
Placental regulation of peptide hormone and growth factor activity by proMBP
10
BIOLOGY OF REPRODUCTION 84, 1077–1086 (2011) Published online before print 26 January 2011. DOI 10.1095/biolreprod.110.090209 Minireview Placental Regulation of Peptide Hormone and Growth Factor Activity by proMBP 1 Kathrin Weyer 3 and Simon Glerup 2,4 Departments of Anatomy 3 and Medical Biochemistry, 4 University of Aarhus, Aarhus, Denmark ABSTRACT The gene PRG2, encoding the proform of eosinophil major basic protein (proMBP), is one of the most highly expressed genes during human pregnancy, and low proMBP levels predict Down syndrome and poor pregnancy outcome. Reminiscent of a magnet, the primary structure of proMBP is extremely charge polarized, consisting of an N-terminal acidic propiece followed by a highly basic MBP domain in the C-terminal. Many tissues synthesize and secrete full-length proMBP, but only distinct cell types of the immune system process and store mature MBP in intracellular granules. MBP is released upon degranulation of eosinophil leukocytes and is toxic to bacteria, parasites, and mammalian cells. In contrast, proMBP is apparently nontoxic and functions in the inhibition of proteolysis and prohormone conversion. Recent research has revealed the complexity of proMBP biology and shed light on the process of MBP generation. ProMBP specifically forms disulfide-mediated, co- valent complexes with the metzincin metalloproteinase preg- nancy-associated plasma protein A (PAPPA) and the prohormone angiotensinogen (AGT). In both processes, PAPPA and AGT have reduced biological activity in the resulting complexes. In addition, proMBP is a component of high-molecular-weight AGT and, therefore, is potentially involved in the development of preeclampsia and in pregnancy-induced hypertension. angiotensin, growth factors, insulin-like growth factor receptor (IGF/IGF receptors), placenta, pregnancy INTRODUCTION The proteolytically processed eosinophil major basic protein (MBP; also known as PRG2) was first described as the major constituent of the crystalloid core of the eosinophil leukocyte granules. MBP is highly basic and toxic to both mammalian cells and parasites, and it functions in the defense against invading parasites. Hence, it was a surprise when a protein immunochemically similar to MBP was detected in the placenta and in the circulation of pregnant women [1, 2]. The protein was of higher molecular weight compared to MBP isolated from the eosinophil granules and was suggested to be disulfide bonded to other proteins [3]. Later, this protein was identified as the unprocessed form (the proform of eosinophil MBP [proMBP]) in disulfide-linked complexes with the metalloproteinase pregnancy-associated plasma protein A (PAPPA) [4] and the prohormone angiotensinogen (AGT) [5]. The Proform of Eosinophil MBP Major basic protein purified from the granules of eosinophil leukocytes has a molecular weight of 13.8 kDa and is highly basic (pI ¼ 10.9). However, analysis of an isolated cDNA clone revealed that MBP is synthesized as a precursor form of 222 residues, including a 16-residue signal peptide and a highly acidic propiece of 89 residues (Fig. 1) [6]. The propiece of human proMBP isolated from pregnancy serum is extensively and heterogeneously glycosylated with an average carbohy- drate content of 38% (w/w), including several O-linked and one N-linked glycosylation. In addition, the propiece is substituted with a glycosaminoglycan (GAG) of the heparan sulfate type [7]. Based on the amino acid sequence, the pI of the propiece was calculated to be 4.0 [6], but the GAG chain most likely contributes further to its acidity. Mature MBP is generated in the bone marrow by maturing eosinophils, and the propiece is believed to mask the cytotoxicity of MBP before proteolytic processing upon granule condensation [8]. In proMBP, the propiece is chained to the MBP domain by three intramolecular disulfide bonds, and these must consequently be disrupted by an unknown mechanism in maturing eosinophils in the bone marrow for the formation of mature MBP [9]. MBP Is an Unusual C-Type Lectin It is unclear how MBP exerts its cytotoxic and cytostimu- latory functions, but it has been proposed that cells are damaged by a direct interaction of the basic MBP with the bilayer lipid membrane [10], that it modulates enzymatic activity [11–13], or that it binds directly to specific receptors at the cell surface [14]. The MBP domain shares weak sequence identity with the C-type lectin (CTL) superfamily, and its crystal structure resembles canonical CTL domains, known from mannose-binding protein, E-selectin, tetranectin, lung surfactant protein, and the CTL domains of CD94 and CD69 [15]. MBP folds into one globular domain supported by two disulfide bonds and consists of two main a-helices and eight b- strands, with the latter forming three antiparallel b-sheets (Fig. 2). Members of the CTL superfamily generally bind carbohy- drate in a calcium-dependent manner, with the binding site formed by conserved acidic calcium ion-ligating residues brought together in the tertiary structure by three extended loops and a b-strand. However, these positions are occupied by basic residues in the putative CTL ligand-binding region of MBP, and it was suggested that natural ligands for MBP include sulfated sugars of the heparan sulfate type [15]. In fact, 1 Supported by the Danish Medical Research Council. 2 Correspondence: Simon Glerup, Department of Medical Biochemistry, Ole Worms Alle ´, Building 1170, University of Aarhus, DK-8000C, Aarhus, Denmark. FAX: 45 86131160; e-mail: [email protected] Received: 6 December 2010. First decision: 8 January 2011. Accepted: 10 January 2011. Ó 2011 by the Society for the Study of Reproduction, Inc. eISSN: 1529-7268 http://www.biolreprod.org ISSN: 0006-3363 1077 Downloaded from www.biolreprod.org.
Transcript of Placental regulation of peptide hormone and growth factor activity by proMBP

BIOLOGY OF REPRODUCTION 84, 1077–1086 (2011)Published online before print 26 January 2011.DOI 10.1095/biolreprod.110.090209
Minireview
Placental Regulation of Peptide Hormone and Growth Factor Activity by proMBP1
Kathrin Weyer3 and Simon Glerup2,4
Departments of Anatomy3 and Medical Biochemistry,4 University of Aarhus, Aarhus, Denmark
ABSTRACT
The gene PRG2, encoding the proform of eosinophil majorbasic protein (proMBP), is one of the most highly expressedgenes during human pregnancy, and low proMBP levels predictDown syndrome and poor pregnancy outcome. Reminiscent of amagnet, the primary structure of proMBP is extremely chargepolarized, consisting of an N-terminal acidic propiece followedby a highly basic MBP domain in the C-terminal. Many tissuessynthesize and secrete full-length proMBP, but only distinct celltypes of the immune system process and store mature MBP inintracellular granules. MBP is released upon degranulation ofeosinophil leukocytes and is toxic to bacteria, parasites, andmammalian cells. In contrast, proMBP is apparently nontoxicand functions in the inhibition of proteolysis and prohormoneconversion. Recent research has revealed the complexity ofproMBP biology and shed light on the process of MBPgeneration. ProMBP specifically forms disulfide-mediated, co-valent complexes with the metzincin metalloproteinase preg-nancy-associated plasma protein A (PAPPA) and the prohormoneangiotensinogen (AGT). In both processes, PAPPA and AGT havereduced biological activity in the resulting complexes. Inaddition, proMBP is a component of high-molecular-weightAGT and, therefore, is potentially involved in the developmentof preeclampsia and in pregnancy-induced hypertension.
angiotensin, growth factors, insulin-like growth factor receptor(IGF/IGF receptors), placenta, pregnancy
INTRODUCTION
The proteolytically processed eosinophil major basic protein(MBP; also known as PRG2) was first described as the majorconstituent of the crystalloid core of the eosinophil leukocytegranules. MBP is highly basic and toxic to both mammaliancells and parasites, and it functions in the defense againstinvading parasites. Hence, it was a surprise when a proteinimmunochemically similar to MBP was detected in theplacenta and in the circulation of pregnant women [1, 2].The protein was of higher molecular weight compared to MBPisolated from the eosinophil granules and was suggested to bedisulfide bonded to other proteins [3]. Later, this protein was
identified as the unprocessed form (the proform of eosinophilMBP [proMBP]) in disulfide-linked complexes with themetalloproteinase pregnancy-associated plasma protein A(PAPPA) [4] and the prohormone angiotensinogen (AGT) [5].
The Proform of Eosinophil MBP
Major basic protein purified from the granules of eosinophilleukocytes has a molecular weight of 13.8 kDa and is highlybasic (pI¼10.9). However, analysis of an isolated cDNA clonerevealed that MBP is synthesized as a precursor form of 222residues, including a 16-residue signal peptide and a highlyacidic propiece of 89 residues (Fig. 1) [6]. The propiece ofhuman proMBP isolated from pregnancy serum is extensivelyand heterogeneously glycosylated with an average carbohy-drate content of 38% (w/w), including several O-linked andone N-linked glycosylation. In addition, the propiece issubstituted with a glycosaminoglycan (GAG) of the heparansulfate type [7]. Based on the amino acid sequence, the pI ofthe propiece was calculated to be 4.0 [6], but the GAG chainmost likely contributes further to its acidity. Mature MBP isgenerated in the bone marrow by maturing eosinophils, and thepropiece is believed to mask the cytotoxicity of MBP beforeproteolytic processing upon granule condensation [8]. InproMBP, the propiece is chained to the MBP domain by threeintramolecular disulfide bonds, and these must consequently bedisrupted by an unknown mechanism in maturing eosinophilsin the bone marrow for the formation of mature MBP [9].
MBP Is an Unusual C-Type Lectin
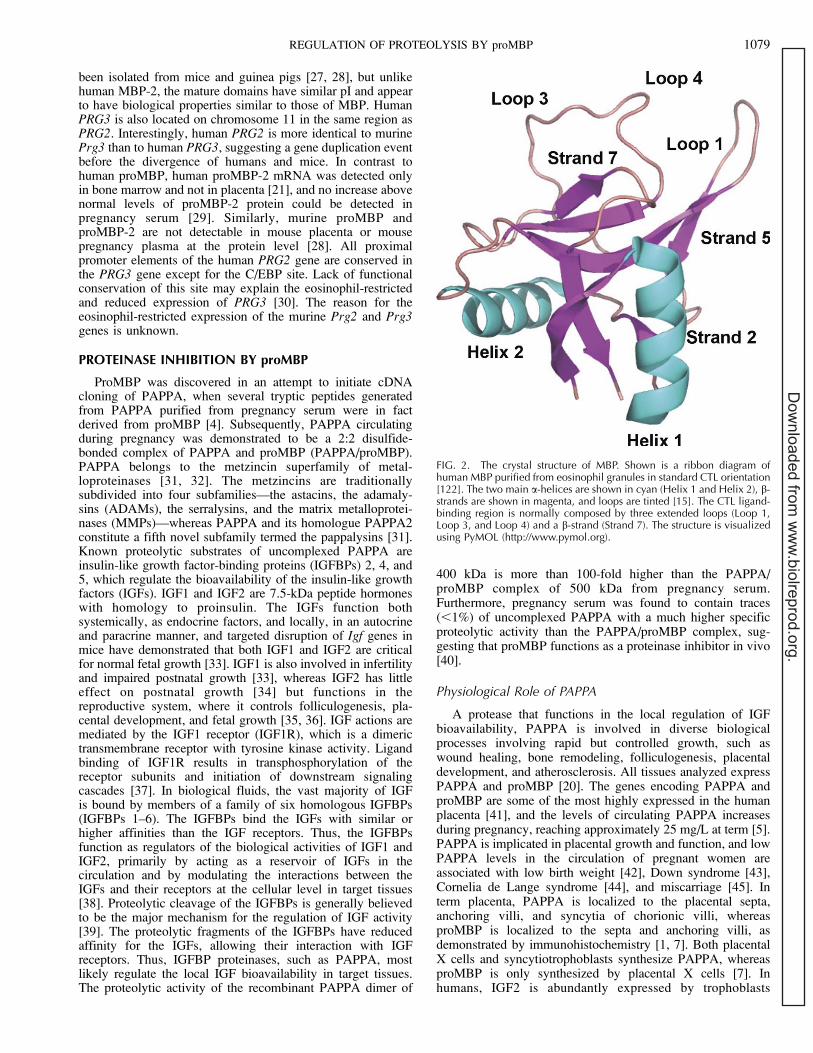
It is unclear how MBP exerts its cytotoxic and cytostimu-latory functions, but it has been proposed that cells aredamaged by a direct interaction of the basic MBP with thebilayer lipid membrane [10], that it modulates enzymaticactivity [11–13], or that it binds directly to specific receptors atthe cell surface [14]. The MBP domain shares weak sequenceidentity with the C-type lectin (CTL) superfamily, and itscrystal structure resembles canonical CTL domains, knownfrom mannose-binding protein, E-selectin, tetranectin, lungsurfactant protein, and the CTL domains of CD94 and CD69[15]. MBP folds into one globular domain supported by twodisulfide bonds and consists of two main a-helices and eight b-strands, with the latter forming three antiparallel b-sheets (Fig.2). Members of the CTL superfamily generally bind carbohy-drate in a calcium-dependent manner, with the binding siteformed by conserved acidic calcium ion-ligating residuesbrought together in the tertiary structure by three extendedloops and a b-strand. However, these positions are occupied bybasic residues in the putative CTL ligand-binding region ofMBP, and it was suggested that natural ligands for MBPinclude sulfated sugars of the heparan sulfate type [15]. In fact,
1Supported by the Danish Medical Research Council.2Correspondence: Simon Glerup, Department of Medical Biochemistry,Ole Worms Alle, Building 1170, University of Aarhus, DK-8000C,Aarhus, Denmark. FAX: 45 86131160; e-mail: [email protected]
Received: 6 December 2010.First decision: 8 January 2011.Accepted: 10 January 2011.� 2011 by the Society for the Study of Reproduction, Inc.eISSN: 1529-7268 http://www.biolreprod.orgISSN: 0006-3363
1077
Dow
nloaded from w
ww
.biolreprod.org.

heparin has previously been found to modulate MBP activity inseveral experimental setups [12–14]. Accordingly, based on x-ray crystallography and surface plasmon resonance experi-ments, the interaction of MBP with heparin and heparan sulfateGAG, but not other sulfated GAGs [16], was shown. Thebinding of cell surface receptors substituted with heparansulfate GAG could be an important step in the cytotoxic andcytostimulatory mechanism of MBP [16]. Such receptors maymediate the accumulation and oligomerization of MBP,promoting its interaction with phospholipids in the plasmamembrane and, thereby, membrane disruption and tissuedamage. Alternatively, a heparan sulfate proteoglycan mayfunction as a coreceptor in the delivery of noncytotoxicconcentrations of MBP to signaling receptors responsible forthe cytostimulatory effects of MBP. It was further proposedthat in addition to the neutralizing effect of the acidic residuesin the propiece, the GAG of the propiece interacts with theCTL ligand-binding region to inhibit its function [15].
ProMBP Cell Surface Binding
Because MBP is cytotoxic, its cell surface-binding proper-ties are difficult to study. However, proMBP was found to binddirectly to the cell surface, and the binding is, indeed, mediatedby a heparan sulfate proteoglycan [17]. It has been proposed inseveral articles and reviews [15] that the proMBP GAG inhibitscell surface-binding of proMBP. The proMBP cell surface-binding site is localized in the MBP domain but is outside theCTL ligand-binding region. This binding site is involved incrystal contacts in the asymmetric unit of the MBP crystal,possibly explaining why this binding was not identified in theanalysis of MBP crystals soaked with heparin disaccharide[16]. Thus, the CTL site is potentially available for otherinteractions. Interestingly, a previous study showed thatproMBP inhibited, in a dose-responsive manner, basophilhistamine release and superoxide anion generation stimulatedby MBP [18], suggesting that proMBP and MBP compete forthe same receptor. Peptide-mapping experiments identified tworegions involved in the cytotoxic and cytostimulatory actions
of MBP [19]. These two regions showed different propertiesregarding cytotoxicity and stimulation of basophil histaminerelease. The cytotoxicity of the region, corresponding toresidues 123–150 in proMBP (Fig. 1), was comparable to MBPand markedly higher than that in the second region, containedwithin the CTL ligand-binding region and corresponding toresidues 195–222 in proMBP. In contrast, this region was moreeffective in stimulating basophil histamine release.
The proMBP Gene
Although all tissues analyzed express human proMBP, thehighest expression by far is found in the bone marrow and inthe placenta [20, 21], suggesting specifically controlled geneexpression in the developing eosinophil and placenta [22]. Thehuman PRG2 gene encoding proMBP is localized to thecentromere of the 11q12 region of chromosome 11. The gene is3.3 kb and composed of six exons [23, 24], and the proximalpromoter sequence contains a TATA and CCAAT box, aGATA transcriptional factor-binding site, a CCAAT/enhancer-binding protein (C/EBP) site, two Ikaros-2 sites, and a signaltransducer and activator of transcription site (STAT) [22]. Theprimary structure of proMBP is known from humans, mice,rats, and guinea pigs, and it shares 50–65% sequence identityin the MBP domain but low sequence identity in the propiece(Fig. 1). Curiously, a cDNA encoding a protein, proMBP-2,homologous to proMBP, was isolated from umbilical cord stemcells stimulated with interleukin (IL) 5 [21], a cytokine thatcauses eosinophil proliferation, differentiation, and activation[25, 26]. The gene encoding proMBP-2, PRG3, was tran-scribed at a high level and was the fifth most abundanttranscript, corresponding to 1% of the total transcripts isolatedfrom IL5-stimulated umbilical cord stem cells. For comparison,the PRG2 gene was the most abundant and corresponded to 8%of total transcripts [21]. The calculated pI values of the maturehuman proteins differ by more than 100-fold, with pI values of11.4 and 8.7 for MBP and MBP-2, respectively. Accordingly,MBP-2 was found to have similar biological activities as MBPbut diminished potency [21]. ProMBP homologues have also
FIG. 1. Alignment of proMBP and proMBP-2 from human (hproMBP1 and hproMBP2), mouse (mproMBP1 and mproMBP2), rat (only rproMBP1), andguinea pig (gproMBP1 and gproMBP2). Conserved residues are shaded, and cysteines residues are in bold. Disulfide pairings of human proMBP [9] areindicated by connecting lines. Arrows indicate sites in the propiece of proteolytic cleavage during processing in the eosinophils [8].
1078 WEYER AND GLERUP
Dow
nloaded from w
ww
.biolreprod.org.
been isolated from mice and guinea pigs [27, 28], but unlikehuman MBP-2, the mature domains have similar pI and appearto have biological properties similar to those of MBP. HumanPRG3 is also located on chromosome 11 in the same region asPRG2. Interestingly, human PRG2 is more identical to murinePrg3 than to human PRG3, suggesting a gene duplication eventbefore the divergence of humans and mice. In contrast tohuman proMBP, human proMBP-2 mRNA was detected onlyin bone marrow and not in placenta [21], and no increase abovenormal levels of proMBP-2 protein could be detected inpregnancy serum [29]. Similarly, murine proMBP andproMBP-2 are not detectable in mouse placenta or mousepregnancy plasma at the protein level [28]. All proximalpromoter elements of the human PRG2 gene are conserved inthe PRG3 gene except for the C/EBP site. Lack of functionalconservation of this site may explain the eosinophil-restrictedand reduced expression of PRG3 [30]. The reason for theeosinophil-restricted expression of the murine Prg2 and Prg3genes is unknown.
PROTEINASE INHIBITION BY proMBP
ProMBP was discovered in an attempt to initiate cDNAcloning of PAPPA, when several tryptic peptides generatedfrom PAPPA purified from pregnancy serum were in factderived from proMBP [4]. Subsequently, PAPPA circulatingduring pregnancy was demonstrated to be a 2:2 disulfide-bonded complex of PAPPA and proMBP (PAPPA/proMBP).PAPPA belongs to the metzincin superfamily of metal-loproteinases [31, 32]. The metzincins are traditionallysubdivided into four subfamilies—the astacins, the adamaly-sins (ADAMs), the serralysins, and the matrix metalloprotei-nases (MMPs)—whereas PAPPA and its homologue PAPPA2constitute a fifth novel subfamily termed the pappalysins [31].Known proteolytic substrates of uncomplexed PAPPA areinsulin-like growth factor-binding proteins (IGFBPs) 2, 4, and5, which regulate the bioavailability of the insulin-like growthfactors (IGFs). IGF1 and IGF2 are 7.5-kDa peptide hormoneswith homology to proinsulin. The IGFs function bothsystemically, as endocrine factors, and locally, in an autocrineand paracrine manner, and targeted disruption of Igf genes inmice have demonstrated that both IGF1 and IGF2 are criticalfor normal fetal growth [33]. IGF1 is also involved in infertilityand impaired postnatal growth [33], whereas IGF2 has littleeffect on postnatal growth [34] but functions in thereproductive system, where it controls folliculogenesis, pla-cental development, and fetal growth [35, 36]. IGF actions aremediated by the IGF1 receptor (IGF1R), which is a dimerictransmembrane receptor with tyrosine kinase activity. Ligandbinding of IGF1R results in transphosphorylation of thereceptor subunits and initiation of downstream signalingcascades [37]. In biological fluids, the vast majority of IGFis bound by members of a family of six homologous IGFBPs(IGFBPs 1–6). The IGFBPs bind the IGFs with similar orhigher affinities than the IGF receptors. Thus, the IGFBPsfunction as regulators of the biological activities of IGF1 andIGF2, primarily by acting as a reservoir of IGFs in thecirculation and by modulating the interactions between theIGFs and their receptors at the cellular level in target tissues[38]. Proteolytic cleavage of the IGFBPs is generally believedto be the major mechanism for the regulation of IGF activity[39]. The proteolytic fragments of the IGFBPs have reducedaffinity for the IGFs, allowing their interaction with IGFreceptors. Thus, IGFBP proteinases, such as PAPPA, mostlikely regulate the local IGF bioavailability in target tissues.The proteolytic activity of the recombinant PAPPA dimer of
400 kDa is more than 100-fold higher than the PAPPA/proMBP complex of 500 kDa from pregnancy serum.Furthermore, pregnancy serum was found to contain traces(,1%) of uncomplexed PAPPA with a much higher specificproteolytic activity than the PAPPA/proMBP complex, sug-gesting that proMBP functions as a proteinase inhibitor in vivo[40].
Physiological Role of PAPPA
A protease that functions in the local regulation of IGFbioavailability, PAPPA is involved in diverse biologicalprocesses involving rapid but controlled growth, such aswound healing, bone remodeling, folliculogenesis, placentaldevelopment, and atherosclerosis. All tissues analyzed expressPAPPA and proMBP [20]. The genes encoding PAPPA andproMBP are some of the most highly expressed in the humanplacenta [41], and the levels of circulating PAPPA increasesduring pregnancy, reaching approximately 25 mg/L at term [5].PAPPA is implicated in placental growth and function, and lowPAPPA levels in the circulation of pregnant women areassociated with low birth weight [42], Down syndrome [43],Cornelia de Lange syndrome [44], and miscarriage [45]. Interm placenta, PAPPA is localized to the placental septa,anchoring villi, and syncytia of chorionic villi, whereasproMBP is localized to the septa and anchoring villi, asdemonstrated by immunohistochemistry [1, 7]. Both placentalX cells and syncytiotrophoblasts synthesize PAPPA, whereasproMBP is only synthesized by placental X cells [7]. Inhumans, IGF2 is abundantly expressed by trophoblasts
FIG. 2. The crystal structure of MBP. Shown is a ribbon diagram ofhuman MBP purified from eosinophil granules in standard CTL orientation[122]. The two main a-helices are shown in cyan (Helix 1 and Helix 2), b-strands are shown in magenta, and loops are tinted [15]. The CTL ligand-binding region is normally composed by three extended loops (Loop 1,Loop 3, and Loop 4) and a b-strand (Strand 7). The structure is visualizedusing PyMOL (http://www.pymol.org).
REGULATION OF PROTEOLYSIS BY proMBP 1079
Dow
nloaded from w
ww
.biolreprod.org.

invading the maternal decidua [46], and cell culture experi-ments have suggested that PAPPA and proMBP may play akey role in syncytiotrophoblast formation and extravilloustrophoblast invasion [47, 48]. Also indicating an important rolein fetal development, Pappa knockout mice are born at 60%the size of their wild-type littermates [49]. This phenotype issimilar to mice with IGF2-null mutations [34], and IGF2 hasbeen shown to be critical early in embryogenesis for optimalbody size [50]. Interestingly, IGF2 is an imprinted gene andonly expressed by the maternal allele, but disruption of IGF2imprinting rescues the dwarf phenotype of the Pappa knockoutmouse [51].
Outside pregnancy, PAPPA expression is particularlystimulated by proinflammatory cytokines. Treatment of humandermal fibroblast with tumor necrosis factor (TNF) and IL1Bincreased both PAPPA mRNA levels and IGFBP4 proteolyticactivity in the medium, while interferon-c treatment markedlyinhibited PAPPA expression [52]. These data could suggest apotential role of PAPPA in the acute inflammatory response ofwound-healing processes [53]. Indeed, PAPPA expression wasinduced in healing human skin wounds in dermal granulationtissue within and adjacent to the injury and colocalized withactivated macrophages, myofibroblasts, and new blood vessels.In uninjured skin, strong immunohistochemical staining forPAPPA was present in the epidermis, sweat, and sebaceousgland epithelial cells, hair follicles, and blood vessels [54].
The involvement of IGFs and IGFBPs, and of IGFBPs 2, 4,and 5 in particular, in ovarian folliculogenesis has beenextensively studied. Membrana granulosa cells of healthyfollicles express PAPPA, and the expression is positivelycorrelated with luteinizing hormone-induced processes ofdominant follicle development [55–57], suggesting thatPAPPA functions in the selection of the dominant follicle bythe regulation of IGF bioavailability. Indeed, female PAPPA-deficient mice display reduced fertility and ovarian steroido-genesis [58]. ProMBP has been detected in estrogen-dominantfollicular fluid but has not been detected in the medium ofcultured granulosa cells or luteinizing granulosa cells [59], andwhether it is associated with PAPPA is unknown.
Expression of PAPPA has been found in both primarycultures of ovarian surface epithelial cells and in unculturedovarian brushings, suggesting that PAPPA could function toinduce the proliferation of ovarian epithelial surface cells afterovulation and subsequent repair of the ovulation-associatedwound [60]. Interestingly, proMBP expression is induced uponthe transformation of normal ovarian surface epithelial cells tomalignant ovarian epithelial cells in a model for ovarianepithelial cancer, resulting in decreased IGFBP4 proteolyticactivity [60]. In addition, ovarian epithelial tumor samples, butnot uncultured ovarian brushings from healthy individuals,contained measurable proMBP mRNA levels. The physiolog-ical relevance of this observation is unclear, but tumorpromoters can increase cell differentiation rather than cellproliferation [61], and higher IGFBP4 levels through theinhibition of PAPPA by proMBP could reflect a morespecialized function of the tumor cells. On the other hand,invasive tumor cells could have decreased growth requirementsfor IGF-1, and high levels of IGFBP4 would limit the growthof the surrounding cells and enhance the invasive properties ofthe tumor cells.
Expression of PAPPA in human coronary artery smoothmuscle cells is also stimulated by TNF and IL1B [62] and alsoby IL6 [32], suggesting a mechanism for the regulation ofPAPPA expression contributing to the increased IGF1 activityobserved in intimal hyperplasia and atherosclerotic plaquedevelopment. Curiously, resveratrol, a polyphenol found in the
skin of grapes and associated with the beneficial cardiovasculareffects of red wine, inhibited TNF- and IL1B-induced PAPPAexpression [62]. PAPPA expression is also increased in pigcoronary arteries following balloon angioplasty [63], andsimilar up-regulation was observed upon vascular injury inmice [52]. Furthermore, intense immunostaining for PAPPAwas observed in ruptured and eroded human atheroscleroticplaques in association with activated macrophages and smoothmuscle cells [64]. Plaque rupture was accompanied by anincrease in plasma levels of the PAPPA dimer [64, 65]. Thus,PAPPA has the characteristics of a potential early diagnosticmarker for unstable angina. Importantly, Pappa knockout micehave increased life span [66] and are protected againstcardiovascular disease [67], whereas the development ofatherosclerosis is accelerated upon transgenic overexpressionof PAPPA in mice [68].
Both PAPPA and proMBP are also involved in boneformation and homeostasis. TNF and IL1B, in addition to IL4and TGFB, stimulate PAPPA expression and increase theIGFBP4 proteolytic activity in cultured human osteoblasts [69,70]. Interestingly, untreated osteoblasts expressed low levels ofproMBP, but the proMBP mRNA levels were completelysuppressed upon TGFB treatment [70], suggesting that TGFBcan amplify the biological response by both increasing PAPPAexpression and decreasing proMBP expression. TGFB and theIGFs belong to a subset of growth factors that are stored in thebone matrix and released during osteoclast resorption tomediate local coupling of bone formation and resorption[71]. The role of PAPPA in the formation of new bone hasrecently been verified in vivo in two studies describingincreased osteoblast proliferation in transgenic mice thatoverexpress PAPPA in osteoblasts [72] and delayed bonefracture repair in Pappa knockout mice [73]. Interestingly, thetreatment of fibroblasts and osteoblasts with phorbol estertumor promoters and their transformation with simian virus 40large T antigen resulted in inhibition of IGFBP4 proteolyticactivity without a reduction in PAPPA expression [74, 75].This was later demonstrated to be the result of inducedexpression of proMBP [76].
The Inhibitory Mechanism of proMBP
The PAPPA/proMBP complex contains close to 200cysteine residues. Of the 82 cysteine residues of each PAPPAsubunit in the PAPPA/proMBP complex, 78 participate inintramolecular disulfide bonds, whereas one participates in anintersubunit PAPPA/PAPPA disulfide bond and two participatein intermolecular disulfide bonds with proMBP [54]. Mutationof these cysteines in proMBP results in a proMBP variant thatis unable to form the covalent PAPPA/proMBP complex andinhibit the proteolytic activity of PAPPA [77]. Thus, theformation of intermolecular proteinase-inhibitor disulfidebonds is required for inhibition of proteolytic activity.Proteinase inhibitors normally function through noncovalentinteractions with their target proteinase, either by binding to theactive site in a substrate-like manner, by binding adjacent to theactive site or to an exosite, or by binding outside the active sitecausing inhibition through an allosteric mechanism. Theserpins and a
2-macroglobulin form covalent complexes with
their target proteinases [78, 79]. To our knowledge, however,an inhibitory mechanism based on the formation of intermo-lecular proteinase-inhibitor disulfide bonds has not beendescribed previously. Tissue inhibitors of metalloproteinases(TIMPs 1–4) are broad-spectrum endogenous inhibitors of allmembers of the MMP subfamily of the metzincin superfamilyin addition to some members of the ADAM subfamily [80].
1080 WEYER AND GLERUP
Dow
nloaded from w
ww
.biolreprod.org.

They function by forming a tight, binding, noncovalent 1:1complex through interactions with both the active site andexosites of the MMPs [81]. Only a few additional endogenousinhibitors have been described for the metzincin metal-loproteinases, and proMBP is the first described inhibitor ofa member of the pappalysin subfamily. Interestingly, thezymogens of many metzincin metalloproteinases are inhibitedby their propeptides through the interaction of a cysteine side-chain sulfur atom with the active-site zinc atom [82]. Severalcysteines of proMBP could potentially function in such acysteine-switch mechanism, but their substitution for serine didnot increase the proteolytic activity of the resulting mutantPAPPA/proMBP complex [77]. The inhibitory mechanism ofproMBP most likely is based on a combination of sterichindrance and allosteric inhibition. In fact, one of the twoPAPPA/proMBP disulfide bonds engages a cysteine locateddirectly in a Lin12-Notch module within the proteolyticdomain of PAPPA and, thereby, potentially obstructs asubstrate-specificity regulatory unit consisting of three Lin12-Notch modules organized in trans within the active PAPPAdimer [83, 84].
PAPPA/proMBP Complex Formation
Disulfide bonds are normally inserted into proteins duringfolding in the endoplasmic reticulum assisted by disulfideisomerases, and because of the oxidizing nature of theextracellular space, disulfide bonds in secreted proteins areconsidered to be very stable. Still, several examples ofregulatory mechanisms involving the reduction or formationof specific disulfide bonds in extracellular proteins are found[85]. The target cysteines in PAPPA are unpaired, whereas thereacting cysteines of proMBP engage in a disulfide bond.However, inspection of the PAPPA/proMBP disulfide structureshows that six cysteine residues of the proMBP subunit andtwo cysteine residues of the PAPPA subunit change their statusfrom the uncomplexed to the complexed states [9]. Thus, theseries of covalent events leading to the final PAPPA/proMBPcomplex is quite complex. First, the proMBP disulfide bondCys-51-Cys-169 is broken, exposing the reactive Cys-169,which then forms the first interchain disulfide with the PAPPAproteolytic domain cysteine Cys-652. Second, disruption of theproMBP disulfide Cys-89-Cys-128 is followed by Cys-51,forming the interchain disulfide with the PAPPA proteolyticdomain cysteine Cys-381. ProMBP is initially a monomer, butit dimerizes during the process of complex formation throughformation of the two interchain bonds, Cys-104-Cys-104 andCys-107-Cys-107, from two Cys-104-Cys-107 intrachaindisulfides [9]. The total redox potential is unchanged uponcomplex formation as each bond broken is accompanied byanother bond formed. Complex formation is dramaticallyaccelerated in a reducing environment, and in vivo, it mayinvolve a specific or a nonspecific catalyst, such as a disulfidereductase or a low-molecular-weight thiol compound. Such acatalyst could function to reduce disulfide bonds that arerendered susceptible as a consequence of an initial noncovalentinteraction between PAPPA and proMBP.
Detachment of PAPPA from the Cell Surface
The binding of PAPPA to a heparan sulfate proteoglycandirects its proteolytic activity to the surface of cells [86],suggesting that surface binding controls PAPPA-mediated IGFrelease and, consequently, IGF signaling spatially. The surface-binding site is localized to complement control protein (CCP)modules 3 and 4 in the C-terminal region of PAPPA, which
contains five consecutive CCP modules [86, 87]. Thesemodules, originally known from proteins associated with thecomplement system, are responsible for interactions withGAGs in several other proteins [88, 89]. The PAPPA/proMBPcomplex does not exhibit cell surface binding, but surfacebinding can be regained following treatment of the PAPPA/proMBP complex with heparinase [86]. As mentioned,proMBP is substituted with a heparan sulfate GAG chain[90], but unlike the formation of another covalent proteinase-inhibitor pair, thrombin-antithrombin-III, which criticallydepends on the binding of heparin-like GAG, creating atemplate for proteinase-inhibitor interaction [91, 92], theproMBP GAG does not mediate the initial recognition ofPAPPA. Instead, the recognition of PAPPA by proMBP isbased on protein-protein interactions and involves proMBPresidues located in the extended Loop 2 and in the loopbetween Strand 2 and Helix 2 of the MBP domain [93] (Fig. 2).The covalent PAPPA/proMBP complex is formed at the cellsurface by localizing the proMBP GAG in close proximity toCCP modules 3 and 4, allowing it to effectively compete withthe PAPPA receptor. Apparently, proMBP also loses affinityfor its surface receptor upon complex formation. Cys-169 islocated in the loop we identified as part of the proMBP surface-binding site. As mentioned above, Cys-169 switches fromparticipating in an intramolecular proMBP disulfide bond toforming the first intermolecular PAPPA/proMBP disulfidebond. This could potentially cause a conformational change ofthe loop, resulting in loss of proMBP surface binding.
Covalent complex formation between the serpins or a2-
macroglobulin and their target proteinases results in rapidclearance from the circulation [79, 94]. Surprisingly, the half-life of PAPPA and proMBP in complex is increased by several-fold compared to the rapid clearance of the uncomplexedcomponents. The slow clearance of PAPPA/proMBP maytherefore reflect an unknown function for the circulatingcomplex, and this observation is potentially interesting in lightof the depressed levels of total circulating PAPPA associatedwith Down syndrome [43] and low birth weight [42]. Hence,distinguishing between both forms of circulating PAPPA(uncomplexed PAPPA vs. PAPPA/proMBP) may be ofdiagnostic value for pregnancy outcome.
The rapid removal of circulating uncomplexed PAPPA andproMBP could indicate that the PAPPA/proMBP complex inhuman pregnancy is formed at the surface of placental cellsrather than in the circulation. Notably, approximately 1% ofPAPPA in pregnancy serum is uncomplexed and proteolyti-cally active [40]. The high clearance rate indicates that theturnover of uncomplexed PAPPA is higher than would beexpected from this fraction. Taken together, PAPPA inhibitionmay be important for the regulation of IGF activity in vivo, andthe growth-promoting effect of PAPPA possibly is onlyproperly antagonized by proMBP on cells also expressing theproMBP receptor. It can further be speculated that pathologicalconditions of altered redox potential, such as placental hypoxiain preeclampsia, may change the balance between complexedand uncomplexed PAPPA and, consequently, IGF signaling.
HIGH-MOLECULAR-WEIGHT AGT
ProMBP circulating during pregnancy exists in a molarexcess compared to PAPPA, and some of this was purified andidentified as disulfide-bonded 2:2 complexes of proMBP/AGTand 2:2:2 complexes of proMBP/AGT/C3dg [5]. AGT is aprohormone, and proteolytic cleavage of the N-terminal by theaspartyl proteinase renin results in the generation of thedecapeptide angiotensin-1 (Ang-1), which is the first and rate-
REGULATION OF PROTEOLYSIS BY proMBP 1081
Dow
nloaded from w
ww
.biolreprod.org.

limiting step of the renin-angiotensin system (RAS) (Fig. 3)[95]. Cells of the liver continuously secrete circulating AGT atconcentrations close to the Michaelis constant (K
m) of the
reaction [96, 97]. Renin, in turn, is stored in granules of thejuxtaglomerular cells in the kidney and is secreted in responseto decreased blood volume or sodium levels. Angiotensin-converting enzyme, primarily present in the lung endothelium,further cleaves Ang-1, leading to the generation of theoctapeptide angiotensin-2 (Ang-2), the physiologically activecomponent of the system. Ang-2 regulates blood pressuredirectly by stimulating kidney Ang-2 type 1 receptors (AT-1),thereby modulating renal sodium and water reabsorption, andindirectly by stimulating the production and release ofaldosterone from the adrenal glands and by stimulating thesensation of thirst in the central nervous system [98]. Theproteins associated with RAS are linked to a number ofcardiovascular and renal disorders, in particular hypertension.In contrast to AT-1 receptors that are present in adultcardiovascular tissue, the Ang-2 type 2 receptors are highlyexpressed during fetal development and may mediate vasodi-lator responses. In addition to their function in the cardiovas-cular system, all RAS components are also synthesized locallyin numerous organs, and Ang-2 and derived peptides actdirectly at the cellular level in a paracrine or autocrine manner,influencing processes such as cell growth, proliferation, andapoptosis [99].
Normally, the majority of AGT circulates as a monomer ofapproximately 54 kDa, in addition to a minor fraction of highermolecular weight (HMW AGT). The HMW AGT fractionincreases in the circulation of women on oral contraceptives(10%) and during pregnancy (16%), and it may become thepredominant form in pathological conditions, such as hyper-tension and preeclampsia [100–102]. HMW AGT is a majorcomponent of the placental-specific RAS and is the predom-inant form in amniotic fluid and placental extracts, and theincreased fraction of circulating HMW AGT is believed to beof placental origin [103, 104]. Although the HMW AGT formwas first described 30 years ago, its formation, physiologicalfunction, and relation to disease are poorly understood. TheHMW AGT fraction has typically been monitored by gelfiltration of biological fluids or tissue extracts followed by theincubation of fractions with renin and quantification of thegenerated Ang-1 by a radioimmunoassay [101–103, 105–107].The molecular weight of the HMW AGT species is in the rangeof 200–500 kDa, but upon reduction, monomeric AGT isgenerated, suggesting that it consists of at least one subunit ofthe AGT monomer covalently linked through disulfide bondsto additional unknown proteins or other AGT molecules [104,108, 109].
The presence of proMBP/AGT complexes has beendemonstrated in plasma of both normal and pregnantindividuals [5, 110]. The complex formation between AGTand proMBP has been studied and was shown to form uponcoexpression in mammalian cells and to depend on Cys-232 ofAGT [111]. Interestingly, Ang-1 generation by renin from therecombinant AGT/proMBP complex was slower than that fromthe AGT monomer [111], rendering proMBP the first exampleof a natural inhibitor of Ang-1 generation. Ang-1 generation isthe first step of RAS, and regulation at this level willconsequently affect all endocrine and paracrine functions ofAng-2 and additional peptides derived from Ang-1 (Fig. 3).HMW AGT complex formation may function as a clearancemechanism for AGT in response to increased Ang-2 signaling.Thus, complex formation of AGT/proMBP/C3dg and subse-quent binding and internalization by complement receptor 2[112] potentially assist in removal of excess AGT. In this way,the increased fraction of circulating HMW AGT associatedwith hypertension and preeclampsia might be a response to,rather than the cause of, the pathological condition.
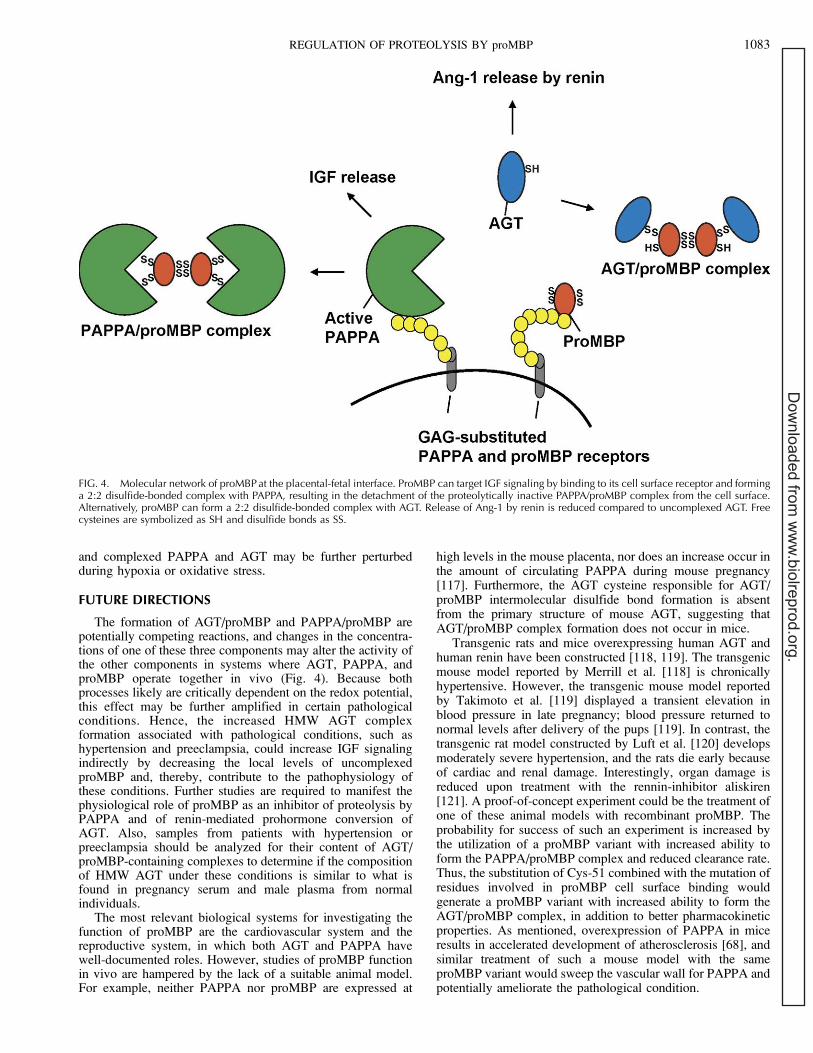
The origin of circulating HMW AGT in normal orhypertensive individuals is unknown, but most tissues knownto express local RAS also synthesize proMBP [20, 99]. HMWAGT circulating during pregnancy is probably derived from theplacental RAS, and AGT is associated with the surface ofplacental and renal-derived cells [113, 114]. However, proMBPinteracts with a cell surface heparan sulfate proteoglycan [17].Thus, it is tempting to speculate that HMW AGT complexformation operates locally at the surface of cells to regulate theparacrine and autocrine functions of RAS and that its formationcould be induced in response to increased Ang-2 signaling. Inthis view, it is interesting that local C3 production is increasedin Ang-2-induced organ damage [115]. Furthermore, it hasbeen speculated that the formation of the PAPPA/proMBPcomplex and AGT/proMBP complex are competing reactions[48, 116]. As a consequence, increased HMW AGT complexformation in tissues where PAPPA and AGT operate together,such as the cardiovascular system and the reproductive system,could contribute indirectly to the pathophysiology of condi-tions like hypertension and preeclampsia by increasing theamount of uncomplexed active PAPPA and, hence, IGFsignaling (Fig. 4). Such a natural balance of uncomplexed
FIG. 3. The renin-angiotensin system (RAS). In response to low bloodpressure, the kidneys release the aspartyl proteinase renin. Renin cleavescirculating AGT at a specific site at the N-terminal, releasing thedecapeptide Ang-1. Ang-1 is further cleaved by the metalloproteinaseangiotensin-converting enzyme (ACE) to the octapeptide Ang-2. Ang-2causes vasoconstriction and triggers the release of aldosterone from theadrenal gland. Aldosterone causes the kidney to retain sodium and excretepotassium. The components of RAS are also synthesized locally in tissuesand Ang-2, and derived peptides act in a paracrine or autocrine manner toinfluence a number of cellular processes [99].
1082 WEYER AND GLERUP
Dow
nloaded from w
ww
.biolreprod.org.
and complexed PAPPA and AGT may be further perturbedduring hypoxia or oxidative stress.
FUTURE DIRECTIONS
The formation of AGT/proMBP and PAPPA/proMBP arepotentially competing reactions, and changes in the concentra-tions of one of these three components may alter the activity ofthe other components in systems where AGT, PAPPA, andproMBP operate together in vivo (Fig. 4). Because bothprocesses likely are critically dependent on the redox potential,this effect may be further amplified in certain pathologicalconditions. Hence, the increased HMW AGT complexformation associated with pathological conditions, such ashypertension and preeclampsia, could increase IGF signalingindirectly by decreasing the local levels of uncomplexedproMBP and, thereby, contribute to the pathophysiology ofthese conditions. Further studies are required to manifest thephysiological role of proMBP as an inhibitor of proteolysis byPAPPA and of renin-mediated prohormone conversion ofAGT. Also, samples from patients with hypertension orpreeclampsia should be analyzed for their content of AGT/proMBP-containing complexes to determine if the compositionof HMW AGT under these conditions is similar to what isfound in pregnancy serum and male plasma from normalindividuals.
The most relevant biological systems for investigating thefunction of proMBP are the cardiovascular system and thereproductive system, in which both AGT and PAPPA havewell-documented roles. However, studies of proMBP functionin vivo are hampered by the lack of a suitable animal model.For example, neither PAPPA nor proMBP are expressed at
high levels in the mouse placenta, nor does an increase occur inthe amount of circulating PAPPA during mouse pregnancy[117]. Furthermore, the AGT cysteine responsible for AGT/proMBP intermolecular disulfide bond formation is absentfrom the primary structure of mouse AGT, suggesting thatAGT/proMBP complex formation does not occur in mice.
Transgenic rats and mice overexpressing human AGT andhuman renin have been constructed [118, 119]. The transgenicmouse model reported by Merrill et al. [118] is chronicallyhypertensive. However, the transgenic mouse model reportedby Takimoto et al. [119] displayed a transient elevation inblood pressure in late pregnancy; blood pressure returned tonormal levels after delivery of the pups [119]. In contrast, thetransgenic rat model constructed by Luft et al. [120] developsmoderately severe hypertension, and the rats die early becauseof cardiac and renal damage. Interestingly, organ damage isreduced upon treatment with the rennin-inhibitor aliskiren[121]. A proof-of-concept experiment could be the treatment ofone of these animal models with recombinant proMBP. Theprobability for success of such an experiment is increased bythe utilization of a proMBP variant with increased ability toform the PAPPA/proMBP complex and reduced clearance rate.Thus, the substitution of Cys-51 combined with the mutation ofresidues involved in proMBP cell surface binding wouldgenerate a proMBP variant with increased ability to form theAGT/proMBP complex, in addition to better pharmacokineticproperties. As mentioned, overexpression of PAPPA in miceresults in accelerated development of atherosclerosis [68], andsimilar treatment of such a mouse model with the sameproMBP variant would sweep the vascular wall for PAPPA andpotentially ameliorate the pathological condition.
FIG. 4. Molecular network of proMBP at the placental-fetal interface. ProMBP can target IGF signaling by binding to its cell surface receptor and forminga 2:2 disulfide-bonded complex with PAPPA, resulting in the detachment of the proteolytically inactive PAPPA/proMBP complex from the cell surface.Alternatively, proMBP can form a 2:2 disulfide-bonded complex with AGT. Release of Ang-1 by renin is reduced compared to uncomplexed AGT. Freecysteines are symbolized as SH and disulfide bonds as SS.
REGULATION OF PROTEOLYSIS BY proMBP 1083
Dow
nloaded from w
ww
.biolreprod.org.

REFERENCES
1. Maddox DE, Kephart GM, Coulam CB, Butterfield JH, Benirschke K,Gleich GJ. Localization of a molecule immunochemically similar toeosinophil major basic protein in human placenta. J Exp Med 1984;160:29–41.
2. Maddox DE, Butterfield JH, Ackerman SJ, Coulam CB, Gleich GJ.Elevated serum levels in human pregnancy of a molecule immunochem-ically similar to eosinophil granule major basic protein. J Exp Med 1983;158:1211–1226.
3. Wasmoen TL, McKean DJ, Benirschke K, Coulam CB, Gleich GJ.Evidence of eosinophil granule major basic protein in human placenta. JExp Med 1989; 170:2051–2063.
4. Oxvig C, Sand O, Kristensen T, Gleich GJ, Sottrup-Jensen L. Circulatinghuman pregnancy-associated plasma protein-A is disulfide-bridged to theproform of eosinophil major basic protein. J Biol Chem 1993;268:12243–12246.
5. Oxvig C, Haaning J, Kristensen L, Wagner JM, Rubin I, Stigbrand T,Gleich GJ, Sottrup-Jensen L. Identification of angiotensinogen andcomplement C3dg as novel proteins binding the proform of eosinophilmajor basic protein in human pregnancy serum and plasma. J Biol Chem1995; 270:13645–13651.
6. Barker RL, Gleich GJ, Pease LR. Acidic precursor revealed in humaneosinophil granule major basic protein cDNA. J Exp Med 1988;168:1493–1498.
7. Bonno M, Oxvig C, Kephart GM, Wagner JM, Kristensen T, Sottrup-Jensen L, Gleich GJ. Localization of pregnancy-associated plasmaprotein-A and colocalization of pregnancy-associated plasma protein-Amessenger ribonucleic acid and eosinophil granule major basic proteinmessenger ribonucleic acid in placenta. Lab Invest 1994; 71:560–566.
8. Popken-Harris P, Checkel J, Loegering D, Madden B, Springett M,Kephart G, Gleich GJ. Regulation and processing of a precursor form ofeosinophil granule major basic protein (ProMBP) in differentiatingeosinophils. Blood 1998; 92:623–631.
9. Glerup S, Boldt HB, Overgaard MT, Sottrup-Jensen L, Giudice LC,Oxvig C. Proteinase inhibition by proform of eosinophil major basicprotein (pro-MBP) is a multistep process of intra- and intermoleculardisulfide rearrangements. J Biol Chem 2005; 280:9823–9832.
10. Abu-Ghazaleh RI, Gleich GJ, Prendergast FG. Interaction of eosinophilgranule major basic protein with synthetic lipid bilayers: a mechanism fortoxicity. J Membr Biol 1992; 128:153–164.
11. Hastie AT, Loegering DA, Gleich GJ, Kueppers F. The effect of purifiedhuman eosinophil major basic protein on mammalian ciliary activity. AmRev Respir Dis 1987; 135:848–853.
12. Temkin V, Aingorn H, Puxeddu I, Goldshmidt O, Zcharia E, Gleich GJ,Vlodavsky I, Levi-Schaffer F. Eosinophil major basic protein: firstidentified natural heparanase-inhibiting protein. J Allergy Clin Immunol2004; 113:703–709.
13. Weiler JM, Gleich GJ. Eosinophil granule major basic protein regulatesgeneration of classical and alternative-amplification pathway C3convertases in vitro. J Immunol 1988; 140:1605–1610.
14. Jacoby DB, Gleich GJ, Fryer AD. Human eosinophil major basic proteinis an endogenous allosteric antagonist at the inhibitory muscarinic M2receptor. J Clin Invest 1993; 91:1314–1318.
15. Swaminathan GJ, Weaver AJ, Loegering DA, Checkel JL, Leonidas DD,Gleich GJ, Acharya KR. Crystal structure of the eosinophil major basicprotein at 1.8 A. An atypical lectin with a paradigm shift in specificity. JBiol Chem 2001; 276:26197–26203.
16. Swaminathan GJ, Myszka DG, Katsamba PS, Ohnuki LE, Gleich GJ,Acharya KR. Eosinophil-granule major basic protein, a C-type lectin,binds heparin. Biochemistry 2005; 44:14152–14158.
17. Glerup S, Kloverpris S, Oxvig C. The proform of the eosinophil majorbasic protein binds the cell surface through a site distinct from its C-typelectin ligand-binding region. J Biol Chem 2006; 281:31509–31516.
18. Popken-Harris P, McGrogan M, Loegering DA, Checkel JL, Kubo H,Thomas LL, Moy JN, Sottrup-Jensen L, Snable JL, Kikuchi MT.Expression, purification, and characterization of the recombinant proformof eosinophil granule major basic protein. J Immunol 1995; 155:1472–1480.
19. Thomas LL, Kubo H, Loegering DJ, Spillard K, Weaver AJ, McCormickDJ, Weiler C, Gleich GJ. Peptide-based analysis of amino acid sequencesimportant to the biological activity of eosinophil granule major basicprotein. Immunol Lett 2001; 78:175–181.
20. Overgaard MT, Oxvig C, Christiansen M, Lawrence JB, Conover CA,Gleich GJ, Sottrup-Jensen L, Haaning J. Messenger ribonucleic acidlevels of pregnancy-associated plasma protein-A and the proform of
eosinophil major basic protein: expression in human reproductive andnonreproductive tissues. Biol Reprod 1999; 61:1083–1089.
21. Plager DA, Loegering DA, Weiler DA, Checkel JL, Wagner JM, ClarkeNJ, Naylor S, Page SM, Thomas LL, Akerblom I, Cocks B, Stuart S,Gleich GJ. A novel and highly divergent homolog of human eosinophilgranule major basic protein. J Biol Chem 1999; 274:14464–14473.
22. Plager DA, Weiler DA, Loegering DA, Johnson WB, Haley L, Eddy RL,Shows TB, Gleich GJ. Comparative structure, proximal promoterelements, and chromosome location of the human eosinophil majorbasic protein genes. Genomics 2001; 71:271–281.
23. Li MS, Sun L, Satoh T, Fisher LM, Spry CJ. Human eosinophil majorbasic protein, a mediator of allergic inflammation, is expressed byalternative splicing from two promoters. Biochem J 1995; 305(pt 3):921–927.
24. Yamaguchi Y, Ackerman SJ, Minegishi N, Takiguchi M, Yamamoto M,Suda T. Mechanisms of transcription in eosinophils: GATA-1, but notGATA-2, transactivates the promoter of the eosinophil granule majorbasic protein gene. Blood 1998; 91:3447–3458.
25. Enokihara H, Nagashima S, Noma T, Kajitani H, Hamaguchi H, Saito K,Furusawa S, Shishido H, Honjo T. Effect of human recombinantinterleukin 5 and G-CSF on eosinophil colony formation. Immunol Lett1988; 18:73–76.
26. Ten RM, Butterfield JH, Kita H, Weiler DA, Fischkoff S, Ishizaka T,Sanderson CJ, Gleich GJ. Eosinophil differentiation of human umbilicalcord mononuclear cells and prolonged survival of mature eosinophils bymurine EL-4 thymoma cell conditioned medium. Cytokine 1991; 3:350–359.
27. Aoki I, Shindoh Y, Nishida T, Nakai S, Hong YM, Mio M, Saito T,Tasaka K. Sequencing and cloning of the cDNA of guinea pig eosinophilmajor basic protein. FEBS Lett 1991; 279:330–334.
28. Macias MP, Welch KC, Denzler KL, Larson KA, Lee NA, Lee JJ.Identification of a new murine eosinophil major basic protein (mMBP)gene: cloning and characterization of mMBP-2. J Leukoc Biol 2000;67:567–576.
29. Plager DA, Loegering DA, Checkel JL, Tang J, Kephart GM, Caffes PL,Adolphson CR, Ohnuki LE, Gleich GJ. Major basic protein homolog(MBP2): a specific human eosinophil marker. J Immunol 2006;177:7340–7345.
30. Plager DA, Adolphson CR, Gleich GJ. A novel human homolog ofeosinophil major basic protein. Immunol Rev 2001; 179:192–202.
31. Boldt HB, Overgaard MT, Laursen LS, Weyer K, Sottrup-Jensen L,Oxvig C. Mutational analysis of the proteolytic domain of pregnancy-associated plasma protein-A (PAPP-A): classification as a metzincin.Biochem J 2001; 358(pt 2):359–367.
32. Boldt HB, Conover CA. Pregnancy-associated plasma protein-A (PAPP-A): a local regulator of IGF bioavailability through cleavage of IGFBPs.Growth Horm IGF Res 2007; 17:10–18.
33. Liu JP, Baker J, Perkins AS, Robertson EJ, Efstratiadis A. Mice carryingnull mutations of the genes encoding insulin-like growth factor I (Igf-1)and type 1 IGF receptor (Igf1r). Cell 1993; 75:59–72.
34. DeChiara TM, Efstratiadis A, Robertson EJ. A growth-deficiencyphenotype in heterozygous mice carrying an insulin-like growth factorII gene disrupted by targeting. Nature 1990; 345(6270):78–80.
35. Constancia M, Hemberger M, Hughes J, Dean W, Ferguson-Smith A,Fundele R, Stewart F, Kelsey G, Fowden A, Sibley C, Reik W. Placental-specific IGF-II is a major modulator of placental and fetal growth. Nature2002; 417:945–948.
36. Giudice LC. Insulin-like growth factor family in graafian follicledevelopment and function. J Soc Gynecol Investig 2001; 8(1suppl):S26–S29.
37. Dupont J, LeRoith D. Insulin and insulin-like growth factor I receptors:similarities and differences in signal transduction. Horm Res 2001;55(suppl 2):22–26.
38. Firth SM, Baxter RC. Cellular actions of the insulin-like growth factorbinding proteins. Endocr Rev 2002; 23:824–854.
39. Bunn RC, Fowlkes JL. Insulin-like growth factor binding proteinproteolysis. Trends Endocrinol Metab 2003; 14:176–181.
40. Overgaard MT, Haaning J, Boldt HB, Olsen IM, Laursen LS, Christian-sen M, Gleich GJ, Sottrup-Jensen L, Conover CA, Oxvig C. Expressionof recombinant human pregnancy-associated plasma protein-A andidentification of the proform of eosinophil major basic protein as itsphysiological inhibitor. J Biol Chem 2000; 275:31128–31133.
41. Miner D, Rajkovic A. Identification of expressed sequence tagspreferentially expressed in human placentas by in silico subtraction.Prenat Diagn 2003; 23:410–419.
42. Smith GC, Stenhouse EJ, Crossley JA, Aitken DA, Cameron AD,
1084 WEYER AND GLERUP
Dow
nloaded from w
ww
.biolreprod.org.

Connor JM. Early-pregnancy origins of low birth weight. Nature 2002;417:916.
43. Wald NJ, Huttly WJ. Validation of risk estimation using the quadrupletest in prenatal screening for Down syndrome. Prenat Diagn 1999;19:1083–1084.
44. Arbuzova S, Nikolenko M, Krantz D, Hallahan T, Macri J. Low first-trimester pregnancy-associated plasma protein-A and Cornelia de Langesyndrome. Prenat Diagn 2003; 23:864.
45. Tong S, Marjono B, Mulvey S, Wallace EM. Low levels of pregnancy-associated plasma protein-A in asymptomatic women destined formiscarriage. Fertil Steril 2004; 82:1468–1470.
46. Han VK, Bassett N, Walton J, Challis JR. The expression of insulin-likegrowth factor (IGF) and IGF-binding protein (IGFBP) genes in thehuman placenta and membranes: evidence for IGF-IGFBP interactions atthe feto-maternal interface. J Clin Endocrinol Metab 1996; 81:2680–2693.
47. Guibourdenche J, Frendo JL, Pidoux G, Bertin G, Luton D, Muller F,Porquet D, Evain-Brion D. Expression of pregnancy-associated plasmaprotein-A (PAPP-A) during human villous trophoblast differentiation invitro. Placenta 2003; 24:532–539.
48. Giudice LC, Conover CA, Bale L, Faessen GH, Ilg K, Sun I, Imani B,Suen LF, Irwin JC, Christiansen M, Overgaard MT, Oxvig C.Identification and regulation of the IGFBP-4 protease and its physiolog-ical inhibitor in human trophoblasts and endometrial stroma: evidence forparacrine regulation of IGF-II bioavailability in the placental bed duringhuman implantation. J Clin Endocrinol Metab 2002; 87:2359–2366.
49. Conover CA, Bale LK, Overgaard MT, Johnstone EW, Laursen UH,Fuchtbauer EM, Oxvig C, van Deursen J. Metalloproteinase pregnancy-associated plasma protein A is a critical growth regulatory factor duringfetal development. Development 2004; 131:1187–1194.
50. Burns JL, Hassan AB. Cell survival and proliferation are modified byinsulin-like growth factor 2 between days 9 and 10 of mouse gestation.Development 2001; 128:3819–3830.
51. Bale LK, Conover CA. Disruption of insulin-like growth factor-IIimprinting during embryonic development rescues the dwarf phenotypeof mice null for pregnancy-associated plasma protein-A. J Endocrinol2005; 186:325–331.
52. Resch ZT, Oxvig C, Bale LK, Conover CA. Stress-activated signalingpathways mediate the stimulation of pregnancy-associated plasmaprotein-A expression in cultured human fibroblasts. Endocrinology2006; 147:885–890.
53. Resch ZT, Chen BK, Bale LK, Oxvig C, Overgaard MT, Conover CA.Pregnancy-associated plasma protein A gene expression as a target ofinflammatory cytokines. Endocrinology 2004; 145:1124–1129.
54. Chen BK, Leiferman KM, Pittelkow MR, Overgaard MT, Oxvig C,Conover CA. Localization and regulation of pregnancy-associatedplasma protein A expression in healing human skin. J Clin EndocrinolMetab 2003; 88:4465–4471.
55. Mazerbourg S, Overgaard MT, Oxvig C, Christiansen M, Conover CA,Laurendeau I, Vidaud M, Tosser-Klopp G, Zapf J, Monget P. Pregnancy-associated plasma protein-A (PAPP-A) in ovine, bovine, porcine, andequine ovarian follicles: involvement in IGF binding protein-4proteolytic degradation and mRNA expression during follicular devel-opment. Endocrinology 2001; 142:5243–5253.
56. Hourvitz A, Kuwahara A, Hennebold JD, Tavares AB, Negishi H, LeeTH, Erickson GF, Adashi EY. The regulated expression of thepregnancy-associated plasma protein-A in the rodent ovary: a proposedrole in the development of dominant follicles and of corpora lutea.Endocrinology 2002; 143:1833–1844.
57. Hourvitz A, Widger AE, Filho FL, Chang RJ, Adashi EY, Erickson GF.Pregnancy-associated plasma protein-A gene expression in humanovaries is restricted to healthy follicles and corpora lutea. J ClinEndocrinol Metab 2000; 85:4916–4920.
58. Nyegaard M, Overgaard MT, Su YQ, Hamilton AE, Kwintkiewicz J,Hsieh M, Nayak NR, Conti M, Conover CA, Giudice LC. Lack offunctional pregnancy-associated plasma protein-A (PAPPA) compromis-es mouse ovarian steroidogenesis and female fertility. Biol Reprod 2010;82:1129–1138.
59. Conover CA, Faessen GF, Ilg KE, Chandrasekher YA, Christiansen M,Overgaard MT, Oxvig C, Giudice LC. Pregnancy-associated plasmaprotein-A is the insulin-like growth factor binding protein-4 proteasesecreted by human ovarian granulosa cells and is a marker of dominantfollicle selection and the corpus luteum. Endocrinology 2001; 142:2155–2158
60. Kalli KR, Chen BK, Bale LK, Gernand E, Overgaard MT, Oxvig C,Cliby WA, Conover CA. Pregnancy-associated plasma protein-A (PAPP-A) expression and insulin-like growth factor binding protein-4 protease
activity in normal and malignant ovarian surface epithelial cells. Int JCancer 2004; 110:633–640.
61. Nishizuka Y. Studies and perspectives of protein kinase C. Science 1986;233:305–312.
62. Conover CA, Bale LK, Harrington SC, Resch ZT, Overgaard MT, OxvigC. Cytokine stimulation of pregnancy-associated plasma protein Aexpression in human coronary artery smooth muscle cells: inhibition byresveratrol. Am J Physiol Cell Physiol 2006; 290:C183–C188.
63. Bayes-Genis A, Schwartz RS, Lewis DA, Overgaard MT, ChristiansenM, Oxvig C, Ashai K, Holmes DR Jr, Conover CA. Insulin-like growthfactor binding protein-4 protease produced by smooth muscle cellsincreases in the coronary artery after angioplasty. Arterioscler ThrombVasc Biol 2001; 21:335–341.
64. Bayes-Genis A, Conover CA, Overgaard MT, Bailey KR, ChristiansenM, Holmes DR Jr, Virmani R, Oxvig C, Schwartz RS. Pregnancy-associated plasma protein A as a marker of acute coronary syndromes. NEngl J Med 2001; 345:1022–1029.
65. Qin QP, Kokkala S, Lund J, Tamm N, Voipio-Pulkki LM, Pettersson K.Molecular distinction of circulating pregnancy-associated plasma proteinA in myocardial infarction and pregnancy. Clin Chem 2005; 51:75–83.
66. Conover CA, Bale LK. Loss of pregnancy-associated plasma protein Aextends life span in mice. Aging Cell 2007; 6:727–729.
67. Harrington SC, Simari RD, Conover CA. Genetic deletion of pregnancy-associated plasma protein-A is associated with resistance to atheroscle-rotic lesion development in apolipoprotein E-deficient mice challengedwith a high-fat diet. Circ Res 2007; 100:1696–1702.
68. Conover CA, Mason MA, Bale LK, Harrington SC, Nyegaard M, OxvigC, Overgaard MT. Transgenic overexpression of pregnancy-associatedplasma protein-A in murine arterial smooth muscle acceleratesatherosclerotic lesion development. Am J Physiol Heart Circ Physiol2010; 299:H284–H291.
69. Conover CA, Chen BK, Resch ZT. Regulation of pregnancy-associatedplasma protein-A expression in cultured human osteoblasts. Bone 2004;34:297–302.
70. Ortiz CO, Chen BK, Bale LK, Overgaard MT, Oxvig C, Conover CA.Transforming growth factor-beta regulation of the insulin-like growthfactor binding protein-4 protease system in cultured human osteoblasts. JBone Miner Res 2003; 18:1066–1072.
71. Farley JR, Tarbaux N, Murphy LA, Masuda T, Baylink DJ. In vitroevidence that bone formation may be coupled to resorption by release ofmitogen(s) from resorbing bone. Metabolism 1987; 36:314–321.
72. Qin X, Wergedal JE, Rehage M, Tran K, Newton J, Lam P, Baylink DJ,Mohan S. Pregnancy-associated plasma protein-A increases osteoblastproliferation in vitro and bone formation in vivo. Endocrinology 2006;147:5653–5661.
73. Miller BS, Bronk JT, Nishiyama T, Yamagiwa H, Srivastava A, BolanderME, Conover CA. Pregnancy associated plasma protein-A is necessaryfor expeditious fracture healing in mice. J Endocrinol 2007; 192:505–513.
74. Conover CA, Kiefer MC, Zapf J. Posttranslational regulation of insulin-like growth factor binding protein-4 in normal and transformed humanfibroblasts. Insulin-like growth factor dependence and biological studies.J Clin Invest 1993; 91:1129–1137.
75. Durham SK, Riggs BL, Harris SA, Conover CA. Alterations in insulin-like growth factor (IGF)-dependent IGF-binding protein-4 proteolysis intransformed osteoblastic cells. Endocrinology 1995; 136:1374–1380.
76. Chen BK, Overgaard MT, Bale LK, Resch ZT, Christiansen M, Oxvig C,Conover CA. Molecular regulation of the IGF-binding protein-4 proteasesystem in human fibroblasts: identification of a novel inducible inhibitor.Endocrinology 2002; 143:1199–1205.
77. Overgaard MT, Glerup S, Boldt HB, Rodacker V, Olsen IM, ChristiansenM, Sottrup-Jensen L, Giudice LC, Oxvig C. Inhibition of proteolysis bythe proform of eosinophil major basic protein (proMBP) requirescovalent binding to its target proteinase. FEBS Lett 2004; 560:147–152.
78. Ye S, Cech AL, Belmares R, Bergstrom RC, Tong Y, Corey DR, KanostMR, Goldsmith EJ. The structure of a Michaelis serpin-proteasecomplex. Nat Struct Biol 2001; 8:979–983.
79. Sottrup-Jensen L. Alpha-macroglobulins: structure, shape, and mecha-nism of proteinase complex formation. J Biol Chem 1989; 264:11539–11542.
80. Visse R, Nagase H. Matrix metalloproteinases and tissue inhibitors ofmetalloproteinases: structure, function, and biochemistry. Circ Res 2003;92:827–839.
81. Bode W, Huber R. Structural basis of the endoproteinase-proteininhibitor interaction. Biochim Biophys Acta 2000; 1477:241–252.
82. Stocker W, Bode W. Structural features of a superfamily of zincendopeptidases: the metzincins. Curr Opin Struct Biol 1995; 5:383–390.
REGULATION OF PROTEOLYSIS BY proMBP 1085
Dow
nloaded from w
ww
.biolreprod.org.

83. Weyer K, Boldt HB, Poulsen CB, Kjaer-Sorensen K, Gyrup C, Oxvig C.A substrate specificity-determining unit of three Lin12-Notch repeatmodules is formed in trans within the pappalysin-1 dimer and requires asequence stretch C-terminal to the third module. J Biol Chem 2007;282:10988–10999.
84. Boldt HB, Kjaer-Sorensen K, Overgaard MT, Weyer K, Poulsen CB,Sottrup-Jensen L, Conover CA, Giudice LC, Oxvig C. The Lin12-notchrepeats of pregnancy-associated plasma protein-A bind calcium anddetermine its proteolytic specificity. J Biol Chem 2004; 279:38525–38531.
85. Hogg PJ. Disulfide bonds as switches for protein function. TrendsBiochem Sci 2003; 28:210–214.
86. Laursen LS, Overgaard MT, Weyer K, Boldt HB, Ebbesen P, Christian-sen M, Sottrup-Jensen L, Giudice LC, Oxvig C. Cell surface targeting ofpregnancy-associated plasma protein A proteolytic activity. Reversibleadhesion is mediated by two neighboring short consensus repeats. J BiolChem 2002; 277:47225–47234.
87. Weyer K, Overgaard MT, Laursen LS, Nielsen CG, Schmitz A,Christiansen M, Sottrup-Jensen L, Giudice LC, Oxvig C. Cell surfaceadhesion of pregnancy-associated plasma protein-A is mediated by fourclusters of basic residues located in its third and fourth CCP module. EurJ Biochem 2004; 271:1525–1535.
88. Kirkitadze MD, Barlow PN. Structure and flexibility of the multipledomain proteins that regulate complement activation. Immunol Rev2001; 180:146–161.
89. Ganesh VK, Smith SA, Kotwal GJ, Murthy KH. Structure of vacciniacomplement protein in complex with heparin and potential implicationsfor complement regulation. Proc Natl Acad Sci U S A 2004; 101:8924–8929.
90. Oxvig C, Haaning J, Hojrup P, Sottrup-Jensen L. Location and nature ofcarbohydrate groups in proform of human major basic protein isolatedfrom pregnancy serum. Biochem Mol Biol Int 1994; 33:329–336.
91. Dementiev A, Petitou M, Herbert JM, Gettins PG. The ternary complexof antithrombin-anhydrothrombin-heparin reveals the basis of inhibitorspecificity. Nat Struct Mol Biol 2004; 11:863–867.
92. Li W, Johnson DJ, Esmon CT, Huntington JA. Structure of theantithrombin-thrombin-heparin ternary complex reveals the antithrom-botic mechanism of heparin. Nat Struct Mol Biol 2004; 11:857–862.
93. Glerup S, Kloverpris S, Laursen LS, Dagnaes-Hansen F, Thiel S,Conover CA, Oxvig C. Cell surface detachment of pregnancy-associatedplasma protein-A requires the formation of intermolecular proteinase-inhibitor disulfide bonds and glycosaminoglycan covalently bound to theinhibitor. J Biol Chem 2007; 282:1769–1778.
94. Pizzo SV. Serpin receptor 1: a hepatic receptor that mediates theclearance of antithrombin III-proteinase complexes. Am J Med 1989;87:10S–14S.
95. Ondetti MA, Cushman DW. Enzymes of the renin-angiotensin systemand their inhibitors. Annu Rev Biochem 1982; 51:283–308.
96. Gould AB, Green D. Kinetics of the human renin and human substratereaction. Cardiovasc Res 1971; 5:86–89.
97. Slater EE, Strout HV Jr. Pure human renin. Identification andcharacterization of two major molecular weight forms. J Biol Chem1981; 256:8164–8171.
98. Lavoie JL, Sigmund CD. Minireview: overview of the renin-angiotensinsystem—an endocrine and paracrine system. Endocrinology 2003;144:2179–2183.
99. Paul M, Poyan Mehr A, Kreutz R. Physiology of local renin-angiotensinsystems. Physiol Rev 2006; 86:747–803.
100. Eggena P, Hidaka H, Barrett JD, Sambhi MP. Multiple forms of humanplasma renin substrate. J Clin Invest 1978; 62:367–372.
101. Gordon DB, Sachin IN. Chromatographic separation of multiple reninsubstrates in women: effect of pregnancy and oral contraceptives. ProcSoc Exp Biol Med 1977; 156:461–464.
102. Tewksbury DA, Dart RA. High molecular weight angiotensinogen levelsin hypertensive pregnant women. Hypertension 1982; 4:729–734.
103. Tewksbury DA, Tryon ES, Burrill RE, Dart RA. High molecular weight
angiotensinogen: a pregnancy associated protein. Clin Chim Acta 1986;158:7–12.
104. Tewksbury DA, Tryon ES. Immunochemical comparison of highmolecular weight angiotensinogen from amniotic fluid, plasma of men,and plasma of pregnant women. Am J Hypertens 1989; 2(5 pt 1):411–413.
105. Tewksbury DA, Goodman JR, Kaiser SJ, Burrill RE, Brown HL.Quantitation of the five forms of plasma high molecular weightangiotensinogen in women with pregnancy-induced hypertension. AmJ Hypertens 2000; 13:221–225.
106. Tewksbury DA. Quantitation of five forms of high molecular weightangiotensinogen from human placenta. Am J Hypertens 1996; 9(10 pt1):1029–1034.
107. Tetlow HJ, Broughton Pipkin F. Changing renin substrates in humanpregnancy. J Endocrinol 1986; 109:257–262.
108. Campbell DJ, Bouhnik J, Coezy E, Menard J, Corvol P. Characterizationof precursor and secreted forms of human angiotensinogen. J Clin Invest1985; 75:1880–1893.
109. Sato Y, Hiwada K, Kokubu T. Physicochemical characteristics of humanhigh molecular weight angiotensinogen. Life Sci 1985; 37:371–377.
110. Christiansen M, Jaliashvili I, Overgaard MT, Ensinger C, Obrist P, OxvigC. Quantification and characterization of pregnancy-associated complex-es of angiotensinogen and the proform of eosinophil major basic proteinin serum and amniotic fluid. Clin Chem 2000; 46(8 pt 1):1099–1105.
111. Gimenez-Roqueplo AP, Celerier J, Schmid G, Corvol P, Jeunemaitre X.Role of cysteine residues in human angiotensinogen. Cys232 is requiredfor angiotensinogen-pro major basic protein complex formation. J BiolChem 1998; 273:34480–34487.
112. Muller-Eberhard HJ. Molecular organization and function of thecomplement system. Annu Rev Biochem 1988; 57:321–347.
113. Pan N, Luo J, Kaiser SJ, Frome WL, Dart RA, Tewksbury DA. Specificreceptor for angiotensinogen on human renal cells. Clin Chim Acta 2006;373:32–36.
114. Tewksbury DA, Pan N, Kaiser SJ. Detection of a receptor forangiotensinogen on placental cells. Am J Hypertens 2003; 16:59–62.
115. Shagdarsuren E, Wellner M, Braesen JH, Park JK, Fiebeler A, Henke N,Dechend R, Gratze P, Luft FC, Muller DN. Complement activation inangiotensin II-induced organ damage. Circ Res 2005; 97:716–724.
116. Ramaha A, Celerier J, Patston PA. Characterization of different highmolecular weight angiotensinogen forms. Am J Hypertens 2003; 16:478–483.
117. Soe R, Overgaard MT, Thomsen AR, Laursen LS, Olsen IM, Sottrup-Jensen L, Haaning J, Giudice LC, Conover CA, Oxvig C. Expression ofrecombinant murine pregnancy-associated plasma protein-A (PAPP-A)and a novel variant (PAPP-Ai) with differential proteolytic activity. Eur JBiochem 2002; 269:2247–2256.
118. Merrill DC, Thompson MW, Carney CL, Granwehr BP, Schlager G,Robillard JE, Sigmund CD. Chronic hypertension and altered baroreflexresponses in transgenic mice containing the human renin and humanangiotensinogen genes. J Clin Invest 1996; 97:1047–1055.
119. Takimoto E, Ishida J, Sugiyama F, Horiguchi H, Murakami K, FukamizuA. Hypertension induced in pregnant mice by placental renin andmaternal angiotensinogen. Science 1996; 274:995–998.
120. Luft FC, Mervaala E, Muller DN, Gross V, Schmidt F, Park JK, SchmitzC, Lippoldt A, Breu V, Dechend R, Dragun D, Schneider W, Ganten D,Haller H. Hypertension-induced end-organ damage: a new transgenicapproach to an old problem. Hypertension 1999; 33(1 pt 2):212–218.
121. Pilz B, Shagdarsuren E, Wellner M, Fiebeler A, Dechend R, Gratze P,Meiners S, Feldman DL, Webb RL, Garrelds IM, Jan Danser AH, LuftFC. Muller DN. Aliskiren, a human renin inhibitor, ameliorates cardiacand renal damage in double-transgenic rats. Hypertension 2005; 46:569–576.
122. Weis WI, Kahn R, Fourme R, Drickamer K, Hendrickson WA. Structureof the calcium-dependent lectin domain from a rat mannose-bindingprotein determined by MAD phasing. Science 1991; 254:1608–1615.
1086 WEYER AND GLERUP
Dow
nloaded from w
ww
.biolreprod.org.




















