
Development and morphology of secretory trichomes of Calceolaria volckmanni (Scrophulariaceae
Upload
independentCategory
view
4download
0
Autosomal Dominant Growth Hormone DeficiencyDisrupts Secretory Vesicles in Vitro and in Vivo inTransgenic Mice
LINDSAY MCGUINNESS, CHARALAMBOS MAGOULAS*, ABDUL K. SESAY, KATHLEEN MATHERS,DANIELLE CARMIGNAC, JEAN-BAPTISTE MANNEVILLE, HELEN CHRISTIAN,JOHN A. PHILLIPS III, AND IAIN C. A. F. ROBINSON
National Institute for Medical Research (L.M., C.M., K.M., D.C., J.-B.M., I.C.A.F.R., A.K.S.), Mill Hill, London NW7 1AA,United Kingdom; Department of Human Anatomy and Genetics (H.C.), Oxford OX1 3QX, United Kingdom; and Departmentof Pediatrics (J.A.P.), Vanderbilt University School of Medicine, Nashville, Tennessee 37235
Autosomal dominant GH deficiency type II (IGHDII) is oftenassociated with mutations in the human GH gene (GH1) thatgive rise to products lacking exon-3 (�exon3hGH). In the het-erozygous state, these act as dominant negative mutationsthat prevent the release of human pituitary GH (hGH). Todetermine the mechanisms of these dominant negative ef-fects, we used a combination of transgenic and morphologicalapproaches in both in vitro and in vivo models. Rat GC celllines were generated expressing either wild-type GH1 (WT-hGH-GC) or a genomic GH1 sequence containing a G->A tran-sition at the donor splice site of IVS3 (�exon3hGH-GC). WT-hGH-GC cells grew normally and produced equivalentamounts of human and rGH packaged in dense-cored secre-tory vesicles (SVs). In contrast, �exon3hGH-GC cells showedfew SVs but accumulated secretory product in amorphouscytoplasmic aggregates. They produced much less rGH andgrew more slowly than WT-hGH-GC cells. When cotransfectedwith an enhanced green fluorescent protein construct (GH-eGFP), which copackages with GH in SVs, WT-hGH-GC cellsshowed normal electron microscopy morphology and SV
movements, tracked with total internal reflectance fluores-cence microscopy. In contrast, coexpression of �exon3hGH withGH-eGFP abolished the vesicular targeting of GH-eGFP,which instead accumulated in static aggregates. Transgenicmice expressing �exon3hGH in somatotrophs showed an IGHD-II phenotype with mild to severe pituitary hypoplasia anddwarfism, evident at weaning in the most severely affectedlines. Hypothalamic GHRH expression was up-regulated andsomatostatin expression reduced in �exon3hGH transgenicmice, consistent with their profound GHD. Few SVs were de-tectable in the residual pituitary somatotrophs in �exon3hGHtransgenic mice, and these cells showed grossly abnormalmorphology. A low copy number transgenic line showed amild effect relatively specific for GH, whereas two severelyaffected lines with higher transgene copy numbers showedearly onset, widespread pituitary damage, macrophage inva-sion, and multiple hormone deficiencies. These new in vitroand in vivo models shed new light on the cellular mechanismsinvolved in IGHDII, as well as its phenotypic consequences invivo. (Endocrinology 144: 720–731, 2003)
HUMAN PITUITARY GH (hGH) is encoded by exons1–5 of the GH1 gene. The major bioactive product is
a 22-kDa form, but alternative splicing can give rise to minorforms, the most prominent of which is a bioactive 20-kDahGH that results from use of a cryptic 3� splice site in exon3, deleting amino acids 32–46 (1). Rarely, however, muta-tions in GH1 can give rise to alternative spliced products thatinduce a form of GH deficiency (IGHDII) inherited as anautosomal dominant trait. Such individuals show variablyreduced plasma GH levels and growth rates but usuallyrespond positively to exogenous GH therapy (2). This fre-quently occurs with mutations at, or close to, the splice sitesaround exon 3 (3, 4), which generate mRNAs lacking se-quences coding for this exon. The resulting exon-3 skip prod-uct (�exon3hGH) generates a 17.5-kDa hGH that lacks aminoacids 32–71 (2, 4), which includes the loop connecting the firsttwo helices of the wild-type GH (WT-hGH) structure (5); it
also disrupts an internal disulfide bridge, by deletion of53Cys in WT-hGH.
Although the 17.5-kDa protein produced by such IGHDIImutations is unlikely to activate the GH receptor, the dom-inant nature and severity of the phenotype, together with thelack of 22-kDa hGH in the circulation, suggests that the�exon3hGH product from the mutant allele efficiently pre-vents the production, storage, or release of WT-hGH from theother, normal allele. Because donor and acceptor splice sitesare well conserved throughout vertebrates (6), it is possibleto reproduce the misplicing by expressing such humangenomic sequences in rodent cell lines, and when coex-pressed with WT-hGH, �exon3hGH products exert a domi-nant negative effect on WT-hGH production (7, 8) and causevariety of morphological abnormalities in heterologous celllines (9). Many possible intracellular mechanisms have beeninvoked to explain the dominant negative effects of mis-folded protein hormones such as �exon3hGH (10–12), fre-quently based on acute transfection studies in heterologouscell lines. The mechanisms by which �exon3hGH suppressesWT-GH production and release, and lead to progressivedysfunction of somatotrophs in vivo, remain unclear.
To address some of these issues, we have expressed
Abbreviations: CMV, Cytomegalovirus; eGFP, enhanced green flu-orescent protein; EM, electron microscopy; hGH, human pituitary GH;IGHDII, autosomal dominant GH deficiency type II; GH1, human GHgene; LCR, locus control region; NT, nontransgenic; rGH, rat GH; SV,secretory vesicle; TIRF, total internal reflection fluorescence; WT-hGH,wild-type GH.
0013-7227/03/$15.00/0 Endocrinology 144(2):720–731Printed in U.S.A. Copyright © 2003 by The Endocrine Society
doi: 10.1210/en.2002-220847
720

�exon3hGH in a GH-producing rat cell line with or without acoexpressed and copackaged fluorescent secretory vesicle(SV) marker (13) to study the effects of �exon3hGH on cell andSV morphology and movements, using confocal, total inter-nal reflection fluorescence (TIRF) and immunoelectron mi-croscopy. Secondly, we have generated several lines of trans-genic mice expressing �exon3hGH, to create and characterizea murine model of human IGHDII and to study the effectsof �exon3hGH in primary somatotrophs for the first time.
Materials and MethodsConstructs
The constructs used in this study are shown in Fig. 1. A SacI/BglIIfragment containing the IVS3 (�1G3A) mutation in a GH1 genomicclone (4) was exchanged into pKS-GH.M, a 2.6-kb genomic constructcontaining 5�-and 3� untranslated GH1 sequences flanked by MluI sites(13). The �1G-�A mutation generates an additional NlaIII site, notpresent in WT-GH1 DNA sequences. Both the WT and mutant IVS3 �1G3A constructs were confirmed by restriction mapping and DNAsequencing. For transfection studies, a BamH1/Not1 fragment of pKS-GH.M was subcloned into pCDNA3.1 (Invitrogen, Paisley, UK) con-taining a cytomegalovirus (CMV) promoter and a zeocin selection cas-sette (Fig. 1A); a similar expression vector was generated for WT-GH1,but with a neomycin selection cassette. For transgenesis, the pKS-GH.Minsert was subcloned into a 40-kb cosmid (14) containing the entire GH1locus control region (LCR) (Fig. 1B), using an Mlu1 site engineered aspreviously described (13). The full-length insert was released from thecosmid as a Not1 fragment and purified for microinjection into oocytes.For simplicity, the DNA sequence containing the GH1-IVS3 (�1G3A)mutation will subsequently be termed �exon3hGH. To generate GC cellswith fluorescent GH SVs, they were also transfected with a CMV ex-pression vector [p48hGH-enhanced green fluorescent protein (eGFP)]containing sequences coding for the signal peptide and first 22 residuesof mature GH1 followed by eGFP (13). We have previously shown thatthis generates a fluorescent product which is copackaged with GH in SVsboth in rat GC cells and in somatotrophs of transgenic mice (13).
Culture of rat GC cells
Rat GC cells (15, 16) were cultured at 37 C in complete medium[DMEM, 15% horse serum, 2.5% fetal calf serum (PAA, Weiner Strasse,Linz, Austria)], 2 mm l-glutamine, and 1� antibiotic-antimycotic solu-tion [penicillin/streptomycin/amphotericin]) and transfected with plas-mids expressing either p48 hGH-eGFP, WT-hGH or �exon3hGH alone or
in combination, using Lipofectamine (Life Technologies, Inc., Gaithers-burg, MD). Stably transfected cell lines were selected for neomycinresistance (G-418, 250 �g/ml, Life Technologies, Inc.) and/or zeocinresistance (zeocin 200 �g/ml, Invitrogen BV, Breda, The Netherlands)for at least 21 d.
Generation of �exon3hGH transgenic animals.
All animal work was carried out under UK Home Office guidelines.Transgenic mice were generated by microinjection of the LCR-�exon3hGH Not1 fragment (Fig. 1B) into the pronucleus of fertilizedone-cell Cba/Ca x C57Bl/10 mouse oocytes followed by oviductal trans-fer into pseudopregnant recipients. Tail DNA from the resulting pupswere screened by Southern blot for GH1 and by PCR using primers thatspanned the first GH1 exon/intron pair, as previously described (13).
RT-PCR
RNA was extracted from GC cells (SNAP total RNA Isolation Kit(Invitrogen BV) and 500 ng reverse transcribed with 200 U of Moloney-murine leukemia virus reverse transcriptase (Roche Molecular Bio-chemicals, Lewes, UK) supplemented with 1 �g random primers(Invitrogen), 0.3 mm deoxynucleotide triphosphates (AmershamPharmacia Biotech), 40 U ribonuclease inhibitor (Promega Corp.,Southampton, UK), and 5 mm dithiothreitol. After incubation at 37 C for2 h the resulting cDNAs were cloned, amplified (TOPO Cloning Kit,Invitrogen BV) and sequenced. To distinguish WT-hGH and �exon3hGHtranscripts, two sets of primer pairs were used, both of which spannedexon 3 and should give 120-bp larger products for WT-hGH than for�exon3hGH transcripts.
Southern blotting
After restriction digestion, DNA (10 �g of genomic DNA or 0.5 �g ofplasmid/cosmid DNA) was separated by electrophoresis in 0.6% aga-rose gels, transferred onto Hybond-N� nylon membranes (AmershamPharmacia Biotech) in 0.4 m NaOH by capillary transfer and hybridizedwith a random primed [�32P]-labeled full-length GH1 genomic DNAprobe (Prime-a-Gene, Promega Corp.) for 65 C overnight. Blots werewashed and exposed to phosphoimager screens or to Kodak BioMax filmat �70 C.
RIA
Pituitaries were dissected and homogenized in 1 ml PBS. Proteincontents were measured with the BCA Protein Assay Reagent (PierceChemical Co., Rockford, IL) using BSA as standard (Sigma, Poole, UK).Aliquots were taken for assay of GH, PRL, LH, and TSH using specificreagents kindly provided by Dr. A. L. Parlow and by the NIDDK (17,18); hGH was assayed using an antibody that detects 22-kDa hGH butnot mouse or rat GH (rGH) (18).
In situ hybridization
In situ hybridization was performed as previously described (19) on12-�m cryostat coronal sections of mouse hypothalamus using [�35S]-uridine triphosphate-labeled sense and antisense riboprobes for GHRHand SRIF generated using an SP6/T7 transcription kit (Roche Diagnos-tics). The GHRH riboprobe represented a full-length mouse cDNA (Im-age clone 1496474), whereas the SRIF riboprobe corresponded to nu-cleotides 280–556 of rat SRIF cDNA. Following overnight hybridization,sections were washed, dried and exposed to autoradiographic film(BioMax MR, Kodak, Rochester, NY) for up to 7 d before measuringintegrated densities, using NIH Image as previously described (19). Foreach transcript, comparisons were made with the same batch of labeledriboprobe on sections from all animals processed at the same time.
Electron microscopy (EM)
After initial fixation (2.5% glutaraldehyde in 0.1 m sodium phosphatebuffer for 2 h at room temperature, then 0.25% overnight at 4 C) cells orpituitary segments were post-fixed in osmium tetroxide (1% wt/vol in0.1 m sodium phosphate buffer), contrasted with uranyl acetate (2%
FIG. 1. Constructs used in this study. A, A CMV expression vector(pKS-GH-M) was constructed, containing a 2.6-kb GH1 genomic se-quence with a single base �1G-�A substitution in intron 3 (*), andflanked by Mlu1 restriction sites (M). B, The Mlu1 fragment of A wascloned into a 40-kb cosmid containing the GH1 LCR, via a Mlu1cloning site to give LCR �exon3hGH (13). The insert was released as aNot1 fragment (N) for oocyte microinjection.
McGuinness et al. • Autosomal Dominant GH Deficiency Endocrinology, February 2003, 144(2):720–731 721

wt/vol in distilled water), dehydrated through increasing concentra-tions of ethanol (70–100%) and embedded in Spurr’s resin (Agar Sci-entific UK, Stansted, UK). For immunogold detection of GH, ultrathinsections (50–80 nm) were incubated at room temperature with rabbitantimouse GH 1:2000 (NIDDK), for 2 h, followed by protein A 15 nmgold (British Biocell, Cardiff, UK) for 1 h at room temperature. In controlsections, the primary antibody was replaced by nonimmune rabbit se-rum. Sections were counter-stained with lead citrate and uranyl acetateand examined with a transmission electron microscope (JEM-1010,JEOL, Peabody, MA).
TIRF microscopy
GC cells stably transfected with p48hGH-eGFP, with or without co-transfected WT-hGH or �exon3hGH were cultured on glass slides, andexamined with a TIRF microscope. Light from TIRF images were passedthrough a dichroic (505DRLP02, Omega Optical, Brattleboro, VT) and anemission filter (530DF30) and collected with an intensified CCD camera(Remote Head Darkstar, S25 Intensifier, Photonics Science, Roberts-bridge, UK). Images were digitized and stored in memory at 25 frames/sec by a frame grabber (IC-PCI 4Mb (AMVS), Imaging Technology,Bedford, MA) and then saved to disk. Image processing was carried outusing Optimas version 6.5 (Optimas Corp., Bothwell, WA).
Data analysis
Unless otherwise stated, data are shown as mean � sem. Differencesbetween groups were analyzed by ANOVA followed by Student’s t testor Mann-Whitney U test as appropriate. Differences of P � 0.05 wereconsidered significant.
ResultsGC cell transfection
Wild-type GH1 (WT-hGH) and IVS-3 � 1 G-�A mutantGH1 constructs (�exon3hGH) were transfected into rat GCcells and stable lines established. RNA was extracted, andRT-PCR performed with two sets of primers spanning exon3 (Fig. 2A). As expected, cells transfected with the �exon3-hGHplasmid (�exon3hGH-GC cells) generated a major transcriptthat yielded an RT-PCR product 120-bp smaller than in cellstransfected with WT-hGH (WT-hGH-GC cells). No detect-able WT-hGH product was amplified from �exon3hGH-GCcells, and neither product was detected in control GC cells.Sequencing the cDNA products obtained by RT-PCR fromfour different �exon3hGH-GC cell isolates confirmed an exactdeletion of exon 3 sequences. Culture flasks were inoculatedwith 250,000 cells and their growth monitored by cell count-ing over 14 d. Untransfected GC cells and WT-hGH-GC cellsaccumulated at similar rates, whereas there were 80% fewer�exon3hGH-GC cells remaining after 14 d.
rGH and hGH contents were compared in extracts fromcultures of GC, WT-hGH-GC, and �exon3hGH-GC cells (Fig.2B). rGH production was similar in both GC and WT-hGH-GC cells (15.2 � 0.6 �g/105 cells and 14.4 � 0.4 �g/105
cells, respectively) and WT-hGH-GC cells contained similaramounts of hGH (17.2 � 1 �g/105 cells). In contrast, no hGHimmunoreactivity could be detected in �exon3hGH-GC cells,and their rGH contents were markedly lower (0.56 � 0.19�g/105 cells, P � 0.005).
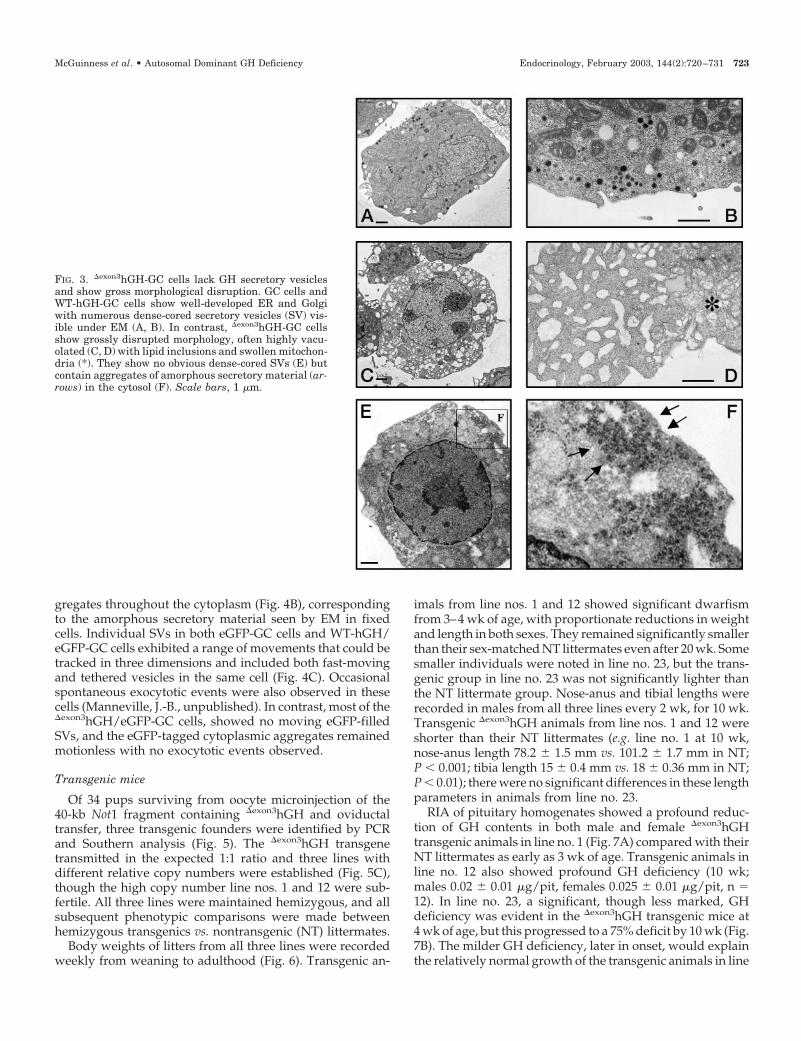
No morphological differences were apparent between GCand WT-hGH-GC cells in culture. In contrast, many more�exon3hGH-GC cells were detached and had a shriveled ap-pearance, with more cell debris evident in these cultures.Both WT-hGH-GC and �exon3hGH-GC cells were examinedby EM. Abnormal morphology was obvious in the
�exon3hGH-GC cell cultures (Fig. 3, C–F) compared with GCor WT-hGH-GC cells (Fig. 3, A and B). The majority of�exon3hGH-GC cells showed a grossly abnormal cytology,with a notable absence of dense-cored SVs, but instead acollection of amorphous electron dense cytoplasmic aggre-gates without any obvious vesicular structure (Fig. 3, E andF). Many �exon3hGH-GC cells showed a highly vacuolatedcytoplasm and lipid accumulations, fragmented or lobularnuclei, and swollen organellar structures, including ER,Golgi apparatus, and mitochondria (Fig. 3, C–F). Immuno-gold EM stained the dense cored SVs in GC cells and WT-hGH-GC cells, but gave only diffuse labeling of the cyto-plasmic aggregates in the �exon3hGH-GC cells (not shown).
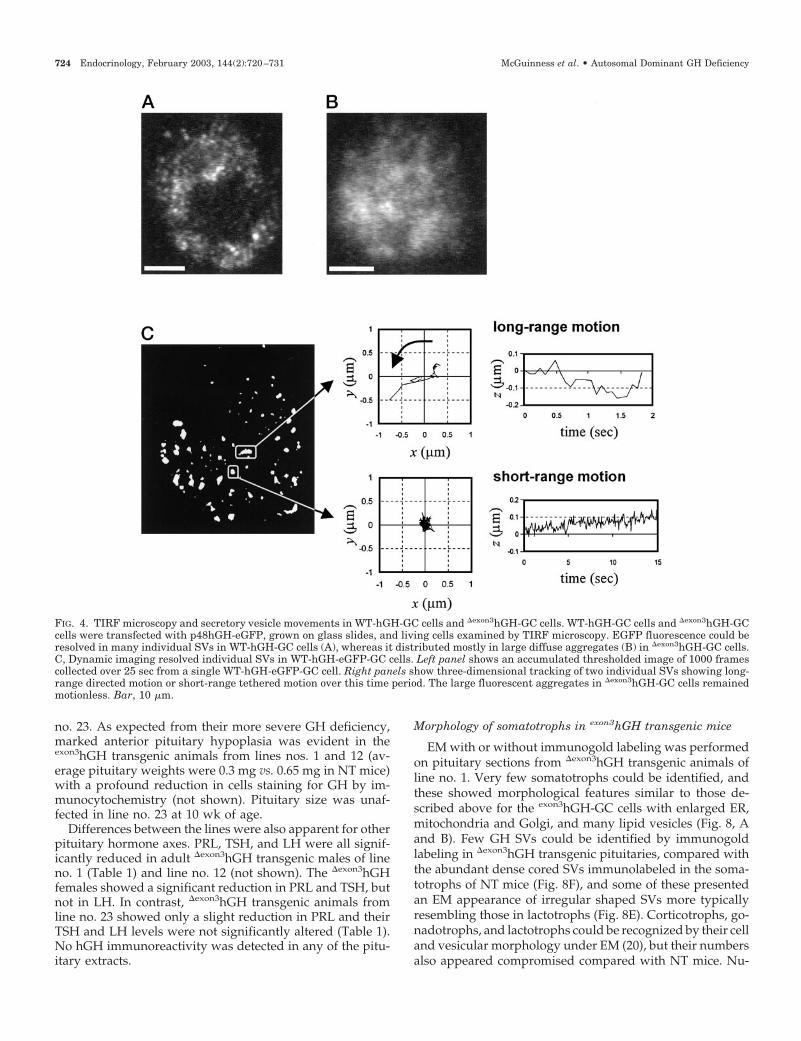
To study GH SVs dynamically in living cells, we exploitedan eGFP construct (p48-hGH-eGFP) whose product is co-packaged with GH in the same SVs, rendering them fluo-rescent (13). When this was coexpressed with WT-hGH or�exon3hGH, bright eGFP fluorescence resulted but in mark-edly different patterns visible under confocal or TIRF mi-croscopy. WT-hGH/eGFP GC cells showed the punctate dis-tribution of eGFP fluorescence typical of SVs (Fig. 4A),whereas �exon3hGH/eGFP-GC cells exhibited few fluores-cent SVs, but the eGFP formed large diffuse fluorescent ag-
FIG. 2. pKS-GH-M generates a �exon3hGH transcript after stabletransfection in GC cells and suppresses endogenous rGH production.A, RT-PCR of RNA extracts from GC cells stably transfected withCMV vectors expressing WT hGH (first six lanes), �exon3hGH (nextfour lanes), or untransfected GC cells (last three lanes). Two differentprimer pairs that spanned exon 3 sequences amplified major products120 bp shorter in �exon3hGH-transfected cells, which when cloned andsequenced proved to lack exon 3 sequences. B, rGH (open bars) andhGH (solid bars) were measured by RIA in extracts of GC cells withouttransfection (GC) or stably transfected with WT-hGH (WT-hGH-GC)or �exon3hGH (�exon3hGH-GC). ***, P � 0.001 vs. WT-hGH-GC or GC.
722 Endocrinology, February 2003, 144(2):720–731 McGuinness et al. • Autosomal Dominant GH Deficiency
gregates throughout the cytoplasm (Fig. 4B), correspondingto the amorphous secretory material seen by EM in fixedcells. Individual SVs in both eGFP-GC cells and WT-hGH/eGFP-GC cells exhibited a range of movements that could betracked in three dimensions and included both fast-movingand tethered vesicles in the same cell (Fig. 4C). Occasionalspontaneous exocytotic events were also observed in thesecells (Manneville, J.-B., unpublished). In contrast, most of the�exon3hGH/eGFP-GC cells, showed no moving eGFP-filledSVs, and the eGFP-tagged cytoplasmic aggregates remainedmotionless with no exocytotic events observed.
Transgenic mice
Of 34 pups surviving from oocyte microinjection of the40-kb Not1 fragment containing �exon3hGH and oviductaltransfer, three transgenic founders were identified by PCRand Southern analysis (Fig. 5). The �exon3hGH transgenetransmitted in the expected 1:1 ratio and three lines withdifferent relative copy numbers were established (Fig. 5C),though the high copy number line nos. 1 and 12 were sub-fertile. All three lines were maintained hemizygous, and allsubsequent phenotypic comparisons were made betweenhemizygous transgenics vs. nontransgenic (NT) littermates.
Body weights of litters from all three lines were recordedweekly from weaning to adulthood (Fig. 6). Transgenic an-
imals from line nos. 1 and 12 showed significant dwarfismfrom 3–4 wk of age, with proportionate reductions in weightand length in both sexes. They remained significantly smallerthan their sex-matched NT littermates even after 20 wk. Somesmaller individuals were noted in line no. 23, but the trans-genic group in line no. 23 was not significantly lighter thanthe NT littermate group. Nose-anus and tibial lengths wererecorded in males from all three lines every 2 wk, for 10 wk.Transgenic �exon3hGH animals from line nos. 1 and 12 wereshorter than their NT littermates (e.g. line no. 1 at 10 wk,nose-anus length 78.2 � 1.5 mm vs. 101.2 � 1.7 mm in NT;P � 0.001; tibia length 15 � 0.4 mm vs. 18 � 0.36 mm in NT;P � 0.01); there were no significant differences in these lengthparameters in animals from line no. 23.
RIA of pituitary homogenates showed a profound reduc-tion of GH contents in both male and female �exon3hGHtransgenic animals in line no. 1 (Fig. 7A) compared with theirNT littermates as early as 3 wk of age. Transgenic animals inline no. 12 also showed profound GH deficiency (10 wk;males 0.02 � 0.01 �g/pit, females 0.025 � 0.01 �g/pit, n 12). In line no. 23, a significant, though less marked, GHdeficiency was evident in the �exon3hGH transgenic mice at4 wk of age, but this progressed to a 75% deficit by 10 wk (Fig.7B). The milder GH deficiency, later in onset, would explainthe relatively normal growth of the transgenic animals in line
FIG. 3. �exon3hGH-GC cells lack GH secretory vesiclesand show gross morphological disruption. GC cells andWT-hGH-GC cells show well-developed ER and Golgiwith numerous dense-cored secretory vesicles (SV) vis-ible under EM (A, B). In contrast, �exon3hGH-GC cellsshow grossly disrupted morphology, often highly vacu-olated (C, D) with lipid inclusions and swollen mitochon-dria (*). They show no obvious dense-cored SVs (E) butcontain aggregates of amorphous secretory material (ar-rows) in the cytosol (F). Scale bars, 1 �m.
McGuinness et al. • Autosomal Dominant GH Deficiency Endocrinology, February 2003, 144(2):720–731 723
no. 23. As expected from their more severe GH deficiency,marked anterior pituitary hypoplasia was evident in theexon3hGH transgenic animals from lines nos. 1 and 12 (av-erage pituitary weights were 0.3 mg vs. 0.65 mg in NT mice)with a profound reduction in cells staining for GH by im-munocytochemistry (not shown). Pituitary size was unaf-fected in line no. 23 at 10 wk of age.
Differences between the lines were also apparent for otherpituitary hormone axes. PRL, TSH, and LH were all signif-icantly reduced in adult �exon3hGH transgenic males of lineno. 1 (Table 1) and line no. 12 (not shown). The �exon3hGHfemales showed a significant reduction in PRL and TSH, butnot in LH. In contrast, �exon3hGH transgenic animals fromline no. 23 showed only a slight reduction in PRL and theirTSH and LH levels were not significantly altered (Table 1).No hGH immunoreactivity was detected in any of the pitu-itary extracts.
Morphology of somatotrophs in exon3hGH transgenic mice
EM with or without immunogold labeling was performedon pituitary sections from �exon3hGH transgenic animals ofline no. 1. Very few somatotrophs could be identified, andthese showed morphological features similar to those de-scribed above for the exon3hGH-GC cells with enlarged ER,mitochondria and Golgi, and many lipid vesicles (Fig. 8, Aand B). Few GH SVs could be identified by immunogoldlabeling in �exon3hGH transgenic pituitaries, compared withthe abundant dense cored SVs immunolabeled in the soma-totrophs of NT mice (Fig. 8F), and some of these presentedan EM appearance of irregular shaped SVs more typicallyresembling those in lactotrophs (Fig. 8E). Corticotrophs, go-nadotrophs, and lactotrophs could be recognized by their celland vesicular morphology under EM (20), but their numbersalso appeared compromised compared with NT mice. Nu-
FIG. 4. TIRF microscopy and secretory vesicle movements in WT-hGH-GC cells and �exon3hGH-GC cells. WT-hGH-GC cells and �exon3hGH-GCcells were transfected with p48hGH-eGFP, grown on glass slides, and living cells examined by TIRF microscopy. EGFP fluorescence could beresolved in many individual SVs in WT-hGH-GC cells (A), whereas it distributed mostly in large diffuse aggregates (B) in �exon3hGH-GC cells.C, Dynamic imaging resolved individual SVs in WT-hGH-eGFP-GC cells. Left panel shows an accumulated thresholded image of 1000 framescollected over 25 sec from a single WT-hGH-eGFP-GC cell. Right panels show three-dimensional tracking of two individual SVs showing long-range directed motion or short-range tethered motion over this time period. The large fluorescent aggregates in �exon3hGH-GC cells remainedmotionless. Bar, 10 �m.
724 Endocrinology, February 2003, 144(2):720–731 McGuinness et al. • Autosomal Dominant GH Deficiency

merous macrophages were noted throughout the �exon3hGHtransgenic anterior pituitaries (Fig. 8D) but not in NT pitu-itaries, with many perivascular macrophages at the bound-ary of the anterior lobe with the intermediate lobe; Fig. 8C).
Hypothalamic GHRH and somatostatin expression
The profound reduction in pituitary GH in �exon3hGHtransgenic mice would be expected to remove GH feedback,thereby increasing hypothalamic GHRH expression and de-creasing SRIF expression. To study this, in situ hybridizationwas performed for these transcripts in hypothalamic sectionsprepared from �exon3hGH transgenic and NT animals. Asexpected, arcuate GHRH mRNA abundance was signifi-cantly higher in the �exon3hGH transgenic mice, and SRIFmRNA abundance was significantly lower in their periven-tricular nuclei, compared with that seen in NT mice (Fig. 9).
Discussion
Familial IGHD-II is often associated with dominant-negative mutations of the third intron of the GH1 gene, eitherby direct interference at the 5� splice site, or by compromisingsplice enhancer sequences; these mutations cause exon-3skipping, which generates a 17.5-kDa hGH protein variant(12, 21, 22). We made use of a human genomic GH1 constructcontaining the IVS3 � 1 G3A donor splice site mutationwhich causes familial IGHDII (4). It gave rise to �exon3hGHbut not WT-hGH when transfected in rodent GC cells andexerted a dominant negative effect on endogenous rGH pro-duction in these cells. The same genomic sequence was theninserted into a human LCR transgene construct that drivescopy number-, position-independent transgene expressionspecifically in somatotrophs in transgenic mice (13, 14). Thisgenerated the first murine model of IGHDII in which it was
possible to examine both the cellular and physiological con-sequences of �exon3hGH expression in vivo.
How the expression of a dominant negative protein hormonesuppresses the secretion of the normal product from an unaf-fected allele is unclear. Such mechanisms could include generaldefects in protein folding or aggregation (11), accumulation ofmutant protein in the ER, or mis-sorting to degradative com-partments (23, 24), a toxic effect of the mutant protein per se oncell function (9) or to a more specific interference with theproduction of the normal allele product (25, 26).
Using metabolic labeling in nonsecretory cell lines, Ha-yashi et al. (7) showed that coexpression of �exon3hGH did notinhibit WT-hGH production nor did it affect cell viability,whereas in secretory pituitary cell lines, it inhibited WT-hGHsecretion in a concentration-dependent manner. Lee et al. (8)found similar results and showed some specificity for hGH,because coexpression of �exon3hGH with PRL did not affectPRL production and secretion. However, in COS7 cells, ex-pression of �exon3hGH did disrupt PRL production and dis-rupted ER to Golgi trafficking (9). An Arg183His hGH mu-tation that also causes IGHDII (26) gives rise to a protein thatwas efficiently secreted from a neuroendocrine cell line whenexpressed alone but greatly impaired secretion when coex-pressed with WT-hGH. Taken together, these results suggestthat mutant GHs do not have a general toxic effect per se butcan exert a powerful dominant negative effect when able tointeract with WT-hGH in a cell with a prominent regulatedsecretory pathway.
Our morphological data from both cell lines and primarysomatotrophs shed some light on this because expression of�exon3hGH prevented or destabilized SV formation in a dom-inant negative fashion and prevented the normal packagingof endogenous mouse or rGH into dense-cored SVs. Instead,
FIG. 5. Generation of �exon3hGH transgenic mice. Three lines of transgenic mice were established following oocyte microinjection with the 40-kbLCR �exon3hGH Not1 insert. A, Transgenic mice (T) were distinguished from NT littermates by a PCR assay of tail DNA, which amplifies acrossthe first hGH intron (see Materials and Methods) and gives a 50-bp larger product for human vs. mouse GH sequences. B, Southern blottingwith a full-length hGH genomic probe comparing WT-GH1 DNA (cosmid K2B, left panel) and DNA from an �exon3hGH transgenic mouse (T,right panel). The �1G-�A mutation generates an additional NlaIII site not present in WT-GH1 DNA sequences, giving two smaller fragmentsafter digestion of the transgenic mice DNA with NlaIII, compared with the 1.1-kb band in WT GH1 DNA. C, To estimate relative copy numbersin the three transgenic lines, DNA was extracted, adjusted to the same DNA concentration, and analyzed on a phosphoimager after gelelectrophoresis and hybridizing with the hGH genomic probe. Line no. 23 had least 4- to 8-fold fewer transgene copies than line nos. 1 and 12.
McGuinness et al. • Autosomal Dominant GH Deficiency Endocrinology, February 2003, 144(2):720–731 725

endogenous GH gradually accumulated in aggregates in thecytosol, unavailable for exocytotic release. In addition toswollen and disrupted ER and Golgi structures, severelyaffected �exon3hGH-GC cells showed an abundance of intra-cellular lipid, also suggesting a substantial increase in mem-brane recycling.
These observations were made on fixed cells, but by usingTIRF microscopy and an eGFP construct that copackageswith GH in SVs we could observe this process dynamicallyat the level of individual SVs in single GC cells. TIRF, orevanescent wave, microscopy, illuminates fluorophores in a100-nm-thick optical slice between a glass surface and thebasal membrane of an adhering cell (27) and can thus be usedto image individual SV movements in living cells (28). WhenTIRF was used to image eGFP-transfected GC cells, individ-ual SVs could be resolved and their range of fast or slowmovements tracked in either GC or WT-hGH-GC cells. How-ever, very few eGFP-containing SVs were present in�exon3hGH-GC cells. Instead, most of the eGFP fluorescencewas localized in amorphous cytoplasmic aggregates corre-
sponding to the diffuse secretory material seen under EM.This material remained motionless in the cell and did notgive rise to exocytotic events. Because the eGFP constructdoes not require WT-hGH for packaging into SVs in othersecretory cells (Robinson, I. C. A. F., unpublished), the lackof fluorescent SVs in �exon3hGH/GC-eGFP cells providesfurther evidence that the mutant hGH disrupts SV formationin these cells. We suggest that this is the key process by which�exon3hGH eventually destroys the somatotroph.
Packaging of hGH into SVs represent the culmination ofmany protein assembly processes that proceed throughoutthe secretory pathway (29). Protein folding, dimerization,and oligomeric association of SV proteins begins in the ER(30) and occur before trafficking via the Golgi to form con-densing SVs (31). Secretory proteins must also be foldedcorrectly to exit the ER (29, 32). Because deletion of exon 3sequences includes Cys53, the resulting unpaired Cys165
could result in aberrant intra- or inter-molecular disulfide-linked misfolded aggregates. However, this cannot be theonly explanation because a dominant negative effect is still
FIG. 6. Growth curves in three lines of �exon3hGH transgenic mice. Body weights were recorded from males (M) and females (F) for transgenic(T) and NT littermates from all three lines of �exon3hGH transgenic mice (n 12 per group). Both T males and females became significantlysmaller and lighter than their NT littermates in line nos. 1 and 12 around 3–4 wk of age, but not in line no. 23. The effects were larger in malesthan in females, and the reduction in body size was proportionate to body weight (inset figure, animals from line no. 1). ***, P � 0.001 vs. NTmales; ##, P � 0.01 vs. NT females.
726 Endocrinology, February 2003, 144(2):720–731 McGuinness et al. • Autosomal Dominant GH Deficiency

observed after mutating the unpaired Cys165 partner (8).Furthermore, dominant negative suppression of WT-hGHproduction is also seen with a single point missense mutationin GH1 not affecting disulfide bridges (26), and largeamounts of WT-hGH are secreted in a patient bearing aheterozygous Arg77Cys mutation, which also generates anadditional unpaired Cys in a mutant hGH that is secreted butwhich then antagonizes WT-hGH at the GH receptor (33).
Misfolded proteins detected in the ER are transported backto the cytosol where they are degraded by proteasomes (34,35) and do not usually cause a dominant negative effect (36)unless the production rate exceeds the proteasome degra-
dation capacity. For example, there are other GH1 frameshiftmutations (25, 37) that also produce misfolded proteins, butthese are phenotypically recessive. The proteasome pathwayhas been implicated in the degradation of �exon3hGH becauseits accumulation is enhanced when expressed in AtT20 cellstreated with proteasome inhibitors (8).
Concentration, oligomerization, and condensation to formdense SV protein cores is promoted by mild acidification inthe presence of high concentrations of Ca2� or Zn2� duringtransit from ER through the Golgi (38, 39). WT-hGH containswell defined Zn2� binding sites that induce cooperativedimerization of hGH (40), and these may facilitate the same
FIG. 7. GH contents in �exon3hGH transgenic mice. Pituitary GH contents were measured by RIA in pituitaries harvested from males (left panels)and females (right panels) at two different ages. Upper panels, Line no. 1; lower panels, line no. 23. In both lines, and for both sexes, transgenicanimals (T, dark bars) had lower pituitary GH contents than their NT littermates (open bars). GH deficiency was evident earlier and was muchmore profound in line no. 1, but also developed with time in line no. 23. **, P � 0.01, ***, P � 0.001 vs. age-matched NT animals. #, P � 0.05,3 wk vs. 10 wk in T animals.
TABLE 1. Pituitary hormone contents in �exon3hGH transgenic mice
Hormonecontent (�g/pit) Age (wk) NT T line no. 1 T line no. 23
Mouse PRL 3–4 1.5 � 0.5 0.75 � 0.23a 1.05 � 0.58–10 2.4 � 0.8 0.25 � 0.1b 1.32 � 0.25a
Mouse TSH 3–4 0.352 � 0.047 0.153 � 0.06a 0.307 � 0.0958–10 0.587 � 0.052 0.079 � 0.02b 0.595 � 0.07
Mouse LH 3–4 0.362 � 0.07 0.339 � 0.075 0.395 � 0.098–10 0.7 � 0.095 0.450 � 0.042a 0.756 � 0.12
PRL, TSH, and LH were measured by RIAs in pituitary homogenates from groups of male transgenic animals (T) and nontransgeniclittermates (NT, line no. 1) at different ages and expressed as �g/pit (mean � SEM, n 6 per group, a P � 0.05, b P � 0.001 vs. NT).
McGuinness et al. • Autosomal Dominant GH Deficiency Endocrinology, February 2003, 144(2):720–731 727

oligomerization/insolubilization process for GH SV cores, ashas been well established for insulin hexamers (41). The(Zn2�-hGH)2 complex is stable and resistant to denaturationduring storage (40), and all of the identified Zn2� bindingresidues in hGH are present in �exon3hGH, though recentwork suggests these may not be a prerequisite for conden-sation and aggregation (42).
We propose the following hypothesis to explain the dom-inant negative effects of �exon3hGH: the 17.5-kDa hGH prod-uct progresses through the regulated secretory pathway,where it can form heterodimers with WT-hGH. Some WT-hGH homodimers may also form, but further oligomeriza-tion with heterodimers occurs poorly so efficient packingand condensation cannot occur, blocking the formation ofdense-cored GH SVs from the trans-Golgi. Because the�exon3hGH:WT-hGH complexes are stable, but cannot exitefficiently via SVs, both WT-hGH and �exon3hGH accumulatein the Golgi, and back up in the ER. This triggers the mis-folded protein response and the complexes are transportedinto the cytosol. Once the production of �exon3hGH:WT-hGHcomplexes exceeds the degradative capacity of the protea-some pathway, they begin to accumulate as aggregates in thecytosol, ER and Golgi, eventually proving toxic to the cell.For GH-producing cells in culture, cell death occurs as anautolytic process; in vivo the process is greatly accelerated by
increased trophic drive from GHRH to increase expression ofboth hGH and �exon3hGH, to replace and expand the (de-fective) somatotroph population, and by an invasion of ac-tivated macrophages to destroy defective cells.
This hypothesis would explain why there is no toxic effectof �exon3hGH when expressed alone or with proteins otherthan GH in other cell types, and also why a dominant effectdoes not usually occur when coexpressed with WT-hGH innonsecretory cells (7). Were �exon3hGH unable to interactwith WT-hGH, then the WT-hGH proteins should homo-oligomerize from the mixture and form a population of nor-mal dense cored SVs containing WT-hGH. Furthermore, therate or extent of cellular damage would be proportional to theamount and rate of traffic of both WT-hGH and �exon3hGHfor SV packaging, being milder in the less granular GC cells,and more severe in highly granulated somatotrophs. In the�exon3hGH-GC cultures, it was possible to observe cells atintermediate stages of morphological disruption, and thoughtheir growth rates were compromised, stable �exon3hGH celllines could be obtained. In contrast, the same features ofdegranulation and morphological disruption caused by�exon3hGH had catastrophic consequences for the soma-totroph population in the transgenic animals.
Our hypothesis would also predict that the onset, severityand rate of progression would be proportional to the relative
FIG. 8. Somatotroph disruption in �exon3hGH transgenic mice. Immunogold EM of pituitary sections from transgenic mice of line no. 1 showeda massive depletion of recognizable somatotrophs, and the remaining GH cells were grossly abnormal with few if any dense cored SVs comparedwith the abundant somatotrophs filled with many GH-containing SVs clearly labeled with immunogold (arrows) in NT mice (F). Many cellswere clearly disintegrating (A) or had dilated mitochondria or vacuolated cytoplasm (B). Many macrophages were evident throughout thepituitary (C and D), many filled with lipid accumulations and immunostained remnants (D). A few cells contained dense vesicular structuresthat immunostained for mouse GH, but these were irregular and misshapen (E). n, Nucleus; L, lipid; arrows, anti-GH immunogold; M,macrophage; m, mitochondria; v, vacuole. Scale bar, 1 �m.
728 Endocrinology, February 2003, 144(2):720–731 McGuinness et al. • Autosomal Dominant GH Deficiency

amounts of exon3-hGH vs. WT-hGH expressed. Analysis of thedifferent lines of transgenic mice was consistent with this,because the onset, severity, and specificity of their IGHDIIphenotype was proportionate to their transgene copy num-ber. We could not establish the relative RNA or �exon3hGHprotein levels directly in pituitary extracts because many ofthe cells were destroyed, and we were unable to detect the17.5-kDa protein. However, we can assume that there is more�exon3hGH expression in the high copy lines because thistransgene LCR shows reliable copy number-dependent andposition-independent expression (14, 43). The most severelyaffected lines were already GH deficient at weaning anddeveloped proportionate reductions in weight and lengthand bone growth, more marked in males than in females, asin other transgenic dominant dwarf animals (18). Althoughthe low copy number line showed a milder phenotype, withrelatively normal postweaning growth, all animals eventu-ally developed pituitary GH deficiency with time.
It is interesting to compare this murine phenotype withhuman IGHDII, which also shows variability in onset, se-verity, and progression, even within the same family (44). Inhuman IGHDII, the allele ratio of WT-hGH:mutant hGH is1:1, and it is assumed that each allele is transcribed equiv-
alently in hemizygous individuals, although it has not beenestablished that they generate equivalently stable or trans-latable RNA products. Severe short stature was only presentin one third of the affected individuals at diagnosis in thestudy by Binder et al. (44), although children with splice sitemutations were on average younger and shorter at diagnosisthan those with the missense mutations. This argues for amore pronounced effect of a dominant negative mutationthan of haploinsufficiency per se, although the severity ofphenotype will also clearly depend on the nature of themissense mutation and the degree to which it disrupts GHstructure.
Variability in effects on pituitary size also appears in hu-man subjects with IGHDII; MR imaging in four childrenshowed a normal adenohypophysis in two cases and mildhypoplasia in two others (44). However, a variety of differentmutations are associated with IGHDII (3, 4, 21, 22, 44), in-cluding alterations or deletions in splice enhancers, whichcan give rise to different ratios of WT-hGH and �exon3hGHtranscripts (Ryther, R., L. McGuinness, J. Phillips, C. Mose-ley, C. Magoulas, I. C. Robinson, and J. Patton, in prepara-tion). Although the correlation between the various GHmRNA isoforms, the amounts of their protein products, and
FIG. 9. GHRH and somatostatin expression in �exon3hGH transgenic mice. In situ hybridization was carried out on hypothalamic sections fromNT and �exon3hGH transgenic mice (line no. 1), using radiolabeled antisense riboprobes for GHRH (left panels) and somatostatin (right panels).Individual examples are shown in the upper panel, and the results from image analysis on arcuate (ARC) and periventricular nuclei (PeN) onsimilar sections from groups of animals are shown in the lower panels. ARC GHRH expression was significantly up-regulated, whereas PeNsomatostatin expression significantly reduced in the same group of animals. ZI, Zona incerta. **, P � 0.01 vs. NT controls (n 6 per group).
McGuinness et al. • Autosomal Dominant GH Deficiency Endocrinology, February 2003, 144(2):720–731 729

the extent of pituitary damage caused remains to be estab-lished, our results suggest that variability in the ratios andamounts of 17.5- to 22-kDa isoforms produced could well bean important contributor to the variability of individual phe-notype in some forms of human IGHDII.
Unexpectedly, both lines 1 and 12 were subfertile. This isunlikely to be due to GH deficiency or body size per se,because other equally small, equally GH-deficient mice (ex-pressing different transgenes on the same genetic back-ground) are normally fertile (Robinson, I. C. A. F., unpub-lished). It is more likely due to the other pituitary hormonedeficiencies that developed in the most severely affectedlines. A fall in PRL was expected because most models of GHcell hypoplasia or ablation also show reduced PRL (18, 45).One or two surviving GH-immunopositive cells in the�exon3hGH mice presented an appearance under EM moreresembling lactotrophs, typified by irregularly shaped SVs.There is a small population of pituitary mammosoma-totrophs that express both GH and PRL, and these mayexpress less GH (and hence less �exon3hGH) per cell than dosomatotrophs. Because �exon3hGH does not block PRL secre-tion, mammosomatotrophs might be able to package somePRL into SVs and with less material accumulating in thecytosol, could perhaps survive longer than somatotrophs.
More surprising was the loss of other pituitary hormonesin the high copy lines. Snell and Jackson dwarf mice withmutations in Pit-1 (46) are deficient in TSH as well as GH andPRL, and it is possible that multiple copies of Pit-1 elementsin the GH1 transgene promoters compete for the availablePit-1 and reduce the transcription of other endogenousPit-1-dependent genes, such as TSH. However, this expla-nation would not explain the fall in LH and gonadotropenumbers visible by EM. Large numbers of activated macro-phages were evident in the high copy lines, especially at theintermediate lobe/anterior pituitary boundary where newGH cells first appear during development (47) so these arewell placed to destroy newly emerging defective somato-tropes as they differentiate from progenitor cells. Becausethis transgene LCR reliably restricts transgene expression tothe somatotrope (13, 14, 43), we suggest that the massive andrapid autodestruction of GH cells induced by high expres-sion of �exon3hGH activates an inflammatory macrophageresponse resulting in significant bystander endocrine cellkilling in these high copy lines.
The phenotype in the line no. 23 is probably a closer modelfor human IGHDII, in which the hormone deficiency appearslargely confined to the GH axis (4, 21, 26, 44). Most reportssuggest normal thyroid and adrenal function and normalplasma PRL levels in IGHDII. However, because multiplepituitary hormone deficiencies may evolve in some childreninitially diagnosed with isolated GHD, it may be importantto reinvestigate older subjects with severe IGHDII diagnosedand treated in childhood, to see whether further pituitaryhormone deficits emerge with time.
Our in vivo model also allowed us to investigate hypo-thalamic changes in IGHDII for the first time. GH normallyregulates its own production by both direct and indirectfeedback, repressing GHRH and increasing somatostatin ex-pression, respectively (16, 19, 48). As expected, lack of GHfeedback in the �exon3hGH transgenic mice was associated
with increased arcuate GHRH and decreased periventricularsomatostatin expression compared with their NT littermatecontrols. We believe this may be an important additionalfactor that accelerates the rate of progression of IGHDII invivo. The increased GHRH drive that stimulates somatotrophproliferation and WT-GH transcription will also increasetranscription of �exon3hGH, compounding the cellular block-ade. Progressive GH deficiency and GHRH up-regulationwould then form a vicious cycle to accelerate the productionand autodestruction of the GH cell population, rapidly ex-hausting the capacity to generate new GH cells.
A reduction in GH cell number may ultimately be moreimportant than compromised GH cell function in the longerterm. Early treatment of IGHDII with exogenous GH re-placement therapy may be important in rescuing a degree ofpituitary function by providing a feedback signal to reducethe GHRH drive, reducing somatotroph proliferation andrate of self-destruction. A secondary benefit could be to re-duce rate of the pituitary damage and hence loss of otherendocrine cell types in IGHDII. If so, precipitate withdrawalof GH treatment following attainment of adult height inIGHDII could be deleterious.
Acknowledgments
We are very grateful to Dr. A. L. Parlow and to the NIDDK for thecontinued provision of assay reagents, and to Dr. Nancy Cooke forproviding us with the original hGH LCR cosmid.
Received August 13, 2002. Accepted October 4, 2002.Address all correspondence and requests for reprints to: Professor
Iain C. A. F. Robinson, Division of Molecular Neuroendocrinology,National Institute for Medical Research, The Ridgeway, Mill Hill, Lon-don NW7 1AA, United Kingdom. E-mail: [email protected].
* Current address: Department of Neurosurgery, Barts, and The Lon-don School of Medicine and Dentistry, Turner Street, London E1 2AD,United Kingdom.
References
1. Baumann G 1991 Growth hormone heterogeneity: genes, isohormones, vari-ants, and binding proteins. Endocr Rev 12:424–449
2. Binder G, Ranke MB 1995 Screening for growth hormone (GH) gene splice-sitemutations in sporadic cases with severe isolated GH deficiency using ectopictranscript analysis. J Clin Endocrinol Metab 80:1247–1252
3. Phillips 3rd JA, Cogan JD 1994 Genetic basis of endocrine disease. 6. Molecularbasis of familial human growth hormone deficiency. J Clin Endocrinol Metab78:11–16
4. Cogan JD, Ramel B, Lehto M, Phillips 3rd J, Prince M, Blizzard RM, de RavelTJ, Brammert M, Groop L 1995 A recurring dominant negative mutationcauses autosomal dominant growth hormone deficiency—a clinical researchcenter study. J Clin Endocrinol Metab 80:3591–3595
5. Ultsch MH, Somers W, Kossiakoff AA, de Vos AM 1994 The crystal structureof affinity-matured human growth hormone at 2 A resolution. J Mol Biol236:286–299
6. Krawczak M, Reiss J, Cooper DN 1992 The mutational spectrum of singlebase-pair substitutions in mRNA splice junctions of human genes: causes andconsequences. Hum Genet 90:41–54
7. Hayashi Y, Yamamoto M, Ohmori S, Kamijo T, Ogawa M, Seo H 1999Inhibition of growth hormone (GH) secretion by a mutant GH-I gene productin neuroendocrine cells containing secretory granules: an implication for iso-lated GH deficiency inherited in an autosomal dominant manner. J Clin En-docrinol Metab 84:2134–2139
8. Lee MS, Wajnrajch MP, Kim SS, Plotnick LP, Wang J, Gertner JM, Leibel RL,Dannies PS 2000 Autosomal dominant growth hormone (GH) deficiency typeII: the Del32-71-GH deletion mutant suppresses secretion of wild-type GH.Endocrinology 141:883–890
9. Graves TK, Patel S, Dannies PS, Hinkle PM 2001 Misfolded growth hormonecauses fragmentation of the Golgi apparatus and disrupts endoplasmic retic-ulum-to-Golgi traffic. J Cell Sci 114:3685–3694
10. Dannies PS 1999 Protein hormone storage in secretory granules: mechanismsfor concentration and sorting. Endocr Rev 20:3–21
730 Endocrinology, February 2003, 144(2):720–731 McGuinness et al. • Autosomal Dominant GH Deficiency

11. Dannies PS 2000 Protein folding and deficiencies caused by dominant-negative mutants of hormones. Vitam Horm 58:1–26
12. Binder G, Brown M, Parks J 1996 Mechanisms responsible for dominantexpression of human growth hormone gene mutations. J Clin EndocrinolMetab 81:4047–4050
13. Magoulas C, McGuinness L, Balthasar N, Carmignac DF, Sesay AK, MathersKE, Christian H, Candeil L, Bonnefont X, Mollard P, Robinson IC 2000 Asecreted fluorescent reporter targeted to pituitary growth hormone cells intransgenic mice. Endocrinology 141:4681–4689
14. Jones BK, Monks BR, Liebhaber SA, Cooke NE 1995 The human growthhormone gene is regulated by a multicomponent locus control region. Mol CellBiol 15:7010–7021
15. Tashjian Jr AH, Yasumura Y, Levine L, Sato GH, Parker ML 1968 Establish-ment of clonal strains of rat pituitary tumor cells that secrete growth hormone.Endocrinology 82:342–352
16. Pellegrini E, Carmignac DF, Bluet-Pajot MT, Mounier F, Bennett P, Epel-baum J, Robinson IC 1997 Intrahypothalamic growth hormone feedback: fromdwarfism to acromegaly in the rat. Endocrinology 138:4543–4551
17. Carmignac DF, Robinson ICAF 1990 Growth hormone (GH) secretion in thedwarf rat: release, clearance and responsiveness to GH-releasing factor andsomatostatin. J Endocrinol 127:69–75
18. Flavell DM, Wells T, Wells SE, Carmignac DF, Thomas GB, Robinson ICAF1996 Dominant dwarfism in transgenic rats by targeting human growth hor-mone (GH) expression to hypothalamic GH-releasing factor neurons. EMBOJ 15:3871–3879
19. Bennett PA, Levy A, Sophokleous S, Robinson ICAF, Lightman SL 1995Hypothalamic GH receptor gene expression in the rat: effects of altered GHstatus. J Endocrinol 147:225–234
20. Nakane PK 1975 Identification of anterior pituitary cells by electron micros-copy. In: Tixier-Vidal A, Farquhar M, eds. The anterior pituitary gland. NewYork: Academic Press; 134–158
21. Missarelli C, Herrera L, Mericq V, Carvallo P 1997 Two different 5� splice sitemutations in the growth hormone gene causing autosomal dominant growthhormone deficiency. Hum Genet 101:113–117
22. Hayashi Y, Kamijo T, Yamamoto M, Ohmori S, Phillips 3rd JA, Ogawa M,Igarashi Y, Seo H 1999 A novel mutation at the donor splice site of intron 3of the GH-I gene in a patient with isolated growth hormone deficiency. GrowthHorm IGF Res 9:434–437
23. Kim PS, Arvan P 1998 Endocrinopathies in the family of endoplasmic retic-ulum (ER) storage diseases: disorders of protein trafficking and the role of ERmolecular chaperones. Endocr Rev 19:173–202
24. Olias G, Richter D, Schmale H 1996 Heterologous expression of humanvasopressin-neurophysin precursors in a pituitary cell line: defective transportof a mutant protein from patients with familial diabetes insipidus. DNA CellBiol 15:929–935
25. Cogan JD, Phillips 3rd JA, Schenkman SS, Milner RD, Sakati N 1994 Familialgrowth hormone deficiency: a model of dominant and recessive mutationsaffecting a monomeric protein. J Clin Endocrinol Metab 79:1261–1265
26. Deladoey J, Stocker P, Mullis PE 2001 Autosomal dominant GH deficiencydue to an Arg183His GH-1 gene mutation: clinical and molecular evidence ofimpaired regulated GH secretion. J Clin Endocrinol Metab 86:3941–3947
27. Thompson NL, Lagerholm BC 1997 Total internal reflection fluorescence:applications in cellular biophysics. Curr Opin Biotechnol 8:58–64
28. Steyer JA, Almers W 1999 Tracking single secretory granules in live chro-maffin cells by evanescent-field fluorescence microscopy. Biophys J 76:2262–2271
29. Arvan P, Castle D 1998 Sorting and storage during secretory granule biogen-esis: looking backward and looking forward. Biochem J 332:593–610
30. Hammond C, Helenius A 1995 Quality control in the secretory pathway. CurrOpin Cell Biol 7:523–529
31. Huang XF, Arvan P 1995 Intracellular transport of proinsulin in pancreatic�-cells. Structural maturation probed by disulfide accessibility. J Biol Chem270:20417–20423
32. Pelham HR 1989 Control of protein exit from the endoplasmic reticulum. AnnuRev Cell Biol 5:1–23
33. Takahashi Y, Kaji H, Okimura Y, Goji K, Abe H, Chihara K 1996 Brief report:short stature caused by a mutant growth hormone. N Engl J Med 334:432–436
34. Werner ED, Brodsky JL, McCracken AA 1996 Proteasome-dependent endo-plasmic reticulum-associated protein degradation: an unconventional route toa familiar fate. Proc Natl Acad Sci USA 93:13797–801
35. Gething MJ, Sambrook J 1990 Transport and assembly processes in the en-doplasmic reticulum. Semin Cell Biol 1:65–72
36. Schwartz AL, Ciechanover A 1999 The ubiquitin-proteasome pathway andpathogenesis of human diseases. Annu Rev Med 50:57–74
37. Igarashi Y, Ogawa M, Kamijo T, Iwatani N, Nishi Y, Kohno H, MasumuraT, Koga J 1993 A new mutation causing inherited growth hormone deficiency:a compound heterozygote of a 6.7 kb deletion and a two base deletion in thethird exon of the GH-1 gene. Hum Mol Genet 2:1073–1074
38. Colomer V, Kicska GA, Rindler MJ 1996 Secretory granule content proteinsand the luminal domains of granule membrane proteins aggregate in vitro atmildly acidic pH. J Biol Chem 271:48–55
39. Orci L, Ravazzola M, Amherdt M, Perrelet A, Powell SK, Quinn DL, MooreHP 1987 The trans-most cisternae of the Golgi complex: a compartment forsorting of secretory and plasma membrane proteins. Cell 51:1039–1051
40. Cunningham BC, Mulkerrin MG, Wells JA 1991 Dimerization of humangrowth hormone by zinc. Science 253:545–548
41. Whittingham JL, Chaudhuri S, Dodson EJ, Moody PC, Dodson GG 1995X-ray crystallographic studies on hexameric insulins in the presence of helix-stabilizing agents, thiocyanate, methylparaben, and phenol. Biochemistry 34:15553–15563
42. Sankoorikal B-J, Zhu Y, Hodsdon ME, Lolis E, Dannies PS 2002 Aggregationof human wild-type and H27A-prolactin in cells and in solution: roles of Zn2�,Cu2�, and pH. Endocrinology 143:1302–1309
43. Su Y, Liebhaber SA, Cooke NE 2000 The human growth hormone gene clusterlocus control region supports position-independent pituitary- and placenta-specific expression in the transgenic mouse. J Biol Chem 275:7902–7909
44. Binder G, Keller E, Mix M, Massa GG, Stokvis-Brantsma WH, Wit JM, RankeMB 2001 Isolated GH deficiency with dominant inheritance: new mutations,new insights. J Clin Endocrinol Metab 86:3877–3881
45. Behringer RR, Mathews LS, Palmiter RD, Brinster RL 1988 Dwarf miceproduced by genetic ablation of growth hormone-expressing cells. Genes Dev2:453–461
46. Li S, Crenshaw 3rd EB, Rawson EJ, Simmons DM, Swanson LW, RosenfeldMG 1990 Dwarf locus mutants lacking three pituitary cell types result frommutations in the POU-domain gene pit-1. Nature 347:528–533
47. Lin SC, Lin CR, Gukovsky I, Lusis AJ, Sawchenko PE, Rosenfeld MG 1993Molecular basis of the little mouse phenotype and implications for cell type-specific growth. Nature 364:208–213
48. Burton K, Kabigting E, Steiner R, Clifton D 1995 Identification of target cellsfor growth hormone’s action in the arcuate nucleus. Am J Physiol 269:E716–E722
McGuinness et al. • Autosomal Dominant GH Deficiency Endocrinology, February 2003, 144(2):720–731 731
Copyright © 2022 FDOKUMEN




















