) or mutated to aspartic acid (Ser214D) progression in late G2 by inhibiting Cdc2 (Figure 5E).Because the Cdc2/Cdc13 complex remains in the nu-(Figure 4D, lanes 4, 5, and 6), suggesting that phosphory-
lation of Ser214 prevents phosphorylation of Ser216 by cleus throughout the cell cycle, the ability of Srk1 tocause nuclear exclusion of Cdc25 may contribute to itsSrk1, as observed with Chk1 (Bulavin et al., 2003). These
data suggest Srk1 has a remarkably similar substrate ability to cause cell cycle arrest.specificity as the Chk1 and Chk2 kinases.
Inhibition of Cdc25 by Srk1 Is Dependent on Rad24,a 14-3-3 Protein Family MemberSrk1 Causes Cdc25 to Accumulate in the Cytoplasm
We next examined the effect of overexpressing Srk1 on Phosphorylation of Cdc25 by the kinases Cds1 and Chk1provides a binding site for the 14-3-3 protein Rad24,the abundance and location of Cdc25 in vivo. Surpris-
ingly, we found that the level of Cdc25 protein is greatly and this stabilizes the Cdc25 protein (Al-Khodairy et al.,1994; Furnari et al., 1997; Sanchez et al., 1997; Lindsayincreased in cells overexpressing Srk1 (Figure 5A). This
effect is dependent on Srk1 kinase activity (Figure 5B) et al., 1998; Zeng et al., 1998; Furnari et al., 1999). Be-cause Srk1 phosphorylates some of the same residuesbut is not due to increased expression of cdc25�, be-
cause mRNA levels were unaffected (our unpublished of Cdc25 as the Chk1 and Cds1 kinases, we examinedwhether Rad24 is necessary for Srk1 to impose a celldata) nor is it due to cell cycle-dependent accumulation
of Cdc25 though the cell cycle (Moreno et al., 1990), cycle arrest and to stabilize Cdc25 protein levels. Asshown in Figure 6A, cell cycle arrest from Srk1 overex-because Cdc25 failed to accumulate to the same levels
in G2-arrested cells overexpressing the Wee1 tyrosine pression was not observed in rad24� cells (Figure 6A),and Cdc25 protein failed to accumulate (Figure 6B). Wekinase (Figures 5C and 5D). At first glance, the observa-
tion that Srk1 causes Cdc25 accumulation, an inducer note that Srk1 causes some cell elongation in �rad24cells, possibly because Srk1 promotes binding of Cdc25of mitotic entry, appears contradictory to the effect of
Srk1 as an inhibitor of the G2/M transition. This to Rad25, another 14-3-3 protein (Ford et al., 1994).GST-Cdc25 was incubated in buffer containing ATP andprompted us to examine the cellular localization of
Cdc25 when Srk1 is overexpressed. In cycling cells, either catalytically active or inactive Srk1 kinase and
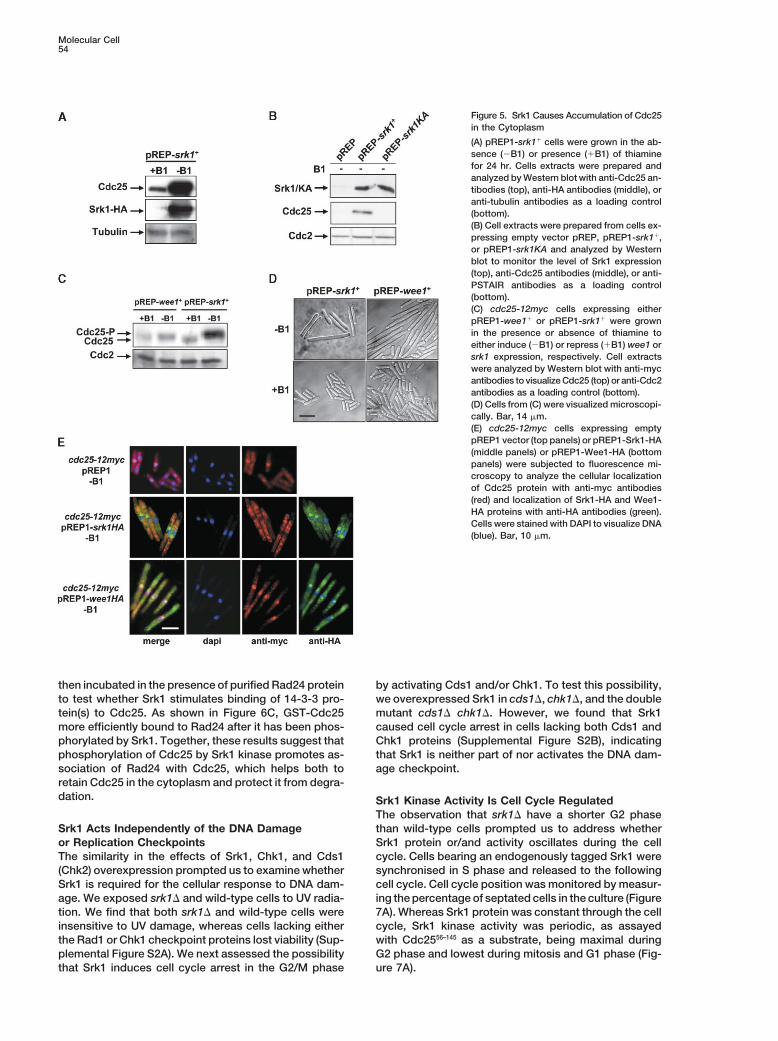
Molecular Cell54
Figure 5. Srk1 Causes Accumulation of Cdc25in the Cytoplasm
(A) pREP1-srk1� cells were grown in the ab-sence (�B1) or presence (�B1) of thiaminefor 24 hr. Cells extracts were prepared andanalyzed by Western blot with anti-Cdc25 an-tibodies (top), anti-HA antibodies (middle), oranti-tubulin antibodies as a loading control(bottom).(B) Cell extracts were prepared from cells ex-pressing empty vector pREP, pREP1-srk1�,or pREP1-srk1KA and analyzed by Westernblot to monitor the level of Srk1 expression(top), anti-Cdc25 antibodies (middle), or anti-PSTAIR antibodies as a loading control(bottom).(C) cdc25-12myc cells expressing eitherpREP1-wee1� or pREP1-srk1� were grownin the presence or absence of thiamine toeither induce (�B1) or repress (�B1) wee1 orsrk1 expression, respectively. Cell extractswere analyzed by Western blot with anti-mycantibodies to visualize Cdc25 (top) or anti-Cdc2antibodies as a loading control (bottom).(D) Cells from (C) were visualized microscopi-cally. Bar, 14 �m.(E) cdc25-12myc cells expressing emptypREP1 vector (top panels) or pREP1-Srk1-HA(middle panels) or pREP1-Wee1-HA (bottompanels) were subjected to fluorescence mi-croscopy to analyze the cellular localizationof Cdc25 protein with anti-myc antibodies(red) and localization of Srk1-HA and Wee1-HA proteins with anti-HA antibodies (green).Cells were stained with DAPI to visualize DNA(blue). Bar, 10 �m.
then incubated in the presence of purified Rad24 protein by activating Cds1 and/or Chk1. To test this possibility,we overexpressed Srk1 in cds1�, chk1�, and the doubleto test whether Srk1 stimulates binding of 14-3-3 pro-
tein(s) to Cdc25. As shown in Figure 6C, GST-Cdc25 mutant cds1� chk1�. However, we found that Srk1caused cell cycle arrest in cells lacking both Cds1 andmore efficiently bound to Rad24 after it has been phos-
phorylated by Srk1. Together, these results suggest that Chk1 proteins (Supplemental Figure S2B), indicatingthat Srk1 is neither part of nor activates the DNA dam-phosphorylation of Cdc25 by Srk1 kinase promotes as-
sociation of Rad24 with Cdc25, which helps both to age checkpoint.retain Cdc25 in the cytoplasm and protect it from degra-dation. Srk1 Kinase Activity Is Cell Cycle Regulated
The observation that srk1� have a shorter G2 phasethan wild-type cells prompted us to address whetherSrk1 Acts Independently of the DNA Damage
or Replication Checkpoints Srk1 protein or/and activity oscillates during the cellcycle. Cells bearing an endogenously tagged Srk1 wereThe similarity in the effects of Srk1, Chk1, and Cds1
(Chk2) overexpression prompted us to examine whether synchronised in S phase and released to the followingcell cycle. Cell cycle position was monitored by measur-Srk1 is required for the cellular response to DNA dam-
age. We exposed srk1� and wild-type cells to UV radia- ing the percentage of septated cells in the culture (Figure7A). Whereas Srk1 protein was constant through the celltion. We find that both srk1� and wild-type cells were
insensitive to UV damage, whereas cells lacking either cycle, Srk1 kinase activity was periodic, as assayedwith Cdc2556–145 as a substrate, being maximal duringthe Rad1 or Chk1 checkpoint proteins lost viability (Sup-
plemental Figure S2A). We next assessed the possibility G2 phase and lowest during mitosis and G1 phase (Fig-ure 7A).that Srk1 induces cell cycle arrest in the G2/M phase
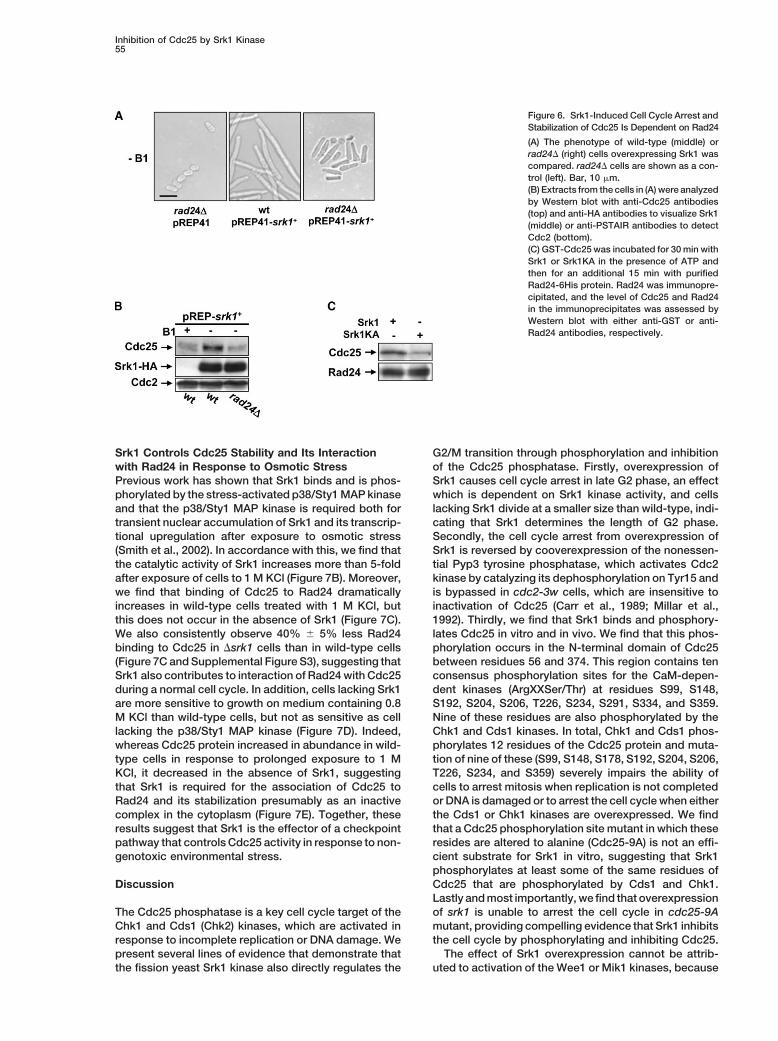
Inhibition of Cdc25 by Srk1 Kinase55
Figure 6. Srk1-Induced Cell Cycle Arrest andStabilization of Cdc25 Is Dependent on Rad24
(A) The phenotype of wild-type (middle) orrad24� (right) cells overexpressing Srk1 wascompared. rad24� cells are shown as a con-trol (left). Bar, 10 �m.(B) Extracts from the cells in (A) were analyzedby Western blot with anti-Cdc25 antibodies(top) and anti-HA antibodies to visualize Srk1(middle) or anti-PSTAIR antibodies to detectCdc2 (bottom).(C) GST-Cdc25 was incubated for 30 min withSrk1 or Srk1KA in the presence of ATP andthen for an additional 15 min with purifiedRad24-6His protein. Rad24 was immunopre-cipitated, and the level of Cdc25 and Rad24in the immunoprecipitates was assessed byWestern blot with either anti-GST or anti-Rad24 antibodies, respectively.
Srk1 Controls Cdc25 Stability and Its Interaction G2/M transition through phosphorylation and inhibitionof the Cdc25 phosphatase. Firstly, overexpression ofwith Rad24 in Response to Osmotic Stress
Previous work has shown that Srk1 binds and is phos- Srk1 causes cell cycle arrest in late G2 phase, an effectwhich is dependent on Srk1 kinase activity, and cellsphorylated by the stress-activated p38/Sty1 MAP kinase
and that the p38/Sty1 MAP kinase is required both for lacking Srk1 divide at a smaller size than wild-type, indi-cating that Srk1 determines the length of G2 phase.transient nuclear accumulation of Srk1 and its transcrip-
tional upregulation after exposure to osmotic stress Secondly, the cell cycle arrest from overexpression ofSrk1 is reversed by cooverexpression of the nonessen-(Smith et al., 2002). In accordance with this, we find that
the catalytic activity of Srk1 increases more than 5-fold tial Pyp3 tyrosine phosphatase, which activates Cdc2kinase by catalyzing its dephosphorylation on Tyr15 andafter exposure of cells to 1 M KCl (Figure 7B). Moreover,
we find that binding of Cdc25 to Rad24 dramatically is bypassed in cdc2-3w cells, which are insensitive toinactivation of Cdc25 (Carr et al., 1989; Millar et al.,increases in wild-type cells treated with 1 M KCl, but
this does not occur in the absence of Srk1 (Figure 7C). 1992). Thirdly, we find that Srk1 binds and phosphory-lates Cdc25 in vitro and in vivo. We find that this phos-We also consistently observe 40% � 5% less Rad24
binding to Cdc25 in �srk1 cells than in wild-type cells phorylation occurs in the N-terminal domain of Cdc25between residues 56 and 374. This region contains ten(Figure 7C and Supplemental Figure S3), suggesting that
Srk1 also contributes to interaction of Rad24 with Cdc25 consensus phosphorylation sites for the CaM-depen-dent kinases (ArgXXSer/Thr) at residues S99, S148,during a normal cell cycle. In addition, cells lacking Srk1
are more sensitive to growth on medium containing 0.8 S192, S204, S206, T226, S234, S291, S334, and S359.Nine of these residues are also phosphorylated by theM KCl than wild-type cells, but not as sensitive as cell
lacking the p38/Sty1 MAP kinase (Figure 7D). Indeed, Chk1 and Cds1 kinases. In total, Chk1 and Cds1 phos-phorylates 12 residues of the Cdc25 protein and muta-whereas Cdc25 protein increased in abundance in wild-
type cells in response to prolonged exposure to 1 M tion of nine of these (S99, S148, S178, S192, S204, S206,T226, S234, and S359) severely impairs the ability ofKCl, it decreased in the absence of Srk1, suggesting
that Srk1 is required for the association of Cdc25 to cells to arrest mitosis when replication is not completedor DNA is damaged or to arrest the cell cycle when eitherRad24 and its stabilization presumably as an inactive
complex in the cytoplasm (Figure 7E). Together, these the Cds1 or Chk1 kinases are overexpressed. We findthat a Cdc25 phosphorylation site mutant in which theseresults suggest that Srk1 is the effector of a checkpoint
pathway that controls Cdc25 activity in response to non- resides are altered to alanine (Cdc25-9A) is not an effi-cient substrate for Srk1 in vitro, suggesting that Srk1genotoxic environmental stress.phosphorylates at least some of the same residues ofCdc25 that are phosphorylated by Cds1 and Chk1.DiscussionLastly and most importantly, we find that overexpressionof srk1 is unable to arrest the cell cycle in cdc25-9AThe Cdc25 phosphatase is a key cell cycle target of the
Chk1 and Cds1 (Chk2) kinases, which are activated in mutant, providing compelling evidence that Srk1 inhibitsthe cell cycle by phosphorylating and inhibiting Cdc25.response to incomplete replication or DNA damage. We
present several lines of evidence that demonstrate that The effect of Srk1 overexpression cannot be attrib-uted to activation of the Wee1 or Mik1 kinases, becausethe fission yeast Srk1 kinase also directly regulates the
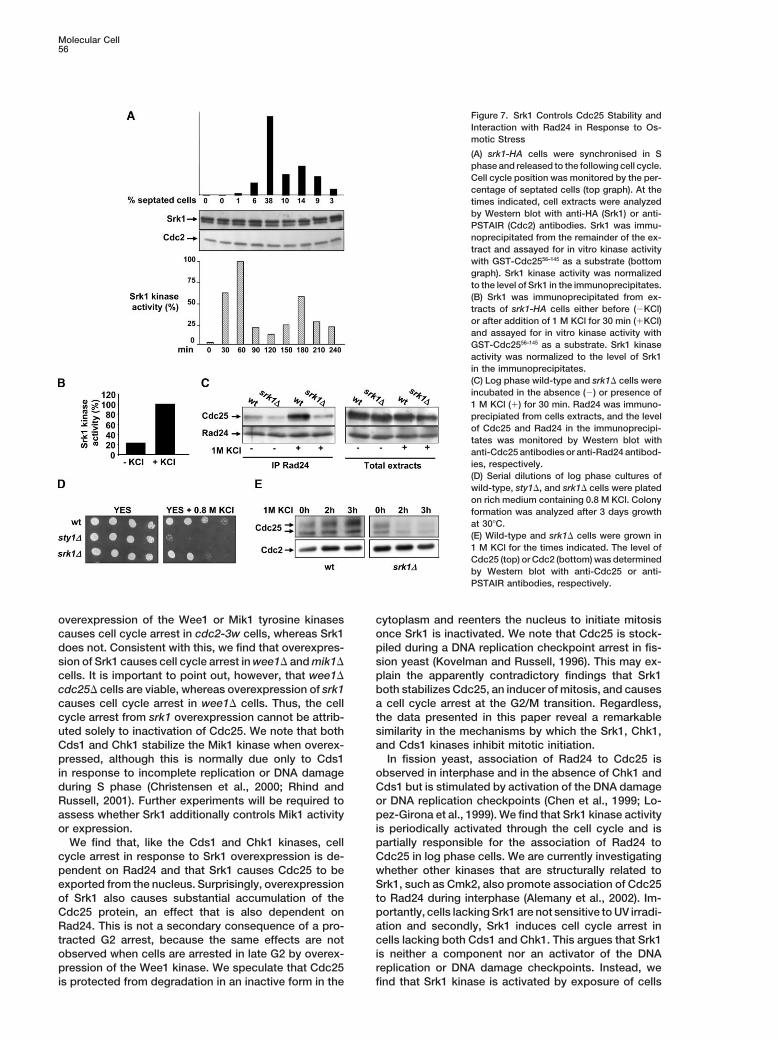
Molecular Cell56
Figure 7. Srk1 Controls Cdc25 Stability andInteraction with Rad24 in Response to Os-motic Stress
(A) srk1-HA cells were synchronised in Sphase and released to the following cell cycle.Cell cycle position was monitored by the per-centage of septated cells (top graph). At thetimes indicated, cell extracts were analyzedby Western blot with anti-HA (Srk1) or anti-PSTAIR (Cdc2) antibodies. Srk1 was immu-noprecipitated from the remainder of the ex-tract and assayed for in vitro kinase activitywith GST-Cdc2556–145 as a substrate (bottomgraph). Srk1 kinase activity was normalizedto the level of Srk1 in the immunoprecipitates.(B) Srk1 was immunoprecipitated from ex-tracts of srk1-HA cells either before (�KCl)or after addition of 1 M KCl for 30 min (�KCl)and assayed for in vitro kinase activity withGST-Cdc2556–145 as a substrate. Srk1 kinaseactivity was normalized to the level of Srk1in the immunoprecipitates.(C) Log phase wild-type and srk1� cells wereincubated in the absence (�) or presence of1 M KCl (�) for 30 min. Rad24 was immuno-precipiated from cells extracts, and the levelof Cdc25 and Rad24 in the immunoprecipi-tates was monitored by Western blot withanti-Cdc25 antibodies or anti-Rad24 antibod-ies, respectively.(D) Serial dilutions of log phase cultures ofwild-type, sty1�, and srk1� cells were platedon rich medium containing 0.8 M KCl. Colonyformation was analyzed after 3 days growthat 30�C.(E) Wild-type and srk1� cells were grown in1 M KCl for the times indicated. The level ofCdc25 (top) or Cdc2 (bottom) was determinedby Western blot with anti-Cdc25 or anti-PSTAIR antibodies, respectively.
overexpression of the Wee1 or Mik1 tyrosine kinases cytoplasm and reenters the nucleus to initiate mitosisonce Srk1 is inactivated. We note that Cdc25 is stock-causes cell cycle arrest in cdc2-3w cells, whereas Srk1
does not. Consistent with this, we find that overexpres- piled during a DNA replication checkpoint arrest in fis-sion yeast (Kovelman and Russell, 1996). This may ex-sion of Srk1 causes cell cycle arrest in wee1� and mik1�
cells. It is important to point out, however, that wee1� plain the apparently contradictory findings that Srk1both stabilizes Cdc25, an inducer of mitosis, and causescdc25� cells are viable, whereas overexpression of srk1
causes cell cycle arrest in wee1� cells. Thus, the cell a cell cycle arrest at the G2/M transition. Regardless,the data presented in this paper reveal a remarkablecycle arrest from srk1 overexpression cannot be attrib-
uted solely to inactivation of Cdc25. We note that both similarity in the mechanisms by which the Srk1, Chk1,and Cds1 kinases inhibit mitotic initiation.Cds1 and Chk1 stabilize the Mik1 kinase when overex-
pressed, although this is normally due only to Cds1 In fission yeast, association of Rad24 to Cdc25 isobserved in interphase and in the absence of Chk1 andin response to incomplete replication or DNA damage
during S phase (Christensen et al., 2000; Rhind and Cds1 but is stimulated by activation of the DNA damageor DNA replication checkpoints (Chen et al., 1999; Lo-Russell, 2001). Further experiments will be required to
assess whether Srk1 additionally controls Mik1 activity pez-Girona et al., 1999). We find that Srk1 kinase activityis periodically activated through the cell cycle and isor expression.
We find that, like the Cds1 and Chk1 kinases, cell partially responsible for the association of Rad24 toCdc25 in log phase cells. We are currently investigatingcycle arrest in response to Srk1 overexpression is de-
pendent on Rad24 and that Srk1 causes Cdc25 to be whether other kinases that are structurally related toSrk1, such as Cmk2, also promote association of Cdc25exported from the nucleus. Surprisingly, overexpression
of Srk1 also causes substantial accumulation of the to Rad24 during interphase (Alemany et al., 2002). Im-portantly, cells lacking Srk1 are not sensitive to UV irradi-Cdc25 protein, an effect that is also dependent on
Rad24. This is not a secondary consequence of a pro- ation and secondly, Srk1 induces cell cycle arrest incells lacking both Cds1 and Chk1. This argues that Srk1tracted G2 arrest, because the same effects are not
observed when cells are arrested in late G2 by overex- is neither a component nor an activator of the DNAreplication or DNA damage checkpoints. Instead, wepression of the Wee1 kinase. We speculate that Cdc25
is protected from degradation in an inactive form in the find that Srk1 kinase is activated by exposure of cells

Inhibition of Cdc25 by Srk1 Kinase57
residue lysine 153 to alanine was performed by PCR with overlap-to osmotic stress and that Srk1 is required for viabilityping oligonucleotides to construct srk1KA. srk1� and srk1KA wereunder these conditions. Moreover, we find that in re-cloned into the bacterial expression plasmid pGEX-KG, which allowssponse to prolonged exposure to osmotic stress Srk1the expression of GST fused proteins in E. coli. An srk1::kanMX6
promotes the association of Cdc25 to Rad24 and causes strain was constructed by PCR-amplification with srk1�-specificCdc25 stabilization, presumably as an inactive complex primers and plasmid pFA6a-kanMX6 as a template. Recombinants
were selected by their ability to grow on G418/geneticin (Invitrogen),in the cytoplasm. This suggests that Srk1 is the effectoras described previously (Bahler et al. 1998).of a signal transduction pathway that inhibits Cdc25 in
response to nongenotoxic environmental stress.In fission yeast, Srk1 binds the stress-activated Sty1 Expression and Purification of GST Fusion Proteins
The GST fusion proteins were expressed in E. coli BL21(D3)pLysMAP kinase, a member of the p38 MAP kinase familycells. Log phase cultures grown at 37�C were treated with 0.5 mM(Smith et al., 2002). Srk1 is not only complexed withisopropyl-s-thio-D-galactoside (IPTG) and grown for additional 4 hrand phosphorylated by Sty1 but is also transcriptionallyat 25�C. Cells were harvested and lysed in NETN buffer (20 mM Tris-
induced in a manner dependent on activation of Sty1. HCl [pH 8.0], 100 mM NaCl, 1 mM EDTA, 0.5% NP-40, 20 �g/mlGenetic experiments suggest a role for the Sty1 MAP aprotinin, 40 �g/ml leupeptin, 20 �g/ml pepstatin A, and 1 mMkinase pathway in controlling cell cycle arrest, adapta- PMSF). GST-proteins were purified by adsorption to glutathione-
Sepharose beads (Amersham-Pharmacia) according to the manu-tion, and recovery in response to environmental stress.facturer’s instructions, and proteins were eluted into 50 mM TrisOur future experiments will address the role of Srk1 in(pH 8.5), 100 mM NaCl, 10 mM glutathione, and 2 mM DTT. Untaggedregulation of the cell cycle by the p38/Sty1 MAP kinaseSrk1 and Srk1KA proteins were cleaved from GST fusion proteins
pathway. In mammalian cells, the G2/M transition can be bound to GST Sepharose beads by digestion with thrombin (10delayed by activation of the p38 MAP kinase pathway, in units/mg protein) (Amersham-Biosciences).a process known as the antephase checkpoint (Wanget al., 2000; Bulavin et al., 2001; Matsusaka and Pines, Immunoprecipitation and Western Blotting2004; Mikhailov et al., 2004). The p38 MAP kinase has 1 � 108 cells were lysed in buffer (150 mM NaCl, 50 mM Tris-HClbeen reported to bind and phosphorylate Cdc25B at [pH 8.0], 5 mM EDTA, 0.1% Triton X-100, 10% glycerol, 50 mM NaF,
1 mM PMSF, 1 mM NaVO4, 5 �g/ml aprotinin, 5 �g/ml leupeptin).Ser309 and Ser361 and Cdc25C at Ser216. This is curi-Proteins were immunoprecipitated from cell extracts with eitherous, because MAP kinases are exclusively proline di-protein A or protein G Sepharose beads, and immunoprecipitatesrected, and none of these serines is followed by a prolinewere washed four times in lysis buffer prior to analysis. Proteins
at the �1 position (Gonzalez et al., 1991). Because the were resolved by SDS-polyacrylamide gel electrophoresis (SDS-activated p38 MAP kinase used to phosphorylate Cdc25 PAGE) and analyzed by Western blotting. The following primaryin vitro in these studies was immunoprecipitated from antibodies were used: monoclonal anti-HA (12CA5) from ascites
fluid (1/1000), monoclonal y anti- tubulin (Sigma) (1/500), polyclonalcell lysates, it is possible that a kinase that associatesanti-Cdc25 (1/1000), polyclonal anti-PSTAIR (Upstate Biotechnol-with p38 phosphorylates Cdc25B and Cdc25C at theseogy) (1/2000), polyclonal anti-rad24 (1/10000), and monoclonal anti-residues (Bulavin et al., 2001). In this paper, we show thatGST (Santa Cruz) (1/2000). Horseradish peroxidase-conjugated anti-
bacterially expressed Srk1 phosphorylate a synthetic mouse or anti-mouse antibodies (Bio Rad) were used as secondarypeptide corresponding to amino acids 211–222 of hu- antibodies. Membranes were developed by enhanced chemilumi-man Cdc25C on Ser216, whereas activated Sty1 MAP nescence (ECL kit, Amersham-Pharmacia).kinase purified from bacteria is unable to do so (ourunpublished data). However, Srk1 copurifies with a ki- In Vitro Kinase Assaysnase from cell extracts that phosphorylates full-length GST-Cdc25 fusion proteins were incubated in kinase buffer (50 mM
Tris-HCl [pH 8.0], 10 mM MgCl2, 5 mM -Mercaptoethanol, 50 �MCdc25 on distinct sites to that phosphorylated by Srk1.ATP, and 0.1 �Ci/�l [�-32P]ATP) and 350 nM GST-Srk1 or GST-It will be of interest to determine whether this copurifyingSrk1KA for 30 min at 30�C. Immunoprecipitates from pombe cellkinase is Sty1, and if so, what is the role of this phosphor-lysates were incubated in kinase buffer containing 0.5 �g/�l GST-
ylation. These data raise the possibility that in mamma- Cdc25 and GST-Cdc2556–145 for 30 min at 30�C. Labeled proteinslian cells, p38 MAPK imposes a G2 arrest by association were resolved by SDS-PAGE and detected by autoradiograph. Forto an as-yet-unidentified kinase that phosphorylates peptide phosphorylation, purified GST-Srk1 was incubated at 30�C
for 60 min in buffer containing 20 mM Tris-HCl [pH 7.5], 200 �MCdc25 rather than by direct phosphorylation of Cdc25ATP, 20 mM MgCl2, 5 mM DTT, 2 �Ci [�-32P]ATP and 200 �M peptide.by p38. We suggest that this kinase may be an effectorPhosphorylated peptides were separated by thin-layer chromatog-of the antephase checkpoint that regulates mitotic com-raphy (TLC) on cellulose plates (Merck) in n-butanol:pyridine:acetic
mitment in mammalian cells. acid:water (3:2:1:4).
Experimental ProceduresImmunofluorescence MicroscopyImmunofluorescence microscopy was performed by methanol fixa-Fission Yeast Strains, Media, and Techniquestion as described (http://pingu.salk.edu/forsburg/lab.html#info).The strains used are listed in Supplemental Table S1. Media andMonoclonal anti-myc (9E10) and rabbit affinity-purified anti-HA anti-genetic methods for studying S. pombe were as described by Mor-bodies (Sigma, H6908) were detected with fluor 488 and 594 conju-eno et al. (1991).gated goat anti-mouse and anti-rabbit antibodies, respectively (Mo-lecular Probes-Alexa). Cells were examined with a Leica JCS-NT
Plasmid and Strain Constructionconfocal laser-scanning microscope.
Plasmids pREP1-srk1(6HisHA), pREP41-srk1(6HisHA), and pREP81-srk1(6HisHA), containing the entire srk1� ORF under the induciblepromoters nmt1, nmt41, and nmt81 (Maundrell, 1993) fused to two Treatment with UV Irradiation
Log phase cells were UV-irradiated at 254 nm with a Stratalinkerhemagglutinin (HA) epitopes and a hexahistidine tag at the C-ter-minal end, were constructed by PCR amplification from genomic (Stratagene) at different J/m2. 1000 cells from each culture were
plated, and colony number was determined after 4 days of growthDNA. The srk1 locus was tagged by integration of pRIP2-srk1(6HisHA) to create an srk1-6HisHA:ura4 strain. Mutagenesis of at 30�C.

Molecular Cell58
Acknowledgments Furnari, B., Blasina, A., Boddy, M.N., McGowan, C.H., and Russell,P. (1999). Cdc25 inhibited in vivo and in vitro by checkpoint kinasesCds1 and Chk1. Mol. Biol. Cell 10, 833–845.We thank Paul Russell and Miguel Rodriguez-Gabriel for their helpful
advice and for gifts of plasmids and strains, Helen Piwnica-Worms Gabrielli, B.G., Clark, J.M., McCormack, A.K., and Ellem, K.A. (1997).and Pedro San-Segundo for providing pGEX-Cdc25-9A plasmid and Hyperphosphorylation of the N-terminal domain of Cdc25 regulatescdc25-9A strain and Dmitry Bulavin for providing the synthetic hu- activity toward cyclin B1/Cdc2 but not cyclin A/Cdk2. J. Biol. Chem.man Cdc25C peptides, and Nancy Walworth for providing Rad24 272, 28607–28614.polyclonal antisera and for her helpful comments. We thank Sergio Gautier, J., Solomon, M.J., Booher, R.N., Bazan, J.F., and Kirschner,Moreno for providing Cdc25 polyclonal antisera and Antonia Lopez- M.W. (1991). Cdc25 is a specific tyrosine phosphatase that directlyGirona and Elena Hidalgo for helpful comments. We are also grateful activates p34cdc2. Cell 67, 197–211.to Anna Bosch (Serveis Cientıfic-Tecnics) for excellent assistance
Goldstone, S., Pavey, S., Forrest, A., Sinnamon, J., and Gabrielli, B.in confocal microscopy. This work was supported by grants SAF(2001). Cdc25-dependent activation of cyclin A/cdk2 is blocked in2002-00452 and BMC2003-00408 from the Direccion General deG2 phase arrested cells independently of ATM/ATR. OncogeneInvestigacion Cientıfica y Tecnica (Ministry of Science and Technol-20, 921–932.ogy, Spain). S.L.-A. is the recipient of a Formacio InvestigadorsGonzalez, F.A., Raden, D.L., and Davis, R.J. (1991). Identificationpredoctoral fellowship from the Generalitat de Catalunya, Spain.of substrate recognition determinants for human ERK1 and ERK2protein kinases. J. Biol. Chem. 266, 22159–22163.Received: June 16, 2004Graves, P.R., Lovly, C.M., Uy, G.L., and Piwnica-Worms, H. (2001).Revised: October 1, 2004Localization of human Cdc25C is regulated both by nuclear exportAccepted: November 22, 2004and 14-3-3 protein binding. Oncogene 20, 1839–1851.Published: January 6, 2005Hoffmann, I., Clarke, P.R., Marcote, M.J., Karsenti, E., and Draetta,G. (1993). Phosphorylation and activation of human cdc25-C byReferencescdc2-cyclin B and its involvement in the self-amplification of MPFat mitosis. EMBO J. 12, 53–63.Alemany, V., Sanchez-Piris, M., Bachs, O., and Aligue, R. (2002).
Cmk2, a novel serine/threonine kinase in fission yeast. FEBS Lett. Kovelman, R., and Russell, P. (1996). Stockpiling of Cdc25 during a524, 79–86. DNA replication checkpoint arrest in Schizosaccharomyces pombe.
Mol. Cell. Biol. 16, 86–93.Al-Khodairy, F., Fotou, E., Sheldrick, K.S., Griffiths, D.J., Lehmann,A.R., and Carr, A.M. (1994). Identification and characterization of Kumagai, A., and Dunphy, W.G. (1992). Regulation of the cdc25new elements involved in checkpoint and feedback controls in fis- protein during the cell cycle in Xenopus extracts. Cell 70, 139–151.sion yeast. Mol. Biol. Cell 5, 147–160. Kumagai, A., and Dunphy, W.G. (1999). Binding of 14-3-3 proteinsAligue, R., Wu, L., and Russell, P. (1997). Regulation of Schizosac- and nuclear export control the intracellular localization of the mitoticcharomyces pombe Wee1 tyrosine kinase. J. Biol. Chem. 272, inducer Cdc25. Genes Dev. 13, 1067–1072.13320–13325. Lindsay, H.D., Griffiths, D.J., Edwards, R.J., Christensen, P.U., Mur-
ray, J.M., Osman, F., Walworth, N., and Carr, A.M. (1998). S-phase-Bahler, J., Wu, J.Q., Longtine, M.S., Shah, N.G., McKenzie, A., 3rd,specific activation of Cds1 kinase defines a subpathway of theSteever, A.B., Wach, A., Philippsen, P., and Pringle, J.R. (1998).checkpoint response in Schizosaccharomyces pombe. Genes Dev.Heterologous modules for efficient and versatile PCR-based gene12, 382–395.targeting in Schizosaccharomyces pombe. Yeast 14, 943–951.
Lopez-Girona, A., Furnari, B., Mondesert, O., and Russell, P. (1999).Bulavin, D.V., Higashimoto, Y., Popoff, I.J., Gaarde, W.A., Basrur,Nuclear localization of Cdc25 is regulated by DNA damage and aV., Potapova, O., Appella, E., and Fornace, A.J., Jr. (2001). Initiation14-3-3 protein. Nature 397, 172–175.of a G2/M checkpoint after ultraviolet radiation requires p38 kinase.
Nature 411, 102–107. Lopez-Girona, A., Kanoh, J., and Russell, P. (2001). Nuclear exclu-sion of Cdc25 is not required for the DNA damage checkpoint inBulavin, D.V., Amundson, S.A., and Fornace, A.J. (2002). p38 andfission yeast. Curr. Biol. 11, 50–54.Chk1 kinases: different conductors for the G(2)/M checkpoint sym-
phony. Curr. Opin. Genet. Dev. 12, 92–97. Lundgren, K., Walworth, N., Booher, R., Dembski, M., Kirschner, M.,and Beach, D. (1991). Mik1 and Wee1 cooperate in the inhibitoryBulavin, D.V., Higashimoto, Y., Demidenko, Z.N., Meek, S., Graves,tyrosine phosphorylation of Cdc2. Cell 64, 1111–1122.P., Phillips, C., Zhao, H., Moody, S.A., Appella, E., Piwnica-Worms,
H., and Fornace, A.J., Jr. (2003). Dual phosphorylation controls Matsusaka, T., and Pines, J. (2004). Chfr acts with the p38 stressCdc25 phosphatases and mitotic entry. Nat. Cell Biol. 5, 545–551. kinases to block entry to mitosis in mammalian cells. J. Cell Biol.
166, 507–516.Carr, A.M., MacNeill, S.A., Hayles, J., and Nurse, P. (1989). Molecularcloning and sequence analysis of mutant alleles of the fission yeast Maundrell, K. (1993). Thiamine-repressible expression vectors pREPcdc2 protein kinase gene: implications for cdc2� protein structure and pRIP for fission yeast. Gene 123, 127–130.and function. Mol. Gen. Genet. 218, 41–49. Mikhailov, A., Shinohara, M., and Rieder, C.L. (2004). Topoisomerase
II and histone deacetylase inhibitors delay the G2/M transition byChen, L., Liu, T.H., and Walworth, N.C. (1999). Association of Chk1triggering the p38 MAPK checkpoint pathway. J. Cell Biol. 166,with 14-3-3 proteins is stimulated by DNA damage. Genes Dev.517–526.13, 675–685.
Millar, J.B., McGowan, C.H., Lenaers, G., Jones, R., and Russell, P.Christensen, P.U., Bentley, N.J., Marthinho, R.G., Nielsen, O., and(1991). p80cdc25 mitotic inducer is the tyrosine phosphatase thatCarr, A.M. (2000). Mik1 levels accumulate in S phase and may medi-activates p34cdc2 kinase in fission yeast. EMBO J. 10, 4301–4309.ate an intrinsic link between S phase and mitosis. Proc. Natl. Acad.
Sci. USA 97, 2579–2584. Millar, J.B., Lenaers, G., and Russell, P. (1992). Pyp3� PTPase actsas a mitotic inducer in fission yeast. EMBO J. 11, 4933–4941.Dahlkvist, A., Kanter-Smoler, G., and Sunnerhagen, P. (1995). The
RCK1 and RCK2 protein kinase genes from Saccharomyces cerevis- Millar, J.B., Buck, V., and Wilkinson, M.G. (1995). Pyp1 and Pyp2iae suppress cell cycle checkpoint mutations in Schizosaccharomy- PTPases dephosphorylate an osmosensing MAP kinase controllingces pombe. Mol. Gen. Genet. 246, 316–326. cell size at division in fission yeast. Genes Dev. 9, 2117–2130.
Ford, J.C., al-Khodairy, F., Fotou, E., Sheldrick, K.S., Griffiths, D.J., Moreno, S., Nurse, P., and Russell, P. (1990). Regulation of mitosisand Carr, A.M. (1994). 14-3-3 protein homologs required for the DNA by cyclic accumulation of p80cdc25 mitotic inducer in fission yeast.damage checkpoint in fission yeast. Science 265, 533–535. Nature 344, 549–552.
Moreno, S., Klar, A., and Nurse, P. (1991). Molecular genetic analysisFurnari, B., Rhind, N., and Russell, P. (1997). Cdc25 mitotic inducertargeted by Chk1 DNA damage checkpoint kinase. Science 277, of fission yeast Schizosaccharomyces pombe. Methods Enzymol.
194, 795–823.1495–1497.

Inhibition of Cdc25 by Srk1 Kinase59
Peng, C.Y., Graves, P.R., Thoma, R.S., Wu, Z., Shaw, A.S., andPiwnica-Worms, H. (1997). Mitotic and G2 checkpoint control: regu-lation of 14-3-3 protein binding by phosphorylation of Cdc25C onserine-216. Science 277, 1501–1505.
Qian, Y.W., Erikson, E., Taieb, F.E., and Maller, J.L. (2001). The polo-like kinase Plx1 is required for activation of the phosphatase Cdc25Cand cyclin B-Cdc2 in Xenopus oocytes. Mol. Biol. Cell 12, 1791–1799.
Rhind, N., and Russell, P. (2001). Roles of the mitotic Wee1 andMik1 in the G2 DNA damage and replication checkpoints. Mol. Cell.Biol. 21, 1499–1508.
Russell, P., and Nurse, P. (1987). Negative regulation of mitosis bywee1�, a gene encoding a protein kinase homolog. Cell 49, 559–567.
Russell, P., and Nurse, P. (1986). cdc25� functions as an inducerin the mitotic control of fission yeast. Cell 45, 145–153.
Sanchez, Y., Wong, C., Thoma, R.S., Richman, R., Wu, Z., Piwnica-Worms, H., and Elledge, S.J. (1997). Conservation of the Chk1 check-point pathway in mammals: linkage of DNA damage to Cdk regula-tion through Cdc25. Science 277, 1497–1501.
Sanchez-Piris, M., Posas, F., Alemany, V., Winge, I., Hidalgo, E.,Bachs, O., and Aligue, R. (2002). The serine/threonine kinase Cmk2is required for oxidative stress response in fission yeast. J. Biol.Chem. 277, 17722–17727.
Shiozaki, K., and Russell, P. (1995). Cell cycle control linked toextracellular environment by MAP kinase pathway in fission yeast.Nature 378, 739–743.
Shiozaki, K., and Russell, P. (1996). Conjugation, meiosis, and theosmotic stress response are regulated by Spc1 kinase through Atf1transcription factor in fission yeast. Genes Dev. 10, 2276–2288.
Smith, D.A., Toone, W.M., Chen, D., Bahler, J., Jones, N., Morgan,B.A., and Quinn, J. (2002). The Srk1 protein kinase is a target forthe Sty1 stress-activated MAPK in fission yeast. J. Biol. Chem.277, 33411–33421.
Walworth, N.C. (2001). DNA damage: Chk1 and Cdc25, more thanmeets the eye. Curr. Opin. Genet. Dev. 11, 78–82.
Wang, X., McGowan, C.H., Zhao, M., He, L., Downey, J.S., Fearns,C., Wang, Y., Huang, S., and Han, J. (2000). Involvement of theMKK6-p38gamma cascade in gamma-radiation-induced cell cyclearrest. Mol. Cell. Biol. 20, 4543–4552.
Wilkinson, M.G., Samuels, M., Takeda, T., Toda, T., Toone, M.W.,Shieh, J.-C., Millar, J.B.A., and Jones, N.C. (1996). The Atf1 transcrip-tion factor is a target for the Sty1 stress activated MAP kinasepathway in fission yeast. Genes Dev. 10, 2289–2301.
Zeng, Y., and Piwnica-Worms, H. (1999). DNA damage and replica-tion checkpoints in fission yeast require nuclear exclusion of theCdc25 phosphatase via 14-3-3 binding. Mol. Cell. Biol. 19, 7410–7419.
Zeng, Y., Forbes, K.C., Wu, Z., Moreno, S., Piwnica-Worms, H., andEnoch, T. (1998). Replication checkpoint requires phosphorylationof the phosphatase Cdc25 by Cds1 or Chk1. Nature 395, 507–510.
Copyright © 2022 FDOKUMEN




















