Hydrogen Sulfide Improves Neutrophil Migration and Survival in Sepsis via K+ATP Channel Activation
46
HYDROGEN SULFIDE IMPROVES NEUTROPHIL MIGRATION AND SURVIVAL IN SEPSIS VIA K + ATP CHANNEL ACTIVATION Fernando Spiller 1 , Maria I.L. Orrico 1 , Daniele C. Nascimento 1 , Paula G. Czaikoski 1 , Fabrício O. Souto 3 , José C. Alves-Filho 1 , Andressa Freitas 1 , Daniela Carlos 1 , Marcelo F. Montenegro 1 , Alberto F. Neto 4 , Sergio H. Ferreira 1 , Marcos A. Rossi 2 , John S. Hothersall 1 , Jamil Assreuy 5 and Fernando Q. Cunha 1 . Departments of 1 Pharmacology, 2 Pathology, and 3 Surgery and Anatomy, School of Medicine of Ribeirao Preto; 4 Department of Pharmaceutical Science, Faculty of Pharmaceutical Sciences, University of Sao Paulo, Ribeirao Preto, Sao Paulo and 5 Department of Pharmacology, Federal University of Santa Catarina, Florianopolis, SC, Brazil. Corresponding author: Prof. Dr. Fernando Q. Cunha, Department of Pharmacology, Faculty of Medicine of Ribeirao Preto, University of Sao Paulo, Avenida Bandeirantes, 3900, 14049-900, Ribeirao Preto, Sao Paulo, Brazil. Tel. 55 16 3602 3324, Fax. 55 16 3633 2301. E-mail: [email protected] This work was supported by grants from the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Pesquisa e Desenvolvimento Tecnológico (CNPq) and Programa de Núcleos de Excelência (PRONEX). Running Head: H 2 S improves survival in sepsis Subject descriptor number: 4.12 Word count: 3764 This article has an online data supplement, which is accessible from this issue's table of contents online at www.atsjournals.org. Scientific Knowledge on the Subject During the inflammatory process, the cystathionine γ-lyase (CSE)/hydrogen sulfide (H 2 S) pathway has been shown to be involved in the control of leukocyte infiltration into inflammatory sites. However, the role of this pathway on the regulation of leukocyte recruitment to infection sites during sepsis has been not investigated. What This Study Adds to the Field H 2 S restores neutrophil migration to the infectious focus and improves survival outcome in severe sepsis by a K + ATP channel-dependent mechanism. Page 1 of 46 Media embargo until 2 weeks after above posting date; see thoracic.org/go/embargo AJRCCM Articles in Press. Published on March 25, 2010 as doi:10.1164/rccm.200907-1145OC Copyright (C) 2010 by the American Thoracic Society.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Hydrogen Sulfide Improves Neutrophil Migration and Survival in Sepsis via K+ATP Channel Activation

HYDROGEN SULFIDE IMPROVES NEUTROPHIL MIGRATION AND SURVIVAL IN
SEPSIS VIA K+
ATP CHANNEL ACTIVATION
Fernando Spiller1, Maria I.L. Orrico
1, Daniele C. Nascimento
1, Paula G. Czaikoski
1, Fabrício O.
Souto3, José C. Alves-Filho
1, Andressa Freitas
1, Daniela Carlos
1, Marcelo F. Montenegro
1,
Alberto F. Neto4, Sergio H. Ferreira
1, Marcos A. Rossi
2, John S. Hothersall
1, Jamil Assreuy
5 and
Fernando Q. Cunha1.
Departments of 1Pharmacology,
2Pathology, and
3Surgery and Anatomy, School of Medicine of
Ribeirao Preto; 4Department of Pharmaceutical Science, Faculty of Pharmaceutical Sciences,
University of Sao Paulo, Ribeirao Preto, Sao Paulo and 5Department of Pharmacology, Federal
University of Santa Catarina, Florianopolis, SC, Brazil.
Corresponding author: Prof. Dr. Fernando Q. Cunha, Department of Pharmacology, Faculty of Medicine of Ribeirao
Preto, University of Sao Paulo, Avenida Bandeirantes, 3900, 14049-900, Ribeirao Preto, Sao
Paulo, Brazil. Tel. 55 16 3602 3324, Fax. 55 16 3633 2301. E-mail: [email protected]
This work was supported by grants from the Fundação de Amparo à Pesquisa do Estado de São
Paulo (FAPESP), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES),
Conselho Nacional de Pesquisa e Desenvolvimento Tecnológico (CNPq) and Programa de
Núcleos de Excelência (PRONEX).
Running Head: H2S improves survival in sepsis
Subject descriptor number: 4.12
Word count: 3764
This article has an online data supplement, which is accessible from this issue's table of contents
online at www.atsjournals.org.
Scientific Knowledge on the Subject
During the inflammatory process, the cystathionine γ-lyase (CSE)/hydrogen sulfide (H2S)
pathway has been shown to be involved in the control of leukocyte infiltration into inflammatory
sites. However, the role of this pathway on the regulation of leukocyte recruitment to infection
sites during sepsis has been not investigated.
What This Study Adds to the Field
H2S restores neutrophil migration to the infectious focus and improves survival outcome in
severe sepsis by a K+
ATP channel-dependent mechanism.
Page 1 of 46 Media embargo until 2 weeks after above posting date; see thoracic.org/go/embargo
AJRCCM Articles in Press. Published on March 25, 2010 as doi:10.1164/rccm.200907-1145OC
Copyright (C) 2010 by the American Thoracic Society.

1
ABSTRACT
Rationale: Recovering the neutrophil migration to the infectious focus improves survival in
severe sepsis. Recently, we demonstrated that the cystathionine γ-lyase (CSE)/hydrogen sulfide
(H2S) pathway increased neutrophil recruitment to inflammatory focus during sterile
inflammation. Objectives and methods: Evaluate if H2S administration increases neutrophil
migration to infectious focus and survival of mice with sepsis induced by cecal ligation and
puncture (CLP). Results: The pre-treatments of mice with H2S donors (NaHS or Lawesson’s
reagent) improved leukocyte rolling/adhesion in the mesenteric microcirculation as well as
neutrophil migration. Consequently, bacteremia levels were reduced, hypotension and lung
lesions were prevented and the survival rate was enhanced from ~13% to ~80%. Notably, even
when treatment was delayed (6 h post-CLP), a highly significant reduction in mortality compared
to untreated mice was still observed. Moreover, H2S-pretreatment prevented the down-regulation
of CXCR2 and L-selectin and the up-regulation of CD11b and GRK2 in neutrophils during
sepsis. H2S also prevented the reduction of ICAM-1 expression in the endothelium of the
mesenteric microcirculation in severe sepsis. Confirming the critical role of H2S on sepsis
outcome, pretreatment with dl-propargylglycine (PAG, a CSE inhibitor) inhibited neutrophil
migration to the infectious focus, enhanced lung lesions and induced high mortality in mice
subjected to non-severe sepsis (from 0% to ~80%). Finally, the beneficial effects of H2S were
blocked by glibenclamide (a K+
ATP channel blocker). Conclusion: These results showed that H2S
restores neutrophil migration to the infectious focus and improves survival outcome in severe
sepsis by a K+
ATP channel-dependent mechanism.
Word count: 244
Keywords: neutrophil migration failure; sepsis; hydrogen sulfide; survival; cecal ligation and
puncture.
Page 2 of 46

2
INTRODUCTION
Sepsis is one of the major causes of death in intensive care units and has a mortality rate of
30 to 50%. Therapy for severe sepsis and septic shock is still largely symptomatic and supportive
(1).
We have shown that severe sepsis induced by cecal ligation and puncture (CLP) (2) or by
Gram-positive (3) or Gram-negative (4) intraperitoneal (ip) bacterial inoculation is associated
with inhibition of neutrophil recruitment to the sites of infection, bacteremia and high mortality
rates. Similarly, neutrophils isolated from non-surviving septic patients exhibited a suppressed
chemotactic response when compared to neutrophils from surviving septic patients or healthy
volunteers (5, 6). In investigating the mechanisms involved in the reduction of neutrophil
migration in severe experimental sepsis, we have shown that circulating cytokines and
chemokines and/or systemic TLR2/4 activation reduce the rolling, adhesion and chemotaxis of
neutrophils (7). The reduction of these events either in experimental or septic patients is due to
CXCR2 internalization and modulation of the levels of adhesion molecules on neutrophils (8-
10). The internalization of CXCR2 is associated with up-regulation of G protein-coupled
receptor kinase 2 (GRK2) expression in neutrophils (8, 11).
Hydrogen sulfide (H2S) is synthesized from L-cysteine mainly by cystathionine β-synthase
(CBS) and cystathionine γ-lyase (CSE) (12). The CSE/H2S pathway has been demonstrated to
play important roles in different biological systems. It modulates blood pressure (13) and
leukocyte trafficking in several inflammatory models such as acute pancreatitis (14), hindpaw
edema (15) and endotoxemia (16). In experimental sepsis induced by CLP, the CSE/H2S
pathway has also been shown to be involved in the control of leukocyte infiltration into distant
tissues such as lung and liver (17, 18).
Page 3 of 46

3
The role of the CSE/H2S pathway in regulating leukocyte recruitment to inflammatory sites
is not completely understood and sometimes contradictory. It has been shown that H2S donors
(NaHS and Na2S) inhibited aspirin-induced leukocyte adherence in mesenteric venules (19, 20).
However, Zhang et al. (17) have shown that the pretreatment of mice with dl-propargylglycine
(PAG), the CSE inhibitor, inhibited CLP-induced leukocyte-endothelial interactions in
mesenteric venules, whereas NaHS improved these parameters. In agreement with this, we have
shown that treatment of mice with H2S synthesis inhibitors reduced neutrophil migration into the
peritoneal cavities or femur/tibial joints of mice. Predictably, treatment of animals with H2S
donors enhanced these parameters (11). Therefore, our objective was to evaluate if exogenous
H2S increases neutrophil migration to the infectious focus and the survival of mice with sepsis
induced by CLP. Moreover, we also investigated whether the mechanism of action of H2S on
neutrophil migration was mediated via the ATP-dependent K+ (K
+ATP) channel pathway because
many of the other effects associated with H2S have been attributed to K+
ATP channel activation
(12).
MATERIALS AND METHODS
Experimental details are provided in the online supplement.
Word count: 500
Animals
The research was approved by the Animal Research Ethics Committee of the FMRP. Male
Swiss mice (23–26 g) were used.
Sepsis model
Page 4 of 46

4
Sepsis was induced by a CLP model, as described (21). We have standardized 2 punctures
in the cecum with a 26-gauge needle as non-severe (NS) sepsis induction or 2 punctures with an
18-gauge needle as severe (S) sepsis induction.
Neutrophil migration to the peritoneal cavity
Neutrophil migration was assessed 6 h after CLP, as reported (2).
Bacterial counts
The bacterial count was determined in the blood 6 h after CLP, as described (4).
Intravital microscopy
Leukocyte rolling and adhesion were examined in the mesenteric microcirculation by
intravital microscopy 2 h and 4 h after CLP, respectively, as described (22).
Cystathionine Gamma Lyase Assay
Total leukocytes were purified 2 h after CLP, and CSE activity measured as described (23).
Determination of Cytokine Levels in the Peritoneal Exudates and Blood
Animals were sacrificed 6 h after CLP, and the peritoneal lavage and blood were collected.
Cytokine concentrations were determined by ELISA.
Histological examination
The animals were sacrificed 12 h after CLP, and after isolation the lungs were fixed by
immersion in 5% paraformaldehyde, dehydrated and embedded in paraffin wax. Sections of 3-
µm thickness were stained with hematoxylin and eosin for histological examination. A
pathologist performed blind histological assessment.
Isolation of bone marrow neutrophils
Bone marrow cells were collected, and neutrophils were isolated by differential
centrifugation on a Percoll gradient, as described (8).
Page 5 of 46

5
Chemotaxis Assays
Mouse bone marrow neutrophils were incubated in the absence (control) or in the presence
of glibenclamide (GLB, a K+
ATP channel blocker) or the H2S donor sodium hydrogen sulfite
(NaHS, 30, 100 or 300 µM). In addition, one group was incubated (37°C) with GLB (100 µM,
30 min) and then NaHS (300 µM) for 1 h. Following this, neutrophil chemotaxis was stimulated
by MIP-2 in a Boyden chamber.
Flow Cytometry Analysis of CD11b, CD62L and CXCR2 expression
Flow cytometry was analyzed with a flow cytometer (BD FACSort; Mountain View, CA)
according to a previously described procedure (9). Total blood was collected 6 h after CLP and
stained with labeled antibodies: PE-anti-CXCR2 (R&D Systems), PE-anti-CD62L, FITC-CD11b
and PerCP-anti-Gr-1 (BD Biosciences).
Immunofluorescence assay for ICAM-1/CD54 or G protein-coupled receptor kinase 2
(GRK2)
Six hours after CLP, an ICAM-1/CD54 immunofluorescence assay was performed in
mesenteric and lung tissues or GRK2 in neutrophils according to a previously described method
(11).
Statistical analysis
The data were reported as the mean ± SEM of values obtained from two or three
independent experiments (n = 5 in each experiment; exception for survival analyses, n = 10-12).
The means between different groups were compared by analysis of variance (ANOVA). If
significance was determined, individual comparisons were subsequently tested with Bonferroni's
t test for unpaired values. Moreover, the Kolmogorov-Smirnov test was employed to test for
normality. Because the rolling and adhesion results failed to reach normality, the data were log
Page 6 of 46

6
transformed to reach this assumption, and then ANOVAs were performed with the modified
data.
RESULTS
Hydrogen sulfide protects against CLP-induced lethality
Firstly, we evaluated CSE activity in blood leukocyte extracts 2 hours after sepsis induction
by CLP. Fig. 1A shows that in mice subjected to severe (S) sepsis, there was significantly
increased CSE activity when compared to sham-operated (Sham) mice, and PAG (1 mM)
abrogated this activity.
Following this, the effect of H2S-synthesis inhibition on survival during NS sepsis (animals
experiencing local infection) was determined (Fig. 1B). Mice pretreated with PAG (50 mg/kg,
ip) and subjected to NS sepsis exhibited a significant reduction (~80%) in survival rate when
compared with NS animals pretreated with saline. Moreover, PAG treatment in mice subjected to
moderate sepsis (~50% mortality rate) also increased the mortality rate (~100%, data not shown).
Furthermore, animals submitted to S sepsis and pretreated with saline or PAG failed to survive
beyond the second day after CLP. All Sham mice survived for 8 days after CLP (data not
shown).
In addition, PAG pretreatment inhibited neutrophil migration to the focus of infection (Fig.
2A), increased blood bacterial counts (Fig. 2B) and the levels of serum TNF-α, MIP-2 and KC
(Table E1) in NS septic mice. As shown in Fig. 2A, pretreatment of S septic mice with PAG did
not significantly change the number of neutrophils in the peritoneal cavity.
The effects of exogenous H2S on the survival rate of mice subjected to S sepsis were then
investigated. The sc pretreatment of mice subjected to S sepsis with 100 µmol/kg of NaHS (Fig.
Page 7 of 46

7
1C) or Lawesson’s reagent (Fig. 1E) improved the survival rate from 10-15% to ~80% 8 days
after CLP. The therapeutic treatment of S CLP mice with these H2S donors 6, 12, and 24 hours
post S sepsis induced a delay in mortality and a significant increase in the survival rate (~35%;
Fig. 1D and 1F).
As previously demonstrated (6), mice subjected to S sepsis showed a significantly reduced
neutrophil migration compared to the NS group (Fig. 2A). The sc pretreatment of mice subjected
to S sepsis with 100 µmol/kg of H2S donors prevented the failure of neutrophil migration to the
focus of infection (Fig. 2A), reduced blood bacterial count (Fig. 2B) and prevented hypotension
(Fig. E2B).
The sc administration of NaHS at a dose of 100 µmol/kg in naive mice did not induce
significant changes in mean arterial pressure (MAP, Fig. E2A). However, in accordance with
previous descriptions (24), the intravenous bolus injection of the same NaHS dose either in naive
(Fig. E2A) or septic mice (data not shown) provoked a significant decline in MAP compared to
saline injected mice.
Using a sulfide-sensitive electrode, we confirmed that plasma H2S levels were increased
after experimentally induced severe sepsis (Fig. E1A). Moreover, NaHS and Lawesson’s reagent
were able to release H2S in a concentration-dependent manner in vitro (data not shown). In
addition, the plasma levels of H2S increased significantly in mice sc treated with 1000 µmol/kg
NaHS (Fig. E1B).
Leukocyte rolling/adhesion to mesenteric microcirculation is improved by NaHS
PAG pretreatment of NS septic mice increased TNF-α and MIP-2 levels in peritoneal
exudates when compared to Sham or NS septic mice (Fig. E3A and E3B, respectively). The
levels of KC were significantly increased in S, NS and NS-PAG-pretreated septic mice when
Page 8 of 46

8
compared to Sham mice (Fig. E3C). Treatment with NaHS did not change the levels of these
cytokines in the peritoneal cavities of S septic mice (Fig. E3). These data strongly suggested that
the reduction of neutrophil migration to the site of infection after PAG treatment or in S sepsis
was not a consequence of low concentrations of cytokines and chemokines in the peritoneal
cavity. However, treatment with PAG inhibited the significant increase in rolling and adhesion of
leukocyte to endothelial cells of mesenteric venules in NS septic mice (Fig. 2C). This reduction
in leukocyte rolling and adhesion was also observed in S septic mice. Interestingly, when S mice
were pretreated with NaHS, there was a significant improvement in leukocyte rolling and
adhesion (Fig. 2C).
Hydrogen sulfide prevents acute lung injury in sepsis
Sepsis led to significant changes in pulmonary histology consistent with acute lung injury.
This included septal thickening, edema and congestion, hemorrhage and both neutrophil and
mononuclear cell accumulation. Animals submitted to S sepsis (Fig. E4E) showed more
pronounced changes in the lungs compared with the lungs of mice submitted to NS septic injury
(Fig. E4C). Following PAG treatment, these same groups presented significant aggravation of
lung alterations (Fig. E4F and E4D). Pretreatment of mice submitted to S sepsis with NaHS
significantly abrogated the development of acute lung injury (Fig. E4B), and the lung structure of
these mice appeared similar to the normal structure observed in Sham mice (Fig. E4A), except
for discrete interstitial neutrophil and mononuclear cell infiltration and edema.
KATP channel blocking prevents protective effects of H2S during sepsis
We evaluated if H2S improves neutrophil migration in a KATP channel-dependent fashion
using both in vivo and in vitro models. Bone marrow neutrophils incubated for 1 h (37°C) with
NaHS (300 µM) showed a significant increase (P < 0.001) in MIP-2-induced neutrophil
Page 9 of 46

9
chemotaxis when compared to the RPMI control group (Fig. 3A). When neutrophils were pre-
incubated with glibenclamide (GLB 100 µM, a selective KATP channel blocker), the MIP-2-
induced chemotaxis and the NaHS enhancement of MIP-2-induced chemotaxis were blocked
(Fig. 3A).
In vivo, pretreatment with GLB (40 µmol/kg, sc) inhibited neutrophil migration (Fig. 3B)
and increased mortality (Fig. 3C) of mice subjected to NS sepsis. Moreover, GLB treatment prior
to NaHS or Lawesson’s reagent treatment inhibited the improvement of neutrophil migration
(Fig. 3B) and survival (Fig. 3C) evoked by these H2S donors in S septic mice.
Because CD11b, CD62L and CXCR2 expression in leukocytes is necessary for neutrophil
rolling, adhesion and migration to the focus of infection (9), the expression of these receptors on
blood neutrophils was evaluated 6 h after CLP. S septic mice had significantly (P < 0.05)
decreased CD62L (Fig. 3E) and CXCR2 (Fig. 3F) expression, while CD11b (Fig. 3D) expression
increased in neutrophils when compared to neutrophils from NS or Sham mice. In contrast,
pretreatment of S septic animals with Lawesson’s reagent partially inhibited the down-regulation
of CD62L and CXCR2 and the up-regulation of CD11b in neutrophils. These effects of the H2S
donor were prevented by GLB pretreatment. Furthermore, when NS mice were pretreated with
PAG or GLB, the neutrophil expression of CD62L (Fig. E5B) and CXCR2 (Fig. E5C) was
significantly lower when compared to neutrophils from NS mice. PAG pretreatment of NS mice
also significantly raised the expression of CD11b, whereas GLB had no significant effect (Fig.
E5A).
High expression of GRK2 is involved in the internalization of chemokine receptors in
leukocytes (8). Six hours after severe sepsis induction, we observed a significant augmentation
of GRK2 expression in blood neutrophils (Fig. 4E). Pretreatment of NS mice with PAG (Fig. 4C)
Page 10 of 46

10
or GLB (Fig. 4D) enhanced, whereas Lawesson’s reagent (Fig. 4F) pretreatment reduced, S
sepsis-induced GRK2 expression. Moreover, GLB prevented the inhibitory effect of H2S on
GRK2 expression (Fig. 4G). These results suggest that H2S prevents CXCR2 internalization via
inhibition of GRK2 expression by a KATP channel-dependent mechanism.
H2S prevented severe sepsis-induced ICAM-1 down-modulation by KATP channel activation
To further clarify the mechanisms by which H2S increased neutrophil recruitment to an
infectious focus, we evaluated ICAM-1 levels in the endothelium of mesenteric microcirculation
after sepsis induction. Compared with Sham animals (Fig. 5A), NS sepsis (Fig. 5B) induction
increased ICAM-1 expression in mesenteric vessels, whereas pretreatment of these mice with
PAG (Fig. 5C) or GLB (Fig. 5D) or S sepsis induction (Fig. 5E) inhibited ICAM-1 expression.
Pretreatment of S septic mice with NaHS significantly enhanced ICAM-1 expression in
endothelial cells (Fig. 5F), and GLB blocked this increase in ICAM-1 expression (Fig. 5G).
We also evaluated the levels of ICAM-1 expression in lung vessels after sepsis induction.
Similar to mesenteric vessels, NS sepsis induced a significantly rise in ICAM-1 expression in
lung vessels, which was inhibited by PAG pretreatment. In contrast, S sepsis reduced ICAM-1
expression and pretreatment with NaHS resulted in a significant increase in ICAM-1 expression
levels (Fig. E6). These data are in accordance with previous reports showing that there is an up-
regulation of ICAM-1 in the lungs during sepsis and that PAG treatment is able to prevent this
increase, while H2S donors increase the ICAM-1 expression (17).
DISCUSSION
In the present study, we demonstrate that pretreatment of mice submitted to severe sepsis
with H2S donors prevented the failure of neutrophil migration, reduced bacterial spreading,
Page 11 of 46

11
prevented hypotension and improved the survival of the mice from ~12% to ~80%. Furthermore,
and more clinically relevant, the delayed treatment of mice with these donors improved the
survival rate by up to ~35%.
The CSE H2S-forming enzyme activity and mRNA levels have been reported as increased
in the liver and kidneys after endotoxin- or CLP-induced sepsis (16, 18). However, the role of
H2S in systemic inflammatory response syndrome (SIRS) is not understood. There is evidence
that the inhibition of H2S synthesis by PAG decreases leukocyte infiltration in the liver and/or
lungs after acute pancreatitis or endotoxemia induction (16, 25). Moreover, Zhang et al. (18)
have shown that administration of PAG at the same dose and route as those used in the present
study (50 mg/kg, ip) significantly reduced leukocyte accumulation in the lungs and livers of
CLP-induced septic animals and attenuated the mortality of animals. Subsequently, the same
group showed that treatment of CLP-septic mice with PAG inhibited rolling and adhesion of
leukocytes to the mesenteric microcirculation (17). Rolling and adhesion are critical steps for
neutrophil migration from the bloodstream into tissues during the inflammatory process (26).
Therefore, it is reasonable to conclude that the reduction of rolling and adhesion of leukocytes in
the mesenteric microcirculation also reduces neutrophil migration into the peritoneal cavity in
CLP-induced sepsis (focus of the infection). In fact, herein we showed that the significant
reduction of leukocyte rolling and adhesion on the mesenteric microcirculation induced by
inhibiting H2S biosynthesis resulted in the reduction of neutrophil migration to the focus of
infection. Therefore, the decrease in available H2S allowed bacterial spread, increased systemic
inflammatory response (measured by cytokines TNF-α, MIP-2, KC in serum), and contrary to
the findings of Zhang’s group (23), aggravated lung injury and induced higher mortality. The
data emphasize the importance of CSE activity and consequent H2S production in infection
Page 12 of 46

12
control and host survival. Thus, the rise of H2S production observed in severe sepsis might be a
protective mechanism triggered by the host aimed at enhancing neutrophil migration to the focus
of infection in an attempt to confine and combat the infection. Similarly, we observed a
significant increase in CSE activity in blood leukocytes and plasma H2S levels after severe sepsis
induction. However, it would appear that the amount of H2S generated in this condition is not
sufficient to protect against mortality. Thus, when we administered pharmacological
concentrations of H2S to S septic animals, they presented 80% survival, reinforcing our
hypothesis that the level of endogenously produced H2S was insufficient in this
pathophysiological condition. Experimental differences, including health status, age and gender
of the animals, commercial source of PAG and level of severity of sepsis, may explain the
opposing effects of CSE inhibition on survival and lung injury of mice exposed to CLP-induced
sepsis, being the deleterious effects described in the present study and the protective effects
described by Zhang´s group (18).
Typically, migration of neutrophils to a site of infection involves rolling on activated
endothelium (mediated by selectins) and subsequently firm adherence to the endothelium via
interaction of activated neutrophil integrins with the endothelial cell intercellular adhesion
molecules (ICAMs) (27). Here, we observed that mice subjected to NS sepsis, which had
efficient rolling/adhesion and neutrophil migration, expressed CD62L, CD11b and ICAM-1 at
similar levels to that found in Sham mice, whereas mice with S sepsis, which presented failure of
neutrophil migration, had a significant shedding of CD62L, high expression of CD11bin
circulating neutrophils and low expression of ICAM-1 in the mesenteric endothelium (28, 29).
Treatment with H2S donors prevented L-selectin shedding, down-modulation of ICAM-1 and the
increase of CD11b during S sepsis. Consequently, mice treated with H2S donors presented
Page 13 of 46

13
leukocyte rolling and adherence on the endothelium similar to that observed in mice with NS
sepsis.
The microbicidal activity of the migrating cells is fundamental to the containment of
infection (30). In view of this, we observed that both in vivo and in vitro treatments with PAG or
H2S donors did not change neutrophil phagocytic or killing capacities. Moreover, H2S did not
show antimicrobial activity as a single agent (data not shown). These results reinforce that the
beneficial effects of H2S on S sepsis are mainly the result of the reestablishment of neutrophil
migration to the focus of infection.
As discussed above, the inability of the host to restrict the infection locally triggers a
systemic inflammatory process, characterized, among other events, by the accumulation of
leukocytes into peripheral organs including the lungs, which contributes to tissue lesion and
overall organ dysfunction (31). Accordingly, treatments of the animals with an H2S donor or
PAG reduced and increased the lung lesion, respectively. Interestingly, H2S treatment of severe
septic mice was able to increase ICAM-1 expression in lung vessels. However, in this group,
there was a dramatic reduction in leukocyte infiltration. The direct explanation for this apparent
contradiction is that H2S treatment also reduced the increase of CD11b expression in the
neutrophils; which is necessary to trigger neutrophil accumulation in the lung and other organs
during systemic inflammation (32).
The aggravation of sepsis syndrome correlates with progressive hypotension that is
unresponsive to vasoconstrictors (33). Several studies have reported an important role of H2S in
the cardiovascular system (12). Genetic deletion of CSE increased blood pressure, whereas
intravenous bolus injection of H2S reduced blood pressure dose-dependently (2.8-39 µmol/Kg)
(13, 24). In our experiments, we also observed that intravenous bolus administration of NaHS
decreased blood pressure. On the other hand, subcutaneous administration of NaHS did not
Page 14 of 46

14
reduce the mean arterial pressure of naïve mice; however, it prevented CLP-induced
hypotension. This valuable effect of H2S on hypotension is unlikely to be due to a direct effect
on the vasculature. Probably, it is a consequence of the reestablishment of infection control
evoked by H2S in the septic animals. Alternatively, it is possible that there is an interaction of
H2S with the NO/iNOS system, because during endotoxic shock H2S was able to prevent the
expression of iNOS and, consequently, LPS-induced hypotension (34).
The CXC-ELR+ chemokines and their receptors are key factors for host defense against
infection (35, 36). In this context, CXCR2 antagonists, which block neutrophil influx, are shown
to increase septic lethality (9). Interestingly, during S sepsis, the down-regulation of CXCR2 in
blood neutrophils was associated with reduction of adhesion and chemotaxis of leukocytes (8, 9,
37). GRK2 has been implicated in the down-regulation of chemokine receptors (CXCRs and
CCRs) (8, 11, 38). Here, we have shown that the treatment of mice subjected to S sepsis with
H2S donors was able to prevent up-regulation of GRK2 expression and down-regulation of
CXCR2 in blood neutrophils. These data are in accordance with previous reports that CXCR2 is
up-regulated in blood neutrophils from naïve mice after NaHS treatment (17).
Studies have reported that several pharmacological effects of H2S are related to the opening
of KATP channels (12). In the present work, we have shown that the H2S-stimulated improvement
in survival during sepsis is mediated by KATP channel activation. KATP channel activity has
already been shown to be important to neutrophil migration. In fact, treatments of animals with
inhibitors or openers of KATP channels inhibit and enhance neutrophil migration in different
experimental models, respectively (39). In the present study, it was observed that GLB treatment
mimicked the action of PAG and prevented the effects of H2S on neutrophil migration, CXCR2
internalization and GRK2 or ICAM-1 expression. Taken together, our results suggest that the
effect of H2S in sepsis is mediated by KATP channel activation. In this context, Tang et al. (40),
Page 15 of 46

15
using a whole-cell and single-channel patch-clamp technique, demonstrated that exogenous H2S
activated KATP channels and hyperpolarized cell membranes in rat mesenteric artery vascular
smooth muscle cells. Inhibition of endogenous H2S production with PAG also reduced whole-
cell KATP currents. Moreover, H2S modifies a large number of proteins, including actin and the
Kir6.1 subunit of the KATP channel, by S-Sulfhydration, a process that may regulate their
function (41).
In summary, our data show a critical role of endogenous H2S production in efficient
neutrophil migration to the infectious focus in NS sepsis. Additionally, exogenous H2S prevents
the failure of neutrophil migration to the focus of infection, and both pre- and post-treatment
ultimately improved the survival in S sepsis. In this context, H2S post-treatment has also been
shown to significantly decrease mortality in a murine model of acute lung injury (42). Therefore,
H2S donors could be important agents in a new approach toward sepsis treatment.
Acknowledgments
We thank GB Francisco for technical assistance.
REFERENCES
1. Angus DC, Linde-Zwirble WT, Lidicker J, Clermont, Pinsky MR. Epidemiology of
severe sepsis in the united states: Analysis of incidence, outcome, and associated costs of care.
Crit Care Med 2001;29:1303-1310.
2. Benjamim CF, Ferreira SH, Cunha FQ. Role of nitric oxide in the failure of neutrophil
migration in sepsis. J Infect Dis 2000;182:214-223.
3. Crosara-Alberto DP, Darini AL, Inoue RY, Silva JS, Ferreira SH, Cunha FQ.
Involvement of no in the failure of neutrophil migration in sepsis induced by staphylococcus
aureus. Br J Pharmacol 2002;136:645-658.
Page 16 of 46

16
4. Alves-Filho JC, de Freitas A, Russo M, Cunha FQ. Toll-like receptor 4 signaling leads to
neutrophil migration impairment in polymicrobial sepsis. Crit Care Med 2006;34:461-470.
5. Tavares-Murta BM, Zaparoli M, Ferreira RB, Silva-Vergara ML, Oliveira CH, Murta EF,
Ferreira SH, Cunha FQ. Failure of neutrophil chemotactic function in septic patients. Crit Care
Med 2002;30:1056-1061.
6. Arraes SM, Freitas MS, da Silva SV, de Paula Neto HA, Alves-Filho JC, Martins MA,
Basile-Filho A, Tavares-Murta BM, Barja-Fidalgo C, Cunha FQ. Impaired neutrophil
chemotaxis in sepsis associates with grk expression and inhibition of actin assembly and tyrosine
phosphorylation. Blood 2006;108:2906-2913.
7. Alves-Filho JC, de Freitas A, Spiller F, Souto FO, Cunha FQ. The role of neutrophils in
severe sepsis. Shock 2008;30 Suppl 1:3-9.
8. Alves-Filho JC, Freitas A, Souto FO, Spiller F, Paula-Neto H, Silva JS, Gazzinelli RT,
Teixeira MM, Ferreira SH, Cunha FQ. Regulation of chemokine receptor by toll-like receptor 2
is critical to neutrophil migration and resistance to polymicrobial sepsis. Proc Natl Acad Sci U S
A 2009;106:4018-4023.
9. Rios-Santos F, Alves-Filho JC, Souto FO, Spiller F, Freitas A, Lotufo CM, Soares MB,
Dos Santos RR, Teixeira MM, Cunha FQ. Down-regulation of cxcr2 on neutrophils in severe
sepsis is mediated by inducible nitric oxide synthase-derived nitric oxide. Am J Respir Crit Care
Med 2007;175:490-497.
10. Cummings CJ, Martin TR, Frevert CW, Quan JM, Wong VA, Mongovin SM, Hagen TR,
Steinberg KP, Goodman RB. Expression and function of the chemokine receptors cxcr1 and
cxcr2 in sepsis. J Immunol 1999;162:2341-2346.
11. Dal-Secco D, Cunha TM, Freitas A, Alves-Filho JC, Souto FO, Fukada SY, Grespan R,
Alencar NM, Neto AF, Rossi MA, et al. Hydrogen sulfide augments neutrophil migration
through enhancement of adhesion molecule expression and prevention of cxcr2 internalization:
Role of atp-sensitive potassium channels. J Immunol 2008;181:4287-4298.
12. Szabo C. Hydrogen sulphide and its therapeutic potential. Nat Rev Drug Discov
2007;6:917-935.
13. Yang G, Wu L, Jiang B, Yang W, Qi J, Cao K, Meng Q, Mustafa AK, Mu W, Zhang S, et
al. H2s as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine
gamma-lyase. Science 2008;322:587-590.
14. Bhatia M, Wong FL, Fu D, Lau HY, Moochhala SM, Moore PK. Role of hydrogen
sulfide in acute pancreatitis and associated lung injury. Faseb J 2005;19:623-625.
15. Bhatia M, Sidhapuriwala J, Moochhala SM, Moore PK. Hydrogen sulphide is a mediator
of carrageenan-induced hindpaw oedema in the rat. Br J Pharmacol 2005;145:141-144.
16. Li L, Bhatia M, Zhu YZ, Zhu YC, Ramnath RD, Wang ZJ, Anuar FB, Whiteman M,
Salto-Tellez M, Moore PK. Hydrogen sulfide is a novel mediator of lipopolysaccharide-induced
inflammation in the mouse. Faseb J 2005;19:1196-1198.
17. Zhang H, Zhi L, Moochhala SM, Moore PK, Bhatia M. Endogenous hydrogen sulfide
regulates leukocyte trafficking in cecal ligation and puncture-induced sepsis. J Leukoc Biol
2007;82:894-905.
18. Zhang H, Zhi L, Moore PK, Bhatia M. Role of hydrogen sulfide in cecal ligation and
puncture-induced sepsis in the mouse. Am J Physiol Lung Cell Mol Physiol 2006;290:L1193-
1201.
19. Fiorucci S, Antonelli E, Distrutti E, Rizzo G, Mencarelli A, Orlandi S, Zanardo R, Renga
B, Di Sante M, Morelli A, et al. Inhibition of hydrogen sulfide generation contributes to gastric
injury caused by anti-inflammatory nonsteroidal drugs. Gastroenterology 2005;129:1210-1224.
Page 17 of 46

17
20. Zanardo RC, Brancaleone V, Distrutti E, Fiorucci S, Cirino G, Wallace JL. Hydrogen
sulfide is an endogenous modulator of leukocyte-mediated inflammation. Faseb J 2006;20:2118-
2120.
21. Wichterman KA, Baue AE, Chaudry IH. Sepsis and septic shock--a review of laboratory
models and a proposal. J Surg Res 1980;29:189-201.
22. Benjamim CF, Silva JS, Fortes ZB, Oliveira MA, Ferreira SH, Cunha FQ. Inhibition of
leukocyte rolling by nitric oxide during sepsis leads to reduced migration of active microbicidal
neutrophils. Infect Immun 2002;70:3602-3610.
23. Steegborn C, Clausen T, Sondermann P, Jacob U, Worbs M, Marinkovic S, Huber R,
Wahl MC. Kinetics and inhibition of recombinant human cystathionine gamma-lyase. Toward
the rational control of transsulfuration. J Biol Chem 1999;274:12675-12684.
24. Zhao W, Zhang J, Lu Y, Wang R. The vasorelaxant effect of h(2)s as a novel endogenous
gaseous k(atp) channel opener. Embo J 2001;20:6008-6016.
25. Collin M, Anuar FB, Murch O, Bhatia M, Moore PK, Thiemermann C. Inhibition of
endogenous hydrogen sulfide formation reduces the organ injury caused by endotoxemia. Br J
Pharmacol 2005;146:498-505.
26. Simon SI, Green CE. Molecular mechanics and dynamics of leukocyte recruitment during
inflammation. Annu Rev Biomed Eng 2005;7:151-185.
27. Smith CW. Endothelial adhesion molecules and their role in inflammation. Can J Physiol
Pharmacol 1993;71:76-87.
28. Turunen R, Andersson S, Nupponen I, Kautiainen H, Siitonen S, Repo H. Increased
cd11b-density on circulating phagocytes as an early sign of late-onset sepsis in extremely low-
birth-weight infants. Pediatr Res 2005;57:270-275.
29. Seidelin JB, Nielsen OH, Strom J. Soluble l-selectin levels predict survival in sepsis.
Intensive Care Med 2002;28:1613-1618.
30. Chertov O, Yang D, Howard OM, Oppenheim JJ. Leukocyte granule proteins mobilize
innate host defenses and adaptive immune responses. Immunol Rev 2000;177:68-78.
31. Cohen J. The immunopathogenesis of sepsis. Nature 2002;420:885-891.
32. Doerschuk CM. Leukocyte trafficking in alveoli and airway passages. Respir Res
2000;1:136-140.
33. Parrillo JE. Pathogenetic mechanisms of septic shock. N Engl J Med 1993;328:1471-
1477.
34. Li L, Salto-Tellez M, Tan CH, Whiteman M, Moore PK. Gyy4137, a novel hydrogen
sulfide-releasing molecule, protects against endotoxic shock in the rat. Free Radic Biol Med
2009.
35. Zabel BA, Zuniga L, Ohyama T, Allen SJ, Cichy J, Handel TM, Butcher EC.
Chemoattractants, extracellular proteases, and the integrated host defense response. Exp Hematol
2006;34:1021-1032.
36. Mehrad B, Strieter RM, Moore TA, Tsai WC, Lira SA, Standiford TJ. Cxc chemokine
receptor-2 ligands are necessary components of neutrophil-mediated host defense in invasive
pulmonary aspergillosis. J Immunol 1999;163:6086-6094.
37. Quaid GA, Cave C, Robinson C, Williams MA, Solomkin JS. Preferential loss of cxcr-2
receptor expression and function in patients who have undergone trauma. Arch Surg
1999;134:1367-1371; discussion 1371-1362.
38. Prossnitz ER, Kim CM, Benovic JL, Ye RD. Phosphorylation of the n-formyl peptide
receptor carboxyl terminus by the g protein-coupled receptor kinase, grk2. J Biol Chem
1995;270:1130-1137.
Page 18 of 46

18
39. Da Silva-Santos JE, Santos-Silva MC, Cunha Fde Q, Assreuy J. The role of atp-sensitive
potassium channels in neutrophil migration and plasma exudation. J Pharmacol Exp Ther
2002;300:946-951.
40. Tang G, Wu L, Liang W, Wang R. Direct stimulation of k(atp) channels by exogenous
and endogenous hydrogen sulfide in vascular smooth muscle cells. Mol Pharmacol
2005;68:1757-1764.
41. Mustafa AK, Gadalla MM, Sen N, Kim S, Mu W, Gazi SK, Barrow RK, Yang G, Wang
R, Snyder SH. H2s signals through protein s-sulfhydration. Sci Signal 2009;2:ra72.
42. Esechie A, Kiss L, Olah G, Horvath EM, Hawkins H, Szabo C, Traber DL. Protective
effect of hydrogen sulfide in a murine model of acute lung injury induced by combined burn and
smoke inhalation. Clin Sci (Lond) 2008;115:91-97.
FIGURES LEGENDS
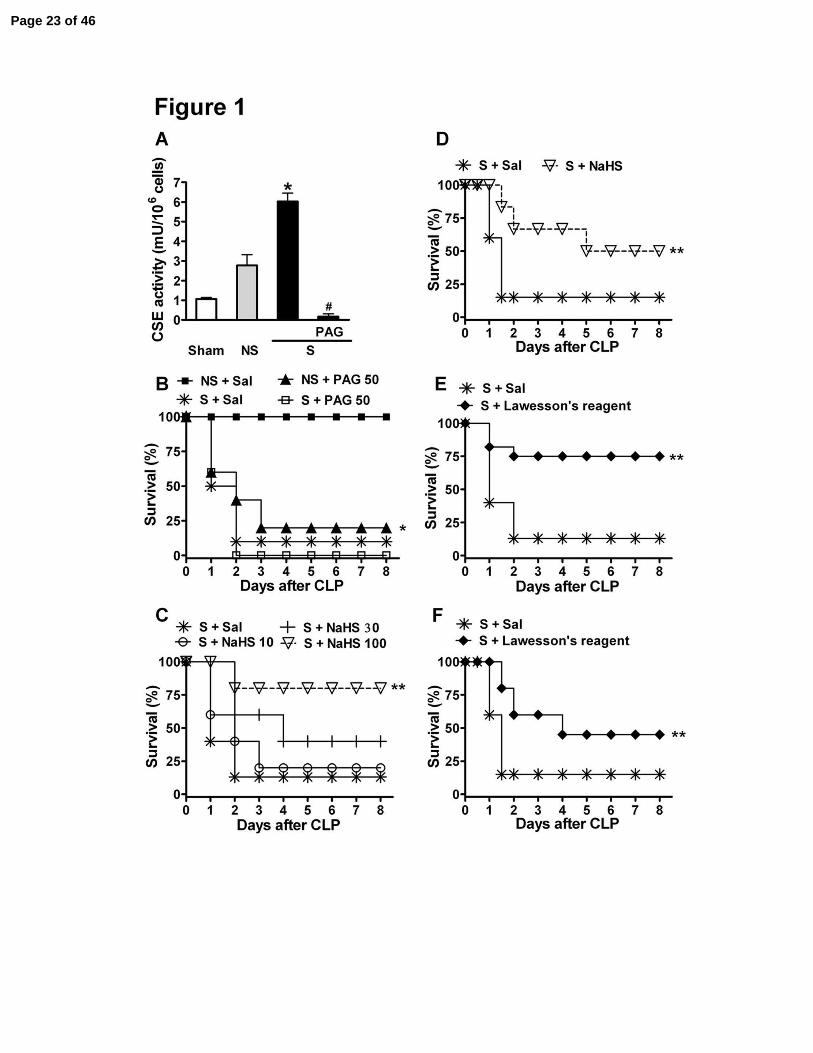
Figure 1. Survival rate deterioration with CSE inhibitor dl-propargylglycine (PAG) and
improvement with hydrogen sulfide donors (NaHS and Lawesson’s reagent) in mice
subjected to sepsis. (A) CSE activity was evaluated in blood leukocytes 2 h after surgery in
mice subjected to sham-operation (Sham), non-severe (NS) or severe (S) sepsis by cecal ligation
and puncture (CLP). CSE activity was also evaluated in blood leukocytes from S septic mice in
the presence of PAG (1 mM). *P < 0.01 compared to the Sham group and #P < 0.01 compared to
the S group (ANOVA, followed by Bonferroni’s test). One hour before induction of Sham, NS or
S sepsis, mice were given saline (Sal), (B) PAG (50 mg/kg, ip), (C) NaHS (10, 30 or 100
µmoles/kg, sc) or (E) Lawesson’s reagent (100 µmoles/kg, sc). Six h, twelve h and twenty-four h
after S sepsis induction, previously untreated mice were randomly given (D) NaHS (100
µmol/kg, sc), (F) Lawesson’s reagent (100 µmoles/kg, sc) or Sal (sc). Their survival rates were
monitored over 8 days. The results are expressed as percent of survival. *P < 0.05 compared to
NS plus Sal, and **P < 0.05 compared to S plus Sal. Mantel-Cox log-rank test (n = 12–20).
Page 19 of 46

19
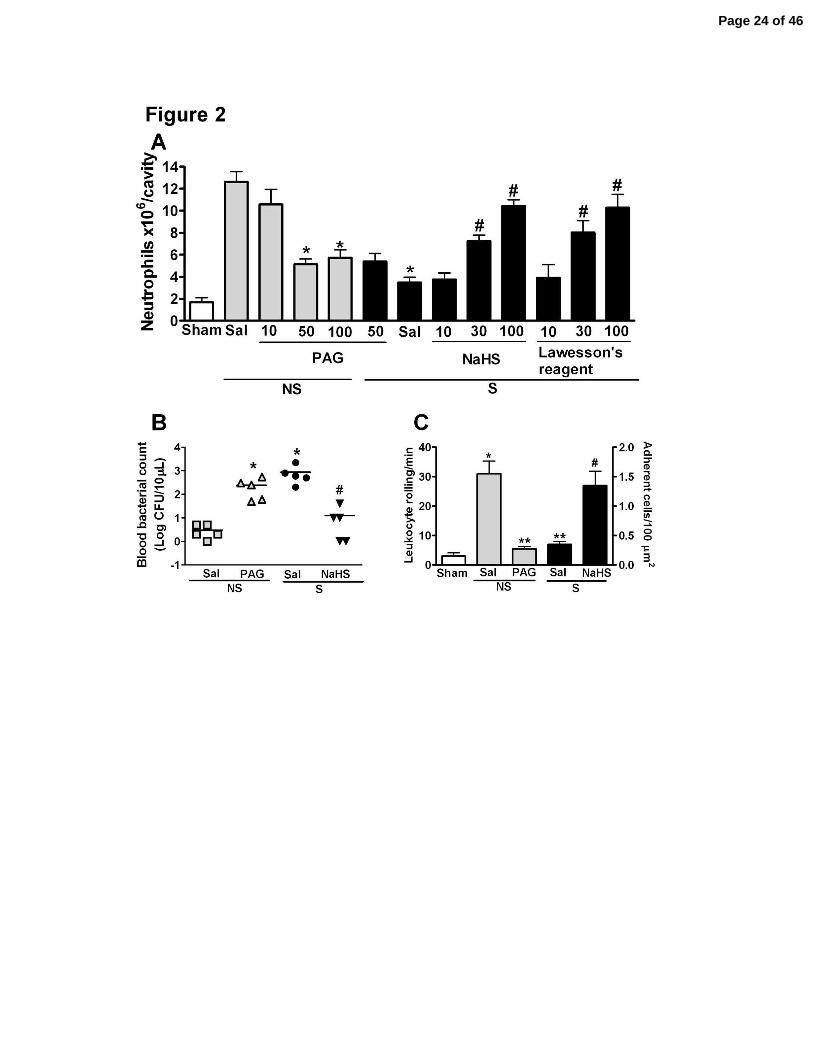
Figure 2. The H2S donors (NaHS and Lawesson’s reagent) prevented the failure of
neutrophil migration to the infectious focus, while PAG inhibited neutrophil migration in
mice subjected to sepsis. One hour before induction of sham-operation (Sham), non-severe (NS)
or severe (S) sepsis, mice were given PAG (10, 50 or 100 mg/kg, ip), NaHS (10, 30 or 100
µmoles/kg, sc), Lawesson’s reagent (10, 30 or 100 µmoles/kg, sc) or saline (Sal). (A) Six hours
after surgery, neutrophil migration into the peritoneal cavity was determined. The data are
reported as the mean ± SEM x 106 neutrophils/cavity. *P < 0.001 compared to the NS plus Sal
group and #P < 0.001 compared to the S plus Sal group. (ANOVA, followed by Bonferroni’s
test, n = 5). (B) Bacterial counts in the blood of mice pretreated with PAG (50 mg/kg, ip), NaHS
(100 µmol/kg, sc) or Sal were determined 6 h after CLP. The data are reported as the mean of the
Log of CFU per 10 µL. *P < 0.01 compared to NS plus Sal; #P < 0.01 compared to the group S
plus Sal (t test, followed by Mann-Whitney U test, n = 5). (C) Animals were treated with Sal,
PAG (50 mg/kg, ip) or NaHS (100 µmol/kg, sc) 1 hour before Sham, NS or S sepsis induction.
Rolling and adherent leukocytes in the mesenteric microcirculation were evaluated 2 h and 4 h
after surgery, respectively. The data are reported as the mean ± SEM of leukocyte rolling per min
and adherent cells per 100 µm2 of vessel. *P < 0.05 compared to Sham; **P < 0.05 compared to
NS plus Sal; and #P < 0.05 compared to S plus Sal (ANOVA, followed by Bonferroni’s test, n =
5). ND = not detectable.
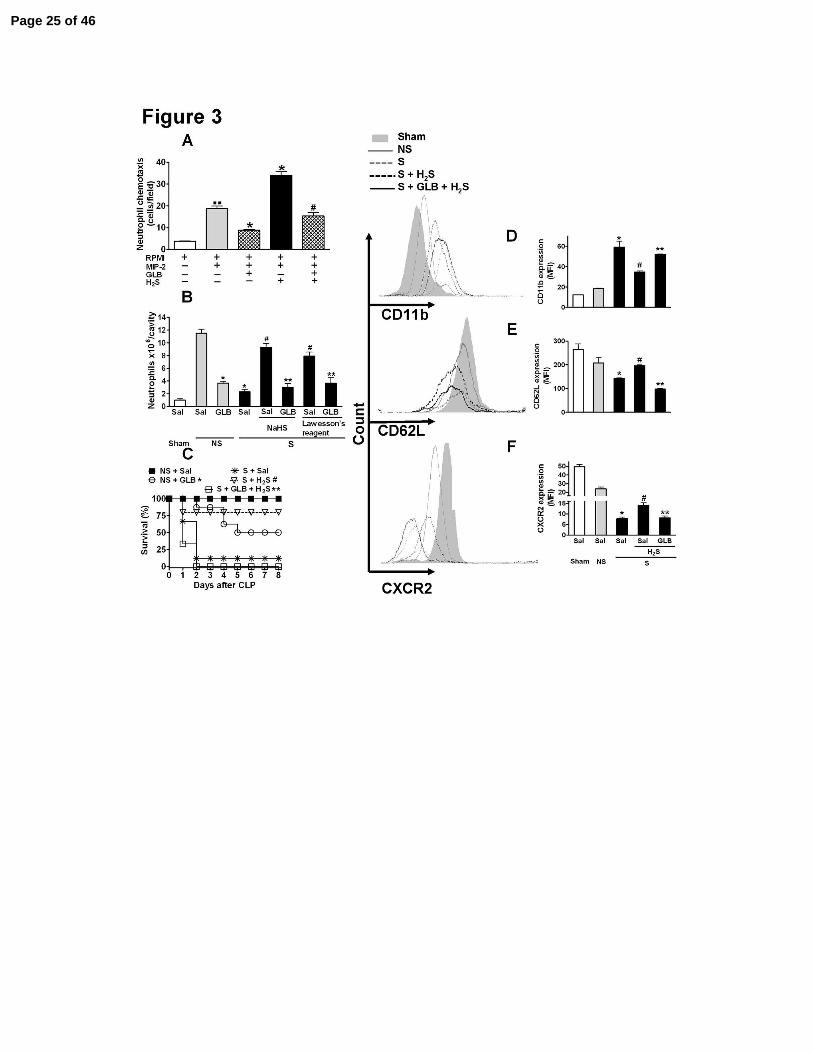
Figure 3. KATP channel blockage prevented the H2S effects in vivo and in vitro. (A) Isolated
neutrophils were incubated (1 h) with H2S, (NaHS, 300 µM), glibenclamide (GLB 100 µM) or
GLB (100 µM) plus H2S (NaHS, 300 µM) for another 30 min before measuring chemotaxis
induced by MIP-2 (20 ng/mL). The data are reported as the mean ± SEM of cells per field. ..P <
Page 20 of 46

20
0.001 compared with RPMI; *P < 0.001 compared with RPMI plus MIP-2 and #P < 0.001
compared with NaHS 300 µM plus MIP-2; n = 5. One hour before the sham-operation (Sham),
non-severe (NS) or severe (S) sepsis induction by CLP surgery, mice were given saline (Sal),
NaHS (100 µmoles/kg, sc), Lawesson’s reagent (100 µmoles/kg, sc), GLB (40 µmoles/kg, sc),
GLB (40 µmoles/kg, sc) plus NaHS (100 µmoles/kg, sc) for another 30 min or GLB (40
µmoles/kg, sc) plus Lawesson’s reagent (100 µmoles/kg, sc) for another 30 min. (B) Six hours
after surgery, neutrophil migration to the infectious focus was determined. The data are reported
as the mean ± SEM x 106 neutrophils/cavity. *P < 0.01 compared to the NS plus Sal group; #P <
0.01 compared to the S plus Sal group; and **P < 0.01 compared to the S plus NaHS group
(ANOVA, followed by Bonferroni’s test, n = 5). (C) Survival rates were monitored over 8 days.
The results are expressed as percent of survival. *P < 0.05 compared to the NS plus Sal group;
#P < 0.01 compared to the S plus Sal group; and **P < 0.01 compared to the S plus H2S group.
Mantel-Cox log-rank test (n = 10-12). Flow cytometry of surface expression of CD11b (D),
CD62L (E) and CXCR2 (F) in blood neutrophils (Gr-1 positive cells) was evaluated 6 h after
surgery. The data are reported as the mean ± SEM of mean fluorescence intensity (MFI) and are
the result of the three independent experiments carried out in triplicate. *P < 0.01 compared to
the NS plus Sal group; #P < 0.01 compared to the S plus Sal group; and **P < 0.01 compared to
the S plus H2S group (ANOVA, followed by Bonferroni’s test, n = 5).
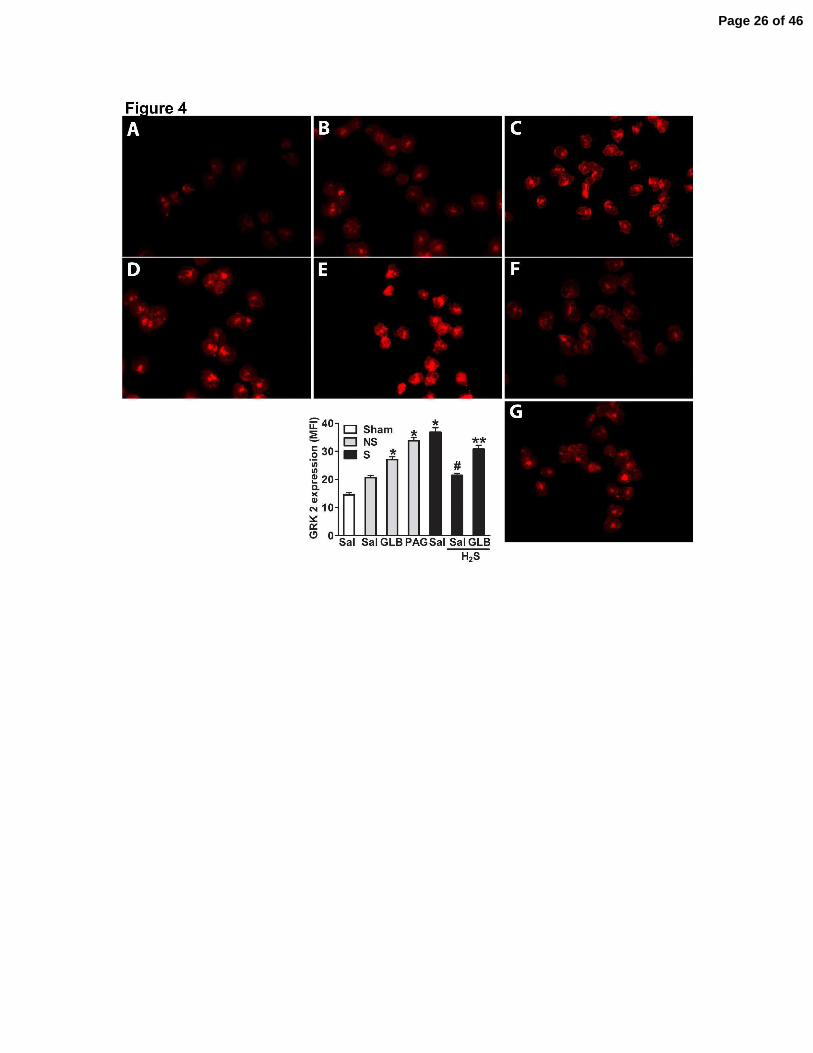
Figure 4. H2S prevented high GRK2 expression in blood neutrophils after severe sepsis via
activation of KATP channels. One hour before sham-operation (Sham), non-severe (NS) or
severe (S) sepsis induction by CLP, mice were given saline (Sal), PAG (50 mg/Kg, ip), GLB (40
µmoles/kg, sc), Lawesson’s reagent (H2S, 100 µmoles/kg, sc) or GLB (40 µmoles/kg, sc) + H2S
Page 21 of 46

21
(Lawesson’s reagent 100 µmoles/kg, sc, for another 30 min). Neutrophils were isolated from
total blood 6 hours after surgery, and GRK2 expression was evaluated by immunofluorescence in
Sham (A), NS (B), NS plus PAG (C), NS plus GLB (D), S (E), S plus H2S (F) and S plus GLB
plus H2S (G) groups. These semi-quantitative analyses showed the means of fluorescence
intensity (MFI). The number of animals per groups was 5, and we used 3 slices per animal and
determined the MFI of at least 200 neutrophils per slice. The experiment was repeated three
times. *P < 0.01 compared to the NS plus Sal group; #P < 0.01 compared to the S plus Sal
group; and **P < 0.01 compared to the S plus H2S group (ANOVA, followed by Bonferroni’s
test).
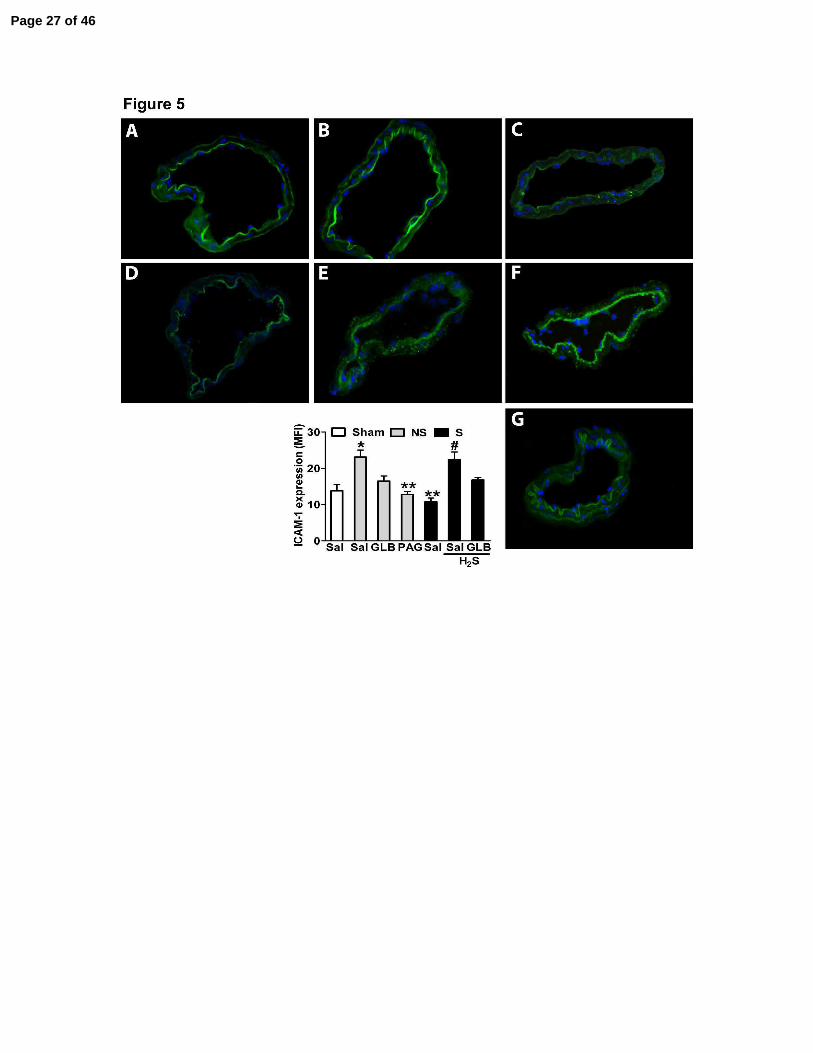
Figure 5. H2S donors enhanced ICAM-1 expression in mesenteric vessels of mice subjected
to severe sepsis by KATP channel activation. One hour before the sham-operation (Sham), non-
severe (NS) or severe (S) sepsis induction by CLP, mice were given saline (Sal), PAG (50
mg/Kg, ip), GLB (40 µmoles/kg, sc), Lawesson’s reagent (H2S, 100 µmoles/kg, sc) or GLB (40
µmoles/kg, sc) + H2S (Lawesson’s reagent 100 µmoles/kg, sc, for another 30 min).
Immunofluorescence staining for ICAM-1 on mesenteric vessels from Sham (A), NS (B), NS
plus PAG (C), NS plus GLB (D), S (E), S plus H2S (F) and S plus GLB plus H2S (G) was
performed. The number of animals per experimental group was 5 (n = 5), and four histological
slices were prepared from each animal. These semi-quantitative analyses showed the means of
fluorescence intensity (MFI) for ICAM-1 on the venular endothelium only. The experiment was
repeated two times. The data represent the mean ± SEM (n = 3). *P < 0.01 compared to the
Sham plus Sal group; **P < 0.01 compared to the NS plus Sal group; and #P < 0.01 compared to
the S plus Sal group (ANOVA, followed by Bonferroni’s test).
Page 22 of 46
Page 23 of 46
Page 24 of 46
Page 25 of 46
Page 26 of 46
Page 27 of 46

1
HYDROGEN SULFIDE IMPROVES NEUTROPHIL MIGRATION AND
SURVIVAL IN SEPSIS VIA K+
ATP CHANNEL ACTIVATION
Fernando Spiller1, Maria I.L. Orrico
1, Daniele C. Nascimento
1, Paula G. Czaikoski
1,
Fabrício O. Souto3, José C. Alves-Filho
1, Andressa Freitas
1, Daniela Carlos
1, Marcelo
F. Montenegro1, Alberto F. Neto
4, Sergio H. Ferreira
1, Marcos A. Rossi
2, John S.
Hothersall1, Jamil Assreuy
5 and Fernando Q. Cunha
1.
Online Data Supplement
Page 28 of 46
2
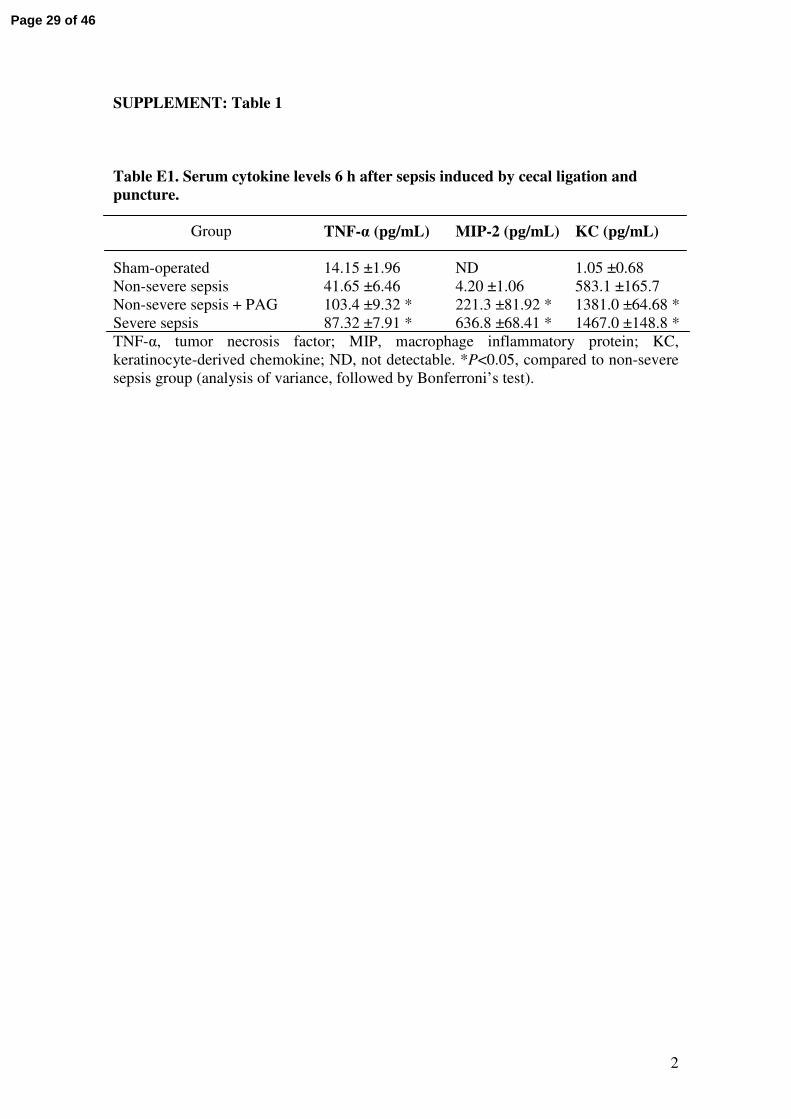
SUPPLEMENT: Table 1
Table E1. Serum cytokine levels 6 h after sepsis induced by cecal ligation and
puncture.
Group
TNF-α (pg/mL) MIP-2 (pg/mL) KC (pg/mL)
Sham-operated 14.15 ±1.96 ND 1.05 ±0.68
Non-severe sepsis 41.65 ±6.46 4.20 ±1.06 583.1 ±165.7
Non-severe sepsis + PAG 103.4 ±9.32 * 221.3 ±81.92 * 1381.0 ±64.68 *
Severe sepsis 87.32 ±7.91 * 636.8 ±68.41 * 1467.0 ±148.8 *
TNF-α, tumor necrosis factor; MIP, macrophage inflammatory protein; KC,
keratinocyte-derived chemokine; ND, not detectable. *P<0.05, compared to non-severe
sepsis group (analysis of variance, followed by Bonferroni’s test).
Page 29 of 46

3
SUPPLEMENT: Figure Legends
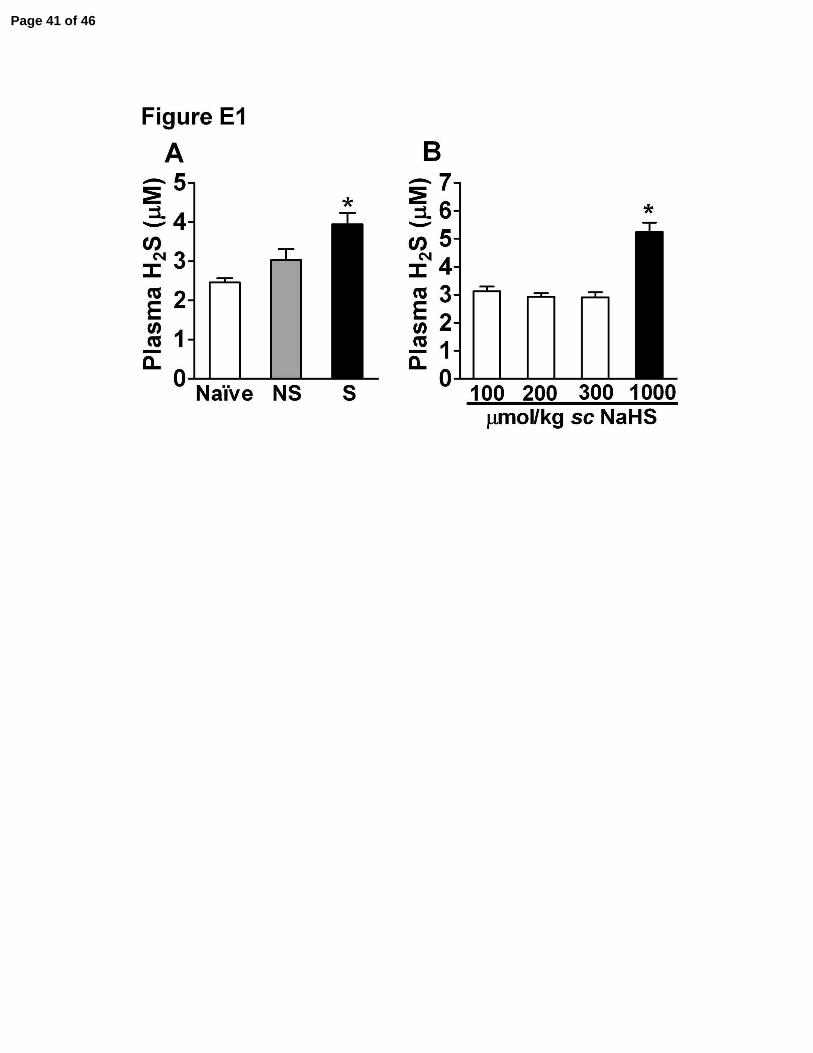
Figure E1. Plasma H2S levels increase after severe sepsis. (A) Plasma was collected
from mice subjected to non-severe (NS) or severe (S) sepsis after 6 h or from naïve
mice. *P<0.05 compared to naïve mice (ANOVA, followed by Bonferroni’s test, n=5)
(B) NaHS (100 – 1000 µmol/kg) were sc administrated and blood collected 30 min after
this administration. Data are expressed as mean of µM H2S. *P<0.05 compared to doses
of 100 to 300 µmol/kg (ANOVA, followed by Bonferroni’s test, n=5).
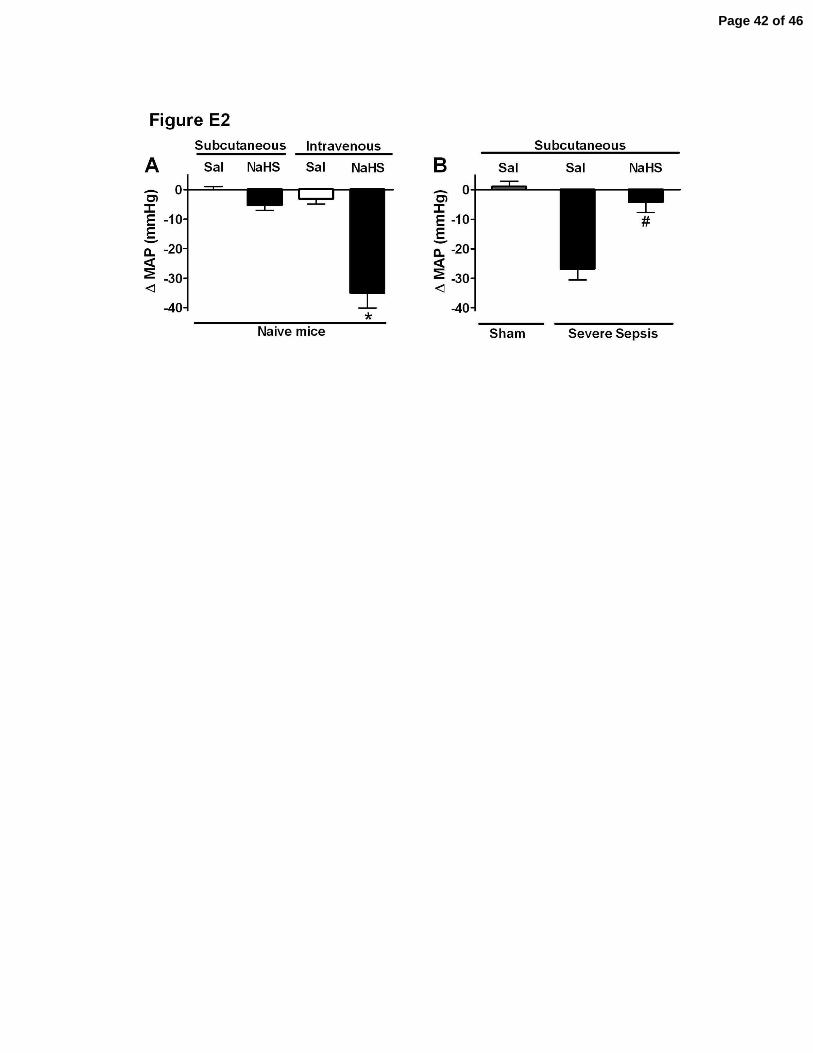
Figure E2. Effect of NaHS on MAP of naïve mice and mice with severe sepsis. (A)
Naïve mice were subcutaneous treated with NaHS (100 µmol/Kg) or saline (Sal) and the
mean arterial pressure (MAP) was measured after 6 h. In another set of experiment,
naïve mice were treated intravenously with NaHS (100 µmol/Kg) or Sal and the MAP
monitored for 20 min. *P<0.01 compared to Naive mice treated intravenously with Sal
(ANOVA, followed by Bonferroni’s test, n=5). (B) ∆ of Mean arterial pressure (MAP)
was determined 6 h after severe sepsis induction in mice subcutaneously pretreated (1 h
before surgery) with NaHS (100 µmol/kg, sc) or Sal. #P<0.01 compared to S plus Sal
group (t test, followed by Mann-Whitney U test). Data are reported as ∆ of
means ± SEM of two independent experiments (n=5).
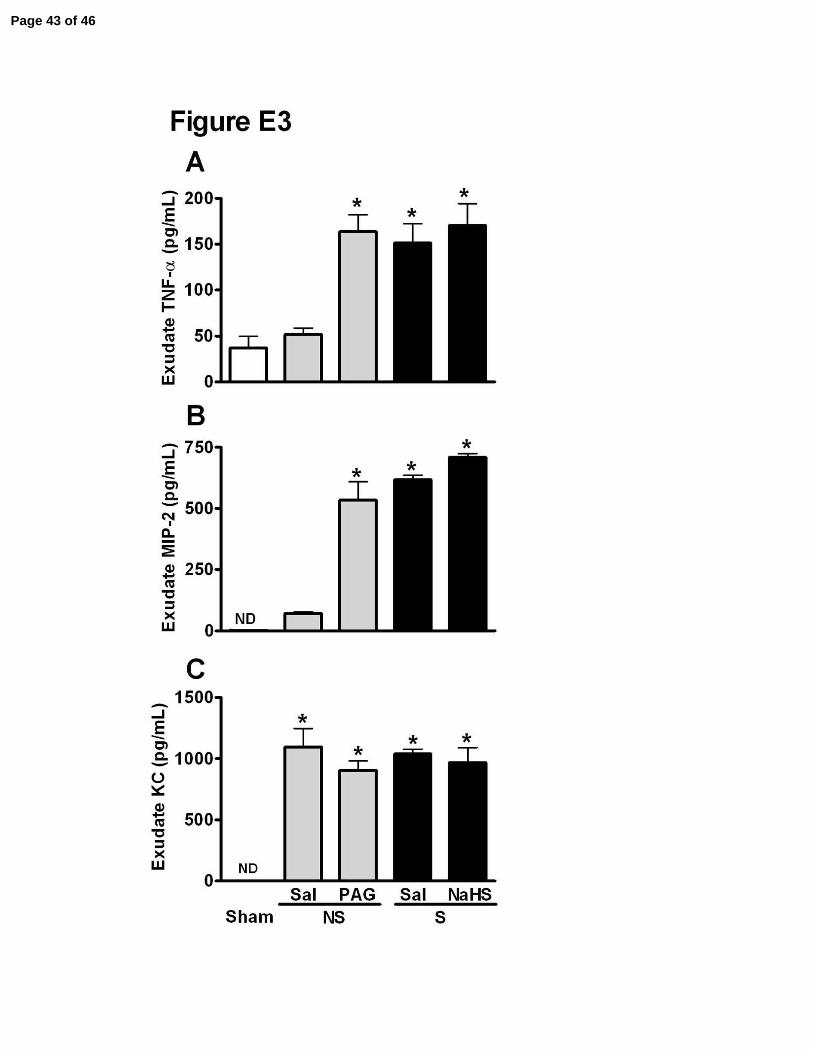
Figure E3. Inhibition of CSE increased the levels of TNF-α and MIP-2 in
peritoneal exudates. Animals were treated with saline (Sal), PAG (50 mg/kg, ip) or
NaHS (100 µmol/kg, sc) 1 hour before sham-operation (Sham), non-severe (NS) or
severe (S) sepsis induction. Six hours after surgery, the peritoneal exudate levels of (A)
TNF-α, (B) MIP-2, and (C) KC in peritoneal exudates were determined by ELISA. Data
Page 30 of 46

4
are reported as mean ± SEM of two independent experiments (n=5). *P<0.05 compared
to Sham (ANOVA, followed by Bonferroni’s test).
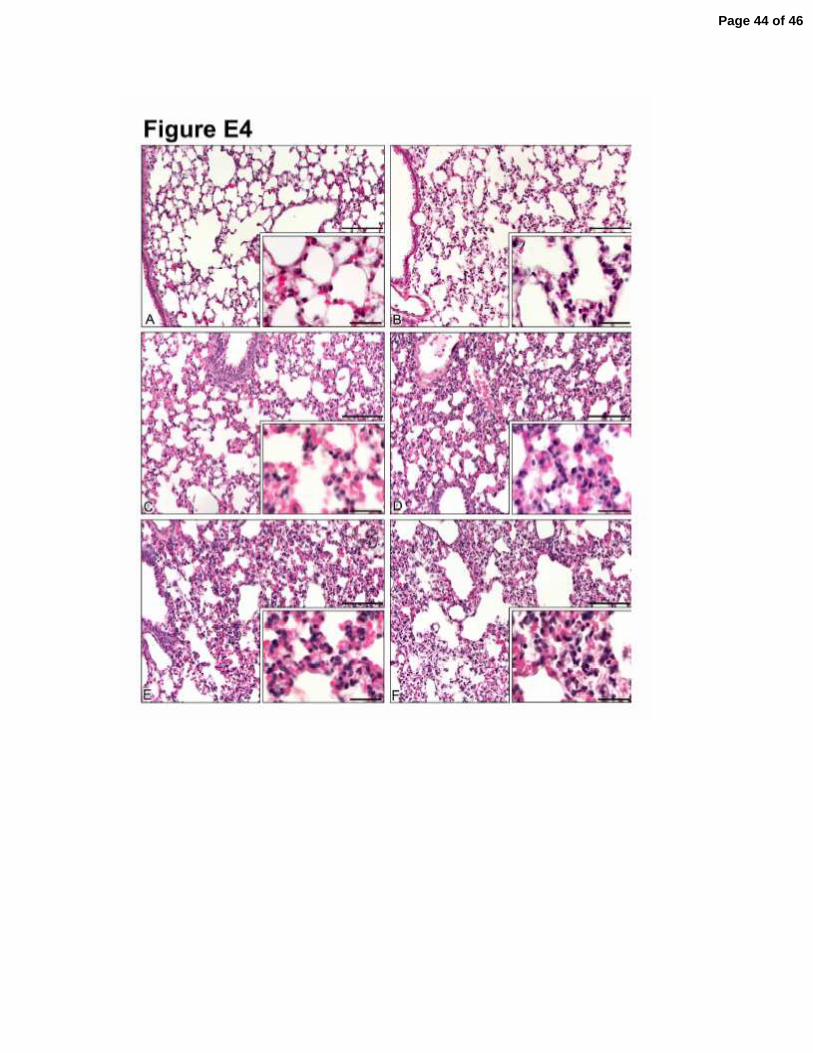
Figure E4. Morphological changes in lung after CLP-induced sepsis. Lungs were
harvested 12 hours after CLP surgery and the tissue sections were stained with
hematoxylin and eosin. Lung sections from mice subjected to sham-operated (A), non-
severe (C) and severe (E) sepsis. Lung sections from mice pretreated (1 h) with PAG
(50 mg/Kg, ip) and subjected to non-severe and severe sepsis are showed in (D) and (F),
respectively. One group of mice were pretreated (1 h) with NaHS (100 µmol/kg, sc) and
subjected to severe sepsis (B). These image of illustrative lung histology are
representative of three independent experiments (n=5). Main panel bars = 100 µm;
insert bars = 25 µm.
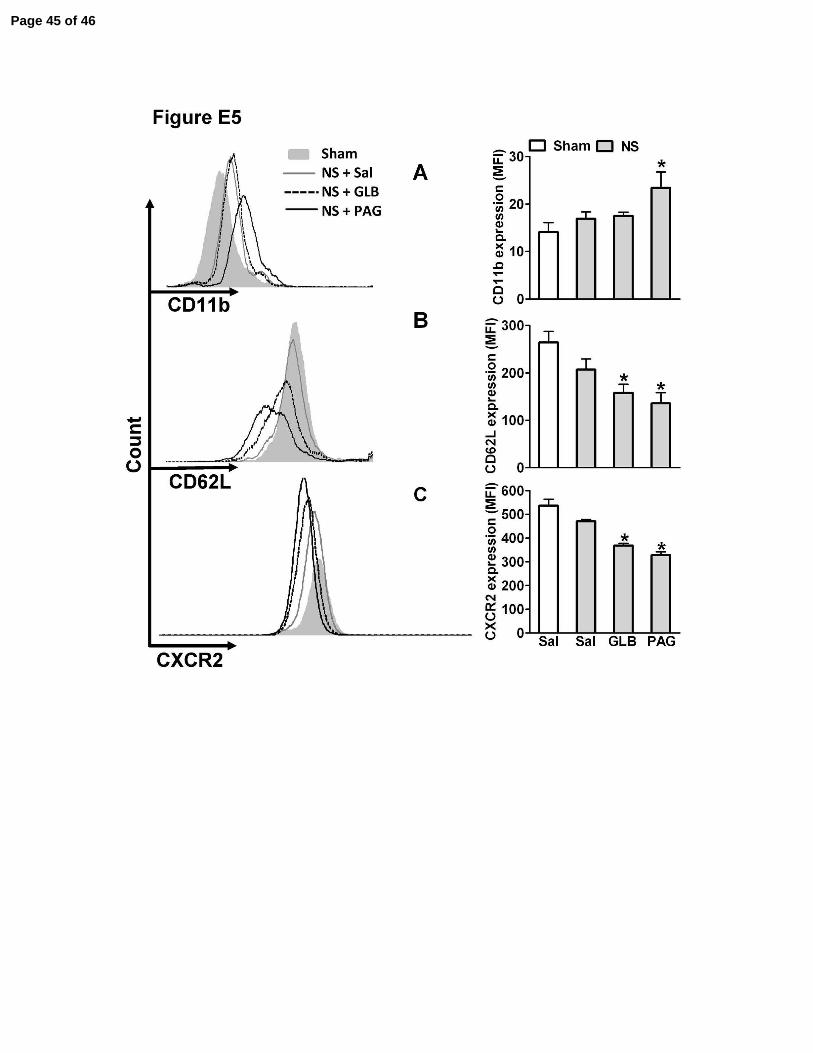
Figure E5. Pretreatment of mice subjected to non-severe sepsis with PAG induces
down-modulation of CD62L and CXCR2 and up-regulation of CD11b. One hour
before the sham-operated (Sham), non-severe (NS) or severe (S) sepsis induction by
CLP, mice were given saline (Sal), GLB (40 µmoles/kg, sc) or PAG (50 mg/kg, ip). Six
hours after surgery surface expression of CD11b (A), CD62L (B) and CXCR2 (C) on
blood neutrophils (Gr-1 positive cells) were evaluated by flow cytometry. *P<0.01
compared to NS plus Sal group (ANOVA, followed by Bonferroni’s test, n=5).
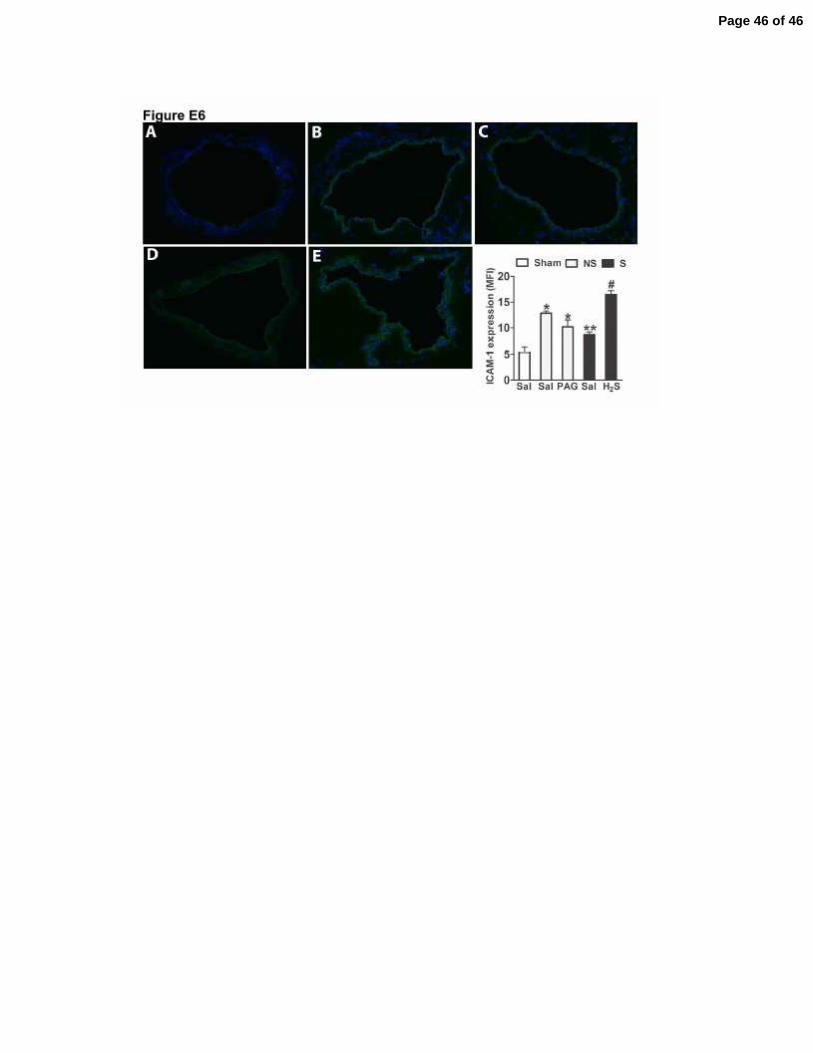
Figure E6. H2S donors increased ICAM-1 expression on lung vessels of mice
subjected to severe sepsis. One hour before sham-operation (Sham), and non-severe
(NS) or severe (S) sepsis induction by CLP, mice were given saline (Sal), PAG (50
mg/Kg, ip) or Lawesson’s reagent (H2S, 100 µmoles/kg, sc). Immunofluorescence
Page 31 of 46

5
staining for ICAM-1 on lung vessels from Sham (A), NS (B), NS plus PAG (C), S (D),
and S plus H2S (E) was performed. The number of animals per experimental group was
5 (n=5) and four histological slices were prepared for each animal. These semi-
quantitative analyses shows the mean fluorescence intensity (MFI) for ICAM-1 on
venular endothelium only. The experiment was repeated two times. *P<0.01 compared
to Sham plus Sal group; **P<0.01 compared to NS plus Sal group; #P<0.01 compared
to S plus Sal group (ANOVA, followed by Bonferroni’s test).
Page 32 of 46

6
SUPPLEMENT: Detailed Method Sections
Animals
The care and treatment of the animals was based on the Guide for the Care and
Use of Laboratory Animals (1), and the research was approved by the Animal Research
Ethics Committee of the FMRP. Male Swiss mice (22–25 g) were used. The mice were
obtained from the Animal Facility of the FMRP and housed in the Animal Room of the
Department of Pharmacology at 23–25°C with free access to water and food.
Sepsis model
Sepsis was induced by cecal ligation and puncture (CLP model), as described (2).
Mice were anesthetized with tribromoethanol (250 mg/kg, ip), a 1 cm midline incision
was made in the anterior abdomen, and the cecum was exposed and ligated below the
ileocecal junction. The cecum was punctured twice with a 26-gauge needle (for NS
sepsis induction) or twice with an 18-gauge needle (for S sepsis induction), and the
cecum was squeezed to allow its contents to be released through the punctures. Sham
animals underwent identical laparotomy but without cecal puncture. The cecum was
repositioned in the abdomen, and the peritoneal wall was closed. All animals received 1
mL of saline sc immediately after CLP.
Neutrophil migration to the peritoneal cavity
Neutrophil migration was assessed 6 h after induction of sepsis (3). The animals
were killed, and the cells present in the peritoneal cavity were harvested by introducing
1.5 ml of PBS containing 1mM EDTA. Total cell counts from the lavage were
performed with a cell counter (Coulter AC T series analyzer; Coulter Corp., Miami,
USA), and differential cell counts were carried out on cytocentrifuge slides (Cytospin 3;
Shandon Southern Products, Astmoore, UK) stained by the May-Grünwald-Giemsa
Page 33 of 46

7
(Rosenfeld) method. The results are expressed as the mean ± SEM of the number of
neutrophils per cavity.
Bacterial counts in blood
The bacterial count was determined as previously described (4). Briefly, mice
were killed 6 h after induction of sepsis. To measure bacteremia, blood was collected
under sterile conditions and 10 µl of blood was plated on Muller-Hinton agar dishes
(Difco Laboratories, Detroit, USA) and incubated at 37°C; CFU were analyzed after 12
h. The results were expressed as the mean ± SEM log of CFU per 10 µl of blood.
Intravital microscopy of mesenteric microcirculation
Leukocyte-endothelium interactions (rolling and adhesion) were examined as
previously described (5) in sham-operated or mice subjected to NS or S sepsis by CLP
(mice were pre-treated with saline, NaHS or PAG). Briefly, 2 and 4h after CLP
procedure, mice was anesthetized with tribromoethanol (250 mg/kg) and rolling and
adhesion were analyzed as follow. The mesenteric tissue was exteriorized for
microscopic examination in situ. Animals were maintained on a special board
thermostatically controlled at 37°C, with a transparent platform on which the tissue to
be transilluminated was placed. The preparation was kept moist by irrigating the tissue
with warmed (37°C) Ringer Locke's solution, pH 7.2 to 7.4, containing 1% gelatin. A
500-line television camera was incorporated onto a triocular Zeiss microscope to
facilitate observation of the enlarged image (x 3.400) on the video screen. Images were
recorded on a video recorder with a long distance objective (x 40) with a 0.65 numerical
aperture. The number of leukocyte adhering was determined at 10-min intervals and a
leukocyte was considered to be adherent to the venular endothelium if it remained
stationary for >60 s (6). Adherent cells are expressed as mean ± SEM of the number of
adherent cells per 100 µm2
of venule. Rolling leukocytes were defined as those white
Page 34 of 46

8
blood cells that moved at a lower velocity than erythrocytes in the
same stream and were
measured at 10 min intervals and are expressed as mean ± SEM of the leukocyte rolling
per min.
Cystathionine Gamma Lyase Assay
CSE activity was measured in total purified leukocytes by a modification of the
method of Steegborn et al. (7) following sham or sepsis induction (2 h). In brief, sample
lysates were prepared by 2 x sonication/freeze/thaw cycles from 106 cells in 100 µl of 5
mM phosphate buffer pH 7.4. Assays (final volume 200 µl) were carried out in 50 mM
phosphate buffer pH 7.4, containing pyridoxal P (0.5 mM) and with or without PAG (1
mM) and samples were pre-incubated at 25°C for 10 min. To start the reaction 5,5’-
dithio-bis(2-nitrobenzoic acid) (DTNB, 5mM) and cystathionine 2 mM were added and
the linear portion of the time course (60 -120 s) was monitored at 412 nm. Activity was
calculated by the OD difference after subtraction of OD change in the PAG containing
sample and calculated using an extinction coefficient Σ412 of 13600 M-1
cm-1
. The
activity of the CSE was expressed as mU/106 cells.
H2S assay by sulfide-sensitive electrode
The blood of mice were collected 6 h after severe sepsis induction or from naïve
mice in tubes with sodium citrate. Measurement of H2S concentration in plasma
samples involved use of a sulfide-sensitive electrode (Innovative Instruments, Inc, FL,
USA). In brief, 1.8 mL of miliQ water was supplemented with 0.2 mL of NaHS
standard solutions (final concentration of the 1, 3, 10, 30 and 100 µM). After each
measurement the electrode was washed with milliQ water. After standard calibration the
samples were measured in the same way. The sample were quantifies from the
calibrated standards.
Page 35 of 46

9
Determination of Cytokine Levels in the Peritoneal Exudates and Blood
Animals were sacrificed 6 h after sepsis induction, and the peritoneal lavage and
blood collected. Cytokines concentrations were determined by ELISA. The results were
expressed as the mean ± SEM as pg/mL.
Mean arterial pressure
Animals were anaesthetized by ip injection of ketamine (35 mg/Kg) and xilasine
(5 mg/Kg). Left carotid artery was cannulated and connected to a pressure transducer
(TRA 021), and this was coupled to a Power Lab/415 amplifier (MacLab System). One
femoral vein was cannulated for delivering chemicals. A heating pad was used to keep
the body temperature stable at 37°C. After 60 min of equilibration, H2S or saline was
injected in bolus intravenously and MAP was monitored for 20 min.
Isolation of bone marrow neutrophils
Bone marrow cells was collected and neutrophils were isolated by differential
centrifugation on Percoll gradient (8) with minor modifications. Gradient was prepared
in 15-ml polystyrene tube by layering 3 ml each of 72% and 65% Percoll solutions.
After centrifugation at 1200 g for 30 min at 25°C, the cell layer at the 72% upper
interface was collected as the neutrophil fraction. Erythrocytes were removed by lysis in
(NH4Cl 0.075 M) and the remaining neutrophil fractions were washed twice in
HANKS. The pelleted cells were resuspended in 1mL of RPMI-1640 medium (Sigma
Chemical Co., St Louis) and the number of neutrophils determined (90% purity and
95% viable cells).
Chemotaxis Assays
The neutrophil chemotaxis assay was performed using a 48-well modified Boyden
chamber (Neuro Probe Inc., Bethesda, MD, U.S.A.) and 5 µm pore size PVP-free
polycarbonate filters (Nuclepore: Costar Co., Cambridge, MA, U.S.A.). Mice bone
Page 36 of 46

10
marrow neutrophils (106 cells/mL in RPMI-0.01%) were incubated at 37°C for 1 h
either in the absence (control) or in the presence of GLB or the H2S donor sodium
hydrogen sulfide (NaHS, 30, 100 or 300 µM). In addition, one group was incubated
with GLB (100 µM, 37°C, 30 min) and then NaHS (300 µM) for a further 1 h. After
these respective incubations, neutrophils were allowed to migrate in the Boyden
chamber for 1 h (37°C) in response to RPMI (random migration) or the chemotactic
stimuli MIP-2 (20 ng/mL). Filters were removed, fixed, stained, and neutrophils that
had migrated through the membrane were counted in at least 5 randomly selected fields.
Each sample was assayed in triplicate. Results are expressed as the mean ± SEM
number of neutrophils per field.
Flow Cytometry Analysis of CD11b, CD62L and CXCR2 expression
Flow cytometry was analyzed according to the procedure previously described (9).
Total blood was collected 6 h after CLP and incubated for 30 minutes at 4°C with
fluorescent labeled antibodies: phycoerythrin (PE)-conjugated anti-CXCR2 mAb (1:50;
R&D Systems), phycoerythrin (PE)-conjugated anti-CD62L mAb (1:100; BD
Biosciences), fluorescein (FITC)-conjugated anti-CD11b mAb (1:100; BD
Biosciences) and peridinin chlorophyll protein (PerCP)-conjugated anti-Gr-1 mAb
(1:200; BD Biosciences). The appropriate conjugated mouse isotype control was used in
parallel (BD Biosciences). The cells were then washed twice with lyses buffer (NH4)
and subsequently washed twice in staining buffer (PBS with 5% BSA), fixed in 2%
formaldehyde and analyzed by flow cytometry. The analysis was performed in a
FACSort flow cytometer using CellQuest software (BD Biosciences) by setting a gate
for granulocyte cells derived from a side versus forward scatter dot plot. Neutrophils
were identified by their light-scatter properties and expression of Gr-1 in granulocytes
cells and the expression of CD62L, CD11b and CXCR2 were determined in this
Page 37 of 46

11
population. Three to five separate experiments were performed for each analyzed
population of neutrophils and approximately 10,000 gated events were collected in each
analysis.
Immunofluorescence assay for ICAM-1/CD54 or G protein-coupled receptor
kinase 2 (GRK2)
Six hours after CLP, an ICAM-1/CD54 immunofluorescence assay in mesenteric
and lung tissue or GRK2 on neutrophils was carried out according to the procedure
previously described (10). Slices were prepared using 5.0 x104 neutrophils. The
mesenteric and lung tissue was removed, washed in PBS and after freezing serial
sections were mounted on poly-L-lysine covered glass slides. For immunofluorescence
staining, 5 micron frozen tissue sections were fixed with paraformaldehyde (4%) in a
wet chamber at room temperature. The slides were incubated with PBS containing 1%
BSA (PBS-BSA), and then slices were incubated for 1 h with FITC-conjugated anti-
mouse CD54-ICAM-1 (1/200; BD Pharmingen) in PBS-BSA. The neutrophils slices
were then incubated with rabbit anti-mouse GRK2 mAb (1/200; Santa Cruz
Biotechnology) overnight, and then with red-fluorescence Alexa Fluor 594 (goat anti-
rabbit; 1/400; Invitrogen). Subsequently, slides were mounted using 4’,6’-diamidino-2-
phenylindole (Vector Laboratories) and sealed with enamel. The results of qualitative
analysis are expressed as fluorescence intensity of stained venules (x40) or neutrophils
(x40) present in the fluorescence microscopic field.
Statistical analysis
The data are reported as the means ± SEM of values obtained from two to three
independent experiments. We used five mice per experimental group with the exception
of survival analyses in which ten to twelve mice were used. The means value for
different groups were compared by analysis of variance (ANOVA). If significance was
Page 38 of 46

12
determined, individual comparisons were subsequently tested with Bonferroni's t test for
unpaired values. Bacterial counts were analyzed by the Mann-Whitney U test. The
survival rate was expressed as the percentage of live animals, and the Fisher´s exact test
was used to determine differences in survival curves. Moreover, the Kolmogorov-
Smirnov test was employed to test for normality. Since the rolling and adhesion results
failed to reach normality, data were log transformed to reach this assumption and then
an ANOVAs were performed with the modified data. A P<0.05 was considered
significant. The data were analyzed using GraphPad Prism version 5.00 for Windows
(GraphPad Software, USA) or SPSS 12.0 for Windows (SPSS, Chicago, IL).
REFERENCES FOR THE SUPPLEMENT
E1. Clark D. Guide for the care and use of laboratory animals. Washington, D.C.
USA; 1996.
E2. Wichterman KA, Baue AE, Chaudry IH. Sepsis and septic shock--a review of
laboratory models and a proposal. J Surg Res 1980;29:189-201.
E3. Benjamim CF, Ferreira SH, Cunha FQ. Role of nitric oxide in the failure of
neutrophil migration in sepsis. J Infect Dis 2000;182:214-223.
E4. Alves-Filho JC, de Freitas A, Russo M, Cunha FQ. Toll-like receptor 4 signaling
leads to neutrophil migration impairment in polymicrobial sepsis. Crit Care Med
2006;34:461-470.
E5. Benjamim CF, Silva JS, Fortes ZB, Oliveira MA, Ferreira SH, Cunha FQ.
Inhibition of leukocyte rolling by nitric oxide during sepsis leads to reduced migration
of active microbicidal neutrophils. Infect Immun 2002;70:3602-3610.
E6. Granger DN, Benoit JN, Suzuki M, Grisham MB. Leukocyte adherence to
venular endothelium during ischemia-reperfusion. Am J Physiol 1989;257:G683-688.
E7. Steegborn C, Clausen T, Sondermann P, Jacob U, Worbs M, Marinkovic S,
Huber R, Wahl MC. Kinetics and inhibition of recombinant human cystathionine
gamma-lyase. Toward the rational control of transsulfuration. J Biol Chem
1999;274:12675-12684.
E8. Alves-Filho JC, Freitas A, Souto FO, Spiller F, Paula-Neto H, Silva JS,
Gazzinelli RT, Teixeira MM, Ferreira SH, Cunha FQ. Regulation of chemokine
receptor by toll-like receptor 2 is critical to neutrophil migration and resistance to
polymicrobial sepsis. Proc Natl Acad Sci U S A 2009;106:4018-4023.
E9. Rios-Santos F, Alves-Filho JC, Souto FO, Spiller F, Freitas A, Lotufo CM,
Soares MB, Dos Santos RR, Teixeira MM, Cunha FQ. Down-regulation of cxcr2 on
neutrophils in severe sepsis is mediated by inducible nitric oxide synthase-derived nitric
oxide. Am J Respir Crit Care Med 2007;175:490-497.
E10. Dal-Secco D, Cunha TM, Freitas A, Alves-Filho JC, Souto FO, Fukada SY,
Grespan R, Alencar NM, Neto AF, Rossi MA, et al. Hydrogen sulfide augments
Page 39 of 46

13
neutrophil migration through enhancement of adhesion molecule expression and
prevention of cxcr2 internalization: Role of atp-sensitive potassium channels. J
Immunol 2008;181:4287-4298.
Page 40 of 46
Page 41 of 46
Page 42 of 46
Page 43 of 46
Page 44 of 46
Page 45 of 46
Page 46 of 46