
A broad spectrum monoclonal antibody to alpha-tubulin does not recognize all protozoan tubulins
7
Protoplasma 128,201-207 (1985) I IOTOPI.ASNA by Springer-Verlag 1985 A Broad Spectrum Monoclonal Antibody to Alpha-Tubulin Does not Recognize all Protozoan Tubulins P. DRABER l, S. RUBINO 2, E. DRABEROVA 1, V. VIKLICK~, and P. CAPPUCCINELLI 2' * 1 Institute of Molecular Genetics, Czechoslovak Academy of Sciences, Prague 2 Institute of Medical Microbiology, University of Sassari, Sassari Received April 4, 1985 Accepted April 24, 1985 Summary Monoclonal antibodies able to recognize single antigenic determi- nants are a powerful tool for the study of immunological heterogene- ity of antigens. In this paper we have used a monoclonal antibody against the a-subunit of pig brain tubulin (TU-01) to investigate the immunoreactivity of tubulins from mammals, avians, amphibia, echinodermata, plathelmints, slime moulds and protozoa. Immuno- reactivity was detected using immunobloning and indirect im- munofluorescence of isolated cells. Our results show that the antigenic determinant recognized by the TU-01 antibody is present in all metazoan tubulin tested and among the eukaryotic microorgan- isms only in the flagellate Trichornonas vaginalis. Indirect im- munofluorescence also reveals that not all Trichomonas microtubules are stained by TU-01 antibody indicating the presence of different tubulins within a single ceil. This results are consistent with the multi- tubulin hypothesis (FULTON and SIMPSON 1976). Keywords: Monoclonal antibody; Microtubules; Immunofluores- cence; Immunoblotting. 1. Introduction Tubulin, the major component of microtubules, is composed of two different subunits a and 13 (for review see DUSTJN 1978). The amino acid sequences of tubulin subunits (KRAuHS et al. 1981, POST~NGL et al. 1981, VALENZUELA et al. 1981) show that tubulin is a highly conserved protein. It seems clear that not all micro- tubules in an organism are identical, since tissue- specific tubulin has been identified in several organisms (B~BRING et al. 1976, STEPHENS 1978). Peptide maps of * Correspondence and Reprints: Istituto di Microbiologia, Viale Mancini 5, 1-07100 Sassari, Italy. tubulins from various species have shown that the 13 tubulin of unicellular and metazoan eukaryotes are extremely similar (LITTLE et al. 1981), but peptide maps of a tubulin from sea urchin egg and sperm flagellar doublets are markedly different (LITTLE et al. 1981) as are the a tubulin of sheep brain compared with that of Physarum (CLAYTON et al. 1980). The multi-tubulin hypothesis of FULTONand SIMPSON (1976) has recently received several confirmations. Many organisms contain multiple tubulin genes (for review see CLEVELAYD 1983). Multiple forms of tubulin have been identified in the cytoskeletal and flagellar microtubules of the alga Polytomella (McKE~THAN and ROSEMBAUM 1981, McK~ITHAN et al. 1983), in a single neurone (GOZESand SWEADNER 1981), in Trypanosoma (GALLO and ANDERTON 1983) and in other eukaryotic microorganisms (manuscript in preparation). By using monoclonal antibodies it is possible to re- cognize different domains of tubulin in different species in order to study in more detail the structure and the heterogeneity of tubulin. Monoclonal antibodies have been obtained that react with the ct subunit of brain tubulin (AsAI et al. 1982, KILMARTIN et al. 1982) as well as to the 13 subunit (GozEs and BARNSTABLE 1982). In this paper, we report that a mouse monoclonal antibody raised against pig brain tubulin (NIKLICKY et al. 1982) reacts with the a subunit of porcine brain and the a tubulin band of electrophoretically separated mammalian cell lysates. This antibody reacts with microtubules from diverse species (mammalian, bird,
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A broad spectrum monoclonal antibody to alpha-tubulin does not recognize all protozoan tubulins

Protoplasma 128, 201-207 (1985) I IOTOPI.ASNA �9 by Springer-Verlag 1985
A Broad Spectrum Monoclonal Antibody to Alpha-Tubulin Does not Recognize all Protozoan Tubulins
P. DRABER l, S. RUBINO 2, E. DRABEROVA 1, V. VIKLICK~, and P. CAPPUCCINELLI 2' *
1 Institute of Molecular Genetics, Czechoslovak Academy of Sciences, Prague 2 Institute of Medical Microbiology, University of Sassari, Sassari
Received April 4, 1985 Accepted April 24, 1985
Summary
Monoclonal antibodies able to recognize single antigenic determi- nants are a powerful tool for the study of immunological heterogene- ity of antigens. In this paper we have used a monoclonal antibody against the a-subunit of pig brain tubulin (TU-01) to investigate the immunoreactivity of tubulins from mammals, avians, amphibia, echinodermata, plathelmints, slime moulds and protozoa. Immuno- reactivity was detected using immunobloning and indirect im- munofluorescence of isolated cells. Our results show that the antigenic determinant recognized by the TU-01 antibody is present in all metazoan tubulin tested and among the eukaryotic microorgan- isms only in the flagellate Trichornonas vaginalis. Indirect im- munofluorescence also reveals that not all Trichomonas microtubules are stained by TU-01 antibody indicating the presence of different tubulins within a single ceil. This results are consistent with the multi- tubulin hypothesis (FULTON and SIMPSON 1976).
Keywords: Monoclonal antibody; Microtubules; Immunofluores- cence; Immunoblotting.
1. Introduction
Tubulin, the major component of microtubules, is composed of two different subunits a and 13 (for review see DUSTJN 1978). The amino acid sequences of tubulin subunits (KRAuHS et al. 1981, POST~NGL et al. 1981, VALENZUELA et al. 1981) show that tubulin is a highly conserved protein. It seems clear that not all micro- tubules in an organism are identical, since tissue- specific tubulin has been identified in several organisms (B~BRING et al. 1976, STEPHENS 1978). Peptide maps of
* Correspondence and Reprints: Istituto di Microbiologia, Viale Mancini 5, 1-07100 Sassari, Italy.
tubulins from various species have shown that the 13 tubulin of unicellular and metazoan eukaryotes are extremely similar (LITTLE et al. 1981), but peptide maps of a tubulin from sea urchin egg and sperm flagellar doublets are markedly different (LITTLE et al. 1981) as are the a tubulin of sheep brain compared with that of Physarum (CLAYTON et al. 1980). The multi-tubulin hypothesis of FULTON and SIMPSON (1976) has recently received several confirmations. Many organisms contain multiple tubulin genes (for review see CLEVELAYD 1983). Multiple forms of tubulin have been identified in the cytoskeletal and flagellar microtubules of the alga Poly tome l la (McKE~THAN and ROSEMBAUM 1981, McK~ITHAN et al. 1983), in a single neurone (GOZES and SWEADNER 1981), in Trypanosoma
(GALLO and ANDERTON 1983) and in other eukaryotic microorganisms (manuscript in preparation). By using monoclonal antibodies it is possible to re- cognize different domains of tubulin in different species in order to study in more detail the structure and the heterogeneity of tubulin. Monoclonal antibodies have been obtained that react with the ct subunit of brain tubulin (AsAI et al. 1982, KILMARTIN et al. 1982) as well as to the 13 subunit (GozEs and BARNSTABLE 1982). In this paper, we report that a mouse monoclonal antibody raised against pig brain tubulin (NIKLICKY et al. 1982) reacts with the a subunit of porcine brain and the a tubulin band of electrophoretically separated mammalian cell lysates. This antibody reacts with microtubules from diverse species (mammalian, bird,

202 P. D~BER et al. : A Broad Spectrum Monoclonal Antibody to Alpha-Tubulin Does not Recognize all Protozoan Tubulins
a m p h i b i a n , fung i , e c h i n o d e r m , p l a t h e l m i n t , s l ime
m o u l d s a n d p r o t o z o a n cells), b u t d o e s n o t r e c o g n i z e al l
p r o t o z o a n t u b u l i n s .
2. Mater ia l s and Methods
2.1. Cells
Cells used in this work are summarized in Tab. 1. The human lung fibroblast line LEP and mouse neuroectodermal line C 1300 were obtained from Dr. M. Baudygov/t (Institute of Physiology, Prague); the rat kangaroo fibroblast line PtK 2 was obtained from Dr. J. Cells (Aarhus University, Denmark). Human T-lymphoblastoid line MOLT3, mouse B-lymphoblastoid line P3X63-Ag8.653, mouse embryonal carcinoma line F9 and mouse L-A9 fibroblasts are maintained in the bank of cell lines of Institute of Molecular Genetics, Prague. Primary cultures of chicken fibroblasts were prepared from embryos, and primary cultures of Leydig's ceils were prepared from testes of adult mice. All cells were grown at 37 ~ in 5% CO 2 air in Eagle's minimal essential medium supplemented with I0% fetal calf or bovine serum. Echinoeoccus granulosus hydatid cyst cells were obtained as previously reported (FIop, I e t al. 1982), Cell ghosts of adult crested newt erythrocytes (Triturus eristatus) were obtained as described (MONACO et al. 1982). Shedding of sea urchin spermatozoa (Strongylocentrotus purpuratus) was produced by injection of 0.1 M KC1. Dictyostelium discoideum amebae were grown at 22 ~ in H1-5 medium (WATTS and ASI-IWORTH 1970). Trichomonas vaginalis were grown axenically at 37 ~ in a CO 2 incubator in a Diamond medium without agar (DIAMOND 1957) supplemented with 10% (v/v) heat-inactivated bovine serum and 200 U/ml penicillin and 100 gg/ml streptomycin. Leishmania tropica were grown at 22 ~ in a medium containing 50% of 199 medium and 50% of DME supplemented with 10% of human erythrocytes lysate and i 000 U penicillin and 500 gg/ml streptomycin. Herpetomonas muscarum were grown at 37 ~ in DME supplemented with heat inactivated fetal calf serum and 100 U/ml penicillin and 50 gg/ml streptomycin.
2.2. Monoclonal Antibody
Mouse monoclonal antibody TU-01 (IgGl, kappa), whose prep- aration and partial characterization was described (V~KHCK'? et al. 1982), was precipitated from ascitic fluid by 50% ammonium sulfate. After dialysis against 10 mM Tris-HC1 (pH 7.8) buffer, the antibody was further purified on a DEAE cellulose (Whatman DE-52) column equilibrated in the same buffer. Proteins were eluted by linear gradient of NaC1 (20-500 mM NaC1 in I0 mM Tris-HCl, pH 7.8). For indirect immunofluorescence the antibody was purified only by ammonium sulfate precipitation.
2.3. Tubulin Preparation
Microtubule proteins were prepared from porcine brain by two cycles of polymerization-depolymerization according to Shelanski (SHELANSKI et al. 1973). Tubulin was separated from the microtubule associated proteins by phosphocellulose chromatography (WEINGARTEN et al. 1975).
2.4. Gel Electrophoresis
SDS-polyacrylamide gel electrophoresis (SDS-PAGE) was carried out as described (LAEMMLI 1970). The ct- and 13-subunits of tubulin
were effectively separated on 7.5% gels in the presence of SDS from Sigma (BEST et al. 1981) or alternatively with 8 M urea in the running gels. Whole cell lysates were separated on 10% gels. Protein concentration was determined according to (LOWRY el al. I951) with bovine serum albumin (Sigma) as standard.
2.5. Immunoblotting
Proteins from SDS-PAGE gels were electrophoretically transferred (TowmN et al. I979) onto nitrocellulose sheets (Synpore 6 or Millipore) for 3 hours at room temperature. To visualize protein bands, portions of the blots were stained with amido black. Unstained blots were incubated in 5% normal bovine serum (NBS) in phosphate buffered saline (PBS) plus 0.1% sodium azide overnight at 4~ to block remaining protein-binding sites. The blots were incubated for 1 hour at room temperature with 50gg/ml of the monoelonal antibody in 5% NBS and then extensively washed with 200raM NaC1 in 50raM Tris-HC1, pH7.4. The second antibody, affinity purified swine anti-mouse Ig labeled with horseradish peroxidase (O'SULLIVAN and MARKS I981) was added at a dilution 1 : 2,000 in 5% NBS without azide. After 1 hour incubation at room temperature the blots were washed and bound peroxidase was detected by 4-chloro-l-naphtol (Sigma) as a chromogen within 1-2 minutes (HAWKES 1982). For controls mouse monoclonal antibody directed against human transferrin (BARTEK et al. 1984) was used.
2.6. Indirect Immunofluorescence Microscopy
Mammalian and avian cells were grown on coverslips and extracted with 0.15% Triton X-100, fixed in glutaraldehyde as previously described (ScHLIWA 1981). MOLT 3 cells were attached to 0.02% polybrene (Sigma) coated coverslips during 1 hour incubation at 37~ T. vaginalis, L. tropiea, H. muscarum, D. discoideum, E. granulosus cells, cell ghosts of newt erythrocytes and sea urchin spermatozoa were fixed in 3 % formaldehyde-0.4 % Triton X- 100 in a cytoskeletal buffer (SMALL and CELLS 1978) for 30 minutes. Monoclonal antibody was added at concentration of 50 ~tg/ml in 2% BSA for 45 minutes. After rinsing in PBS, fluorescein isothiocyanate- conjugated swine anti-mouse (SEVAC, Prague) or goat anti-mouse (Dako) Ig antibodies was added for further 45 minutes. Finally, ceils were rinsed in PBS and mounted in 90% glycerol or Gelvatol (Monsanto Co., U.S.A.). In control experiments mouse monoclonal antibody against human transferrin (BARTEK 1984) was used. Cells were examined with a Leitz Orthoplan microscope or with a Leitz Dialux EB20 and photographed with FOMAPAN film or with Kodak TRI-X pan film.
3. Results
3.1. I m m u n o b l o t t i n g
Spec i f i c i ty o f t he m o n o c l o n a l a n t i b o d y to p o r c i n e b r a i n
t u b u l i n s u b u n i t s was i n v e s t i g a t e d b y i m m u n o b l o t t i n g .
P u r i f i e d p ig b r a i n t u b u l i n was s e p a r a t e d i n t o a- a n d [3-
s u b u n i t s o n S D S - P A G E s l ab gels a n d e l e c t r o p h o r e t -
ica l ly t r a n s f e r r e d o n t o n i t r o c e l l u l o s e shee ts . A s s h o w n
in Fig. 1 a b o t h s u b u n i t s we re e f f ic ien t ly t r a n s f e r r e d a n d
m o n o c l o n a l a n t i b o d y l a b e l e d t he a - s u b u n i t o f t u b u l i n .
T h e s a m e r e su l t s were o b t a i n e d u s i n g 8 M u r e a in the
r u n n i n g gels fo r t u b u l i n s u b u n i t s e p a r a t i o n . I f t he b l o t s
we re i n c u b a t e d in t he c h r o m o g e n s o l u t i o n fo r a l o n g

P. DIL~BER et al. : A Broad Spectrum Monoclonal Antibody to Alpha-Tubulin Does not Recognize all Protozoan Tubulins
Table 1. Reactivity o f the TU-01 antibody with tubulin from different species
203
Species Cell types Name of line Reactivity
IF IB
Human fibroblasts LEP +
T-lymphoblastoid line MOLT 3 +
Mouse fibroblasts L-A 9 +
Leydig's cells primary culture +
B-lymphoblastoid line P 3 X 63-Ag 8.653 +
embryonat carcinoma F 9 +
neuroblasts C 1300 ND
Hamster fibroblasts BHK +
Rat kangaroo fibroblasts PtK 2 +
Chicken fibroblasts primary culture +
Amphibia newt erythrocytes ( T. cristatus) + Echinodermata sea urchin spermatozoa (S. purpuratus) + Plathelmints Echinococcus granulosus + Slime molds Dictyostelium discoideum Ax 2 - -
Protozoa Triehomonas vaginalis + Herpetomonas muscarum Leishmania tropica
IF = immunofluorescence, IB = immunoblotting, ND = not determined, + = positive, - - = negative.
+ + + + + + +
ND
ND +
+
+
ND
+
time, very weak labeling of ~-subunit were observed (not shown). In control experiments no binding was detected. Immunoblotting was also used to determine antibody binding specificity in whole lysates. TU-01 antibody labeled only the tubulin band in whole lysates of MOLT 3 and LEP cell lines (Fig. 1 b). Even when 6.5% SDS-PAGE slab gels and overnight transfer onto nitrocellulose sheets resulting in better separation and transfer of high molecular proteins was used, the same results were obtained. The antibody interacted only with the tubulin band from several cell lines (Tab. 1) and pig brain homogenate (not shown). Tab. 1 shows the specificity of monoclonal antibody blotted with different microorganisms and with sea urchin sper- matozoa and newt erythrocytes. A positive reaction was visible with both whole extract and cytoskeleton preparation of T. vaginalis, with sea urchin sper- matozoa and with ghost of adult crested newt erythrocytes. No reaction was detected with D. dis~ coideum, H. muscarum, and L. tropica.
3.2. Indirect Irnrnunofluorescence
By using indirect immunofluorescence technique differ- ent cell types of different origins (mammals, amphibia, bird, echinodermata, plathelmints and protozoa) were stained with the monoclonal antibody against pig brain tubulin (Tab. 1). In well spread cultured mammalian
cells an abundant array of microtubules in interphase and in mitotic figures were seen (Figs. 2a and b). Microtubules networks originating from microtubule- organizing centres were also revelead in lymphoblast- oid cells (MOLT 3) (Fig. 2 c). In addition this antibody stains the tail of the sea urchin S. purpuratus sperm (Fig. 3 b) and in cell ghost of newt erythrocytes, the fluorescence was closely localized to the periphery of the cell where the marginal band is located (Fig. 3 a). The fluorescent pattern of Echinococcus hydatid cyst cells with curvilinear microtubules radiating from the nuclear region is in agreement with previous results obtained using monospecific antibodies against sheep brain tubulin (RuBINo et al. 1984). The staining pattern revealed by the monoclonal antibody is very typical in the flagellate T. vaginalis where only axostyle, mitotic spindle and the distal part of flagella are fluorescent (Fig. 3d); other antitubulin antibodies both mono- clonal and polyclonal (data not shown) stain complete- ly the flagellar axoneme. Regarding the other eukaryo- tic microorganisms considered in this study, no flu- orescence was evident with flagellate Herpetornonas and Leishmania and in amebas of the slime mould Dictyostelium the pattern of cytoplasmic microtubules originating from a nucleus associated centre previously demonstrated with other antibodies, was not seen (CAvPUCC~NELLI et al. 1981, 1982, RUBINO et al. 1984, and in press).
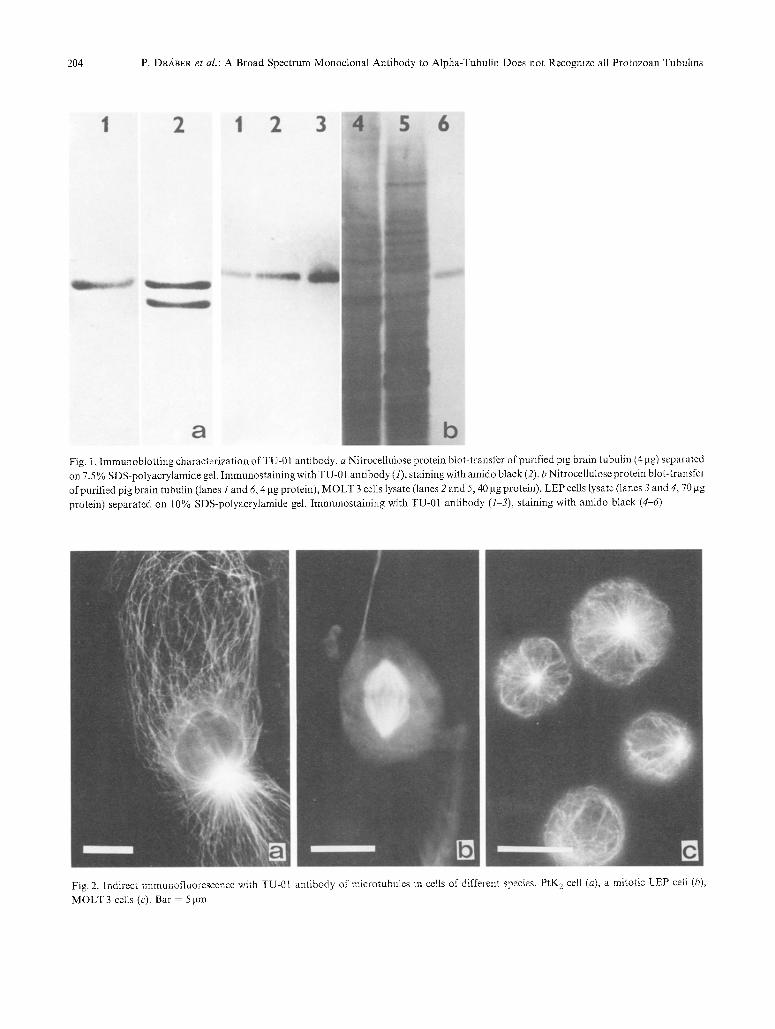
204 P. D~ABER et al.: A Broad Spectrum Monoclonal Antibody to Alpha-Tubulin Does not Recognize all Protozoan Tubulins
Fig. I. Immunobtot t ing characterization of TU-01 antibody, a Nitrocellulose protein blot-transfer of purified pig brain tubulin (4 lag) separated on 7.5 % SDS-polyacrylamide gel. Immunostaining with TU-01 antibody (1), staining with amido black (2). b Nitrocellulose protein blot-transfer of purified pig brain tubulin (lanes i and 6, 4 gg protein), MOLT 3 cells lysate (lanes 2 and 5, 40 lag protein), LEP cells lysate (lanes 3 and 4, 70 lag protein) separated on 10% SDS-polyacrylamide gel. Immunostaining with TU-01 antibody (1-3), staining with amido black (4-6)
Fig. 2. Indirect immunofluorescence with TU-01 antibody of microtubules in cells of different species. PtK 2 cell (a), a mitotic LEP celt (b),
MOLT 3 cells (c). Bar - 5 >m
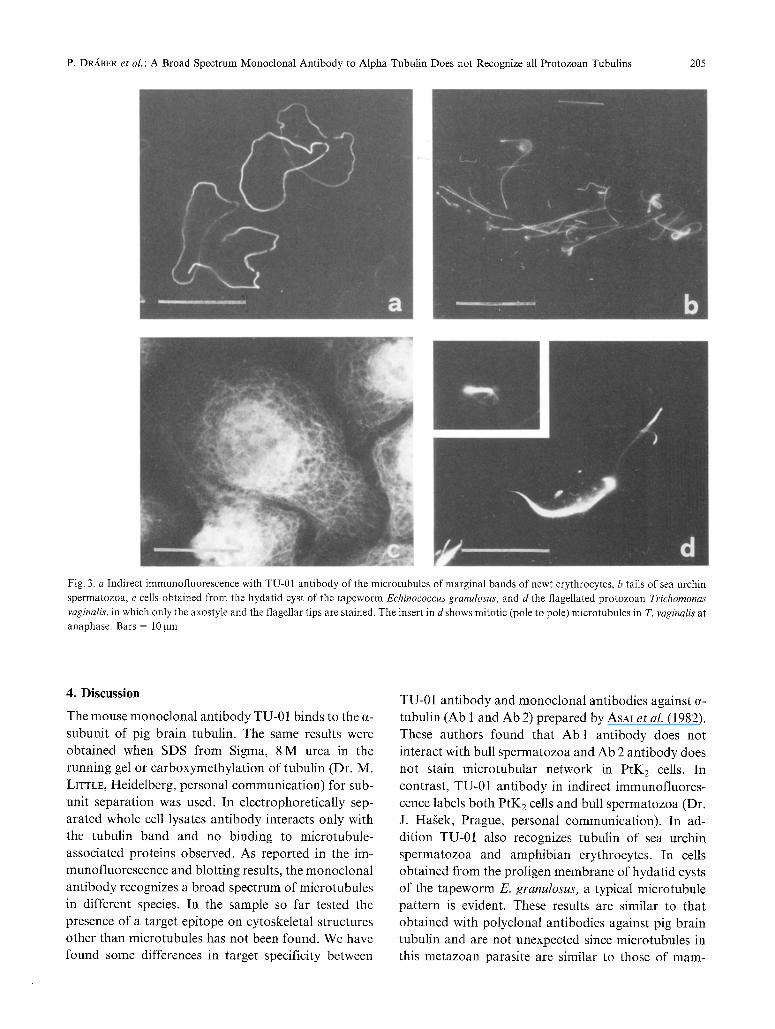
P. DRABER et al. : A Broad Spectrum Monoclonal Antibody to Alpha-Tubulin Does not Recognize all Protozoan Tubulins 205
Fig. 3. a Indirect immunofluorescence with TU-01 antibody of the microtubules of marginal bands of newt erythrocytes, b tails of sea urchin spermatozoa, c cells obtained from the hydatid cyst of the tapeworm Echinococcus granulosus, and d the flagellated protozoan Trichornonas vaginalis, in which only the axostyle and the flagellar tips are stained. The insert in d shows mitotic (pole to pole) microtubules in T. vaginalis at anaphase. Bars = 10gm
4. Discussion
The mouse monoclonal antibody TU-01 binds to the ~-
subunit of pig brain tubulin. The same results were obtained when SDS from Sigma, 8 M urea in the
running gel or carboxymethylation of tubulin (Dr. M. LITTLE, Heidelberg, personal communication) for sub-
unit separation was used. In electrophoretically sep- arated whole cell lysates antibody interacts only with
the tubulin band and no binding to microtubule- associated proteins observed. As reported in the im-
munofluorescence and blotting results, the monoclonal antibody recognizes a broad spectrum of microtubules in different species. In the sample so far tested the
presence of a target epitope on cytoskeletal structures other than microtubules has not been found. We have found some differences in target specificity between
TU-01 antibody and monoclonal antibodies against ~- tubulin (Ab 1 and Ab 2) prepared by ASAI et al. (1982).
These authors found that Ab 1 antibody does not
interact with bull spermatozoa and Ab 2 antibody does
not stain microtubular network in PtK 2 cells. In
contrast, TU-01 antibody in indirect immunofluores- cence labels both PtK 2 cells and bull spermatozoa (Dr.
J. Hagek, Prague, personal communication). In ad- dition TU-01 also recognizes tubulin of sea urchin
spermatozoa and amphibian erythrocytes. In cells obtained from the proligen membrane of hydatid cysts of the tapeworm E. granulosus, a typical microtubule pattern is evident. These results are similar to that
obtained with polyclonal antibodies against pig brain tubulin and are not unexpected since microtubules in this metazoan parasite are similar to those of mam-

206 P. DRA.BER et al.: A Broad Spectrum Monoclonal Antibody to Alpha-Tubulin Does not Recognize ai1 Protozoan Tubulins
malian cells (RuBINo et al. 1983). It seems evident that the TU-01 antibody recognizes an epitope of tubulin that is well represented in higher eukaryotes. This antigenic determinant is not expressed in some eu- karyotic microorganisms (i.e., flagellates such as Herpetomonas and Leishmania and the slime mould Dictyostelium) and is present in only some forms of tubulin in Trichomonas.
Although tubulin is an extremely conserved protein (KALFAYAN and WENSlNK 1981, LUDUENA and WOODWARD 1973, S~LELOW and ROSEMBAUM 1981, VALENZUELA et al. 1981) and it seems certain that a- and ~-tubulin arose from the same ancestral gene (LITTLE et al. 1982), several data on the heterogeneity of tubulin within different cell species and also within the same cell are accumulating. In particular, heterogeneity has been demonstrated in lower eukaryotes and STEPIqENS (1978) has showed that the tubulin subunits of cilia, flagella and cytoplasm have some nonidentical peptides in- dicating differences in primary structure. Many other cell types including eukaryotic microorganisms contain multiple tubulin genes (CLEVELAND 1983, FULTON and LAI 1982, KALF+YAN and WENSINK 1981, RUBINO et al. 1984, SANCHEZ et al. 1980) and multiple forms of tubulin have been recently identified in neurons (GozEs and SWEADNER 1981), Polytornella (McKEITHAN and ROSEMBAUM 1981, McKEITHAN et al. 1983) and Naeg- leria (FULTON and LAX 1982). GALLO and ANDERTON (1983) have revealed subpopulation of microtubules in the protozoan Trypanosoma brucei using monoclonal antibody against bovine brain tubulin. The existence of an immunologically unrelated tubulin in different eu- karyotic microorganisms (Leishmania, Herpetomonas, Euplotes, Trichomonas, Naegleria) has been shown using monoclonal and polyclonal antibodies against tubulins (manuscript in preparation). At the moment data on the number of tubulin genes and the chemical complexity of the tubulin system of Trichomonas are not available. Our results showing that TU-01 antibody recognizes only determinants present in microtubules of the axostyle, mitotic spindle and the distal part of the flagellum indicate that in T. vaginalis at least two different tubulin isotypes are present. The fact that TU-01 do not visualize the proximal flagellar micro- tubules could be explained if T. vaginalis utilizes some cytoplasmic tubulin for the assembly ofmicrotubules at the flagellar tip. However, since the specificity of antibody is not absolute the two tubulin isotypes shown in Trichomonas flagellum could still be different from the cytoplasmic tubulin.
In conclusion the monoclonal antibody against pig
brain tubulin recognizes a broad spectrum of micro- tubules in different species and in Trichomonas reveals multiple forms of tubulins. The determination of the exact isotype specificity of the TU-01 antibody is required for further study of tubulin heterogeneity.
Acknowledgements
Thanks are due to Dr. CHANDLER FULTON for criticism in prep- aration of the manuscript. Partially supported by grants from the Italian Research Council (C.N.R.), research contract no. 83.0063.52, and the Ministry of Education (M.P.I.).
References
ASAI, D. J., BROKAW, C. J., THOMPSON, W. C., WILSON, L., 1982: Two different monoclonal antibodies to alpha tubulin inhibit the bending of reactivated sea urchin spermatozoa. Cell Motility 2, 599-614.
BARTEK, J., VIKLICK'~(, V., HOI~EJgi, V., VERLOVA, H., DRABER, P., 1984: Production of monoclonal antibodies against human transferrin. Fol. Biol. (Prague) 30, 137-140.
BEST, D., WARR, P. J., GULL, K., 1981: Influence of the composition of commercial sodium dodecyl sulfate preparations on the separation of u and 13 tubulin during polyacrylamide gel electro- phoresis. Anal. Biochem. 114, 281-284.
BIBRING, J., BAXANDALL, J., DENSLOW, J., WALKER, B., 1976: Heterogeneity of the subunit of tubulin and the variability of tubulin within a single organism. J. Cell Biol. 69, 301-312.
CAPPUCCINELLI, P., UNGER, E., RUBINO, S., 1981: Immunofluores- cence of microtubular structures during the cell cycle of Dic- tyostelium discoideum. J. gen. Microbiol. 124, 207-21 I.
- - RUBINO, S., FIGHETTI, M., UNGER, E., 1982: Organization of the microtubular system in Dictyostelium amoebae. In: Microtubules in microorganisms (CAPPUCCINELLI, P., MORRIS, N. R.~ eds.), p. 71. New York: Marcel Dekker.
CLAYTON, L., QUINLAN, R. A., ROOBOL, A., POGSON, C. I., GULL, K., 1980: A comparison of tubulin from mammalian brain and Physarum polycephalum using SDS-polyacrylamide gel electro- phoresis and peptide mapping. FEBS Letters 115, 301-305.
CLEVELAND, D. W., LOPATA, M. A., MACDONALD, R. J., COWAN, N. J., RUTTER, W. J., KIRSCHNER, M. W., i980: Number and evolutionary conservation of the c~- and [3-tubulin and cytoplasmic 13- and y-actin genes using specific cloned cDNA probes. Cell 20, 95-105.
- - 1983: The tubulins: from DNA to RNA to protein and back again. Cell 34, 330-332.
DIAMOND, L. S., 1957: The establishment of various trichomonads of animal and man in axenic cultures. J. Parasitol. 43, 488~490.
DUSTIN, P., 1978: Microtubules. Berlin-Heidelberg-New York: Springer.
FIORI, L.,RuBINO, S., LUmNU, G., PUGLIESE, A., CAPPUCCINELLI, P., 1982: In vitro susceptibility of cell cultures from Echinococcus granulosus to antimicrotubular drugs. In: Current chemotherapy and immunotherapy (PERTITI, P., GIALDRONI-GRASSl, G., eds.), p. 1402. Washington: American Society for Microbiology.
FULTON, C., SIMPSON, P. A., 1976: Selective synthesis and utilization of flagellar tubulin. The multi-tubulin hypothesis. In: Cell motility (GOLDMAN, R., POLLARD, T., ROSEMBAUM, J., eds.), p. 987. New York: Cold Spring Harbor Laboratory.

P. DR~_BEe, et al.: A Broad Spectrum Monoclonal Antibody to Alpha-Tubulin Does not Recognize all Protozoan Tubulins 207
FULTON, C., LAI, E. Y., 1982: Questions about tubulin synthesis in Naegleria. In: Microtubules in microorganisms (CAPPUCCINELLI, P., MORRIS, N. R., eds.), p. 235. New York: Marcel Dekker.
GALLO, J. M., ANDERTON, B. H., 1983: A subpopulation of trypanosome microtubules recognized by a monoclonal antibody to tubulin. E.M.B.O.J. 2. 479-483.
GOZES, I., SWEADNER, K. J., 198t: Multiple tubulin forms are expressed by a single neurone. Nature 294, 477-479.
- - BARNSTABLE, C., 1982: Monoclonal antibodies that recognize discrete forms of tubulin. Proc. Nat. Acad. Sci. U.S.A. 79, 2579- 2583.
HAWKES, R., 1982: Identification ofconcanavalin A-binding proteins after sodium dodecyl sulfate-gel electrophoresis and protein blotting. Anal. Biochem. 123, 143-146.
KALFAYAN, L., WENSINK, P. C., 1981: ct-tubulin genes of Drosophila. Cell 24, 97-106.
KILMARTIN, J. V., WRIGHT, B., MILSTEIN, C., 1982: Rat monoclonal antitubulin antibodies derived by using a new nonsecreting rat cell line. J. Cell Biol. 93, 576-582.
KRAUHS, E., LITTLE, M., KEMPF, T., HOFER-WARB1NEKWADE, R., PONST1NGL, H., 1981 : Complete amino acid sequence of 13 tubulin from porcine brain. Proc. Nat. Acad. Sci. U.S.A. 78, 4156-4160.
LAEMMLI, U. K., 19/0: Cleavage of structural proteins during the assembly of the head of bacteriophage 14. Nature 227, 680-685.
LITTLE, M., LUDUENA, R. F., LANGFORD, G. M., ASNES, C. F., FARRELL, K., 1981 : Comparison of proteolytic cleavage patterns of ct-tubulins and 13-tubulins from taxonomically distant species. J. Mol. Biol. 149, 95-107.
- - KRAUHS, E., PONSTINGL, H., LUDUENA, R., RICE, N., 1982: Tubulin subunits from higher and lower eukaryotes. A structural comparison. In: Microtubules in microorganisms (CAPPUCCINELLI, P., MORRIS, N. R., eds.), p. 1. New York: Marcel Dekker.
LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L., RANDALL, R. J., 1951: Protein measurement with the Folin phenol reagent. J. biol. Chem. 193, 265275.
LUDUENA, R. F., WOODWARD, D. O., I973: Isolation and partial characterization of a and 13 tubulin from outer doublets of sea urchin sperm and microtubules of chick embryo brain. Proc. Nat. Acad. Sci. U.S.A. 70, 3594-3598.
MCKEITHAN, T. Wi, ROSEMBAUM, J. L., 1981: Multiple forms of tubulin in the cytoskeletal and flagellar microtubules of Polytomella. J. Cell Biol. 91, 352-360.
- - LEFEBVRE, P. A., SILFLOW, C. D., ROSEMSAUM, J. L., 1983: Multiple forms of tubulin in Polytornella and Chlamydomonas: evidence for a precursor of flagellar ct-tubulin. J. Cell Biol. 96, I056-1063.
MONACO, G., SALUSTm, A., BERTOLINI, B., 1982: Observations on the molecular components stabilizing the microtubular system of the marginal band in the newt erythrocyte. J. Cell Sci. 58, 149 163.
O'SULLIVAN, M. J., MARKS, V., 1981: Methods for the preparation of enzyme antibody conjugates for use in enzyme immunoassay. In: Methods in enzymology, Vol. 73 (LANGONE, J. J., VAN VUNAKIS, H.. eds.), p. 147. New York: Academic Press.
PONSTINGL, H., KRAUHS, E., LITTLE, M., KEMPF, T., 1981: Complete amino acid sequence of a tubulin from porcine brain. Proc. Nat. Acad. Sci. U.S.A. 78, 2757-2761.
RUBINO, S., FIOR1, P. L., LUBINU, G., MONACO, G., CAPPUCCINELLI, P., 1983: The cytoskeleton of hydatid cyst cultured cells and its sensitivity to inhibitors. Eur. J. Cell Biol. 30, I82-190.
- - FIGHETTI, M., UNOER, E., CAPPUCCINELLI, P., 1984: Location of actin, myosin, and microtubular structures during directed locomotion of Dietyostelium amebae. J. Cell Biol. 98, 382-390.
- - SMALL, V. J., CLAVIETZ, M., SELLITTO, C., CAPPUCCINELLI, P.: Actin filament meshworks and microtubules in Dictyostelium discoideum. In: Advances in contractile proteins (ALIA, E. E., ARENA, N., RuSSO, M., eds.). New York: Praeger Publ. Co., in press.
SANCHEZ, F., NATZLE, J. E., CLEVELAND, D. W., KIRSCHNER, M. W., MCCARTHY, B. J., 1980: A dispersed multigene family encoding tubulin in Drosophila melanogaster. Cell 20, 845-854.
SCHLIWA, M., EUTENEUR, U., BULINSK1, J. C., IZANT, J. G., 1981: Calcium lability of cytoplasmic microtubules and its modulation by microtubule associated proteins. Proc. Nat. Acad. Sci. U.S.A. 78, 1037-104I.
SHELANSKI, M. L., GASKIN, F., CANTOR, C. R., 1973: Microtubule assembly in the absence of added nucleotide. Proc. Nat. Acad. Sci. U.S.A. 70, 765-768.
SILFLOW, C. D., ROSEMBAUM, J. L., 1981: Multiple u and 13 tubulin genes in Chlamydomonas and regulation of tubulin mRNA level after defiagellation. Cell 24, 81-88.
SMALL, J. V., CELLS, J. E., 1978: Filament arrangements in negatively stained cultured cells: the organisation of actin. Cytobiologie 16, 308-325.
STEPHENS, R. E., 1978: Primary structural differences among tubulin subunits from flagella, cilia and the cytoplasm. Biochemistry 17, 2882-2891.
TOWBIN, H. T., STAEHEL1N, T., GORDON, J., 1979: Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedures and some application. Proc. Nat. Acad. Sci. U.S.A. 74, 4350-4354.
UNGER, E., RUBINO, S., WEINERT, T., CAPPUCCINELLI, P., 1979: Immunofluorescence of the tubulin system in cellular slime moulds. F.E.M.S. Microbiol. Lett. 6, 317-320.
VALENZUELA, P., QUIROGA, M., ZALDIVAR, J., RUTTER, W. 1., KIRSCHNER, M. W., CLEVELAND, D. W., 1981: Nucleotide and corresponding amino acid sequences encoded by u and 13 tubulin mRNA. Nature 289, 650-655.
VIKLICK?, V., DRABER, P., HAgEK, J., B~RTEK, J., 1982: Production and characterization of a monoclonal antitubulin antibody. Cell Biol. int. Rep. 6, 726-731.
WATTS, D. J., ASHWORTH, J. M., 1970: Growth ofmyxamoebae of the cellular slime mould Dictyostelium discoideum in axenic culture. Biochem. J. 119, 171-174.
WEINGARTEN, M. D., LOCKWOOD, A. H., Hwo, S. Y., KIRSCHNER, M. W., 1975: A protein factor essential for microtubules as- sembly. Proc. Nat. Acad. Sci. U.S.A. 72, 1858-1862.
WHITE, E., TOLBERT, E. M., KATZ, E. R., 1983: Identification of tubulin in Dictyostelium discoideum: Characterization of some unique properties. J. Cell Biol. 97, 1015-1019.




















