
Skin diseases associated with Malassezia yeasts: Facts and controversies
Upload
independentCategory
view
0download
0
Cell, Vol. 57, 295-303, April 21, 1989, Copyright 0 1989 by Cell Press
Conservation of M itotic Controls in F ission and Budding Yeasts
Paul Russell,’ Sergio Moreno,t and Steven I. Reed* * Department of Molecular Biology Research Institute of Scripps Clinic La Jolla, California 92037 tlCRF Cell Cycle Control Laboratory Microbiolgy Unit Department of Biochemistry University of Oxford Oxford OXlSQU England
Summary
In fission yeast, the initiation of mitosis is regulated by a control network that integrates the opposing activi- ties of mitotic inducers and inhibitors. To evaluate whether this control system is likely to be conserved among eukaryotes, we have investigated whether a similar mitotic control operates in the distantly related budding yeast S. cerevisiae. We have found that the protein kinase encoded by the mitotic inhibitor gene weeI+ of fission yeast, which acts to delay mitosis, is able also to delay the initiation of mitosis when ex- pressed in S. cerevisiae. The wee7+ activity is coun- teracted in S. cerevisiae by the gene product of M/HI, a newly identified gene capable of encoding a protein of M W 54,000, which is a structural and functional homolog of the cdc25+ mitotic inducer of fission yeast. Expression of weel+ in a mihf- strain prevents the initiation of mitosis. These data indicate that im- portant features of the cdc25+-weel+ mitotic control network identified in S. pombe are conserved in S. cerevisiae, and therefore are also likely to be generally conserved among eukaryotic organisms.
Introduction
Increasing molecular evidence indicates that some of the components of eukaryotic cell cycle control are evolution- arily conserved among widely divergent species, even be- tween yeast and man. Homologs of the fission yeast p34CdC2 protein kinase (called p34CDC28 in budding yeast), the interacting protein ~13~~~‘. and cyclin have been im- plicated to have roles in cell cycle control in a broad range of organisms (Beach et al., 1982; Draetta et al., 1987; Goebl and Byers, 1988; Lee and Nurse, 1987; Solomon et al., 1988; Swenson et al., 1986; Hadwiger et al., sub- mitted). The exact functions of these elements are as yet unclear. However, recent evidence suggests that ~34~~~~ homologs are quite closely involved in stimulating mitotic events (Arion et al., 1988; Dunphy et al., 1988; Gautier et al., 1988; Labbe et al., 1988).
While it now appears very likely that ~34~~~ is a central component of mitotic control in most, if not all eukaryotes,
it is also clear that, at least in fission yeast, ~34~~~~ is but one element of a complex regulatory network that deter- mines the timing of mitosis. Genetic studies in S. pombe have identified three higher order regulatory elements, the gene products of c&25+, niml+, and weel+, having roles quite distinct from that of ~34~~~~ (Russell and Nurse, 1986a, 1987a, 1987b). The timing of mitosis is acutely sensitive to the level of expression of these higher order regulatory elements. The c&25+ and nim7+ gene products act to stimulate the initiation of mitosis, whereas the weel+ gene product has the opposite function of in- hibiting the initiation of mitosis. The timing of mitosis is de- termined by the competition between the positive and negative regulatory elements. The niml+ and weel+ ele- ments are likely to be protein kinases, based on their pri- mary structure homology to known protein kinases. Genetic evidence indicates that the hierarchical control activities of the c&25+, niml+, and weelf elements are integrated in a pathway that regulates the activity of the p34cdc2 protein kinase.
In view of the apparent conservation of p34cdc2, ~13~~~‘~ and cyclin cell cycle control elements among eukaryotes, it is important to determine whether mitotic control net- works similar to the Schizosaccharomyces pombe cdc25+-niml+-weel+ control serve to regulate progres- sion through the cell cycle in other eukaryotic species. The budding yeast, S. cerevisiae, has a cell cycle that ap- pears to be organized and regulated quite differently from that in S. pombe (Nurse, 1985), which is consistent with their high degree of evolutionary divergence. Although there is some evidence indicating that a cell cycle control that regulates the initiation of mitosis exists in S. cere- visiae, very little is known about how this control operates (Piggott et al., 1982; Pringle and Hartwell, 1981; Weinert and Hartwell, 1988). Therefore, a comparison of these two yeast species is a useful criterion for assessing conserva- tion of mitotic controls.
If the c&25+-niml+-weel+ control network is largely conserved between fission and budding yeasts, two straightforward predictions should be fulfilled. First, S. pombe mitotic control elements should be able to regulate mitosis when expressed in S. cerevisiae. Here we show this to be true, by demonstrating that the weel+ protein kinase is able to delay the initiation of mitosis in S. cerevisiae. The second prediction is that S. cerevisiae should have homologs of the S. pombe mitotic control ele- ments. We have confirmed this by identifying a new S. cerevisiae gene called MIHl, a homolog of the S. pombe c&25+ mitotic inducer. As in S. pombe, the S. cerevisiae MIHl/cdc25+ class of mitotic inducer can counteract the mitotic inhibitor function of the weel+ gene product. These data show that S. cerevisiae has a mitotic control that responds to regulatory elements of the types encoded by cdc25+ and weel+. Therefore, it is likely that similar mitotic control networks using conserved regulatory ele- ments will be present in other eukaryotic species.
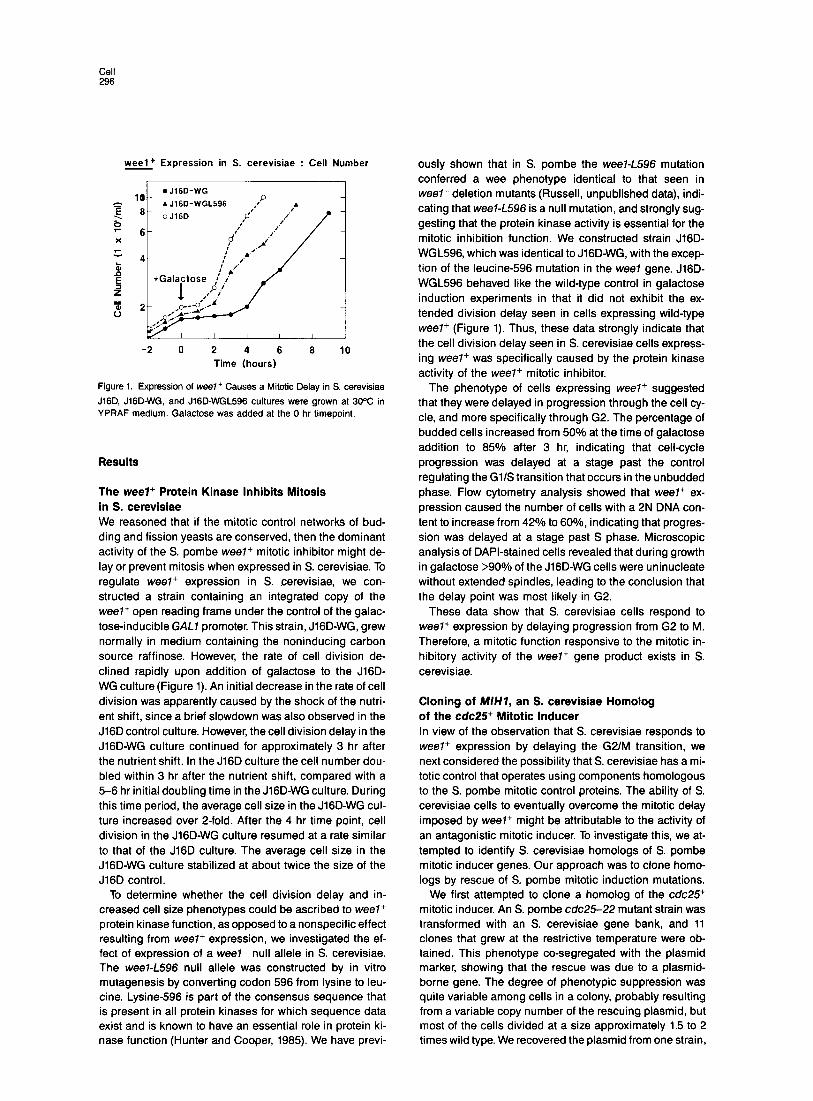
Cell 296
weel+ Expression in S. cerevisiae : Cell Number
. lo-
J16D-WG
. J16D-WGL596 P
-2 0 2 4 6 8 10 Time (hours)
Figure 1. Expression of weel+ Causes a Mitotic Delay in S. cerevisiae
J16Q JIGD-WG, and J16D-WGL596 cultures were grown at 30% in YPRAF medium. Galactose was added at the 0 hr timepoint.
Results
The weel+ Protein Kinase Inhibits Mitosis in S. cerevisiae We reasoned that if the mitotic control networks of bud- ding and fission yeasts are conserved, then the dominant activity of the S. pombe weel+ mitotic inhibitor might de- lay or prevent mitosis when expressed in S. cerevisiae. To regulate weel+ expression in S. cerevisiae, we con- structed a strain containing an integrated copy of the weel+ open reading frame under the control of the galac- tose-inducible GAL7 promoter. This strain, J lGD-WG, grew normally in medium containing the noninducing carbon source raffinose. However, the rate of cell division de- clined rapidly upon addition of galactose to the J16D- W G culture (Figure 1). An initial decrease in the rate of cell division was apparently caused by the shock of the nutri- ent shift, since a brief s lowdown was also observed in the J16D control culture. However, the cell division delay in the J lGD-WG culture continued for approximately 3 hr after the nutrient shift. In the J16D culture the cell number dou- bled within 3 hr after the nutrient shift, compared with a 5-6 hr initial doubling time in the J lGD-WG culture. During this time period, the average cell size in the J lGD-WG cul- ture increased over P-fold. After the 4 hr time point, cell division in the J lGD-WG culture resumed at a rate similar to that of the J16D culture. The average cell size in the J lGD-WG culture stabilized at about twice the size of the J16D control.
To determine whether the cell division delay and in- creased cell size phenotypes could be ascribed to weel+ protein kinase function, as opposed to a nonspecific effect resulting from weelf expression, we investigated the ef- fect of expression of a weel- null allele in S. cerevisiae. The weel-7596 null allele was constructed by in vitro mutagenesis by converting codon 596 from lysine to leu- tine. Lysine-596 is part of the consensus sequence that is present in all protein kinases for which sequence data exist and is known to have an essential role in protein ki- nase function (Hunter and Cooper, 1965). We have previ-
ously shown that in S. pombe the weel-L596 mutation conferred a wee phenotype identical to that seen in weel- deletion mutants (Russell, unpublished data), indi- cating that weel-L596 is a null mutation, and strongly sug- gesting that the protein kinase activity is essential for the mitotic inhibition function. We constructed strain J16D- WGL596, which was identical to J lGD-WG, with the excep- tion of the leucine-596 mutation in the wee7 gene. J16D- WGL596 behaved like the wild-type control in galactose induction experiments in that it did not exhibit the ex- tended division delay seen in cells expressing wild-type weel+ (Figure 1). Thus, these data strongly indicate that the cell division delay seen in S. cerevisiae cells express- ing weel+ was specifically caused by the protein kinase activity of the weel+ mitotic inhibitor.
The phenotype of cells expressing weel+ suggested that they were delayed in progression through the cell cy- cle, and more specifically through G2. The percentage of budded cells increased from 50% at the time of galactose addition to 65% after 3 hr, indicating that cell-cycle progression was delayed at a stage past the control regulating the GllS transition that occurs in the unbudded phase. Flow cytometry analysis showed that weel+ ex- pression caused the number of cells with a 2N DNA con- tent to increase from 42% to 60%, indicating that progres- sion was delayed at a stage past S phase. Microscopic analysis of DAPI-stained cells revealed that during growth in galactose >90% of the J lGD-WG cells were uninucleate without extended spindles, leading to the conclusion that the delay point was most likely in G2.
These data show that S. cerevisiae cells respond to weel+ expression by delaying progression from G2 to M. Therefore, a mitotic function responsive to the mitotic in- hibitory activity of the weel+ gene product exists in S. cerevisiae.
Cloning of MIH7, an S. cerevisiae Homolog of the c&25+ Mitotic Inducer In view of the observation that S. cerevisiae responds to weel+ expression by delaying the G2/M transition, we next considered the possibility that S. cerevisiae has a mi- totic control that operates using components homologous to the S. pombe mitotic control proteins. The ability of S. cerevisiae cells to eventually overcome the mitotic delay imposed by weel+ might be attributable to the activity of an antagonistic mitotic inducer. To investigate this, we at- tempted to identify S. cerevisiae homologs of S. pombe mitotic inducer genes. Our approach was to clone homo- logs by rescue of S. pombe mitotic induction mutations.
We first attempted to clone a homolog of the cdc25+ mitotic inducer. An S. pombe cdc25-22 mutant strain was transformed with an S. cerevisiae gene bank, and 11 clones that grew at the restrictive temperature were ob- tained. This phenotype co-segregated with the plasmid marker, showing that the rescue was due to a plasmid- borne gene. The degree of phenotypic suppression was quite variable among cells in a colony, probably resulting from a variable copy number of the rescuing plasmid, but most of the cells divided at a size approximately 1.5 to 2 times wild type. We recovered the plasmid from one strain,

cdc25+-weeI+ Mitotic Control 297
Rescue Of cdcX-22
pCE25-Sl + t I
-s2 I 1
-53 - H -s4 + t i
Figure 2. Restriction Enzyme Map of pEC25
The restriction enzyme map of the insert DNA in pCE25 is shown at the top. The 2.2 kb region that was sequenced is indicated by the solid line below the restriction map. The extent of subclones and their ability to complement cdc2522 is shown in the lower part of the figure.
naming it pCE25. The insert DNA was purified and used as a probe in a Southern hybridization with total DNAfrom the other ten transformants. This showed that the other ten transformants had plasmids containing the same in- sert DNA or sequences that overlapped it (data not shown).
We wished to determine whether pCE25 could provide all the activities of cdc25+. To do this, we introduced pCE25 into a diploid strain containing one wild-type and one deleted allele of cdc25. After sporulation, we found that all viable colonies that contained the deleted version of cdc25 also contained pCE25. These cells were unable to divide without pCE25, showing that pCE25 provided all the essential functions normally performed by cdc25+.
By analyzing the ability of subclones to rescue cdc25-22, we were able to localize the critical sequences to a 1.1 kb region bordered by Sstl and Xbal sites (Figure 2). We sequenced this fragment and adjoining regions and found a 1.5 kb open reading frame (Figure 3). Near the beginning of the open reading frame, there is a methi- onine codon that could be used to initiate translation of a protein 474 amino acids long with a molecular weight of 54,000. Upon comparison of the predicted protein se- quence to those currently available in databases, a signifi- cant degree of sequence identity to only one protein was found: the S. pombe cdc25+ gene product (Russell and Nurse, 1986a). The homology was located in a 150 amino acid region in the C-terminal halves of the two proteins, where there was 40% sequence identity (Figure 4). In- terestingly, this correlated with the subcloning analysis that showed that subclone pCE25-Sl, which lacked the amino-terminal 25% of the open reading frame, rescued cdc25-22. Thus, a truncated version of the protein is syn- thesized and is functional in S. pombe.
In recognition of the structural and apparent functional similarity between the S. pombe cdc25+ mitotic inducer and the complementing S. cerevisiae gene present on pCE25, we named the S. cerevisiae gene MIHl (mitotic in- ducer homolog)
M/H7 Functions as a Mitotic inducer in S. cerevisiae The observation that expression of weel+ delays mitosis in S. cerevisiae, together with the cloning of a S. cere-
visiae homolog of the S. pombe cdc25+ mitotic inducer, strongly suggested that the mitotic controls of the two yeasts are largely conserved. To establish whether the M/H1 gene product has a role in S. cerevisiae mitotic con- trol, we examined the effect of loss of M/H7 function. We introduced a deletion into one copy of the gene in a diploid that was then induced to sporulate (see Experimental Procedures). All five tetrads that were analyzed yielded four viable clones having equal growth rates, showing that M/H1 function is not required for vegetative growth. How- ever, flow cytometry analysis showed that the percentage of cells with a 2N DNA content increased from 41% in wild- type cells to 58% in mihl- cells (Figure 5). This shift to a greater proportion of cells in the latter half of the cell cycle in mihl- cells was accompanied by a 15% increase in av- erage cell size.
The phenotype exhibited by mihl- cells is very similar to that seen in S. pombe mutants that partly lack mitotic induction activities, such as temperature-sensit ive cdc25 mutants grown at a semi-permissive temperature, as well as S. pombe mutants lacking the mitotic inducer niml+ (Russell and Nurse, 1987b). These data indicate that the M/H1 gene product functions as a mitotic inducer in S. cerevisiae, and like its homolog cdc25+ in S. pombe, M/H1 may provide only part of the mitotic induction activi- ties present in the cell.
In S. cerevisiae, the M/H7 Mitotic Induction Activity Is Necessary to Counteract the wee7+ Mitotic Inhibitor To clarify the role of M/H7 in S. cerevisiae mitotic control, we investigated the relationship between its function and that of the mitotic inhibitor weel+. If M/H1 acts as a mi- totic inducer in a manner similar to that of cdc25+ in S. pombe, it should function to counteract the mitotic inhibi- tory activity of weel+. We tested this hypothesis by deter- mining whether the effect of weel+ expression was ac- centuated in cells lacking MIHl. The M/H1 gene was disrupted in a J lGD-WG strain. This strain, J lGD-WGCD, grew normally in raffinose medium, but cell division ceased soon after the shift to galactose media. Like the J lGD-WG strain, these cells were delayed in the budded phase of the cell cycle with a 2N DNA content (Figure 6). However, in the J lGD-WGCD strain, the cell cycle arrest was complete: no cells recovered from the cell cycle ar- rest. Microscopic observation showed that the cells ini- tially arrested with a single long bud (Figure 7). Observa- tion of these cells stained with the DNA-specif ic dye DAPI showed that they had a single nucleus that was located near the neck of the bud (data not shown). Upon pro- longed incubation, some of the cells acquired two buds or exhibited a branched structure. At the later timepoints the nucleus had often migrated further into the bud and ac- quired an elongated appearance, but in no cells was nu- clear division observed.
These data showed that MlHl function is required to overcome the cell division cycle delay imposed by weel+. Consistent with these results, we observed that overex- pression of M/H1 in cells expressing weel+ largely neu- tralized the morphological and cell size phenotypes con- ferred by weel+ (data not shown), suggesting that MlHl

Cell 298
CTG TCA ATA TGC CTG TCA GTT TGA AAA TCG AAG ATA CCA CAT CGT ACA AAA GCA AGA GGG GTG CCC CTA ATG GGA GAA GAT TAC CGT TTT
TAT TTT GGG GTA GCT AAC TGC GCC ACA AAT AGT ATC TAC CAG TTT GCT ACT TTT AGA ACT GTA TTA TGT ATC TTT GCC GAA TAC TTA ATT
TCC TAG GCC TCA TTT TCA GGA CTA AGG ATG AAG AGG GGC TTC AAT GTT TTA TAT GGC GCT TTT TCT GTC CAT TGT ATA CGT CAA AAA AGC
ATA GAT ACT CCA CTT ATT AGC GGT AAA GCC CTT TTT GAT ACA TTG TTC CGG CGT TTA AAT ATT CTC ATT ATG ATA TTT CCC CAA CAG TTA
CAC TCT GTT ATT TCC TTC TTT TTT ATG CTT TAT CTG TTT TTT CCC TGT TTC GAG TGC CGT ATA TCA GAA AAC TTA AAG TTA CAA GCG AAA
ACA AAG AAA AGA GAG TTA ACT TGA AAG GAG ACA AAA GAT AAA CAG AGC AGT GGA CAA ACC AGG ATT GAA GTC AGC GAG GGT GAA GAA ACC
ATG AAC AAT ATA TTT CAT GGA ACT GAA GAT GAA TGT GCC AAT GAA GAC GTT CTT AGT TTC CA?, AAA ATT TCC TTG AAA AGT CCC TTT GGT MNNIFHGTEDECANEDVLSFQKISLKSPFG
AAG AAG AAG AAC ATA TTT AGA AAT GTT CAG ACC TTC TTT AAG TCA AAA AGC AAA CAT TCG AAT GTC GAC GAT GAT TTA ATC AAT ?@A GAG KKKNIFRNVQTFFKSKSKHSNVDDDLINKE
AAT CTT GCC TTT GAT AAA TCT CCA TTG TTA ACA AAT CAC AGG AGT AAG GAA ATT GAT GGT CCT TCA CCG AAT ATA AAG CAG CTT GGC CAT NLAFDKSPLLTNHRSKEIDGPSPNIKQLGH
CGC GAT GAG TTA GAT GAA AAT GA?+ AAT GAA AAT GAT GAT ATA GTC TTA AGC ATG CAT TTT GCT TCT CAA ACC TTA CAA AGT CCA PICA AGA RDELDENENENDDIVLSMHFASQTLQSPTR
AAC TCA TCA AGA AGA TCT TTG ACC AAT AAT AGG GAT AAT GAT CTT TTA AGT AGA ATC AAG TAT CCT GGC AGT CCC CAG AGG TCG AGC TCT NSSRRSLTNNRDNDLLSRIKYPGSPQRSSS
TTT TCA AGG TCA AGG TCC TTG TCG AGA AAG CCA AGC ATG AAT AGC AGT AGC AAT TCA AGC AGG AGG GTG CAA CGG CAA GAT GGG AAA ATT FSRSRSLSRKPSMNSSSNSSRRVQRQDGKI
CCG AGG TCA TCG AGA AAA AGT TCA CAG AAA TTT TCT &AC ATT ACT CAG AAT ACT TTG AAC TTC ACG TCT GCG TCA TCC AGT CCA CTA GCT PRSSRKSSQKFSNITQNTLNFTSASSSPLA
CCT AAT TCG GTA GGG GTG AAA TGT TTT GAA TCA TGC CTA GCC AAA ACA CAG ATA CCA TAT TAT TAT GAC GAT CGT AAC TCA ATG ACT TTT PNSVGVKCFESCLAKTQIPYYYDDRNSMTF
TCC CTC GAA TTT CTC CAG AAA CGT TTG AAA AAT ATT CTG CAG AAT AAT ATG TGC GAG TCA TTT TAT AAT TCA TGT CGT ATT ATA GAT TGT SLEFLQKRLKNILQNNMCESFYNSCRIIDC
CGG TTT GAA TAC GAA TAT ACT GGC GGC CAT ATA ATA AAC TCC GTT AAC ATA CAC TCG AGA GAT GAG TTA GAG TAC GAA TTT ATC CAT AAA RFEYEYTGGHIINSVNIHSRDELEYEFIHK
GTT TTG CAT AGC GAT ACT AGC AAC AAC AAC ACT CTA CCA ACT CTT TTA ATT ATC CAT TGC GAA TTT AGT TCC CAC CGG GGA CCT TCA TTG VLHSDTSNNNTLPTLLIIHCEFSSHRGPSL
GCC TCT CAT TTG AGA AAT TGT GAT CGT ATA ATA AAT CAG GAT CAT TAT CCC AAA TTA TTC TAT CCC GAC ATA TTG ATA TTG GAT GGC GGA ASHLRNCDRIINQDHYPKLFYPDILILDGG
TAC AAA GCT GTT TTG ACA TTT CCT GAA CTA TGT TAT CCA CGT CAG TAC GTG GGA ATG AAC TCA CAA GAA AAT TTG TTG AAT TGT GAA CAA YKAVLTFPELCYPRQYVGMNSQENLLNCEQ
GAA ATG GAT AAG TTT AGG AGA GAA TCT AAA AGA TTT GCT ACT AAA AAT AAT TCT TTT AGA AAA CTA GCT TCC CCT TCA AAT CCT AAC TTC EMDKFRRESKRFATKNNSFRKLASPSNPNF
TTT TAT AGA GAT AGC CAT CAA TCT TCA ACA ACA ATG GCA TCT TCT GCA CTA TCG TTC AGG TTT GAG CCG CCT CCG AAG CTG TCA TTA AAC FYRDSHQSSTTMASSALSFRFEPPPKLSLN
CAC AGA AGA GTC TCT AGT GGT TCA TCA TTA AAT TCA AGT GAG AGT ACA G G C GAC GAA AAT TTC TTC CCA TCT TAA GTA AGT CGT CTA TGA HRRVSSGSSLNSSESTGDENFFPSX
GTA GTA ACA GTA ACC TAA GCA CTT CAC ATA TGC TAC TAA TGG ACG GAC TGG ATA CCC CAT CAT ATT TTA GTT TCG AAG ATG AGA GAG GCA
ATC ATC AGC AAG TAA GTG GAG ATG AAG AGC AAG ACG GAG ATT TTA CTT TCG TTG GTT CCG ACC GAG AAG ATT TAC CCA G G C CCG CAA GAA
GAT CGT TA
Figure 3. DNA Sequence of the pCE25 Open Reading Frame, the M/HI Gene
The GenBank accession number for this sequence is JO4846
90
180
270
360
450
540
630
720
810
900
990
1080
1170
1260
1350
1440
1530
1620
1710
1800
1890
1980
2070
2160
2168
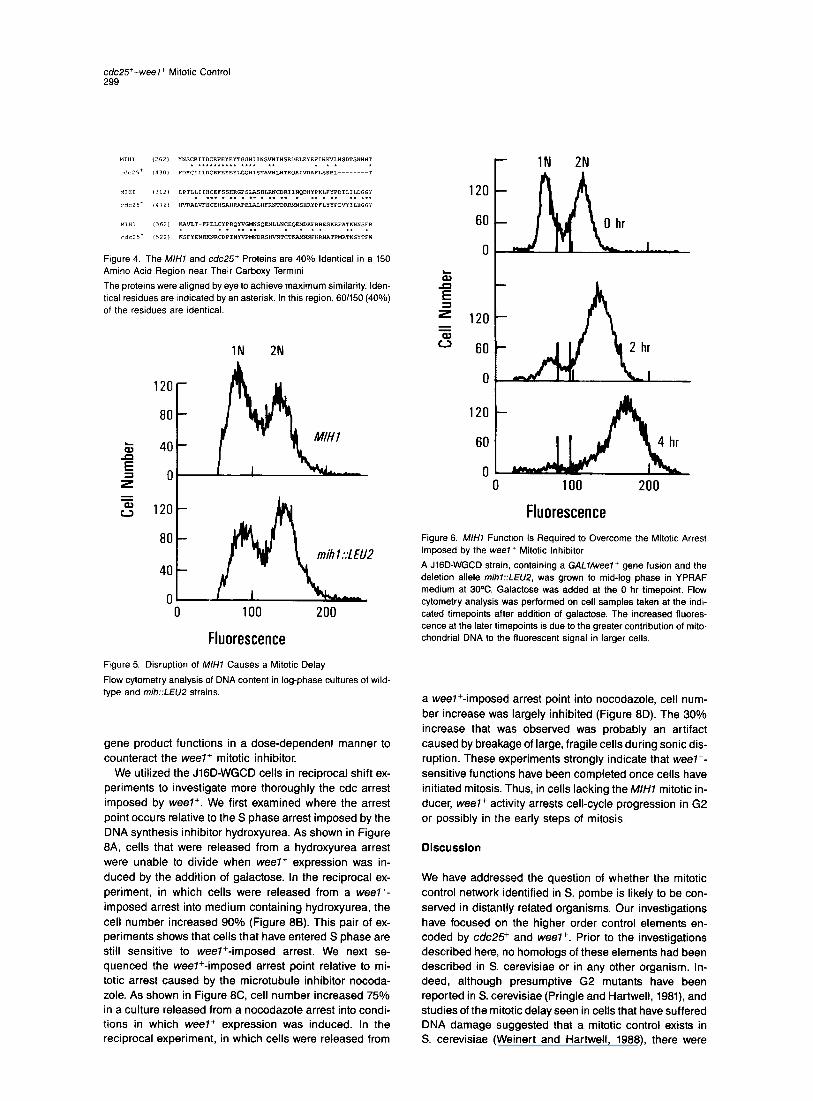
c&25+-weel+ Mitotic Control 299
Figure 4. The M/H7 and cdc25+ Proteins are 40% Identical in a 150 Amino Acid Region near Their Carboxy Termini
The proteins were aligned by eye to achieve maximum similarity. Iden- tical residues are indicated by an asterisk. In this region, 601150 (40%) of the residues are identical.
1N 2N
120-
80 -
40 -
0'
Fluorescence Figure 5. Disruption of M/H7 Causes a Mitotic Delay
Flow cytometry analysis of DNA content in log-phase cultures of wild- type and mih::LEUP strains.
gene product functions in a dose-dependent manner to counteract the weel+ mitotic inhibitor.
We utilized the J lGD-WGCD cells in reciprocal shift ex- periments to investigate more thoroughly the cdc arrest imposed by weel+. We first examined where the arrest point occurs relative to the S phase arrest imposed by the DNA synthesis inhibitor hydroxyurea. As shown in Figure 6A, cells that were released from a hydroxyurea arrest were unable to divide when weel+ expression was in- duced by the addition of galactose. In the reciprocal ex- periment, in which cells were released from a weeli- imposed arrest into medium containing hydroxyurea, the cell number increased 90% (Figure 88). This pair of ex- periments shows that cells that have entered S phase are still sensitive to weel+-imposed arrest. We next se- quenced the wee7+-imposed arrest point relative to mi- totic arrest caused by the microtubule inhibitor nocoda- zole. As shown in Figure 8C, cell number increased 75% in a culture released from a nocodazole arrest into condi- tions in which weel+ expression was induced. In the reciprocal experiment, in which cells were released from
r 1N 2N
120 -
60 -
0 m I
120
60
0
120
60
0 0 100 200
Fluorescence Figure 6. M//-/l Function Is Required to Overcome the Mitotic Arrest Imposed by the weel+ Mitotic Inhibitor
A J lGD-WGCD strain, containing a GAL7Aveel+ gene fusion and the deletion allele mihl::LEU2, was grown to mid-log phase in YPRAF medium at 30%. Galactose was added at the 0 hr timepoint. Flow cytometry analysis was performed on cell samples taken at the indi- cated timepoints after addition of galactose. The increased fluores- cence at the later timepoints is due to the greater contribution of mito- chondrial DNA to the fluorescent signal in larger cells.
a weel+-imposed arrest point into nocodazole, ceil num- ber increase was largely inhibited (Figure 8D). The 30% increase that was observed was probably an artifact caused by breakage of large, fragile cells during sonic dis- ruption. These experiments strongly indicate that weel+- sensitive functions have been completed once cells have initiated mitosis. Thus, in cells lacking the M/H7 mitotic in- ducer, weel+ activity arrests cell-cycle progression in G2 or possibly in the early steps of mitosis.
Discussion
We have addressed the question of whether the mitotic control network identified in S. pombe is likely to be con- served in distantly related organisms. Our investigations have focused on the higher order control elements en- coded by c&25+ and weel+. Prior to the investigations described here, no homologs of these elements had been described in S. cerevisiae or in any other organism. In- deed, although presumptive G2 mutants have been reported in S. cerevisiae (Pringle and Hartwell, 1981), and studies of the mitotic delay seen in cells that have suffered DNA damage suggested that a mitotic control exists in S. cerevisiae (Weinert and Hartwell, 1988) there were
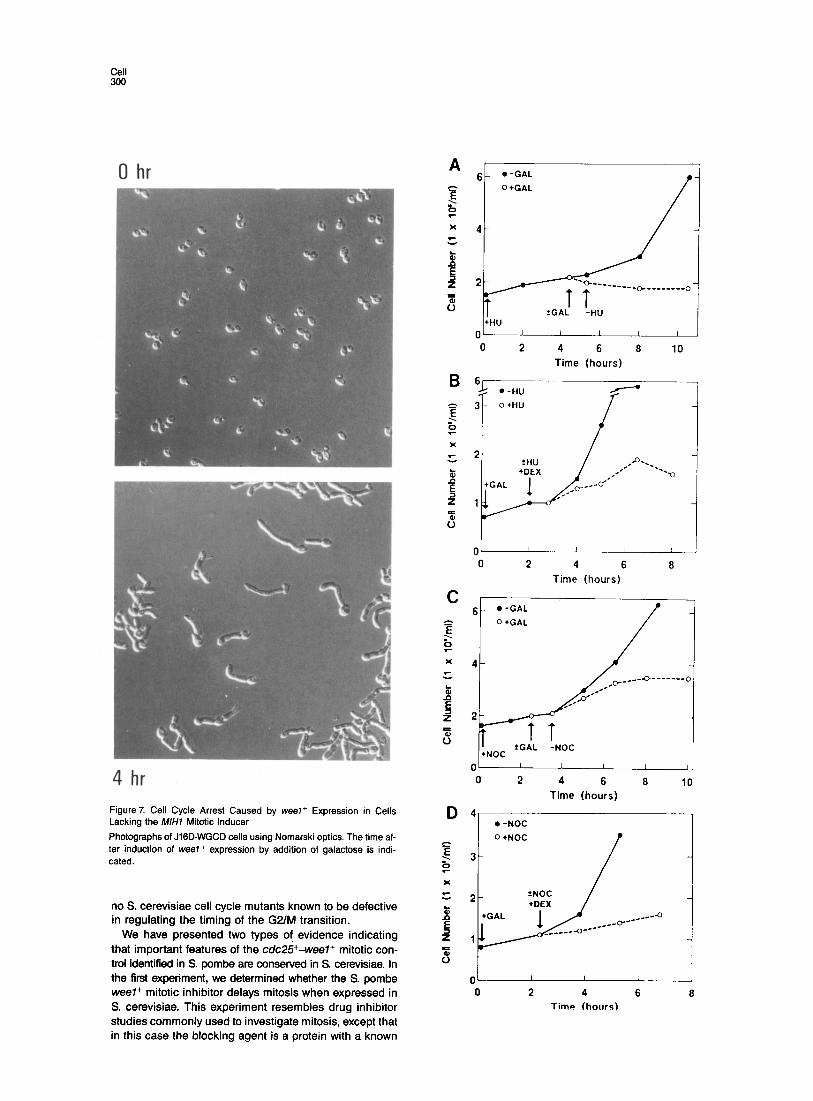
0 hr
4 hr Figure7. Cell Cycle Arrest Caused by weeI+ Expression in Cells Lacking the M/H7 Mitotic Inducer
Photographs of JIGD-WGCD cells using Nomarski optics. The time af- ter induction of weel+ expression by addition of galactose is indi- cated.
no S. cerevisiae cell cycle mutants known to be defective in regulating the timing of the G2/M transition.
We have presented two types of evidence indicating that important features of the c&25+-weel+ mitotic con- trol identified in S. pombe are conserved in S. cerevisiae. In the first experiment, we determined whether the S. pombe weel+ mitotic inhibitor delays mitosis when expressed in S. cerevisiae. This experiment resembles drug inhibitor studies commonly used to investigate mitosis, except that in this case the blocking agent is a protein with a known
6
I
.-GAL o+GAL f
:I ..,/ I ?GAi -NJ +HU
II I I I I I
0 2 4 6 a 10 Time (hours)
0' 1 I I 1 I
0 2 4 6 a Time (hours)
6- O-GAL 0 +GAL
4- --,-C.-------o
l OC
0' I I 1 I I
0 2 4 6 a 10 Time (hours)
4 4 l -NOC l -NOC
0 woe 0 woe
3- 3-
-4 -4
0' I I 1
0 2 4 6 8 Time (hours)

cdc25+-weel+ Mitotic Control 301
mitotic regulatory function. The finding that the weel+ mi- totic inhibitor is functional in S. cerevisiae shows that, at the very least, a mitotic initiation process capable of re- sponding to weel+ is present in S. cerevisiae.
The cloning of MIHl, a structural homolog of c&25+, provides the second type of evidence that important aspects of the S. pombe mitotic control are conserved in S. cerevisiae. The role of M/H1 in S. cerevisiae appears to be similar to that of cdc25+ in S. pombe: the M/H1 gene product functions as a mitotic inducer that is capable of counteracting the effect of the weel+ mitotic inhibitor. To- gether, these data show that in S. cerevisiae the mitotic ini- tiation mechanism responds to the activity of rate-limiting mitotic inducers and inhibitors of the types encoded by weel+ and cdc25+IMIHl. There is no direct evidence that a weel+ homolog exists in S. cerevisiae, although this must be considered a strong possibility. Moreover, in view of the large degree of evolutionary divergence between fission and budding yeasts, these results suggest it is very likely that homologs of cdc25+ and weel+ function in the mitotic controls of higher cells.
Unique Features of Mitotic Control in S. cerevisiae It is apparent that certain features of S. cerevisiae have made the analysis of mitotic control in this organism more difficult. In particular, our experiments have shown that at least one S. cerevisiae mitotic inducer gene, MIHl, would not have been detected by cdc mutant screens. This is be- cause, unlike S. pombe cdc25- mutants, S. cerevisiae mihl+ mutants do not have a cdc phenotype. While it is possible that S. cerevisiae can undergo mitosis without the mitotic induction activity of a cdc25+ homolog, it is more likely that M/H1 is nonessential because it provides only a portion of the mitotic induction activities present in the cell. In recent years it has become apparent that in S. cerevisiae essential proteins are frequently encoded by two or more similar genes (Kataoka et al., 1984; Tatchell et al., 1984; Toda et al., 1987). The failure to identify other S. cerevisiae mitotic inducer genes in the cdc25-22 res- cue experiment may have been due to problems as- sociated with the divergence of gene expression mecha- nisms between the two yeasts, since it is known that S. cerevisiae genes are often poorly expressed in S. pombe (Russell, 1989). However, it is likely that other S. cerevisiae mitotic inducer genes could be cloned by suppression of the cdc arrest phenotype of the J lGD-WGCD strain.
Divergent Mitotic Roles of the p34 Protein Kinases in Fission and Budding Yeasts? In S. pombe, genetic analysis has suggested that the cdc25+/weel+ control system acts by regulating the activ-
Figure 8. Reciprocal Shift Experiments
Strain J lGD-WGCD was grown at 30C in YPRAF medium to mid-log phase. + signifies that the indicated drug or carbon source was added; - signifies that the indicated drug or carbon source was washed out by centrifuging cells and resuspending in fresh, pre- warmed medium; + signifies that the culture was divided into two flasks and the indicated drug or carbon source was added to one flask. Abbreviations: GAL, galactose; HlJ, hydroxyurea; DEX, dextrose; NOC, nocodazole.
ity of the ~34~~~~ protein kinase. Although mutational analysis has shown that ~34~~~~ has two essential func- tions in the cell cycle, being required for transiting both GllS and G2/M, the cdc25+/weel+ regulatory system ap- pears to regulate only the G2 function. In S. cerevisiae, the ~34~~~~ homolog is encoded by CDC28. Mutational analy- sis has indicated that CDC28 functions are completed early in the cell cycle, near the GllS boundary. This poses a paradox, since by analogy to the mitotic control of S. pombe, the weel+ gene product would be expected to function by controlling CDC28 activity. We have re- examined the role of CDC28 in mitosis and have found that all functions performed by ~34~~~~ can be com- pleted in cells that are arrested in S-phase by the drug hydroxyurea (Reed, unpublished data). On the other hand, our experiments reported here have shown that in mihl- cells, mitotic functions that are sensitive to weel+ inhibition are not completed during a hydroxyurea arrest. These results suggest that p34coc2* mitotic functions, if they exist, are not sensitive to the mitotic inhibitory activi- ties of the weel+ gene product. A caveat to this conclu- sion is that it is based on the presumption that in mihl- cells, p34CDC28 functions are completed on schedule dur- ing a hydroxyurea arrest. It has previously been sug- gested that p34CDC2* may have mitotic functions that are normally completed soon after commitment to the GUS transition (Piggott et al., 1982; Russell and Nurse, 1986b). It is possible that completion of these functions is delayed in cells defective in M/H1 function. This question is cur- rently under investigation.
Conclusions We have described two experimental approaches that have extended the understanding of mitotic control from fission yeast to budding yeast. One approach, in which we used cross-species complementation to identify and clone the M/H1 mitotic inducer gene, had earlier been suc- cessfully employed to clone the human homolog of cdc2+ (Lee and Nurse, 1987) The second approach, in which we examined whether the S. cerevisiae mitotic control responded to the S. pombe weel+ mitotic inhibitor, is con- ceptually similar to studies showing that injection of clam cyclin mFlNA into Xenopus oocytes stimulated progres- sion through meiosis (Swenson et al., 1986). Applied to- gether, these approaches have allowed us to identify a mi- totic control in S. cerevisiae that had not been discovered by more traditional genetic analyses. We were able to use the combined approaches to construct an S. cerevisiae strain that exhibits a conditional cdc arrest at the G2/M boundary. This strain can now be used in pseudorever- sion and suppression studies to identify additional inter- acting elements and to acquire a more detailed under- standing of mitotic control in budding yeast. Our data strongly indicate that the S. pombe mitotic control should serve as a useful model system for these studies. In view of these results, we think it is likely that the cdc25+/weel+ mitotic control system is conserved in higher eukaryotes, and where possible it should be profitable to apply the ap- proach described here to the analysis of mitotic control in these organisms.

Cell 302
Experimental Procedures
Plasmid and Strain Constructions To construct strain JlGD-WG, we integrated plasmid pWG at the URAB locus in strain J16D. Plasmid pWG was constructed in two steps. First, a 4.2 kb BamHI-Xhol DNA fragment containing the weel+ ORF was inserted into BamHI-Sall-digested pUC16URA3. The BamHl site is im- mediately upstream of the weel+ ORF. Plasmid pUC16URA3 has a 1.2 kb Hindlll fragment containing URA.3 ligated into the Hindlll site of a modified pUC16, in which the Smal and Pstl sites in the polylinker have been removed. The 0.7 kb BamHI-EcoRI GAL7 promoter frag- ment was subsequently ligated into the plasmid so that it directs ex- pression of weel+. Integration of pWG into the genome of J16D was targeted by cutting at the Smal site inside the URA3 fragment. The genotype of J16D is : MATa frp1 leu2 ura3 his-.
To construct plasmid pWG-L596, the oligonucleotide S-AGTATG- CAGTACTAAAGCTCAAG-3’was used to introduce a Seal restriction en- zyme site (underlined) and to change wee1 codon 596 from AAA-lysine to CTA-leucine. The method described by Kunkel (1965) was used.
To clone MIH7. an S. pombe c&25-22 leul-32 strain was trans- formed with an S. cerevisiae library constructed in the vector YEpl3 (Nasmyth and Reed, 1960), which carries the S. cerevisiae LEUPgene that complements the S. pombe leu7-32 mutation. The transformants were initially grown at the permissive temperature of 25OC, and then shifted to the restrictive temperature of 35OC.
Toconstruct pmihl::LEUP, the4 kb EcoRI-BamHIfragmentcontain- ing M/HI was cloned into pBR322. followed by replacement of the 0.5 kb Bglll-Xhol fragment internal to the ORF with a 2.9 kb Bglll-Xhol fragment containing LEU2. Plasmid pmihl::LEU2 was digested with Nhel to generate a 4 kb fragment containing the mihl::LEU2 allele. This fragment was used to replace one copy of M/H7 in diploid strain J15D (al trpplltrpl leu211eu2 his-/his-adelladel).
Galactose Induction Experiments Yeast cells were grown in YPRAF (1% Difco Yeast Extract, 2% Difco Bactopeptone, 2% raffinose) at 30°C for approximately 16 hr until the cultures reached mid-log phase. Cell concentration was adjusted to approximately 1 x lo6 cells per ml with YPRAF, and then galactose was added to a final concentration of 2.5%. Cell number was moni- tored by diluting samples into an isotonic saline solution and counting in a Coulter Cell Counter Model ZM. Cell size distribution was mea- sured by the Single Threshold Analysis Technique (Coulter Counter Model ZM Product Reference Manual, 4235160). To measure the rela- tive cell size of mihl:tLEUP mutants and congenic M/H7 strains, a Bec- ton Dickinson FACS IV Analyzer was used. Cells were prepared for flow cytometry by the method of Hutter and Eipel (1979) using propidium iodide.
Cell-Cycle Arrest Point and Reciprocal Shift Experiments In the reciprocal shift experiments with strain J lGD-WGCD, cultures were grown at 30°C. Hydroxyurea was used at 0.2 M and nocodazole at 10 vglml. Where appropriate, weel+ was induced by adding galac- tose to 1% to cultures grown in 2% raffinose (YPRAF). To repress weeI+, dextrose was added to 1% to a galactose-induced culture. For shifts from either hydroxyurea or nocodazole to galactose, galactose was added when the culture was uniformly arrested as determined by microscopic observation. After 1 hr of induction or mock induction (no galactose), the drug was washed out by centrifugation and cells were resuspended in the appropriate prewarmed medium. The time course of arrest in response to hydroxyurea was extended because high con- centrations of hydroxyurea retard growth. In shifts from galactose to ei- ther hydroxyurea or nocodazole, the drug was added simultaneously with 1% dextrose. Controls without the drug received only dextrose. Cells were observed using a Leitz SM-Lux phase-contrast microscope and a hemacytometer after fixing in 10% formalin and disaggregating using a Braun sonifier.
Acknowledgments
We thank Bruce Edgar, Jim Hicks, Paul Nurse, Aresa Toukdarian, and Curt Wittenburg for their advice, Don McQuitty for assistance with flow cytometry, and Matt Schibler for assistance with photomicroscopy. P R. was supported by a fellowship from The Imperial Cancer Re-
search Fund and received funding from the Research Institute of Scripps Clinic and National Institutes of Health grant GM41261; S. M. was supported by a fellowship from the Consejo Superior de Investiga- ciones Cientificas; and S. I. R. was supported by NIH grant GM36326.
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received December 2, 1969; revised February 3, 1969.
References
Arion, D., Meijer, L., Brizuela, L., and Beach, D. (1966). cdc2 is a com- ponent of the M phase-specific histone Hl kinase: evidence for iden- tity with MPF. Cell 55, 371-376.
Beach, D.. Durkacz, B., and Nurse, P (1962). Functionally homologous cell cycle control genes in budding and fission yeast. Nature 300, 706-709.
Brizuela. L., Draetta, G., and Beach, D. (1967). ~13 sucl acts in the fis- sion yeast cell division cycle as a component of the ~34~“~’ protein ki- nase. EMBO J. 6. 3507-3514. Draetta, G., Brizuela, L., Potashkin, J., and Beach, D. (1987). Identifica- tion of ~34 and ~13. human homologs of the cell cycle regulators of fis- sion yeast encoded by cdc2+ and sucl+. Cell 50, 319-325.
Dunphy, W. G., Brizuela, L., Beach, D.. and Newport, J. (1988). The Xenopus cdc2 protein is a component of MPF, a cytoplasmic regulator of mitosis. Cell 54, 423-431.
Gautier, J., Norbury, C., Lohka, M., Nurse, P., and Mailer, J. (1988). Puri- fied maturation-promoting factor contains the product of the Xenopus homolog of the fission yeast cell cycle control gene cdc2+. Cell 54, 433-439.
Goebl, M., and Byers. B. (1988). Cyclin in fission yeast. Cell 54, 739-740.
Hunter, T., and Cooper, J. A. (1985). Protein-tyrosine kinases. Annu. Rev. Biochem. 54, 897-930.
Hutter, K.-J., and Eipel. H. E. (1979). DNA determination of yeast by flow cytometry. J. Gen. Microbial. 773, 369-375.
Kataoka, T., Powers, S., McGill, C., Fasono. O., Strathern. J., Broach, J., and Wigler, M. (1984). Genetic analysis of yeast RASl and RAS2 genes. Cell 37; 437-445.
Kunkel. T. A. (1985). Rapid and efficient site-specific mutagenesis with- out phenotypic selection. Proc. Natl. Acad. Sci. USA 82, 488-492.
Labbe, J. C., Lee, M. G., Nurse, P., and Doree, M. (1988). Activation at M-phase of a protein kinase encoded by a starfish homologue of the cell cycle control gene cd&. Nature 335, 251-254.
Lee, M. G., and Nurse, P (1987). Complementation used to clone a hu- man homologue of the fission yeast cell cycle control gene cdc2. Na- ture 327, 31-35. Nasmyth, K. A., and Reed, S. I. (1980). The isolation of genes by com- plementation in yeast: the molecular cloning of a cell cycle control gene. Proc. Natl. Acad. Sci. USA 77, 2119-2123.
Nurse, l? (1985). Cell cycle control genes in yeast. Trends Genet. 7, 51-55. Piggott, J. R.. Rai, R., and Carter, B. L. A. (1982). A bifunctional gene product involved in two phases of the yeast cell cycle. Nature 298. 391-393. Pringle, J. R., and Hartwell, L. H. (1981). The Saccharomyces cere- visiae cell cycle. In The Molecular Biology of the Yeast Saccharomyces cerevisiae: Life Cycle and Inheritance, J. N. Strathern, E. W. Jones, and J. R. Broach, eds. (Cold Spring Harbor, New York: Cold Spring Harbor Laboratory), pp. 97-142. Russell, I? (1989). Gene cloning and expression in fission yeast. In The Molecular Biology of the Fission Yeast, A. Nasim, P Young, and B. F. Johnson, eds. (San Diego, California: Academic Press), in press. Russell, P., and Nurse, I? (1986a). cdc25+ functions as an inducer in the mitotic control of fission yeast. Cell 45. 145-153. Russell, i?, and Nurse, P. (198613). Schizosaccharomyces pombe and Saccharomyces cerevisiae: a look at yeasts divided. Cell 45, 781-782.

cdc25+-weel+ Mitotic Control 303
Russell, P, and Nurse, P (1987a). Negative regulation of mitosis by wee7 +, a gene encoding a protein kinase homolog. Cell 49, 559-567
Russell, P, and Nurse, f? (1987b). The mitotic inducer niml+ functions in a regulatory network of protein kinase homologs controlling the initi- ation of mitosis. Cell 49, 569-576.
Solomon, M., Booher, Ft., Kirschner, M., and Beach, D. (1988). Cyclin in fission yeast. Cell 54, 738-739.
Swenson, K. I., Farrell, K. M., and Ruderman, J. V. (1986). The clam embryo protein cyclin A induces entry into M phase and the resump- tion of meiosis in Xenopus oocytes. Cell 47, 881-870.
Tatchell, K., Chaleff, D. T., DeFoe-Jones, D., and Scolnick, E. M. (1984). Requirement of either of a pair of ras-related genes of Saccharomyces cerevisiae for spore viability. Nature 309, 523-527.
Toda. T., Cameron, S., Sass, P, Zoller, M., and Wigler, M. (1987). Three different genes in S. cerevisiae encode the catalytic subunits of the CAMP-dependent protein kinase. Cell 50, 277-287.
Weinert, T. A., and Hartwell, L. H. (1988). The RAD9 gene controls the cell cycle response to DNA damage in Saccharomyces cerevisiae. Science 247. 317-322.
Copyright © 2022 FDOKUMEN




















