
Inn1 couples contraction of the actomyosin ring to membrane ingression during cytokinesis in budding...
23
© 2008 Nature Publishing Group ARTICLES Inn1 couples contraction of the actomyosin ring to membrane ingression during cytokinesis in budding yeast Alberto Sanchez-Diaz 1 , Vanessa Marchesi 1 , Stephen Murray 1 , Richard Jones 2,3 , Gislene Pereira 1,4 , Ricky Edmondson 2,5 , Terry Allen 1 and Karim Labib 1.6 By rapidly depleting each of the essential budding yeast proteins of unknown function, we identified a novel factor that we call Inn1, which associates with the contractile actomyosin ring at the end of mitosis and is needed for cytokinesis. We show that Inn1 has a C2 domain at the amino terminus of the protein that is required for ingression of the plasma membrane, whereas the remainder of the protein recruits Inn1 to the actomyosin ring. The lethal effects of deleting the INN1 gene can be suppressed by artificial fusion of the C2 domain to other components of the actomyosin ring, restoring membrane ingression on contraction of the actomyosin ring. Our data indicate that recruitment of the C2 domain of Inn1 to the contractile actomyosin ring is crucial for ingression of the plasma membrane during cytokinesis in budding yeast. The process of cytokinesis whereby a eukaryotic cell divides in two provides a unique series of challenges for the regulation of membrane dynamics 1,2 . In animal cells and fungi, the plasma membrane extends into the cytoplasm at the end of mitosis in a process called ingression, and division of the cytoplasm is then completed by a step termed abscis- sion. Ingression of the plasma membrane must be regulated in time and space to ensure that cell division takes place after nuclear division has been completed, and occurs between the two divided nuclei. Before the end of mitosis, a contractile ring of actin and type II myosin assembles underneath the plasma membrane, usually in the middle of the cell, and this defines the site at which ingression will occur 3–5 . The contractile ring contains many other factors in addition to actin and type II myosin, including adaptor proteins that control the recruitment of actin and of other components 6,7 . It is unclear how the contractile ring is attached to the plasma membrane, but this could involve the interaction of ring proteins with peripheral or integral membrane proteins, as well as with membrane lipids. Contraction of the actomyosin ring is thought to cause ingression of the membrane 1,2,8 , but the underlying mechanisms remain unclear and previous studies have not identified factors that are specifically required for this stage of cytokinesis. To try to identify novel factors that are required for cell division in budding yeast, we began a systematic screen for new cell-cycle proteins. Previous work identified all the proteins that are essential for viability in Saccharomyces cerevisiae 9,10 , and we focused on the subset of these proteins that were of unknown function. We previously reported the first part of this work, resulting in the identification of new proteins required for chromosome replication 11 . Here, we describe the completion of the screen, leading to the identification of a novel factor that is required specifically for ingression of the plasma membrane during cytokinesis. RESULTS A novel cytokinesis factor identified by functional proteomics For each of the remaining essential proteins of unknown function, a yeast strain was generated in which the ‘heat-inducible degron’ cassette was fused to the amino terminus of the target protein 12 . This allowed rapid inactivation of the fusion protein at 37°C after expression of the E3 ubiquitin ligase UBR1 from the GAL1,10 promoter 13 (see Supplementary Information, Fig. 1a). An asynchronous culture of each strain was grown at 24°C and the consequences of rapidly depleting the target protein at 37°C were then examined (see Supplementary Information, Fig. 1b). Inactivation of four proteins caused a defect in mitosis, and others identified these same proteins as the kinetochore components Dad1, Ask1, Nsl1 and Dsn1 (refs 14–20). Inactivation of Ynl152w, which we named Inn1 (as it is required for Ingression), produced a distinct phenotype. The nuclear cycle of S-phase and mitosis continued in the absence of cell division, producing multi-budded cells with a DNA content of 2C, 4C or 8C (Fig. 1a, b). 1 Cancer Research U.K., Paterson Institute for Cancer Research, University of Manchester, Wilmslow Road, Manchester, M20 4BX, UK. 2 FDA-NCTR, 3900 NCTR Road, Jefferson, AR 72079, USA. 3 Current address: NextGen Sciences, Inc., 4401 Varsity Drive, Suite E, Ann Arbor, MI 48108, USA. 4 Current address: German Cancer Research Centre, Im Neuenheimer Feld 581, 69120 Heidelberg, Germany. 5 Current address: Myeloma Institute for Research and Therapy, University of Arkansas for Medical Sciences, 4301 W.Markham #776, Little Rock, AR 72205, USA. 6 Correspondence should be addressed to K.L. (email: [email protected]) Received 8 January 2008; accepted 21 February 2008; published online 16 March 2008; DOI: 10.1038/ncb1701 NATURE CELL BIOLOGY VOLUME 10 | NUMBER 4 | APRIL 2008 395
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Inn1 couples contraction of the actomyosin ring to membrane ingression during cytokinesis in budding...

© 2008 Nature Publishing Group
A RT I C L E S
Inn1 couples contraction of the actomyosin ring to membrane ingression during cytokinesis in budding yeastAlberto Sanchez-Diaz1, Vanessa Marchesi1, Stephen Murray1, Richard Jones2,3, Gislene Pereira1,4, Ricky Edmondson2,5, Terry Allen1 and Karim Labib1.6
By rapidly depleting each of the essential budding yeast proteins of unknown function, we identified a novel factor that we call Inn1, which associates with the contractile actomyosin ring at the end of mitosis and is needed for cytokinesis. We show that Inn1 has a C2 domain at the amino terminus of the protein that is required for ingression of the plasma membrane, whereas the remainder of the protein recruits Inn1 to the actomyosin ring. The lethal effects of deleting the INN1 gene can be suppressed by artificial fusion of the C2 domain to other components of the actomyosin ring, restoring membrane ingression on contraction of the actomyosin ring. Our data indicate that recruitment of the C2 domain of Inn1 to the contractile actomyosin ring is crucial for ingression of the plasma membrane during cytokinesis in budding yeast.
The process of cytokinesis whereby a eukaryotic cell divides in two provides a unique series of challenges for the regulation of membrane dynamics1,2. In animal cells and fungi, the plasma membrane extends into the cytoplasm at the end of mitosis in a process called ingression, and division of the cytoplasm is then completed by a step termed abscis-sion. Ingression of the plasma membrane must be regulated in time and space to ensure that cell division takes place after nuclear division has been completed, and occurs between the two divided nuclei.
Before the end of mitosis, a contractile ring of actin and type II myosin assembles underneath the plasma membrane, usually in the middle of the cell, and this defines the site at which ingression will occur3–5. The contractile ring contains many other factors in addition to actin and type II myosin, including adaptor proteins that control the recruitment of actin and of other components6,7. It is unclear how the contractile ring is attached to the plasma membrane, but this could involve the interaction of ring proteins with peripheral or integral membrane proteins, as well as with membrane lipids. Contraction of the actomyosin ring is thought to cause ingression of the membrane1,2,8, but the underlying mechanisms remain unclear and previous studies have not identified factors that are specifically required for this stage of cytokinesis.
To try to identify novel factors that are required for cell division in budding yeast, we began a systematic screen for new cell-cycle proteins. Previous work identified all the proteins that are essential for viability in Saccharomyces cerevisiae9,10, and we focused on the subset of these
proteins that were of unknown function. We previously reported the first part of this work, resulting in the identification of new proteins required for chromosome replication11. Here, we describe the completion of the screen, leading to the identification of a novel factor that is required specifically for ingression of the plasma membrane during cytokinesis.
ResulTsA novel cytokinesis factor identified by functional proteomicsFor each of the remaining essential proteins of unknown function, a yeast strain was generated in which the ‘heat-inducible degron’ cassette was fused to the amino terminus of the target protein12. This allowed rapid inactivation of the fusion protein at 37°C after expression of the E3 ubiquitin ligase UBR1 from the GAL1,10 promoter13 (see Supplementary Information, Fig. 1a).
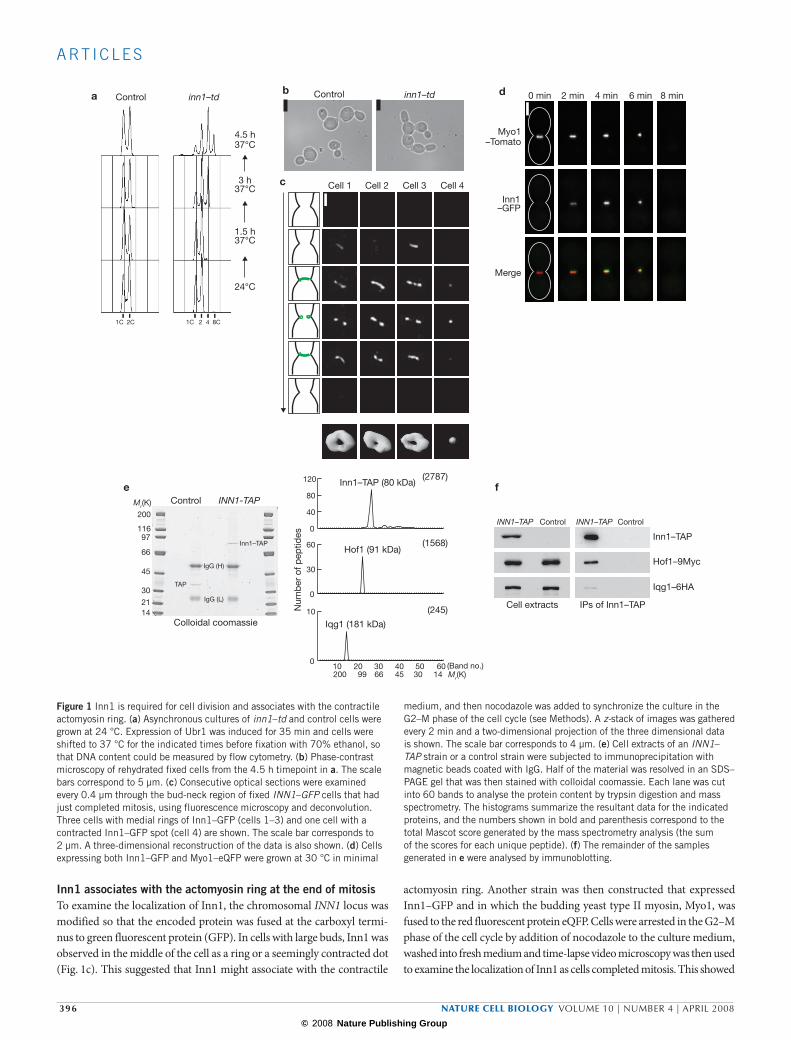
An asynchronous culture of each strain was grown at 24°C and the consequences of rapidly depleting the target protein at 37°C were then examined (see Supplementary Information, Fig. 1b). Inactivation of four proteins caused a defect in mitosis, and others identified these same proteins as the kinetochore components Dad1, Ask1, Nsl1 and Dsn1 (refs 14–20). Inactivation of Ynl152w, which we named Inn1 (as it is required for Ingression), produced a distinct phenotype. The nuclear cycle of S-phase and mitosis continued in the absence of cell division, producing multi-budded cells with a DNA content of 2C, 4C or 8C (Fig. 1a, b).
1Cancer Research U.K., Paterson Institute for Cancer Research, University of Manchester, Wilmslow Road, Manchester, M20 4BX, UK. 2FDA-NCTR, 3900 NCTR Road, Jefferson, AR 72079, USA. 3Current address: NextGen Sciences, Inc., 4401 Varsity Drive, Suite E, Ann Arbor, MI 48108, USA. 4Current address: German Cancer Research Centre, Im Neuenheimer Feld 581, 69120 Heidelberg, Germany. 5Current address: Myeloma Institute for Research and Therapy, University of Arkansas for Medical Sciences, 4301 W.Markham #776, Little Rock, AR 72205, USA.6Correspondence should be addressed to K.L. (email: [email protected])
Received 8 January 2008; accepted 21 February 2008; published online 16 March 2008; DOI: 10.1038/ncb1701
nature cell biology volume 10 | number 4 | APrIl 2008 395
© 2008 Nature Publishing Group
A RT I C L E S
Inn1 associates with the actomyosin ring at the end of mitosisTo examine the localization of Inn1, the chromosomal INN1 locus was modified so that the encoded protein was fused at the carboxyl termi-nus to green fluorescent protein (GFP). In cells with large buds, Inn1 was observed in the middle of the cell as a ring or a seemingly contracted dot (Fig. 1c). This suggested that Inn1 might associate with the contractile
actomyosin ring. Another strain was then constructed that expressed Inn1–GFP and in which the budding yeast type II myosin, Myo1, was fused to the red fluorescent protein eQFP. Cells were arrested in the G2–M phase of the cell cycle by addition of nocodazole to the culture medium, washed into fresh medium and time-lapse video microscopy was then used to examine the localization of Inn1 as cells completed mitosis. This showed
b Control inn1–tda Control inn1–td
1C 2C 1C 2 4 8C
24°C
1.5 h37°C
3 h37°C
4.5 h37°C
d
f
0 min 2 min 4 min 6 min 8 min
Myo1–Tomato
Inn1–GFP
Merge
Cell 1 Cell 2 Cell 3 Cell 4c
eINN1-TAPControl
200
11697
66
45
30
2114
Inn1–TAP
TAP
IgG (H)
IgG (L)
Colloidal coomassie
Inn1–TAP
Cell extracts
Hof1–9Myc
Iqg1–6HA
IPs of Inn1–TAP
INN1–TAP Control INN1–TAP Control
Inn1–TAP (80 kDa)(2787)
0
40
80
120
Hof1 (91 kDa)
0
30
60 (1568)
Iqg1 (181 kDa)
10 20 30 40 50 60 (Band no.)200 99 66 45 30 14
(245)
0
10Num
ber
of p
eptid
es
Mr(K)
Mr(K)
Figure 1 Inn1 is required for cell division and associates with the contractile actomyosin ring. (a) Asynchronous cultures of inn1–td and control cells were grown at 24 °C. Expression of Ubr1 was induced for 35 min and cells were shifted to 37 °C for the indicated times before fixation with 70% ethanol, so that DNA content could be measured by flow cytometry. (b) Phase-contrast microscopy of rehydrated fixed cells from the 4.5 h timepoint in a. The scale bars correspond to 5 µm. (c) Consecutive optical sections were examined every 0.4 µm through the bud-neck region of fixed INN1–GFP cells that had just completed mitosis, using fluorescence microscopy and deconvolution. Three cells with medial rings of Inn1–GFP (cells 1–3) and one cell with a contracted Inn1–GFP spot (cell 4) are shown. The scale bar corresponds to 2 µm. A three-dimensional reconstruction of the data is also shown. (d) Cells expressing both Inn1–GFP and Myo1–eQFP were grown at 30 °C in minimal
medium, and then nocodazole was added to synchronize the culture in the G2–M phase of the cell cycle (see Methods). A z-stack of images was gathered every 2 min and a two-dimensional projection of the three dimensional data is shown. The scale bar corresponds to 4 µm. (e) Cell extracts of an INN1–TAP strain or a control strain were subjected to immunoprecipitation with magnetic beads coated with IgG. Half of the material was resolved in an SDS–PAGE gel that was then stained with colloidal coomassie. Each lane was cut into 60 bands to analyse the protein content by trypsin digestion and mass spectrometry. The histograms summarize the resultant data for the indicated proteins, and the numbers shown in bold and parenthesis correspond to the total Mascot score generated by the mass spectrometry analysis (the sum of the scores for each unique peptide). (f) The remainder of the samples generated in e were analysed by immunoblotting.
396 nature cell biology volume 10 | number 4 | APrIl 2008
© 2008 Nature Publishing Group
A RT I C L E S
that Inn1 was recruited to the actomyosin ring just before contraction, and disappeared with similar kinetics to Myo1 (Fig. 1d and see Supplementary Information, Movie 1; 20 cells examined). The observed colocalization of the GFP and eQFP signals was equivalent to that observed in a diploid strain expressing both Myo1–eQFP and Myo1–GFP (data not shown).
To determine whether the Inn1 protein associates with components of the actomyosin ring in vivo, a yeast strain was generated in which Inn1 was fused to the tandem affinity purification (TAP) tag. The resultant fusion protein was isolated from cell extracts and mass spectrometry was used to identify proteins that copurified specifically with Inn1–TAP (Fig. 1e). The most abundant such factor was the Hof1 protein, which was previously found to interact with the actomyosin ring21,22. In addi-tion, Inn1–TAP copurified with a second component of the actomyosin ring, the IQGAP protein called Iqg1 or Cyk1 (Fig. 1e). These interactions were confirmed subsequently by immunoblotting (Fig. 1f).
To determine whether the localization of Inn1 is dependent on other components of the actomyosin ring, yeast strains were generated in which Myo1 or Iqg1 were fused to the degron cassette. We first con-firmed that Myo1 and Iqg1 were essential in the budding yeast strain
used for our experiments. Spores that lack Myo1, Iqg1 or Inn1 were not able to form colonies (see Supplementary Information, Fig. S2a) and died after germination as chains of a few cells (see Supplementary Information, Fig. S2b), indicating a defect in cell division.
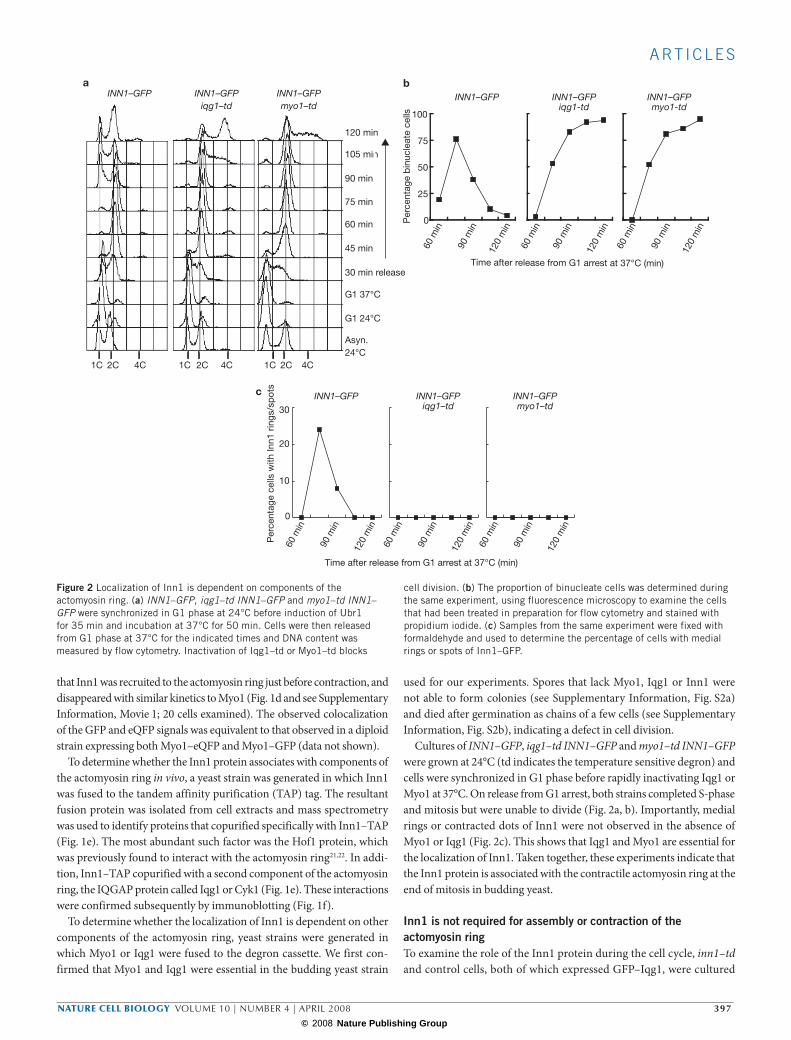
Cultures of INN1–GFP, iqg1–td INN1–GFP and myo1–td INN1–GFP were grown at 24°C (td indicates the temperature sensitive degron) and cells were synchronized in G1 phase before rapidly inactivating Iqg1 or Myo1 at 37°C. On release from G1 arrest, both strains completed S-phase and mitosis but were unable to divide (Fig. 2a, b). Importantly, medial rings or contracted dots of Inn1 were not observed in the absence of Myo1 or Iqg1 (Fig. 2c). This shows that Iqg1 and Myo1 are essential for the localization of Inn1. Taken together, these experiments indicate that the Inn1 protein is associated with the contractile actomyosin ring at the end of mitosis in budding yeast.
Inn1 is not required for assembly or contraction of the actomyosin ringTo examine the role of the Inn1 protein during the cell cycle, inn1–td and control cells, both of which expressed GFP–Iqg1, were cultured
aINN1–GFP
1C 2C 4C
Asyn.24°C
G1 24°C
30 min release
45 min
60 min
75 min
90 min
105 min
120 min
G1 37°C
0
25
50
75
100
Time after release from G1 arrest at 37°C (min)
b
120
min
90 m
in
60 m
in
120
min
90 m
in
60 m
in
120
min
90 m
in
60 m
in12
0 m
in
90 m
in
60 m
in
120
min
90 m
in
60 m
in
120
min
90 m
in
60 m
in
0
10
20
30
Per
cent
age
cells
with
Inn1
rin
gs/s
pot
s c
1C 2C 4C
INN1–GFPiqg1–td
1C 2C 4C
INN1–GFPmyo1–td
INN1–GFP INN1–GFPiqg1-td
INN1–GFPmyo1-td
INN1–GFP INN1–GFPiqg1–td
INN1–GFPmyo1–td
Per
cent
age
bin
ucle
ate
cells
Time after release from G1 arrest at 37°C (min)
Figure 2 Localization of Inn1 is dependent on components of the actomyosin ring. (a) INN1–GFP, iqg1–td INN1–GFP and myo1–td INN1–GFP were synchronized in G1 phase at 24°C before induction of Ubr1 for 35 min and incubation at 37°C for 50 min. Cells were then released from G1 phase at 37°C for the indicated times and DNA content was measured by flow cytometry. Inactivation of Iqg1–td or Myo1–td blocks
cell division. (b) The proportion of binucleate cells was determined during the same experiment, using fluorescence microscopy to examine the cells that had been treated in preparation for flow cytometry and stained with propidium iodide. (c) Samples from the same experiment were fixed with formaldehyde and used to determine the percentage of cells with medial rings or spots of Inn1–GFP.
nature cell biology volume 10 | number 4 | APrIl 2008 397

© 2008 Nature Publishing Group
A RT I C L E S
and synchronized in G1 phase at 24°C. Inn1 was inactivated in the degron strain and the cells were then released from G1 arrest at 37°C. Bud formation, chromosome replication and mitosis were not affected by the absence of Inn1, but cells were subsequently unable to divide (Fig. 3a). Recruitment of actin to the contractile ring did not require Inn1 (Fig. 3b). Moreover, medial rings and contracted dots of Iqg1 were observed with similar kinetics in both strains (Fig. 3b). These findings indicated that Inn1 is not required for assembly of the
actomyosin ring or for the initiation of contraction. However, the pro-portion of cells with contracted dots of Iqg1 was higher in the absence of Inn1 (Fig. 3b), suggesting that the final stages of contraction or disassembly of the actomyosin ring might take slightly longer in the absence of Inn1. To confirm that the localization of Iqg1 is indeed dependent on the actomyosin ring, we showed that rapid inactivation of Myo1–td prevents medial localization of Iqg1 (see Supplementary Information, Fig. S3).
c
Control
inn1–td
0 min 1 min 2 min 3 min 4 min 5 min 6 min
inn1–td
Start End
d
0
20
40
60
80
CDC12–GFP
0
25
50
75
100
0
25
50
75
100
inn1–td CDC12-GFPBinucleate cells Binucleate cells
Time after release from G1 arrest at 37°C (min)
Cdc12–GFP Phase
CDC12–GFP
inn1–td CDC12–GFP
bGFP–IQG1 inn1–td GFP–IQG1
Time after release from G1 arrest at 37°C (min)
0
10
20
30
105
min
75 m
in
60 m
in
Actin rings
90 m
in10
5 m
in12
0 m
in
75 m
in
60 m
in
90 m
in
105
min
120
min
75 m
in
60 m
in
90 m
in
105
min
120
min
75 m
in
60 m
in
90 m
in
105
min
120
min
75 m
in
60 m
in
90 m
in
105
min
120
min
75 m
in
60 m
in
90 m
in
105
min
120
min
75 m
in
60 m
in
90 m
in
105
min
120
min
75 m
in
60 m
in
90 m
in
105
min
120
min
75 m
in
60 m
in
90 m
in
105
min
75 m
in
60 m
in
90 m
in
Actin rings
0
10
20
30
40
50
Iqg1 spots Iqg1 spotsIqg1 rings Iqg1 rings
GFP–IQG1 inn1–td GFP–IQG1
Per
cent
age
cells
Per
cent
age
cells
a
Asyn.24°C
G1 24°C
30 min release
45 min
60 min
75 min
90 min
105 min
120 min
G1 37°C
1C 2C 4C 1C 2C 4C
GFP–IQG1 inn1–td GFP–IQG1
Asyn.24°C
G1 24°C
30 min release
45 min
60 min
75 min
90 min
105 min
120 min
G1 37°C
Binucleate cells
0
25
50
75
100Binucleate cells
Time after release from G1 arrest at 37°C (min)
**
Per
cent
age
cells
Per
cent
age
cells
Per
cent
age
cells
Split septin rings Split septin rings
Figure 3 Inn1 is not required for assembly or contraction of the actomyosin ring, or for correct localization of the septin component Cdc12. (a) GFP–IQG1 and inn1–td GFP–IQG1 were grown and analysed as described in Fig. 2. The budding index was determined by bright-field microscopy and the asterisk denotes the first timepoint at which 95% of the cells had budded. (b) Samples from the same experiment were fixed with formaldehyde and used to determine the proportion of cells with actin rings (by staining with rhodamine–phalloidin), or with either medial rings or spots of GFP–Iqg1. Examples of the latter are shown in the lower panels. The scale bar corresponds to 4 µm. (c) Control and inn1–td cells expressing both Spc42–eQFP and Myo1–Tomato were synchronized in G2–M phase at 37°C after inactivation of Inn1–td. Cells were then washed into minimal medium and processed for time-lapse video microscopy as in Fig. 1d, except that the
temperature was maintained at 37 °C. A z-stack of images was gathered every minute and a two-dimensional projection of the three-dimensional data that correspond to a region around the actomyosin ring is shown. The scale bar corresponds to 1 µm. A grey circle denotes the timepoint at which the actomyosin ring started to contract and a black circle marks the timepoint at which the contracted ring disappeared. (d) An analogous experiment to that in a was performed with CDC12–GFP and inn1–td CDC12–GFP strains, and fixed samples were used to determine the proportion of binucleate cells throughout the experiment, together with the proportion of cells with split rings of the septin protein Cdc12. Examples from the 90 min timepoint are shown of cells with split rings of Cdc12–GFP. The left panels show deconvolved images that correspond to a single optical section from the z-stack. Phase contrast images are shown in the right panels. The scale bar corresponds to 4 µm.
398 nature cell biology volume 10 | number 4 | APrIl 2008

© 2008 Nature Publishing Group
A RT I C L E S
Time-lapse video microscopy was then used to examine live cells in which progression through mitosis could be visualized at the same time as contraction of the actomyosin ring. The Spc42 component of spindle pole bodies was fused to eQFP in control or inn1–td cells that also expressed Myo1–Tomato. Cells were synchronized in G1 phase, Inn1–td was inactivated at 37°C, and cells were then released from G1 arrest in fresh medium at 37°C. After the cells had budded and completed S-phase, nocodazole was added to synchronize the cultures once again, this time in G2–M phase. The cells were then washed into minimal medium, transferred to a glass chamber at 37°C and examined every minute by time-lapse video microscopy. Twenty movies each were examined for control and inn1–td cells, and formation, contraction and disassembly of the actomyosin ring was observed in all cells at the end of mitosis (Fig. 3c and see Supplementary Information, Movie 2). The average period from the initiation of contraction to the final disappearance of the ring was
slightly longer in the inn1–td strain (a mean value of 6.6 min in inn1–td compared with 4.2 min in the control strain). This was caused by a slight delay in the final stages of disassembly of the ring, as the period from the initiation of contraction to the initial generation of a contracted ‘spot’ was similar in both strains (a mean value of 2.9 min for inn1–td, 2.7 min for the control). This finding agrees with the increased proportion of contracted Iqg1–GFP dots that was observed in the inn1–td strain in the experiment described above (Fig. 3b).
Taken together, these experiments demonstrate that Inn1 is not required for the actomyosin ring to form or subsequently to contract and disassemble. It was also confirmed that Inn1 is not required for assembly or splitting of the septin ring (Fig. 3d), which limits the diffusion of membrane proteins away from the cleavage site during cytokinesis23. We therefore considered whether Inn1 might be necessary for contraction of the ring to be coupled to ingression of the plasma membrane.
b = area bleached by laser
8 min 10 min 12 min 14 min 16 min
0 min 2 min 4 min 6 minControl
aControl Bud-neck
n
n
inn1–td Bud-neck
n
n
n
n
nn
n
n
*
0 min 4 min 6 mininn1–td
8 min 10 min 12 min 14 min 16 min
* * * * *
* * * * *
* * * * *
2 min
** * * *
Figure 4 Inn1 is required for division of the cytoplasm at the end of mitosis. (a) inn1–td and control cells were synchronized in G1 phase at 24°C, before induction of Ubr1 and incubation of cells at 37°C for 50 min to deplete the Inn1–td protein. Cells were then released from G1 arrest at 37°C for 105 min, and processed for transmission electron microscopy. By examining serial sections through each sample, cells that had just completed nuclear division (n indicates the position of the two divided nuclei) were identified. Control cells completed cytokinesis and formation of a new region of cell wall, whereas 100% of inn1–td cells retained a single cytoplasm, indicating
that the plasma membrane did not ingress during cytokinesis (serial sections through 50 cells examined). The scale bars correspond to 1 µm. The right-hand panels show an enlarged region of the bud-neck. (b) Control or inn1–td cells expressing GFP were grown as in Fig. 1a and incubated at 37 °C for 2.5 h. A laser was used to bleach a small region of the cytoplasm on one side of the cell (circle) for the indicated times. The GFP signal on the opposite side of the cell (marked by an asterisk) was measured throughout the experiment using ImageJ software. The GFP signal decreased by 25% in control cells and 90% in inn1–td cells. The scale bar corresponds to 5 µm.
nature cell biology volume 10 | number 4 | APrIl 2008 399

© 2008 Nature Publishing Group
A RT I C L E S
Inn1 is required for ingression of the plasma membraneTransmission electron microscopy was used to determine whether ingres-sion of the plasma membrane could occur in cells that complete mitosis after inactivation of Inn1. Cultures of inn1–td and control strains were synchronized, as described for the experiment in Fig. 3a, and samples were fixed 105 min after release from G1 arrest at 37°C (most control cells had just completed cytokinesis at this time). By analysing serial sec-tions of 60 nm through the samples, binucleate cells were identified and the region of the bud-neck was then examined. Control cells completed
cytokinesis and septation (Fig. 4a), whereas inn1–td cells retained a single cytoplasm, indicating that ingression of the plasma membrane did not occur (Fig. 4a; serial sections through fifty cells were examined).
To confirm this observation in live cells, two different approaches were used. First, GFP was expressed in control and inn1–td strains before incubating the cultures at 37°C so that the inn1–td cells failed to divide and subsequently formed a new bud in the next cell cycle. A laser was then used to repeatedly bleach the GFP signal in a small portion of the cytoplasm on one side of the cell. In control cells that had just completed
a GFP–RAS2 MYO1–TOMATO
Gre
enR
edM
erge
0 min 2 min 4 min 6 min 8 min 10 min
b0 min 2 min 4 min 6 min 8 min 10 min
inn1-td GFP-RAS2 MYO1-TOMATO SPC42-eQFP
Gre
enR
edM
erge
inn1–td GFP–RAS2 MYO1–TOMATO SPC42–eQFP0 min 2 min 4 min 6 min 8 min 10 min
Gre
enR
edM
erge
Start End
Start End
Start End
Figure 5 Inn1 is required for ingression of the plasma membrane during cytokinesis. (a, b) GFP–RAS2 MYO1–TOMATO (a) and inn1–td GFP–RAS2 MYO1–TOMATO SPC42–eQFP (b) were grown as described in Fig. 3c and released from G2–M arrest into minimal medium. The cultures were then mixed, transferred to a timelapse chamber at 37 °C and a z-stack of images was gathered every 2 min for both GFP and TOMATO–eQFP signals. The data were deconvolved, and all optical sections (z-levels) were examined at each timepoint, to determine whether ingression of the plasma membrane occurred during
contraction and disassembly of the actomyosin ring. A single optical section for the GFP signal that corresponds to the region of the bud-neck and a two dimensional projection of the three-dimensional data for the TOMATO and eQFP signals (so that the spindle poles can be observed as well as the actomyosin ring) are shown. A region of ingressed plasma membrane was observed in control cells (red arrows in a) after disappearance of the contracted actomyosin ring, but this was not observed after contraction and disassembly of the actomyosin ring in the inn1–td strain (b). The scale bars correspond to 4 µm.
400 nature cell biology volume 10 | number 4 | APrIl 2008

© 2008 Nature Publishing Group
A RT I C L E S
cytokinesis, the GFP signal on the opposite side of the cell was largely unaffected throughout the experiment (Fig. 4b). However, a very dif-ferent result was observed in the inn1–td strain: bleaching of a small area in the newest bud caused depletion of the GFP signal throughout the cytoplasm (Fig. 4b; 30 cells examined). This shows that inactivation of Inn1–td prevents division of the cytoplasm, in agreement with the electron microscopy data.
Second, time-lapse video microscopy was used to examine the kinet-ics of ring contraction and membrane ingression in cells that expressed Myo1–Tomato, and in which the small G-protein Ras2 was fused to
GFP. Ras2 provides a useful marker in live cells for the plasma mem-brane throughout the cell cycle24,25. Control and inn1–td cells were grown as described for the experiment in Fig. 3c, so that the dynamics of the actomyosin ring and plasma membrane could be examined as the cells completed mitosis. To ensure that both strains were treated in an identi-cal fashion, the cultures were mixed before the cells were placed in the time-lapse chamber at 37°C (the inn1–td cells expressed Spc42–eQFP and could thus be distinguished from the control). A series of optical sections through the cells were examined every 2 min to determine the behaviour of Myo1–Tomato and Ras2-coated plasma membrane at the
a Inn1 (Ynl152w)
1-
-409
71-
141-
211-
281-
351-
C2 domain
PXXP motif
c
Loop 1
Loop 3
1
23
4
5
6
7 8
dS. pombe (SPBC83.18c) C.glabrata (CAG60903)
C2 domain
PXXP motif
Inn1 C2 domain (type II) 1 2
7 8
3 4
65
SynIII C2 domain (type I)1 2 3 4 5
6 7 8
b “Loop 1”
“Loop 3”
10 20 30 40 50 60 70 80
90 100 110 120 130 140 150 160
Inn1C. glabrataA. gossypiiC. albicansS. pombeSynIII
11111
294
Inn1C. glabrataA. gossypiiC. albicansS. pombeSynIII
6969718565
686870846477
134134134153126127
* *
Figure 6 The Inn1 protein has a C2 domain at its N-terminus, and the rest of the protein is rich in PXXP motifs. (a) Amino-acid sequence of the budding yeast Inn1 protein. The first 134 amino acids correspond to the C2 domain, and remaining 275 amino acids until the C-terminus of the protein contain eight PXXP motifs (for comparison, note that the Myo1 protein comprises 1929 amino acids and yet only has two PXXP motifs). (b) Alignment of the C2 domain of Inn1 with the corresponding region of Inn1 orthologues from the indicated species (Candida glabrata protein CAG60903, Ashbya gossypii protein AAS53428; Candida albicans protein Ca019_10350 and Schizosaccharomyces pombe protein SPBC83.18c). The bottom sequence represents the C2 domain of the mouse protein synaptotagmin III (not an orthologue of Inn1). The black (SynIII) and green (Inn1) lines above the alignment indicate the location of the eight β-strands that typify C2 domains. For SynIII, these were observed in the crystal structure, whereas the secondary structure of the C2 domain of
Inn1 was predicted with the software PredictProtein45. The β-strands of C2 domains are arranged in one of two ways in the primary sequence without altering the tertiary structure: 1-2-3-4-5-6-7-8 (type I) or 2-3-4-5-6-7-8-1 (type II)46. Inn1 corresponds to the type II group of C2 domains and strands 1–7 of Inn1 thus correspond to strands 2–8 of synaptotagmin III (the latter is a type I C2 domain). The red asterisks mark two lysine residues in Loop 1 that were mutated to alanine (inn1-AA). (c) Three dimensional-structure of a type II C2 domain (phospholipase A2, PDB identification code 1BCI), illustrating that the top sheet of the β-sandwich is formed from strands 2-3-5-6 and the bottom sheet from strands 1-4-7-8, and Loops 1 and 3 protude from the top corner of the β-sandwich. The image was generated with PDB ProteinWorkshop 1.50 software. (d) Amino-acid sequence of the orthologues of Inn1 in Schizosaccharomyces pombe and Candida glabrata, showing the presence of the conserved C2 domain and the PXXP motifs.
nature cell biology volume 10 | number 4 | APrIl 2008 401
© 2008 Nature Publishing Group
A RT I C L E S
bud-neck during cytokinesis (ten movies were examined for each strain). Contraction and disassembly of the actomyosin ring in the control strain led to the formation of a region of ingressed plasma membrane (Fig. 5a; the yeast bud-neck is very narrow, so the ingressed membrane is most clearly observed when contraction of the ring is complete). However, in the inn1–td cells, contraction and disassembly of the actomyosin ring did not cause ingression of the plasma membrane, regardless of whether the actomyosin ring had contracted in a seemingly symmetrical fashion or had contracted asymmetrically (Fig. 5b). In the latter case, the ring seemed to remain attached to one side of the plasma membrane, but detached from the other side during contraction and disassembly (Fig. 5b). Taken together, these experiments show that Inn1 is required for ingression of the plasma membrane during cytokinesis in budding yeast.
essential roles for the C2 domain and the C-terminus of Inn1 during cytokinesisPsi-BLAST searches and secondary structure analysis were used to show that the amino terminal 134 amino acids of Inn1 correspond to a so-called ‘C2 domain’ (Fig. 6 a, b). Previous work has shown that C2 domains represent a unique class of membrane targeting module that is formed by a sandwich of two β-sheets, each containing four β-strands26
(Fig. 6c). Amino acids 135–409 of Inn1 are predicted to be relatively unstructured and contain eight PXXP motifs that may mediate the inter-action of Inn1 with other proteins that contain SH3 domains (Fig. 6a).
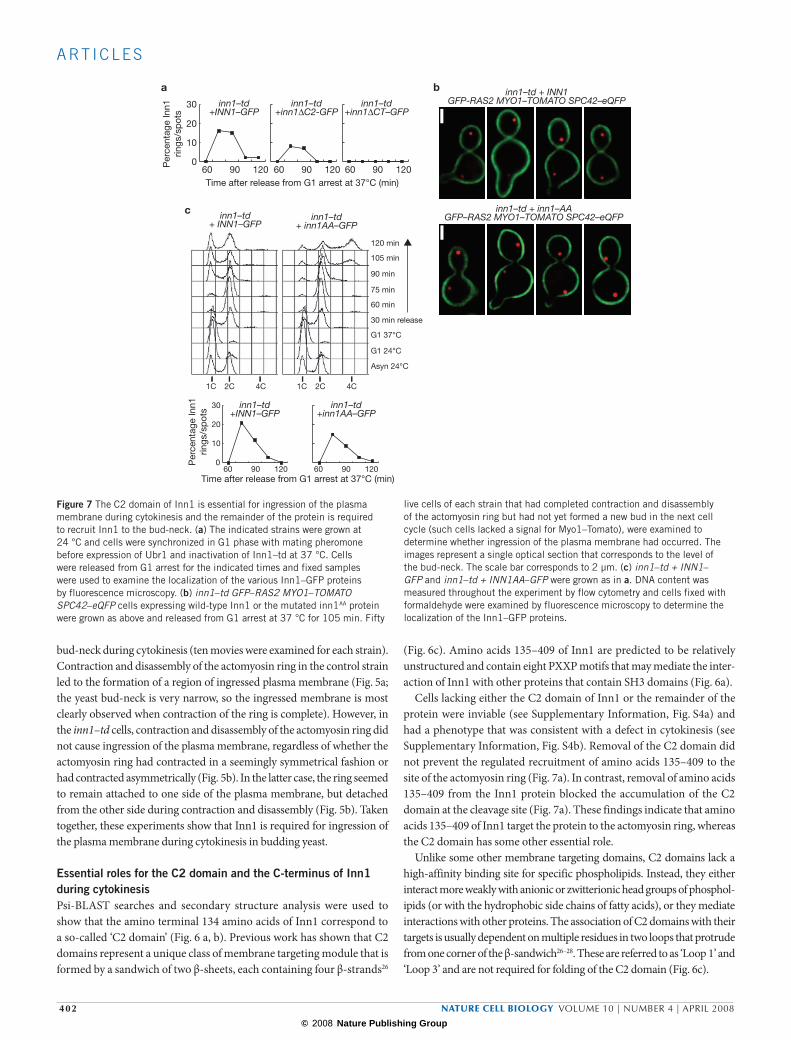
Cells lacking either the C2 domain of Inn1 or the remainder of the protein were inviable (see Supplementary Information, Fig. S4a) and had a phenotype that was consistent with a defect in cytokinesis (see Supplementary Information, Fig. S4b). Removal of the C2 domain did not prevent the regulated recruitment of amino acids 135–409 to the site of the actomyosin ring (Fig. 7a). In contrast, removal of amino acids 135–409 from the Inn1 protein blocked the accumulation of the C2 domain at the cleavage site (Fig. 7a). These findings indicate that amino acids 135–409 of Inn1 target the protein to the actomyosin ring, whereas the C2 domain has some other essential role.
Unlike some other membrane targeting domains, C2 domains lack a high-affinity binding site for specific phospholipids. Instead, they either interact more weakly with anionic or zwitterionic head groups of phosphol-ipids (or with the hydrophobic side chains of fatty acids), or they mediate interactions with other proteins. The association of C2 domains with their targets is usually dependent on multiple residues in two loops that protrude from one corner of the β-sandwich26–28. These are referred to as ‘Loop 1’ and ‘Loop 3’ and are not required for folding of the C2 domain (Fig. 6c).
c
Asyn 24°C
G1 24°C
G1 37°C
30 min release
60 min
75 min
90 min
105 min
120 min
inn1–td+ inn1AA–GFP
inn1–td+ INN1–GFP
1C 2C 4C 1C 2C 4C
0
10
20
30 inn1–td+INN1–GFP
60 90 120
inn1–td+inn1AA–GFP
60 90 120Time after release from G1 arrest at 37°C (min)
a
0
10
20
30
60 90 120 60 90 120 60 90 120Time after release from G1 arrest at 37°C (min)
inn1–td+INN1–GFP
inn1–td+inn1∆C2-GFP
inn1–td+inn1∆CT–GFP
Per
cent
age
Inn1
rings
/sp
ots
Per
cent
age
Inn1
rings
/sp
ots
b inn1–td + INN1GFP-RAS2 MYO1–TOMATO SPC42–eQFP
inn1–td + inn1–AAGFP–RAS2 MYO1–TOMATO SPC42–eQFP
Figure 7 The C2 domain of Inn1 is essential for ingression of the plasma membrane during cytokinesis and the remainder of the protein is required to recruit Inn1 to the bud-neck. (a) The indicated strains were grown at 24 °C and cells were synchronized in G1 phase with mating pheromone before expression of Ubr1 and inactivation of Inn1–td at 37 °C. Cells were released from G1 arrest for the indicated times and fixed samples were used to examine the localization of the various Inn1–GFP proteins by fluorescence microscopy. (b) inn1–td GFP–RAS2 MYO1–TOMATO SPC42–eQFP cells expressing wild-type Inn1 or the mutated inn1AA protein were grown as above and released from G1 arrest at 37 °C for 105 min. Fifty
live cells of each strain that had completed contraction and disassembly of the actomyosin ring but had not yet formed a new bud in the next cell cycle (such cells lacked a signal for Myo1–Tomato), were examined to determine whether ingression of the plasma membrane had occurred. The images represent a single optical section that corresponds to the level of the bud-neck. The scale bar corresponds to 2 µm. (c) inn1–td + INN1–GFP and inn1–td + INN1AA–GFP were grown as in a. DNA content was measured throughout the experiment by flow cytometry and cells fixed with formaldehyde were examined by fluorescence microscopy to determine the localization of the Inn1–GFP proteins.
402 nature cell biology volume 10 | number 4 | APrIl 2008

© 2008 Nature Publishing Group
A RT I C L E S
To examine the role of the C2 domain in more detail, two basic amino acids in Loop 1, which are well conserved in orthologues of Inn1 in other eukaryotic species, were mutated (Fig. 6b)26. Wild-type Inn1 or the mutated Inn1AA protein were expressesd in an inn1–td strain that also expressed
GFP–Ras2 and Myo1–Tomato. Both strains were then grown as described for the experiments in Fig. 3a and cells that had just completed contraction and disassembly of the actomyosin ring were examined. In cells express-ing wild-type Inn1, contraction and disassembly of the actomyosin ring
= inn1∆= C2–HOF1
a
INN1 / inn1∆HOF1 / C2(AA)-HOF1
b
INN1 / inn1∆HOF1 / C2-HOF1
= inn1∆= MYO1–C2
INN1 / inn1∆MYO1 / MYO1-C2
d CHS2-GFP MYO1-TOMATO
Gre
enR
ed
0 min 2 min 4 min 6 min 8 min 10 min
Mer
geG
reen
Red
0 min 2 min 4 min 6 min 8 min 10 min
Mer
geG
reen
Red
0 min 2 min 4 min 6 min 8 min 10 min
Mer
ge
Start End
Start End
Start End
Control inn1∆ C2–HOF1 inn1∆ MYO1–C2c
1C 2C 4C 1C 2C 4C 1C 2C 4C
C2–HOF1 inn1∆ CHS2–GFP MYO1–TOMATO SPC42–eQFP
C2-HOF1 inn1∆ CHS2-GFP MYO1-TOMATO SPC42-eQFP
Figure 8 Artificial recruitment of the C2 domain of Inn1 to the actomyosin ring is sufficient to allow ingression of the plasma membrane in cells lacking the INN1 gene. (a) Tetrad analysis of diploid INN1/inn1∆ HOF1/C2–HOF1 after 4 days growth at 24 °C and tetrad analysis of diploid INN1/inn1∆ HOF1/C2(AA) –HOF1 after 3 days growth at 24 °C. (b) Tetrad analysis of INN1/inn1∆ MYO1/MYO1–C2 after 4 days growth at 24 °C. (c) Asynchronous cultures of the indicated strains were grown at 24 °C. Cells were fixed with 70% ethanol and then either rehydrated and examined by phase contrast microscopy or processed for flow cytometry. The scale bars correspond to
5 µm. (d) CHS2–GFP MYO1–TOMATO and C2–HOF1 inn1∆ CHS2–GFP MYO1–TOMATO SPC42–eQFP were grown at 30 °C as described in Fig. 1d, and released from G2–M arrest into minimal medium. The cultures were then mixed and transferred to a time-lapse chamber. A z-stack of images was gathered every 2 min and a two-dimensional projection of the three-dimesional data is shown. The scale bars correspond to 4 µm. Red arrows denote the region of ingressed plasma membrane that was observed in both strains after disappearance of the contracted actomyosin ring. The grey and black circles denote the timing of ring contraction, as in Fig. 3c.
nature cell biology volume 10 | number 4 | APrIl 2008 403

© 2008 Nature Publishing Group
A RT I C L E S
was associated with ingression of the plasma membrane (Fig. 7b). In con-trast, ingression of the plasma membrane did not occur in cells expressing Inn1AA (Fig. 7b; 50 cells of each strain were examined). A similar experi-ment was performed with inn1–td cells that expressed either Inn1–GFP or Inn1AA–GFP. This confirmed that mutation of residues in Loop 1 blocked cell division, but did not prevent the mutated protein from accumulating at the cleavage site as cells completed mitosis (Fig. 7c). These experiments indicate that the C2 domain of Inn1 is required for membrane ingression during cytokinesis in budding yeast, but is not essential for recruitment of the Inn1 protein to the actomyosin ring at the end of mitosis.
Taken together, these findings indicate that amino acids 135–409 of Inn1 represent a module that targets the protein to the actomyosin ring, whereas amino acids 1–134 represent a C2 domain that is essential for membrane ingression during cytokinesis in budding yeast.
Artificial recruitment of the C2 domain to the actomyosin ring bypasses the requirement for the remainder of Inn1The simplest model for the function of Inn1 during cytokinesis would be that amino acids 135–409 target the protein to the actomyosin ring where the C2 domain then has an essential role during ingression of the plasma membrane. To test this hypothesis, we investigated whether artificial recruitment of the C2 domain to the actomyosin ring was suf-ficient to supply Inn1 function in cells lacking the INN1 gene. Diploid cells were created that lacked one copy of INN1 and in which one copy of either MYO1 or HOF1 had been modified so that the encoded protein was fused to the C2 domain of Inn1. The meiotic progeny of the resultant strains were then analysed by tetrad analysis (Fig. 8a, b). Remarkably, the lethal effects of deleting the INN1 gene could be suppressed by fusion of the C2 domain of Inn1 to the amino terminus of Hof1 (Fig. 8a; 100% of inn1∆C2–HOF1 spores formed viable colonies), and the resultant cells grew as rapidly as a wild-type strain and did not display any detectable defects in cell division (Fig. 8c). Fusion of the C2 domain of Inn1 to the C-terminus of Myo1 also restored viability to cells lacking the INN1 gene (Fig. 8b; 76% of inn1∆ MYO1-C2 spores were able to form viable colo-nies), although inn1∆ MYO1-C2 cells grew more slowly than wild-type cells and were partially defective in cell division (Fig. 8c). We confirmed that the function of Loop1 is still essential under these conditions, by showing that a version of the C2 domain with the ‘AA’ mutations in Loop 1 was not able to suppress deletion of the INN1 gene when fused to the amino terminus of Hof1 (Fig. 8a).
Time-lapse video microscopy was used to examine contraction of the actomyosin ring and ingression of the plasma membrane in cells expressing C2–Hof1 but lacking the endogenous INN1 gene. The chi-tin synthase Chs2 has multiple transmembrane domains and is trans-ported to the cleavage site in secretory vesicles at the end of mitosis, just before the actomyosin ring starts to contract29,30. After incorpora-tion into the plasma membrane, Chs2 accumulates adjacent to the actomyosin ring during contraction and disassembly of the latter. It then persists at the same site for a couple of minutes31, and provides a specific marker at this time for the ingressed portion of the plasma membrane. Subsequently, Chs2 is removed into endocytic vesicles before being degraded in the vacuole32. Chs2–GFP was recruited to the plasma membrane at the end of mitosis, both in control cells and also in inn1∆ cells that expressed the C2–Hof1 fusion protein, and then accumulated at the site of the contracting ring (Fig. 8d). Most importantly, Chs2–GFP was present in a region of ingressed plasma
membrane in both strains after disappearance of the contracted acto-myosin ring (Fig. 8d). This confirms that artificial recruitment of the C2 domain to the actomyosin ring is sufficient to support ingression of the plasma membrane in cells that lack the INN1 gene.
Taken together, these data indicate that recruitment of the C2 domain of Inn1 to the actomyosin ring is central to the process of membrane ingression during cytokinesis in budding yeast. The C-terminal 67% of the Inn1 protein is normally required for recruitment of the C2 domain to the actomyosin ring (Fig. 7a), but has no other essential role during cytokinesis.
DIsCussIOnOur data highlight a key role for the C2 domain of Inn1 during ingression of the plasma membrane in budding yeast, and provide the first evidence in any species to indicate that recruitment of a C2 domain to the actomyosin ring is important during cytokinesis. Assembly of the actomyosin ring does not require Inn1, indicating that Inn1 is not required to establish the link between the ring and the membrane. Instead, the C2 domain of Inn1 may help to preserve this link during contraction of the actomyosin ring, and thus con-tribute both to membrane ingression, as well as to stability of the contracting ring. Alternatively, Inn1 might induce curvature of the plasma membrane adjacent to the contracting ring, thereby promot-ing ingression of the membrane. A previous study showed that the C2 domain of the human protein synaptotagmin induces curvature in target membranes and thereby contributes to fusion of these mem-branes with synaptic vesicles33.
Inn1 interacts with the SH3 protein Hof1, which is a component of the actomyosin ring. The amino terminus of Hof1 forms a ‘PCH’ domain (‘pombe cdc15 homology’ – Hof1 denotes ‘homologue of cdc15’), which is also thought to represent a domain that can induce membrane curva-ture and deformation34. It will be interesting to determine how the PCH domain of Hof1 and the C2 domain of Inn1 may jointly contribute to the regulation of membrane dynamics during cytokinesis in budding yeast.
We showed that Schizosaccharomyces pombe and other yeasts contain an uncharacterized orthologue of Inn1 (Fig. 6b, d). Each protein has a C2 domain at the N-terminus (Fig. 6b), and the remainder of each orthologue is predicted to be relatively unstructured, with 8–11 PXXP motifs, analogous to amino acids 135–409 of Inn1. It will be of inter-est, therefore, to determine whether these proteins also play a role in cytokinesis and interact with the SH3 proteins that are the orthologues of Hof1 in budding yeast.
Ingression of the plasma membrane is poorly understood in mam-malian cells and, at present, it is unclear whether specific factors are required for this stage of cytokinesis. The anillin protein was origi-nally thought to play such a role35,36, but later studies contradicted this view, as depletion of anillin by siRNA in primate or Drosophila cells does not prevent the initiation of furrow ingression, but instead blocks cytokinesis at a later stage37,38,39,40. Mammalian genomes encode mul-tiple uncharacterized proteins with C2 domains, and it remains to be determined whether any of these play an analogous role to Inn1 during cytokinesis. Amino acids 135–409 of Inn1 are dispensable if the C2 domain is fused artificially to components of the actomyosin ring, and so it is possible that other eukaryotic species might recruit a C2 domain to the actomyosin ring by a mechanism that is different to that used by amino acids 135–409 of Inn1.
404 nature cell biology volume 10 | number 4 | APrIl 2008

© 2008 Nature Publishing Group
A RT I C L E S
Finally, it will be interesting to determine whether Inn1 is also required for abscission of the ingressed plasma membrane during cytokinesis. The localization of the protein would be consistent with such a role, but the requirement of Inn1 for ingression currently precludes us from deter-mining whether it is also needed for abscission.
MeThODsConstruction and growth of yeast strains. All the strains used in this study are based on the W303 genetic background and are described in the Supplementary Information, Table S1. Cells were grown in rich medium for all experiments except those involving time-lapse video microscopy, for which minimal medium was used either throughout or at the end of the experiment. Cells were arrested in the G1 phase of the cell cycle by addition of alpha factor mating pheromone to the medium at a final concentration of 7.5 µg ml–1, followed by incubation for at least one generation time. Cells were synchronized in G2–M phase by addition of nocodazole to the medium to a final concentration of 5 µg ml–1 for rich medium and 3.4 µg ml–1 for minimal medium.
Microscopy. To observe proteins that were fused to GFP, eQFP or Tomato, cells were either viewed live or were fixed with 8% formaldehyde, as specified in the figure legends. Time-lapse video microscopy was performed using a DeltaVision system with Olympus BX71 microscope and Photometrics Cascade II 512B EMCCD camera. The objective lens was a Zeiss alpha plan ×100 1.45 NA with a Chroma ET Sedat Quad filter set (89000). The illumination source was the Sutter 300w xenon system with liquid light guide, and Softworx acquisition software (Applied Precision Instruments) was used. Cells were grown in a BIOPTECHS Focht Chamber System 2 (FCS2, Photometrics UK) to allow precise control of temperature. The base of the chamber is formed by a glass coverslip that was coated with a 2 mg ml–1 solution of the lectin concanavalin A (Sigma; L7647), and then washed with water and dried for 30 min under vacuum, as described previously41. A suspension of cells was then placed on the glass coverslip and incubated for 5 min at the appropriate temperature for the experiment, to allow cells to attach. The coverslip was then washed with pre-warmed minimal medium and the cham-ber was assembled in the presence of 0.4 ml pre-warmed minimal medium with appropriate supplements. The cells were thus separated by a thin layer of liquid from the glass top of the BIOPTECHS FCS2 chamber, which was heated directly by metal electrodes and monitored to ensure that it remained at the desired tem-perature for the experiment. The objective lens was also heated to the appropriate temperature with a BIOPTECHS Objective Heater Controller (Photometrics), and the surrounding air temperature was controlled with a weather station temperature controller (Precision Control). For all experiments, except those involving GFP–Ras2, eight z-sections (with a spacing of 0.4 µm) were analysed. For GFP–Ras2, 16 z-sections (with a spacing of 0.2 µm) were analysed.
The photobleaching experiment in Fig. 4b was performed using the Deltavision QLM 100 mW 488 nm laser system, which was adjusted so that the diameter of the laser beam was reduced to its smallest aperture (approximately 800 nm). For other experiments, cells were viewed with a Zeiss Axiovert 200M microscope and a Cool Snap HQ camera (Photometrics). The objective lens was a Zeiss alpha plan ×100 1.45 NA with a Chroma ET GFP filter set (49002). The illumination source was the Sutter 300w xenon system with liquid light guide, and Metamorph acqui-sition software (Molecular Devices) was used. Stacks of z-sections were analysed as described above, except that for GFP–Ras2 30 sections of 0.2 µm were used to facilitate the examination of whole cells, as bleaching was less of a concern than in time-lapse experiments with multiple timepoints.
To visualize the actin cytoskeleton in fixed cells, the samples were washed twice with PBS, and then incubated them in 0.05 U µl–1 rhodamine–phalloidin (Cambridge Biosciences) for 90 min before further washing and viewing. The microscopy data were deconvolved using Huygens (SVI) according to the quick maximum likelihood-estimation method and a measured point spread function. The deconvolved data set was viewed with Imaris software (Bitplane). At least 100 cells were examined for each sample unless specified otherwise in the text. Further details of equipment and settings for fluorescence microscopy can be found in the Supplementary Information, Methods.
To perform transmission electron microscopy, cell pellets were frozen in a Balt-Tec HPM010 high-pressure freezer (Bal-Tec). Samples were then substituted with 2% osmium tetroxide and 0.1% uranyl acetate, before infiltration and embedding
with Spurr’s resin42. Serial sections (60 nm) were cut on a Leica UCS ultramicrotome (Leica Microsystems), stained with Reynolds’ lead citrate43, and then observed with a JEOL 1220 TEM (JEOL) operated at 80 KV. Individual cells were tracked through each section in the series and images were acquired with a SiS Megaview II CCD camera and ANALYSiS software v3.2. (Olympus Soft Imaging Systems).
Immunoprecipitation of proteins. Cell extracts were prepared by grinding 2.5 g of cell ‘popcorn’ with dry ice in a coffee mill, as described previously44. The lysis buffer contained 100 mM HEPES–KOH at pH 7.9, 300 mM potassium acetate, 10 mM magnesium acetate, 10% glycerol, 0.1% NP-40, 2 mM EDTA, 2 mM β-glycerophosphate, 2 mM sodium fluoride, 1 mM DTT, 1× protease inhibitor cocktail (Roche 11575400), and 1% protease inhibitors (Sigma P 8215). To isolate tagged proteins, 1.8 × 107 of dynabeads M-270 Epoxy (Dynal; 143.01) coupled to rabbit anti-sheep IgG (Sigma S-1265) were used. The beads were washed with an appropriate buffer (similar to the lysis buffer but with 150 mM potassium acetate and lacking glycerol) and then the bound proteins were released by heating the beads in presence of 1× Laemmli buffer.
Detection of proteins. The c-Myc and HA epitopes were detected with the mouse monoclonal antibodies 9E10 and 12CA5 at a final concentration of 5 µg ml–1. The TAP tag was detected using rabbit peroxidase–anti-peroxidase complex (Sigma; P-2026, 1:5000). For the experiment shown in Fig. 1d, each gel lane was cut into 60 × 1.0 mm bands with a razor blade, and transferred each band to a 96-well plate (PRO10003; Genomic Solutions). Gel bands were processed robotically using a ProGest instrument (Genomic Solutions), analysed using nano LC-MSMS on a LCQ Deca XP Plus ion trap mass spectrometer (Thermo), and the data were processed as described previously44.
Flow cytometry. DNA content was measured by flow cytometry, as described previously11.
Note: Supplementary Information is available on the Nature Cell Biology website.
ACKNOWLEDGEMENTSThis work was funded by Cancer Research U.K., from whom K.L. received a Senior Cancer Research Fellowship, and by the EMBO Young Investigator Programme. A.S.D. received a Marie Curie personal training fellowship from the European Union. We are very grateful to S. Bagley for much advice and help with time-lapse video microscopy, B. Glick for advice regarding the use of concanavalin A, J. Wiedenmann for the eQFP construct, and E. Schiebel for plasmids and helpful comments on the manuscript. A.S.D. thanks A. Sanchez-Sanfructuoso and J. Diaz-Marcos for their support.
AUTHOR CONTRIBUTIONSV. M. generated the strains that are described in Supplementary Information, Fig. S1a and b, and screened these strains for defects in cell cycle progression to generate the corrsponding data in Supplementary Information, Fig. 1c. S. M. performed the electron microscopy that is summarized in Fig. 4a. R. J. was responsible for the mass spectrometry data in Fig. 1e. G. P. made the initial observations of the localization of Inn–GFP and taught A. S. D. how to prepare samples for fluorescence microscopy. K. L. made the strains shown in Fig. 8a, b, c. A. S. D. performed all other experiments.
Published online at http://www.nature.com/naturecellbiology/ Reprints and permissions information is available online at http://npg.nature.com/reprintsandpermissions
1. Albertson, R., Riggs, B. & Sullivan, W. Membrane traffic: a driving force in cytokinesis. Trends Cell Biol. 15, 92–101 (2005).
2. Strickland, L. I. & Burgess, D. R. Pathways for membrane trafficking during cytokinesis. Trends Cell Biol. 14, 115–118 (2004).
3. Burgess, D. R. & Chang, F. Site selection for the cleavage furrow at cytokinesis. Trends Cell Biol. 15, 156–162 (2005).
4. D’Avino, P. P., Savoian, M. S. & Glover, D. M. Cleavage furrow formation and ingres-sion during animal cytokinesis: a microtubule legacy. J. Cell Sci. 118, 1549–1558 (2005).
5. Tolliday, N., Bouquin, N. & Li, R. Assembly and regulation of the cytokinetic apparatus in budding yeast. Curr. Opin. Microbiol. 4, 690–695 (2001).
6. Balasubramanian, M. K., Bi, E. & Glotzer, M. Comparative analysis of cytokinesis in budding yeast, fission yeast and animal cells. Curr. Biol. 14, R806–R818 (2004).
7. Glotzer, M. The molecular requirements for cytokinesis. Science 307, 1735–1739 (2005).
8. Barr, F. A. & Gruneberg, U. Cytokinesis: placing and making the final cut. Cell 131, 847–860 (2007).
nature cell biology volume 10 | number 4 | APrIl 2008 405

© 2008 Nature Publishing Group
A RT I C L E S
9. Giaever, G. et al. Functional profiling of the Saccharomyces cerevisiae genome. Nature 418, 387–391 (2002).
10. Winzeler, E. A. et al. Functional characterization of the S. cerevisiae genome by gene deletion and parallel analysis. Science 285, 901–906 (1999).
11. Kanemaki, M., Sanchez-Diaz, A., Gambus, A. & Labib, K. Functional proteomic identifi-cation of DNA replication proteins by induced proteolysis in vivo. Nature 423, 720–725 (2003).
12. Dohmen, R. J., Wu, P. & Varshavsky, A. Heat-inducible degron: a method for construct-ing temperature-sensitive mutants. Science 263, 1273–1276 (1994).
13. Labib, K., Tercero, J. A. & Diffley, J. F. X. Uninterrupted MCM2-7 function required for DNA replication fork progression. Science 288, 1643–1647 (2000).
14. Cheeseman, I. M. et al. Implication of a novel multiprotein Dam1p complex in outer kinetochore function. J. Cell Biol. 155, 1137–1145 (2001).
15. Enquist-Newman, M. et al. Dad1p, third component of the Duo1p/Dam1p complex involved in kinetochore function and mitotic spindle integrity. Mol. Biol. Cell 12, 2601–2613 (2001).
16. Euskirchen, G. M. Nnf1p, Dsn1p, Mtw1p, and Nsl1p: a new group of proteins important for chromosome segregation in Saccharomyces cerevisiae. Eukaryot. Cell 1, 229–240 (2002).
17. Janke, C., Ortiz, J., Tanaka, T. U., Lechner, J. & Schiebel, E. Four new subunits of the Dam1–Duo1 complex reveal novel functions in sister kinetochore biorientation. EMBO J. 21, 181–193 (2002).
18. Li, Y. et al. The mitotic spindle is required for loading of the DASH complex onto the kinetochore. Genes Dev. 16, 183–197 (2002).
19. Pinsky, B. A., Tatsutani, S. Y., Collins, K. A. & Biggins, S. An Mtw1 complex promotes kinetochore biorientation that is monitored by the Ipl1/Aurora protein kinase. Dev. Cell 5, 735–745 (2003).
20. Scharfenberger, M. et al. Nsl1p is essential for the establishment of bipolarity and the localization of the Dam–Duo complex. EMBO J. 22, 6584–6597 (2003).
21. Lippincott, J. & Li, R. Dual function of Cyk2, a cdc15/PSTPIP family protein, in regulat-ing actomyosin ring dynamics and septin distribution. J. Cell Biol. 143, 1947–1960 (1998).
22. Vallen, E. A., Caviston, J. & Bi, E. Roles of Hof1p, Bni1p, Bnr1p, and myo1p in cyto-kinesis in Saccharomyces cerevisiae. Mol. Biol. Cell 11, 593–611 (2000).
23. Dobbelaere, J. & Barral, Y. Spatial coordination of cytokinetic events by compartmen-talization of the cell cortex. Science 305, 393–396 (2004).
24. Norden, C. et al. The NoCut pathway links completion of cytokinesis to spindle midzone function to prevent chromosome breakage. Cell 125, 85–98 (2006).
25. Whistler, J. L. & Rine, J. Ras2 and Ras1 protein phosphorylation in Saccharomyces cerevisiae. J. Biol. Chem. 272, 18790–18800 (1997).
26. Cho, W. & Stahelin, R. V. Membrane binding and subcellular targeting of C2 domains. Biochim. Biophys. Acta 1761, 838–849 (2006).
27. Cho, W. & Stahelin, R. V. Membrane-protein interactions in cell signaling and membrane trafficking. Annu. Rev. Biophys. Biomol. Struct. 34, 119–151 (2005).
28. Hurley, J. H. Membrane binding domains. Biochim. Biophys. Acta 1761, 805–811 (2006).
29. VerPlank, L. & Li, R. Cell cycle-regulated trafficking of Chs2 controls actomyosin ring stability during cytokinesis. Mol. Biol. Cell 16, 2529–2543 (2005).
30. Zhang, G., Kashimshetty, R., Ng, K. E., Tan, H. B. & Yeong, F. M. Exit from mitosis triggers Chs2p transport from the endoplasmic reticulum to mother-daughter neck via the secretory pathway in budding yeast. J. Cell Biol. 174, 207–220 (2006).
31. Roh, D. H., Bowers, B., Schmidt, M. & Cabib, E. The septation apparatus, an autono-mous system in budding yeast. Mol. Biol. Cell 13, 2747–2759 (2002).
32. Chuang, J. S. & Schekman, R. W. Differential trafficking and timed localization of two chitin synthase proteins, Chs2p and Chs3p. J. Cell Biol. 135, 597–610 (1996).
33. Martens, S., Kozlov, M. M. & McMahon, H. T. How synaptotagmin promotes membrane fusion. Science 316, 1205–1208 (2007).
34. Aspenstrom, P., Fransson, A. & Richnau, N. Pombe Cdc15 homology proteins: regula-tors of membrane dynamics and the actin cytoskeleton. Trends Biochem. Sci. 31, 670–679 (2006).
35. Field, C. M. & Alberts, B. M. Anillin, a contractile ring protein that cycles from the nucleus to the cell cortex. J. Cell Biol. 131, 165–178 (1995).
36. Oegema, K., Savoian, M. S., Mitchison, T. J. & Field, C. M. Functional analysis of a human homologue of the Drosophila actin binding protein anillin suggests a role in cytokinesis. J. Cell Biol. 150, 539–552 (2000).
37. Echard, A., Hickson, G. R., Foley, E. & O’Farrell, P. H. Terminal cytokinesis events uncovered after an RNAi screen. Curr. Biol. 14, 1685–1693 (2004).
38. Somma, M. P., Fasulo, B., Cenci, G., Cundari, E. & Gatti, M. Molecular dissection of cytokinesis by RNA interference in Drosophila cultured cells. Mol. Biol. Cell 13, 2448–2460 (2002).
39. Straight, A. F., Field, C. M. & Mitchison, T. J. Anillin binds nonmuscle myosin II and regulates the contractile ring. Mol. Biol. Cell 16, 193–201 (2005).
40. Zhao, W. M. & Fang, G. Anillin is a substrate of anaphase-promoting complex/cyclosome (APC/C) that controls spatial contractility of myosin during late cytokinesis. J. Biol. Chem. 280, 33516–33524 (2005).
41. Losev, E. et al. Golgi maturation visualized in living yeast. Nature 441, 1002–1006 (2006).
42. Spurr, A. R. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 26, 31–43 (1969).
43. Reynolds, E. S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 17, 208–212 (1963).
44. Gambus, A. et al. GINS maintains association of Cdc45 with MCM in replisome pro-gression complexes at eukaryotic DNA replication forks. Nature Cell Biol. 8, 358–366 (2006).
45. Rost, B., Yachdav, G. & Liu, J. The PredictProtein server. Nucleic Acids Res 32, W321–W326 (2004).
46. Rizo, J. & Sudhof, T. C. C2-domains, structure and function of a universal Ca2+-binding domain. J. Biol. Chem. 273, 15879–15882 (1998).
406 nature cell biology volume 10 | number 4 | APrIl 2008
© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 1
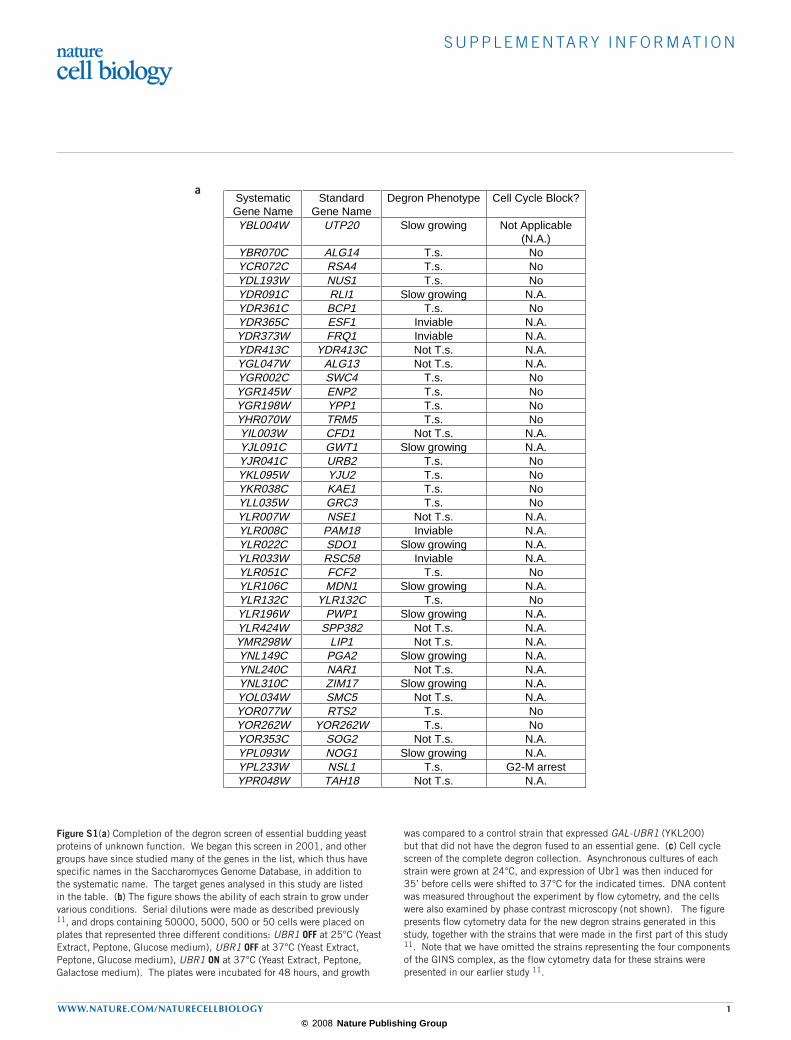
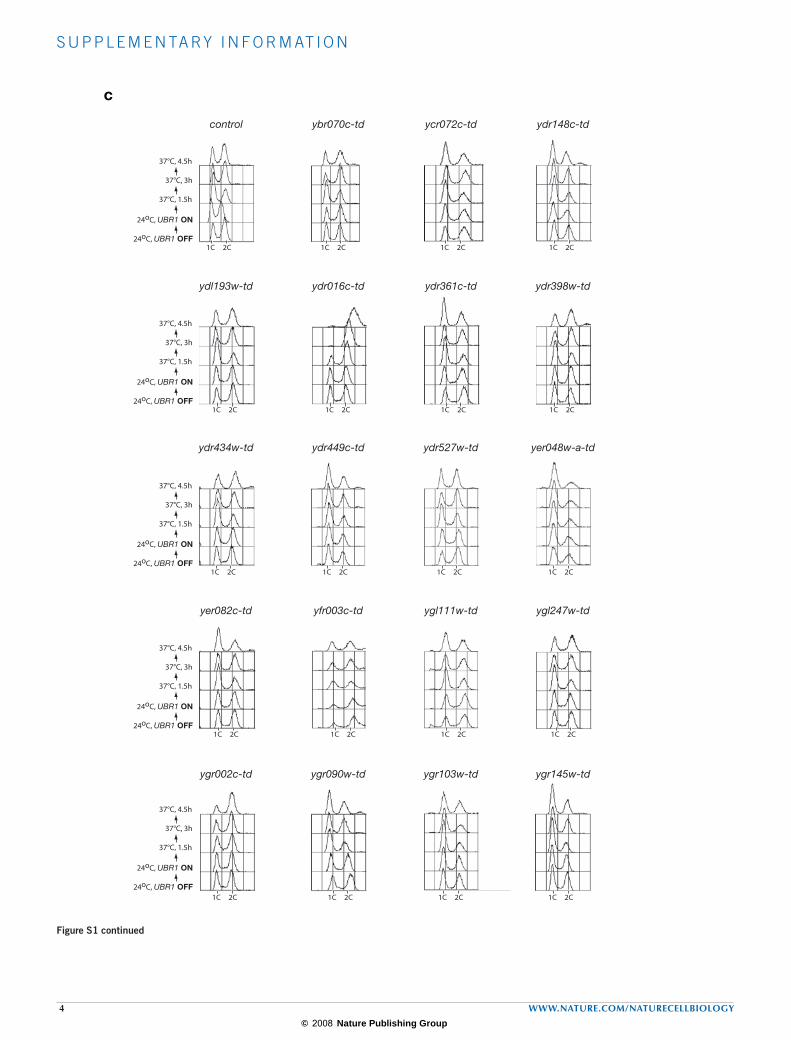
Figure S1(a) Completion of the degron screen of essential budding yeast proteins of unknown function. We began this screen in 2001, and other groups have since studied many of the genes in the list, which thus have specific names in the Saccharomyces Genome Database, in addition to the systematic name. The target genes analysed in this study are listed in the table. (b) The figure shows the ability of each strain to grow under various conditions. Serial dilutions were made as described previously 11, and drops containing 50000, 5000, 500 or 50 cells were placed on plates that represented three different conditions: UBR1 OFF at 25°C (Yeast Extract, Peptone, Glucose medium), UBR1 OFF at 37°C (Yeast Extract, Peptone, Glucose medium), UBR1 ON at 37°C (Yeast Extract, Peptone, Galactose medium). The plates were incubated for 48 hours, and growth
was compared to a control strain that expressed GAL-UBR1 (YKL200) but that did not have the degron fused to an essential gene. (c) Cell cycle screen of the complete degron collection. Asynchronous cultures of each strain were grown at 24°C, and expression of Ubr1 was then induced for 35’ before cells were shifted to 37°C for the indicated times. DNA content was measured throughout the experiment by flow cytometry, and the cells were also examined by phase contrast microscopy (not shown). The figure presents flow cytometry data for the new degron strains generated in this study, together with the strains that were made in the first part of this study 11. Note that we have omitted the strains representing the four components of the GINS complex, as the flow cytometry data for these strains were presented in our earlier study 11.
SystematicGene Name
StandardGene Name
Degron Phenotype Cell Cycle Block?
YBL004W UTP20 Slow growing Not Applicable(N.A.)
YBR070C ALG14 T.s. NoYCR072C RSA4 T.s. NoYDL193W NUS1 T.s. NoYDR091C RLI1 Slow growing N.A.YDR361C BCP1 T.s. NoYDR365C ESF1 Inviable N.A.YDR373W FRQ1 Inviable N.A.YDR413C YDR413C Not T.s. N.A.YGL047W ALG13 Not T.s. N.A.YGR002C SWC4 T.s. NoYGR145W ENP2 T.s. NoYGR198W YPP1 T.s. NoYHR070W TRM5 T.s. NoYIL003W CFD1 Not T.s. N.A.YJL091C GWT1 Slow growing N.A.YJR041C URB2 T.s. NoYKL095W YJU2 T.s. NoYKR038C KAE1 T.s. NoYLL035W GRC3 T.s. NoYLR007W NSE1 Not T.s. N.A.YLR008C PAM18 Inviable N.A.YLR022C SDO1 Slow growing N.A.YLR033W RSC58 Inviable N.A.YLR051C FCF2 T.s. NoYLR106C MDN1 Slow growing N.A.YLR132C YLR132C T.s. NoYLR196W PWP1 Slow growing N.A.YLR424W SPP382 Not T.s. N.A.YMR298W LIP1 Not T.s. N.A.YNL149C PGA2 Slow growing N.A.YNL240C NAR1 Not T.s. N.A.YNL310C ZIM17 Slow growing N.A.YOL034W SMC5 Not T.s. N.A.YOR077W RTS2 T.s. NoYOR262W YOR262W T.s. NoYOR353C SOG2 Not T.s. N.A.YPL093W NOG1 Slow growing N.A.YPL233W NSL1 T.s. G2-M arrestYPR048W TAH18 Not T.s. N.A.
a

© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
2 WWW.NATURE.COM/NATURECELLBIOLOGY
control ybr070cw-td ycr072c-td
ydr361c-td ydr413c-td
yll035w-td ylr007w-td
ydl193w-td
ygl247w-td ygr002c-td
ygr198w-td
ygr145w-td
yhr070w-td
ykl095w-tdyjr041c-td
yil003w-td
UBR1 OFF, 25oC
UBR1 OFF, 37oC
UBR1 ON, 37oC
ykr038c-td
ylr051c-td
UBR1 OFF, 25oC
UBR1 OFF, 37oC
UBR1 ON, 37oC
UBR1 OFF, 25oC
UBR1 OFF, 37oC
UBR1 ON, 37oC
UBR1 OFF, 25oC
UBR1 OFF, 37oC
UBR1 ON, 37oC
UBR1 OFF, 25oC
UBR1 OFF, 37oC
UBR1 ON, 37oC
UBR1 OFF, 25oC
UBR1 OFF, 37oC
UBR1 ON, 37oC
b
Figure S1 continued

© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 3
yor353c-td
ypr048c-td
ylr132c-td ylr424w-td
yol034w-tdynl240c-td
yor262w-td
ymr298w-td
yor077w-td
ypl233w-td
UBR1 OFF, 25oC
UBR1 OFF, 37oC
UBR1 ON, 37oC
UBR1 OFF, 25oC
UBR1 OFF, 37oC
UBR1 ON, 37oC
UBR1 OFF, 25oC
UBR1 OFF, 37oC
UBR1 ON, 37oC
UBR1 OFF, 25oC
UBR1 OFF, 37oC
UBR1 ON, 37oC
Figure S1 continued
© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
4 WWW.NATURE.COM/NATURECELLBIOLOGY
control
ydr434w-td
37oC, 1.5 h
37oC, 3 h
37oC, 4.5 h
24oC, UBR1 ON
ydr449c-td ydr527w-td yer048w-a-td
yer082c-td yfr003c-td
ygr002c-td
ydl193w-td
ybr070c-td
ydr016c-td ydr361c-td ydr398w-td
ygl111w-td ygl247w-td
ygr090w-td ygr103w-td ygr145w-td
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C1C 2C
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
1C 2C
ycr072c-td ydr148c-td
1C 2C
c
Figure S1 continued

© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 5
yhr036w-td
ygr156w-td ygr198w-td ygr211w-td
yhr070w-td
yjl010c-td yjl069c-td
yil083c-td yil091c-td
yhr088w-td yhr122w-td
yjr041c-td
ygr280c-td
yhr083w-td yhr085w-td
yhr196w-td yhr197w-td
yil104c-td yir010w-td
yjr072c-td
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
Figure S1 continued

© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
6 WWW.NATURE.COM/NATURECELLBIOLOGY
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
ykl052c-td ykl082c-td
yjr136c-td ykl018w-tdyjr141w-td
ykl095w-td
ykr038c-td yll035w-td ylr002c-td ylr051c-td
ylr132c-td ylr186w-td ylr215c-td ylr243w-td
ylr409c-td ylr440c-td ylr459w-td yml093w-td
ykl033w-td
ykl099c-td
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
Figure S1 continued

© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 7
ymr093w-td ynl038w-td
ynl132w-td
ynl245c-td ynl258c-td
ynl110c-td
ynl263c-td
yor145c-td
yor077w-td
ypl146c-td
yor262w-td
ypl233w-td
ynl152w-td ynl181w-td ynl182c-td
ynl124w-td
yor287c-td ypl012w-td
ypr143w-td ypr144c-td
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 2C1C 2C
1C 2C 1C 2C1C 21C 2C
1C 2C 1C 2C1C 2C1C 2C
4 8C24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
24oC, UBR1 OFF
24oC, UBR1 ON
37°C, 1.5h
37°C, 3h
37°C, 4.5h
Figure S1 continued

© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
8 WWW.NATURE.COM/NATURECELLBIOLOGY
Figure S2 The INN1, IQG1 and MYO1 genes are all essential in the budding yeast strain background W303 and are required for cell division. (a) Tetrad analysis of germinating spores from diploid cells lacking one copy of the INN1 (YASD788), MYO1 (YASD782) or IQG1 (YASD836) genes. The plates were incubated at 24°C for 3 days (INN1 and MYO1) or 4 days (IQG1). (b) Phase contrast images illustrating the phenotype of germinated spores with the indicated genotype, after growth for 24 hours.
Figure S3 Rapid inactivation of Myo1 blocks the localisation of Iqg1. A myo1-td GFP-IQG1 strain (YASD690) was grown as in Figure 2, and DNA content (a), the proportion of binucleate cells (b) and the proportion of cells with medial rings or spots of GFP-Iqg1 (c) were determined throughout the experiment.
ainn1
inn1
myo1
myo1
iqg1
iqg1b
0
20
40
60
80
100
120'105'90'75'60'
Binucleate cells
% c
ells
myo1-td GFP-IQG1
Asyn.24°C
G1 24°C
30' release
45'
60'
75'
90'
105'
120'
G1 37°C
1C 2C 4C
a
b
Time after releasefrom G1 arrest at 37°C / minutes
c
0
10
20
30
40
50
120'105'90'75'60'
Iqg1 Rings/Spots
Time after releasefrom G1 arrest at 37°C / minutes
% c
ells

© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 9
Figure S4 Both the C2 domain and the remainder of the Inn1 protein are required for viability and cell division. (a) Tetrad analysis of diploid INN1/inn1-ΔC2 cells (YASD1211 in which one copy of the INN1 gene expresses a protein lacking amino acids 1-134) and INN1/inn1-ΔCT
cells (YASD1214 in which one copy of the INN1 gene expresses a protein lacking amino acids 135-409), after three days growth at 24°C. (b) Phase contrast images of cells from the same experiment after growth for 24 hours.
aINN1 / inn1- C2 INN1 / inn1- CT
inn1- C2 inn1- CTControl
b
© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
10 WWW.NATURE.COM/NATURECELLBIOLOGY
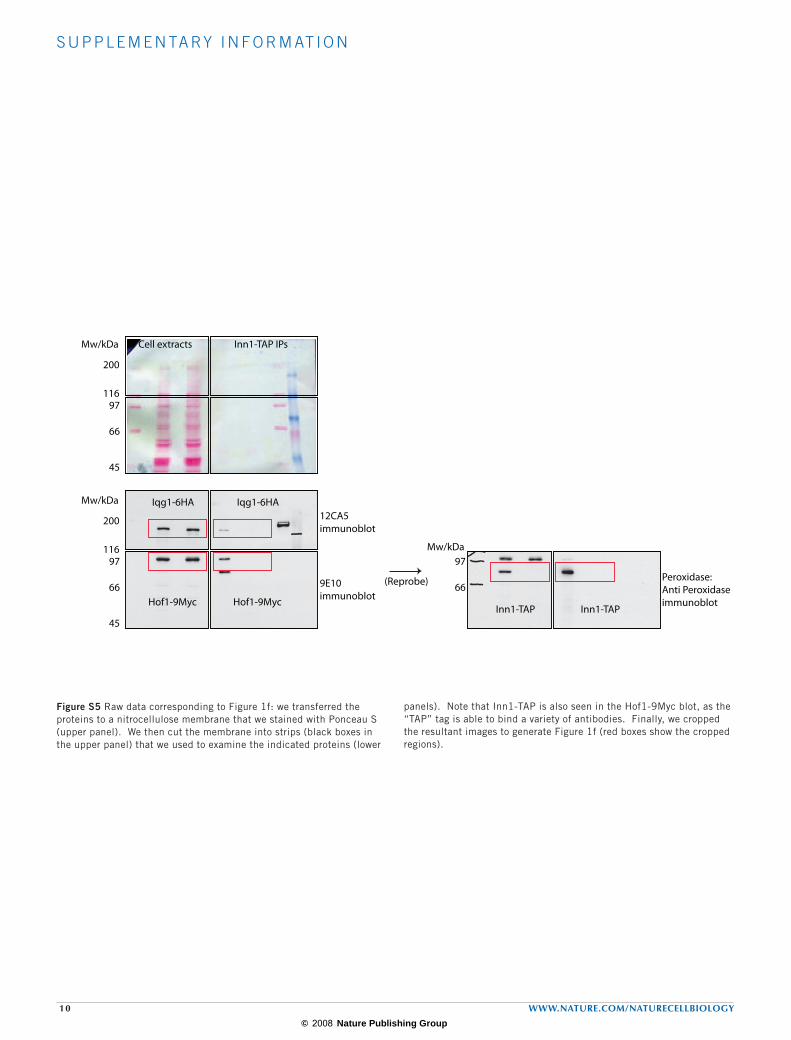
Figure S5 Raw data corresponding to Figure 1f: we transferred the proteins to a nitrocellulose membrane that we stained with Ponceau S (upper panel). We then cut the membrane into strips (black boxes in the upper panel) that we used to examine the indicated proteins (lower
panels). Note that Inn1-TAP is also seen in the Hof1-9Myc blot, as the “TAP” tag is able to bind a variety of antibodies. Finally, we cropped the resultant images to generate Figure 1f (red boxes show the cropped regions).
Mw/kDa
116
200
97
66
45
Cell extracts Inn1-TAP IPs
Iqg1-6HA
Hof1-9Myc
Iqg1-6HA
Hof1-9Myc
12CA5immunoblot
9E10immunoblot
Peroxidase:Anti Peroxidaseimmunoblot
Inn1-TAP Inn1-TAP
Mw/kDa
116
200
97
66
45
97
66
Mw/kDa
(Reprobe)

© 2008 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 11
Movie S1 The localisation of Inn1 is coincident with the actomyosin ring. Cells expressing both Inn1-GFP and Myo1-eQFP (YASD705) were grown at 30°C and treated as described in the legend to Figure 1d. Inn1-GFP is recruited to the actomyosin ring at the end of mitosis and co-localises with Myo1-eQFP, so that the red signal changes to yellow just before the ring contracts.
Movie S2 Inn1 is not required for contraction of the actomyosin ring at the end of mitosis. Control (YASD1155) and inn1-td (YASD1144) cells expressing both Spc42-eQFP and Myo1-Tomato were filmed at the end of mitosis as described in the legend to Figure 3c. The movies show six control cells (left-hand side) and six inn1-td cells (right-hand side), and correspond to a period of 12 minutes at the end of mitosis, during which time the actomyosin ring contracted in each cell.
Supplementary movie legends




















