
Percolative conductivity in alkaline-earth silicate melts and glasses
Upload
independentCategory
view
3download
0
EXPERIMENTAL NEUROLOGY 54,128-140 (1977)
Increased Alkaline Phosphatase Specific Activity in
Reeler Cerebella 1
BARRY D. SHUR, G. ROBERT DELONG, AND STEPHEN ROTH 2
Department of Biology, The Johns Hopkins Utiversity, Baltimore, Maryland 21218, and Department of Pediatric Neurology, Massachusetts General Hospital,
Boston, Massachusetts 02114
Received June 21,1976; revision received August 23,1976
The neurological mutant, reeler, is characterized by faulty cellular asso- ciations in various brain regions in mice. A cell surface enzyme, alkaline phosphatase, was examined in an attempt to understand the nature of this morphogenetic lesion. The specific activity of alkaline phosphatase in 13-day postnatal reeler cerebella is three times higher than in phenotypically normal littermates. Cerebella taken from 4-day postnatal reelers show enzyme activ- ity more similar to normal controls. Alkaline phosphatase specific activity decreases with cerebellar development much more in unaffected animals than in reelers, which maintain a higher level during the same development period. Phosphatase activities are indistinguishable in normal and reeler cerebral isocortices and brain stems at all developmental stages tested. Enzyme assays are linear throughout the range of protein concentrations and the times of incubation used. No divalent cation stimulation can be demonstrated. K, values toward p-nitrophenylphosphate as a substrate have been deter- mined and are not significantly different for the normal and reeler cerebellar enzymes. Histochemical localization of alkaline phosphatase suggests that this enzyme is concentrated in one particular area of the cerebellum. The relationship between this specific localization and the increased activity in reeler cerebella is discussed.
INTRODUCTION
The mechanisms which govern cell recognition during development have
yet to be firmly established. Complex carbohydrates and glycosyltrans-
1 This work was supported by a research grant from the National Institute of Child Health and Human Development to S. R. This i’s Contribution No. 886 from the McCollum-Pratt Institute.
* Drs. Shur and Roth are at the Johns Hopkins University ; and Dr. DeLong is at Massachusetts General Hospital.
1977 by Academic Press form'

INCREASED PROSPHATASE ACTIVITY IN REELER 129
ferases, both on the cell surface and within the extracellular matrix, have been implicated in this process [for review, see (7) 1. The murine neuro- logical mutant, reeler, shows a morphogenetic defect in which the role of glycosyltransferases was being tested. During the course of the investiga- tion it was noted that when reeler brain homogenates were incubated with uridine diphosphate N-acetylgalactosamine, free N-acetylgalactosamine was formed at a higher rate than in homogenates of incubated normal mouse brain. The rate of formation of N-acetylgalactosamine-PO4 was high but equal in both of these preparations. An analysis of the higher phosphatase activity in this mutant was initiated, and the data are reported here.
The reeler mutant, originally isolated by Falconer (2)) is characterized by faulty cell alignments in regions of the brain where histotypic laminations normally occur (4, 8, 9). Cerebral isocortex, hippocampus, and cerebellum all show disturbed cellular associations. The cerebellum, which is affected most, is greatly reduced in size due to the lack of granule cell proliferation and subsequent migration into the cerebellar cortex (9). Purkinje cells do not align themselves into a uniform layer, and neurons of the deep cerebellar nuclei exist scattered throughout the white matter (4). The few existing granule cells remain at the surface of the cerebellum. Unlike the cerebellum, other affected areas of the reeler brain are similar to normal in size and shape although the arrangement of their cells is abnormal (1, 9). In vitro studies (1) by DeLong and Sidman have shown that cells dis- sociated from reeler brain areas fail to sort out into histotypic patterns characteristic of normal aggregates. Cells from normal cerebral isocortex produce aggregates with three well-defined layers when removed from 17.5 to l&S-gestation-day embryos. Reeler isocortical cells of similar age produce aggregates with random cell orientations. Similarly, normal cere- bellar cells from 3- to S-day-old pups reaggregate into patterns analogous to in V&O cell associations. Reeler cerebellar cells from these ages yield aggregates with poorly defined cellular layers and random orientations. These studies (1) suggest that reeler cells may possess an abnormal surface component capable of disturbing morphogenesis.
The data presented here show that at least one cell surface component, alkaline phosphatase, is distributed abnormally in reeler cerebella as com- pared to normal cerebella.
EXPERIMENTAL PROCEDURE
Mice. C57B1/6J mice heterozygous at the reeler (r-1) locus were obtained from the Jackson Laboratories and maintained through heterozygote matings. Cerebella, cerebral isocortices, and brain stems from pheno- typically normal (+/+, rl/+) and reeler (rl/rl) littermates were re- moved and frozen. Phenotypes were determined in lZday, or older, animals

130 SHUR, DELONG AND ROTH
by noting the presence of disturbed muscular coordination in reelers as well as by microscopic confirmation. Younger animals were diagnosed histologically.
Honzogenization. Brains were homogenized (tissue grinder with a Teflon pestle, size AA, A. H. Thomas Co., Philadelphia, Pennsylvania 19105) with 15 strokes in 20 volumes of the following homogenization medium: 0.15 M NaCI, 1% Triton X-100, 0.03 M Hepes [4-(2-hydroxyethyl)-l- piperazine ethanesulfonate] buffer (Sigma Chemical Co., St. Louis, Missouri 63178), pH 7.2. When the effect of the detergent concentration was assayed, cerebella were homogenized in Triton-free medium.
Assays. Fifty microliters homogenate were removed and assayed for determination of protein concentration (5) using bovine serum albumin as a standard.
In a total of 235 ~1, alkaline phosphatase reaction mixtures contained the following components: 200 ~1 0.1 M glycine-NaOH buffer, pH 10.6; 20 ~1 71.4 mM p-nitrophenylphosphate (Sigma 104 substrate, 5.0 mg tablet per 0.25 ml H,O) to give a final concentration of 6.1 mM ; and 15 ~1 homogenate diluted threefold with homogenization medium. These condi- tions were shown to be optimal by studies described in Results. Incubations
PH
FIG. 1. The pH-dependent phosphatase activity in normal ( l - l ) and reeler
co- 0) cerebellar homogenates. Reaction components in this and subsequent figures are as in Materials and Methods except that for this assay the following buffers were used: 0.1 M citrate, pH 2.9, 4.0, 4.8, 5.6, and 6.0; 0.1 M Tris-maleate, pH 6.0, 6.9, 8.0, and 8.6; 0.1 M glycine-NaOH, pH 8.6, 8.9, 9.7, 10.5, and 11.6. Assay tubes contained 67.0 (normal) and 44.0 (reeler) pg protein.
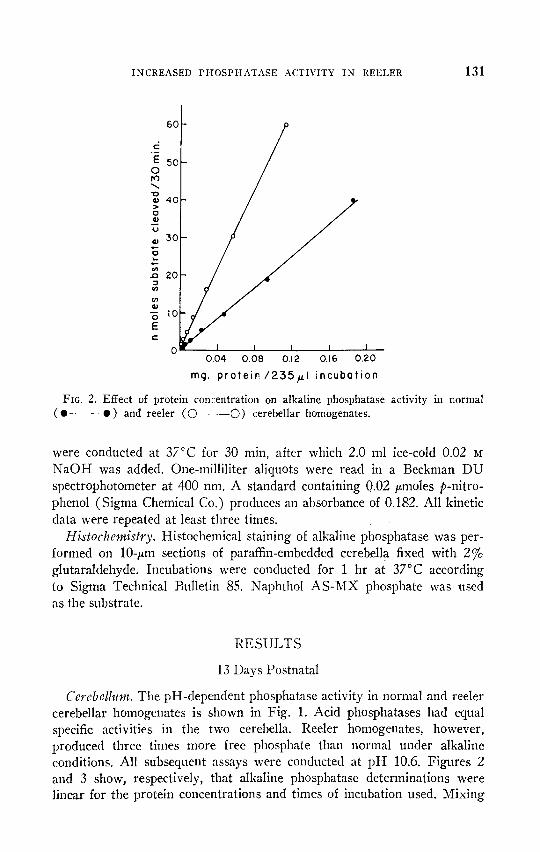
INCREASED PJIOSPIIAT.4SE ACTIVITY IN REELER 131
mg. protein /235Pl incubation
FIG. 2. Effect of protein concentration on alkaline phosphatase activity in normal (0 -0) and reeler (0 -0) cerebellar homogenates.
were conducted at 37°C for 30 min, after which 2.0 ml ice-cold 0.02 M
NaOH was added. One-milliliter aliquots were read in a Beckman DU spectrophotometer at 400 nm. A standard containing 0.02 pmoles p-nitro- phenol (Sigma Chemical Co.) produces an absorbance of 0.182. All kinetic data were repeated at least three times.
Histoclze&sfr~~. Histochemical staining of alkaline phosphatase was per- formed on lo-pm sections of paraffin-embedded cerebella fixed with 2% glutaraldehyde. Incubations were conducted for 1 hr at 37°C according to Sigma Technical Bulletin 85. Naphthol AS-MX phosphate was used as the substrate.
RESULTS
13 Days Postnatal
Cerebsllztwa. The pH-dependent phosphatase activity in normal and reeler cerebellar homogenates is shown in Fig. 1. Acid phosphatases had equal specific activities in the two cerebella. Reeler homogenates, however, produced three times more free phosphate than normal under alkaline conditions. All subsequent assays were conducted at pII 10.6. Figures 2 and 3 show, respectively, that alkaline phosphatase determinations were linear for the protein concentrations and times of incubation used. Mixing

132 SFITJR, DELONG AND ROTH
0 15 30 45 60
minutes
FIG. 3. Time course of normal ( l -0) and reeler (0 -0) cerebellar homogenate alkaline phosphatase activity. 0.05 ml aliquots of normal (146 pg protein) and reeler (130 pg protein) homogenate were mixed and incubated (X -x>.
normal and reeler aliquots of equal protein concentration yielded the expected intermediate value (Fig. 3). These data are inconsistent with the presence of any reeler phosphatase stimulator or normal phosphatase
S
FIG. 4. S (millimolar concentration)/V (micromoles of substrate cleaved per milli- gram of protein per 30 min) vs S (millimolar) plot of normal reeler (0
(0 -0)
-0) and cerebellar homogenate alkaline phosphatase activity. The amounts
of protein per assay were 26.6 (normal) and 39.0 (reeler) pg. K, values were dcter- mined as 1.3 (normal) and 1.1 m&z (reeler) for $-nitrophenylphosphate as the sub- strate.
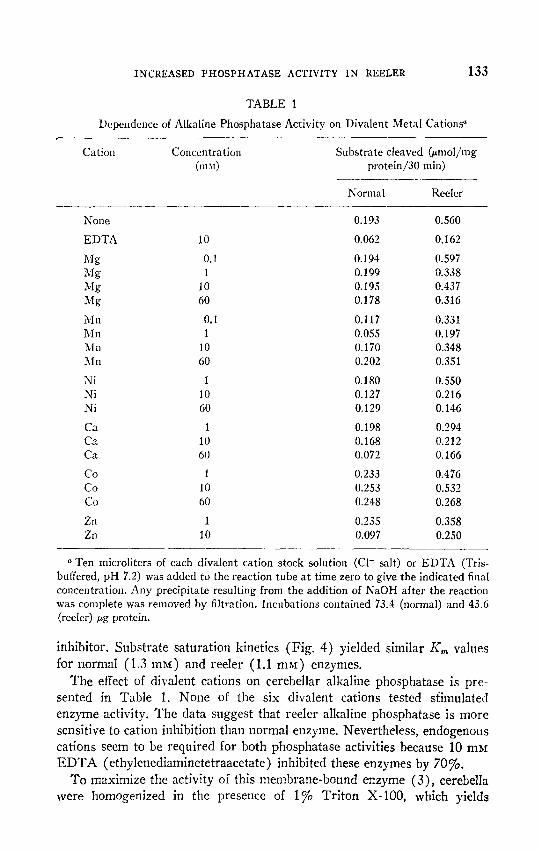
INCREASED PHOSPHATASE ACTIVITY IN REELER 133
TABLE 1
Dependence of Alkaline Phosphatase Activity on Divalent Metal Cations”
Cation Concentration (ml)
None
EDTA
mx Mg w? b Mn RI 11 Ml1 MI1
Ni Ni Ni
Ca Ca Ca
co co co
Zn Zn
10
0.1 1
10 60
0.1 1
10 60
1 10 60
1 10 60
1 10 60
1 10
Substrate cleaved &mol/mg protein/30 min)
Normal Reeler
0.193 0.560
0.062 0.162
0.194 0.597 0.199 0.338 0.195 0.437 0.178 0.316
0.117 0.331 0.055 0.197 0.170 0.348 0.202 0.351
0.180 0.550 0.127 0.216 0.129 0.146
0.198 0.294 0.168 0.212 0.072 0.166
0.233 0.476 0.253 0.532 0.248 0.268
0.235 0.358 0.097 0.250
- a Ten microliters of each divalent cation stock solution (Cl- salt) or EDTA (Tris-
buffered, pH 7.2) was added to the reaction tube at time zero to give the indicated final concentration. Any precipitate resulting from the addition of NaOH after the reaction was complete was removed by filtration. Incubations contained 73.4 (normal) and 43.6 (reeler) pg protein.
inhibitor. Substrate saturation kinetics (Fig. 4) yielded similar k:, values for normal (1.3 mM) and reeler (1.1 mM) enzymes.
The effect of divalent cations on cerebellar alkaline phosphatase is pre- sented in Table 1. None of the six divalent cations tested sfimulated enzyme activity. The data suggest that reeler alkaline phosphatase is more sensitive to cation inhibition than normal enzyme. Nevertheless, endogenous cations seem to be required for both phosphatase activities because 10 mM EDTA (ethylenediaminetetraacetate) inhibited these enzymes by 70%.
To maximize the activity of this membrane-bound enzyme (3), cerebella were homogenized in the presence of 1% Triton X-100, which yields
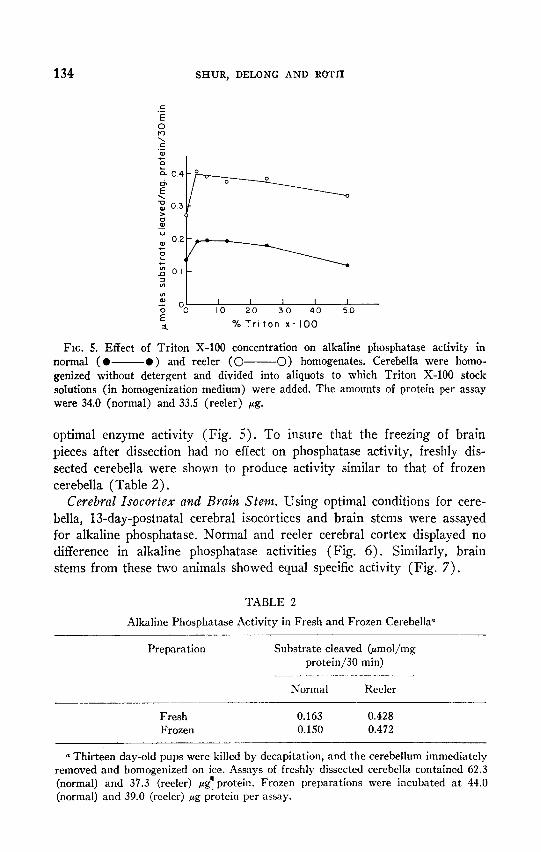
134 SHUR, DELONG AND ROTH
E 3. % Trl ion x- 100
FIG. 5. Effect of Triton X-100 concentration on alkaline phosphatase activity in normal ( l - l ) and reeler (O- 0) homogenates. Cerebella were homo- genized without detergent and divided into aliquots to which Triton X-100 stock solutions (in homogenization medium) were added. The amounts of protein per assay were 34.0 (normal) and 33.5 (reeler) pg.
optimal enzyme activity (Fig. 5). To insure that the freezing of brain pieces after dissection had no effect on phosphatase activity, freshly dis- sected cerebella were shown to produce activity similar to that of frozen cerebella (Table 2).
Cerebral Isocortex and Brain Stem. Using optimal conditions for cere- bella, 13-day-postnatal cerebral isocortices and brain stems were assayed for alkaline phosphatase. Normal and reeler cerebral cortex displayed no difference in alkaline phosphatase activities (Fig. 6). Similarly, brain stems from these two animals showed equal specific activity (Fig. 7).
TABLE 2
Alkaline Phosphatase Activity in Fresh and Frozen Cerebella”
Preparation Substrate cleaved (Nmol/mg protein/30 min)
Normal Reeler
Fresh 0.163 0.428 Frozen 0.150 0.472
a Thirteen day-old pups were killed by decapitation, and the cerebellum immediately removed and homogenized on ice. Assays of freshly dissected cerebella contained 62.3
(normal) and 37.3 (reeler) rg: protein. Frozen preparations were incubated at 44.0 (normal) and 39.0 (reeler) rg protein per assay.

INCREASED PIIOSPIIATASE ACTIVITY IN REELER 135
30 45
minutes
FIG. 6. Time course of normal (a----- O) and reeler (O- 0) cereberal
isocortex homogenate alkaline phosphatase activity. The amounts of protein per incubation tube were 120 (normal) and 112 (reeler) pg.
Age Dependence
Cerebella. Alkaline phosphatase activities in normal and reeler cerebella were determined for a variety of ages. Figure 8 indicates that normal ad reeler enzyme activity per milligram protein was similar early in cerebellar
minutes
1~~ 7. .Time course of normal (O---O) and reeler (O---- 0) brain stcn~
homogenate aIkaline phosphatase activity. The amounts of protein per assay were 49.8 (normal) and 43.8 (reeler) pg.
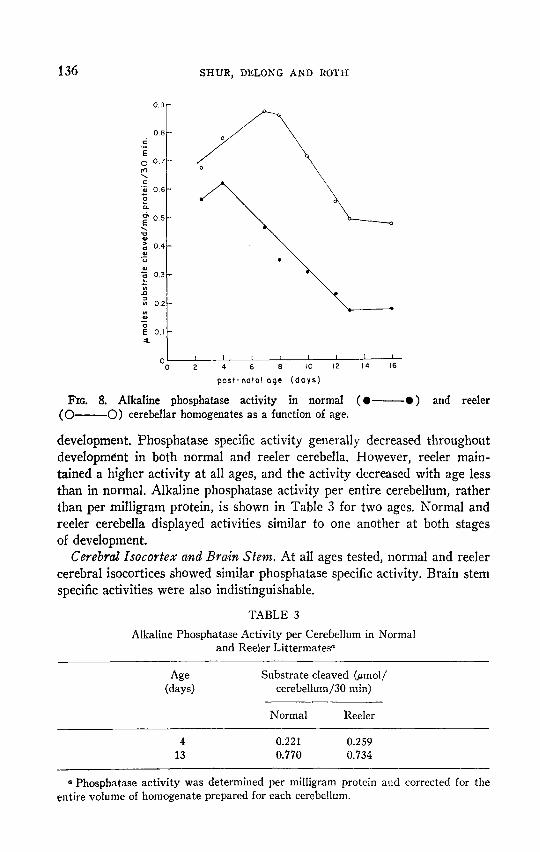
136 SHUR, DELONG AND ROTH
os- .c E
Cl o’-
2 c
‘5 0.6- z k E 0.5-
\ z ; 0.4-
z
s 0.3 - L ITI 0 : 0.2-
: z E 0.1 - P
post-natal age (days)
FIG. 8. Alkaline phosphatase activity in normal ( l - l ) and reeler
(0-O) cerebellar homogenates as a function of age.
development. Phosphatase specific activity generally decreased throughout development in both normal and reeler cerebella, However, reeler main- tained a higher activity at all ages, and the activity decreased with age less than in normal. Alkaline phosphatase activity per entire cerebellum, rather than per milligram protein, is shown in Table 3 for two ages. Normal and reeler cerebella displayed activities similar to one another at both stages of development.
Cerebrd Isocortex and Brain Stem, At all ages tested, normal and reeler cerebral isocortices showed similar phosphatase specific activity. Brain stem specific activities were also indistinguishable.
TABLE 3
Alkaline Phosphatase Activity per Cerebellum in Normal and Reeler Littermates
Age Substrate cleaved (Irmol/
Wvs) cerebellum/30 min)
4 13
Normal Reeler
0.221 0.259 0.770 0.734
0 Phosphatase activity was determined per milligram protein aud corrected for the
entire volume of homogenate prepared for each cerebellum.

INCREASED PHOSPHATASE ACTIVITY IN REELER 137
Histochemistry
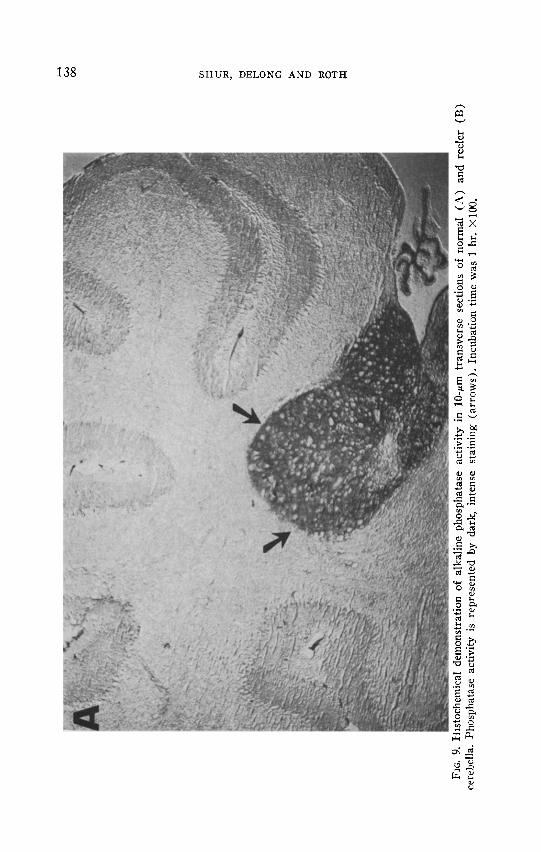
Histochemical localization of alkaline phosphatase in 13-day cerebella, using naphthol AS-MX phosphate as the substrate, is pictured in Fig. 9. Sections were cut perpendicular to the long axis of the cerebellum. In normal cerebellum (A), the region of the deep cerebellar nuclei near the union of the cerebellum and brain stem was stained predominantly. In reeler (B), enzyme activity persisted throughout the entire cerebellum, being more concentrated in the presumptive medullary region. The outer membrane which covers normal and reeler cerebella also possesses alkaline phosphatase. A very faint staining is detectable in the molecular layers and fiber tracts. The histochemical patterns suggest that cell processes, fibers, or both, rather than cell bodies, possess the enzyme activity.
Unlike 13-day cerebella, normal and reeler 4-day-postnatal cerebella show similar alkaline phosphatase staining patterns (not shown). Similar ceil types stain in 13- and 4-day cerebella but occupy a greater percentage of the total cerebellum at 4 days than at 13 days postnatal. Reeler enzyme activity appears more diffuse than does wild type, similar to 13-day-old cerebella.
DISCUSSION
The neurological mutant, reeler, shows severe ataxia, retarded cerebellar development, and abnormal cell lamination in the cerebral isocortex, hippo- campus, and cerebellum (2, 4, 8, 9). The data presented here show that reeler cerebella have three times more alkaline phosphatase activity per milligram protein than do wild-type or heterozygous cerebella. Normal and reeler cerebral isocortices have equal specific activities, as do brain stems from these animals.
Histochemical determination of alkaline phosphatase activity in 13-day normal cerebella shows staining primarily in the deep cerebellar nuclei. In reeler cerebella, the constituent cells of the deep cerebellar nuclei remain scattered throughout the white matter, not aggregating to form these struc- tures (4). Consequently, enzyme activity remains dispersed throughout the tissue.
The threefold increase in reeler alkaline phosphatase specific activity is probably due to the abundance of phosphatase-free tissue in normal cerebella. Phosphatase activity per cerebellum is similar in 13-day normal and reeler, and the high reeler specific activity can be attributed to its small size with retention of the deep nuclei cells. Three results are con- sistent with this explanation of cerebellar size-dependent specific activity. First are the similar phosphatase specific activities in normal and reeler cerebral isocortex. Cells are misaligned in reeler isocortex, but this organ
138 SHUR, DELONG AND ROTH

INCREASED PIIOSI’HATASE ACTIVITY IN REELER 139

140 SHUR, DELONG AND ROTH
retains its normal size (1, 8, 9). Second, similar K,,, values suggest that increased reeler activity is probably not due to a modified form of the enzyme. Finally, normal and reeler enzyme specific activities are similar early in development. However, as phosphatase-free tissue proliferates more rapidly in normal than in reeler cerebella, the specific activity diminishes with development to a lower level in the wild type.
Murine cerebellar alkaline phosphatase is similar to many mammalian alkaline phosphatases with one exception. Exogenous divalent cations fail to stimulate the activity of these enzymes. Why these cations inhibit reeler phosphatases more than normal phosphatases is not understood. Marked inhibition of alkaline phosphatases by EDTA implies the necessity of some endogenous cation.
Preliminary observations on another neurological mutant, weaver, shows this mutant to possess increased alkaline phosphatase specific activity, as in reeler. Homozygous weaver cerebella are also grossly reduced in size (6). Unlike reeler heterozygotes, weaver heterozygotes show intermediate alkaline phosphatase activities, although they do not show any significant reduction in the size of the cerebellum (6).
REFERENCES
1. DELONG, G. R., AND R. L. SIDMAN. 1970. Alignment defect of reaggregating cells in cultures of developing brains of Reeler mutant mice. Develop. Biol. 22: 584-600.
2. FALCONER, D. S. 1951. Two new mutants, “Trembler” and “Reeler,” with neuro- logical actions in the house mouse (Mus mzcscuZus L.). J. Genet. 50: 192-201.
3. FERNLEY, H. N. 1971. Mammalian alkaline phosphatases. Pages 417-447. irt P. BOYER, Ed., The Enzymes, Vol. IV. Academic Press, New York.
4. HAMBURGH, M. 1963. Analysis of the postnatal developmental effects of “Reeler,” a neurological mutation in mice. A study in developmental genetics. Develop.
Biol. 8: 165-185. 5. LAYNE, E. 1957. Spectrophotometric and turbidimetric methods for measuring
proteins. Pages 447-454 ilt S. P. COLOWICK AND N. 0. KAPLAN, Eds., Methods in Enzymology Vol. III. Academic Press, New York.
6. RAKIC, P., AND R. L. SIDMAN. 1973. Weaver mutant mouse cerebellum: Defective neuronal migration secondary to abnormalities of Bergmann glia. Prcx. Nat.
Acad. Sci. USA 71: 240-244. 7. SHUR, B. D., AND S. ROTH. 1975. Cell surface glycosyltransferases. Biochim.
Biophys. Acta 415 : 473-512. 8. SIDMAN, R. L. 1968. Development of interneuronal connections in brains of mutant
mice. Pages 163-193 ilz F. D. CARLSON, Ed., Physiological and Biochemical Aspects of Nervous Integration. Prentice-Hall, Englewood Cliffs, New Jersey.
9. SIDMAN, R. L. 1972. Cell interactions in developing mammalian central nervous system. Pages 1-13 in L. G. SILVESTRI, Ed., Cell Interactions, Third Leptit
Colloquium. American Elsevier, New York.
Copyright © 2022 FDOKUMEN




















