
Perkinsozoa, a well-known marine protozoan flagellate parasite group, newly identified in lacustrine...
12
DISREGARDED DIVERSITY AND ECOLOGICAL POTENTIALS Review Article Perkinsozoa, a well-known marine protozoan flagellate parasite group, newly identified in lacustrine systems: a review Jean-Franc ¸ois Mangot • Didier Debroas • Isabelle Domaizon Received: 19 August 2009 / Accepted: 12 April 2010 / Published online: 16 May 2010 Ó Springer Science+Business Media B.V. 2010 Abstract The recurrent detection of parasitic zoo- spores among aquatic heterotrophic flagellates (HFs) has recently modified our view of how the microbial loop is organized, and called into question the role of eukaryotic parasites in the aquatic trophic food web. The Perkinsozoa group, already known to play a significant role as parasite in marine systems, is of special interest here, since it has recently been detected in several lakes by constructing clone libraries. In marine systems, this group is known to consist solely of intracellular parasites of molluscs or phytoplanktonic species, but their hosts in freshwater environments are still unknown, and little is yet known about their functional importance in plank- tonic systems. This review summarizes the main information currently available about Perkinsozoa through a description of their phylogenetic position, their life cycles, and regulatory factors, and the consideration of the specificities of their hosts in marine systems, and the few data recently acquired in lakes. Keywords Perkinsozoa Á Eukaryotic parasites Á Marine and lacustrine systems Introduction Numerous molecular studies have recently character- ized picoplanktonic assemblages by amplifying and sequencing the gene coding for 18S rRNA gene, both in marine and freshwater systems (Dı ´ez et al., 2001; Moreira & Lo ´pez-Garcı ´a, 2002; Lefranc et al., 2005; Lepe `re et al., 2006, 2008; Lefe `vre et al., 2008). These data have called into question the role of eukaryotic parasites in aquatic systems. More precisely, the importance of putative parasites within small eukary- otes has made it clear that the concept of a microbial loop within which heterotrophic flagellates (HFs) being thought to be mainly bacterivorous (Azam et al., 1983; Sanders et al., 1989) does not adequately describe the various interactions within the microbial food web. The presence of parasitic zoospores among HFs has led us to envisage greater taxonomic and functional heterogeneity among these heterotrophs, Guest editors: T. Sime-Ngando & N. Niquil / Disregarded Microbial Diversity and Ecological Potentials in Aquatic Systems J.-F. Mangot (&) Á I. Domaizon INRA – UMR 42 CARRTEL, Centre Alpin de Recherche sur les Re ´seaux Trophiques des Ecosyste `mes Limniques, Thonon-les-bains Cedex 74203, France e-mail: [email protected] J.-F. Mangot Á D. Debroas LMGE, Laboratoire ‘‘Microorganismes: Ge ´nome & Environnement’’, UMR CNRS 6023, Campus des Ce ´zeaux, 24, av. des Landais, Aubie `re Cedex 63177, France 123 Hydrobiologia (2011) 659:37–48 DOI 10.1007/s10750-010-0268-x
-
Upload
nationalagriculturalresearchinra -
Category
Documents
-
view
4 -
download
0
Transcript of Perkinsozoa, a well-known marine protozoan flagellate parasite group, newly identified in lacustrine...

DISREGARDED DIVERSITY AND ECOLOGICAL POTENTIALS Review Article
Perkinsozoa, a well-known marine protozoan flagellateparasite group, newly identified in lacustrine systems:a review
Jean-Francois Mangot • Didier Debroas •
Isabelle Domaizon
Received: 19 August 2009 / Accepted: 12 April 2010 / Published online: 16 May 2010
� Springer Science+Business Media B.V. 2010
Abstract The recurrent detection of parasitic zoo-
spores among aquatic heterotrophic flagellates (HFs)
has recently modified our view of how the microbial
loop is organized, and called into question the role of
eukaryotic parasites in the aquatic trophic food web.
The Perkinsozoa group, already known to play a
significant role as parasite in marine systems, is of
special interest here, since it has recently been
detected in several lakes by constructing clone
libraries. In marine systems, this group is known to
consist solely of intracellular parasites of molluscs or
phytoplanktonic species, but their hosts in freshwater
environments are still unknown, and little is yet
known about their functional importance in plank-
tonic systems. This review summarizes the main
information currently available about Perkinsozoa
through a description of their phylogenetic position,
their life cycles, and regulatory factors, and the
consideration of the specificities of their hosts in
marine systems, and the few data recently acquired in
lakes.
Keywords Perkinsozoa � Eukaryotic parasites �Marine and lacustrine systems
Introduction
Numerous molecular studies have recently character-
ized picoplanktonic assemblages by amplifying and
sequencing the gene coding for 18S rRNA gene, both
in marine and freshwater systems (Dıez et al., 2001;
Moreira & Lopez-Garcıa, 2002; Lefranc et al., 2005;
Lepere et al., 2006, 2008; Lefevre et al., 2008). These
data have called into question the role of eukaryotic
parasites in aquatic systems. More precisely, the
importance of putative parasites within small eukary-
otes has made it clear that the concept of a microbial
loop within which heterotrophic flagellates (HFs)
being thought to be mainly bacterivorous (Azam
et al., 1983; Sanders et al., 1989) does not adequately
describe the various interactions within the microbial
food web. The presence of parasitic zoospores among
HFs has led us to envisage greater taxonomic and
functional heterogeneity among these heterotrophs,
Guest editors: T. Sime-Ngando & N. Niquil / Disregarded
Microbial Diversity and Ecological Potentials in Aquatic
Systems
J.-F. Mangot (&) � I. Domaizon
INRA – UMR 42 CARRTEL, Centre Alpin de Recherche
sur les Reseaux Trophiques des Ecosystemes Limniques,
Thonon-les-bains Cedex 74203, France
e-mail: [email protected]
J.-F. Mangot � D. Debroas
LMGE, Laboratoire ‘‘Microorganismes: Genome &
Environnement’’, UMR CNRS 6023, Campus des
Cezeaux, 24, av. des Landais, Aubiere Cedex 63177,
France
123
Hydrobiologia (2011) 659:37–48
DOI 10.1007/s10750-010-0268-x

particularly if parasitism is taken into account.
Parasitism is indeed a model of interaction, which
increases the complexity of the trophic network by
extending food chains, increasing connectance, and
the efficiency of transfers; particularly as these
parasites have relatively complex life cycles, and
are known to infect organisms of various trophic
levels (Arias-Gonzales & Morand, 2006).
Most aquatic organisms are known to contain
intracellular symbionts (viruses, archaea, bacteria,
fungi, and other eukaryotes parasites) (Canter &
Lund, 1969; Cole, 1982; Embley et al., 1992;
Holfeld, 1998; Erard-Le Denn et al., 2000; Brussaard,
2004; Park et al., 2004). Also, while viral parasitism
on bacterioplankton in aquatic systems has been well
documented for the last 10 years (Fuhrman, 1999;
Wommack & Colwell, 2000; Bettarel et al., 2003,
2004; Weinbauer, 2004), studies focusing on eukary-
otic parasites have generally been overlooked. Prob-
ably because little is yet known about the identity and
diversity of these parasites in aquatic systems,
eukaryotic parasitism is often poorly taken into
account in functional models (Arias-Gonzales &
Morand, 2006; Lafferty et al., 2006). However, some
significant regulating effects have been reported in
the literature, and have highlighted the significant
impact of parasitism on algal community control in
marine systems (Johansson & Coats, 2002; Park
et al., 2002; Ibelings et al., 2004; Kagami et al., 2004,
2010). For instance, Syndiniales parasites are able to
infect marine dinoflagellates, which are responsible
for toxic red tides, also known as harmful algal
blooms (HAB), which can cause illness and even
human death (Zingone & Enevoldsen, 2000). Cham-
bouvet et al. (2008) recently revealed that the dino-
flagellate parasitoid successions in a natural estuary
are correlated to the rapid development of four major
species of phytoplanktonic dinoflagellate populations
(Heterocapsa rotundata, Scrippsiella trochoidea,
Alexandrium minutum, and Heterocapsa triquetra).
On the basis of such examples, it is obvious that
planktonic food web models must now take the role
of eukaryotic parasites into account (Arias-Gonzales
& Morand, 2006; Lafferty et al., 2006, 2008).
Parasitic interactions increase the complexity of the
trophic network both through direct effects (host–cell
mortality, parasitic zoospores used as a resource by
grazers) and indirect impacts (cascade effects on the
competition between phytoplanktonic species, and on
the redistribution and recycling of organic matter
after cell lysis) that modify connectance and the
efficiency of transfers (Kagami et al., 2004, 2010;
Arias-Gonzales & Morand, 2006).
In aquatic ecosystems, three major eukaryotic
lineages are known to include potentially parasitic
organisms. Organisms belonging to the fungal group
of Chytridiomycota are known mainly as eukaryotic
parasites of phytoplankton (Goldstein, 1960; Canter
& Lund, 1969; Ibelings et al., 2004). Some para-
sites have also been reported within the Cercozoa
(Rhizaria phyla), in which a handful of taxa are
parasites of diatoms (Cavalier-Smith & Chao, 2003).
However, alveolates exhibit the most considerable
diversity of known eukaryotic parasites, including
various morphologically distinctive parasites among
the three traditional phyla of Ciliophora, Dinoflagel-
lata, and Apicomplexa (Cavalier-Smith, 1993) and
the newly included Chromerida (Moore et al., 2008;
Obornık et al., 2009). Parasitic activities of alveolates
have been reported in a wide range of ecosystems
(soil, aquatic, etc.). In order to begin with, apicom-
plexans (ex-sporozoan) and ciliates are major para-
sites of marine and terrestrial populations
(crustaceans, fish, or mammals), and can cause
extensive health and economic damage to human
populations, particularly those in the developing
world (Hakimi & Deitsch, 2007). Leaving aside
these two alveolates phyla, Dinoflagellata constitute
the main source of aquatic parasites (Blastodiniales,
Syndiniales). Among them, Syndiniales constitute an
atypical order of dinoflagellates, consisting of well-
known and exclusively marine parasites (Hematodi-
nium spp., Amoebophrya spp.), which display a host-
specific infection strategy (Groisillier et al., 2006;
Chambouvet et al., 2008; Guillou et al., 2008).
Finally, Chromera velia, a newly cultured photosyn-
thetic symbiont of the stony coral Plesiastrea purpu-
rea, constitutes the latest symbiotic alveolates defined
until now (Moore et al., 2008). Based on the available
sequences, chromerids may form a sister group to the
parasitic apicomplexans and predatory colpodellids
rather than the photoautotrophic dinoflagellates
(Moore et al., 2008; Obornık et al., 2009), and its
discovery allows to speculate on the origin of
symbiosis and parasitism among the Alveolata.
The Perkinsozoa group, a sister-group of the
dinoflagellates (also known as perkinsids or perkins-
eans), is structured around two well-known marine
38 Hydrobiologia (2011) 659:37–48
123

parasites, Perkinsus marinus (Mackin et al., 1950)
and Parvilucifera infectans (Noren et al., 1999)
(Fig. 1). Perkinsids are intracellular parasites that
form a relatively close sister lineage to generally free-
living dinoflagellates (Noren et al., 1999; Leander &
Keeling, 2003). Little is known about Perkinsozoa
diversity, especially in freshwater, and this consti-
tutes a real handicap for attempts to understand the
role of parasites in aquatic systems more precisely.
Since the middle of the 2000s, based on recent
descriptive studies (Lefranc et al., 2005; Richards
et al., 2005; Lefevre et al., 2007; Lepere et al., 2008)
attempts have been made to characterize the compo-
sition of the picoeukaryotic population (\5 lm) in
several lakes using cloning–sequencing approaches.
The recurrent detection of sequences of putative
parasites in these studies explains the new interest in
this group of eukaryotic parasites in lacustrine
systems where their presence had hitherto been
unknown. We propose to review the main informa-
tion available about this phylogenetic group in both
marine and freshwater systems.
The phylogenetic position of Perkinsozoa phyla
among the lineage of alveolates
Perkinsids have mainly been studied in marine
environments (Mackin et al., 1950; Noren et al.,
1999; Park et al., 2002, 2004, 2006; Casas et al.,
Fig. 1 Phylogenetic tree showing the position of each clades
of Perkinsozoa among the alveolates based on their small
subunit ribosomal RNA (SSU rRNA) sequences. The names of
the lakes in which sequences were identified are indicated with
their access number. Numbers at branch nodes indicate
significant bootstrap confidence values in percent (BV
[90%). The tree was constructed (maximum parsimony
criteria), and the BVs for terminal nodes were calculated both
using the ARB software package (Ludwig et al., 2004)
Hydrobiologia (2011) 659:37–48 39
123

2008). Although there is now strong evidence that
perkinsozoans constitute a sister group of dinoflagel-
lates (Moore et al., 2008), their systematic position
has long been controversial. They have been alterna-
tively linked to Apicomplexa or to Dinoflagellata
(Coss et al., 2001; Brugerolle, 2002). Initially, the
classification of organisms within this genus was
based on a variety of morphological criteria, espe-
cially at the ultrastructural scale. The debate about
their phylogenetic position is illustrated by the
example of Perkinsus marinus, the best-known
parasite in this group. Initially described as a fungus
named Dermocystidium marinus by Mackin et al.
(1950), on the basis of the observation of the position
of the nucleus close to the cell wall and the presence
of a centrally located endosome, a subsequent
reexamination of its morphological features led to
its being transferred to the protozoan phylum Laby-
rinthomorpha, and designated as Labyrinthomixa
marina (Mackin & Ray, 1966). The evaluation of
zoospore ultrastructure by Levine (1978) led to a
further reconsideration of the phylogenetic position
of Perkinsus marinus and the establishment of a new
Perkinsean taxon in the protozoan phylum Apicom-
plexa. Indeed, zoospores of this species differ from
those of all other apicomplexans in having a mastig-
oneme-bearing anterior flagellum, and having an
atypical conoid apparatus on the apical complex
structure. More recently, studies based on morpho-
logical observations and phylogenetic analyses of
alveolate DNA sequences, and especially actin gene
sequences (Reece et al., 1997; Siddall et al., 1997),
led to their exclusion from the phylum Apicomplexa,
and the conclusion that Perkinsus was related to the
dinoflagellates. However, on the basis of ultrastruc-
tural features and phylogenetic studies of the 18S
rRNA sequence of a new parasitic species, Parvilu-
cifera infectans, Noren et al. (1999) established a new
phylum, Perkinsozoa, within the Alveolata. However,
the apparent widespread serological affinity between
Perkinsus marinus and diverse parasitic dinoflagel-
lates suggested a closer phylogenetic link to the
syndinean dinoflagellate lineage (Bushek et al.,
2002). This newly established phylum is assumed to
be entirely parasitic (Moreira & Lopez-Garcıa, 2002),
and it consists of two freshwater clades as defined by
Lepere et al. (2008) by applying the criteria of Zwart
et al. (2002). According to these authors, an envi-
ronmental clade comprises at least two sequences that
are at least 95% identical and originate from at least
two different aquatic sites. Thus, we have defined two
clades inside the phylum of Perkinsozoa (named
clade 1 and clade 2), each structured around two
marine species of parasites, Perkinsus marinus (par-
asite of molluscs) and Parvilucifera infectans (para-
site of dinoflagellates) (Fig. 1). Indeed, in order to
determine the phylogenetic affiliation of perkinsozo-
an species reported here with other congeneric
species, a phylogenetic analysis thanks to ARB
software was performed (Ludwig et al., 2004). The
rooted tree identified two clades highly supported by
bootstrap value (BV) of 99% (Fig. 1). Among the
second clade of Perkinsozoa named clade 2, Parvilu-
cifera infectans with other environmental sequences
reported here constituted a higher-order clade with a
strong BV (98%). Perkinsus marinus, often consid-
ered as part of the Perkinsozoa clade 1, seems to form
only an additional branch apart of this clade which is
constituted by environmental sequences supported by
a strong BV (100%).
Perkinsozoa in marine systems: a natural cause
of diseases in oyster cultures and of toxic
dinoflagellate proliferations
Perkinsus spp. are known to be parasites of molluscs.
These hosts are easily sampled in situ, moreover,
problems involving infected molluscs have an eco-
nomic impact as many of these species are commer-
cialized; consequently Perkinsus spp. are the most
studied perkinsozoan species. Recently, an assay tool
based on a polymerase chain reaction coupled with
restriction fragment length polymorphism (PCR–
RFLP) of the rRNA internal-transcribed spacers (ITS
region) has made it possible to target these parasites.
This methodology was validated to discriminate
between the various Perkinsus species (P. marinus,
P. olseni, P. mediterraneus, and P. chesapeaki), to
provide rapid identification, and the final goal was to
prevent recurrent diseases (Abollo et al., 2006).
Perkinsus marinus is a well-known agent of the
disease known as ‘‘Dermo’’, which is the main cause
of mortality of bivalve molluscs in American coastal
waters. The eastern oyster, Crassostrea virginica, and
the Pacific oyster, Crassostrea cortenziensis, are both
parasited by Perkinsus marinus, along the Gulf of
Mexico and Atlantic coast of North America
40 Hydrobiologia (2011) 659:37–48
123

(Burreson et al., 1994; Soniat, 1996), and along the
Pacific coast of Mexico, respectively (Caceres-Martı-
nez et al., 2008). In European coastal waters, some
species of the Perkinsus genus are also able to infect
other bivalve hosts. This is the case of Perkinsus
mediterraneus, a parasite of the European flat oyster
Ostrea edulis (Casas et al., 2008), and P. atlanticus, a
parasite of the Portuguese clam Ruditapes decussatus
(Azevedo, 1989; Ordas & Figueras, 1998).
As illustrated by these examples, Perkinsus spe-
cies are very widely geographically distributed,
which partly reflects their host distribution (Fig. 2).
It seems that a variety of levels of host specificities
can be observed. Perkinsus marinus seems to solely
infect the oyster Crassostrea, while, the clam Rudi-
tapes decussatus known to be infected by P. atlan-
ticus in European waters can also be parasited by P.
qugwadi on the west coast of Canada (Blackbourn
et al., 1998).
Most of the Perkinsus spp. seem to be specific to
one type of bivalve (clams or oysters); however,
some of them are able to infect various types of host,
such as Perkinsus olseni which is a parasite of both
clams and abalones. Depending on its geographic
location, P. olseni, which is known to infect the
Spanish Ruditapes decussatus in Mediterranean
waters (Elandaloussi et al., 2009), elsewhere also
infects the Korean Venus clam Protothaca jedoensis
(Park et al., 2006) and the Australian abalone species,
Haliotis rubra and Haliotis laevigata (Lester et al.,
1990). Perkinsid infection of species other than
molluscs has been reported with the case of
P. chabelardi, a parasite of sardine eggs (Gestal
et al., 2006). The prevalence of P. marinus, P.
atlanticus, and P. olseni on their respective hosts are
highly variable, and can sometimes reach very high
values (up to 100%) (Burreson et al., 1994; Ngo &
Choi, 2004). Seasonal variations have been reported
by several authors (Ngo & Choi, 2004; Gullian-
Klanian et al., 2008); these changes in prevalence are
linked to various environmental factors (temperature,
salinity, phosphorous, silica variations, etc.).
The Perkinsozoa group also includes various
parasites of protistan species. Parvilucifera infectans
is known to infect 26 different microalgal species, 17
of which belong to 10 different dinoflagellate genera
(Park et al., 2004). The distribution of this parasite
extends from Australian to Norwegian seas, where
they commonly infect photosynthetic and heterotro-
phic dinoflagellates (Park et al., 2004) (Fig. 2).
Although the rare mortality rates estimated until
now were low (\0.2% of the host population per day,
Park et al. 2004), several authors suggest that
Parvilucifera infectans could have a significant
regulatory effect on its host, contributing to bloom
dissipation and controlling dinoflagellate populations
(Gisselson et al., 2002; Park et al., 2004).
Other species phylogenetically close to P infectans
have recently been isolated and studied. They include
Parvilucifera sinerae sp. nov., a perkinsid isolated
from a bloom of the toxic dinoflagellate Alexandrium
minutum in Mediterranean Sea (Figueroa et al.,
Fig. 2 Global distribution
of Perkinsus spp. (whiteellipsoidal symbol) and of
Parvilucifera spp. (grayellipsoidal symbol). Each
symbol notes locations
where infections have been
observed in molluscs or
phytoplanktonic organisms
Hydrobiologia (2011) 659:37–48 41
123
2008), and Parvilucifera prorocentri sp. nov., a novel
parasite found to infect the marine benthic dinofla-
gellate Prorocentrum fukuyoi (Leander & Hoppen-
rath, 2008). The pathogenicity of Perkinsus spp. has
been relatively thoroughly investigated and quanti-
fied, but this is not the case for Parvilucifera
spp. which constitute a newly described group of
parasites for which no quantitative prevalence data
are available.
Life cycle and regulatory factors of Perkinsozoa
phyla in aquatic systems
The life cycles of the three genera of perkinsids,
Perkinsus spp., Parvilucifera spp., and Cryptophagus
spp., display some common characteristics. For
instance, like chytrids, perkinsids have a free living
stage (a zoosporic flagellate) during their life cycle
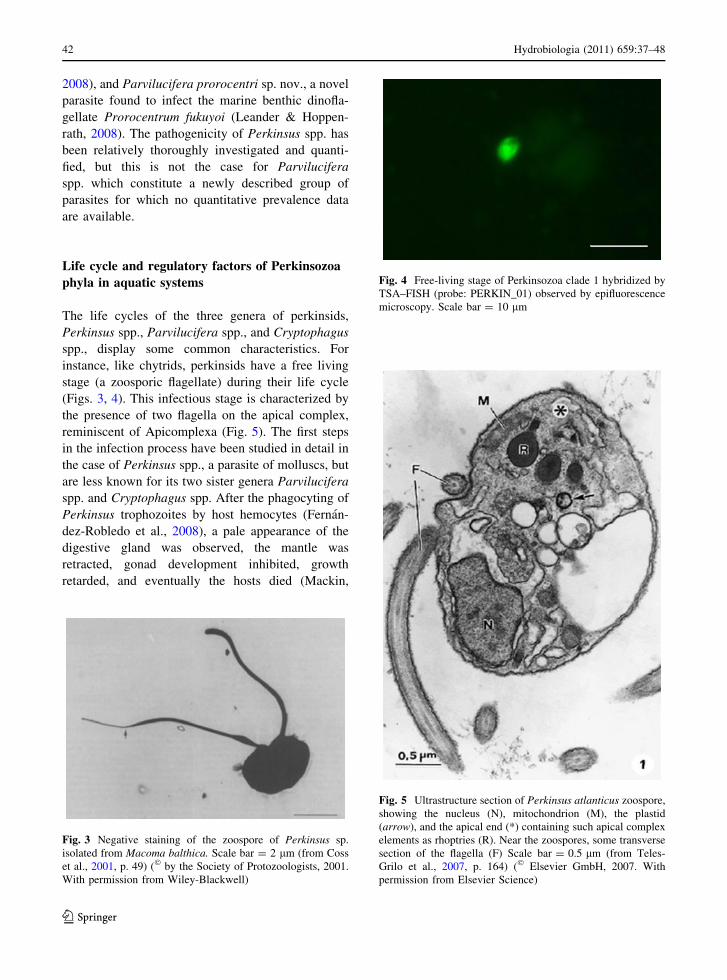
(Figs. 3, 4). This infectious stage is characterized by
the presence of two flagella on the apical complex,
reminiscent of Apicomplexa (Fig. 5). The first steps
in the infection process have been studied in detail in
the case of Perkinsus spp., a parasite of molluscs, but
are less known for its two sister genera Parvilucifera
spp. and Cryptophagus spp. After the phagocyting of
Perkinsus trophozoites by host hemocytes (Fernan-
dez-Robledo et al., 2008), a pale appearance of the
digestive gland was observed, the mantle was
retracted, gonad development inhibited, growth
retarded, and eventually the hosts died (Mackin,
Fig. 3 Negative staining of the zoospore of Perkinsus sp.
isolated from Macoma balthica. Scale bar = 2 lm (from Coss
et al., 2001, p. 49) (� by the Society of Protozoologists, 2001.
With permission from Wiley-Blackwell)
Fig. 4 Free-living stage of Perkinsozoa clade 1 hybridized by
TSA–FISH (probe: PERKIN_01) observed by epifluorescence
microscopy. Scale bar = 10 lm
Fig. 5 Ultrastructure section of Perkinsus atlanticus zoospore,
showing the nucleus (N), mitochondrion (M), the plastid
(arrow), and the apical end (*) containing such apical complex
elements as rhoptries (R). Near the zoospores, some transverse
section of the flagella (F) Scale bar = 0.5 lm (from Teles-
Grilo et al., 2007, p. 164) (� Elsevier GmbH, 2007. With
permission from Elsevier Science)
42 Hydrobiologia (2011) 659:37–48
123

1951). The rest of the life cycle of Perkinsozoa shows
some similarities among the three genera, such as the
presence of a similar intracellular stage, the trophont.
Perkinsus spp. and Parvilucifera spp. trophonts
secrete a ‘‘sporangium’’ inside the host cell, within
which the sporozoites differentiate (Noren et al.,
1999; Coss et al., 2001), whereas Cryptophagus spp.
zoospores grow and directly divide within the host
cytoplasm and are not surrounded by a sporangium or
a cyst envelope (Brugerolle, 2002). Thanks to these
studies, a general life cycle scenario of perkinsids
parasites can now be proposed: After the adhesion of
the sporozoite to the host cell surface, the zoospore
penetrates into the host cytoplasm. The trophont then
grows within the host cytoplasm, and undergoes
multiple divisions either in sporangia (Perkinsus spp.
and Parvilucifera spp.) or within the host cytoplasm
(Cryptophagus spp.). After 4–5 successive nuclear
mitoses by schizogony, trophozoite/sporozoite differ-
entiation occurs, and buds onto the host cell surface
(Fig. 6). Sporozoites acquire flagella, micronemes
and other apical structures, and are then release
outside the host structure. Sometimes, a resting cyst-
like stage can occur during the trophozoite stage.
It seems likely that the development of populations
of Perkinsozoa parasites could be controlled by
various abiotic factors, notably temperature. Indeed,
the proliferation of all the Perkinsus species is
correlated with warm summer water temperatures
(above 20�C), which is when pathogenicity and the
associated mortality are the highest (Soudant et al.,
2005). The same authors report a slight, but signif-
icant, decrease in the percentage of viable cells
(PVC) after cold or heat treatments (-80, -20, and
40�C). Furthermore, temperature may have also an
impact on the development of the hosts of Perkinsus.
The gonadal development of Ruditapes philippina-
rum appeared to be more efficient at higher temper-
ature (18�C, Delgado & Camacho, 2007). As a result,
temperature may have direct and indirect effects, on
the viability of Perkinsozoa species and the metab-
olism of their hosts, respectively. However, some of
them are more tolerant to variations in various
environmental factors. P. atlanticus is able to adapt
to very different culture media, salinity, and temper-
ature conditions, and its development seems to be
independent of density (Ordas & Figueras, 1998).
Detection and recent interest in the Perkinsozoa
phylum in lacustrine systems
The characterization of small-eukaryote diversity
(\5 lm) conducted in various lakes with differing
trophic status (Lefranc et al., 2005) has recently
revealed the presence of Perkinsozoa in freshwater
systems. This study showed that sequences affiliated to
Perkinsozoa can account for up to 60% of the clone
library (18S rRNA gene) obtained from the eutrophic
lake Aydat (summer sampling in the euphotic zone).
The sequences obtained for this lake were affiliated
with low similarities (86–90%) to Perkinsus marinus.
The unexpected qualitative importance of perkinsids
in lacustrine systems led to the emergence of numer-
ous questions about their functional roles in lacustrine
food webs. Several recent studies also dealing with the
Fig. 6 Light micrograph of the different live stages of cultured
Perkinsus mediterraneus cells in JL-ODRP-2F medium.
Trophozoites (T) showing a granular cytoplasm with a large
vacuole (v) and a prominent vacuoplast (vp). Perkinsusmediterraneus divides by schizogony, and schizonts (S) vary
in size. Scale bar: 20 lm (from Casas et al., 2008, p. 37) (� by
the International Society of Protistologists, 2008. With
permission from Wiley-Blackwell)
Hydrobiologia (2011) 659:37–48 43
123

characterization of lacustrine picoeukaryotic diversity
have confirmed the recurrent presence and importance
of this group. Perkinsozoa have been detected in two
geographically distinct, oligotrophic lakes, the Amer-
ican Lake George (euphotic zone) (Richards et al.,
2005) and the French Lake Pavin, respectively (oxic
and oxycline zones) (Lefevre et al., 2007, 2008). More
recently, Lepere et al. (2008) have reported their
presence in the 18S rRNA gene library of perkinsids
sequences in the epilimnion of Lake Bourget in spring
and summer, providing semi-quantitative information
about this group. In this mesotrophic system,
sequences of Perkinsozoa account for 15.2% of the
different OTUs obtained during the summer (August)
(29.2% of the clones), whereas in May, the proportion
was only about 5% of both sequences and clones. All
these perkinsid sequences obtained from sequencing in
various lacustrine systems can be divided to form two
clades of Perkinsozoa (named clade 1 and clade 2).
Recently, on the basis of these molecular data, specific
oligonucleotide probes have been designed to target
each clade of Perkinsozoa to quantify and estimate the
in situ dynamics of this group in Lake Bourget
(Mangot et al., 2009). The temporal dynamics of the
free-living stage (zoospore) of clade 1 and clade 2 of
Perkinsozoa (size fraction \5 lm), as analyzed by
epifluorescence microscopy and TSA-FISH (Fig. 4),
have all revealed an increase in abundance in summer
in the epilimnion of the Lake Bourget (up to 30% of all
eukaryotes targeted by the EUK1209R probe in July
and August), possibly corresponding to an infectious
peak. As already reported in marine systems (Soudant
et al., 2005), the dynamics of freshwater perkinsozo-
ans are strongly correlated to temperature (Lefranc
et al., 2005, Lefevre et al., 2007, 2008; Lepere et al.,
2008; Mangot et al., 2009). In Lake Bourget, the
dynamics of Perkinsozoa clade 1 coincide significantly
(RDA analysis, P \ 0.05) with the abundance pat-
terns of Peridinium and Ceratium (Dinoflagellates),
whereas the dynamics of Perkinsozoa clade 2 are
correlated to the presence of Dinobryon (Chrysophy-
ceae). Although some putative phytoplanktonic host–
Perkinsozoa links could be suggested on the basis of
this study, no definite association (host/parasite) was
demonstrated. Only one study has reported the pres-
ence of an algal-parasitic Perkinsozoa, Rastrimonas
subtilis gen. et sp. nov (Brugerolle, 2003) in freshwater
environments. This taxon was previously known as
Cryptophagus subtilis (Brugerolle, 2002), a parasite of
cryptophytes. The ultrastructure and life cycle of this
freshwater perkinsid have been studied using cultures
of the cryptophyte Chilomonas paramaecium, origi-
nally collected from a river. Because no genetic
characterization of this taxon was obtained by
sequencing, its affiliation to one of the two clades of
Perkinsozoa defined here is not possible. However, the
description of its ultrastructure given by Brugerolle
strongly supports the affiliation of R. subtilis to the
phylum of Perkinsozoa (Brugerolle, 2002).
Conclusion
It is now obvious that eukaryotic parasitism, hitherto
generally overlooked in aquatic studies, could in fact
play a critical role in the functioning of these
ecosystems. For instance, the Perkinsozoa, a sister-
group of dinoflagellates, are a good example of
organisms that have generally been classified as
unidentified HFs, but which could affect planktonic
populations. These parasites have mainly been stud-
ied in marine environments, where they constitute
one of the main causes of mortality in some molluscs
(oysters, clams, abalones, etc.) and phytoplanktonic
species (especially, dinoflagellates) worldwide. Valu-
able knowledge has been acquired about the role of
some Perkinsozoa in marine systems, but very little
information is known about this parasite group in
freshwater systems. It is only recently, thanks to the
development of molecular techniques, that Perkinso-
zoa has been detected and quantified in lakes. Some
tools are now available that can target these groups
(notably specific oligonucleotide probes), but further
research is now needed to connect the potential
quantitative importance of these parasites to their
precise functional roles in lacustrine systems, notably
through cell–scale approaches. Isolating infected
hosts, coupled with molecular analysis (genome
amplification) of these single cells could reveal the
association between lacustrine perkinsids and their
hosts. The use of double staining (host/parasite) using
TSA–FISH would, in a second step, provide some
essential information about prevalence and host/
parasite in situ dynamics. A combination of conven-
tional methods (isolation, microscopic observations)
and molecular techniques should soon help elucidate
questions such as the diversity of Perkinsozoa in
lacustrine systems, their real function, and the
44 Hydrobiologia (2011) 659:37–48
123

identity of their hosts in this system, or even make it
possible to draw a parallel between their roles in
marine and in freshwater systems.
References
Abollo, E., S. M. Casas, G. Ceschia & A. Villalba, 2006.
Differential diagnosis of Perkinsus species by polymerase
chain reaction-restriction fragment length polymorphism
assay. Molecular and Cellular Probes 20: 323–329.
Arias-Gonzales, J. E. & S. Morand, 2006. Trophic functioning
with parasites: a new insight for ecosystem analysis.
Marine Ecology Progress Series 320: 43–53.
Azam, F., T. Fenchel, J. G. Field, J. S. Gray, L. A. Meyer-Reil
& F. Thingstad, 1983. The ecological role of water col-
umn microbes in the sea. Marine Ecology Progress Series
10: 257–263.
Azevedo, C., 1989. Fine structure of Perkinsus atlanticus n. sp.
(Apicomplexa, Perkinsea) parasite of the clam Ruditapesdecussatus from Portugal. Journal of Parasitology 75:
627–635.
Bettarel, Y., C. Amblard, T. Sime-Ngando, J. F. Carrias, D.
Sargos, F. Garabetian & P. Lavandier, 2003. Viral lysis,
flagellate grazing potential and bacterial production in
Lake Pavin. Microbial Ecology 45: 119–127.
Bettarel, Y., T. Sime-Ngando, C. Amblard & J. Dolan, 2004.
Viral activity in two contrasting lake ecosystems. Applied
and Environmental Microbiology 70: 2941–2951.
Blackbourn, J., S. M. Bower & G. R. Meyer, 1998. Perkinsusqugwudi sp. nov. (incertae sedis), a pathogenic protozoan
parasite of Japanese scallops, Patinopecten yessoensis,
cultured in British Columbia, Canada. Canadian Journal
of Zoology 76: 942–953.
Brugerolle, G., 2002. Cryptophagus subtilis: a new parasite of
cryptophytes affiliated with the Perkinsozoa lineage.
European Journal of Protistology 37: 379–390.
Brugerolle, G., 2003. Apicomplexan parasite Cryptophagusrenamed Rastrimonas gen. nov. European Journal of
Protistology 39: 101.
Brussaard, C. P. D., 2004. Viral control of phytoplankton
populations – a review. The Journal of Eukaryotic
Microbiology 51: 125–138.
Burreson, E. M., R. S. Alvarez, V. Vidal, M. Leopoldina & A.
Macedo, 1994. Perkinsus marinus (Apicomplexa) as a
potential source of oyster Crassostrea virginica mortality
in coastal lagoons of Tabasco, Mexico. Diseases of
Aquatic Organisms 20: 73–82.
Bushek, D., C. F. Dungan & A. J. Lewitus, 2002. Serological
affinities of the Oyster Pathogen Perkinsus marinus(Apicomplexa) with some dinoflagellates (Dinophyceae).
The Journal of Eukaryotic Microbiology 49: 11–16.
Caceres-Martınez, J., R. Vasquez-Yeomans, G. Padilla-
Lardizabal, M. A. del Rıo & Portilla., 2008. Perkinsusmarinus in pleasure oyster Crassostrea corteziensis from
Nayarit, Pacific coast of Mexico. Journal of Invertebrate
Pathology 99: 66–73.
Canter, H. M. & J. W. G. Lund, 1969. The parasitism of
planktonic desmids by Fungi. Plant Systematics and
Evolution 116: 351–377.
Casas, S. M., K. S. Reece, Y. Li, J. A. Moss, A. Villalba & J. F.
La Peyre, 2008. Continuous culture of Perkinsus medi-terraneus, a parasite of the European Flat Oyster Ostreaedulis, and characterization of its morphology, propaga-
tion, and extracellular proteins in vitro. The Journal of
Eukaryotic Microbiology 55: 34–43.
Cavalier-Smith, T., 1993. Kingdom protozoa and its 18 phyla.
Microbiological Reviews 57: 953–994.Cavalier-Smith, T. & E. Y. Chao, 2003. Phylogeny and clas-
sification of phylum Cercozoa (Protozoa). Protist 154:
341–358.
Chambouvet, A., P. Morin, D. Marie & L. Guillou, 2008.
Control of toxic marine dinoflagellate blooms by serial
parasitic killers. Science 322: 1254–1257.
Cole, J. J., 1982. Interactions between bacteria and algae in
aquatic ecosystems. Annual Review of Ecology, Evolu-
tion and Systematics 13: 291–314.
Coss, C. A., J. A. F. Robledo & G. R. Vasta, 2001. Fine
structure of clonally propagated in vitro life stages of a
Perkinsus sp. isolated from the Baltic Clam Macomabalthica. The Journal of Eukaryotic Microbiology 48: 38–
51.
Delgado, M. & A. P. Camacho, 2007. Influence of temperature
on gonadal development of Ruditapes philippinarum(Adams and Reeve, 1850) with special reference to
ingested food and energy balance. Aquaculture 264: 398–
407.
Dıez, B., C. Pedros-Alio, T. L. Marsh & R. Massana, 2001.
Application of denaturing gradient gel electrophoresis
(DGGE) to study the diversity of marine picoeukaryotic
assemblages and comparison of DGGE with other
molecular techniques. Applied in Environmental Micro-
biology 67: 2942–2951.
Elandaloussi, L. M., N. Carrasco, A. Roque, K. Andree & M.
D. Furones, 2009. First record of Perkinsus olseni, a
protozoan parasite infecting the commercial clam Rudi-tapes decussatus in Spanish Mediterranean waters. Jour-
nal of Invertebrate Pathology 100: 50–53.
Embley, T. M., B. J. Finlay, R. H. Thomas & P. L. Dyal, 1992.
The use of rRNA sequences and fluorescent probes to
investigate the phylogenetic positions of the anaerobic
ciliate Metopus pdueformis and its archaeobacterial
endosymbiont. Journal of General Microbiology 138:
1479–1487.
Erard-Le Denn, E., M. J. Chretiennot-Dinet & I. Probert, 2000.
First report of parasitism on the toxic Dinoflagellate Al-exandrium minutum Halim. Estuarine, Coastal and Shelf
Science 50: 109–113.
Fernandez-Robledo, J. A., E. J. Schott & G. R. Vasta, 2008.
Perkinsus marinus superoxide dismutase 2 (PmSOD2)
localizes to single-membrane subcellular compartments.
Biochemical and Biophysical Research Communications
375: 215–219.
Figueroa, R. I., E. Garces, R. Massana & J. Camp, 2008.
Description, host-specificity, and strain selectivity of the
dinoflagellate parasite Parvilucifera sinerae sp. nov.
(Perkinsozoa). Protist 159: 563–578.
Fuhrman, J. A., 1999. Marine viruses and their biogeochemical
and ecological effects. Nature 399: 541–548.
Gestal, C., B. Novoa, D. Posada, A. Figueras & C. Azevedo,
2006. Perkinsoide chabelardi n. gen., a protozoan parasite
Hydrobiologia (2011) 659:37–48 45
123

with an intermediate evolutionary position: possible cause
of the decrease of sardine fisheries? Environmental
Microbiology 8: 1105–1114.
Gisselson, L. A., P. Carlsson, E. Graneli & J. Pallon, 2002.
Dinophysis blooms in the deep euphotic zone of the Baltic
Sea: do they grow in the dark? Harmful Algae 1: 401–418.
Gullian-Klanian, M., J. A. Herrera-Silveira, R. Rodriguez-Ca-
nul & L. Aguirre-Macedo, 2008. Factors associated with
the prevalence of Perkinsus marinus in Crassostreavirginica from the southern Gulf of Mexico. Diseases of
Aquatic Organisms 79: 237–247.
Goldstein, S., 1960. Physiology of aquatic fungi: nutrition of
two monocentric chytrids. Journal of Bacteriology 80:
701–707.
Groisillier, A., R. Massana, K. Valentin, D. Vaulot & L.
Guillou, 2006. Genetic diversity and habitats of two
enigmatic marine alveolate lineages. Aquatic Microbial
Ecology 42: 277–291.
Guillou, L., M. Viprey, A. Chambouvet, R. M. Welsh, A. R.
Kirkham, R. Massana, D. J. Scanlan & A. Z. Worden,
2008. Occurrence and genetic diversity of marine para-
sitoids belonging to Syndiniales (Alveolata). Environ-
mental Microbiology 10: 3349–3365.
Hakimi, M. A. & K. W. Deitsch, 2007. Epigenetics in Api-
complexa: control of gene expression during cell cycle
progression, differentiation and antigenic variation. Cur-
rent Opinion in Microbiology 10: 357–362.
Holfeld, H., 1998. Fungal infections of the phytoplankton:
seasonality, minimal host density, and specificity in a
mesotrophic lake. New Phytologist 138: 507–517.
Ibelings, B. W., A. De Bruin, M. Kagami, M. Rijkeboer, M.
Brehm & E. Van Donk, 2004. Review: host parasite
interactions between freshwater phytoplankton and chy-
trid fungi (Chytridiomycota). Journal of Phycology 40:
437–453.
Johansson, M. & D. W. Coats, 2002. Ciliate grazing on the
parasite Amoebophrya sp. decreases infection of the red-
tide dinoflagellate Akashiwo sanguinea. Aquatic Micro-
bial Ecology 28: 69–78.
Kagami, M., E. Van Donk, A. De Bruin, M. Rijkeboer & B. W.
Ibelings, 2004. Daphnia can protect diatoms from fungal
parasitism. The American Society of Limnology and
Oceanography 49: 680–685.
Kagami M., N. R. Helmsing & E. van Donk, 2010. Parasitic
chytrids could promote copepod survival by mediating
material transfer from inedible diatoms. Hydrobiologia.
doi:10.1007/s10750-010-0274-z.
Lafferty, K. D., A. P. Dobson & A. M. Kuris, 2006. Parasites
dominate food web links. PNAS 103: 11211–11216.
Lafferty, K. D., S. Allesina, M. Arim, C. J. Briggs, G. De Leo, A.
P. Dobson, J. A. Dunne, P. T. J. Johnson, A. M. Kuris, D. J.
Marcogliese, N. D. Martinez, J. Memmott, P. A. Marquet, J.
P. McLaughlin, E. A. Mordecai, M. Pascual, R. Poulin & D.
W. Thieltges, 2008. Parasites in food webs: the ultimate
missing links. Ecology Letters 11: 533–546.
Leander, B. S. & P. J. Keeling, 2003. Morphostasis in alveolate
evolution. Trends in Ecology and Evolution 18: 395–402.
Leander, B. S. & M. Hoppenrath, 2008. Ultrastructure of a
novel tube-forming, intracellular parasite of dinoflag-
ellates: Parvilucifera prorocentri sp. nov. (Alveolata,
Myzozoa). European Journal of Protistology 44: 55–70.
Lefevre, E., C. Bardot, C. Noel, J. F. Carrias, E. Viscogliosi, C.
Amblard & T. Sime-Ngando, 2007. Unveiling fungal
zooflagellates as members of freshwater picoeukaryotes:
evidence from a molecular diversity study in a deep
meromictic lake. Environmental Microbiology 9: 61–71.
Lefevre, E., B. Roussel, C. Amblard & T. Sime-Ngando, 2008.
The molecular diversity of freshwater picoeukaryotes
reveals high occurrence of putative parasitoids in the
plankton. PLoS ONE 3: 2324–2333.
Lefranc, M., A. Thenot, C. Lepere & D. Debroas, 2005.
Genetic diversity of small eukaryotes in lakes differing by
their trophic status. Applied in Environmental Microbi-
ology 71: 5935–5942.
Lepere, C., D. Boucher, L. Jardillier, I. Domaizon & D. Deb-
roas, 2006. Succession and regulation factors of small
eukaryote community composition in a lacustrine eco-
system (Lake Pavin). Applied in Environmental Micro-
biology 72: 2971–2981.
Lepere, C., I. Domaizon & D. Debroas, 2008. Unexpected
importance of potential parasites in the composition of the
freshwater small-eukaryote community. Applied in Envi-
ronmental Microbiology 74: 2940–2949.
Lester, R. J. G., C. L. Goggin & K. B. Sewell, 1990. Perkinsusin Australia. In Cheng, T. C. & F. O. Perkins (eds),
Pathology in Marine Aquaculture. Academic Press, New
York: 189–199.
Levine, N. D., 1978. Perkinsus gen. n. and other new taxa in
the protozoan phylum Apicomplexa. Journal of Parasi-
tology 64: 549.
Ludwig, W., O. Strunk, R. Westram, L. Richter, H. Meier, B.
Yadhukumar, A. Buchner, T. Lai, S. Steppi, G. Jobb, W.
Forster, I. Brettske, S. Gerber, A. W. Ginhart, O. Gross, S.
Grumann, S. Hermann, R. Jost, A. Konig, T. Liss, R.
Lussmann, M. May, B. Nonhoff, B. Reichel, R. Strehlow,
A. Stamatakis, N. Stuckmann, A. Vilbig, M. Lenke, T.
Ludwig, A. Bode & K. H. Schleifer, 2004. ARB: a soft-
ware environment for sequence data. Nucleic Acids
Research 32: 1363–1371.
Mackin, J. G., 1951. Histopathology of infection of Crassos-trea virginica Gmelin by Dermocystidium marinumMackin, Owen and Collier. Bulletin of Marine Science of
the Gulf and Caribbean 1: 72–87.
Mackin, J. G. & S. M. Ray, 1966. The taxonomic relationships
of Dermocystidium marinum Mackin. Owen and Collier.
Journal of Invertebrate Pathology 8: 544–545.
Mackin, J. G., H. M. Owen & A. Collier, 1950. Preliminary
note on the occurrence of a new protistan parasite,
Dermocystidium marinum n. sp. in Crassostrea virginica(Gmelin). Science 111: 328–329.
Mangot, J. F., C. Lepere, C. Bouvier, D. Debroas & I.
Domaizon, 2009. Community structure and dynamics of
small eukaryotes (\5 lm) targeted by new oligonucleo-
tide probes: a new insight into the lacustrine microbial
food web. Applied in Environmental Microbiology 75:
6373–6381.
Moore, R. B., M. Obornik, J. Janouskovec, T. Chrudimsky, M.
Vancova, D. H. Green, S. W. Wright, N. W. Davies, C. J.
Bolch, K. Heimann, J. Slapeta, O. Hoegh-Guldberg, J. M.
Logsdon & D. A. Carter, 2008. A photosynthetic alveolate
closely related to apicomplexan parasites. Nature 451:
959–963.
46 Hydrobiologia (2011) 659:37–48
123

Moreira, D. & P. Lopez-Garcıa, 2002. The molecular ecology
of microbial eukaryotes unveils a hidden world. Trends in
Microbiology 10: 31–38.
Ngo, T. T. T. & K. S. Choi, 2004. Seasonal changes of Perk-insus and Cercaria infections in the Manila clam Rudi-tapes philippinarum from Jeju, Korea. Aquaculture 239:
57–68.
Noren, F., O. Moestrup & A. S. Rehnstam-Holm, 1999. Par-vilucifera infectans Noren et Moestrup gen. et sp. nov.
(Perkinsozoa phylum nov.): a parasitic flagellate capable
of killing toxic microalgae. European Journal of Protis-
tology 35: 233–254.
Obornık, M., J. Janouskovec, T. Chrudimsky & J. Lukes, 2009.
Evolution of the apicoplast and its hosts: from heterotro-
phy to autotrophy and back again. International Journal
for Parasitology 39: 1–12.
Ordas, M. C. & A. Figueras, 1998. In vitro culture of Perkinsusatlanticus, a parasite of the carpet shell clam Ruditapesdecussatus. Diseases of Aquatic Organisms 33: 129–136.
Park, M. G., S. K. Cooney, W. Yih & D. W. Coats, 2002.
Effects of two strains of the parasitic dinoflagellate
Amoebophrya on growth, photosynthesis, light absorption,
and quantum yield of bloom-forming dinoflagellates.
Marine Ecology Progress Series 227: 281–292.
Park, M. G., W. Yih & D. W. Coats, 2004. Parasites and phy-
toplankton, with special emphasis on dinoflagellate infec-
tions. The Journal of Eukaryotic Microbiology 51: 144–155.
Park, K. I., T. T. T. Ngo, S. D. Choi, M. Cho & K. S. Choi,
2006. Occurrence of Perkinsus olseni in the Venus clam
Protothaca jedoensis in Korean waters. Journal of Inver-
tebrate Pathology 93: 81–87.
Reece, K. S., M. E. Siddall, E. M. Burreson & J. E. Graves,
1997. Phylogenetic analysis of Perkinsus based on actin
gene sequences. Journal of Parasitology 83: 417–423.
Richards, T. A., A. A. Vepritskiy, D. E. Gouliamova & S. A.
Nierzwicki-Bauer, 2005. The molecular diversity of
freshwater picoeukaryotes from an oligotrophic lake
reveals diverse, distinctive and globally dispersed lin-
eages. Environmental Microbiology 7: 1413–1425.
Sanders, R. W., K. G. Porter, S. J. Bennet & A. E. DeBiase,
1989. Seasonal patterns of bacterivory by flagellates, cil-
iates, rotifers, and cladocerans. Limnology and Ocean-
ography 34: 673–687.
Siddall, M. E., K. S. Reece, J. E. Graves & E. M. Burreson,
1997. ‘‘Total evidence’’ refutes the inclusion of Perkinsusspecies in the phylum Apicomplexa. Parasitology 115:
165–167.
Soniat, T. M., 1996. Epizootiology of Perkinsus marinus dis-
ease of eastern oysters in the Gulf of Mexico. The Journal
of Shellfish Research 15: 35–43.
Soudant, P., F. L. E. Chu & E. D. Lund, 2005. Assessment of
the cell viability of cultured Perkinsus marinus (Perkin-
sea), a parasitic protozoan of the Eastern Oyster, Cras-sostrea virginica, using SYBRgreen–propidium iodide
double staining and flow cytometry. The Journal of
Eukaryotic Microbiology 52: 492–499.
Teles-Grilo, M. L., J. Tato-Costa, S. M. Duarte, A. Maia, G.
Casal & C. Azevedo, 2007. Is there a plastid in Perkinsus
atlanticus (Phylum Perkinsozoa)? European Journal of
Protistology 43: 163–167.
Weinbauer, M. G., 2004. Ecology of prokaryotic viruses.
FEMS Microbiology Reviews 28: 127–181.
Wommack, K. E. & R. R. Colwell, 2000. Virioplankton:
viruses in aquatic ecosystems. Microbiology and Molec-
ular Biology Reviews 64: 69–114.
Zingone, A. & H. O. Enevoldsen, 2000. The diversity of
harmful algal blooms: a challenge for science and
management. Ocean & Coastal Management 43: 725–
748.
Zwart, G., B. C. Crump, M. P. Kamst-van Agterveld, F. Hagen
& S. K. Han, 2002. Typical freshwater bacteria: an
analysis of available 16S rNRA gene sequences from
plankton of lakes and rivers. Aquatic Microbial Ecology
28: 141–155.
Author Biographies
Jean-Francois Mangot is
a PhD student in lacustrine
microbial ecology at Blaise
Pascal (Clermont-Ferrand)
and Savoie Universities,
France. His PhD thesis
deals with the study of the
dynamics and the diversity
of potential eukaryotic par-
asite microorganisms in
lacustrine systems, espe-
cially the group of Perkin-
sozoa recently discovered in
freshwater ecosystems.
Didier Debroas is Profes-
sor at Blaise Pascal Univer-
sity in the fields of microbial
ecology and bioinformatics.
He began his scientific
career by studying the
microbial ecology of the
rumen ecosystem and he has
been working since 1993 on
the aquatic ecosystem. At
present, he is the leader of
the group ‘‘Environmental
Microbiology and Bioinfor-
matics’’ at the LMGE lab
(CNRS, Aubiere – http://
www.lmge.univ-bpclermont.fr). Currently, his main research
concerns (i) the taxonomic and functional diversities of lacus-
trine picoplankton and (ii) the community genetic potential of
these microorganisms. His research group is developing (i) some
methodologies for analysing diversity and functions at the cell
level (cell-sorting, metagenomics, etc.) and (ii) bioinformatic
tools for dealing with data sequences originated from massive
parallel sequencing.
Hydrobiologia (2011) 659:37–48 47
123

Isabelle Domaizon is
researcher at the hydrobio-
logical INRA center (CARR-
TEL, Thonon, France). She is
currently the leader of the
group ‘Biodiversity Func-
tioning Evolution of Lacus-
trine Ecosystem’. Her major
research interest deals with
the structure and functioning
of microbial food webs in
lakes; the general objective is
to identify the links between
the structure of planktonic
communities and the func-
tioning of lacustrine pelagic systems. The specific research
questions addressed concern (i) the diversity of microbial
eukaryotes, (ii) the trophic interactions and pathways within
microbial trophic networks and up to higher trophic levels, and (iii)
the regulatory factors involved in the dynamics, structure, diversity
and activity of planktonic micro-organisms.
48 Hydrobiologia (2011) 659:37–48
123




















