
Corporates Look to Digital Solutions as COVID Disrupts Trade ...
Upload
independentCategory
view
3download
0
www.e l sev i e r . com/ l oca te /eu roneu ro
Caffeine modulates P50 auditory sensory gating inhealthy subjects
Eduardo S. Ghisolfi a,b,*, Alice Schuch b, Ivo M. Strimitzer Jr.b, GustavoLuersen a, Fabıola F. Martins a, Fernanda L.P. Ramos a, Jefferson Becker c,d,Diogo R. Lara a
a Departamento de Ciencias Fisiologicas, Faculdade de Biociencias, PUCRS, Porto Alegre, Brazilb Departamento de Bioquımica, ICBS, UFRGS, Porto Alegre, Brazilc Servico de Neurologia, HCPA, Porto Alegre, Brazild Unidade de Neurofisiologia, ULBRA, Canoas, Brazil
Received 17 May 2005; received in revised form 5 September 2005; accepted 15 September 2005
0924-977X/$ - see front matter D 200doi:10.1016/j.euroneuro.2005.09.001
* Corresponding author. FaculdadeIpiranga, 6681, Predio 12 A, Caixa PoBrazil, 90619-900. Fax: +55 51 3320 36
E-mail address: [email protected] (
KEYWORDSSensory;Gating;Evoked potentials;Caffeine;Adenosine;Auditory;Schizophrenia
Abstract The P50 suppression paradigm is an index of sensory gating assumed to reflect aninhibitory process. Adenosine is a neuromodulator with mostly inhibitory activity that isreleased by physiological stimuli and can be blocked by non-selective adenosine receptorantagonists such as theophylline and caffeine. A previous study showed that a single dose oftheophylline decreased P50 suppression in healthy volunteers. Here we investigated the effectof caffeine (0, 100, 200 and 400 mg p.o.) on P50 sensory gating in 24 healthy volunteers (15habitual caffeine high-users and 9 low-users). The 200 and 400 mg doses reduced P50 gating,whereas 100 mg produced a non-significant effect. The effect of caffeine was independent ofgender and habitual caffeine intake. High caffeine users also showed baseline differences, withlower S2 amplitudes compared to low-users. These results reinforce the participation ofadenosine in the modulation of P50 sensory gating and suggest that caffeine ingestion should becontrolled for in the P50 sensory gating paradigm.D 2005 Elsevier B.V. and ECNP. All rights reserved.
5 Elsevier B.V. and ECNP. All right
de Biociencias, PUC-RS, Av.stal 1429, Porto Alegre, RS,12.E.S. Ghisolfi).
1. Introduction
Caffeine, the most commonly used psychoactive drug,exerts mild stimulant effects attributed to its antagonisticaction at adenosine receptors, particularly A1 and A2A(Fredholm et al., 1999). Effects of caffeine includeincreased alertness and reduced fatigue, improved sus-tained attention and mood, as well as anxiety in susceptible
European Neuropsychopharmacology (2006) 16, 204—210
s reserved.

Caffeine modulates P50 auditory sensory gating in healthy subjects 205
individuals (Smith, 2002). Habitual doses of caffeine rangebetween 60 and 120 mg per dose, which are most commonlytaken 2 to 6 times a day. In humans, the dose—responsepattern to acute caffeine is usually linear up to 300 mg, withflattening or even a negative effect at higher doses(Fredholm et al., 1999; Smith, 2002). Also, responses areconsiderably influenced by interindividual differences, suchas regular caffeine consumption or personality traits (Alseneet al., 2003).
The P50 auditory event-related potential has been usedas an index of sensory gating particularly studied inneuropsychiatric research (Freedman et al., 1983; Adler etal., 1998, Boutros et al., 1999). It is thought to reflect a pre-attentive and central inhibitory mechanism related to theability of filtering irrelevant sensory information. The P50wave is a small amplitude, positive wave occurring about 50ms after an auditory stimulus. In the P50 suppressionparadigm, when two stimuli are presented 500 ms apart,the amplitude of the second peak (S2), compared to the first(S1), is usually attenuated in healthy subjects, whereas inpatients with schizophrenia or acute mania this suppressionis impaired (Adler et al., 1998). The neurochemical basis ofthe P50 suppression is not yet completely understood, butcholinergic, GABAergic and monoaminergic systems havebeen proposed to modulate this phenomenon (Adler et al.,1998; Hershman et al., 1995; Light et al., 1999). We haverecently shown that a moderate dose of the non-selectiveadenosine receptor antagonist theophylline (6 mg/kg)induced a suppression deficit in P50 sensory gating of normalsubjects (Ghisolfi et al., 2002) who had abstained fromcaffeine at least one week prior to neurophysiologicassessment. The pattern produced was a combination ofincreased S2 and reduced S1 wave and both changes havebeen reported in schizophrenic patients.
In the present study we further investigated the effectsof xanthines in sensory gating in healthy subjects submittedto the P50 evaluation before and after oral administration of0, 100, 200 and 400 of caffeine. In contrast to our previousstudy, we included subjects who are regular caffeine userswithout a long washout as well as low-users.
2. Experimental procedures
This study was approved by the Institutional Review Board ofthe Hospital de Clınicas de Porto Alegre (HCPA), carried outin accordance with the Declaration of Helsinki and all theparticipants signed an informed consent form after com-plete explanation about the potential risks involved in thisprotocol as well as the purpose of this study in lay terms.
2.1. Subjects
Twenty-five healthy volunteers were recruited for this studyamong university students and employees (12 females and13 males, mean age of 25F1.7 years, all Caucasian). Theywere submitted to a semi-structured interview by a trainedpsychiatrist (E.S.G.). Exclusion criteria were DSM-IV in axis Idiagnosis (evaluated with MINI), clinical illness and anycurrent use of illicit drugs of abuse or medicines, except fororal contraceptives. Subjects with familiar history ofschizophrenia or other psychotic disorders in first degree
were also excluded. They were also assessed on their typicalpattern of caffeine intake similarly to the protocol bySchicatano and Blumenthal (1995) and were divided in agroup of regular users of at least moderate caffeine doses(at least 200 mg/day every day —named caffeine high-users—n =15) or a group of low caffeine use (less than 100 mg/dayand necessarily not every day —named caffeine low-users —n =10). In fact, most of the subjects labeled as caffeine low-users did not use caffeinated beverages or foods at all.
2.2. Pharmacological challenge
Volunteers were instructed to abstain from xanthine-con-taining drinks (coffee, teas and colas) for at least 6 h beforethe first recording, which is an interval of time that allowssignificant clearance of circulating caffeine (half-life be-tween 2.5 and 4.5 h) without being associated withwithdrawal symptoms, which usually start at least 12 h afterthe last dose (Fredholm et al., 1999). Because nicotine canaffect P50 suppression (Adler et al., 1998), only non-smokerswere included (23 were never smokers and 2 had not smokedfor more than 1 year).
Subjects were submitted to neurophysiologic recordingsessions on 4 separate occasions 6—8 days apart. All subjectscompleted the four recording sessions. All recording sessionswere performed between 4:00 and 8:00 p.m. The P50 event-related potential of subjects was assessed at baseline and15, 30, 60 and 90 min after treatment administration. Undersingle blind conditions, they were randomly assigned toreceive pills of identical appearance containing 0, 100, 200or 400 mg of caffeine taken with plain water.
As the study design consists of four planned consecutiveweekly P50 examinations that are randomized for dose, wedid not expect for the 12 female subjects any kind of non-balanced hormonal influence of the menstrual cycle whencomparing the effect of the dose on P50 sensory gating.Additionally, regarding their menstrual cycle at entrance inthe study, 4 started in the first week of their cycle, 2 in thesecond one, 3 in the third week and the remaining 3 in thelast week of the cycle. Moreover, all female subjects were inuse of low dosage oral contraceptives, providing minorhormonal variation along the month.
2.3. Electrophysiological recordings
The method for electrophysiological recordings was aspreviously described (Ghisolfi et al., 2002). In brief, subjectswere recorded seated, relaxed, and awake with eyes openand fixed on a distant target during the recording.Electroencephalographic activity was recorded from a diskelectrode affixed to the vertex (Cz) and referenced to eachmastoid, generating two separated waves, corresponding toboth derivations A1—Cz and A2—Cz. The positive electrodewas placed at Cz and Fz position was used as earth referenceelectrode of the system. Eletrooculogram (EOG) wasobtained to minimize the influence of possible muscularartifacts such as eye blinks, rejecting contaminated waves.Electroencephalogram (EEG) and EOG was provided usingthe 4-channel system Nihon—Kohden MEM-4104K for record-ing of evoked responses integrated with auditory stimulator.The experiment was performed in a specially designed room
E.S. Ghisolfi et al.206
provided of a Faraday’s cage in order to minimize externalelectromagnetic noise. The mean signal was registered intwo channels, one for each side of the cranium, andamplified 20,000 times with a bandpass filter between 1Hz and 10 kHz. EEG was collected during 1 s for each pairedstimulus presented. Trials were rejected if they containedartifacts as indicated by either an EEG or an EOG tension ofF50 AV within the 1000 ms that included both P50 waves,starting 100 ms before the first stimulus. The rejection ratewas typically less than 20% and the recording session lasted 5to 7 min.
No semi-automatic control was performed to excludepossible P30 influence over the trials. In fact, all grandaverages were manually analyzed for possible contaminantartifacts, non-identifiable P50 waves or great P30 influ-ences. Twelve (1.2%) grand averages were excluded from atotal number of 1000 collected (25 subjects on 4 occasions in5 measurements —1 pre-treatment and 4 post-treatment —each one generating two grand averages). Auditory stimuliwere presented in a conditioning—testing paradigm with aninterpair interval of 500 ms and interstimuli interval of 10 s.A 0.04 ms square wave pulse was amplified in the auditoryfrequencies (20—12,000 Hz) and delivered through ear-phones that produce a 2.5 ms sound with an intensity of60 dB sound pressure level above the auditory acuitythreshold, which was measured 15 min before the record-ings. Thirty non-rejected waves were added together to givea grand average signal. Two consecutive grand averagewaves were collected and used together for analysis. Themost positive peak between 40 and 90 ms after theconditioning stimulus (S1) was selected as the P50 finallatency and the wave amplitude was measured relative tothe previous negativity, determining the initial latency andthe first P50 wave. The test wave (S2) was determined usingthe corresponding peak between 500F10 ms away from S1latency and its amplitude also measured relative to theprevious negative peak. Test/conditioning ratios werecalculated by dividing the test P50 amplitude (S2) by theconditioning P50 amplitude (S1) and multiplied by 100 torepresent a percentage (P50=100 S2/S1). The data werecollected by an unblind researcher (E.S.G.) and analyzed byan independent trained ratter blind to treatment (J.B.).Once obtained the S1 and S2 amplitudes for each grand
0102030405060708090
100
-15 15 30 60 90time (minutes)
P50
Rat
io (%
)
placebo
100 mg
200 mg
400 mgtreatment
0102030405060708090
100110
plac
P50
Rat
io (
%)
A. P50 Ratio B. M
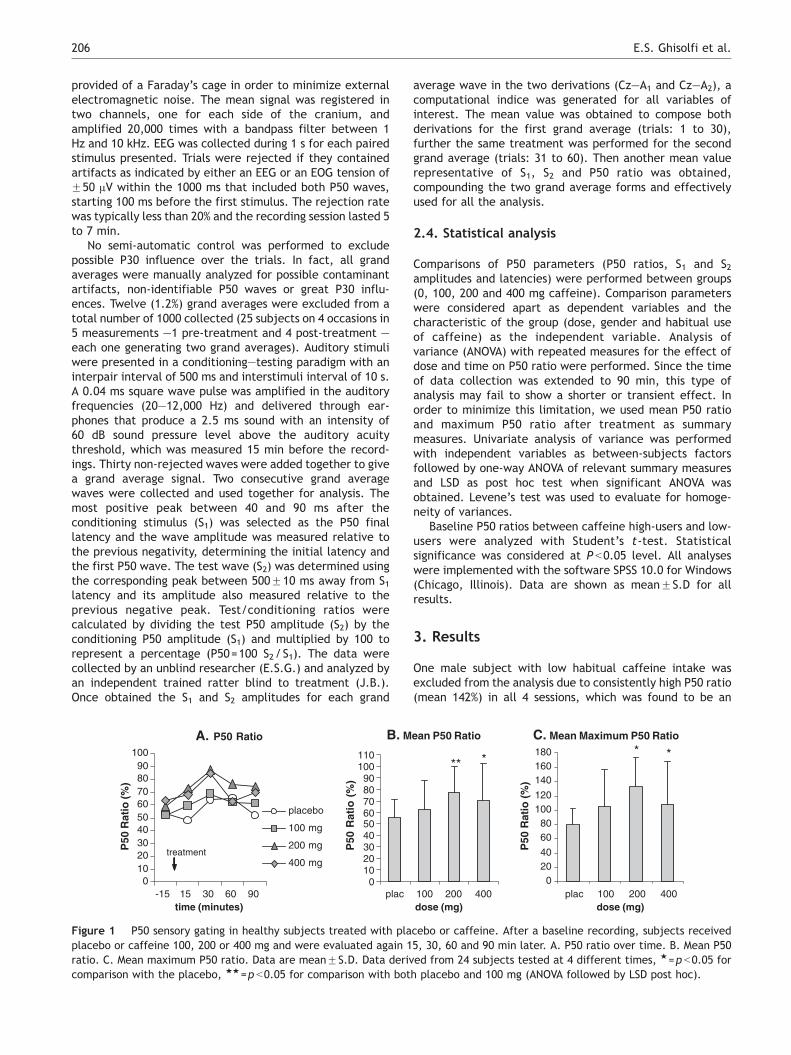
Figure 1 P50 sensory gating in healthy subjects treated with plaplacebo or caffeine 100, 200 or 400 mg and were evaluated again 1ratio. C. Mean maximum P50 ratio. Data are meanFS.D. Data derivcomparison with the placebo, 11=p b0.05 for comparison with bot
average wave in the two derivations (Cz—A1 and Cz—A2), acomputational indice was generated for all variables ofinterest. The mean value was obtained to compose bothderivations for the first grand average (trials: 1 to 30),further the same treatment was performed for the secondgrand average (trials: 31 to 60). Then another mean valuerepresentative of S1, S2 and P50 ratio was obtained,compounding the two grand average forms and effectivelyused for all the analysis.
2.4. Statistical analysis
Comparisons of P50 parameters (P50 ratios, S1 and S2amplitudes and latencies) were performed between groups(0, 100, 200 and 400 mg caffeine). Comparison parameterswere considered apart as dependent variables and thecharacteristic of the group (dose, gender and habitual useof caffeine) as the independent variable. Analysis ofvariance (ANOVA) with repeated measures for the effect ofdose and time on P50 ratio were performed. Since the timeof data collection was extended to 90 min, this type ofanalysis may fail to show a shorter or transient effect. Inorder to minimize this limitation, we used mean P50 ratioand maximum P50 ratio after treatment as summarymeasures. Univariate analysis of variance was performedwith independent variables as between-subjects factorsfollowed by one-way ANOVA of relevant summary measuresand LSD as post hoc test when significant ANOVA wasobtained. Levene’s test was used to evaluate for homoge-neity of variances.
Baseline P50 ratios between caffeine high-users and low-users were analyzed with Student’s t-test. Statisticalsignificance was considered at P b0.05 level. All analyseswere implemented with the software SPSS 10.0 for Windows(Chicago, Illinois). Data are shown as meanFS.D for allresults.
3. Results
One male subject with low habitual caffeine intake wasexcluded from the analysis due to consistently high P50 ratio(mean 142%) in all 4 sessions, which was found to be an
100 200 400dose (mg)
***
02040
6080
100120
140160180
plac 100 200 400dose (mg)
P50
Rat
io (
%)
* *ean P50 Ratio C. Mean Maximum P50 Ratio
cebo or caffeine. After a baseline recording, subjects received5, 30, 60 and 90 min later. A. P50 ratio over time. B. Mean P50ed from 24 subjects tested at 4 different times, 1=p b0.05 forh placebo and 100 mg (ANOVA followed by LSD post hoc).
0
1
2
3
4
5
6
-15 15 30 60 90
time
mic
rovo
lts placebo
100 mg
200 mg
400 mgtreatment
S1
S2
0
1
2
3
4
5
6
7
plac 100 200 400
dose (mg)
mic
rovo
lts
0
1
2
3
4
plac 100 200 400
dose (mg)
mic
rovo
lts
A. S1 and S2 Amplitudes B. Mean S1 Amplitudes C. Mean S2 Amplitudes
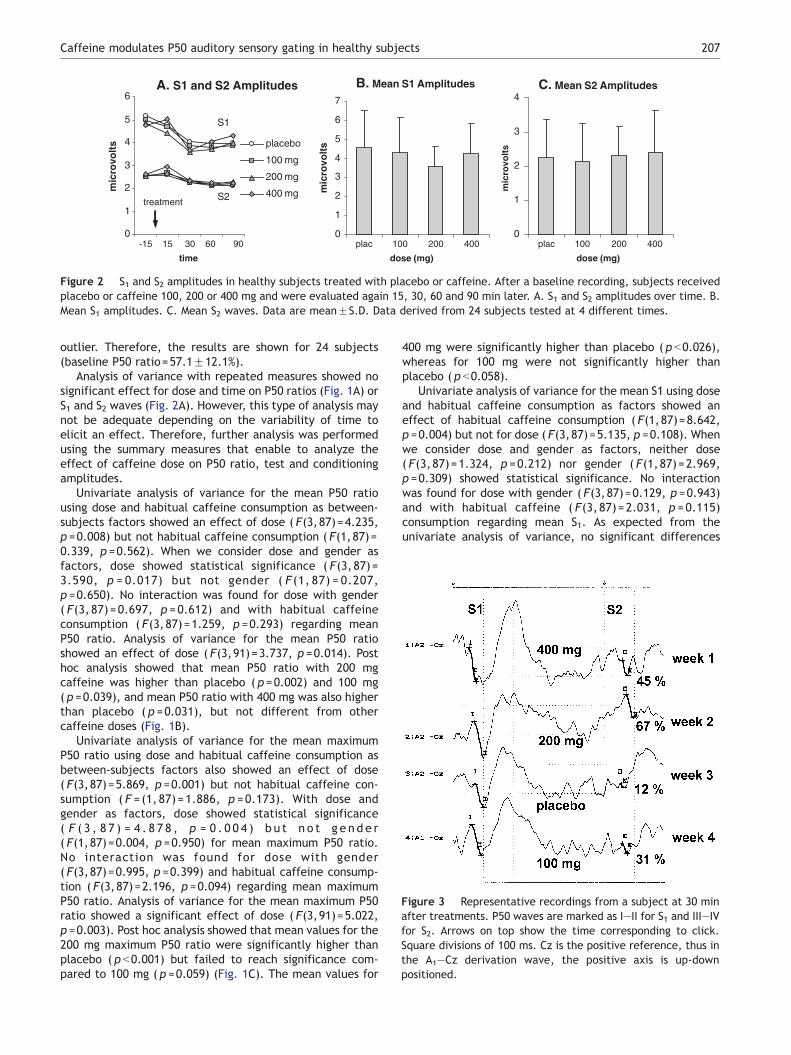
Figure 2 S1 and S2 amplitudes in healthy subjects treated with placebo or caffeine. After a baseline recording, subjects receivedplacebo or caffeine 100, 200 or 400 mg and were evaluated again 15, 30, 60 and 90 min later. A. S1 and S2 amplitudes over time. B.Mean S1 amplitudes. C. Mean S2 waves. Data are meanFS.D. Data derived from 24 subjects tested at 4 different times.
Figure 3 Representative recordings from a subject at 30 minafter treatments. P50 waves are marked as I—II for S1 and III—IVfor S2. Arrows on top show the time corresponding to click.Square divisions of 100 ms. Cz is the positive reference, thus inthe A1—Cz derivation wave, the positive axis is up-downpositioned.
Caffeine modulates P50 auditory sensory gating in healthy subjects 207
outlier. Therefore, the results are shown for 24 subjects(baseline P50 ratio=57.1F12.1%).
Analysis of variance with repeated measures showed nosignificant effect for dose and time on P50 ratios (Fig. 1A) orS1 and S2 waves (Fig. 2A). However, this type of analysis maynot be adequate depending on the variability of time toelicit an effect. Therefore, further analysis was performedusing the summary measures that enable to analyze theeffect of caffeine dose on P50 ratio, test and conditioningamplitudes.
Univariate analysis of variance for the mean P50 ratiousing dose and habitual caffeine consumption as between-subjects factors showed an effect of dose ( F(3,87)=4.235,p =0.008) but not habitual caffeine consumption ( F(1,87)=0.339, p =0.562). When we consider dose and gender asfactors, dose showed statistical significance ( F(3,87)=3.590, p = 0.017) but not gender ( F (1, 87) = 0.207,p =0.650). No interaction was found for dose with gender( F(3, 87)=0.697, p =0.612) and with habitual caffeineconsumption ( F(3,87)=1.259, p =0.293) regarding meanP50 ratio. Analysis of variance for the mean P50 ratioshowed an effect of dose ( F(3,91)=3.737, p =0.014). Posthoc analysis showed that mean P50 ratio with 200 mgcaffeine was higher than placebo ( p =0.002) and 100 mg( p =0.039), and mean P50 ratio with 400 mg was also higherthan placebo ( p =0.031), but not different from othercaffeine doses (Fig. 1B).
Univariate analysis of variance for the mean maximumP50 ratio using dose and habitual caffeine consumption asbetween-subjects factors also showed an effect of dose( F(3,87)=5.869, p =0.001) but not habitual caffeine con-sumption ( F = (1, 87)=1.886, p =0.173). With dose andgender as factors, dose showed statistical significance( F ( 3 , 8 7 ) = 4 . 8 7 8 , p = 0 . 0 0 4 ) b u t n o t g e n d e r( F(1,87)=0.004, p =0.950) for mean maximum P50 ratio.No interaction was found for dose with gender( F(3,87)=0.995, p =0.399) and habitual caffeine consump-tion ( F(3,87)=2.196, p =0.094) regarding mean maximumP50 ratio. Analysis of variance for the mean maximum P50ratio showed a significant effect of dose ( F(3,91)=5.022,p =0.003). Post hoc analysis showed that mean values for the200 mg maximum P50 ratio were significantly higher thanplacebo ( p b0.001) but failed to reach significance com-pared to 100 mg ( p =0.059) (Fig. 1C). The mean values for
400 mg were significantly higher than placebo ( p b0.026),whereas for 100 mg were not significantly higher thanplacebo ( p b0.058).
Univariate analysis of variance for the mean S1 using doseand habitual caffeine consumption as factors showed aneffect of habitual caffeine consumption ( F(1,87)=8.642,p =0.004) but not for dose ( F(3,87)=5.135, p =0.108). Whenwe consider dose and gender as factors, neither dose( F(3,87)=1.324, p =0.212) nor gender ( F(1,87)=2.969,p =0.309) showed statistical significance. No interactionwas found for dose with gender ( F(3,87)=0.129, p =0.943)and with habitual caffeine ( F(3, 87)=2.031, p =0.115)consumption regarding mean S1. As expected from theunivariate analysis of variance, no significant differences
0
10
20
30
40
50
60
70
80
90
100
high-user low-user high-user low-user
S2/
S1
(%)
0
1
2
3
4
5
6
mic
rovo
lts
*A. P50 Ratio B. Test Amplitude (S2)
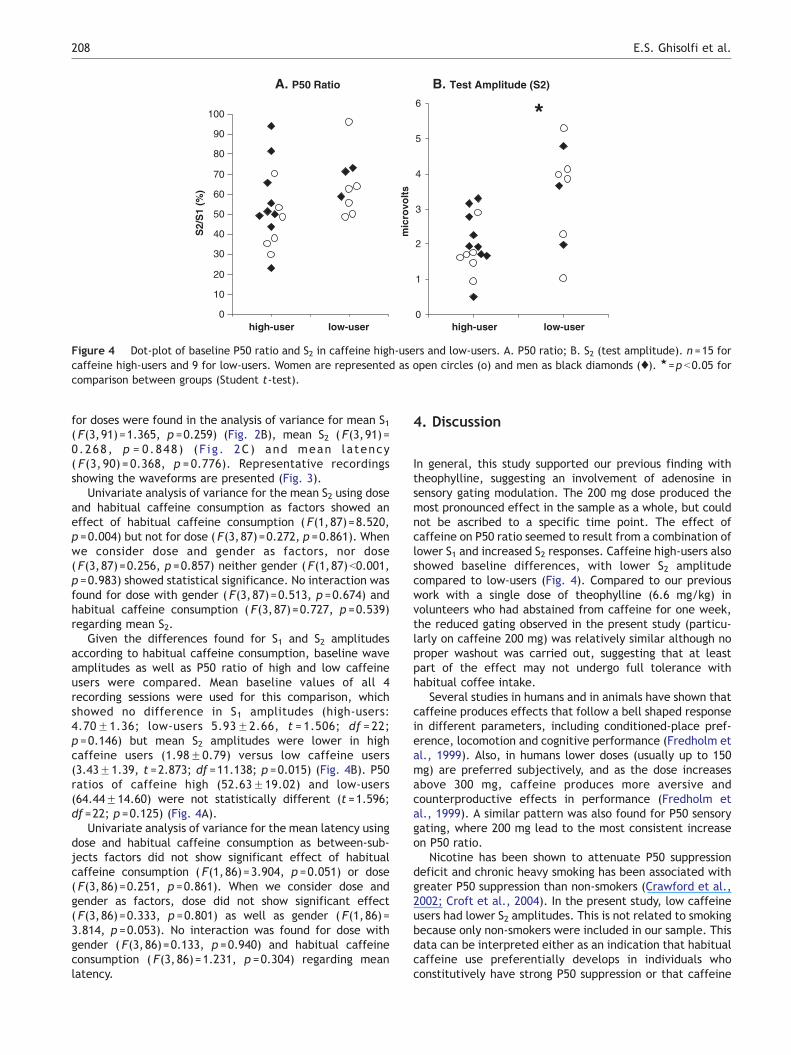
Figure 4 Dot-plot of baseline P50 ratio and S2 in caffeine high-users and low-users. A. P50 ratio; B. S2 (test amplitude). n =15 forcaffeine high-users and 9 for low-users. Women are represented as open circles (o) and men as black diamonds (x). 1=p b0.05 forcomparison between groups (Student t-test).
E.S. Ghisolfi et al.208
for doses were found in the analysis of variance for mean S1( F(3,91)=1.365, p =0.259) (Fig. 2B), mean S2 ( F(3,91)=0.268, p = 0.848) (F ig. 2C) and mean latency( F(3, 90) = 0.368, p =0.776). Representative recordingsshowing the waveforms are presented (Fig. 3).
Univariate analysis of variance for the mean S2 using doseand habitual caffeine consumption as factors showed aneffect of habitual caffeine consumption ( F(1,87)=8.520,p =0.004) but not for dose ( F(3,87)=0.272, p =0.861). Whenwe consider dose and gender as factors, nor dose( F(3,87)=0.256, p =0.857) neither gender ( F(1,87)b0.001,p =0.983) showed statistical significance. No interaction wasfound for dose with gender ( F(3,87)=0.513, p =0.674) andhabitual caffeine consumption ( F(3,87)=0.727, p =0.539)regarding mean S2.
Given the differences found for S1 and S2 amplitudesaccording to habitual caffeine consumption, baseline waveamplitudes as well as P50 ratio of high and low caffeineusers were compared. Mean baseline values of all 4recording sessions were used for this comparison, whichshowed no difference in S1 amplitudes (high-users:4.70F1.36; low-users 5.93F2.66, t = 1.506; df = 22;p =0.146) but mean S2 amplitudes were lower in highcaffeine users (1.98F0.79) versus low caffeine users(3.43F1.39, t =2.873; df =11.138; p =0.015) (Fig. 4B). P50ratios of caffeine high (52.63F19.02) and low-users(64.44F14.60) were not statistically different (t =1.596;df =22; p =0.125) (Fig. 4A).
Univariate analysis of variance for the mean latency usingdose and habitual caffeine consumption as between-sub-jects factors did not show significant effect of habitualcaffeine consumption ( F(1,86)=3.904, p =0.051) or dose( F(3,86)=0.251, p =0.861). When we consider dose andgender as factors, dose did not show significant effect( F(3,86)=0.333, p =0.801) as well as gender ( F(1,86)=3.814, p =0.053). No interaction was found for dose withgender ( F(3,86)=0.133, p =0.940) and habitual caffeineconsumption ( F(3,86)=1.231, p =0.304) regarding meanlatency.
4. Discussion
In general, this study supported our previous finding withtheophylline, suggesting an involvement of adenosine insensory gating modulation. The 200 mg dose produced themost pronounced effect in the sample as a whole, but couldnot be ascribed to a specific time point. The effect ofcaffeine on P50 ratio seemed to result from a combination oflower S1 and increased S2 responses. Caffeine high-users alsoshowed baseline differences, with lower S2 amplitudecompared to low-users (Fig. 4). Compared to our previouswork with a single dose of theophylline (6.6 mg/kg) involunteers who had abstained from caffeine for one week,the reduced gating observed in the present study (particu-larly on caffeine 200 mg) was relatively similar although noproper washout was carried out, suggesting that at leastpart of the effect may not undergo full tolerance withhabitual coffee intake.
Several studies in humans and in animals have shown thatcaffeine produces effects that follow a bell shaped responsein different parameters, including conditioned-place pref-erence, locomotion and cognitive performance (Fredholm etal., 1999). Also, in humans lower doses (usually up to 150mg) are preferred subjectively, and as the dose increasesabove 300 mg, caffeine produces more aversive andcounterproductive effects in performance (Fredholm etal., 1999). A similar pattern was also found for P50 sensorygating, where 200 mg lead to the most consistent increaseon P50 ratio.
Nicotine has been shown to attenuate P50 suppressiondeficit and chronic heavy smoking has been associated withgreater P50 suppression than non-smokers (Crawford et al.,2002; Croft et al., 2004). In the present study, low caffeineusers had lower S2 amplitudes. This is not related to smokingbecause only non-smokers were included in our sample. Thisdata can be interpreted either as an indication that habitualcaffeine use preferentially develops in individuals whoconstitutively have strong P50 suppression or that caffeine

Caffeine modulates P50 auditory sensory gating in healthy subjects 209
exerts a chronic effect on neural mechanisms involved inP50 sensory gating, e.g. the adenosine system, and after afew hours abstaining from caffeine this inhibitory systemmay be particularly active. Further studies with largersamples are required to confirm and clarify this issue. Also,similarly to nicotine, caffeine intake prior to the recordingsession should be considered as a potential bias in P50studies.
The neuromodulator adenosine exerts potent inhibitoryinfluence on synaptic activity in the CNS (Brundege andDunwiddie, 1997) mostly by adenosine A1 receptors, whichare at high levels in the hippocampus, cerebral cortex,thalamus and cerebellum (Ralevic and Burnstock, 1998). Byacting on presynaptic A1 receptors, adenosine inhibits therelease of several neurotransmitters (including glutamateand dopamine) and activation of post-synaptic A1 receptorsproduces neuronal hyperpolarization (Brundege and Dun-widdie, 1997). In contrast, A2A receptors are mostlyexpressed in dopamine-rich regions, co-localized with D2
receptors (Svenningsson et al., 1999). Activation of A2Areceptors reduces the affinity of D2 receptors for dopamine(Ferre et al., 1991; Ferre, 1997) and at least part of thestimulating effect of caffeine is due to inhibition of A2Areceptors (Fredholm et al., 1999). Both A1 and A2A receptorspotentially modulate P50 sensory gating, but A1 effectsdesensitize after chronic ingestion of caffeine (Quarta etal., 2004; Karcz-Kubicha et al., 2003), which may be relatedto the differences in baseline P50 ratio and S2 amplitude inhigh and low-users. Since conditioning (S1) response seemsto be more regulated by dopamine than S2, the effect ofcaffeine on S1 may be related to A2A receptor blockade. Incontrast, as A1 mediated adenosine actions are regarded asreactive or retaliatory and are related to rapid and transientinhibition of neurotransmitter release and action, test (S2)response is possibly more related to this receptor subtype.Studies in mice lacking the A2A receptor reinforce the role ofthis receptor in sensory gating (Wang et al., 2003; Nagel etal., 2003).Of particular interest regarding the P50 suppres-sion paradigm is the work by Mitchell et al. (1993), whoshowed that when two independent excitatory pathways toCA1 pyramidal neurons are used to evoke field excitatorypost-synaptic potential (EPSP), prior activation of onepathway at a physiological level inhibits the EPSP in theother pathway when evoked immediately after. This activ-ity-dependent inhibition lasted ~750 ms, peaking at 250 ms,which is a quite similar time-course compared to the P50suppression paradigm regarding the interstimuli (test andconditioning) intervals (Adler et al., 1998). Importantly, thisinhibition was mediated by adenosine, since it was blockedby theophylline and the A1 receptor antagonist CPT, andpotentiated by drugs that increase adenosine extracellularavailability by inhibiting adenosine uptake and degradation(Mitchell et al., 1993). This hypothesis can be clarified withselective adenosine ligands in animal models of P50 gating.
We have recently proposed that a deficit of adenosinergicactivity could contribute to the pathophysiology of schizo-phrenia and underlie its sensory gating deficits (Lara andSouza, 2000; Lara, 2002; Ghisolfi et al., 2002). Earlyneurodevelopment events such as hypoxia lead to excessiveadenosine release inducing neurotoxicity via activation of A1receptors (Turner et al., 2003). Excessive adenosine A1receptor activation during early development produces a
reduction in the number of these receptors afterwards(Turner et al., 2002). These events may lead to an adenosineinhibitory deficit that later emerges as reduced control ofdopamine activity and increased vulnerability to excitotoxicglutamate action in the mature brain. In this model,adenosine receptor antagonists, such as caffeine andtheophylline, are pharmacological models of adenosinehypofunction. If correct, pharmacological treatments en-hancing adenosine activity, especially through A1 receptors,could be effective for symptom and course control, as wellas for sensory gating deficits. In line with this proposal,clinical improvement of schizophrenic patients with add-ondipyridamole, an inhibitor of adenosine uptake (Akhondza-deh et al., 2000) and allopurinol, which inhibits degradationof purines (Brunstein et al., 2005; Akhondzadeh et al.,2005), has been shown in double-blind trials. However,allopurinol failed to correct the P50 suppression deficit inthese patients and the contribution of adenosine for sensorygating alterations has not been shown in schizophrenicpatients, only in healthy controls and animal models.
In summary, P50 sensory gating was reduced by relativelylow doses of caffeine and high caffeine users showed lowerbaseline S2 amplitude. These results suggest that habitualcaffeine intake and ingestion prior to evoked potentialrecordings should be taken into account for the P50suppression paradigm.
Acknowledgments
This research line has been supported by PRONEX(#41960904-366/96) and FAPERGS.
References
Adler, L.E., Olincy, A., Waldo, M., Harris, J.G., Griffith, J., Stevens,K., Flach, K., Nagamoto, H., Bickford, P., Leonard, S., Freed-man, R., 1998. Schizophrenia, sensory gating, and nicotinicreceptors. Schizophr. Bull. 24, 189–202.
Akhondzadeh, S., Shasavand, E., Jamilian, H., Shabestari, O.,Kamalipour, A., 2000. Dipyridamole in the treatment of schizo-phrenia: adenosine—dopamine receptor interactions. J. Clin.Pharm. Ther. 25, 131–137.
Akhondzadeh, S., Safarcherati, A., Amini, H., 2005. Beneficialantipsychotic effects of allopurinol as add-on therapy forschizophrenia: a double blind, randomized and placebo con-trolled trial. Prog. Neuro-psychopharmacol. Biol. Psychiatry 29,253–259.
Alsene, K., Deckert, J., Sand, P., de Wit, H., 2003. Associationbetween A2a receptor gene polymorphisms and caffeine-inducedanxiety. Neuropsychopharmacology 28, 1694–1702.
Boutros, N.N., Belger, A., Campbell, D., D’Souza, C., Krystal, J.,1999. Comparison of four components of sensory gating inschizophrenia and normal subjects: a preliminary report.Psychiatry Res. 88, 119–130.
Brundege, J.M., Dunwiddie, T.V., 1997. Role of adenosine as amodulator of synaptic activity in the central nervous system.Adv. Pharmacol. 39, 353–391.
Brunstein, M.G., Ghisolfi, E.S., Ramos, F.L.P., Lara, D.R., 2005.Clinical trial of allopurinol adjuvant therapy for moderatelyrefractory schizophrenia. J. Clin. Psychiatry 66, 213–219.
Crawford, H.J., McClain-Furmanski, D., Castagnoli, N. Jr., Castag-noli, K., 2002. Enhancement of auditory sensory gating and

E.S. Ghisolfi et al.210
stimulus-bound gamma band (40 Hz) oscillations in heavytobacco smokers. Neurosci. Lett. 317, 151–155.
Croft, R.J., Dimoska, A., Gonsalvez, C.J., Clarke, A.R., 2004.Suppression of P50 evoked potential component, schizotypalbeliefs and smoking. Psychiatry Res. 128, 53–62.
Ferre, S., 1997. Adenosine—dopamine interactions in the ventralstriatum: implications for the treatment of schizophrenia.Psychopharmacology 133, 107–120.
Ferre, S., Euler, G.V., Johansson, B., Fredholm, B., Fuxe, K., 1991.Stimulation of high affinity adenosine A2 receptors decreases theaffinity of dopamine D2 receptors in rat striatal membranes.Proc. Natl. Acad. Sci. U. S. A. 88, 7238–7241.
Fredholm, B.B., Batting, K., Holmen, J., Nehlig, A., Zvartau, E.,1999. Actions of caffeine in the brain with special reference tofactors that contribute to its widespread use. Pharmacol. Rev.51, 83–153.
Freedman, R., Adler, L.E., Waldo, M.C., Pachtman, E., Franks, R.D.,1983. Neurophysiological evidence for a defect in inhibitorypathways in schizophrenia: comparison of medicated and drug-free patients. Biol. Psychiatry 18, 537–551.
Ghisolfi, E.S., Prokopiuk, A.S., Becker, J., Ehlers, J.A., Belmonte-de-Abreu, P., Souza, D.O., Lara, D.R., 2002. The adenosineantagonist theophylline impairs p50 auditory sensory gating innormal subjects. Neuropsychopharmacology 27, 629–637.
Hershman, K.M., Freedman, R., Bickford, P.C., 1995. GABABantagonists diminish the inhibitory gating of auditory responsein the rat hippocampus. Neurosci. Lett. 190, 133–136.
Karcz-Kubicha, M., Antoniou, K., Terasmaa, A., Quarta, D., Solinas,M., Justinova, Z., Pezzola, A., Reggio, R., Muller, C.E., Fuxe, K.,Goldberg, S.R., Popoli, P., Ferre, S., 2003. Involvement ofadenosine A1 and A2A receptors in the motor effects of caffeineafter its acute and chronic administration. Neuropsychopharma-cology 28, 1281–1291.
Lara, D.R., 2002. Inhibitory deficit in schizophrenia is not neces-sarily a GABAergic deficit. Cell. Mol. Neurobiol. 22, 239–247.
Lara, D.R., Souza, D.O., 2000. Schizophrenia: a purinergic hypoth-esis. Med. Hypotheses 54, 157–166.
Light, G.A., Malaspina, D., Geyer, M.A., Luber, B.M., Coleman, E.A.,Sackeim, H.A., Braff, D.L., 1999. Amphetamine disrupts P50suppression in normal subjects. Biol. Psychiatry 46, 990–996.
Mitchell, J.B., Lupica, C.R., Dunwiddie, T.V., 1993. Activity-dependent release of endogenous adenosine modulates synapticresponses in the rat hippocampus. J. Neurosci. 13, 3439–3447.
Nagel, J., Schladebach, H., Koch, M., Schwienbacher, I., Muller,C.E., Hauber, W., 2003. Effects of an adenosine A2A receptorblockade in the nucleus accumbens on locomotion, feeding, andprepulse inhibition in rats. Synapse 49, 279–286.
Quarta, D., Ferre, S., Solinas, M., You, Z.B., Hockemeyer, J., Popoli,P., Goldberg, S.R., 2004. Opposite modulatory roles for adeno-sine A1 and A2A receptors on glutamate and dopamine release inthe shell of the nucleus accumbens. Effects of chronic caffeineexposure. J. Neurochem. 88, 1151–1158.
Ralevic, V., Burnstock, G., 1998. Receptors for purines andpyrimidines. Pharmacol. Rev. 50, 413–492.
Schicatano, E.J., Blumenthal, T.D., 1995. The effects of differentdoses of caffeine on habituation of the human acoustic startlereflex. Pharmacol. Biochem. Behav. 52, 231–236.
Smith, A., 2002. Effects of caffeine on human behavior. Food Chem.Toxicol. 40, 1243–1255.
Svenningsson, P., Le Moine, C., Fisone, G., Fredholm, B.B., 1999.Distribution, biochemistry and function of striatal adenosine A2Areceptors. Prog. Neurobiol. 59, 355–396.
Turner, C.P., Yan, H., Schwart, M., Othman, T., Rivkees, S.A., 2002.A1 adenosine receptor activation induces ventriculomegaly andwhite matter loss. NeuroReport 13, 1199–1204.
Turner, C.P., Seli, M., Ment, L., Stewart, W., Yan, H., Johansson, B.,Fredholm, B.B., Blackburn, M., Rivkees, S.A., 2003. A1 adeno-sine receptors mediate hypoxia-induced ventriculomegaly. Proc.Natl. Acad. Sci. U. S. A. 100, 11718–11722.
Wang, J.H., Short, J., Ledent, C., Lawrence, A.J., van den Buuse,M., 2003. Reduced startle habituation and prepulse inhibition inmice lacking the adenosine A2A receptor. Behav. Brain Res. 143,201–207.
Copyright © 2022 FDOKUMEN





![[A brief "update" on renal effects of caffeine.]](https://static.fdokumen.com/doc/165x107/630bdf00dffd330585081b05/a-brief-update-on-renal-effects-of-caffeine.jpg)














