
Growth and endocrine effects of recombinant bovine growth hormone treatment in non-transgenic and...
10
Growth and endocrine effects of recombinant bovine growth hormone treatment in non-transgenic and growth hormone transgenic coho salmon P.A. Raven a , D. Sakhrani a , B. Beckman b , L. Neregård c , L.F. Sundström a , B.Th. Björnsson c , R.H. Devlin a,⇑ a Department of Fisheries & Oceans, Centre for Aquaculture & Environmental Research, West Vancouver, BC, Canada V7V 1N6 b Northwest Fisheries Science Center, Seattle, WA 98112-2097, USA c Fish Endocrinology Laboratory, Department of Biology and Environmental Sciences, University of Gothenburg, Box 463, S-405 30 Göteborg, Sweden article info Article history: Received 19 December 2011 Revised 28 February 2012 Accepted 1 March 2012 Available online 14 March 2012 Keywords: Growth hormone GH Transgenic IGF-I Coho salmon abstract To examine the relative growth, endocrine, and gene expression effects of growth hormone (GH) trans- genesis vs. GH protein treatment, wild-type non-transgenic and GH transgenic coho salmon were treated with a sustained-release formulation of recombinant bovine GH (bGH; Posilac™). Fish size, specific growth rate (SGR), and condition factor (CF) were monitored for 14 weeks, after which endocrine param- eters were measured. Transgenic fish had much higher growth, SGR and CF than non-transgenic fish, and bGH injection significantly increased weight and SGR in non-transgenic but not transgenic fish. Plasma salmon GH concentrations decreased with bGH treatment in non-transgenic but not in transgenic fish where levels were similar to controls. Higher GH mRNA levels were detected in transgenic muscle and liver but no differences were observed in GH receptor (GHR) mRNA levels. In non-transgenic pituitary, GH and GHR mRNA levels per mg pituitary decreased with bGH dose to levels seen in transgenic salmon. Plasma IGF-I was elevated with bGH dose only in non-transgenic fish, while transgenic fish maintained an elevated level of IGF-I with or without bGH treatment. A similar trend was seen for liver IGF-I mRNA lev- els. Thus, bGH treatment increased fish growth and influenced feedback on endocrine parameters in non- transgenic but not in transgenic fish. A lack of further growth stimulation of GH transgenic fish suggests that these fish are experiencing maximal growth stimulation via GH pathways. Crown Copyright Ó 2012 Published by Elsevier Inc. All rights reserved. 1. Introduction Many teleost species show a remarkable capacity for growth enhancement [4,23]. Injections of exogenous growth hormone (GH) have resulted in increased growth in many species, including channel catfish [49,50], tilapia [38,68], striped bass [25], grass pickerel [67], rainbow trout [11,26,29], and coho salmon [14,30– 32,41,59]. Other physiological changes can be associated with in- creased growth rate, including increased feed conversion efficien- cies, greater energy and nutrient utilization, altered tissue shape and composition, and enhanced hypoosmoregulatory ability [4]. Thus, GH treatment has the potential to improve production effi- ciencies for aquaculture, however to date has not been applied in commercial production due to public perception issues and a lack of field trials of sufficient scale to demonstrate efficacy. An alternative to GH injections for growth acceleration is the creation of transgenic fish that over-produce GH from gene con- structs [20,21,24,33,39,40,42,45,52,53]. Transgenic salmon that over express GH show similar physiological changes as are found for GH protein injection, including enhanced growth rates [20,21,24], increased appetite and feeding motivation, enhanced feed conversion efficiencies, increased nutrient and energy utiliza- tion, and acceleration of development and onset of maturation [1,10,12,15,16,19,47,54,55,61–63]. Elevated levels of GH in salmon, whether by injection or transgenesis affect the growth hormone – insulin-like growth factor-I (GH-IGF-I) axis, modifying gene expression and plasma hormone levels and hence modifying growth rates [3,7,17,28,35,57]. Normally, GH is secreted from the pituitary gland, circulates in plasma, and binds growth hormone receptors (GHR) to stimulate IGF-I mRNA transcription and subse- quent IGF-I protein release from the liver and other tissues [51,56,58,66]. IGF-I mediates many GH growth effects and both hormones can feed back on the pituitary to reduce GH mRNA tran- scription and GH protein production [2,7]. In addition, autocrine and paracrine stimulation by GH and IGF-I may be functioning in many tissues, particularly in transgenic animals [8,9,55]. Intraperitoneally injected GH circulates in the body and acts via endocrine mechanisms, whereas transgenesis has the potential to act in both an endocrine and paracrine/autocrine fashion (and potentially intracellularly). In both transgenic and injected cases, expression or treatment can be adjusted such that saturating ef- fects on growth stimulation are achieved - that is, further GH administration, or increasing the dosage of the transgene has little 0016-6480/$ - see front matter Crown Copyright Ó 2012 Published by Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.ygcen.2012.03.002 ⇑ Corresponding author. Fax: +1 604 666 3497. E-mail address: [email protected] (R.H. Devlin). General and Comparative Endocrinology 177 (2012) 143–152 Contents lists available at SciVerse ScienceDirect General and Comparative Endocrinology journal homepage: www.elsevier.com/locate/ygcen
Transcript of Growth and endocrine effects of recombinant bovine growth hormone treatment in non-transgenic and...

General and Comparative Endocrinology 177 (2012) 143–152
Contents lists available at SciVerse ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen
Growth and endocrine effects of recombinant bovine growth hormone treatmentin non-transgenic and growth hormone transgenic coho salmon
P.A. Raven a, D. Sakhrani a, B. Beckman b, L. Neregård c, L.F. Sundström a, B.Th. Björnsson c, R.H. Devlin a,⇑a Department of Fisheries & Oceans, Centre for Aquaculture & Environmental Research, West Vancouver, BC, Canada V7V 1N6b Northwest Fisheries Science Center, Seattle, WA 98112-2097, USAc Fish Endocrinology Laboratory, Department of Biology and Environmental Sciences, University of Gothenburg, Box 463, S-405 30 Göteborg, Sweden
a r t i c l e i n f o
Article history:Received 19 December 2011Revised 28 February 2012Accepted 1 March 2012Available online 14 March 2012
Keywords:Growth hormoneGHTransgenicIGF-ICoho salmon
0016-6480/$ - see front matter Crown Copyright � 2http://dx.doi.org/10.1016/j.ygcen.2012.03.002
⇑ Corresponding author. Fax: +1 604 666 3497.E-mail address: [email protected] (R.H
a b s t r a c t
To examine the relative growth, endocrine, and gene expression effects of growth hormone (GH) trans-genesis vs. GH protein treatment, wild-type non-transgenic and GH transgenic coho salmon were treatedwith a sustained-release formulation of recombinant bovine GH (bGH; Posilac™). Fish size, specificgrowth rate (SGR), and condition factor (CF) were monitored for 14 weeks, after which endocrine param-eters were measured. Transgenic fish had much higher growth, SGR and CF than non-transgenic fish, andbGH injection significantly increased weight and SGR in non-transgenic but not transgenic fish. Plasmasalmon GH concentrations decreased with bGH treatment in non-transgenic but not in transgenic fishwhere levels were similar to controls. Higher GH mRNA levels were detected in transgenic muscle andliver but no differences were observed in GH receptor (GHR) mRNA levels. In non-transgenic pituitary,GH and GHR mRNA levels per mg pituitary decreased with bGH dose to levels seen in transgenic salmon.Plasma IGF-I was elevated with bGH dose only in non-transgenic fish, while transgenic fish maintained anelevated level of IGF-I with or without bGH treatment. A similar trend was seen for liver IGF-I mRNA lev-els. Thus, bGH treatment increased fish growth and influenced feedback on endocrine parameters in non-transgenic but not in transgenic fish. A lack of further growth stimulation of GH transgenic fish suggeststhat these fish are experiencing maximal growth stimulation via GH pathways.
Crown Copyright � 2012 Published by Elsevier Inc. All rights reserved.
1. Introduction
Many teleost species show a remarkable capacity for growthenhancement [4,23]. Injections of exogenous growth hormone(GH) have resulted in increased growth in many species, includingchannel catfish [49,50], tilapia [38,68], striped bass [25], grasspickerel [67], rainbow trout [11,26,29], and coho salmon [14,30–32,41,59]. Other physiological changes can be associated with in-creased growth rate, including increased feed conversion efficien-cies, greater energy and nutrient utilization, altered tissue shapeand composition, and enhanced hypoosmoregulatory ability [4].Thus, GH treatment has the potential to improve production effi-ciencies for aquaculture, however to date has not been applied incommercial production due to public perception issues and a lackof field trials of sufficient scale to demonstrate efficacy.
An alternative to GH injections for growth acceleration is thecreation of transgenic fish that over-produce GH from gene con-structs [20,21,24,33,39,40,42,45,52,53]. Transgenic salmon thatover express GH show similar physiological changes as are foundfor GH protein injection, including enhanced growth rates
012 Published by Elsevier Inc. All r
. Devlin).
[20,21,24], increased appetite and feeding motivation, enhancedfeed conversion efficiencies, increased nutrient and energy utiliza-tion, and acceleration of development and onset of maturation[1,10,12,15,16,19,47,54,55,61–63]. Elevated levels of GH in salmon,whether by injection or transgenesis affect the growth hormone –insulin-like growth factor-I (GH-IGF-I) axis, modifying geneexpression and plasma hormone levels and hence modifyinggrowth rates [3,7,17,28,35,57]. Normally, GH is secreted from thepituitary gland, circulates in plasma, and binds growth hormonereceptors (GHR) to stimulate IGF-I mRNA transcription and subse-quent IGF-I protein release from the liver and other tissues[51,56,58,66]. IGF-I mediates many GH growth effects and bothhormones can feed back on the pituitary to reduce GH mRNA tran-scription and GH protein production [2,7]. In addition, autocrineand paracrine stimulation by GH and IGF-I may be functioning inmany tissues, particularly in transgenic animals [8,9,55].
Intraperitoneally injected GH circulates in the body and acts viaendocrine mechanisms, whereas transgenesis has the potential toact in both an endocrine and paracrine/autocrine fashion (andpotentially intracellularly). In both transgenic and injected cases,expression or treatment can be adjusted such that saturating ef-fects on growth stimulation are achieved - that is, further GHadministration, or increasing the dosage of the transgene has little
ights reserved.

144 P.A. Raven et al. / General and Comparative Endocrinology 177 (2012) 143–152
effect on growth. However, the relative effects of transgenesis vs.GH protein injection have not been directly compared, nor hasthe effect of supplemental GH administration in GH transgenic sal-mon. In the present experiments, transgenic and non-transgenicfish were injected with a slow-release formulation of bovine GH(Posilac™; Monsanto Corporation), previously shown to increasegrowth in salmonids [29,35,38,41,46,49,50,64]. Growth was moni-tored over a 14 week period, and plasma IGF-I and GH hormonelevels, and GH, IGF-I and GHR mRNA levels in several tissues weremeasured to assess effects on the endocrine system.
2. Methods
2.1. Fish culture and sampling
Experiments were conducted on strains of coho salmon(Oncorhynchus kisutch). Non-transgenic (NT) coho salmon were de-rived from crosses using 10 male and 10 female parents from theChehalis River in southwestern British Columbia, Canada. GHtransgenic salmon (T; M77 strain containing the OnMTGH1 con-struct, Devlin et al. 1994; 2004) contain GH transgenes at a singlelocus and was originally developed from the same Chehalis Riverstrain. The T strain is maintained by backcrossing at each genera-tion with 10 females derived from wild Chehalis River populationto retain a wild background genotype. On March 1, 2004, 102 NTand 102 T coho salmon were weighed, measured for fork-length,and intraperitoneally injected with passive integrated transponder(PIT) tags. Following a 3-day recovery period without feeding, 6size-matched groups (n = 17; 17.5 ± 0.1 g, mean fish weight ± SE)were selected for each genotype, and groups were intraperitone-ally injected (at the ventral midline anterior to the anal fin) with(1) a control (CTRL) injection of 0.01 M phosphate buffered saline,pH 7.4, (2) a low (Low) dose of Posilac™ (Monsanto Corporation, St.Louis, MI) at 2.23 mg recombinant bovine GH/g body weight or (3)a high (High) dose of Posilac™ at 6.69 mg recombinant bovine GH/g body weight. Fish group names are described by genotype fol-lowed by dosage, i.e. NT-CTRL, NT-Low, NT-High, T-CTRL, T-Lowand T-High. One group of each treatment ‘‘x’’ genotype was placedinto each of two replicate 3000 L tanks (Tank A and B); thus eachtank held each of the six fish groups. Tanks were supplied withwell water (10.2 mg/L DO2, 10 �C ± 1 �C) at 1 L/min/kg body weightand reared for 98 days on a simulated natural photoperiod (Van-couver, Canada, 45�150N, 123�100W from March to June). Startingon day 1, fish were fed to satiation twice daily using size-appropri-ate commercial salmon feed (Skretting Canada). Fish rearing andexperimentation was conducted as per Canadian Council for Ani-mal Care guidelines and at Fisheries and Oceans Canada’s labora-tory in West Vancouver which possesses physical containmentmeasures to prevent escape of transgenic fish to the naturalenvironment.
At 2-week intervals following injection, fish were anesthetizedin tricane methane-sulphonate (MS 222; 0.2 g/L supplementedwith air and 0.2 g of sodium bicarbonate/L), weighed, and mea-sured for fork-length. At 14 weeks post-injection all fish wereagain assessed for size and 14 NT-CTRL, 13 NT-Low, 12 NT-High,13 T-CTRL, 12 T-Low and 11 T-High fish were obtained from TankA and Tank B and rapidly team sampled for blood, pituitary, muscleand liver tissue, with fish taken from the different groups randomlythroughout the sampling period. Blood was centrifuged at 4000gfor 5 min at 4 �C, and plasma removed and frozen at �80 �C formeasurement of growth hormone (GH) and insulin-like growthfactor-I (IGF-I) protein levels. Pituitary, muscle and liver wereplaced in RNAlater (Ambion) at 4 �C overnight and then frozen at�80 �C. White muscle was dissected from above the lateral lineon the left side of the fish 1 cm behind the operculum, and a liversample was taken from the tip of the posterior lobe.
In a second experiment, NT and T coho salmon fry (4 weeks old,average weight 0.5 g) were injected with either 4 mg recombinantbovine GH/g body weight or a sham injection as described aboveand 60 fish were assigned to each of four groups; NT-CTRL, NT-GH, T-CTRL and T-GH. The fry were held in 6 tanks of 40 fish (10fish from each treatment) at the conditions above and 10 represen-tative fish per treatment were randomly selected and measured forweight and length at 0, 2 and 4 weeks. Specific growth rates (SGR)were calculated as follows: SGR = 100[ln (W2 �W�1
1 )](d2 � d1)�1
where W = weight in grams and d = days.
2.2. Hormone analysis
Plasma growth hormone levels were measured using an estab-lished 125I-based radioimmunoassay [65] that measures salmonGH and does not cross react with exogenous bGH. Plasma IGF-Iconcentrations were determined using an 125I radioimmunoassayIGF-I kit (GroPep IGF-I Fish Kit, Adelaide, Australia) as described[55].
2.3. RNA extraction and quantitative PCR
RNA from frozen muscle and liver samples were extracted, quan-tified and measured for b-actin, GH and IGF-I mRNA levels usingquantitative PCR (Q-PCR) as in Raven et al. [55]. Whole pituitarieswere weighed to the nearest lg (PerkinElmer AD6 Autobalance),and RNA was extracted by homogenization using a micropestleand Micro-Aqueous Kit (Ambion). To allow quantitation of RNArecovery per pituitary, column elution volumes were measuredand RNA concentrations determined using a PicoGreen kit (AppliedBiosystems) on a FLUOROStar Omega plate reader. 500 ng of RNAwas reverse transcribed using a SuperScript III Reverse TranscriptionKit (Invitrogen) and Q-PCR was performed and analyzed on an Ap-plied Biosystems 7500 Real-Time PCR System using a dilution seriesof pooled cDNA. Inter-plate differences within a group were com-pensated for by measuring four RNA samples on each plate (platestandards) and correcting RNA quantities on each plate relative tothe change in the mean of the four samples (mean standard samplequantities from initial plate/mean standard sample quantities onplate of interest X experimental sample on plate of interest). b-actinquantities showed no significant differences between groups formuscle and liver (data not shown) and therefore were used to nor-malize muscle and liver mRNA levels (GH or IGF-I mRNA levels/b-ac-tin mRNA level). Pituitary b-actin did show significant differencesbetween groups and thus was not used to normalize mRNA levels.Pituitary mRNA levels are reported per 0.01 ng RNA for GH and per10 ng RNA for GHR, levels per whole pituitary, levels per whole pitu-itary per g fish, and levels per mg pituitary. We assumed that equalpituitary homogenization and extraction of RNA and reverse tran-scription of RNA occurred between groups, although these technicalvariables could have increased variance and made detection of dif-ferences among groups more difficult.
2.4. Statistical analyses
For all analyses, a two-way ANOVA using the variables of geno-type and bGH dose was used to assess treatment effects, followedby Holm-Sidak multiple comparison tests to identify intergroup dif-ferences. In cases where data were not normal or variances were notequal, a transformation was applied followed by two-way ANOVA. Ifdata transformation did not result in normal data or equal variances,a Kruskal–Wallis one-way ANOVA on ranks was performed on thesix treatment groups (genotype and dose variables combined) fol-lowed by Dunn’s multiple comparison tests. Grubb’s outlier testwas performed on growth hormone data and 2 outlier data pointswere removed. Correlation analyses were performed using Pearson
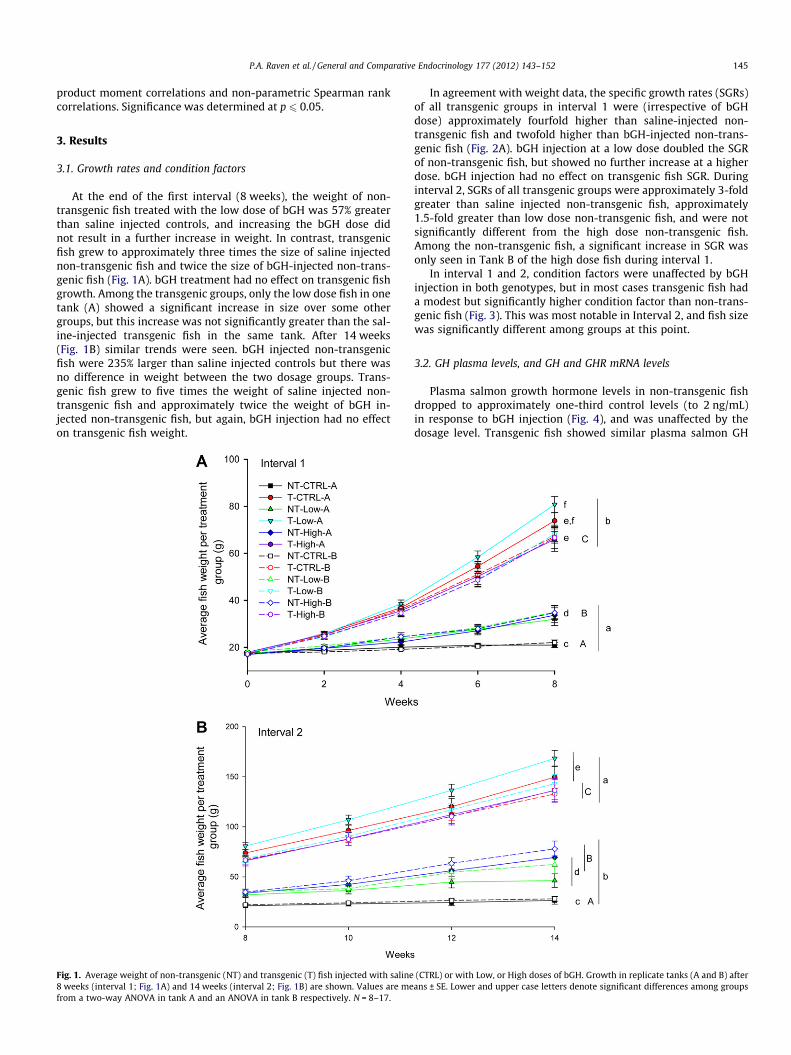
P.A. Raven et al. / General and Comparative Endocrinology 177 (2012) 143–152 145
product moment correlations and non-parametric Spearman rankcorrelations. Significance was determined at p 6 0.05.
3. Results
3.1. Growth rates and condition factors
At the end of the first interval (8 weeks), the weight of non-transgenic fish treated with the low dose of bGH was 57% greaterthan saline injected controls, and increasing the bGH dose didnot result in a further increase in weight. In contrast, transgenicfish grew to approximately three times the size of saline injectednon-transgenic fish and twice the size of bGH-injected non-trans-genic fish (Fig. 1A). bGH treatment had no effect on transgenic fishgrowth. Among the transgenic groups, only the low dose fish in onetank (A) showed a significant increase in size over some othergroups, but this increase was not significantly greater than the sal-ine-injected transgenic fish in the same tank. After 14 weeks(Fig. 1B) similar trends were seen. bGH injected non-transgenicfish were 235% larger than saline injected controls but there wasno difference in weight between the two dosage groups. Trans-genic fish grew to five times the weight of saline injected non-transgenic fish and approximately twice the weight of bGH in-jected non-transgenic fish, but again, bGH injection had no effecton transgenic fish weight.
Fig. 1. Average weight of non-transgenic (NT) and transgenic (T) fish injected with saline8 weeks (interval 1; Fig. 1A) and 14 weeks (interval 2; Fig. 1B) are shown. Values are mefrom a two-way ANOVA in tank A and an ANOVA in tank B respectively. N = 8–17.
In agreement with weight data, the specific growth rates (SGRs)of all transgenic groups in interval 1 were (irrespective of bGHdose) approximately fourfold higher than saline-injected non-transgenic fish and twofold higher than bGH-injected non-trans-genic fish (Fig. 2A). bGH injection at a low dose doubled the SGRof non-transgenic fish, but showed no further increase at a higherdose. bGH injection had no effect on transgenic fish SGR. Duringinterval 2, SGRs of all transgenic groups were approximately 3-foldgreater than saline injected non-transgenic fish, approximately1.5-fold greater than low dose non-transgenic fish, and were notsignificantly different from the high dose non-transgenic fish.Among the non-transgenic fish, a significant increase in SGR wasonly seen in Tank B of the high dose fish during interval 1.
In interval 1 and 2, condition factors were unaffected by bGHinjection in both genotypes, but in most cases transgenic fish hada modest but significantly higher condition factor than non-trans-genic fish (Fig. 3). This was most notable in Interval 2, and fish sizewas significantly different among groups at this point.
3.2. GH plasma levels, and GH and GHR mRNA levels
Plasma salmon growth hormone levels in non-transgenic fishdropped to approximately one-third control levels (to 2 ng/mL)in response to bGH injection (Fig. 4), and was unaffected by thedosage level. Transgenic fish showed similar plasma salmon GH
(CTRL) or with Low, or High doses of bGH. Growth in replicate tanks (A and B) afterans ± SE. Lower and upper case letters denote significant differences among groups

Fig. 2. Specific growth rate in weight (%) of non-transgenic (NT) and transgenic (T)fish injected with saline (CTRL) or with Low, or High doses of bGH. Specific growthrates in replicate tanks (A and B) after 8 weeks (interval 1; Fig. 1A) and 14 weeks(interval 2; Fig. 1B) are shown. Values are means ± SE. Lower and upper case lettersdenote significant differences among groups in tank A and tank B respectively.Horizontal bars denote differences between T and NT where a two-way ANOVA waspossible. N = 8–16.
Fig. 3. Condition factor of non-transgenic (NT) and transgenic (T) fish injected withsaline (CTRL) or with Low, or High doses of bGH. Condition factor in replicate tanks(A and B) after 8 weeks (interval 1; Fig. 1A) and 14 weeks (interval 2; Fig. 1B) areshown. Values are means ± SE. Lower and upper case letters denote significantdifferences among groups in tank A and tank B respectively. Horizontal bars denotedifferences between T and NT where a two-way ANOVA was possible. N = 7–16.
146 P.A. Raven et al. / General and Comparative Endocrinology 177 (2012) 143–152
levels to saline-injected non-transgenic fish despite bGH injection.Due to the lower plasma GH levels in bGH injected non-transgenicfish, the non-transgenic fish as a whole had a significantly lowerplasma salmon GH concentration than transgenic fish.
GH mRNA in liver (Fig. 4B) was undetectable in non-transgenicfish but present in all transgenic fish (as expected by expressionfrom the GH transgene). Non-transgenic fish are not expected tohave GH expression in the liver as the pituitary is the main sourceof endogenous GH. Compared to transgenic fish, non-transgenicfish had an approximately eightfold lower GH mRNA level in mus-cle (Fig. 4C). In both tissues, there was a trend towards bGH sup-pressing GH mRNA levels in a dose-specific fashion, but inneither case was this significant due to the variable data associatedwith the levels of expression detected.
Levels of pituitary GH mRNA have been expressed in severalformats to reflect potential effects at the cellular, pituitary, andorganismal levels. Per unit of assay input RNA (0.01 ng), GH mRNAlevels for transgenic fish were higher than for non-transgenic fish,but there was no effect of bGH injection (Fig. 5A). GH mRNA levelsper whole pituitary showed a similar trend as per unit assay RNA
(Fig. 5B). Pituitary sizes differed significantly between groups. Highdose non-transgenic fish had a larger pituitary compared to con-trols while the low dose non-transgenic fish pituitary size wasintermediate. All transgenic fish groups had pituitary sizes similarto each other and larger than the low dose non-transgenic but notthe high-dose non-transgenic group (data not shown). GH mRNAper mg pituitary tissue (which adjusts for varying pituitary sizesto examine whether negative feedback was occurring; Fig. 5C)showed a reduced level in transgenic compared to non-transgenicfish. Further, bGH treatment reduced GH mRNA levels in non-transgenic fish close to transgenic levels. A similar trend is seenwhen pituitary GH mRNA levels are calculated per gram of fish(to examine the total amount of pituitary GH mRNA available to di-rect effects on a specific number of cells in each group; Fig. 5D). Inthis case, GH mRNA levels were reduced by bGH in non-transgenicfish, but remained unaffected in transgenic fish. No differences dueto genotype or bGH dose were seen in GHR expression levels in li-ver (Fig. 6A) or muscle (Fig. 6B). In the pituitary, no difference inGHR mRNA levels were seen among groups when calculated perassay input RNA or per whole pituitary (data not shown). However,when calculated on a per mg pituitary basis, GHR mRNA was
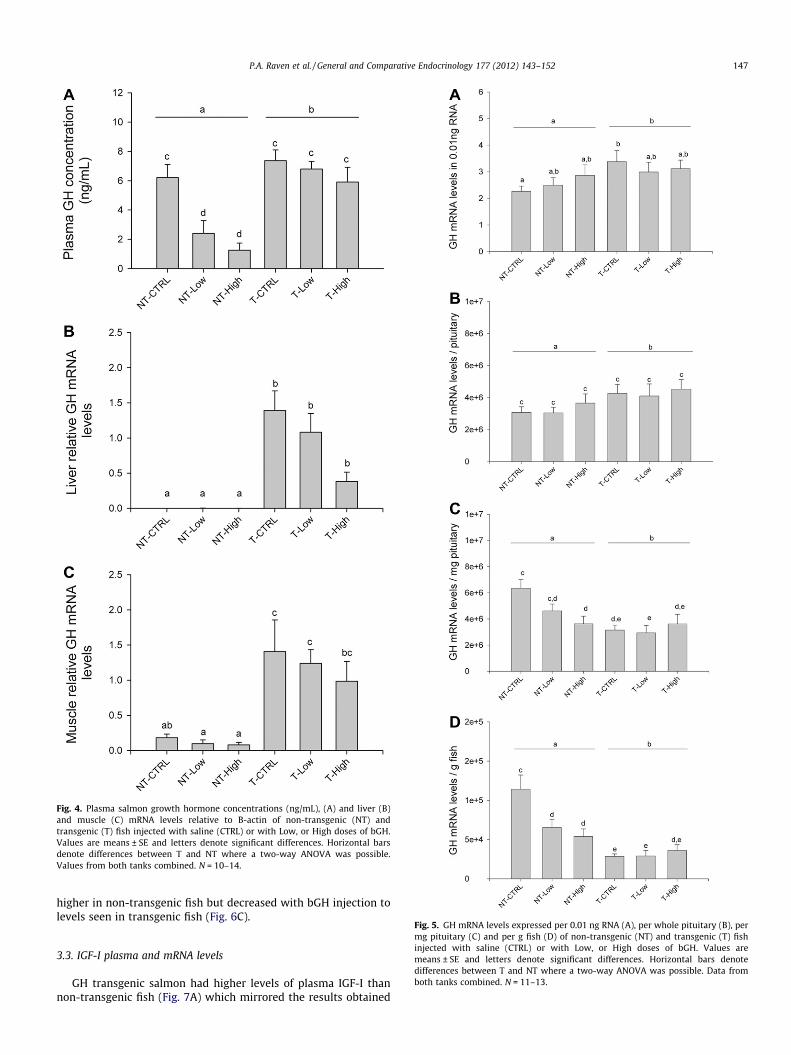
Fig. 4. Plasma salmon growth hormone concentrations (ng/mL), (A) and liver (B)and muscle (C) mRNA levels relative to B-actin of non-transgenic (NT) andtransgenic (T) fish injected with saline (CTRL) or with Low, or High doses of bGH.Values are means ± SE and letters denote significant differences. Horizontal barsdenote differences between T and NT where a two-way ANOVA was possible.Values from both tanks combined. N = 10–14.
P.A. Raven et al. / General and Comparative Endocrinology 177 (2012) 143–152 147
higher in non-transgenic fish but decreased with bGH injection tolevels seen in transgenic fish (Fig. 6C).
Fig. 5. GH mRNA levels expressed per 0.01 ng RNA (A), per whole pituitary (B), permg pituitary (C) and per g fish (D) of non-transgenic (NT) and transgenic (T) fishinjected with saline (CTRL) or with Low, or High doses of bGH. Values aremeans ± SE and letters denote significant differences. Horizontal bars denotedifferences between T and NT where a two-way ANOVA was possible. Data fromboth tanks combined. N = 11–13.
3.3. IGF-I plasma and mRNA levels
GH transgenic salmon had higher levels of plasma IGF-I thannon-transgenic fish (Fig. 7A) which mirrored the results obtained

Fig. 6. GHR mRNA levels relative to B-actin in liver (A) and muscle (B) and GHRmRNA levels per mg pituitary (C) of non-transgenic (NT) and transgenic (T) fishinjected with saline (CTRL) or with Low, or High doses of bGH. Values aremeans ± SE. Horizontal bars denote differences between T and NT where a two-wayANOVA was possible. Data from both tanks combined. N = 11–14.
Fig. 7. Plasma insulin-like growth factor-I concentrations (ng/mL, A) and IGF-ImRNA levels relative to B-actin in liver (B) and muscle (C) of non-transgenic (NT)and transgenic (T) fish injected with saline (CTRL) or with Low, or High doses ofbGH. Values are means ± SE and letters denote significant differences. Horizontalbars denote differences between T and NT where a two-way ANOVA was possible.Data from both tanks combined. N = 10–14.
148 P.A. Raven et al. / General and Comparative Endocrinology 177 (2012) 143–152
for growth and SGR between the genotypes. Although bGH did ap-pear to increase plasma IGF-I in a dosage-dependant way in non-transgenic fish, only the high dose group had significantly higher
IGF-I levels than controls (Fig. 7A). Transgenic fish showed no dif-ference in IGF-I concentrations with bGH injections. IGF-I mRNAlevels in the liver show similar, although less pronounced, relation-ships as for plasma IGF-I concentrations (Fig. 7B). Overall, non-transgenic fish had lower IGF-I mRNA levels than transgenic fish(primarily due to the lower levels found in the saline injected
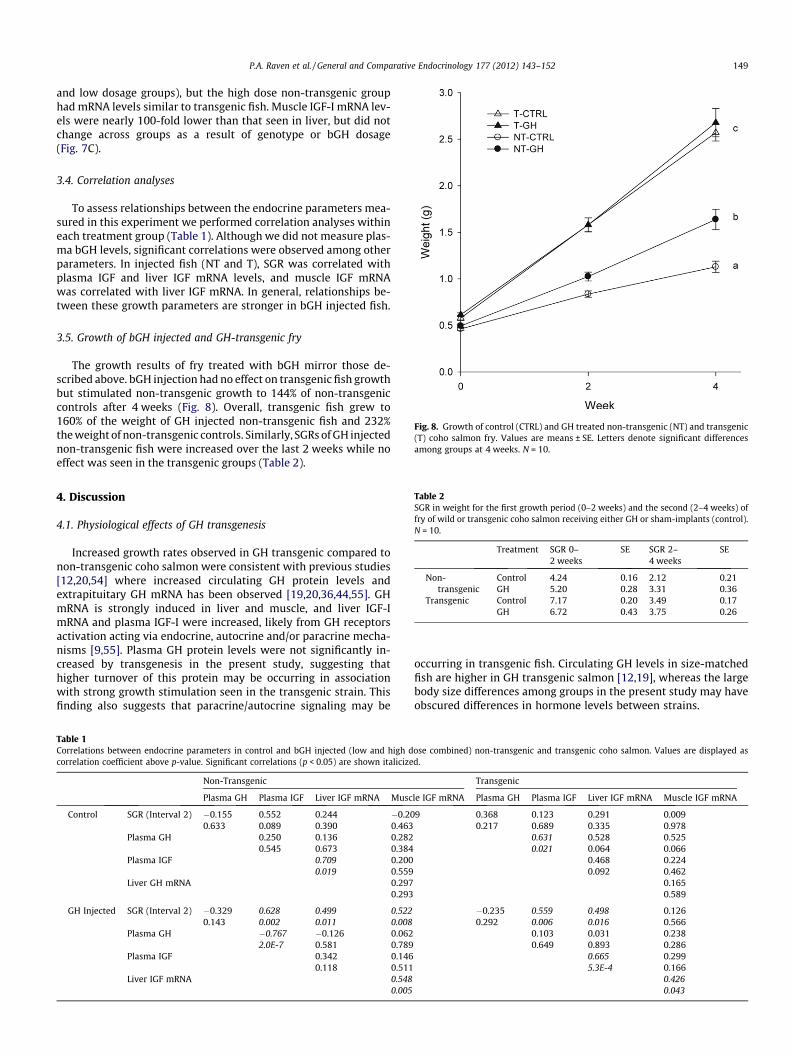
Fig. 8. Growth of control (CTRL) and GH treated non-transgenic (NT) and transgenic(T) coho salmon fry. Values are means ± SE. Letters denote significant differencesamong groups at 4 weeks. N = 10.
P.A. Raven et al. / General and Comparative Endocrinology 177 (2012) 143–152 149
and low dosage groups), but the high dose non-transgenic grouphad mRNA levels similar to transgenic fish. Muscle IGF-I mRNA lev-els were nearly 100-fold lower than that seen in liver, but did notchange across groups as a result of genotype or bGH dosage(Fig. 7C).
3.4. Correlation analyses
To assess relationships between the endocrine parameters mea-sured in this experiment we performed correlation analyses withineach treatment group (Table 1). Although we did not measure plas-ma bGH levels, significant correlations were observed among otherparameters. In injected fish (NT and T), SGR was correlated withplasma IGF and liver IGF mRNA levels, and muscle IGF mRNAwas correlated with liver IGF mRNA. In general, relationships be-tween these growth parameters are stronger in bGH injected fish.
3.5. Growth of bGH injected and GH-transgenic fry
The growth results of fry treated with bGH mirror those de-scribed above. bGH injection had no effect on transgenic fish growthbut stimulated non-transgenic growth to 144% of non-transgeniccontrols after 4 weeks (Fig. 8). Overall, transgenic fish grew to160% of the weight of GH injected non-transgenic fish and 232%the weight of non-transgenic controls. Similarly, SGRs of GH injectednon-transgenic fish were increased over the last 2 weeks while noeffect was seen in the transgenic groups (Table 2).
Table 2SGR in weight for the first growth period (0–2 weeks) and the second (2–4 weeks) offry of wild or transgenic coho salmon receiving either GH or sham-implants (control).N = 10.
Treatment SGR 0–2 weeks
SE SGR 2–4 weeks
SE
Non-transgenic
Control 4.24 0.16 2.12 0.21GH 5.20 0.28 3.31 0.36
Transgenic Control 7.17 0.20 3.49 0.17GH 6.72 0.43 3.75 0.26
4. Discussion
4.1. Physiological effects of GH transgenesis
Increased growth rates observed in GH transgenic compared tonon-transgenic coho salmon were consistent with previous studies[12,20,54] where increased circulating GH protein levels andextrapituitary GH mRNA has been observed [19,20,36,44,55]. GHmRNA is strongly induced in liver and muscle, and liver IGF-ImRNA and plasma IGF-I were increased, likely from GH receptorsactivation acting via endocrine, autocrine and/or paracrine mecha-nisms [9,55]. Plasma GH protein levels were not significantly in-creased by transgenesis in the present study, suggesting thathigher turnover of this protein may be occurring in associationwith strong growth stimulation seen in the transgenic strain. Thisfinding also suggests that paracrine/autocrine signaling may be
Table 1Correlations between endocrine parameters in control and bGH injected (low and high dcorrelation coefficient above p-value. Significant correlations (p < 0.05) are shown italicize
Non-Transgenic
Plasma GH Plasma IGF Liver IGF mRNA Musc
Control SGR (Interval 2) �0.1550.633
0.5520.089
0.2440.390
�0.200.463
Plasma GH 0.2500.545
0.1360.673
0.2820.384
Plasma IGF 0.7090.019
0.2000.559
Liver GH mRNA 0.2970.293
GH Injected SGR (Interval 2) �0.3290.143
0.6280.002
0.4990.011
0.5220.008
Plasma GH �0.7672.0E-7
�0.1260.581
0.0620.789
Plasma IGF 0.3420.118
0.1460.511
Liver IGF mRNA 0.5480.005
occurring in transgenic fish. Circulating GH levels in size-matchedfish are higher in GH transgenic salmon [12,19], whereas the largebody size differences among groups in the present study may haveobscured differences in hormone levels between strains.
ose combined) non-transgenic and transgenic coho salmon. Values are displayed asd.
Transgenic
le IGF mRNA Plasma GH Plasma IGF Liver IGF mRNA Muscle IGF mRNA
9 0.3680.217
0.1230.689
0.2910.335
0.0090.978
0.6310.021
0.5280.064
0.5250.066
0.4680.092
0.2240.4620.1650.589
�0.2350.292
0.5590.006
0.4980.016
0.1260.566
0.1030.649
0.0310.893
0.2380.286
0.6655.3E-4
0.2990.1660.4260.043

150 P.A. Raven et al. / General and Comparative Endocrinology 177 (2012) 143–152
4.2. Physiological effects of bGH treatment
Treatment of salmon with the slow-release formulations of re-combinant bovine growth hormone (bGH) effectively doubledgrowth rate for 8 weeks in non-transgenic fish (a single injectionat 2.23 mg rbGH/g BW resulted in a 57% increase in body weight).A higher dose had little further effect on growth apart from a minorelevation of growth rate beyond week 8. In fry, no effect of a higherdose on growth was seen. bGH injection has previously been foundto promote growth over an extended period (P20 weeks) in cohosalmon [41] and is effective at stimulating growth in rainbow trout[29], brown trout [34,64], Atlantic salmon [46], channel catfish[49,50] and tilapia [38]. On the other hand, GH transgenic fishare already growing at very rapid rates (greater than seen withbGH treatment alone) and did not respond in growth to bGH injec-tion regardless of the dose. Furthermore, endocrine parameterssuch as plasma GH and IGF, mRNA levels of liver and muscle GH,GHR, and IGF-I and mRNA levels of pituitary GH and GHR wereunaffected by bGH treatment. These results suggest that there isphysiological plasticity in the endocrine system of non-transgenicfish to allow for endocrine adjustment and growth enhancement,whereas GH transgenic fish (that already have high levels of sal-mon GH production) are at, or near, the physiological maximumgrowth rate that can be stimulated by GH alone.
Growth effects stimulated by GH in non-transgenic and trans-genic fish are believed to be mediated in part by IGF-I productionby the liver and other tissues [5,55]. In non-transgenic fish, therewas a general up-regulation of the GH-IGF-I axis with bGH dose.This was seen in an increase in plasma IGF-I and liver IGF-I mRNAlevels, and in a correlation between growth and IGF after bGHinjection. These findings are consistent with previous studiesinvolving bGH or bGH injection that have found increases in plas-ma IGF-I in rainbow trout [3,28], channel catfish [50] and tilapia[35], and increases in liver IGF-I mRNA levels in rainbow trout[28,57] and tilapia [35]. There is a negative correlation betweenGH and plasma IGF-I in bGH treated non-transgenic fish due tothe GH assay being specific to salmon GH. Salmon GH levels werereduced, presumably due to negative feedback effects from bGHtreatment. We did not measure total GH (i.e. both salmon andbGH together) which would likely correlate positively with IGF-I.Contrary to Gahr et al. [28] and Kajimura et al. [35] who found thatbGH injection increased muscle IGF-I mRNA, and similar to Peter-son et al. [50] who found it did not, we found no effect of bGHinjection on muscle IGF-I mRNA. This is possibly due to a drop inthe efficacy of the slow-release bGH formulation by week 14, or be-cause of effects of fish size differences among groups.
4.3. Pituitary involvement in GH regulation
Previous studies have shown that GH treatment can feedback tothe pituitary gland to negatively regulate GH mRNA and protein[2,5]. In the present study, GH mRNA levels as measured per assayinput RNA amount and per entire pituitary showed little differ-ences. Generally, pituitary size increases with body size in cohosalmon, but GH transgenic coho salmon have previously beenfound to have smaller pituitaries than non-transgenic fish of thesame size [43]. In the present study, when expressed per mg pitu-itary, GH mRNA levels were found to be less in transgenic andhigh-dose bGH-treated fish. This trend was not seen when GHmRNA is expressed as a proportion of pituitary RNA or on a perpituitary basis. These data are consistent with negative feedbackregulation of GH expression in the pituitary due to exogenousGH occurring at the whole-animal level. These data differ from thatseen by Mori and Devlin [44] with larger fish, where suppression ofpituitary GH mRNA was also seen at the level of proportion of totalRNA. The level of pituitary GH mRNA per g fish is actually less in
bGH-injected and transgenic fish than in non-transgenic fish, andconcomitantly plasma levels of GH were also lower in injectednon-transgenic fish as is seen in GH injected Atlantic salmon [46]and rainbow trout [6]. Transgenic fish do not exhibit a decreasein plasma GH with bGH injection likely due to high GH productionfrom the GH transgene and the lack of transgene promoter respon-siveness to negative feedback regulation of circulating GH and IGF-I. On the other hand, in some studies on non-transgenic speciessuch as rainbow trout [3] and tilapia [38], endogenous GH wasfound not to change due to bGH injection.
In rainbow trout pituitary cells treated with bGH, inhibition ofGH production occurred predominantly through IGF-I rather thanfrom bGH [7]. This mechanism is consistent with the presentexperiments where groups with the lowest GH mRNA/mg pituitarywere generally those with higher plasma IGF-I levels. It has alsobeen found that GH levels return to normal before changes inIGF-I following refeeding in rainbow trout, suggesting that IGF-Iis not responsible for GH production [27]. This appears to be thecase when looking at GH levels on a RNA or whole pituitary bases,but our results differ on a per mg and g fish basis. Thus, accountingfor differences in pituitary and fish size is important in this case.Pituitary GHR mRNA levels were lower in injected fish and loweragain in transgenic fish, possibly due to receptor down-regulationas a physiological response to desensitize the pituitary to thechronically high levels of circulating GH derived from the trans-gene. Other pituitary hormones involved in growth and metabo-lism, such as prolactin and somatolactin a, have been found tochange at the protein level in response to both GH treatment andGH-transgenesis [37]. Further studies on how these proteins havechanged and affect growth in GH-transgenic coho salmon iswarranted.
4.4. Functional differences in GH action
Cellular stimulation by GH is likely to differ between exogenousrecombinant bGH, transgene GH (sockeye salmon GH1), and nativecoho GH1 and GH2 in the experimental fish. Different somatotro-pins are known to affect fish with different potencies. For example,placental lactogen was found to be a strong stimulator of growth incoho salmon [14] perhaps due to its ability to activate GHR withoutrequiring dimerization [60]. Furthermore, bGH binds salmon GHRwith a 30-fold lower affinity than salmon GH [69]. In the presentexperiments, bGH-injected non-transgenic fish in this study didnot have plasma IGF-I nor growth rates elevated to that seen intransgenic fish, despite receiving a high saturating dose (basedon a lack of increase in growth with increasing dose of bGH). Dif-ferences in binding affinities, receptor activation mechanisms, orendocrine vs. paracrine stimulation, may influence the potency ofexogenous hormone treatments [8,9,55].
4.5. Conclusions
The present study has found that saturating doses of exogenousbovine GH delivered by intraperitoneal injection can stimulategrowth of non-transgenic salmon, but not to the extent seen fromendogenous overproduction of salmon GH by transgenesis. Despitedifferences in final body weight between the transgenic and non-transgenic groups we see similar trends as in other studies. Bothapproaches elevate IGF-I production and cause suppression ofendogenous host GH production, indicating significant modifica-tion of the normal GH/IGF-I growth axis. Additional GH did notstimulate the growth of transgenic fish, indicating their growthrates are at or near the maximal achievable by stimulation withGH. Other strains of GH transgenic salmon we have generatedcan show initially very high levels of growth (in wild or domesti-cated backgrounds) but ultimately have poor viability or display

P.A. Raven et al. / General and Comparative Endocrinology 177 (2012) 143–152 151
morphological abnormalities [12,13,21,22,48]. Thus, overexpres-sion of GH within a specific range is likely required to achieve max-imal growth stimulation without induction of pathologicalconditions. Whereas the application of transgenesis may be moreefficacious for growth enhancement, the use of slow-release for-mulations of GH do not carry a significant genetic risk and thusmay be preferable in some circumstances [18].
Acknowledgments
The authors wish to thank Kathy Cooper for performing theplasma GH assays. Funding to RHD from the Canadian RegulatorySystem for Biotechnology is gratefully acknowledged.
References
[1] M.V. Abrahams, A. Sutterlin, The foraging and antipredator behaviour ofgrowth-enhanced transgenic Atlantic salmon, Anim. Behav. 58 (1999) 933–942.
[2] T. Agustsson, B.T. Björnsson, Growth hormone inhibits growth hormonesecretion from the rainbow trout pituitary in vitro, Comp. Biochem. Physiol C(2000) 299–303.
[3] P.R. Biga, B.C. Peterson, G.T. Schelling, R.W. Hardy, K.D. Cain, K. Overturf, et al.,Bovine growth hormone treatment increased IGF-I in circulation and inducedthe production of a specific immune response in rainbow trout (Oncorhynchusmykiss), Aquaculture 246 (2005) 437–445.
[4] B.T. Björnsson, The biology of salmon growth hormone: from daylight todominance, Fish Physiol. Biochem. 17 (1997) 9–24.
[5] B.T. Björnsson, V. Johansson, S. Benedet, I.E. Einarsdottir, J. Hildahl, T.Agustsson, et al., Growth hormone endocrinology of salmonids: regulatorymechanisms and mode of action, Fish Physiol. Biochem. (2002) 227–242.
[6] B.T. Bjornsson, P. Kling, E. Jönsson, T.O. Nilsen, I.E. Einarsdottir, I. Rønnestad,et al., The role of growth hormone in growth, lipid homeostasis, energyutilization and partitioning in rainbow trout: interactions with leptin, ghrelinand insulin-like growth factor Gen Comp Endocrinol. In Press (2011).
[7] O. Blaise, C. Weil, P.Y. Le Bail, Role of IGF-I in the control of GH secretion inrainbow trout (Oncorhynchus mykiss), Growth Regul. 5 (1995) 142–150.
[8] A.A. Butler, D. LeRoith, Minireview: tissue-specific versus generalized genetargeting of the igf1 and igf1r genes and their roles in insulin-like growthfactor physiology, Endocrinology 142 (2001) 1685–1688.
[9] A. Caelers, N. Maclean, G. Hwang, E. Eppler, M. Reinecke, Expression ofendogenous and exogenous growth hormone (GH) messenger (m) RNA in aGH-transgenic tilapia (Oreochromis niloticus), Transgenic Res. 14 (2005) 95–104.
[10] J.T. Cook, M.A. McNiven, G.F. Richardson, A.M. Sutterlin, Growth rate, bodycomposition and feed digestibility/conversion of growth-enhanced transgenicAtlantic salmon (Salmo salar), Aquaculture 188 (2000) 15–32.
[11] R.G. Danzmann, G.J. Van Der Kraak, T.T. Chen, D.A. Powers, Metabolic effects ofbovine growth hormone and genetically engineered rainbow trout growthhormone in rainbow trout (Oncorhynchus mykiss) reared at high temperature,Can. J. Fish. Aquat. Sci. 47 (1990) 1292–1301.
[12] R.H. Devlin, C.A. Biagi, T.Y. Yesaki, Growth, viability and genetic characteristicsof GH transgenic coho salmon strains, Aquaculture 236 (2004) 607–632.
[13] R.H. Devlin, C.A. Biagi, T.Y. Yesaki, D.E. Smailus, J.C. Byatt, Growth ofdomesticated transgenic fish, Nature 409 (2001) 781–782.
[14] R.H. Devlin, J.C. Byatt, E. McLean, T.Y. Yesaki, G.G. Krivi, E.G. Jaworski, et al.,Bovine placental lactogen is a potent stimulator of growth and displays strongbinding to hepatic receptor sites of coho salmon, Gen. Comp. Endocrinol. 95(1994) 31–41.
[15] R.H. Devlin, M. D’Andrade, M. Uh, C.A. Biagi, Population effects of growthhormone transgenic coho salmon depend on food availability and genotype byenvironment interactions, Proc. Natl. Acad. Sci. 101 (2004) 9303–9308.
[16] R.H. Devlin, J.I. Johnsson, D.E. Smailus, C.A. Biagi, E. Jonsson, B.T. Bjornsson,Increased ability to compete for food by growth hormone-transgenic cohosalmon Oncorhynchus kisutch (Walbaum), Aquac. Res. 30 (1999) 479–482.
[17] R.H. Devlin, P.A. Raven, L.F. Sundström, M. Uh, Issues and methodology fordevelopment of transgenic fish for aquaculture with a focus on growthenhancement, in: K. Overturf (Ed.), Molecular Research in Aquaculture, Wiley-Blackwell, Ames, 2009, pp. 217–260.
[18] R.H. Devlin, L.F. Sundstrom, W.M. Muir, Interface of biotechnology and ecologyfor environmental risk assessments of transgenic fish, Trends Biotechnol. 24(2006) 89–97.
[19] R.H. Devlin, P. Swanson, W.C. Clarke, E. Plisetskaya, W. Dickhoff, S. Moriyama,et al., Seawater adaptability and hormone levels in growth-enhancedtransgenic coho salmon, Oncorhynchus kisutch, Aquaculture 191 (2000) 367–385.
[20] R.H. Devlin, T.Y. Yesaki, C.A. Biagi, E.M. Donaldson, P. Swanson, W.-K. Chan,Extraordinary salmon growth, Nature 371 (1994) 209–210.
[21] R.H. Devlin, T.Y. Yesaki, E.M. Donaldson, S.J. Du, C.-L. Hew, Production ofgermline transgenic Pacific salmonids with dramatically increased growthperformance, Can. J. Fish. Aquat. Sci. 52 (1995) 1376–1384.
[22] R.H. Devlin, T.Y. Yesaki, E.M. Donaldson, C.L. Hew, Transmission andphenotypic effects of an antifreeze/GH gene construct in coho salmon(Oncorhynchus kisutch), Aquaculture 137 (1995) 161–170.
[23] E.M. Donaldson, U.H.M. Fagerlund, D.A. Higgs, J.R. McBride, Hormonalenhancement of growth, in: W.S. Hoar, D.J. Randall, J.R. Brett (Eds.), FishPhysiology: Bioenergetics and Growth, Academic Press, New York, 1979, pp.455–597.
[24] S.J. Du, Z. Gong, G.L. Fletcher, M.A. Shears, M.J. King, D.R. Idler, et al., Growthenhancement in transgenic Atlantic salmon by use of an ‘‘all fish’’ chimericgrowth hormone gene construct, Biotechnology 10 (1992) 176–181.
[25] A. Farmanfarmaian, L.Z. Sun, Growth hormone effects on essential amino acidabsorption, muscle amino acid profile, N-retention and nutritionalrequirements of striped bass hybrids, Genet. Anal. Biomol. Eng. 15 (1999)107–113.
[26] A.R. Foster, D.F. Houlihan, C. Gray, F. Medale, B. Fauconneau, S.J. Kaushik, et al.,The effects of ovine growth hormone on protein turnover in rainbow trout,Gen. Comp. Endocrinol. 82 (1991) 111–120.
[27] J.C. Gabillard, B.B. Kamangar, N. Montserrat, Coordinated regulation of the GH/IGF system genes during refeeding in rainbow trout (Oncorhynchus mykiss), J.Endocrinol. 191 (2006) 15–24.
[28] S.A. Gahr, R.L. Vallejo, G.M. Weber, B.S. Shepherd, J.T. Silverstein, C.E. RexroadIII, Effects of short-term growth hormone treatment on liver and muscletranscriptomes in rainbow trout (Oncorhynchus mykiss), Physiol. Genomics. 32(2008) 380–392.
[29] M.J. Garber, K.G. Deyonge, J.C. Byatt, W.A. Lellis, D.C. Honeyfield, R.C. Bull, et al.,Dose-response effects of recombinant bovine somatotropin (Posilac-TM) ongrowth performance and body composition of two-year-old rainbow trout(Oncorhynchus mykiss), J. Anim. Sci. 73 (1995) 3216–3222.
[30] D. Higgs, U. Fagerlund, J. McBride, H. Dye, E. Donaldson, Influence ofcombinations of bovine growth hormone, 17alpha-methyltestosterone, andL-thyroxine on growth of yearling coho salmon (Oncorhynchus kisutch), Can. J.Zool. 55 (1977) 1048–1056.
[31] D.A. Higgs, E.M. Donaldson, H.M. Dye, J.R. McBride, Influence of bovine growthhormone and L-thyroxine on growth, muscle composition and histologicalstructure of the gonads, thyroid, pancreas and pituitary of coho salmon(Oncorhynchus kisutch), J. Fish. Res. Board Can. 33 (1976) 1585–1603.
[32] D.A. Higgs, E.M. Donaldson, J.R. McBride, H.M. Dye, Evaluation of the potentialfor using a chinook salmon Oncorhynchus-tshawytscha pituitary extract vsbovine growth hormone to enhance the growth of coho salmonOncorhynchus-kisutch, Can. J. Zool. 56 (1978) 1226–1231.
[33] Y. Hinits, B. Moav, Growth performance studies in transgenic Cyprinus carpio,Aquaculture 173 (1999) 285–296.
[34] J.I. Johnsson, E. Petersson, E. Jönsson, B.T. Björnsson, T. Järvi, Domestication andgrowth hormone alter growth patterns and antipredator behaviour in juvenilebrown trout, Salmo trutta, Can. J. Fish. Aquat. Sci. 53 (1996) 1546–1554.
[35] S. Kajimura, K. Uchida, T. Yada, L.G. Riley, J.C. Byatt, R.J. Collier, et al.,Stimulation of insulin-like growth factor-I production by recombinant bovinegrowth hormone in Mozambique tilapia, Oreochromis mossambicus, FishPhysiol. Biochem. 25 (2001) 221–230.
[36] D.Y. Kang, R.H. Devlin, Effects of 3,5,30-triiodo-L-thyronine (T3) and 6-n-propyl-2-thiouracil (PTU) on growth of GH-transgenic coho salmon,Oncorhynchus kisutch, Fish Physiol. Biochem. 29 (2004) 77–85.
[37] Y. Kurata, Y. Kimura, Y. Yamanaka, A. Ishikawa, H. Okamoto, T. Masaoka, et al.,Effects of growth hormone on the salmon pituitary proteome, J. Proteomics(2011).
[38] T.A. Leedom, K. Uchida, T. Yada, N.H. Richman, J.C. Byatt, R.J. Collier, et al.,Recombinant bovine growth hormone treatment of tilapia: growth response,metabolic clearance, receptor binding and immunoglobulin production,Aquaculture 207 (2002) 359–380.
[39] J.-K. Lu, B.-H. Fu, J.-L. Wu, T.T. Chen, Production of transgenic silver sea bream(Sparus sarba) by different gene transfer methods, Mar. Biotech. 4 (2002) 328–337.
[40] R. Martinez, M.P. Estrada, J. Berlanga, I. Guillen, O. Hernandez, R. Pimentel,et al., Growth enhancement in transgenic tilapia by ectopic expression oftilapia growth hormone, Mol. Mar. Biol. Biotech. 5 (1996) 62–70.
[41] E. McLean, R.H. Devlin, J.C. Byatt, W.C. Clarke, E.M. Donaldson, Impact of acontrolled release formulation of recombinant bovine growth hormone upongrowth and seawater adaptation in coho (Oncorhynchus kisutch) and chinook(Oncorhynchus tshawytscha) salmon, Aquaculture 156 (1997) 113–128.
[42] R. Morales, M.T. Herrera, A. Arenal, A. Cruz, O. Hernandez, R. Pimentel, et al.,Tilapia chromosomal growth hormone gene expression accelerates growth intransgenic zebrafish (Danio rerio), EJB Electron. J. Biotechnol. 4 (2001) 1–7.
[43] T. Mori, R.H. Devlin, GH transgene affects pituitary size and GH geneexpression in transgenic salmon, Zoological Science (Tokyo) 15 (1998) 7.
[44] T. Mori, R.H. Devlin, Transgene and host growth hormone gene expression inpituitary and nonpituitary tissues of normal and growth hormone transgenicsalmon, Mol. Cell. Endocrinol. 149 (1999) 129–139.
[45] Y.K. Nam, J.K. Noh, Y.S. Cho, H.J. Cho, K.-N. Cho, C.G. Kim, et al., Dramaticallyaccelerated growth and extraordinary gigantism of transgenic mud loachMisgurnus mizolepis, Transgenic Res. 10 (2001) 353–362.
[46] L. Neregård, L. Sundt-Hansen, K.U. Hindar, S. Einum, J.I. Johnsson, R.H. Devlin,et al., Wild Atlantic salmon Salmo salar L. strains have greater growth potentialthan a domesticated strain selected for fast growth, J. Fish Biol. 73 (2008) 79–95.
[47] J.D. Oakes, D.A. Higgs, J.G. Eales, R.H. Devlin, Influence of ration level on thegrowth performance and body composition of non-transgenic and growth-

152 P.A. Raven et al. / General and Comparative Endocrinology 177 (2012) 143–152
hormone-transgenic coho salmon (Oncorhynchus kisutch), Aquaculture 265(2007) 309–324.
[48] T.H. Ostenfeld, E. McLean, R.H. Devlin, Transgenesis changes body and headshape in Pacific salmon, J. Fish Biol. 52 (1998) 850–854.
[49] B.C. Peterson, B.C. Small, B.G. Bosworth, Effects of bovine growth hormone(Posilac(R)) on growth performance, body composition, and IGFBPs in twostrains of channel catfish, Aquaculture 232 (2004) 651–663.
[50] B.C. Peterson, G.C. Waldbieser, L. Bilodeau, Effects of recombinant bovinesomatotropin on growth and abundance of mRNA for IGF-I and IGF-II inchannel catfish (Ictalurus punctatus), J. Anim. Sci. 83 (2005) 816–824.
[51] A.L. Pierce, J.T. Dickey, D.A. Larsen, H. Fukada, P. Swanson, W.W. Dickhoff, Aquantitative real-time RT-PCR assay for salmon IGF-I mRNA, and itsapplication in the study of GH regulation of IGF-I gene expression inprimary culture of salmon hepatocytes, Gen. Comp. Endocrinol. 135 (2004)401–411.
[52] T.I. Pitkaenen, A. Krasnov, H. Teerijoki, H. Moelsae, Transfer of growthhormone (GH) transgenes into Arctic charr (Salvelinus alpinus L.) I. Growthresponse to various GH constructs. Genet. Anal., Biomol. Eng. 15 (1999) 91–98.
[53] M.A. Rahman, N. Maclean, Growth performance of transgenic tilapiacontaining an exogenous piscine growth hormone gene, Aquaculture 173(1999) 333–346.
[54] P.A. Raven, R.H. Devlin, D.A. Higgs, Influence of dietary digestible energycontent on growth, protein and energy utilization and body composition ofgrowth hormone transgenic and non-transgenic coho salmon (Oncorhynchuskisutch), Aquaculture 254 (2006) 730–747.
[55] P.A. Raven, M. Uh, D. Sakhrani, B.R. Beckman, K. Cooper, J. Pinter, et al.,Endocrine effects of growth hormone overexpression in transgenic cohosalmon, Gen. Comp. Endocrinol. 159 (2008) 26–37.
[56] A. Saera-Vila, J.-A. Calduch-Giner, J. Perez-Sanchez, Duplication of growthhormone receptor (GHR) in fish genome: gene organization andtranscriptional regulation of GHR type I and II in gilthead sea bream (Sparusaurata), Gen. Comp. Endocrinol. 142 (2005) 193–203.
[57] M. Shamblott, C. Cheng, D. Bolt, T. Chen, Appearance of insulin-like growthfactor mRNA in the liver and pyloric ceca of a teleost in response to exogenousgrowth hormone, Proc. Nat. Acad. Sci. 92 (1995) 6943–6946.
[58] M. Shimizu, P. Swanson, W.W. Dickhoff, free and protein-bound insulin-likegrowth factor-I (IGF-I) and IGF-binding proteins in plasma of coho salmon,Oncorhynchus kisutch, Gen. Comp. Endocrinol. 3 (1999) 398–405.
[59] J.M. Shrimpton, R.H. Devlin, E. McLean, J.C. Byatt, E.M. Donaldson, D.J. Randall,Increases in gill cytosolic corticosteroid receptor abundance and saltwatertolerance in juvenile coho salmon (Oncorhynchus kisutch) treated withgrowth hormone and placental lactogen, Gen. Comp. Endocrinol. 98 (1995)1–15.
[60] N.R. Staten, J.C. Byatt, G.G. Krivi, Ligand-specific dimerization of theextracellular domain of the bovine growth hormone receptor, J. Biol. Chem.268 (1993) 18467–18473.
[61] L.F. Sundström, R.H. Devlin, J.I. Johnsson, C.A. Biagi, Vertical position reflectsincreased feeding motivation in growth hormone transgenic coho salmon(Oncorhynchus kisutch), Ethology 109 (2003) 701–712.
[62] L.F. Sundström, M. Lõhmus, R.H. Devlin, J.I. Johnsson, C.A. Biagi, T. Bohlin,Feeding on profitable and unprofitable prey: comparing behaviour of growth-enhanced transgenic and normal coho salmon (Oncorhynchus kisutch),Ethology 110 (2004) 381–396.
[63] L.F. Sundström, M. Lohmus, J.I. Johnsson, R.H. Devlin, Growth hormonetransgenic salmon pay for growth potential with increased predationmortality, Proc. R. Soc. Lond. B (Suppl.) 271 (2004) S350–S352.
[64] L. Sundt-Hansen, L. Neregård, S. Einum, J. Höjesjö, B.T. Björnsson, K. Hindar,et al., Growth enhancement increases movement activity of brown trout in thewild, Funct. Ecol. 23 (2009) 551–558.
[65] P. Swanson, Radioimmunoassay of fish growth hormone, prolactin, andsomatolactin, in: P.W. Hochachka, T.P. Mommsen (Eds.), Biochemistry andMolecular Biology of Fishes Analytical Techniques 3, Elsevier Science,Amsterdam, 1994, pp. 545–565.
[66] N.M. Very, J.D. Kittilson, L.A. Norbeck, M.A. Sheridan, Isolation,characterization, and distribution of two cDNAs encoding for growthhormone receptor in rainbow trout (Oncorhynchus mykiss), Comp. Biochem.Physiol. Part B: Biochem. Mol. Biol. 140 (2005) 615–628.
[67] A.H. Weatherley, H.S. Gill, Growth increases produced by bovine growthhormone in grass pickerel Esox-Americanus-Vermiculatus Le Sueur and theunderlying dynamics of muscle fiber growth, Aquaculture 65 (1987) 55–66.
[68] K. Wille, E. McLean, J.S. Goddard, J.C. Byatt, Dietary lipid level and growthhormone alter growth and body conformation of blue tilapia, Oreochromisaureus, Aquaculture 209 (2002) 219–232.
[69] K. Yao, P. Niu, F. Le Gac, P.-Y. Le Bail, Presence of specific growth hormone bindingsites in rainbow trout (Oncorhynchus mykiss) tissues: characterization of thehepatic receptor, Gen. Comp. Endocrinol. 81 (1991) 72–82.




















