
IMMUNOPRECIPITATION OF BIOSYNTHETICALLY-LABELLED PRODUCTS IN THE IDENTIFICATION OF ANTIGENS OF...
7
Aust. J. Exp. Biol. Med. Sci., 60 (Pt. 1) 91-96 (1982) © IMMUNOPRECIPITATION OF BIOSYNTHETICALLY LABELLED, STAGE-SPECIFIC PROTEINS FROM CULTURED PLASMODIUM FALCIPARUM USING INHIBITORY HUMAN SERA by PETER MYLER. SUYING CHIU. JACQUELINE UPCROFT, DANIEL CASTELINO AND C H E V KIDSON (From the Oueensland Institute of Medical Research, Brisbane, Australia 4006.) (Accepted for publication October 28, J981.) Summary. Proteins from synchronized continuous cullures of Ptasmodium falciparum were biosynthetically labelled with [•'•'''S]methionine at ring, trophozoite, and schizont stages. Several proteins appeared to be synthesized predominantly at one stage only, especially the schizont stage. Immunoprecipitation using inhibitory human sera revealed that a variety of proteins, from all stages of parasite growth, were recognized. More proteins, especially those with high molecular weight, were precipitated from the schizont stage than from ring and trophozoite stages, but two major proteins (MW — 70,000 daltons and -— 45.000 daltons) were recognized in all three stages. INTRODUCTION Identification of specific antigens from erythrocytic stages of the malarial parasite Plasmodium falciparum is the concern of a number of investigators as a prerequisite for the development of protein vaccines. Kilejian (1980) reported that serum from an immune Nigerian adult reacted mainly with proteins labelled in the schizont stage of an African (Gambian) P. falciparum strain. More recently, Perrin, Dayal and Rieder (1981) found that immune sera from Africans living in malaria endemic areas in The Gambia and Tanzania precipitated a number of schizont proteins and at least one major protein (MW—76,000 daltons) from both ring and trophozoite stages of another African (Senegal) parasite strain. In addition, sera from individuals recovering from their first infection with P. falciparum appeared to precipitate only some of the schizont proteins. Using naturally synchronous, primary parasite isolates from malaria patients in Papua New Guinea, Brown et al. (1982) observed that immune sera from Papua New Guinea adults recog- nized antigens in all life-cycle stages and that they precipitated a larger number of proteins from schizonts than from rings and trophozoites. In the present study we have analyzed radiolabelled P. falciparum stage-specific antigens derived from highly synchronous cultures of a Papua New Guinea parasite strain, using sera from Papua New Guinean adults which inhibit in vitro invasion of erythrocytes by merozoites.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of IMMUNOPRECIPITATION OF BIOSYNTHETICALLY-LABELLED PRODUCTS IN THE IDENTIFICATION OF ANTIGENS OF...

Aust. J. Exp. Biol. Med. Sci., 60 (Pt. 1) 91-96 (1982)
© IMMUNOPRECIPITATION OF BIOSYNTHETICALLYLABELLED, STAGE-SPECIFIC PROTEINS FROM CULTURED
PLASMODIUM FALCIPARUM USING INHIBITORYHUMAN SERA
by PETER MYLER. SUYING CHIU. JACQUELINE UPCROFT,DANIEL CASTELINO AND C H E V KIDSON
(From the Oueensland Institute of Medical Research, Brisbane, Australia 4006.)
(Accepted for publication October 28, J981.)
Summary. Proteins from synchronized continuous cullures of Ptasmodiumfalciparum were biosynthetically labelled with [•'•'''S]methionine at ring, trophozoite,and schizont stages. Several proteins appeared to be synthesized predominantly atone stage only, especially the schizont stage. Immunoprecipitation usinginhibitory human sera revealed that a variety of proteins, from all stages ofparasite growth, were recognized. More proteins, especially those with highmolecular weight, were precipitated from the schizont stage than from ring andtrophozoite stages, but two major proteins (MW — 70,000 daltons and -— 45.000daltons) were recognized in all three stages.
INTRODUCTIONIdentification of specific antigens from erythrocytic stages of the malarial
parasite Plasmodium falciparum is the concern of a number of investigatorsas a prerequisite for the development of protein vaccines. Kilejian (1980)reported that serum from an immune Nigerian adult reacted mainly withproteins labelled in the schizont stage of an African (Gambian) P. falciparumstrain. More recently, Perrin, Dayal and Rieder (1981) found that immunesera from Africans living in malaria endemic areas in The Gambia andTanzania precipitated a number of schizont proteins and at least one majorprotein (MW—76,000 daltons) from both ring and trophozoite stages ofanother African (Senegal) parasite strain. In addition, sera from individualsrecovering from their first infection with P. falciparum appeared to precipitateonly some of the schizont proteins. Using naturally synchronous, primaryparasite isolates from malaria patients in Papua New Guinea, Brown et al.(1982) observed that immune sera from Papua New Guinea adults recog-nized antigens in all life-cycle stages and that they precipitated a largernumber of proteins from schizonts than from rings and trophozoites.
In the present study we have analyzed radiolabelled P. falciparumstage-specific antigens derived from highly synchronous cultures of a PapuaNew Guinea parasite strain, using sera from Papua New Guinean adultswhich inhibit in vitro invasion of erythrocytes by merozoites.
92 P. MYLER. S. CHIU. J. UPCROFT, D. CASTELINO AND C . KIDSON
MATERIALS AND METHODS
Growth and xynclironizution of P. falciparum
Plasmodium falciparum-infected eryihrocytes were maintained in continuous culture(Lamont. Saul and Kidson, 1981) with erythrocyies at haemalocrits of ~-59c. TheFCQ-27/PNG line (Chen et al, 1980) was used throughout. Cultures were synchronized by amodification of the method of Lambros and Vanderberg (1979). Asynchronous cultures weresuspended in a solution of 5 ^ sorbitol for 5 min at room temperature. This resulted in lysisof all erythrocytes containing parasites older than 33 h after merozoiie invasion (unpublisheddata). A second treatment followed between 7 and 17 h later. A third sorbitol lysis, exactly24 h after the first, resulted in a high degree of synchrony with the next round of invasionoccuring over a period of 6 to 8 h. After this initial round of sorbitol lysis, synchrony wasmaintained by sorbitol treatments as required, either immediately after invasion to eliminateslow growing parasites or 24 h after invasion to eliminate rapidly growing parasites.
Biosynthetic labellinfi of parasite stages
Stage-specific labelling of parasite proteins was carried out using 20 f*Ci/mI[•'•"•Slmethionine I 1160 Ci mmol.. Radiochemical Centre. Amersham. UK.) in methionine-freemedium {RPMI 1640: GIBCO. Grand Island. New York. U.S.A.) modified for malariagrowth (Trager and Jensen. 1976) plus 10% human serum. A 20 ml culture of synchronousP. fiilcipeiruin-'miectcii erythrtKytes was subjected to a sorbitol treatment immediately followinga round of merozoitc invasion and split into four ."i ml aliquots which were labelled asindicated in Table I. After growth in the presence of [•'"'Slmethionine as indicated, thecells in each culture were centrifuged at 700 ,i? for 5 min and the pellet suspended in 5 ml ofphosphate buffered saline (PBS). Parasites were released from the erythrocytes by addition of120 «l of 10% saponin (Sigma. St. Louis. U.S.A.). The parasites were centrifuged at 10,000 gfor 10 min. resuspended in 100 ;'l of ?>''•', Triton X-IOO in 150 mM NaCI. 5 mM EDTA. 50[TiM Tris pH 7 4 (Triton-NET buffer; Kessler, 1975), Triton-insoluble material was removedby centrifugation at i().t)OO a for 10 min. The supernatant containing Triton-soluble[•-•••SImethionine-iabelled parasite material was stored frozen at • 20° until used. The Triton-insoluble pellet contained no different protein species when compared with the Triton-solublefraction (unpublished data).
TABLE 1Biosynthetic luhellinn of P. falciparum cultures with [•'^S]methionine.
Stacelabelled
young ringslate ringstrophozoitesschizonis
time*
0-13 h13-23 h23-35 h35-47 h
parasitaemia when['JSJmethionine added
% rings I
10098
33
% trophs- %
02
9765
schizontS''
000
31
parasitaemia whenharvested
% ring.s
10064
37 ^
9c trophs
09472
2
'~/c schizonts
00
2460
The piirasitaemia of all cultures remained constant ;u 2 9'"r throughout ihe labellingperiod except when parasites labelled as schizonts were harvested by which time theparasitaemia had risen to 5 4%.
Differential parasite counts were obtained from Giemsa-siained thin blood films, byexamining cells close to the thin end of the film.
' vacuolated parasites occupying <309^ of diameter of erythrocyte.-' solid basophiiic staining with <2 nuclei and occupying >30ri of diameter of
er>throcyte,•* parasites with >2 nuclei.* hours after the sorbitol treatment.+ less than 3 h old.

STAGE-SPECIFIC MALARIA PARASITE PROTEINS 93
Immunoprecipitation of labelled proteinsA pool of sera from 5 Papua New Guinean adults was used for immunoprecipitation.
These sera were shown individually to inhibit merozoite invasion of eryihrocytes in vitro,to an extent comparable to that demonstrated by another serum sample tested extensively in aquantitative, automated invasion assay (Saul et at.. 1982). Immunoprecipitation of thelabelled proteins was carried out essentially according to ihe technique described by Kessler(1975), using 0 1 % Triton-NET buffer. The immunoprecipitation was done on 20 ^l ofTriton-soluble supernatant from each stage, and the total proteins precipitated in each casewere used for electrophoretic analysis.
Polyacrylamide gel eleetrophoresis (SOS-PAGE)This was carried out essentially according to the method of Laemmli and Favre (1973),
using 5-15% gradient gels.
RESULTSAnalysis of Triton-soluble [ •' ''S]methionine-labelled proteins and
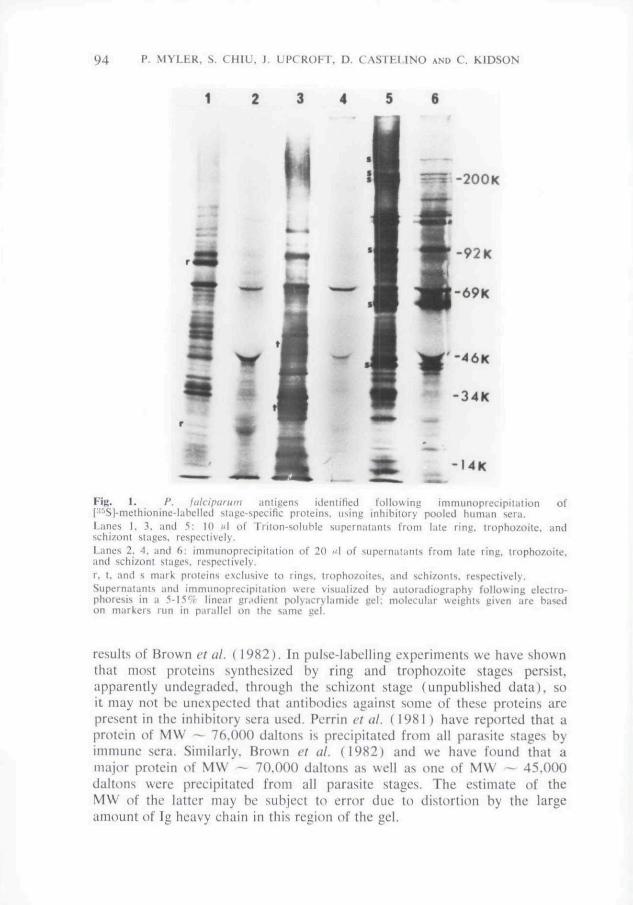
immunoprecipitation of these proteins from late ring (13-23 h), trophozoite(23-35 h) and schizont (35-47 h) stages is shown in Fig. L Several highmolecular weight proteins (MW > 100,000) appeared to be synthesizedalmost exclusively in the schizont stage while other particular proteinsappeared to be synthesized predominantly at ring or trophozoite stages.Examples of ring, trophozoite and schizont-specific proteins arc marked r, t,and s, respectively. At least 28 major protein bands were precipitated fromthe schizont stage using the pooled sera. Fewer proteins from ring andtrophozoite stages appeared to be recognized by the sera but two majorproteins of approximately 70,000 and 45,000 daltons, respectively, wereprecipitated.
Control immunoprecipitations using sera from non-exposed adults orStaphylococcus aureus alone gave a low level of non-specific precipitationin these experiments, less than 10% of the amount precipitated by thepooled inhibitory sera. In general, the level of non-specific precipitation usingcontrol sera was no greater than with 5. aureus alone, indicating that thisbackground was largely due to interaction of parasite proteins with the S.aureus.
DISCUSSIONThere are several proteins which appear to be labelled predominantly at
particular stages of malaria parasite growth. This is in agreement with recentreports (Kilejian, 1980; Perrin et al, 1981; Brown et al, 1982). Thereappear to be several proteins labelled specifically during the ring or tropho-zoite stages only, as well as the high molecular weight proteins labelledduring the schizont stage reported previously (Kilejian, 1980).
The finding of immunoprecipitation by adult sera of a substantial num-ber of proteins synthesized during ring and trophozoite stages as well asthose synthesized by schizonts contrasts somewhat with the results of Kilejian(1980) and Perrin et al (1981), but is essentially in agreement with the
9 4 P- MYLER. S. CHIU. J. UPCROFI. D. CASTELINO AND C . KIDSON
= ^ -200K
"-46K
-34K
- I 4 K
FIR. L /'. fulciporum antigens identified following immunoprecipiiaiion of[•'''Si-methionine-labelleJ stage-specific proteins, using inhibitory pooled human sera.t-anes 1. 3. and .*•: 10 M! of Triton-soluble superniitarits from l:ite ring, trupbozoite. andschizont stages, respectively,Lanes 2. 4. and 6: immunoprecipitation of 20 "I of supematants from late ring, trophozoite,and schizon! stages, respectively.r, t, and s mark proteins exclusive to rings, tropho/oitcs, and schizonts. respectively.Supernaiants and immunoprecipitation were visualized by autoradiography following eleetro-phoresis in a 5-15'̂ f linear grjilient polyacrylamide gel; molecular weights given arc basedon markers run in parallel on the same gel.
results of Brown et al { 1982). In pulsc-Iabclling experiments we have shownthat most proteins synthesized by ring and trophozoite stages persist,apparently undcgradod, through the schi/ont stage (unpublished data), soit may not be unexpected that antibodies against some of these proteins arepresent in the inhibitory sera used. Porrin et al. (1981) have reported that aprotein of MW ~~ 76.000 daltons is precipitated from all parasite stages byimmune sera. Similarly. Brown et al. ( 19K2) and we have found that amajor protein of MW - 70.000 daltons as well as one of MW -- 45.000daltons were precipitated from all parasite stages. The estimate of theMW of the latter may be subject to error due to distortion by the largeamount of Ig heavy chain in this regioti of the gel.

STAGE-SPECIFIC MALARIA PARASITE PROTEINS 95
Several possible explanations have been suggested by Brown et al.(1982) for the apparent differences between their data and those of Kilejian(1980). Brown et al (1982) labelled cultures with [-^Hlleucine. Kilejian(1980) used [-'Hlproline. while Perrin et al (1982) and the present studiesused |-'''Slmethioniiie. We have found that there is essentially no differencein protein bands visualized by SDS-PAGE after labelling with these threelabelled amino acids (unpublished data). Thus, it is unlikely that the use ofthe different labelled atnino acids would account for the differing immuno-precipitation results.
Brown et al (1982) used naturally synchronous primary isolates ofP. falciparum as opposed to our long-term in vitro cultures which weresynchronized artificially. Hence, it would appear that parasites subjectedto long-term culture continue to synthesize a large number of antigens recog-nized by precipitating antibodies present in sera from adults living in malariaendemic areas.
Both the present study and that of Brown et al (1982) have usedpooled sera and parasite strains from Papua New Guinea, whereas Africansera and parasite strains were employed by Kilejian (1980) and by Perrinet al (1981). This may possibly explain the differences seen between antigensprecipitated from ring and trophozoite stages. Clearly, cross-comparisonsare needed to test this possibility.
The results presented here show that several proteins were precipi-tated from ring and trophozoite stages by a pool of inhibitory sera as wellas a large number of proteins from the schizont stage of P. falciparum. Thispoints to the need to consider antigens synthesized by all stages of theparasite when analyzing the human immunological response to malarialinfection.
Acknowledgements. This study received financial support from the UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases and from theNational Health and Medical Research Council of Australia. We would like to thank tbe RedCross Blood Bank, Brisbane, for supplying human erythrocytes and serum. We thank Dr. G.Brown for the opportunity to see his data prior to publication.
REFERENCESBROWN, G. V., COPPEI., R. L.. VRBOVA. H.,
GRUMONT. R. J., and ANDERS, R. F .
(1982): 'Plasmodium falciparum: Com-parative analysis of erythrocyte stage-dependent protein antigens.' Exp. Para-sitol, accepted for publication.
CHEN. P., LAMONT. G . L.. ELLIOT, T.,
KIDSON, C , BROWN, G.. MITCHELL, G.,
STACK, J., and ALPERS, M. (1980): T/«.v-modium fatciparutn strains from PapuaNew Guinea: culture characteristics anddrug sensitivity.' Southeast Asian J.Trop. Med. Puh. filth., t!, 435.
KESSLER, S. W. (t975): 'Cell membraneantigen isolation with staphylococcal pro-tein A-antibody adsorbent." / . Immunol,115. 1617.
KILEJIAN, A. (1980): 'Stage-specific pro-teins and glycoproteins of Plasmodiumfalciparum: Identification of antigensunique to schizonts and merozoites.' Proc.Nat. Acad. Sci., 77, 3695.
LAEMMLI, U . K., and FAVRE, M . (1973):
'Maturation of the head of bacteriophageT4. DNA packaging events.' /. Mol.Biol. 80, 575.

96 p. MYLER, S. CHIU. J. UPCROFT. D. CASTELINO AND C. KIDSON
LAMBROS, C , and VANDERBERG, J. P.(1979): "Synchronization of Plasmodiumfatciparum erythrocytic stages in culture.'/. Para.^itol.. 65, 418.
LAMONT. G., SAUL, A., and KIDSON, C.(1981): 'Plasmodium falciparum: Assayof invasion of erythrocytes.' Exp. Para-sitol. 5L 74.
PERRIN. L. H., DAYAL, R., and RIEDER. H .(1981): 'Characterization of antigensfrom erythrocytic stages of Plasmodium
falciparum reacting with human immunesera.' Trans. R. Soc. Trop. Med. Hyg..75. 163.
SAUL, A., MYLER, P., MANGAN, T., andKIDSON, C . (1982): 'PUumodluni falci-parum: Automated assay of erythrocyteinvasion using flow cytofluorimetry.'Exp. Parasitol. accepted for publication.
TRAGER. W., and JENSEN. J. B. (1976);'Human malaria parasites in continuousculture.' Seienee, 193, 674.





















