
Depositional environment and sequence stratigraphy of the Oligo-Miocene Asmari Formation in SW Iran
Upload
independentCategory
view
1download
0
491
Journal of Andrology, Vol. 16, No. 6, November/December 1995Copyright C American Society of Andrology
Impaired Secretory Function of the Prostate inMen With Oligo-Asthenozoospermia
GORAN AHLGREN,* GIJNNAR RANNEVIK,t AND HANS LILJAt
From the Departments of *Urology and *Clinical Chemistry, Lund University, Malm#{246}General Hospital and the
tFertility Research Institute, Malm#{246},Sweden.
ABSTRACT: The secretory function of the human prostate and theseminal vesicles is a prerequisite for gel formation and liquefaction
of semen, but the relation to poor sperm motility and low spermcount in infertile men remains to be clarifyed. Our aim was to evaluatethe secretory function of the prostate and the seminal vesicles innormozoospermic men (n = 35) and in asthenozoospermic men, whowere all also oligozoospermic (n = 27). All 62 subjects belonged tocouples undergoing routine infertility evaluation. In liquefied seminalfluid we measured the concentrations of fructose and protein Cinhibitor (PCI) contributed by the seminal vesicles, PCI complexedto prostate-specific antigen (PSA), and the prostatic contribution ofzinc, PSA, acid phosphatase (PAP), -microseminoprotein (19-MSP),and Zna2-glycoprotein (Zna2-GP). The concentration of each pro-static secretory protein correlated significantly with that of zinc (P< 0.01) in both the normozoospermic (NZS) and oligo-astheno-zoospermic (OAZS) subgroups, but the PCI concentration did not
correlate significantly with that of fructose. There was no significantdifference between the NZS and OAZS subgroups In ejaculate vol-ume or secretory contribution from the seminal vesicles, whereasthe OAZS subgroup was characterized by significantly lower secre-tory contributions of Zna2-GP (P = 0.001), Zn, PSA, PAP (P < 0.01),and ll-MSP (P < 0.05). The two subgroups did not differ significantlyin the serum concentration of luteinizing hormone (LH), testosterone,or sex hormone-binding globulin (SHBG). The results thus suggestthe secretory contribution of major prostatic proteins and zinc perejaculate to be significantly decreased in ollgo-asthenozoospermlcmen. The importance of this finding in relation to poor sperm countand motility as indicators of Impaired gonadal function requires fur-ther investigation.
Key words: Seminal plasma, infertility, PSA, seminal vesicles.J Androl 1 995;1 6:49 1-498
t ejaculation, the small volume of sperm dense fluid
rom the epididymal cauda mixes with the secretory
contributions from the prostate and the seminal vesicles.The contribution from the seminal vesicles accounts for
about 60% of the normal ejaculate volume of 2-6 ml
(Mann and Lutwak-Mann, 1981; Kvist et al, 1990). The
bulk of the ejaculate is immediately transformed into a
gel as the secretions from the seminal vesicles containing
the gel forming proteins are mixed with the secretion from
the prostate. The gel may provide a neutral pH environ-
ment for the spermatozoa to protect them from exposure
to the acidic pH in the vagina. The gel dissolves and
liquefies within 15-20 minutes when the gel-forming pro-
teins are split into multiple fragments due to the action
of the major prostate-derived protease, prostate-specific
antigen (PSA) (Lilja et al, 1984, 1987; Lilja and Laurell,
1985; McGee and Herr, 1988; Lee et al, 1989; Chris-
tensson et al, 1990). The importance of this process in
normal fertilization is not known, and the secretory func-
tion of the accessory sex glands in infertile men remains
to be clarified.
Correspondence to:Dr. Gdran Ahigren, Department of Urology, Mal-m#{246}University Hospital, S-205 02 MalmO, Sweden.
Received for publication February 6, 1995; accepted for publication
August 7, 1995.
To study the secretory function of the male accessory
sex glands in infertile men with poor sperm motility, we
measured the concentrations of the major secretory com-
ponents of the prostate and the seminal vesicles in the
seminal fluid from normozoospermic and asthenozoo-
spermic men belonging to couples undergoing routine in-
fertility evaluation. In addition, blood serum levels offollicle-stimulating hormone (FSH), luteinizing hormone
(LH), testosterone, and sex hormone-binding globulin(SHBG) were measured to evaluate the androgenic controlof these glands.
Materials and Methods
Subjects and Study Procedure
Semen samples were collected during 1990 and 1991 at theFertility Clinic Ellenbogen, Malm#{246},from males undergoing se-men analysis as part of an infertility evaluation. The sampleswere obtained by masturbation after a period of at least 2 days
of sexual abstinence, and they were allowed to liquefy at roomtemperature. The time of abstinence was (mean ± standard de-
viation [SD]) 5.0 ± 1.8 days in the nonmozoospermic (NZS)
group (range 2-10 days) and 5.2 ± 2.3 days in the oligo-asthen-
ozoospermic (OAZS) group (range 2-10 days).
An aliquot was used for routine semen analysis, which was

492 Journal of Andrology . November/December 1995
Detection limits, intraassay coefficient of variation (CV), and
performed by the same person in all cases, using methods ofsemen assessment based on those described in the WHO Lab-
oratory Manual (World Health Organization, 1987) and com-prising measurement of volume, sperm count, percent motile
sperm, and grading of motility as well as morphological classi-fication of spermatozoa. Data about consistency and sperm vi-
tality are not given. In cases where there was an increased number
of cellular elements other than spermatozoa andlor increasedagglutination, routine semen culture for bacteria was performed.Of 17 semen cultures for bacteria (all from oligo-asthenozoo-spermic men), 5 were positive, and 2 of them had significant
growth of pathogenic bacteria.A capillarytube testwith Sperm
Select#{174}(Pharmacia AB, Sweden) was used as an in vitro pene-
tration test.
After separation of the aliquot,benzamidine was added-within
1 hour afterejaculation-to a concentration of 100 mmolJL to
prevent further degradation of the seminal plasma proteins. Sem-
inal plasma was separated as the supernatant obtained after cen-
trifugation at 1,000 x g for 10 minutes, and the samples were
kept frozen at - 20#{176}Cuntil subsequent biochemical analysis.
Venous blood samples were drawn between 8:00 and 9:00 i,
and sera were used for the assay of hormones and SHBG.
Consecutive samples from asthenozoospermic men were se-
lected for the study, but all of these samples also manifested
oligozoospermia (OAZS subgroup; n = 27; mean age 32.6 ± 5.9years, range 23-44). Samples with severe oligozoospermia (<1x 106 sperm/mI) were excluded because of inaccuracy in motility
measurement. Samples from normozoospermic men from the
same period were included in the study (NZS subgroup; n = 35;
mean age 33.1 ± 5.5 years,range 21-46). Only one sample from
each subject was used. However, to study the intraindividual
variation of the seminal plasma parameters, seminal plasma
from two separate ejaculatesfrom 13 men (10 OAZS and 3 NZS)were analyzed.
Serum Hormone and SHBG Assays
The serum concentrations of hormones as well as SHBG were
determined by radioimmunoassay (RIA). All assays were carried
out in duplicate. Serum concentrations of FSH and LH were
analyzed by means of a double-antibody RIA technique and
measured using human (h) FSH (lot, no. 090982; Akers Hospital,
Oslo, Norway) and hLH (Division of Biological Standards no.
6 8/40 = 1St IRP) and calibrated against WHO FSH 7 8/549 and
WHO LH 68/40, respectively. Testosterone concentrations were
determined after ether extraction, as described previously (Tho-
rell and Larson, 1978). The assay procedures were conducted in
accordance with the original protocols with the following excep-
tions: anti-testosterone-3-(O-carboxy-methyl-oxime) and bo-
vine serum albumin (rabbit) (Euro-Diagnostica AB, Malm#{246},
Sweden) were used as antibody and ‘251-labeled derivative was
used as the tracer.
SHBG was measured with a commercial kit obtained from
Euro-Diagnostica AB, using polyclonal rabbit antiserum and
pure ‘251-labeled, native human SHBG, based on a technique
previously described (Fernlund et al, 1985). SHBG values in
mg/L were multiplied by 10.7 to convert them to SI units (nmoll
interassay CV were as follows: for FSH 1.0 IU/L, 5%, and 8%;
for LH 1.0 IU/L, 6%, and 9%; for testosterone 0.1 nmollL, 6%,
and 10%; for SHBG 5 g/L (corresponding to about 0.2 mg/L
for samples diluted 1:40 in routine procedures), 4%, and 8%.
Biochemical Analysis of Seminal Plasma
The concentration of each variable was measured, and the total
contributed amount was calculated from the volume of the ejac-
ulate. The following methods were used for analysis of the dif-
ferent parameters from the two glands.
Prostate-Prostate-specific antigen (PSA): Seminal plasma di-
luted in Delphia#{174} assay buffer was analyzed with a two-site
immunometric assay using a monoclonal solid phase catching
antibody and a monoclonal Eu-labeled tracer antibody (Delphia,
Wallac), detecting the free form of PSA and that complexed to
protein C inhibitor (PCI) or a 1-antichymotrypsin (ACT) in an
almost equimolar fashion (Lilja et al, 1991; Christensson and
Lilja, 1994). The intraassay CV was <7%, and the interassay
CV was <8%. Acid phosphatase (PAP): Seminal plasma diluted
in assay buffer was analyzed with a commercial two-site im-
munometric kit (Delphia PAP kit, Wallac), based on monoclonal
antibodies. The intraassay CV was <5%, and the interassay CV
was <7%. 3-Microseminoprotein (jI-MSP): Seminal plasma was
diluted and analyzed with a competitive RIA as previously re-
ported, using 1251 9-MSP as tracer (Abrahamsson et al, 1989).
The intraassay CV was <8%. Zna2-glycoprotein (Zna2-GP):
Concentrations of Zna2-GP were analyzed by electroimmu-
noassay in 75 mmolJL sodium barbitalurate, pH 8.6, containing
2 mmolJL Ca2, as outlined by Laurell and McKay (1981), using
a monospecific polyclonal antiserum that was a gift from Dr. A
Grubb, University of Lund. All Zna2-GP-concentrations are giv-
en in percent of the level in a human reference serum (Seronorm
Human, Nycomed#{174}). Zn: The seminal plasma concentration of
Zn was analyzed with a Varian AA-775 atomic absorption spec-
trophotometer, as described by Parker et al (1967). The intraas-
say CV was <5%, and the interassay CV was <3%. PSA-PCI
complex: Seminal plasma diluted in Delphia assay buffer was
analyzed with a two-site immunometric method, using a mono-
clonal catcher anti-PCI antibody and a monoclonal Eu-labeledanti-PSA tracer IgG antibody (Christensson and Lilja, 1994).
The intraassay CV was <3%, and the interassay CV was <11%.
Seminal vesicle-Fructose: Fructose was determined as the
difference between the initial glucose concentration of the sample
and that resulting from treatment with phosphoglycosisomerase
(essentially according to the procedure described by Wetterauer
in 1976). The intraassay CV was <3%, and the interassay CV
was <5%. PCI: Seminal fluid was diluted and analyzed with a
two-site immunoradiometric method with a solid-phase poly-
clonal anti-PCI catcher antibody and a ‘251-labeled monoclonal
anti-PCI antibody (Ml 1-5) as the tracer, as described above.
This method is sensitive for PCI as a native inhibitor, as a
proteolytically modified inhibitor, and as complexed to a pro-
tease such as PSA (Laurell Ct al, 1992). The intraassay CV was
<4%, and the interassay CV was <13%.
Statistics
All results of semen analysis as well as serum hormone concen-
trations and SHBG are given as mean values, using a t-test for

Ahigren et al Impaired Secretory Function in Oligo-Asthenozoospermia 493
Table 1. Standard variables of semen analysis in the two su bgroups of patients
Oligo-asthenozoospermic (nVariables Normozoospermic (n = 35) = 27) P v#{224}luet
Volume (ml) 4.2 ± 1.6 (2.2-10.5) 3.9 ± 1.9 (1.4-8.6) NSTotal sperm count (spermatozoa x 10#{176}/ejaculate) 378 ± 245 (78-940) 14.3 ± 11.9 (3.0-36.0) <0.0001Motility (percent rapidly progressive sperm) 54 ± 12 (30-84) 7.4 ± 7.4 (0-25) <0.0001Morphology (percent of normal forms) 63 ± 10 (34-80) 29 ± 13 (8-67) <0.0001Penetration (score 0-9) 7.5 ± 1.2 (5-9) 1.0 ± 1.6 (0-7) <0.0001
Data are presented as mean ± SD and range.
* Normal values of semen variables (World Health Organization, 1987): volume �2 ml, total sperm count �40 x 10#{176}spermatozoa per ejaculate,
motilIty >25% with rapid progression (category ‘a’), morphology �30% normal forms.
t P values were obtained from t-tests.
Table 2. Serum hormone con centrations in the two subgroups of patients
Variables* Normozcospermic (n = 33) Oligo-asthenozoospermic (n = 27) P valuet
FSH, lU/LLH, lU/LTestosterone, nmol/LSHBG, mg/LT/SHBG ratio
5.4 ± 2.0 (3.0-1 0.8)4.5 ± 1.4 (2.6-7.8)
19.5 ± 6.0 (8.0-32.0)2.2 ± 0.8 (1.1-3.9)9.3 ± 3.0 (4.0-1 7.0)
7.8 ± 4.1 (2.8-22.8)5.0 ± 1.7 (2.6-9.6)
19.6 ± 9.2 (9.5-44.0)2.0 ± 0.9(0.6-4.0)
10.3 ± 3.4 (5.5-20.0)
0.008NSNSNS
NS
Data are presented as means ± SD and ranges.* Reference values. FSH 1 0 lU/L, LH 9 lU/L, testosterone (T) 10 nmol/L, SHBG s4.1 mg/L, T/SHBG ratio >4.
t P values were obtained from t-tests.
statistical comparison. Because the NZS subgroup was too small
to determine the normal distribution of the secretory compo-
nents in seminal plasma, all values for this subgroup are givenas median values, ranges, and percentiles. The statistical signif-
icance of differences for these variables was tested with the Mann-Whitney U-test, and correlation was analyzed with Spearman’s
rank correlation coefficient.
Results
I. Sperm Characteristics and Serum Hormoneconcentrations
The sperm characteristics of the NZS and OAZS sub-
groups are given in Table 1. As expected from patientselection, differences between the two subgroups regarding
all sperm variables were statistically significant except for
the volume of the ejaculate. Mean values of serum hor-
mones and SHBG are given in Table 2. There was no
significant difference in serum concentrations of testos-
terone, LH, SHBG, or testosterone-to-SHBG ratio be-
tween the two subgroups. The mean serum FSH concen-
tration was significantly higher in the OAZS subgroup,mainly due to five men with levels above the reference
range.
II. Evaluation of Secretoty Proteins as SpecificMarkers of Accessory Sex Gland Function
To study covariation of secretory products from each gland
in the NZS subgroup and OAZS subgroup separately, the
concentrations of secretory proteins and other markers
were analyzed (Tables 3 and 4). Zinc and fructose, re-
spectively, were used as reference markers of prostate and
seminal vesicle secretory function.
In both the NZS and OAZS subgroups, the concentra-tion of zinc manifested a strong correlation to those of
PSA and PAP (P < 0.001), but only a moderate corre-
lation to that of i9-MSP (P < 0.01). The concentration of
zinc also manifested a strong correlation to that of Zna2-
OP in the NZS subgroup (P < 0.001), but this correlation
was much weaker in the OAZS subgroup (P < 0.05). There
was no correlation between the concentration of any pros-
tate-derived variable and that of PCI or fructose. There
was no correlation between the concentration of fructose
Table 3. Concentrations in seminal plasma of secretory productsfrom the accessory sex glands in the normozoospermic (NZS) andoligo-asthenozoospermic (OAZS) men
Variables
Concentrations(NZS subgroup
n = 35)
Concentrations(OAZS subgroups
n = 27)
Seminal vesicleFructose (mmol/L)PCI (mg/L)
PCI-PSA (mg/L)
14.2 (2.9-25.0)180 (100-320)
89 (29-192)
14.6 (4.5-29.2)184 (75-383)
73 (22-176)
Prostrate
Zn (mmol/L)PSA (mg/mI)fl-MSP (mg/mI)PAP (mg/mI)Zna2-GP (units/mI)
2.5 (0.17-7.3)1.5 (0.21-5.0)0.94(0.17-3.0)1.2 (0.16-3.2)1.4 (0.16-5.3)
1.6 (0.14-5.8)1.0 (0.22-2.9)0.92(0.20-2.9)0.89(0.17-3.3)0.78(0-2.7)

494 Journal of Andrology . November/December 1995
Table 4. Spearman’s rank correlation coefficient for concentration of seminal plasma variables, NZS subgroup (n = 35, lower left) andOAZS subgroup (n = 27, upper right)
Fructose PCI PCI-PSA Zn PSA fl-MSP PAP Zna2-GP
Seminal vesicleFructose 1 0.15 0.01 -0.38 -0.37 -0.33 -0.40 -0.16
PCI 0.10 1 0.79*** 0.16 0.17 0.03 0.24 0.31PCI-PSA -0.12 0.61*** 1 0.11 0.14 0.03 0.07 0.48*
Prostate
Zn -0.28 -0.01 -0.06 1 0.89*** 0.73*** 0.70*** 0.42*PSA -0.21 -0.06 -0.15 0.77*** 1 0.67*** 0.80*** 0.47-MSP -0.09 -0.40 -0.28 0.53** 0.62*** 1 0.54 0.37PAP -0.20 -0.15 -0.17 0.78*** 0.79*** 0.67*** 1 0.34Zna2-GP -0.34 -0.05 0.24 0.57*** 0.60*** 0.25 0.52** 1
Asterisks Indicate degree of significance, obtained by z-test. ‘P < 0.05, < 0.01, P < 0.001.
and that of PCI, either in the NZS and OAZS subgroup.
However, the concentration of PCI manifested a signifi-
cant correlation to that of the PCI-PSA complex (P <
0.001), in contrast to the concentration of PSA, which did
not correlate to that of the PCI-PSA complex in either of
the two subgroups.
Ill. Accssoiy Sex Gland Function
We compared the two subgroups with regard to the se-
cretory function of the prostate using measurements of
Zn, PSA, PAP, 3-MSP, and Zncr2-GP. Measurements of
fructose and PCI were used to evaluate the function of
the seminal vesicles. The volume of each ejaculate was
used to calculate the total secreted amount of each secre-
tory component in the ejaculate. The intraindividual vari-
ation of the total amounts of the different semen param-
eters was tested in 13 men with an initial and a second
sample. There was no statistical difference (P = 0.3-0.9)
between the median values of each secretory component
in these two samples. The prostatic secretory function was
significantly decreased in the OAZS compared to the NZS
subgroup (Table 5). The most significant reduction was
in the secretion of Zna2-GP (P < 0.001), whereas the
differences between the subgroups were less significant for
the secretion of Zn, PSA, PAP (P < 0.01), and i9-MSP (P
<0.05). There was no significant decrease in the secretion
of fructose or PCI in the OAZS subgroup, indicating that
there was no significant reduction in secretory function
of the seminal vesicles. The amount of PCI-PSA complex
was slightly lower in the OAZS than in the NZS subgroup
(P < 0.05).
The secretory contribution from the prostate was com-
pared to that of the seminal vesicles by calculating the
molar PSA/fructose ratios. The median PSA/fructose ra-
tio was slightly lower in the OAZS than in the NZS sub-
group (0.002 vs. 0.003; P < 0.05), possibly reflecting an
altered balance between the prostatic and seminal vesicle-
derived secretory contributions in the the OAZS subgroup
(not shown).
IV. Decreased Secretory Function of theProstate as a Marker of Oligo-Asthenozoospermia
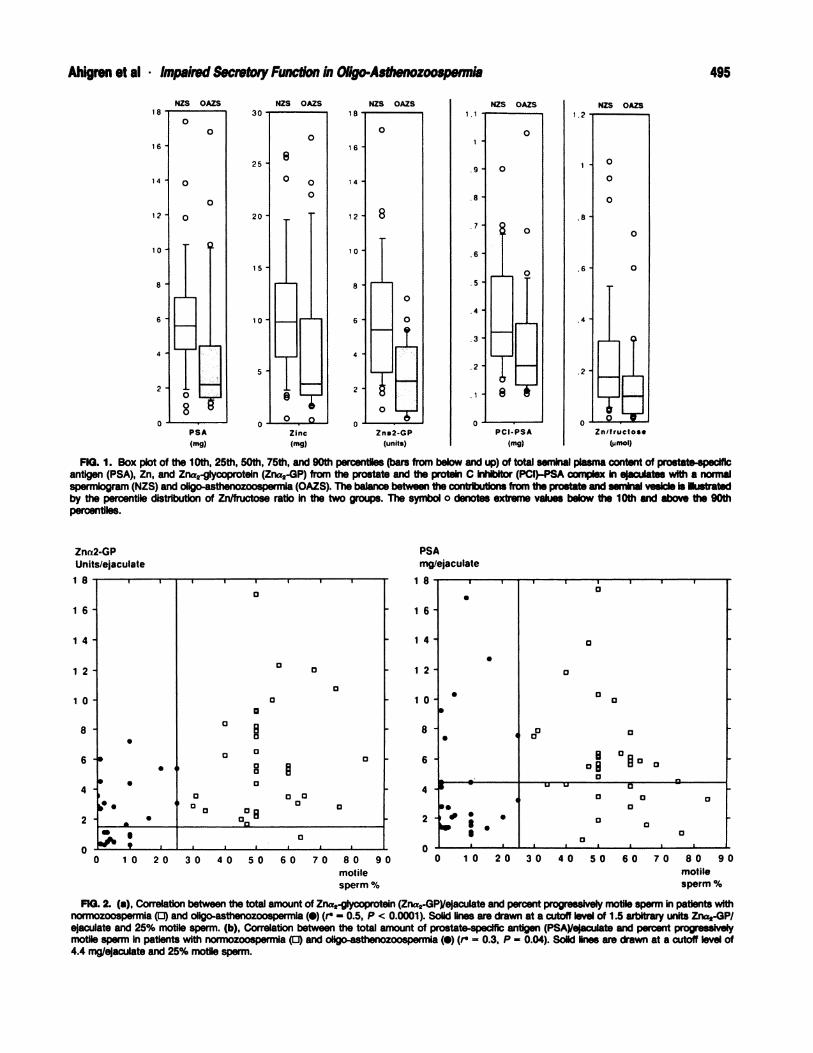
The ability of a decrease in the concentrations of a pro-
static secretory component to indicate oligo-astheno-
zoospermia was calculated for each of the prostate pro-
teins and Zn (Fig. 1). PSA proved to be the most infor-
mative variable, 75% of the men in the OAZS subgroup
having <4.4 mg PSA per ejaculate, whereas 75% of the
men in the NZS subgroup had >4.4 mg PSA per ejaculate.
There was also a significant correlation between per-
centage of progressively motile sperm and the ejaculate
content of PSA, Zn, PSA-PCI (P < 0.005), and Zna2-GP
(P < 0.0001) in the ejaculate, analyzing the results from
both the NZS and OAZS subgroup together. Most of the
normospermic men had >1.5 arbitrary units of Zna2-GP
(specificity 91%), whereas less than half of the men in the
OAZS subgroup had <1.5 arbitrary units of Zna2-GP per
ejaculate (sensitivity 42%) (Fig. 2a). At a cutoff value of
4.4 mg, the sensitivity of PSA was higher (74%) but the
Table 5. Total seminal plasma content of secretory products fromthe accessory sex glands in the two subgroups of patients
Oligo-asthenozoo-Normozoospermic spermic
Variables (n = 35) (n = 27) P valuet
Seminal vesicleFructose
(mol) 49 (28-110) 48 (19-161) 0.92 (NS)PCI (mg) 0.76(0.33-1.19) 0.50(0.31-1.47) 0.21 (NS)PCI-PSA (mg) 0.32(0.16-0.87) 0.20(0.11-0.51) 0.0112*
Prostate
Zn (JLmol) 9.8 (3.2-19.6) 3.8 (1.7-20.2) 0.0026**PSA(mg) 5.6 (1.9-10.3) 2.2 (1.3-10.1) 0.0032*.fl-MSP (mg) 4.4 (1.5-11.1) 2.2 (0.7-10.0) 0.0298’PAP (mg) 4.5 (1.5-11.1) 1.6 (0.9-11.9) 0.0045**Zfla2-GP 5.4 (2.2-10.7) 2.4 (0.3-5.4) 0.0001”
Data are presented as median values and 10th and 90th percentiles.t Data were obtained by Mann-Whitney U-test; the asterisk indicates
the degree of significance: P < 0.05, **p < 0.01, “‘P < 0.001.
11
.9
.8
.7
.6
5
18
16
14
12
10
8
6
4
2
.8
.2
0
.1
0
Zna2-GP
(units)
0
Ahigren et al - Impaired Secretoiy Function in Oligo-Asthenozoospermia 495
NZS OAZS NZS OAZS NZS OAZS NZS OAZS NZS OAZSIS 30 12
0
0 0
16
S25 o 1
14 o 0 o
00
12 0 20
0
Zinc PCI-PSA Zn!t,uctoss
1mg) (mg) (umol)
FIG. 1. Box plot of the 10th, 25th, 50th, 75th, and 90th percentIles (bars from below and up) of total seminal plasma content of prostate-specificantigen (PSA), Zn, and Zna2-glycoprotein (Zna2-GP) from the prostate and the protein C inhibitor (PCI)-PSA complex In eJaculates wIth a normalspermlogram (NZS) and ollgo-asthenozoospermla (OAZS). The balance between the contslbutlons from the prostate and seminal vesicle Is Illustratedby the percentile distribution of Zn/fructose ratio in the two groups. The symbol o denotes extreme values below the 10th and above the 90thpercentiles.
Zn2-GP PSAUnits/ejaculate mg/ejaculate
18 , 18
16 16
14 14
12 12
10 10
8 8
6 6
4 4
2 - 2
0
aS
a a
a0
.
a
.
a
0
a a
a
afi a0
a
0
aDS
a0
0
Doa
aa,.,
a
0 10 20 30 40 50 80 90motilesperm %
10
8
6
4
20
8PSA
(mgI
aa
a
aaaa
.IIIbI
aI I I I
U U 0
a a ai#{149} a‘. .
#{149}
#{149} a
a
a
60 70 80 90 0 10 20 30 40 50 60 70
motilesperm %
FIG. 2. (a), Correlation between the total amount of Zna2-glycoprotein (Zna2-GP)/ejaculate and percent progressively motile sperm in patients withnormozoospermia (0) and oligo-asthenozoospermia (#{149})(r = 0.5, P < 0.0001). SolId lines are drawn at a cutoff level of 1.5 arbitrary units Zna2-GP/elaculate and 25% motile sperm. (b), Correlation between the total amount of prostate-specific antigen (PSA)/ejaculate and percent progressivelymotile sperm In patients with normozoospermia (0) and oligo-asthenozoospermia (#{149})(r = 0.3, P = 0.04). SolId lines are drawn at a cutoff level of4.4 mg/eJaculate and 25% motile sperm.

496 Journal of Andrology November/December 1995
specificity was lower (66%) than that of Zna2-GP as a
marker of oligo-asthenozoospermia (Fig. 2b).
Although the ratio between the secretory contribution
from the prostate and that of the seminal vesicles (PSA-
fructose ratio) did not correlate significantly to progressive
motility (P = 0.06), of a total of seven samples in the
OAZS subgroup with a PSA-fructose ratio of <0.001, six
manifested <5% motile sperm (not shown).
Discussion
In this study we selected samples with asthenozoospermia
that in all cases were associated with oligozoospermia,
because this combination is far more common than pro-
gressive motility disturbances alone. The time of absti-
nence before sampling of the ejaculates varied between 2
and 10 days, with similar mean values in the NZS and
OAZS groups. Previous studies have demonstrated thatthe accessory sex glands replenish their secretions within2-3 days of an ejaculation and that little extra is added
to seminal volume with longer abstinence (Jouannet et
al, 1981).
Because liquefaction of the human coagulum takes place
within 20 minutes of ejaculation, the molecular changes
taking place immediately following ejaculation have to be
taken into consideration in analyzing the components of
seminal plasma. With time, up to 5% of the active form
of PSA in the seminal fluid will complex to the protease
inhibitor PCI (Christensson et al, 1994). The PSA anti-
bodies used in this study are essentially unaffected by these
reactions, because they react with PSA epitopes also ac-
cessible after complex formation has occurred with PCI.
Therefore, measured concentrations of PSA in seminal
fluid may reflect an essentially unbiased total concentra-
tion of PSA (Lilja et al, 1991; Christensson and Lilja
1994). The methods used to measure the secretory pro-
teins from the prostate in this study are unaffected by the
interval between ejaculation and the freezing of seminal
plasma at - 20#{176}C,except for the concentration of PSA-
PCI complexes, that increases during the liquefaction of
semen (Christensson and Lilja, 1994). The previously re-
ported degradation of the secretory proteins from the
prostate and the seminal vesicles by the aspartic protease
gastricin requires an acidic pH to convert progastricin to
the active enzyme and will not take place in samples
obtained by masturbation (Szecsi and Lilja, 1993). It is
not likely that intraindividual variation had any influence
on the results in the study because there was no statistical
difference between the median values of any of the com-
ponents analyzed in two separate samples from different
occasions from 13 men. The validity of our results is also
supported by the comparison of Zn concentrations in the
NZS group with a study on 456 normal men using the
same methodology (Marmar et al, 1980), in which the
mean concentration of Zn was 5.9 mmolJL in expressedprostatic fluid compared to a mean 2.7 mmol/L in whole
semen samples (median 2.5 mmol!L in the NZS group in
this study).The seminal fluid concentrations of Zn manifested good
correlation to those of PSA, i3-MSP, PAP, and Zna2-GP,
findings consistent with those previously obtained using
other techniques and tissue samples to detect the pro-
duction of these most abundant prostate-derived proteins
in seminal fluid (Frenette et al 1987; Lilja and Abra-
hamsson, 1988). By contrast, it is remarkable that the
concentrations of fructose and PCI did not correlate, de-
spite a common major secretory contribution from the
seminal vesicles (Mann and Lutwak-Mann, 1981; Laurell
et a!, 1992). However, the total secreted amount of fruc-tose correlated significantly to that of PCI, probably due
to stronger correlation between the volume of the ejacu-
late and fructose than that between ejaculate volume and
PCI. Neither the concentration of Zn nor that of any of
the secretory proteins from the prostate correlated with
that of fructose or PCI from the seminal vesicles, despite
the fact that both glands are under androgenic regulation
(deWinter et al, 1991).
The control group in our study was composed of men
having a semen analysis performed as a routine workup
in the investigation of infertile couples. It is known that
in >50% of the cases, the cause of infertility is found in
the female by tubal occlusion, and that sperm analysis
was performed before a planned in vitro fertilization (per-
sonal communication Dr. Rannevik). Moreover, the con-
centrations of fructose and Zn in the NZS subgroup were
similar to those found in a study of seminal plasma from
men who had recently become fathers (Cooper et al, 1991).
Secretion of Zn and of all the major prostate proteins
in seminal plasma was significantly lower in the OAZS
subgroup than in the NZS subgroup. Still, there were no
indications of insufficient androgen supply in the OAZS
subgroup compared to the NZS group; there was no dif-
ference in LH, testosterone concentrations, or in the tes-
tosterone/SHBG ratio, which reflects the concentration
of free testosterone. The lack of difference between the
NZS and OAZS subgroups in seminal vesicle contribution
may have been due to the seminal vesicle variables mea-
sured being insufficiently sensitive, or perhaps secretory
function of the seminal vesicles is unaffected in OAZS
men. The latter possibility derives support from the ab-
sence of significant difference in ejaculate volume between
the two subgroups, despite the decrease in secretory con-
tribution from the prostate in the OAZS subgroup. As in
the prostate, the secretory activity of the seminal vesicles
is also dependent upon testosterone after its conversion
to dihydrotestosterone (DHT). This suggests that there

Ahigren et al . Impaired Secretor,’ Function in Oligo-Asthenozoospermia 497
may be a nonandrogenic cause of the reduction in secre-
tory function of the prostate observed in this study. It is
noteworthy that a low Zna2-GP contribution to the ejac-
ulate most strongly correlated to poor sperm motility. The
function of this glycoprotein is unknown, but it is known
to be expressed in the testis and the epididymis as well
as in the prostate (Frenette et al, 1987). Because all the
asthenozoospermic samples also turned out to be oligo-
zoospermic, this may reflect defective function of both
the testis and the prostate, in contrast to the preserved
function of the seminal vesicles. A number of different
growth factors are involved in the regulation of sper-
matogenesis (Lamb, 1993). Lack of yet-unknown endo-
crine or paracrine factors affecting both the testicles and
the accessory sex gland function is another possible ex-
planation for the impaired secretory function of the pros-
tate in OAZS men.
There have been no reports of significant impairment
of the secretory contribution of Zn and the major prostate
proteins in men from infertile couples. An age-dependent
decrease in secretory contribution from the prostate has
been described (Rui et al, 1986). Also, data have been
published supporting an impaired secretory function of
the prostate secondary to prostatitis (Comhaire et al, 1989;Wolff et al, 1991) and varicoceles (Ando et al, 1990),
whereas in another study no difference in PSA or PAP
was found in men with epididymitis or chronic prostatitis
(v der Kammer et al, 1991). The incidence of varicoceles
and prostatitis among the men in this series is not known,
but of 17 bacterial cultures of ejaculates from the OAZS
subgroup, only 2 manifested significant bacterial growth.There have been several reports indicating the impor-
tance of seminal clot dissolution for sperm motility and
fertility. In an immediate postcoital test, sperm were found
in the cervical mucus within 90 seconds. In one case,
where liquefaction of the coagulum was absent, no sperm
were found in the cervix, and in another two cases with
high viscosity, no forward motility was seen (Sobrero and
MacLeod, 1962). In a recent study, separation of seminal
plasma from sperm after liquefaction reduced the pro-
gressive motility of sperm. Addition of seminal plasma
from normozoospermic and asthenozoospermic men
stimulated progressive sperm motility in both instances,
an effect also seen when serum was added. No difference
was seen after heat treatment, indicating the involvement
of a nonorganic motility factor. However, the prostate-
enriched first fraction of a split ejaculate seemed to be
superior in stimulating sperm motility (Magnus et al, 1991).
In conclusion, the present findings suggest the secretory
contribution by the prostate to be significantly reduced in
OAZS men as compared to NZS men. The importance
of this finding in relation to poor sperm count and mo-
tility, as well as indicators of impaired gonadal function,
requires further investigation.
AcknowledgmentsWe thank Mrs. Gun-Britt Enksson for perfonning biochemical analyses
of seminal plasma and Mrs. Mania Winitsky for performing the sperm
analysis. This work was supported by grants from the Swedish Medical
Research Council (project no. 7903), the Faculty of Medicine at LundUniversity, the Research Fund at Malmo University Hospital, the Mag-
nus Bergvall Foundation, the Crafoord Foundation, and the Fundacion
Fedenco S.A.
ReferencesAbrahamsson P-A, Andersson C, Bj#{246}rkT, Fernlund P, Lilja H, Murne
A, Weiber H. Radioimmunoassay of )9-microseminoprotein, a pro-static-secreted protein present in sera of both men and women. C/in
Chem 1989;35: 1497-1 503.
Ando S, Carpino A, Buffone M, Maggiolini M, Giacchetto C, Seidita F.Fructose, prostaticacid phosphatase and zinc levelsin the seminal
plasma of varicoceles.mt i Fertil 1990;35:249-252.
Christensson A, Laurell C-B, Lilja H. Enzymatic activity of prostate
specific antigen and its reactions with extracellular serine proteaseinhibitors. Eur J Biochem 1990;194:755-763.
Christensson A, Lilja H. Complex formation between protein C inhibitorand prostate specific antigen in vitro and in human semen. Eur J
Biochem 1994;220:45-53.
Comhaire F, Vermeulen L, Pieters 0. Study of the accuracy of physicaland biochemical markers in semen to detect infectious dysfunctionof the accessory sex glands. JAndrol 1989;l0:50-53.
CooperT 0, JockenhOvel F,Nieschlag E. Variations in semen parametersfrom fathers. Hum Reprod 199 l;6:859-866.
deWinter JAR, Trapman J, Vermey M, Mulder E, Zegers ND, V derKwast TH. Androgen receptor expression in human tissues: an im-
munohistochemical study. J Histochem Cytochem 1991 ;7:927-936.Ferniund P, Gershagen 5, Larsson I, Rannevik G. Radioimmunoassay
of human sex hormone binding globulin: improved radioiodination
procedure. Scand I C/in Lab Invest 1985;45:46 1-469.Frenette 0, Dub#{233}J Y, Lazure C, Paradis G, Chr#{233}tien M, Tremblay R.
The major 40-kDa glycoprotein in human prostatic fluid is identicalto Zna2-Glycoprotein. Prostate 1987; 11:257-270.
Jouannet P, Czyglik F, David G, Mayaux MJ, Spira A, Moscato ML,Schwartz D. Study of a group of 484 fertile men. Part I: distribution
of semen characteristics. mt j. Androl. 1981 ;4:440-449.Kvist U, Kjellberg S, Bjorndahl L, Soufir J-C, Aever S. Seminal fluid
from men with agenesis of the Wolffian ducts: zinc-binding propertiesand effects on sperm chromation properties. Int I Androl 1990;13:
245-252.Lamb DJ. Growth factors and testicular development. J Urol 1993;l50:
583-592.
Laurell C-B McKay E. Electroimmunoassay methods. Methods Enzymol
1981 ;73:339-369.Laurell M, Christensson A, Abrahamsson PA, Stenflo J, Lilja H. Protein
C inhibitor in human body fluids. J C/in Invest 1992;89: 1094-1101.Lee C, Keefer M, Thao ZW, Kroes R, Berg L, Liu X, Sensibar J. Dem-
onstration of the role of prostatic specific antigen in semen liquefe-cation by two-dimensional electrophoresis. I Androl 1989; 10:432-
438.
Lilja H, Abrahamsson PA. Three predominant proteins secreted by the
human prostate gland. Prostate l988;l2:29-38.Lilja H, Christensson A, Dahl#{233}nU, Matikainen M-T, Nilsson 0, Pet-
tersson K, Lvgren T. Prostate-specific antigen in serum occurs pre-
dominantly in complex withal -antichymotrypsin. C/in Chem 1991;37:1618-1625.
Lilja H, Laurell C-B. A kallikrein-like serine protease in prostatic fluid

498 Journal of Andrology . November/December 1995
cleaves the predominant seminal vesicle protein. I C/in Invest 1985;
76:1899-1903.
Lilja H, Laurell C-B, Jepsson J-0. Characterization of the predominantbasic protein in human seminal plasma, one cleavage product of themajor seminal vesicle protein. Scandf C/in Lab Invest 1 984;44:439-
446.Lilja H, Oldbring J, Rannevik 0, Laurel CB. Seminal vesicle- secreted
proteins and their reactions during gelation and liquefaction of human
semen. IC/in Invest 1987;80:281-285.
Magnus O, Brekke I, Abyholm T, Purvis K. Effects of seminal plasmafrom normal and asthenozoospermic men on the progressive motility
of washed human Sperm. mt J Andro/ 1991; 14:44-51.Mann T, Lutwak-Mann C. Ma/e Reproductive Function and Semen.
Berlin: Springer Verlag; 1981.Marmar JL, Katz 5, Praiss DE, DeBenedicts TJ. Values for zinc in whole
semen, fractions of split ejaculates and expressed prostatic fluid. Uro/
1980; 16:478-480.
McGee RS, Herr R. Human seminal vesicle-specific antigen is a substrate
for prostate specific antigen (or P30). Biol Reprod 1988;39:499-5 10.Parker MM, Humoller FL, Mahler DJ. Determination of copper and
zinc in biological material. C/in Chem 1967;13:40-48.
Rui H, Thomassen Y, Oldereid NB, Purvis K. Accessory sex gland func-
tion in normal young (20-25 years) and middle-aged (50-55 years)
men. JAndro/ 1986;7:93-99.
Sobrero AJ, MacLeod J. The immediate postcoital test. Ferti/Steri/ 1962;
13:184-189.
Szecsi PB, Lilja H. Gastricsin-mediated proteolytic degradation of hu-man seminal fluid proteins at pH levels found in the human vagina.JAndro/ l993;14:35l-358.
Thorell JI, Larson SM. Radioimmunoassay and Re/ated Techniques. St.Louis: Mosby; 1978.
v der Kammer H, Scheit KH, Weidner W, Cooper T G. The evaluation
of markers of prostatic function. Uro/ Res 1991; 19:343-347.
World Health Organization. WHO Laboratory Manual for the Exami-
nation of Human Semen and Sperm-Cervica/ Mucus Interactions.
Cambridge, UK: Cambridge University Press; 1987.Wolff H, Bezold G, Zebhauser M, Meurer M. Impact of clinical silent
inflammation on male genital tract organs as reflected by biochemical
markers in semen. JAndro/ 199l;12:331-334.
Wetterauer. Aktuelle dermatologie 2. Mannheim, Germany. Boehringer
Mannheim; 1976:239-248.
Copyright © 2022 FDOKUMEN




















