
IgA-associated Inhibition of Polymorphonuclear Leukocyte Chemotaxis in Neutrophilic Dermatoses
Upload
independentCategory
view
3download
0
Identification of Distinct Glycoforms of IgA1 inPlasma from Patients with Immunoglobulin A(IgA) Nephropathy and Healthy Individuals*Sylvain Lehoux‡, Rongjuan Mi‡‡‡, Rajindra P. Aryal‡§§, Yingchun Wang‡,Katrine T.-B. G. Schjoldager§, Henrik Clausen§, Irma van Die¶, Yoosun Han�,Arlene B. Chapman�, Richard D. Cummings‡, and Tongzhong Ju‡**
Immunoglobulin A nephropathy (IgAN) is the most com-mon form of glomerulonephritis worldwide and is histo-logically characterized by the deposition of IgA1 and con-sequent inflammation in the glomerular mesangium. Priorstudies suggested that serum IgA1 from IgAN patientscontains aberrant, undergalactosylated O-glycans, for ex-ample, Tn antigen and its sialylated version, SialylTn(STn), but the mechanisms underlying aberrant O-glyco-sylation are not well understood. Here we have used seriallectin separation technologies, Western blot, enzymaticmodifications, and mass spectrometry to explore whetherthere are different glycoforms of IgA1 in plasma frompatients with IgAN and healthy individuals. Although totalplasma IgA in IgAN patients was elevated �1.6-fold com-pared with that in healthy donors, IgA1 in all samples wasunexpectedly separable into two distinct glycoforms: onewith core 1 based O-glycans, and the other exclusivelycontaining Tn/STn structures. Importantly, Tn antigenpresent on IgA1 from IgAN patients and controls wasconvertible into the core 1 structure in vitro by recombi-nant T-synthase. Our results demonstrate that under-galactosylation of O-glycans in IgA1 is not restricted toIgAN and suggest that in vivo inefficiency of T-synthasetoward IgA1 in a subpopulation of B or plasma cells, aswell as overall elevation of IgA, may contribute to IgANpathogenesis. Molecular & Cellular Proteomics 13:10.1074/mcp.M114.039693, 3097–3113, 2014.
Immunoglobulin A (IgA) nephropathy (IgAN)1, also calledBerger’s Disease, was first described by Jean Berger in 1968
(1). More than four decades later, IgAN is the most commonform of primary glomerulonephritis worldwide and leads toterminal renal failure in 20–40% of patients over 20–25 years.The majority of primary IgAN cases are sporadic, and only aminority of patients appear within family clusters, yet, noheritable gene associated with the disease has been identified(2). Histologically, IgAN is characterized by deposition of IgA1and inflammatory lesions in the glomeruli. In contrast to IgA2,human IgA1 contains an extra 13 amino acids in its hinge region(HR) to form a �20 amino acid domain characteristically rich inSer/Thr/Pro residues (3). Six of the 9 Ser/Thr residues are usu-ally modified by the mono- and di-sialylated core 1 structure orT antigen [Neu5Ac�2–3Gal�1–3(�Neu5Ac�2–6)GalNAc�-Ser/Thr] (3). Many studies (4–8) have suggested that undergalacto-sylated O-glycans, that is, Tn antigen (GalNAc�-Ser/Thr) and itssialylated version, SialylTn (STn, Neu5Ac�2–6GalNAc�-Ser/Thr), are enriched in the HR of IgA1 from patients with IgAN incomparison to IgA1 from normal individuals, and might be re-sponsible for the pathogenesis of IgAN. However, the mecha-nism(s) underlying the undergalactosylation of IgA1 from pa-tients with IgAN is unclear.
Mucin type O-glycosylation (O-glycan) is a common proteinpost-translational modification of Ser/Thr residues of secretedand transmembrane glycoproteins and can regulate manyaspects of their functions and recognition properties (9–14).Within human immunoglobulins only IgA1 and IgD are O-glycosylated in their HR domains (15, 16). The biosynthesis ofO-glycans mainly takes place in the Golgi apparatus by serialreactions of a group of glycosyltransferases. In humans, poly-peptide-GalNAc-transferases (ppGalNAcTs) encoded by atleast 20 ppGalNAcTs, initiate O-glycosylation in the Golgi (17,18), and possibly in the ER (19, 20), by transferring a GalNAcfrom UDP-GalNAc to Ser or Thr residues in proteins passingthrough the secretory pathway to form GalNAc�-Ser/Thr (Tnantigen) (Fig. 1). For the generation of the Tn antigen on IgA1ppGalNAcT2 may be the key enzyme responsible (21). Typi-cally, the Tn antigen is subsequently efficiently converted to
From the Departments of ‡Biochemistry and �Medicine, EmoryUniversity School of Medicine, Atlanta, GA 30322; §Department ofCellular and Molecular Medicine, Copenhagen Center for Glycom-ics, Copenhagen, Denmark; ¶Department of Molecular Cell Biology& Immunology, VU University Medical Center, Amsterdam, TheNetherlands.
Received March 19, 2014, and in revised form, July 7, 2014Published, MCP Papers in Press, July 28, 2014, DOI 10.1074/
mcp.M114.039693Author contributions: S.L., R.M., Y.W., R.D.C., and T.J. designed
research; S.L. and R.M. performed research; R.P.A., K.T.S., H.C., I.V.,Y.H., and A.B.C. contributed new reagents or analytic tools; S.L.,R.D.C., and T.J. analyzed data; S.L., R.D.C., and T.J. wrote the paper.
1 The abbreviations used are: IgAN, IgA Nephropathy; HPA, Helixpomatia agglutinin; HR, hinge region; Neu, neuraminidase; PNA, pea-
nut agglutinin; ppGalNAcTs, polypeptide �-GalNAc-transferases;STn, SialylTn.
Research© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.This paper is available on line at http://www.mcponline.org
Molecular & Cellular Proteomics 13.11 3097

the core 1 structure Gal�1–3GalNAc�-Ser/Thr by a singleenzyme, the core 1 �3galactosyltransferase (T-synthase) (22)which utilizes UDP-Gal as the donor and GalNAc�-Ser/Thr inproteins as the acceptor. Moreover, the T antigen is usuallyfurther modified, branched and/or elongated to form complexnormal O-glycans, including sialyl Core 1 (or sialyl-T), Core 2O-glycans and extended Core 1 O-glycans in glycoproteins ofhematopoietic cells (11, 23). Thus, the Tn antigen does notnormally occur in significant amounts in cellular glycoproteins(Fig. 1). Interestingly, biosynthesis of active T-synthase re-quires an ER-localized molecular chaperone Cosmc to pre-vent its aggregation and subsequent proteasomal degrada-tion (24–26). Defects in Cosmc on Xq24 because of somaticmutations (27), gene deletion (28), or epigenetic silencing ofits promoter region (29) result in an inactive T-synthase andconsequent expression of the Tn and STn antigens on glyco-proteins. Such altered O-glycosylation is associated with thepathology of several human diseases such as Tn syndrome(30, 31), in which patients have a sporadic acquired mutationin X-linked Cosmc in hematopoietic precursors, and in neo-plastic transformations (32, 33). However, in the case of IgANno mutation in either Cosmc/T-synthase or other glycosyl-transferases has been identified, although there are conflict-ing studies suggesting that compromised transcription ofCosmc and/or T-synthase, up-regulated expression ofST6GalNAcII, or decreased T-synthase activity in the pe-ripheral blood B cells from patients with IgAN may contrib-ute to aberrant expression of Tn and STn antigens in IgA1HR (8, 34–37).
Although previous studies suggested the expression of Tn/STn antigens in IgA1 of patients with IgAN might correlatewith the disease, we questioned whether plasma IgA1 might
occur in distinct glycoforms and whether differential expres-sion of these glycoforms might correlate with disease. Herewe report the unexpected discovery that human plasmaIgA1 can be separated into distinct glycoforms by seriallectin affinity chromatography: one glycoform contains nor-mal mono/di-sialylated T antigen whereas the other containsexclusive Tn/STn antigens, which was further confirmed by invitro conversion into T antigen by recombinant human T-syn-thase and mass spectrometry analysis. We further exploredthe basis for formation of these distinct glycoforms using celllines and enzymatic modifications. The identification of thedistinct glycoform of plasma IgA1 carrying Tn/STn antigensoffers a new direction for future studies aimed at identifying itspotential contribution to IgAN.
EXPERIMENTAL PROCEDURES
Plasma Samples and Cell Culture—Blood samples from bothbiopsy-proven patients with IgAN and healthy controls were ob-tained from the Emory Clinic under the approved IRB protocol(IRB00008410). Information from all donors is reported in Table I. Theplasma, erythrocytes and leukocytes were separated using Lym-phoprep (StemcellTM Technologies, Vancouver, Canada) following themanufacturer’s protocol. All plasma samples were aliquotted andstored at �80 °C, or �20 °C during experiments. Dakiki cells (ATCC,TIB-206) and Tn4 cells (38) were grown in RPMI1640 (Invitrogen,Carlsbad, CA) containing 20% heat-inactivated fetal bovine serum at37 °C, 5% CO2.
ELISA Assays—Flat-bottomed 96-well ELISA Microplates (Greinerbio-one, Frickenhausen, Germany) were coated overnight at 4 °C with50 �l of 1 �g/ml F(Ab)’2 fragment of goat IgG anti-human IgA (JacksonImmuno-Research Labs, West Grove, PA) in 0.05 M carbonate/bicar-bonate pH 9.6 buffer. Coated plates were blocked with 1% BSA inphosphate-buffered saline (PBS)-0.05%, Tween 20 (PBS-T), 1h atroom temperature (RT) and then washed three times with PBS-T.Plasma samples diluted 1:10,000 in PBS-T were incubated in dupli-
FIG. 1. The O-glycosylation of proteins in hematopoietic cells: Mucin-type O-glycosylation of proteins initiates mainly in the Golgi whenpolypeptide �-GalNAc-transferases (ppGalNAcTs) transfer GalNAc from UDP-Gal to Ser/Thr on a protein going through the secretory pathwayto form Tn antigen (GalNAc�-Ser/Thr). Tn antigen is normally further modified by the T-synthase (Core 1 �3GalT) which transfers Gal fromUDP-Gal to GalNAc�-Ser/Thr in a protein to form core 1 (T antigen) in all cell types. Cosmc is the specific molecular chaperone for theT-synthase. In all cell types, the T antigen is usually sialylated by ST3Gal-I, which transfers the N-acetylneuraminic acid (sialic acid) fromCMP-Neu5Ac to form �3sialyl Core 1 (Neu5Ac�2,3Gal�1,3GalNAc�-Ser/Thr), which can be further sialylated by ST6GalNAc-II to form di-sialylCore 1 [Neu5Ac�2,3Gal�1,3(Neu5Ac�2,6)GalNAc�-Ser/Thr] structures. In hematopoietic cells, besides synthesis of sialyl core 1 structures, theT antigen is usually further modified by core 2 �6-N-acetylglucosaminyltransferase (C2GnT) to form Core 2 [Gal�1,3(GlcNAc�1,6)GalNAc�-Ser/Thr] based O-glycans. Mutations in Cosmc or loss of T-synthase activity results in pathological expression of Tn and/or STn antigens. TheSTn antigen (Neu5Ac�2,6GalNAc�-Ser/Thr) results from the actions of ST6GalNAc-I, which transfers Neu5Ac from CMP-Neu5Ac to the Tnantigen. Because of poor efficiency of ST6GalNAc-I, it is likely that only very high expression of ST6GalNAc-I could outcompete functionalT-synthase to result in pathologic STn expression in the absence of corresponding Tn expression.
O-Glycans of Human IgA1
3098 Molecular & Cellular Proteomics 13.11

cate for 1h at RT, and washed three times with PBS-T. The wells wereincubated with peroxidase labeled goat anti-human IgA (�) (KPL,Gaithersburg, MD) at 0.25 �g/ml in PBS-T solution 1h at RT, and thenwashed 4 times with PBS-T. The signal was revealed with o-phe-nylenediamine dihydrochloride (Thermo Scientific, Waltham, MA) ac-cording to manufacturer’s instructions. Serial dilutions of commercialhuman IgA (InvivoGen, San Diego, CA) were used as a standard forquantification. All sample measurements were done twice.
Pull Down Assays—Peanut agglutinin (PNA)-bound Agarose beadswere purchased from Vector Labs (Burlingame, CA); Helix pomatiaagglutinin (HPA)-beads were generated by coupling free HPA lectin(Sigma Aldrich, St Louis, MO) to Affi-gel 10 beads (Bio-Rad, Hercules,CA) in the presence of GalNAc. Briefly, HPA was resuspended (2mg/ml) into a 50 mM GalNAc solution and 500 �l of this solution wasincubated (1h at RT, in rotation) with 1 ml Affi-gel 10 beads firstwashed with 10 ml of ice cold water. The beads were then pulleddown by centrifugation (1000 rpm, 3min), the supernatant removed,and the beads were incubated 1h with 1 ml of 1 M Glycine at RT, inrotation. The Glycine solution was removed and the beads werewashed 3 times with 10 ml of PBS, and finally resuspended into 2 mlof PBS and stored at 4 °C.
Plasma samples were diluted 1:2 with PBS and then treated or notwith neuraminidase (from Clostridium perfringens, Roche, Germany)(1 �l of neuraminidase for 10 �l of plasma) overnight at 37 °C. PNAand HPA pull downs were performed at RT, 2h, in rotation. Aftercentrifugation (1000 rpm, 3min) the supernatants were collected anddesignated as Unbound fraction (UB). Following two washes withPBS, the beads were eluted by incubation (15min, RT, in rotation) withan equal volume of the UB fraction of 0.2 M GalNAc for the HPA pulldown or 0.2 M Lactose for PNA pull down. The eluted fraction, termedthe Bound fraction (B), was collected after centrifugation at 1000 rpmfor 3min.
The Anti-Human IgA (�-chain specific)-Agarose beads were pur-chased from Sigma-Aldrich. Total IgA pull down was performed at4 °C overnight in rotation. After centrifugation (1000 rpm, 3min) thebeads were transferred into a 5 ml Poly-Prep Chromatography Col-umn (Bio-Rad) and washed 3 times with PBS. Beads were eluted 7times with 100 �l of 0.2 M Glycine pH 2.6. All fractions were pooled,washed and concentrated in water using a centricon unit (Millipore,Billerica, MA), to a final volume of 100 �l. The IgA concentration ofeach sample was determined by Pierce® BCA Protein Assay Kit(Thermo Scientific).
IgA1-Hinge Region Specific-Tn Antibody Western Blot—Mouseanti-human IgA1 (�1 chain specific) FITC-conjugated and mouseanti-human IgA2 (�2 chain specific) FITC-conjugated were purchasedfrom (Southern Biotech, Birmingham, AL). Rabbit anti-FITC HRP-conjugated antibody was purchased from Dako (Denmark). The goatanti-human IgA (� chain specific) HRP-conjugated antibody was pur-chased from KPL. PNA HRP-conjugated and HPA HRP-conjugatedwere purchased from Sigma Aldrich. Goat anti-mouse HRP-conju-gated antibody was purchased from KPL.
The 6E5–7Tn antibody was produced as follow. A 20-mer peptidecovering the Ser/Thr rich hinge region of IgA1 was synthesized bySchafer-N (Denmark) and in vitro glycosylated using recombinanthuman ppGalNAcTs GalNAc-T1 and GalNAc-T2 (39) adding in total 7
mols of GalNAc per peptide (-VPSTPPTPSPSTPPTPSPS-). The gly-cosylation reactions were monitored by MALDI-TOF mass spectrom-etry. The IgA1-HR-7Tn glycopeptide was conjugated to KLH (Pierce)using gluteraldehyde as previously described (40, 41), and conjuga-tion efficiency was evaluated using comparative titration analysis ofthe KLH-conjugated peptide versus the corresponding non-conju-gated glycopeptide in an ELISA. Female Balb/c mice were injectedsubcutaneously with 20 �g KLH-conjugated glycopeptide in a totalvolume of 200 �l (1:1 mix with Freund’s complete (first immunizationonly) and incomplete adjuvant (Sigma) for subsequent injections)under the Danish Animal Experiments Inspectorate approved proto-col. Mice received 3 immunizations 14 days apart and blood wascollected from the orbital sinus 1 week following the third immuniza-tion. After fusion of splenocytes from the immunized mouse with NS-1myeloma cells, the hybridoma clones were screened based on pos-itive reactivity to the glycopeptide immunogen and negative reactivityto the non-glycosylated peptide and various other control Tn-pep-tides in a direct ELISA. The monoclonal mouse IgG1 specificallyreacting to the IgA-HR-7Tn glycopeptide, designated as 6E5–7Tn,was generated, and the purified 6E5–7Tn (from hybridoma mediausing Protein-G column) was used for the experiments.
For all lectin (HPA and PNA) and Western (IgA, IgA1, IgA2, and6E5–7Tn) blots, samples were boiled for 5 min in reducing conditionsand run into 4–20% Mini-PROTEAN® TGXTM precast gels (Bio-Rad,Richmond, VA) and then dry-transferred onto a nitrocellulose mem-brane using the iBlot® Blotting System and the iBlot® Gel TransferStacks, Mini Nitrocellulose (Invitrogen).
For the PNA, HPA, and IgA blots, the membranes were blocked 1hat RT with a Tris-Buffered Saline (TBS) solution containing 0.1%Tween 20 and 5% (w/v) BSA and then incubated with HPA- orPNA-HRP 1 �g/ml in TBS, 0.1% Tween 20, 5% (w/v) BSA for 1h atRT, or incubated with goat anti-human IgA HRP-conjugated antibodyat 0.2 �g/ml in TBS for 10 min at RT. The membranes were finallywashed four times with TBS, 0.1% Tween 20.
For the IgA1, IgA2, and 6E5–7Tn blots, the membranes wereblocked 1h at RT with TBS solution containing 0.1% Tween 20 and5% (w/v) non-fat dried milk and incubated with mouse anti-humanIgA1 or IgA2 FITC-conjugated at 1 �g/ml or with mouse 6E5–7Tnantibody at 1 �g/ml in TBS, 0.1% Tween 20, 5% (w/v) milk overnightat 4 °C. The membranes were next washed four times with TBS, 0.1%Tween 20 followed by incubation with rabbit anti-FITC HRP-conju-gated for IgA1 or IgA2 at 1 �g/ml or with goat anti-mouse HRP-conjugated antibody at 1 �g/ml in TBS, 0.1% Tween 20, 5% (w/v)non-fat dried milk 1h at RT. The membranes were finally washed fourtimes with TBS, 0.1% Tween 20. For all membranes, the detection ofsignal was achieved using enhanced chemiluminescence substrate ofHRP (SuperSignal® West Pico Chemiluminescent Substrate, ThermoScientific).
Quantification of Lectin and Statistical Analysis on Western BlotData—Lectin and Western blot films were scanned using a Cannonscanner and band intensities were quantified using an Alphatechsystem. The intensity of each band was measured and subtractedfrom the background value. Statistical differences were analyzedusing an unpaired Student’s t test. p value � 0.05 were consideredsignificant.
Conversion of Tn Antigen in IgA1 to T Antigen In Vitro by Recom-binant T-Synthase—Soluble recombinant human T-synthase wasproduced and purified as described previously (26). For the T-syn-thase reaction on IgA1: 10 �g of plasma-purified IgA from IgANpatient P7 and from control individual C16 were incubated with gua-nidinium hydrochloride (6 M final) or with the same volume of water for1h at RT. Samples were subsequently washed with water and con-centrated with Centrifugal Filter Units (4 ml, 10,000 Da cut-off, Ami-con® Ultra, Millipore, Billerica, MA) to a final volume of 50 �l. Then 6
TABLE IIgAN patients and healthy controls information
Age (years)Gender
ratio (M:F)
IgAN patients 44.57 � 13.34(S.D.) 8:6Healthy controls 32.33 � 16.94(S.D.) 6:9
O-Glycans of Human IgA1
Molecular & Cellular Proteomics 13.11 3099

�l of recombinant T-synthase, UDP-Gal (Calbiochem, San Diego, CA)(final concentration 1.4 mM) or water, and MnCl2 (final concentration20 mM) were added to the IgA concentrated solution. The mixture wasincubated overnight at 37 °C.
Semiquantitative PCR—Total RNA was extracted from 2 � 106
Dakiki or Tn4 cells with the RNeasy Mini Kit (Qiagen, Valencia, CA)according to manufacturer’s instructions. The primer pairs were syn-thesized by Integrated DNA Technologies (Coralville, IA) and are asfollows:
Cosmc, forward: GCTCCTTTTTGAAGGGTGTG reverse: TACTGC-AGCCCAAAGACTCA, T-synthase, forward: AAGGTTGACACCCAG-CCTAA, reverse: CTTTGACGTGTTTGGCCTTT, ST6GalNAcI, forward:GCAACCACAGCCAAGACGCTCATTCCCAA, reverse: TGTCACGAC-CTTCTGCACCAAGGAGTAGT, ST6GalNAcII, forward: GGAAATGTC-GGTGGAGTGTTCAGCAAG, reverse: AAGCAACTAACCCCCATCAA-GTGCCAGACCCTC. The reaction mixes were composed of 4 �l of5� Phusion GC Buffer, 0.25 �M final of dNTP, 300 nM final of primerpairs, 0.2 �l of Taq Phusion polymerase (New England Biolabs, Ips-wich, MA), and 1 �l of cDNA for a final volume of 20 �l. PCRamplifications were performed using the following cycles program:98 °C 30s, then 37 cycles of 98 °C 10s, 60 °C 20s, and 72 °C 10s,followed by a final extension of 5min at 72 °C. PCR products wereseparated on a 2% agarose gel.
N- and O-Glycan Removal and Mass Spectrometry Analysis—IgAfrom pooled IgAN patient and healthy control individual plasma sam-ples were purified as described above and IgA concentration deter-mined (Pierce® BCA Protein Assay Kit, Thermo Scientific).
N-glycans were released from �70 �g of purified IgA in a totalvolume of 90 �l using the PNGase F enzyme (New England Biolabs),according to manufacturer’s instructions: 10 �l of Denaturing Bufferwas added before boiling samples for 10min. Then, 11 �l of NonidetP-40 was added and let stand at RT for 45min. Finally, 12 �l of G7Buffer and 1.5 �l of PNGase F were added to the mix and incubatedovernight at 37 °C in rotation. Purification of the N-glycans from themixture was performed by passing the reaction mix first over apre-equilibrated C18 Sep-Pak column (Waters, Milford, MA) and thendirectly entering a pre-equilibrated Extra-Clean SPE Carbograph col-umn (Grace Davidson Discovery Science, Deerfield, IL). The C18Sep-Pak column was washed with 2 � 1 ml of Milli-Q water, then theCarbograph column was washed with 3 � 1 ml of 5% ACN � 0.1%Trifluoroacetic Acid (TFA). N-glycans were eluted from Carbographcolumn by washing with 2 ml of 15% ACN � 0.1% TFA followed by1 ml of 30% ACN � 0.1% TFA. The eluted fractions were pooled andlyophilized.
Neuraminidase treated (overnight incubation with neuraminidase,37 °C) IgA samples (�500 �g) were proceeded to HPA chromatog-raphy by the addition 200 �l of HPA-beads and incubated ON at 4 °Cin rotation. Bead mixtures were next loaded into a 2 ml Poly-PrepChromatography Column (Bio-Rad) and washed 3 times with water.The flow-through was collected as the Unbound fraction. Beads wereeluted 10 times with 100 �l of 0.2 M GalNAc. Eluted fractions werepooled, washed and concentrated in water using a centricon unit(Millipore), to a final volume of 100 �l. O-glycans were released asfollows: Pronase (Millipore) was added to the IgA samples to a finalconcentration of 0.2 �g/ml and incubated at 55 °C for 48h. Digestedsamples were lyophilized and permethylated as follows: 200 �l ofDMSO/NaOH slurry solution and 50 �l of RT methyl iodide (Sigma-Aldrich) were added to the dry sample then mixed vigorously andagitated on an automatic shaker for 30min at RT. The mixtures werecentrifuged at 5000 � g for 1min, the supernatants collected andthe precipitates discarded. 500 �l of water and 500 �l of chloroformwere added, shacked briefly and centrifuged at 5000 � g for 1min, theupper aqueous layer was discarded. This washing step was repeated2 times. The resultant chloroform layer was dried down, resuspended
in 200 �l of 50% methanol and loaded onto a pre-equilibrated C18Sep-Pak column (Waters). The column was washed with three columnvolumes of water. O-glycans were eluted with 1 ml each of 15%,35%, 50%, and 75% of ACN. Each eluted fraction was evaporated todryness in a speedvac and then resuspended in 20 �l of 50% meth-anol. Permethylated O-glycans (0.5 �l) were spotted and mixed with0.5 �l of 2,5-dihydroxybenzoic acid (DHB) (Sigma-Aldrich) matrix (10mg/ml in 50:50 ACN/water � 0.1% TFA) on a MALDI plate. MALDI-TOF mass spectra were acquired using an Ultraflex-II TOF/TOF sys-tem from Bruker Daltonics, and using positive reflector mode. Nopeeling side reactions were observed in using these conditions.
RESULTS
Plasma Concentration of IgA is Increased from IgAN Pa-tients, and O-Glycans on Plasma IgA1 From Both IgAN Pa-tients and Control Donors Similarly Contain Tn and STn Struc-tures—A common feature reported in IgAN patients is anincrease of plasma IgA as compared with healthy controls(42). We determined the concentration of plasma IgA byELISA, and found that, consistent with earlier studies, IgA waspresent at an average of 4.77 mg/ml (� 1.09 mg/ml) from 14IgAN patients, whereas in control plasma from 14 healthydonors we found 2.97 mg/ml (� 0.70 mg/ml) IgA, thus repre-senting a low, but significant �1.6 fold elevation of IgA inpatients with IgAN (Fig. 2A).
In profiling the O-glycans on glycoproteins such as IgA1 inplasma samples, we next examined their recognition by lec-tins that can discriminate between Tn and T antigens. Helixpomatia agglutinin (HPA) binds with high affinity to terminal�-GalNAc residues on O-glycans, such as the Tn antigen (43,44). In previous studies, HPA has been used to identify thepresence of Tn-containing glycans in total IgA1 by ELISA (45,46). In our study we prepared immobilized HPA in order to testwhether there are specific glycoforms of IgA1 enriched in Tnand/or STn. Plasma samples were either untreated or treatedwith neuraminidase to expose potential T and Tn antigensprior to pull-down experiments. Both the bound (B) and un-bound (UB) materials were then analyzed by SDS-PAGE andimmunoblotted with an anti-IgA1 antibody. Unexpectedly, theHPA-bound (HPA�) IgA1 was detected in all plasma samples(IgAN patients and control individuals) after desialylation (Fig.2B), as was the HPA-unbound (HPA�) IgA1. Interestingly,among 14 individuals in each group, IgA1 from plasma sam-ples of 3 IgAN patients (P4, P5, and P6) and 5 healthy controls(C10, C14, C16, C20, and C24) were pulled down by HPA onlyafter desialylation, indicating that these glycoforms of IgA1contained mostly STn antigen (Fig. 2B). For the other patientor control samples, the removal of sialic acids resulted in onlya slight increase in IgA1 staining in the HPA� fraction. Theseresults indicated that among the IgA1 molecules that haveunder-galactosylated O-glycans, the Tn and STn antigens arecommon, and that IgA1 from both IgAN patients and controlindividuals contained IgA1 expressing the Tn and STnepitopes.
We next evaluated the percentage of Tn/STn-containingIgA1 among the total IgA1 population with the same method
O-Glycans of Human IgA1
3100 Molecular & Cellular Proteomics 13.11

FIG. 2. Analysis of Tn/STn content of IgA1 from plasma: A, Total IgA plasma level (mg/ml) of IgAN patients and healthy control individuals,determined by ELISA (**, p � 0. 01). B, Western blot analysis of plasma samples after HPA-bead pull down, immunoblotted with anti-humanIgA1 (�1 chain specific) antibody. All samples were treated with (�) or without (�) neuraminidase prior to HPA chromatography. Results of 14IgAN patient samples (labeled P) are shown in the two sets of upper panels, where the top panels show the HPA� fractions and the bottompanels show the HPA� fractions. Results of 14 control samples (labeled C) are shown in the two sets of lower panels, where the top panelsshow the HPA� fractions and the bottom panels show the HPA� fractions. C, Western blot analysis of plasma samples after HPA-bead pulldown, immunoblotted with anti-human IgA1 (�1 chain specific) antibody. All samples were treated with neuraminidase prior to experiments.An equal volume of plasma sample was used as the input (I) and used for HPA pull down experiments. All subsequent unbound (UB) and bound(B) fractions were loaded into the gel for analysis. D, Quantification of HPA�-IgA1 staining (B) and total IgA1 (I). All observable bands in I andB fractions were quantified. The HPA�-IgA1 fraction intensity is expressed as a percentage of the IgA1 in the corresponding input fraction. Thedifference of HPA�-IgA1 percentage between the IgAN patients and control samples was NS. Vertical black lines between Western blot lanesindicate that lanes were not contiguous on the same gel or from different gels. ELISA assays were performed in duplicate, twice independently.
O-Glycans of Human IgA1
Molecular & Cellular Proteomics 13.11 3101
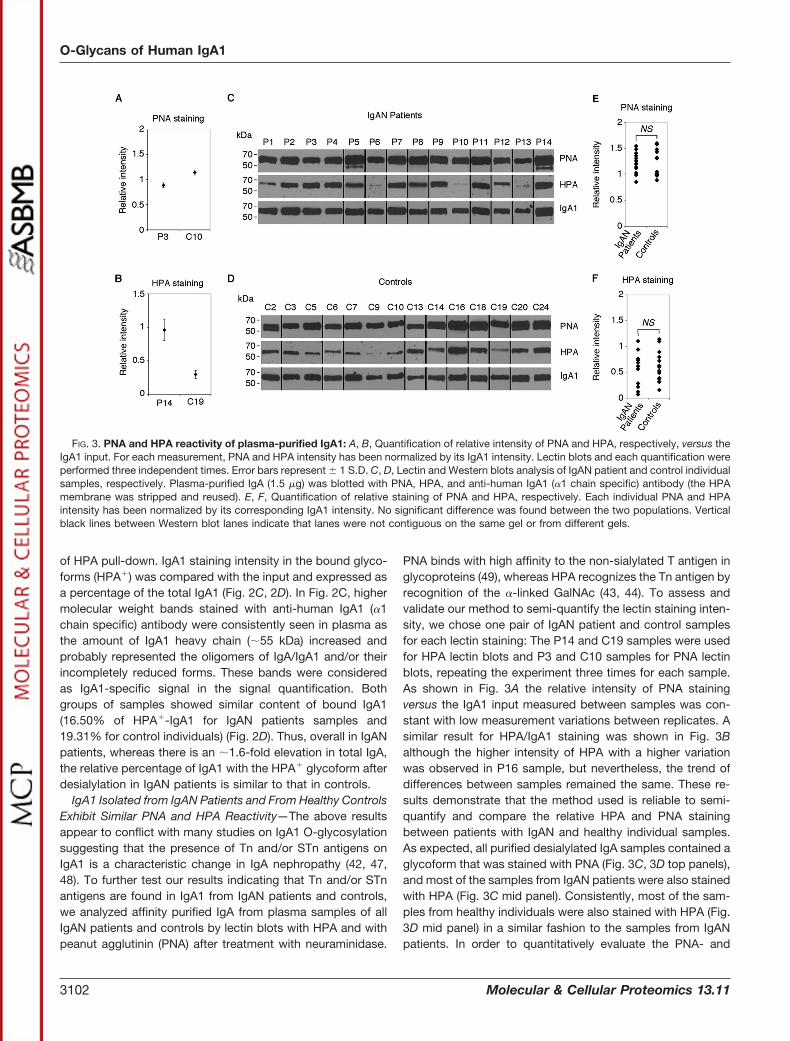
of HPA pull-down. IgA1 staining intensity in the bound glyco-forms (HPA�) was compared with the input and expressed asa percentage of the total IgA1 (Fig. 2C, 2D). In Fig. 2C, highermolecular weight bands stained with anti-human IgA1 (�1chain specific) antibody were consistently seen in plasma asthe amount of IgA1 heavy chain (�55 kDa) increased andprobably represented the oligomers of IgA/IgA1 and/or theirincompletely reduced forms. These bands were consideredas IgA1-specific signal in the signal quantification. Bothgroups of samples showed similar content of bound IgA1(16.50% of HPA�-IgA1 for IgAN patients samples and19.31% for control individuals) (Fig. 2D). Thus, overall in IgANpatients, whereas there is an �1.6-fold elevation in total IgA,the relative percentage of IgA1 with the HPA� glycoform afterdesialylation in IgAN patients is similar to that in controls.
IgA1 Isolated from IgAN Patients and From Healthy ControlsExhibit Similar PNA and HPA Reactivity—The above resultsappear to conflict with many studies on IgA1 O-glycosylationsuggesting that the presence of Tn and/or STn antigens onIgA1 is a characteristic change in IgA nephropathy (42, 47,48). To further test our results indicating that Tn and/or STnantigens are found in IgA1 from IgAN patients and controls,we analyzed affinity purified IgA from plasma samples of allIgAN patients and controls by lectin blots with HPA and withpeanut agglutinin (PNA) after treatment with neuraminidase.
PNA binds with high affinity to the non-sialylated T antigen inglycoproteins (49), whereas HPA recognizes the Tn antigen byrecognition of the �-linked GalNAc (43, 44). To assess andvalidate our method to semi-quantify the lectin staining inten-sity, we chose one pair of IgAN patient and control samplesfor each lectin staining: The P14 and C19 samples were usedfor HPA lectin blots and P3 and C10 samples for PNA lectinblots, repeating the experiment three times for each sample.As shown in Fig. 3A the relative intensity of PNA stainingversus the IgA1 input measured between samples was con-stant with low measurement variations between replicates. Asimilar result for HPA/IgA1 staining was shown in Fig. 3Balthough the higher intensity of HPA with a higher variationwas observed in P16 sample, but nevertheless, the trend ofdifferences between samples remained the same. These re-sults demonstrate that the method used is reliable to semi-quantify and compare the relative HPA and PNA stainingbetween patients with IgAN and healthy individual samples.As expected, all purified desialylated IgA samples contained aglycoform that was stained with PNA (Fig. 3C, 3D top panels),and most of the samples from IgAN patients were also stainedwith HPA (Fig. 3C mid panel). Consistently, most of the sam-ples from healthy individuals were also stained with HPA (Fig.3D mid panel) in a similar fashion to the samples from IgANpatients. In order to quantitatively evaluate the PNA- and
FIG. 3. PNA and HPA reactivity of plasma-purified IgA1: A, B, Quantification of relative intensity of PNA and HPA, respectively, versus theIgA1 input. For each measurement, PNA and HPA intensity has been normalized by its IgA1 intensity. Lectin blots and each quantification wereperformed three independent times. Error bars represent � 1 S.D. C, D, Lectin and Western blots analysis of IgAN patient and control individualsamples, respectively. Plasma-purified IgA (1.5 �g) was blotted with PNA, HPA, and anti-human IgA1 (�1 chain specific) antibody (the HPAmembrane was stripped and reused). E, F, Quantification of relative staining of PNA and HPA, respectively. Each individual PNA and HPAintensity has been normalized by its corresponding IgA1 intensity. No significant difference was found between the two populations. Verticalblack lines between Western blot lanes indicate that lanes were not contiguous on the same gel or from different gels.
O-Glycans of Human IgA1
3102 Molecular & Cellular Proteomics 13.11

HPA-reactive IgA1, the staining intensity of each sample wasmeasured and normalized by its corresponding IgA1 intensity.As shown in Fig. 3E, 3F, normalized PNA and HPA intensitiesfor patient group and controls were plotted. Although varia-tions were observed between individuals, there were no sig-nificant differences in either HPA or PNA staining betweensamples from IgAN patients and controls. Because only theHR of IgA1 but not IgA2 is O-glycosylated (50), our resultsshowed no overall differences in the types of O-glycosylationof IgA1 between IgAN patients and healthy control individuals,and Tn and STn structures naturally occur on plasma IgA1.Consistent with our results in Fig. 2, this data, which arecontrary to prior reports that Tn/STn antigens on IgA1 are thecharacteristics of IgA1 from IgAN, further confirm that theseantigens are present on IgA1 of both IgAN patients and con-trol donors.
Plasma from IgAN Patients and Healthy Controls ContainTwo Pools of IgA1 With Distinct Glycoforms that can beDistinguished by PNA or HPA Lectins—The above resultssuggest that a fraction of IgA1 in the IgA pool from plasmasamples of both IgAN patients and control donors mightcontain O-glycans with Tn and STn structures. Human IgA1HR contains nine potential O-glycosylation sites with four to
�six of them usually occupied (3). Thus, we questionedwhether the O-glycans on the IgA1 with Tn/STn antigens werecomposed of homogeneous or heterogeneous structures. Forexample, a homogeneous aberrant O-glycosylated IgA1 pop-ulation would contain Tn/STn O-glycans on all the glycosyla-tion sites and lack the core 1 (or T antigen) based O-glycans,whereas a microheterogeneous IgA1 population would con-tain all types of glycans on the same IgA1 molecule includingTn/STn and core 1 based O-glycans. To address this ques-tion, we devised an experimental approach that incorporatedserial lectin chromatography. Plasma samples were firsttreated with neuraminidase and then chromatographicallyseparated by HPA-beads into the HPA-bound (HPA�) andHPA-unbound (flow through, HPA�) materials. These two ma-terials were then further chromatographed on PNA-beads toobtain PNA-bound (PNA�) materials, which would representPNA-bound (PNA�) materials HPA�PNA� (B,B) andHPA�PNA� (U,B), as well as PNA-unbound (PNA�) materialsHPA�PNA� (B,U), HPA�PNA� (U,U), as schematically illus-trated in Fig. 4A. These subsequent fractions were analyzed byWestern blot using anti-IgA1 antibody.
Eight randomly selected plasma samples from each group,IgAN patient and control, were treated with neuraminidase
FIG. 4. Separation of IgA1 glycoforms by lectin-based chromatography: A, Scheme of the serial HPA and PNA chromatography assaysused to separate IgA1 glycoforms. B, Unbound fractions after the first round of HPA pull-down were reused for a second round of HPApull-down and then analyzed by Western blot with anti-IgA antibody. C, Western blot analysis of the stripped membranes used in B for P2 andC19 immunoblotted with anti-human IgA2 (�2 chain specific) antibody. D, Western blot analysis after HPA pull-down followed by PNApull-down using anti-human IgA1 (�1 chain specific) antibody. Eight IgAN patient samples are presented in the top panels (P), and 8 controlsamples are presented in the bottom panels (C). For B “UB” refers to the Unbound fraction, and “B” refers to the Bound fraction. For C andD “U,U” refers to the HPA�/PNA� fraction, “U,B” refers to the HPA�/PNA� fraction, “B,U” refers to the HPA�/PNA� fraction, and “B,B” refersto the HPA�/PNA� fraction. Vertical black lines between Western blot lanes indicate that lanes were not contiguous on the same gel or fromdifferent gels.
O-Glycans of Human IgA1
Molecular & Cellular Proteomics 13.11 3103

prior to HPA-bead chromatography. To measure the capacityof the lectin-column and avoid the potential problem of over-loading, in a preliminary experiment, HPA� fraction (flowthrough) from the HPA column was reloaded onto a new HPAcolumn, and HPA bound and unbound fractions were blottedwith anti-IgA1. The HPA� IgA1 was only present in the first-round chromatography, but not in the second-round fraction,demonstrating that the conditions used could exclusively andquantitatively isolate the HPA� IgA1 from HPA� IgA1 in theplasma as shown in Fig. 4B, where most of the remaining IgAstaining is detected in the unbound fractions. Moreover, IgA2molecules which are not decorated with O-glycans (50) arepresent only in the U,U lane, as predicted, corresponding tothe double negative material (HPA�PNA�), as revealed byanti-IgA2 antibody (Fig. 4C). Unexpectedly, the results shownin Fig. 4D demonstrated that the majority of plasma samplesfrom both IgAN patients and control individuals consisted ofmainly two glycoforms of IgA1: one being HPA�PNA� (ex-pressing only Tn/STn antigens and lacking the T antigen), andthe other being HPA�PNA� (expressing only T antigen with orwithout sialic acid). Some of the IgA1 was HPA�PNA�, whichmay represent a glycoform with complex O-glycans beyondsialylT, such as core 2-related structures, or non O-glycosyl-ated, that in either case would not bind either HPA or PNA.This material was not further examined. Interestingly, in addi-tion to those two glycoforms of IgA1, only sample P2 con-tained some of the HPA�PNA� (B,B) material, which wouldrepresent microheterogeneous structures (a combination ofTn/STn and T or Sialylated T). These results indicate thatplasma IgA1 molecules from both patients with IgAN andcontrol donors are distinguished by two major glycoforms:one expressing Tn/STn (B,U fraction), whereas the other ex-pressing normal O-glycans (U,B and U,U fractions). We foundthat both of the glycoforms were present within plasma IgA1from both patients with IgAN and controls. Moreover, therewere no significant differences in terms of ratios of these twodistinct glycoforms of IgA1 between samples from patientswith IgAN and healthy individuals.
Tn Antigen on IgA1 from Control and IgAN Patient SamplesCan be Converted to T Antigen In Vitro by RecombinantHuman T-Synthase—To further verify that the HPA�PNA�-IgA1 glycoform contains the Tn/STn structures and lacks theT antigen, we incubated purified IgA1 with soluble humanrecombinant T-synthase to test whether the Tn structurecould be enzymatically converted in vitro to the T antigen.Afterward, we determined the reactivity of the treated sampleto HPA and PNA, as well as to a monoclonal antibody 6E5–7Tn, which recognizes the Tn antigen in the context of the HRof IgA1 (unpublished data). As shown in Fig. 5A, 5B, 6E5–7Tnbound well to the IgA1 in P2 plasma and to purified plasmaIgA1 from both control C16 and IgAN patient P7. The P2plasma sample was treated with neuraminidase to ensure thata maximum of Tn antigen sites were exposed. The specificexpected signal, at �55 kDa corresponding to the � chain of
IgA1, was seen as a thin band because of compression by themassive amount of albumin in the plasma. Without neuramin-idase treatment, the 6E5–7Tn antibody was also able to rec-ognize the Tn antigen in the context of IgA1-HR, suggestinghere and as in Fig. 2B that the HR of IgA1 was mostlyrepresented by Tn antigens and not STn antigens. Consis-tently, the 6E5–7Tn antibody also bound to the HPA�PNA�
glycoforms of IgA1 from P3 plasma, but did not bind to theHPA�PNA� glycoform from the C14 sample, in accordancewith our previous HPA staining results. Importantly, 6E5–7Tnstaining was inhibited by addition of synthetic IgA1 hingeregion glycopeptides carrying four to �six Tn antigens, butnot by 20 mM GalNAc (Fig. 5A, 5B), demonstrating the spec-ificity of the antibody. The bands at �50 kDa and �100 kDavisible in Fig. 5B, as well as very mildly in Fig. 5A, for the P2plasma sample, were considered as nonspecific plasma-re-lated signal. Indeed, these two bands were only observed inthe plasma sample and not in any of the purified IgA samples.Importantly, the staining of these two bands was unaffectedby the presence of IgA1-HR Tn-glycopeptides, whereas the55 kDa specific signal corresponding to the � chain of IgA1was abolished.
To test as substrates of T-synthase, the two purified IgAsamples, P3 (IgAN patient) and C16 (control), were treatedwith or without guanidinium hydrochloride (Gdm-HCl) whichdenaturates proteins such as IgA1, and allowed us to explorethe efficiency of the T-synthase toward native versus dena-tured IgA1. In the presence of T-synthase and the donorUDP-Gal, but without treatment with Gdm-HCl, IgA1 wasstained by both HPA and 6E5–7Tn (Fig. 5C, 5D). However,when both IgA samples were denatured with Gdm-HCl andincubated with T-synthase in the presence of UDP-Gal,staining of both HPA and 6E5–7Tn on IgA1 was lost (Fig. 5C,5D). Given the fact that the T-synthase is a specific enzymethat recognizes only the Tn antigen acceptor, these resultsdemonstrate that the Tn antigen was present in the IgA1from both patients with IgAN and the healthy controls.Interestingly, staining of HPA and 6E5–7Tn was decreasedbut not completely abolished when the IgA1 samples werenot denatured by treatment of guanidinium hydrochloride,indicating that the conformation of IgA1 might hinder therecognition of IgA1 by T-synthase. Taken together, theseresults confirm that the Tn/STn antigen occurs on a fractionof IgA1 in plasma from both patients with IgAN and healthydonors.
IgA1-Secreting B Cell Line Dakiki Exclusively Produces IgA1With Core 1-Based Glycoform—The Dakiki cell line has beenpreviously characterized as an IgA1-secreting B cell line (29,51). Using ELISA-based assays with the Helix aspersa agglu-tinin (HAA) lectin, previous studies reported that IgA1 secretedby the Dakiki cells was galactose-deficient and mostly sialy-lated, that is, STn O-glycans (51). Thus, we reasoned that theDakiki cell line would be useful to explore the mechanismunderlying the STn on the HR of IgA1. We first examined
O-Glycans of Human IgA1
3104 Molecular & Cellular Proteomics 13.11

the expression of Cosmc, T-synthase, ST6GalNAcI, andST6GalNAcII, whose corresponding chaperone and glycosyl-transferases are involved in the formation of T, STn, andsialylT antigen structures respectively (Fig. 1). As shown inFig. 6A, Cosmc/T-synthase, whose products are responsiblefor the formation of T antigen, and ST6GalNAcII whose prod-uct is primarily responsible for synthesizing the sialylT anddisialylT structures, are expressed in Dakiki cells, whereasST6GalNAcI, which encodes a sialyltransferase primarily re-sponsible for synthesis of STn antigen on O-glycoproteins, isnot expressed. As demonstrated in previous studies (29, 38),the human B cell line Tn4 cells do not express Cosmc andST6GalNAcI genes, but do express ST6GalNAcII. Tn4 cells,which have an epigenetically silenced Cosmc (29), expressedTn antigen but not core 1-based (T antigen) structures (Fig.6B), whereas total Dakiki cell extracts exhibited strong PNAstaining only after neuraminidase treatment, but were onlyweakly stained with HPA. These results demonstrate thatDakiki cells express normal sialylated core 1-based O-glycansin cellular glycoproteins. Isolated IgA1 from the media ofDakiki cells was quantitatively bound by PNA after desialyla-
tion, but not by HPA, demonstrating that IgA1 secreted fromDakiki cells contains normal O-glycans, and little if any Tn/STnantigen (Fig. 6C). This is interesting in light of our finding thatDakiki cells do express ST6GalNAcII, confirming prior studies(29, 51), yet do not synthesize significant amounts of STn onglycoproteins including IgA1. PNA staining of IgA1 isolatedfrom Dakiki was similar to that of IgA1 isolated from both setsof plasma samples whereas HPA staining was totally absent(Fig. 6C, 6D). Consistent with this result, and in contrast to theIgA1 from patients with IgAN and healthy controls, IgA1 fromDakiki cells was exclusively bound by PNA-beads but not byHPA-beads (Figs. 6E, 2B). Taken together, these results dem-onstrated that the IgA1 secreted from Dakiki cells lacks eitherTn or STn antigens, and expresses normal sialylated core1-based O-glycans. These results also strongly argue against arole for ST6GalNAc-II in STn formation on IgA1 as suggested byearlier studies (8, 51, 52). Furthermore, the results show that inthis cell line a single major glycoform (HPA�PNA�) of IgA1 isgenerated. Thus, the formation of different glycoforms of IgA1is rather a cellular phenomenon and not inherent to all IgA1molecules.
FIG. 5. Reactivity of IgA1-hinge region Tn specific antibody and conversion of Tn antigen after incubation with T-synthase: A, Westernblot analysis of different samples with the mouse 6E5–7Tn antibody. Arrowhead indicates the IgA1 � chain size. Neu indicates neuraminidase.B, Western blot analysis. The mouse 6E5–7Tn antibody was incubated with or without 20 mM GalNAc or with or without 2.5 �g/ml of synthetichinge region (HR) glycopeptide containing 4 or more Tn antigens. Arrowheads indicate the IgA1 � chain signal (left gel) and the absence ofsignal (right gel). C, Diagram of the reaction converting Tn-containing IgA1 among total IgA1 population (IgA1-Tn/T) to T-containing IgA1 only(IgA1-T) using recombinant T-synthase in vitro. D, Western bot analyses of samples incubated with T-synthase blotted with HPA and PNAlectins, and 6E5–7Tn and anti-human IgA (� chain specific) antibodies. Healthy control sample C16 and IgAN patient sample P3 were treatedwith guanidinium hydrochloride (Gdm-HCl) prior to incubation with T-synthase with or without the donor substrate UDP-Gal. * indicatesnonspecific binding of the goat anti-mouse HRP conjugated antibody.
O-Glycans of Human IgA1
Molecular & Cellular Proteomics 13.11 3105

Mass Spectrometry Analysis of O-Glycans of IgA1—To fur-ther test our conclusions regarding O-glycosylation and dif-ferent glycoforms of IgA1, we investigated the O- and N-glycans on the distinct glycoforms of IgA1 by MS analysis. Wechose to analyze the total O-glycans after their release fromthe intact IgA or IgA1 rather than from HR glycopeptides,because the heterogeneity of the total HR glycopeptidesmakes the full analysis of all glycopeptides difficult, and inef-ficiencies and instabilities occur regarding glycopeptides intypical MALDI-TOF ionization approaches (53). Becausesome samples were in very limited amounts, we pooled sevenplasma samples from IgAN patients (P3, 4, 5, 7, 8, 11, and 12)and seven plasma samples from healthy individuals (C9, 14,16, 18, 19, 20, and 24). The seven IgAN patient plasmasamples selected correspond to the plasma samples used inFig. 4D with the exception of the sample P2, which signifi-cantly contains the HPA�/PNA� IgA1 glycoform. For the con-trol samples, seven of the eight control samples used in Fig.4D were randomly selected, because none of them signifi-cantly contain the HPA�/PNA� IgA1 glycoform. Total IgA fromthese two pools of samples was purified. The N-glycans re-leased from purified IgA of the IgAN patients and controls, aswell as the purified IgA1 of Dakiki cells, were analyzed byMS. The N-glycan profile from controls and IgAN patientsshowed a very similar pattern (Fig. 7A two upper panels), withcompositions that suggest they are mostly mono- or di-sialy-lated biantennary N-glycans, with or without a core fucose.Purified IgA from plasma include both IgA1 and IgA2, and we
cannot rule out the possibility of contributions of minoramounts of N-glycans from other glycoproteins, including IgGand IgM. Nonetheless, no differences in the N-glycan profileswere observed between IgAN patients and control groups.Interestingly, N-glycans of IgA1 produced in Dakiki cells hadcompositions consistent with non-sialylated, non-fucosylatedbiantennary N-glycans (Fig. 7A lower panel), suggesting sia-lyltransferases acting on IgA1 N-glycans are not expressed ornot functional in these cells. It should be noted that unlike IgAfrom plasma, the IgA1 from Dakiki cells is much more homo-geneous, as these cells clonally produce copious amounts ofIgA1 without significant contamination of other glycoproteins.
Next, we analyzed the whole O-glycan profile of IgA1 puri-fied from pooled plasma of IgAN patients and control individ-uals, as well as IgA1 secreted by Dakiki cells. IgA1 is one ofvery few glycoproteins in human plasma to carry O-glycans,therefore, and unlike the N-glycans, the O-glycans profilefrom purified IgA would represent the O-glycans carriedmainly, if not exclusively by IgA1. The analyses in Fig. 7Bshow that the total O-glycan profiles of IgA1 from pooled IgANpatient and healthy control plasma, which is the mixture of allglycoforms, and from Dakiki cells were similar; the majority ofthe O-glycans were primarily mono-sialylated core 1 struc-ture, with minor amounts of core 1 (T antigen), di-sialylatedcore 1 structures and, unexpectedly, di-sialylated core 2structures which have been previously reported (54).
Overall, we did not observe significant differences in N- andO-glycan MS profiles between IgA1 from IgAN patients and
FIG. 6. Characterization of IgA1 produced by the B cell line Dakiki: A, Semi-quantitative PCR analysis of Cosmc, T-synthase, ST6GalNAcI,and ST6GalNAcII genes in Dakiki and Tn4 cells. B, Western blot analysis of total protein cell extracts from Dakiki and the Cosmc-silenced Tn4cells (29) blotted with HPA and PNA lectins. Protein extracts were treated with or without neuraminidase (� or � Neu) prior to experiments.C, PNA, HPA, and IgA1 staining of 1.5 �g of media-purified IgA from Dakiki cells and plasma-purified IgA from Control and IgAN patientsamples. D, PNA and HPA measured intensities relative to IgA1 intensity, from C for Dakiki and Fig. 2C, 2D for IgAN patients and controls. E,Western blot analysis of IgA1 after PNA (� or � Lactose) and HPA chromatography of 1 �g of neuraminidase-treated IgA1 purified from Dakikicell media. UB represents the unbound fractions, B represent the bound fractions. Error bars represent S.D.
O-Glycans of Human IgA1
3106 Molecular & Cellular Proteomics 13.11

O-Glycans of Human IgA1
Molecular & Cellular Proteomics 13.11 3107

from healthy controls. It should be noted that the releasedmonosaccharide GalNAc from glycoproteins, which repre-sents the Tn antigen, cannot be observed on MALDI underthese conditions, because its molecular weight is �400 Dal-tons and causes it to be buried in the background noise of thematrix used for MS. Next, we confirmed the existence of thetwo glycoforms observed with the serial lectin chromatogra-phy as seen in Fig. 4. The pooled IgA from IgAN patients andhealthy controls were treated with neuraminidase and thenseparated by chromatography on HPA. The presence of IgAwas confirmed in both Unbound (UB, HPA�) and Bound (B,HPA�) fractions (Fig. 7C). Based on the IgA staining in Fig. 7C,an equivalent amount of HPA� and HPA� IgA was then takenfor MS analysis (Fig. 7D). The HPA Unbound (UB, HPA�)fraction of IgA1 from both IgAN patients and healthy controlscontain glycans with the composition indicating they are pri-marily of the core 1 structure (Fig. 7D upper panels), whereasthe HPA Bound (B, HPA�) fraction of IgA1 was highly deficientin such core 1-based structures (Fig. 7D bottom panels). Onlyvery minor signals corresponding to the core 1 structure couldbe observed at m/z 518. The intensity of these signals, mostlyburied in the matrix noise, represents merely about 3% of thesignal intensity observed at m/z 518 in the Fig. 7D upperpanels for the HPA� samples. In addition, these low signalscorresponding to m/z 518 are buried in the matrix and couldnot be verified as the core 1 (T antigen) structure because itcould not be analyzed by fragmentation by MS/MS. Thesedata further indicated that the HPA� glycoform exclusivelycarries Tn or STn in both IgAN patients and control individu-als.
Finally, to unequivocally confirm the presence of Tn antigenon HPA�-IgA1 fraction, we utilized the recombinant T-syn-thase to convert the Tn on HPA�-IgA1 to the core 1 structure,as we did in Fig. 5, and then the O-glycans were released andanalyzed by MS. By definition, the O-glycan structure that canbe converted by the T-synthase into the core 1 structure in thepresence of the sugar donor UDP-Gal is the Tn antigen, thatis, GalNAc�-Ser/Thr. We purified total IgA from a controldonor (C12), treated the material with neuraminidase, andthen isolated the HPA�-IgA1 glycoform by chromatographyon immobilized HPA. After denaturation with Gdm-HCl, theHPA�-IgA1 was incubated with T-synthase with or withoutUDP-Gal, and the O-glycans were then released, permethy-lated and analyzed by MS. Fig. 7E ensured that a similaramount of IgA was being analyzed in both conditions, in thepresence and absence of UDP-Gal for T-synthase reactions.Thus, the same amount of material was used to perform MS
analysis. As expected, in the absence of UDP-Gal, no core 1disaccharide structure was detected in the HPA�-IgA1 frac-tion (Fig. 7F left panel). However, when the same HPA�-IgA1fraction was incubated with UDP-Gal and with the T-syn-thase, the core 1 disaccharide was synthesized, released anddetected by MS (Fig. 7F right panel). Taken together, theseresults demonstrate that the existence of two major glyco-forms of IgA1, one expressing exclusively the Tn/STn anti-gens, and the other being exclusively normal O-glycans (Fig.8), and that both glycoforms exist in plasma of both patientswith IgAN and in normal controls.
DISCUSSION
In the present study we investigated the occurrence of theTn/STn antigens on IgA1 of patients with IgAN and healthyindividuals, and whether there were disease-specific glyco-forms of IgA1. We utilized a combination of quantitative andqualitative approaches to examine potential differences ofplasma IgA1 in their O-glycan structures using lectin blots,lectin chromatography, Western blot with monoclonal anti-body, enzymatic conversion and mass spectrometry. Theseapproaches substantially differ from those in previous studiesin the field, which did not employ methods to separate po-tential glycoforms using specific lectins, but instead usedlectin-based ELISA or MS of HR glycopeptides of total IgA(46, 48, 55–59). Here, our results have both demonstratedunexpectedly that total plasma IgA1 exists as two major andseparable glycoforms, one expressing primarily the Tn/STnantigens, and the other expressing normal sialylated andgalactosylated O-glycans, and that the relative amounts ofIgA1 glycoforms expressing Tn and/or STn antigens occur ata similar percentage in a minor fraction of plasma IgA1 frompatients with IgAN and healthy donors. Furthermore, IgA1secreted from the B cell line Dakiki is exclusively decoratedwith normal sialylated galactosylated O-glycans and lacksTn/STn antigens, despite the expression of ST6GalNAcII inthese cells. Our findings strongly argue that expression per seof Tn/STn on plasma IgA1 is not a hallmark for IgAN.
Overall our results are consistent with several historicalstudies that have detected the presence of IgA1 containingthe Tn antigen in plasma of both IgAN patients and controlindividuals in their published data (42, 45, 46, 48, 56, 58, 60).Several studies have also reported minimal or even no differ-ences between galactosylation of IgA1 from IgAN patientsand IgA1 from control donors, showing in their data similarreactivity with lectins (HAA, HPA or VVA) specific to galac-tose-deficient O-glycans (48, 60, 61). Yet, the conclusions of
Fig. 7. MS analysis of IgA1 glycoforms O-glycans: A, MS analysis of the N-glycans and B, O-glycans of purified IgA1 from plasma ofcontrol individuals and IgAN patients, and Dakiki cell culture media. C, Western blot analysis with anti-IgA antibody of HPA Unbound (UB,HPA�) and HPA Bound (B, HPA�) fractions following HPA chromatography of pooled IgAN patients and control plasma. D, MS analysis of theO-glycans of HPA� and HPA� fractions of plasma IgA1 from controls and IgAN patients. E, Western blot analysis with the anti-IgA antibodyof the HPA� fraction of C12 sample IgA1 after incubation with T-synthase, with or without UDP-Gal. F, MS analysis of the O-glycans of theHPA� fraction of C12 sample IgA1 after incubation with T-synthase, with or without UDP-Gal.
O-Glycans of Human IgA1
3108 Molecular & Cellular Proteomics 13.11

many of these reports identify galactose-deficient IgA1 as ahallmark of IgAN and as a marker to discriminate betweenIgAN patients and healthy individuals. Here, we show thatafter removal of sialic acids from IgA1, HPA can be used toefficiently pull down a fraction (�19%) of plasma IgA1 from allof the IgAN patient samples tested and, unexpectedly, fromall of the control samples as well (Fig. 2B). Without neuramin-idase treatment, only 3 of 14 IgAN patients and 5 of 14 controlsamples did not exhibit significant IgA1 pull down with HPA,suggesting that the Tn antigen in some IgA1 glycoforms inthose plasma is mostly sialylated and occurs as the STnantigen. In addition, the purified IgA1 from all control and IgANpatient samples exhibited direct and similar HPA reactivityafter desialylation. Therefore, based on our results, we con-clude that there is no significant difference in the Tn/STncontent in plasma IgA1 between patients with IgAN andhealthy controls.
Consistent with several earlier findings (42, 48), we foundthat plasma of IgAN patients contains significantly more IgA(�1.6-fold higher) than plasma of control individuals (Fig. 2A).This differential level of IgA1 expression was the only signifi-cant difference between those two groups that we couldidentify. Of course, our results imply that the absolute amountof Tn antigen-containing IgA1 glycoforms is, on average, alsoelevated �1.6-fold in the plasma of IgAN patients, thus, thisoverall elevation of the Tn/STn-containing glycoforms of IgA1could play a role in the pathogenesis. We also noted thatsome healthy individuals have a higher level of plasma galac-
tose-deficient IgA1 (the HPA�/PNA� glycoforms) than someIgAN patients. Therefore, the sole expression of Tn antigen onplasma IgA1 is unlikely to be useful as a unique diagnosticfeature for IgAN.
An unexpected finding in our study is the identification oftwo major and separable glycoforms of IgA1 molecules inplasma of all donors (Fig. 8). By performing serial HPA- andPNA-chromatography on plasma IgA1 of IgAN patients orhealthy individuals, we found that IgA1 molecules from 15 ofthe 16 samples tested could be easily separated into twounique fractions in their reactivity to only one or the otherlectin (HPA�/PNA� or HPA�/PNA�), and only one of those 16samples contained a third glycoform that was the HPA�PNA�
glycoform of IgA1 (Fig. 4D). We confirmed the existence of theHPA�/PNA� glycoform by demonstrating it could be readilyconverted to the core 1 structure by recombinant humanT-synthase in vitro. We also verified by MS analysis that nocore 1 based structures were detected in the HPA�/PNA�
glycoform of IgA1, whereas core 1 O-glycans were found inthe HPA�/PNA� glycoform (Fig. 7D, 7F). These data demon-strate that IgA1 occurs in relatively homogeneous and sepa-rable glycoforms in terms of O-glycan galactosylation, andcan be readily distinguished and isolated by chromatographyon HPA and PNA, and that both glycoforms occur in IgANpatients and healthy controls.
Several past studies have analyzed the O-glycans of IgA1,but the approaches have differed from ours. Previous studiesgenerally utilize the unfractionated IgA1 material, that is often
FIG. 8. Two distinct glycoforms of IgA1 in human plasma: IgA1 is the major O-glycoprotein in human plasma. From this study, results haveshown that two distinct glycoforms of IgA1 occur in plasma, the majority of IgA1 (80�85%) with the normal O-glycans, for example, mono-and/or di-sialyl Core 1 structures which is recognized by PNA but not HPA (PNA�HPA�) after desialylation; the other, a minority of IgA1(15�20%) containing the Tn and STn antigens which can be bound by HPA, but not PNA (PNA�HPA�) after desialylation.
O-Glycans of Human IgA1
Molecular & Cellular Proteomics 13.11 3109

digested by trypsin, followed by MS analysis of the 33-mertryptic glycopeptide corresponding to the hinge region (55,62–65). Such studies have suggested a rather wide hetero-geneity in the O-glycan content of serum IgA1 from IgANpatients and healthy individuals with up to 57 differentiallyglycosylated hinge region glycopeptides; accordingly, most ofthe IgA1 would be predicted by such studies to be mixedglycoforms and would be predicted to be bound by both PNAand HPA lectins after desialylation. To our knowledge, ourwork is the first attempt to separate IgA1 glycoforms from thesame individual’s plasma with two lectins thought to discrim-inate “normal” (PNA�) and “aberrant” (HPA�) O-glycosylatedIgA1.
Indications of these glycoform homogeneities were furtherconfirmed when we investigated the O-glycosylation of IgA1secreted by the immortalized B cell line Dakiki. Unlike theplasma samples tested, IgA1 purified from the culture mediaof Dakiki cells showed reactivity only to PNA after desialyla-tion, suggesting that only sialylated core 1-based O-glycansare present in the IgA1 secreted by these cells (Fig. 6). Dakikicells represent a clonal expansion of B cells and therefore it isnot surprising that they produce a homogeneous populationof IgA1. The presence of a single major glycoform with galac-tosylated O-glycans on IgA1 from Dakiki cells demonstratesthat IgA1 in general can be efficiently galactosylated by T-synthase in cells to form the core 1-based O-glycans. Thisraises the question as to the mechanism for formation of theundergalactosylated glycoforms in plasma we designated asHPA�/PNA�.
Although our work clearly demonstrates that the so-calledaberrant O-glycosylation of IgA1, that is, expression of the Tnand STn antigens, occurs in all individuals including healthycontrols, the cause of this phenomenon is not yet known. Wecan envision three scenarios that might explain the presenceof two glycoforms of IgA1 in the plasma of individuals. InScenario I there may be two different populations of IgA1-producing B cells or plasma cells with one “major” populationthat produces IgA1 with “normal” core 1 based O-glycans,whereas another “minor” population is unable to add galac-tose to the O-glycans of IgA1 and secretes IgA1 carryingTn/STn antigens. In Scenario II the IgA1-producing cells si-multaneously produce both HPA�/PNA� and HPA�/PNA�
glycoforms of IgA1, whereby for either kinetic or other rea-sons, one portion of IgA1 molecules made by the cells is notO-galactosylated, whereas another portion is O-galactosyl-ated. In Scenario III the IgA1 produced in a particular cell maybe folded into such a structural form that the hinge region isonly accessible to the ppGalNAcTs but not to the T-synthase.Our results with the Dakiki cells tend to favor Scenario I,because the cells produce a single glycoform that is HPA�/PNA�. We note that in some samples there was a smallamount of a heterogeneous HPA�/PNA� glycoforms, whichmight arise if there is a limited amount of T-synthase activityor an excess amount of IgA1 in cells, also taking into account
the evidence suggesting that the IgA1 hinge region is not agood substrate for T-synthase, or T-synthase has a very highKm to the Tn antigen on the hinge region of IgA1.
If the different glycoforms we observed arise by Scenario I,it may suggest that the minor population of B cells or plasmacells which produce the HPA�/PNA� (Tn/STn) glycoform ofIgA1 may have a deficiency in T-synthase activity, either tem-porary down-regulation of Cosmc and/or T-synthase, or per-sistently compromised T-synthase activity, such as silencingand mutation of Cosmc as discussed below. The poor kineticactivity of T-synthase to the IgA1 coupled with deficiency ofT-synthase could lead to a deficiency of galactose on IgA1 yetO-glycosylation of cellular glycoproteins could be relativelynormal. Consistent with this are some studies proposing thatpatients with IgAN show decreased expression of Cosmcand/or T-synthase (8, 34, 36). However, we observed no sig-nificant differences in the percentage of IgA1 glycoforms lack-ing O-galactosylation between patients with IgAN and healthycontrols. It is also possible that there could be site-specificdifferences in O-glycosylation among the two glycoforms thatwe observed independent of their total content of galactose orTn/STn antigens, and that such site-specific differences mightcontribute to disease pathogenesis. In any case, using ouraffinity isolation approaches, such site-specific studies shouldbe done on isolated glycoforms of IgA1 from both normalcontrols and IgAN patients.
Finally, our results also strongly argue that ST6GalNAc-IIdoes not compete with the T-synthase to promote theformation of the STn antigen, because we observed thatST6GalNAcII is expressed in the Dakiki cells, yet IgA1 se-creted from these cells is negative to HPA after desialylation.In addition, Dakiki cells do not express, or so little if any, STnon their glycoproteins (Fig. 6B). Suzuki and others (8, 51)concluded, using indirect approaches, that up-regulatedST6GalNAcII is responsible for the synthesis of STn on IgA1.ST6GalNAc-II, however, has been primarily described to sia-lylate the core 1 and �2,3sialyl-core 1 structures to formNeuAc�2,6GalNAc- within sialyl-core 1 and disialyl-core 1structures (66–68), whereas ST6GalNAc-I is mainly involvedin sialylation of the Tn, but not T, antigen to form the STn (69,70). Consistent with these data, we also observed that Tn4cells only express the Tn antigen, but no significant amountsof the STn antigen, although they express ST6GalNAcII, butnot ST6GalNAcI (Fig. 6A). Lastly, we observed in preliminarystudies that the recombinant human ST6GalNAc-II is onlyable to sialylate core 1 and �2,3sialyl-core 1 structures onIgA1 HR peptides in vitro, but not the Tn antigen on IgA1 HRpeptides. Taken together, these data strongly indicate thatST6GalNAc-II is not the sialyltransferase responsible for thesynthesis of the STn antigen on IgA1.
In conclusion, we have demonstrated that plasma IgA1from patients with IgAN and healthy control individuals simi-larly contain IgA1 with Tn/STn antigens. However, we foundthat IgA1 in plasma from both IgAN patients and healthy
O-Glycans of Human IgA1
3110 Molecular & Cellular Proteomics 13.11

controls exists as two distinct glycoforms, one with Tn/STnO-glycans as a minor fraction, and the other as the majorfraction containing the normal sialylated and galactosylatedO-glycans (Fig. 8). Thus, the Tn antigen, considered as amarker of pathological conditions or neoplastic transforma-tions, naturally occurs in healthy individuals on IgA1 at asimilar level as compared with patients with IgAN, and there-fore should not be considered as a unique feature for IgA inIgAN. The only significant difference we found between theIgAN patient samples and the control samples is the totalplasma IgA concentration, being 60% higher in average inIgAN patients than normal controls. Finally, we showed thatT-synthase can fully galactosylate denatured IgA1 in vitro,suggesting that the appearance of the Tn antigen could bebecause of poor efficiency of T-synthase to use the IgA1hinge region as a substrate in vivo. Such a kinetic constraintmight act in addition to the possibility that there are twogeneral populations of B cells generating IgA1, one generatingnormally O-glycosylated IgA1 and another generating the Tn/STn glycoform. Together, these results regarding IgA1 O-gly-cosylation in IgAN strongly suggest that expression of Tnantigen alone is not sufficient to cause the disease and ad-vocate for a reevaluation of the role and importance of O-gly-cosylation of IgA1 in IgA Nephropathy.
Acknowledgments—We thank Drs. Jamie Heimburg-Molinaro andDavid F. Smith for help in editing the manuscript.
* This work was supported by NIH Grant R01DK80876 (to T. J.).** To whom correspondence should be addressed: Department of
Biochemistry, Emory University School of Medicine, 1510 CliftonRoad, Room 4029, Atlanta, GA 30322. E-mail: [email protected].
‡‡ Current address: Department of Molecular and Cell Biology,Boston University, Boston MA 02118.
§§ Current address: Department of Neurobiology, Harvard MedicalSchool, Boston MA 02115.
REFERENCES
1. Berger, J., and Hinglais, N. (1968) [Intercapillary deposits of IgA-IgG].J. Urol. Nephrol. 74, 694–695
2. Beerman, I., Novak, J., Wyatt, R. J., Julian, B. A., and Gharavi, A. G. (2007)The genetics of IgA nephropathy. Nat. Clin. Pract. Nephrol. 3, 325–338
3. Tarelli, E., Smith, A. C., Hendry, B. M., Challacombe, S. J., and Pouria, S.(2004) Human serum IgA1 is substituted with up to six O-glycans asshown by matrix assisted laser desorption ionisation time-of-flight massspectrometry. Carbohydr. Res. 339, 2329–2335
4. Giannakakis, K., Feriozzi, S., Perez, M., Faraggiana, T., and Muda, A. O.(2007) Aberrantly glycosylated IgA1 in glomerular immune deposits ofIgA nephropathy. J. Am. Soc. Nephrol. 18, 3139–3146
5. Hiki, Y. (2009) O-linked oligosaccharides of the IgA1 hinge region: roles ofits aberrant structure in the occurrence and/or progression of IgA ne-phropathy. Clin. Exp. Nephrol. 13, 415–423
6. Mestecky, J., Tomana, M., Moldoveanu, Z., Julian, B. A., Suzuki, H., Ma-tousovic, K., Renfrow, M. B., Novak, L., Wyatt, R. J., and Novak, J. (2008)Role of aberrant glycosylation of IgA1 molecules in the pathogenesis ofIgA nephropathy. Kidney Blood Pressure Res. 31, 29–37
7. Novak, J., Julian, B. A., Tomono, M., and Mestecky, J. (2008) IgA glyco-sylation and IgA immune complexes in the pathogenesis of IgA nephrop-athy. Seminars Nephrol. 28, 78–87
8. Suzuki, H., Moldoveanu, Z., Hall, S., Brown, R., Vu, H. L., Novak, L., Julian,B. A., Tomana, M., Wyatt, R. J., Edberg, J. C., Alarcon, G. S., Kimberly,R. P., Tomino, Y., Mestecky, J., and Novak, J. (2008) IgA1-secreting cell
lines from patients with IgA nephropathy produce aberrantly glycosyl-ated IgA1. J. Clin. Investig. 118, 629–639
9. McEver, R. P., Moore, K. L., and Cummings, R. D. (1995) Leukocytetrafficking mediated by selectin-carbohydrate interactions. J. Biol.Chem. 270, 11025–11028
10. Homeister, J. W., Thall, A. D., Petryniak, B., Maly, P., Rogers, C. E., Smith,P. L., Kelly, R. J., Gersten, K. M., Askari, S. W., Cheng, G., Smithson, G.,Marks, R. M., Misra, A. K., Hindsgaul, O., von Andrian, U. H., and Lowe,J. B. (2001) The alpha(1,3)fucosyltransferases FucT-IV and FucT-VII exertcollaborative control over selectin-dependent leukocyte recruitment andlymphocyte homing. Immunity 15, 115–126
11. Ju, T., Otto, V. I., and Cummings, R. D. (2011) The Tn antigen-structuralsimplicity and biological complexity. Angew Chem Int Ed Engl 50,1770–1791
12. Yeh, J. C., Hiraoka, N., Petryniak, B., Nakayama, J., Ellies, L. G., Rabuka,D., Hindsgaul, O., Marth, J. D., Lowe, J. B., and Fukuda, M. (2001) Novelsulfated lymphocyte homing receptors and their control by a Core1extension beta 1,3-N-acetylglucosaminyltransferase. Cell 105, 957–969
13. Xia, L., Ju, T., Westmuckett, A., An, G., Ivanciu, L., McDaniel, J. M., Lupu,F., Cummings, R. D., and McEver, R. P. (2004) Defective angiogenesisand fatal embryonic hemorrhage in mice lacking core 1-derived O-glycans. J. Cell Biol. 164, 451–459
14. Fu, J., Gerhardt, H., McDaniel, J. M., Xia, B., Liu, X., Ivanciu, L., Ny, A.,Hermans, K., Silasi-Mansat, R., McGee, S., Nye, E., Ju, T., Ramirez, M. I.,Carmeliet, P., Cummings, R. D., Lupu, F., and Xia, L. (2008) Endothelialcell O-glycan deficiency causes blood/lymphatic misconnections andconsequent fatty liver disease in mice. J. Clin. Investig. 118, 3725–3737
15. Mellis, S. J., and Baenziger, J. U. (1983) Structures of the O-glycosidicallylinked oligosaccharides of human IgD. J. Biol. Chem. 258, 11557–11563
16. Field, M. C., Dwek, R. A., Edge, C. J., and Rademacher, T. W. (1989)O-linked oligosaccharides from human serum immunoglobulin A1.Biochem. Soc. Trans. 17, 1034–1035
17. Clausen, H., and Bennett, E. P. (1996) A family of UDP-GalNAc: polypeptideN-acetylgalactosaminyl-transferases control the initiation of mucin-typeO-linked glycosylation. Glycobiology 6, 635–646
18. Ten Hagen, K. G., Fritz, T. A., and Tabak, L. A. All in the family: theUDP-GalNAc:polypeptide N-acetylgalactosaminyltransferases. Glycobi-ology 13:1R-16R, 2003
19. Li, X., Wang, J., Li, W., Xu, Y., Shao, D., Xie, Y., Xie, W., Kubota, T.,Narimatsu, H., and Zhang, Y. (2012) Characterization of ppGalNAc-T18,a member of the vertebrate-specific Y subfamily of UDP-N-acetyl-alpha-D-galactosamine:polypeptide N-acetylgalactosaminyltransferases. Gly-cobiology 22, 602–615
20. Gill, D. J., Clausen, H., and Bard, F. (2011) Location, location, location: newinsights into O-GalNAc protein glycosylation. Trends Cell Biol. 21, 149–158
21. Iwasaki, H., Zhang, Y., Tachibana, K., Gotoh, M., Kikuchi, N., Kwon, Y. D.,Togayachi, A., Kudo, T., Kubota, T., and Narimatsu, H. (2003) Initiation ofO-glycan synthesis in IgA1 hinge region is determined by a single en-zyme, UDP-N-acetyl-alpha-D-galactosamine:polypeptide N-acetyl-galactosaminyltransferase 2. J. Biol. Chem. 278, 5613–5621
22. Ju, T., Brewer, K., D’Souza, A., Cummings, R. D., and Canfield, W. M.(2002) Cloning and expression of human core 1 beta1,3-galactosyltrans-ferase. J. Biol. Chem. 277, 178–186
23. Byrd, J. C., and Bresalier, R. S. (2004) Mucins and mucin binding proteinsin colorectal cancer. Cancer Metastasis Rev. 23, 77–99
24. Ju, T., and Cummings, R. D. (2002) A unique molecular chaperone Cosmcrequired for activity of the mammalian core 1 beta 3-galactosyltrans-ferase. Proc. Natl. Acad. Sci. U.S.A. 99, 16613–16618
25. Ju, T., Aryal, R. P., Stowell, C. J., and Cummings, R. D. (2008) Regulationof protein O-glycosylation by the endoplasmic reticulum-localized mo-lecular chaperone Cosmc. J. Cell Biol. 182, 531–542
26. Aryal, R. P., Ju, T., and Cummings, R. D. (2010) The endoplasmic reticulumchaperone Cosmc directly promotes in vitro folding of T-synthase.J. Biol. Chem. 285, 2456–2462
27. Ju, T., Lanneau, G. S., Gautam, T., Wang, Y., Xia, B., Stowell, S. R., Willard,M. T., Wang, W., Xia, J. Y., Zuna, R. E., Laszik, Z., Benbrook, D. M.,Hanigan, M. H., and Cummings, R. D. (2008) Human tumor antigens Tnand sialyl Tn arise from mutations in Cosmc. Cancer Res. 68, 1636–1646
28. Wang, Y., Ju, T., Ding, X., Xia, B., Wang, W., Xia, L., He, M., and Cummings,R. D. (2010) Cosmc is an essential chaperone for correct protein O-gly-cosylation. Proc. Natl. Acad. Sci. U.S.A. 107, 9228–9233
O-Glycans of Human IgA1
Molecular & Cellular Proteomics 13.11 3111

29. Mi, R., Song, L., Wang, Y., Ding, X., Zeng, J., Lehoux, S., Aryal, R. P., Wang,J., Crew, V. K., van Die, I., Chapman, A. B., Cummings, R. D., and Ju, T.(2012) Epigenetic silencing of the chaperone Cosmc in human leuko-cytes expressing tn antigen. J. Biol. Chem.287, 41523–41533
30. Dahr, W., Uhlenbruck, G., Gunson, H. H., and Van Der Hart, M. (1975)Molecular basis of Tn-polyagglutinability. Vox Sang 29, 36–50
31. Berger, E. G. (1999) Tn-syndrome. Biochim. Biophys. Acta 1455, 255–26832. Springer, G. F. (1997) Immunoreactive T and Tn epitopes in cancer diag-
nosis, prognosis, and immunotherapy. J. Mol. Med. 75, 594–60233. Brockhausen, I., Yang, J., Dickinson, N., Ogata, S., and Itzkowitz, S. H.
(1998) Enzymatic basis for sialyl-Tn expression in human colon cancercells. Glycoconj. J. 15, 595–603
34. Allen, A. C., Topham, P. S., Harper, S. J., and Feehally, J. (1997) Leucocytebeta 1,3 galactosyltransferase activity in IgA nephropathy. Nephrology,Dialysis, Transplantation 12, 701–706
35. Qin, W., Zhou, Q., Yang, L. C., Li, Z., Su, B. H., Luo, H., and Fan, J. M.(2005) Peripheral B lymphocyte beta1,3-galactosyltransferase and chap-erone expression in immunoglobulin A nephropathy. J. Intern. Med. 258,467–477
36. Yamada, K., Kobayashi, N., Ikeda, T., Suzuki, Y., Tsuge, T., Horikoshi, S.,Emancipator, S. N., and Tomino, Y. (2010) Down-regulation of core 1beta1,3-galactosyltransferase and Cosmc by Th2 cytokine alters O-gly-cosylation of IgA1. Nephrology, Dialysis, Transplantation 25, 3890–3897
37. Inoue, T., Sugiyama, H., Hiki, Y., Takiue, K., Morinaga, H., Kitagawa, M.,Maeshima, Y., Fukushima, K., Nishizaki, K., Akagi, H., Narimatsu, Y.,Narimatsu, H., and Makino, H. (2010) Differential expression of glyco-genes in tonsillar B lymphocytes in association with proteinuria and renaldysfunction in IgA nephropathy. Clin. Immunol. 136, 447–455
38. Crew, V. K., Singleton, B. K., Green, C., Parsons, S. F., Daniels, G., andAnstee, D. J. (2008) New mutations in C1GALT1C1 in individuals with Tnpositive phenotype. Br. J. Hematol. 142, 657–667
39. White, T., Bennett, E. P., Takio, K., Sorensen, T., Bonding, N., and Clausen,H. (1995) Purification and cDNA cloning of a human UDP-N-acetyl-alpha-D-galactosamine:polypeptide N-acetylgalactosaminyltransferase.J. Biol. Chem. 270, 24156–24165
40. Sorensen, A. L., Reis, C. A., Tarp, M. A., Mandel, U., Ramachandran, K.,Sankaranarayanan, V., Schwientek, T., Graham, R., Taylor-Papadimi-triou, J., Hollingsworth, M. A., Burchell, J., and Clausen, H. (2006) Che-moenzymatically synthesized multimeric Tn/STn MUC1 glycopeptideselicit cancer-specific anti-MUC1 antibody responses and override toler-ance. Glycobiology 16, 96–107
41. Tarp, M. A., Sorensen, A. L., Mandel, U., Paulsen, H., Burchell, J., Taylor-Papadimitriou, J., and Clausen, H. (2007) Identification of a novel cancer-specific immunodominant glycopeptide epitope in the MUC1 tandemrepeat. Glycobiology 17, 197–209
42. Moldoveanu, Z., Wyatt, R. J., Lee, J. Y., Tomana, M., Julian, B. A.,Mestecky, J., Huang, W. Q., Anreddy, S. R., Hall, S., Hastings, M. C.,Lau, K. K., Cook, W. J., and Novak, J. (2007) Patients with IgA nephrop-athy have increased serum galactose-deficient IgA1 levels. Kidney Int.71, 1148–1154
43. Piller, V., Piller, F., and Cartron, J. P. (1990) Comparison of the carbohy-drate-binding specificities of seven N-acetyl-D-galactosamine-recogniz-ing lectins. Eur. J. Biochem. 191, 461–466
44. Hammarstrom, S. (1974) Structure, specificity, binding properties, andsome biological activities of a blood group A-reactive hemagglutinin fromthe snail Helix pomatia. Ann. NY Acad. Sci. 234, 183–197
45. Moore, J. S., Kulhavy, R., Tomana, M., Moldoveanu, Z., Suzuki, H., Brown,R., Hall, S., Kilian, M., Poulsen, K., Mestecky, J., Julian, B. A., and Novak,J. (2007) Reactivities of N-acetylgalactosamine-specific lectins with hu-man IgA1 proteins. Mol. Immunol. 44, 2598–2604
46. Tomana, M., Matousovic, K., Julian, B. A., Radl, J., Konecny, K., andMestecky, J. (1997) Galactose-deficient IgA1 in sera of IgA nephropathypatients is present in complexes with IgG. Kidney Int. 52, 509–516
47. Hiki, Y., Odani, H., Takahashi, M., Yasuda, Y., Nishimoto, A., Iwase, H.,Shinzato, T., Kobayashi, Y., and Maeda, K. (2001) Mass spectrometryproves under-O-glycosylation of glomerular IgA1 in IgA nephropathy.Kidney Int. 59, 1077–1085
48. Shimozato, S., Hiki, Y., Odani, H., Takahashi, K., Yamamoto, K., andSugiyama, S. (2008) Serum under-galactosylated IgA1 is increased inJapanese patients with IgA nephropathy. Nephrology, Dialysis, Trans-plantation 23, 1931–1939
49. Lotan, R., Skutelsky, E., Danon, D., and Sharon, N. (1975) The purification,composition, and specificity of the anti-T lectin from peanut (Arachishypogaea). J. Biol. Chem. 250, 8518–8523
50. Torano, A., and Putnam, F. W. (1978) Complete amino acid sequence of thealpha 2 heavy chain of a human IgA2 immunoglobulin of the A2m (2)allotype. Proc. Natl. Acad. Sci. U.S.A. 75, 966–969
51. Raska, M., Moldoveanu, Z., Suzuki, H., Brown, R., Kulhavy, R., Andrasi, J.,Hall, S., Vu, H. L., Carlsson, F., Lindahl, G., Tomana, M., Julian, B. A.,Wyatt, R. J., Mestecky, J., and Novak, J. (2007) Identification and char-acterization of CMP-NeuAc:GalNAc-IgA1 alpha2,6-sialyltransferase inIgA1-producing cells. J. Mol. Biol. 369, 69–78
52. Suzuki, H., Raska, M., Yamada, K., Moldoveanu, Z., Julian, B. A., Wyatt,R. J., Tomino, Y., Gharavi, A. G., and Novak, J. (2014) Cytokines AlterIgA1 O-Glycosylation by Dysregulating C1GalT1 and ST6GalNAc-II En-zymes. J. Biol. Chem. 289, 5330–5339
53. Leymarie, N., and Zaia, J. (2012) Effective use of mass spectrometry forglycan and glycopeptide structural analysis. Anal. Chem. 84, 3040–3048
54. Goetz, J. A., Novotny, M. V., and Mechref, Y. (2009) Enzymatic/chemicalrelease of O-glycans allowing MS analysis at high sensitivity. Anal.Chem. 81, 9546–9552
55. Hiki, Y., Tanaka, A., Kokubo, T., Iwase, H., Nishikido, J., Hotta, K., andKobayashi, Y. (1998) Analyses of IgA1 hinge glycopeptides in IgA ne-phropathy by matrix-assisted laser desorption/ionization time-of-flightmass spectrometry. J. Am. Soc. Nephrol. 9, 577–582
56. Allen, A. C., Harper, S. J., and Feehally, J. (1995) Galactosylation of N- andO-linked carbohydrate moieties of IgA1 and IgG in IgA nephropathy. Clin.Exp. Immunol. 100, 470–474
57. Novak, J., Tomana, M., Kilian, M., Coward, L., Kulhavy, R., Barnes, S., andMestecky, J. (2000) Heterogeneity of O-glycosylation in the hinge regionof human IgA1. Mol. Immunol. 37, 1047–1056
58. Lau, K. K., Wyatt, R. J., Moldoveanu, Z., Tomana, M., Julian, B. A., Hogg,R. J., Lee, J. Y., Huang, W. Q., Mestecky, J., and Novak, J. (2007) Serumlevels of galactose-deficient IgA in children with IgA nephropathy andHenoch-Schonlein purpura. Pediatric Nephrol. 22, 2067–2072
59. Zhao, N., Hou, P., Lv, J., Moldoveanu, Z., Li, Y., Kiryluk, K., Gharavi, A. G.,Novak, J., and Zhang, H. (2012) The level of galactose-deficient IgA1 inthe sera of patients with IgA nephropathy is associated with diseaseprogression. Kidney Int. 82, 790–796
60. Smith, A. C., de Wolff, J. F., Molyneux, K., Feehally, J., and Barratt, J. (2006)O-glycosylation of serum IgD in IgA nephropathy. J. Am. Soc. Nephrol.17, 1192–1199
61. Berthoux, F., Suzuki, H., Thibaudin, L., Yanagawa, H., Maillard, N., Mariat,C., Tomino, Y., Julian, B. A., and Novak, J. (2012) Autoantibodies tar-geting galactose-deficient IgA1 associate with progression of IgA ne-phropathy. J. Am. Soc. Nephrol. 23, 1579–1587
62. Iwase, H., Tanaka, A., Hiki, Y., Kokubo, T., Ishii-Karakasa, I., Kobayashi, Y.,and Hotta, K. (1996) Estimation of the number of O-linked oligosaccha-rides per heavy chain of human serum IgA1 by matrix-assisted laserdesorption ionization time-of-flight mass spectrometry (MALDI-TOFMS)analysis of the hinge glycopeptide. J. Biochem. 120, 393–397
63. Odani, H., Hiki, Y., Takahashi, M., Nishimoto, A., Yasuda, Y., Iwase, H.,Shinzato, T., and Maeda, K. (2000) Direct evidence for decreased sialy-lation and galactosylation of human serum IgA1 Fc O-glycosylated hingepeptides in IgA nephropathy by mass spectrometry. Biochem. Biophys.Res. Commun. 271, 268–274
64. Takahashi, K., Smith, A. D., Poulsen, K., Kilian, M., Julian, B. A., Mestecky,J., Novak, J., and Renfrow, M. B. (2012) Naturally occurring structuralisomers in serum IgA1 o-glycosylation. J. Proteome Res. 11, 692–702
65. Odani, H., Yamamoto, K., Iwayama, S., Iwase, H., Takasaki, A., Takahashi,K., Fujita, Y., Sugiyama, S., and Hiki, Y. (2010) Evaluation of the specificstructures of IgA1 hinge glycopeptide in 30 IgA nephropathy patients bymass spectrometry. J. Nephrol. 23, 70–76
66. Marcos, N. T., Pinho, S., Grandela, C., Cruz, A., Samyn-Petit, B., Harduin-Lepers, A., Almeida, R., Silva, F., Morais, V., Costa, J., Kihlberg, J.,Clausen, H., and Reis, C. A. (2004) Role of the human ST6GalNAc-I andST6GalNAc-II in the synthesis of the cancer-associated sialyl-Tn antigen.Cancer Res. 64, 7050–7057
67. Sewell, R., Backstrom, M., Dalziel, M., Gschmeissner, S., Karlsson, H., Noll,T., Gatgens, J., Clausen, H., Hansson, G. C., Burchell, J., and Taylor-Papadimitriou, J. (2006) The ST6GalNAc-I sialyltransferase localizesthroughout the golgi and is responsible for the synthesis of the tumor-
O-Glycans of Human IgA1
3112 Molecular & Cellular Proteomics 13.11

associated Sialyl-Tn O-glycan in human breast cancer. J. Biol. Chem.281, 3586–3594
68. Samyn-Petit, B., Krzewinski-Recchi, M. A., Steelant, W. F., Delannoy, P.,and Harduin-Lepers, A. (2000) Molecular cloning and functional expres-sion of human ST6GalNAc II. Molecular expression in various humancultured cells. Biochim. Biophys. Acta 1474, 201–211
69. Ikehara, Y., Kojima, N., Kurosawa, N., Kudo, T., Kono, M., Nishihara, S., Issiki,S., Morozumi, K., Itzkowitz, S., Tsuda, T., Nishimura, S. I., Tsuji, S., andNarimatsu, H. (1999) Cloning and expression of a human gene encoding an
N-acetylgalactosamine-alpha2,6-sialyltransferase (ST6GalNAc I): a candi-date for synthesis of cancer-associated sialyl-Tn antigens. Glycobiology9, 1213–1224
70. Sewell, R., Backstrom, M., Dalziel, M., Gschmeissner, S., Karlsson, H., Noll,T., Gatgens, J., Clausen, H., Hansson, G. C., Burchell, J., and Taylor-Papadimitriou, J. (2006) The ST6GalNAc-I sialyltransferase localizesthroughout the Golgi and is responsible for the synthesis of the tumor-associated sialyl-Tn O-glycan in human breast cancer. J. Biol. Chem.281, 3586–3594
O-Glycans of Human IgA1
Molecular & Cellular Proteomics 13.11 3113
Copyright © 2022 FDOKUMEN




















