
Growth hormone in the male reproductive tract of the chicken: heterogeneity and changes during...
13
Growth hormone in the male reproductive tract of the chicken: heterogeneity and changes during ontogeny and maturation M. Luna, a L. Huerta, a L. Berumen, a H. Mart ınez-Coria, a S. Harvey, b and C. Ar amburo a, * a Department of Cellular and Molecular Neurobiology, Institute of Neurobiology, Campus UNAM-UAQ Juriquilla, Universidad Nacional Aut onoma de M exico, A.P. 1-1141, Quer etaro, Qro. 76001, M exico b Department of Physiology and Perinatal Research Center, University of Alberta, Edmonton, Alta., Canada T6G 2H7 Received 12 November 2003; revised 10 February 2004; accepted 11 February 2004 Abstract Growth hormone (GH) gene expression is not confined to pituitary somatotrophs and occurs in many extrapituitary tissues. In this study, we describe the presence of GH moieties in the chicken testis. GH-immunoreactivity (GH-IR), determined by ELISA, was found in the testis of immature and mature chickens, but at concentrations <1% of those in the pituitary gland. The immu- noassayable GH concentration in the testis was unchanged between 4 and 66 weeks of age, and approximately 10-fold higher than that at 1-week of age and 25-fold higher than that in 1-day-old chicks and perinatal (embryonic day 18) embryos. This immuno- reactivity was associated with several proteins of different molecular size, as in the pituitary gland, when analyzed by SDS–PAGE under reducing conditions. However, while most of the GH-IR in the pituitary (40 and 15%, respectively) is associated with monomer (26 kDa) or dimer (52 kDa) GH moieties GH-IR in the testis is primarily (30–50%) associated with a 17 kDa moiety. GH bands between 32 and 45 kDa are also relatively more abundant in the testis than in the pituitary. During ontogeny the relative abundance of a 14 kDa GH and 40 kDa GH moieties in the testis significantly declined, whereas the relative abundance of the 17 and 45 kDa moieties increased with advancing age. In adult birds, GH-IR was widespread and intense in the seminiferous tubules. Although the GH-IR was not present in the basal compartment of Sertoli cells, nor in spermatogonia and primary spermatocytes, it was abundantly present in secondary spermatocytes and spermatids in the luminal compartments of the tubules as well as in some surrounding myocytes and interstitial cells. In summary, immunoreactive GH moieties are present in the chicken testis but at concentrations far less than in the pituitary. Age-related changes in the relative abundance of testicular GH variants may be related to local (autocrine/paracrine) actions of testicular GH. The localization of GH in spermatocytes and spermatids suggests hitherto unsuspected roles in gamete development. Ó 2004 Elsevier Inc. All rights reserved. Keywords: Chicken growth hormone; Testis; GH heterogeneity; Testicular GH 1. Introduction Growth hormone (GH) is primarily produced in pi- tuitary somatotrophs, although GH and GH mRNA has been found in many extrapituitary tissues (e.g., in the neural system—Yoshizato et al., 1998; Harvey et al., 2001; immune system—Hull et al., 1996, 2001; Recher et al., 2001; integumentary system—Palmetshofer et al., 1995; Slominski et al., 2000; skeletal system—Harvey et al., 2000; respiratory system—Allen et al., 2000; ali- mentary system—Tresguerres et al., 1999; Kyle et al., 1981; cardiovascular system—Costa et al., 1993; Wu et al., 1996; Recher et al., 2001; orthodontic tissues— Zhang et al., 1997; and in ocular tissues—Baudet et al., 2003). GH moieties and genes have also been detected in some reproductive tissues. Indeed, it is now well estab- lished that GH gene expression occurs extensively in the placenta (Hu et al., 1999). At least two GH genes are expressed in the human placenta, the pituitary hGH-N gene and a placental variant (hGH-V), which can be differentially spliced to produce three proteins (hGH-V 1 , hGH-V 2 , and hGH-V 3 ) (Boguszewski et al., 1998). The hGH-N and hGH-V genes have also been shown to be present in the human ovary (Schw€ arzler et al., 1997) and * Corresponding author. Fax: +52-442-238-1004. E-mail address: [email protected] (C. Ar amburo). 0016-6480/$ - see front matter Ó 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2004.02.005 www.elsevier.com/locate/ygcen General and Comparative Endocrinology 137 (2004) 37–49 GENERAL AND COMPARATIVE ENDOCRINOLOGY
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Growth hormone in the male reproductive tract of the chicken: heterogeneity and changes during...

GENERAL AND COMPARATIVE
ENDOCRINOLOGY
www.elsevier.com/locate/ygcen
General and Comparative Endocrinology 137 (2004) 37–49
Growth hormone in the male reproductive tract of thechicken: heterogeneity and changes during ontogeny and maturation
M. Luna,a L. Huerta,a L. Berumen,a H. Mart�ınez-Coria,a S. Harvey,b and C. Ar�amburoa,*
a Department of Cellular and Molecular Neurobiology, Institute of Neurobiology, Campus UNAM-UAQ Juriquilla,
Universidad Nacional Aut�onoma de M�exico, A.P. 1-1141, Quer�etaro, Qro. 76001, M�exicob Department of Physiology and Perinatal Research Center, University of Alberta, Edmonton, Alta., Canada T6G 2H7
Received 12 November 2003; revised 10 February 2004; accepted 11 February 2004
Abstract
Growth hormone (GH) gene expression is not confined to pituitary somatotrophs and occurs in many extrapituitary tissues. In
this study, we describe the presence of GH moieties in the chicken testis. GH-immunoreactivity (GH-IR), determined by ELISA,
was found in the testis of immature and mature chickens, but at concentrations <1% of those in the pituitary gland. The immu-
noassayable GH concentration in the testis was unchanged between 4 and 66 weeks of age, and approximately 10-fold higher than
that at 1-week of age and 25-fold higher than that in 1-day-old chicks and perinatal (embryonic day 18) embryos. This immuno-
reactivity was associated with several proteins of different molecular size, as in the pituitary gland, when analyzed by SDS–PAGE
under reducing conditions. However, while most of the GH-IR in the pituitary (�40 and 15%, respectively) is associated with
monomer (26 kDa) or dimer (52 kDa) GH moieties GH-IR in the testis is primarily (30–50%) associated with a 17 kDa moiety. GH
bands between 32 and 45 kDa are also relatively more abundant in the testis than in the pituitary. During ontogeny the relative
abundance of a 14 kDa GH and 40 kDa GH moieties in the testis significantly declined, whereas the relative abundance of the 17 and
45 kDa moieties increased with advancing age. In adult birds, GH-IR was widespread and intense in the seminiferous tubules.
Although the GH-IR was not present in the basal compartment of Sertoli cells, nor in spermatogonia and primary spermatocytes, it
was abundantly present in secondary spermatocytes and spermatids in the luminal compartments of the tubules as well as in some
surrounding myocytes and interstitial cells. In summary, immunoreactive GH moieties are present in the chicken testis but at
concentrations far less than in the pituitary. Age-related changes in the relative abundance of testicular GH variants may be related
to local (autocrine/paracrine) actions of testicular GH. The localization of GH in spermatocytes and spermatids suggests hitherto
unsuspected roles in gamete development.
� 2004 Elsevier Inc. All rights reserved.
Keywords: Chicken growth hormone; Testis; GH heterogeneity; Testicular GH
1. Introduction
Growth hormone (GH) is primarily produced in pi-
tuitary somatotrophs, although GH and GH mRNA
has been found in many extrapituitary tissues (e.g., in
the neural system—Yoshizato et al., 1998; Harvey et al.,2001; immune system—Hull et al., 1996, 2001; Recher
et al., 2001; integumentary system—Palmetshofer et al.,
1995; Slominski et al., 2000; skeletal system—Harvey
et al., 2000; respiratory system—Allen et al., 2000; ali-
* Corresponding author. Fax: +52-442-238-1004.
E-mail address: [email protected] (C. Ar�amburo).
0016-6480/$ - see front matter � 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.ygcen.2004.02.005
mentary system—Tresguerres et al., 1999; Kyle et al.,
1981; cardiovascular system—Costa et al., 1993; Wu
et al., 1996; Recher et al., 2001; orthodontic tissues—
Zhang et al., 1997; and in ocular tissues—Baudet et al.,
2003). GH moieties and genes have also been detected in
some reproductive tissues. Indeed, it is now well estab-lished that GH gene expression occurs extensively in the
placenta (Hu et al., 1999). At least two GH genes are
expressed in the human placenta, the pituitary hGH-N
gene and a placental variant (hGH-V), which can be
differentially spliced to produce three proteins (hGH-V1,
hGH-V2, and hGH-V3) (Boguszewski et al., 1998). The
hGH-N and hGH-V genes have also been shown to be
present in the human ovary (Schw€arzler et al., 1997) and

38 M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49
human testis, in which the hGH-V gene is preferentiallytranscribed (Berger et al., 1999; Untergasser et al., 1997,
1998). In the chicken it has been recently described that
GH gene expression can occur in the male reproductive
tract. The presence of GH mRNA in the testis of adult
chickens was assessed by RT-PCR and sequencing of
the testicular GH cDNA showed it had >99.5% ho-
mology with pituitary GH cDNA (Harvey et al., 2002).
It is, however, not known if these genes are translated inreproductive tissues, although GH has been found by
immunocytochemistry in the cells of the Wolffian duct in
fetal mice (Nguyen et al., 1996) and embryonic chickens
(Harvey et al., 2000; Wang, 1989). The possible presence
of GH in testicular tissue has therefore been further
investigated in domestic fowl, during ontogeny and
sexual maturation. Moreover, as pituitary GH is com-
prised of a family of heterogeneous molecular variants(Ar�amburo et al., 1989a, 1990, 2000, 2001a), the possi-
bility that testicular GH may also be heterogenous has
been examined.
2. Materials and methods
2.1. Tissues
Broiler fowl testes were collected during embryogen-
esis (18 embryonic day, ED), after hatch (day 1), and
before (weeks 4–9) and after (15 weeks of age) puberty
and after sexual maturation (at 36 and 66 weeks of age).
The tissues were immediately frozen on dry ice and
stored at )70 �C until used for assay. For comparison,
vas deferens, liver, and pituitary tissues were also col-lected from some of the birds.
2.2. Tissue extracts
Frozen tissues were homogenized (1 g/10ml) in an
EDTA-free protease inhibitor cocktail (Mini-complete,
Roche Mannheim, Germany), containing 0.1M PMSF
and 50 KIU aprotinin/ml, pH 9.0. The homogenateswere then stirred for 2 h at 4 �C and centrifuged
(12,000 rpm at 4 �C, for 15min). The supernatants were
collected and the protein content determined by the
Bradford micro-method (Bio-Rad, Hercules, CA).
2.3. ELISA
The presence of GH-immunoreactivity (GH-IR) inthe extracts was investigated using an enzyme-linked
immunosorbant assay (ELISA) (Martinez-Coria et al.,
2002). This assay showed a close parallelism between
pituitary and testicular extracts with the standard curve
(data not shown). Briefly, 96-well microtiter plates
(Immulon 2HB, Chantully, VA) were coated overnight
at 4 �C, with 12 ng recombinant chicken GH (rcGH) in
100ll 1M carbonate buffer, pH 10.3. The plates wereextensively washed with TPBS (0.01M sodium phos-
phate, 0.15mM NaCl, 0.05% v/v Tween 20, pH 7) five
times using an automatic microplate immunowasher
(Bio-Rad, Mod.1250, Hercules, CA). This procedure
was performed after each incubation step. Tissue ex-
tracts or serial dilutions of rcGH (5120—5 ng/ml) in
TPBS were then incubated for 16 h with 100 ll primary
antibody diluted 1:100,000 with TPBS and 1% w/v non-fat dried milk raised in rabbits against purified pituitary-
derived chicken GH (Ar�amburo et al., 1989b). This
antibody is specific for GH and has no cross-reactivity
(<0.001%) with any other pituitary hormones (Ar�am-
buro et al., 1989b). The samples and standards (100 ll)were then added to the coated wells and incubated for a
further 2 h at room temperature. Horseradish peroxi-
dase–anti-rabbit IgG conjugate (Bio-Rad) was thenadded (at a dilution of 1:3000 in nonfat dry milk, in
TPBS) and incubated for 2 h at room temperature.
Bound secondary antibodies were then developed by
reaction with 2,20-amino-di-[3-ethylbenzothiazoline sul-
fonate] substrate (Roche Mannheim, Germany). The
plates were read 1 h later, in an automatic ELISA Mi-
croplate Reader (Bio-Rad), at a wavelength of 405 nm.
The assay has a sensitivity of 2 ng/well (20 ng/ml) andinter-assay and intra-assay coefficients of variation of
<4% (Martinez-Coria et al., 2002).
2.4. Western blotting
GH-IR in tissue extracts was also investigated by
Western blotting. Samples were analyzed by sodium
dodecyl sulphate–polyacrylamide gel electrophoresis(SDS–PAGE) in 1.0mm thick, 6 cm long, 15% gels;
using the buffer system of Laemmli (1970) in a mini-
Protean II cell (Bio-Rad). Samples were electrophore-
sed, under reducing conditions (RC, in the presence of
5% (w/v) 2-mercaptoethanol) and under non-reducing
conditions (NRC). After electrophoresis, the gels were
equilibrated in transfer buffer (25mM Tris–HCl,
192mM glycine, 20% (v/v) methanol, pH 8.3) for 30–60min and electrotransferred (at 200mA for 60min) to
nitrocellulose membranes (Bio-Rad). Later, the mem-
branes were washed with 30mM Tris, 500mM NaCl,
pH 7.5 (TBS) for 5min, then blocked with 5% (w/v)
nonfat dried milk (Bio-Rad) in TBS, for 2 h at room
temperature. After washing the membranes with TTBS
for 15min, they were incubated overnight at room
temperature with rabbit-anti-chicken GH antisera(Ar�amburo et al., 1989a,b, 1990), diluted 1:10,000 in
TTBS with 1% (w/v) nonfat dried milk. The membranes
were then rinsed three times (for 15min) in TTBS and
antibody binding was visualized using anti-rabbit IgG–
horseradish peroxidase conjugate (Bio-Rad), diluted
1:3000 in 1% (w/v) nonfat dried milk in TTBS for 2 h.
Immunoreactive bands were developed by incubation in

Fig. 1. Growth hormone (GH) concentrations in the pituitary glands
(Pit), testes (Tes), and vas deferens (Vas Def) of 4-, 9-, and 15-week
(w)-old male chickens as determined by ELISA. Results are expressed
as means� SEM, n ¼ 6. The asterisks indicate values significantly
different from the corresponding pituitary control (P < 0:001). The +
symbols indicate groups significantly different from those in the
4-week-old birds (P < 0:05).
M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49 39
ECL chemiluminescent reagent (Amersham–Pharmacia,Buckinghamshire, UK) for 1min and exposure to Ko-
dak Biomax ML film.
2.5. Densitometric analysis
The number, amount, and relative proportion of
immunoreactive GH moieties were determined by den-
sitometric analysis after digitalization of the lumino-grams in a HP scanner, using IP LabGel 2.2 software
(Scanalytics, Fairfax, VA, USA), as previously de-
scribed (Ar�amburo et al., 2000; Martinez-Coria et al.,
2002). This assay has a sensitivity <2 ng for the GH
standard and an inter-assay coefficient of variation of
8.5%.
2.6. Immunohistochemistry
For immunohistochemical analysis, testes were fixed
with Bouin-Hollande sublimate for 48 h, dehydrated in
ethanol, and embedded in paraffin. Tissue sections were
rehydrated with xylene, absolute alcohol, 95% alcohol,
70% alcohol, Lugol�s solution, thiosulphate, and dis-
tilled water. Tissue sections were then incubated with a
polyclonal antibody directed to recombinant chickenGH (1:500 dilution, overnight), and then incubated with
horseradish peroxidase-conjugated goat anti-rabbit IgG
(1:200 dilution for 2 h) and developed with 3,30-diam-
inobenzidine tetrahydrochloride (DAB) and H2O2
(Malamed et al., 1997). The specificity of staining was
determined by preabsorbing the primary antibody with
excess recombinant chicken GH (1.5mg/ml). The pri-
mary antibody was also substituted for normal rabbitserum or omitted, for further negative controls. Pitui-
tary tissue sections were also analyzed as a control for
the specificity of the primary antibody although in this
case a higher dilution was employed (1:3000).
2.7. Data analysis
The results were analyzed by one way ANOVA.Differences between the groups were determined by
Tukey�s Multiple Comparison Test.
Fig. 2. Growth hormone (GH) concentrations in the testes of embry-
onic (18 days of incubation, 18e) and day old (1d) chicks and in
cockerels between 4 and 66 weeks of age, as determined by ELISA
(means�SEM, n ¼ 6). Groups with different letters are significantly
different (P < 0:05).
3. Results
3.1. ELISA
GH-IR was first detected by ELISA in extracts from
the testes of 4-, 9-, and 15-week-old birds (Fig. 1). The
GH content was comparable at all ages and <1%
(P < 0:001) of that in the pituitary glands, in which the
GH concentration at 4 week was higher than that at 9
and 15 week. GH-immunoreactivity was also present in
the vas deferens of the 9- and 15-week-old birds, at
concentrations comparable to those in the testis (Fig. 1).In contrast, GH-IR was not detectable in extracts of
hepatic tissues (data not shown).
Testicular GH-IR was similarly seen in a second
study with embryonic, neonatal, juvenile, pubertal, and
adult chickens (Fig. 2). Testicular GH-IR was present at
all ages and was not significantly different between 4 and
66 weeks of age (Fig. 2). Lower testicular GH concen-
trations (P < 0:05, compared with the older birds) werefound in the embryonic and neonatal (1-week-old)
chicks.

40 M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49
3.2. Western blotting
As expected (Ar�amburo et al., 2000, 2001b), the pi-
tuitary glands of 9-week-old cockerels contained im-
munoreactive GH moieties with molecular weights of
15, 26, 29, 32, 40, 45, and 52 kDa, when separated under
RC (Fig. 3A). Under the same conditions rcGH is
electrophoresed as moieties with molecular weights of
26 kDa (monomeric GH), 44, and 52 kDa (dimer GH).Monomeric GH was the most abundant GH moiety in
the pituitary gland (approximately 40%), although
45 kDa GH accounted for approximately 13% of the
pituitary GH-IR (Fig. 4). The relative abundance of the
15, 29, and 52 kDa GH moieties accounted for ap-
proximately 14, 11, and 15% of the total GH-IR, re-
spectively, while the relative abundance of the 32 and
40 kDa proteins was approximately 4 and 3%, in eachcase (Fig. 4).
Fig. 3. Representative Western blot of GH-IR in extracts of the testes (30lg)week-old chicken. (A) The extracts were separated by SDS–PAGE under red
the proteins cross-reacting with a rabbit anti-chicken GH antisera were vis
nescence. Similar data were obtained for extracts from 5 other 9-week-old b
analyzed by Western blot before (B) and after preabsorption of the primary
Fig. 4. Relative proportion (%) of immunoreactive GH moieties in the testis, v
SDS–PAGE (RC) and Western blotting and densitometry (means�SEM, n
Similar immunoreactive GH moieties (with the ex-ception of the 15 kDa band) were also present in extracts
of the 9-week-old chick testis and vas deferens (Fig. 3A),
together with moieties of 14, 17, 23, and 35 kDa, which
were absent in the pituitary. In contrast with the pitui-
tary, the 17 kDa moiety was the most abundant GH
moiety in the reproductive tissues (approximately 40%),
whereas monomer GH only accounted for approxi-
mately 11% of the total GH-IR (Fig. 4). In anotherexperiment, preabsorption of the primary antibody with
excess rcGH (900 lg/ml) abolished staining of the GH-
IR bands both in the pituitary and testis extracts (Figs.
3B and C). Multiple GH-IR bands were also seen in
extracts of embryonic, neonatal, juvenile, and adult pi-
tuitary glands (Fig. 5). At ED 18, the most abundant
were the monomer and 15 kDa variant (Fig. 6), although
bands of 35–45 kDa were also clearly present. Variantsof 35 and 42 kDa, unique to the embryo, also accounted
(Te), vas deferens (30lg) (Vd), and pituitary gland (10lg) (Pit) of a 9-
ucing conditions (RC) and after transfer to nitrocellulose membranes,
ualized using HRP labeled anti-rabbit IgG and enhanced chemilumi-
irds. In another assay, 1 lg of Pit extract and 10 lg of Te extract were
antibody with excess (900lg) rcGH (C).
as deferens and pituitary glands of 9-week-old chickens, as analyzed by
¼ 6).
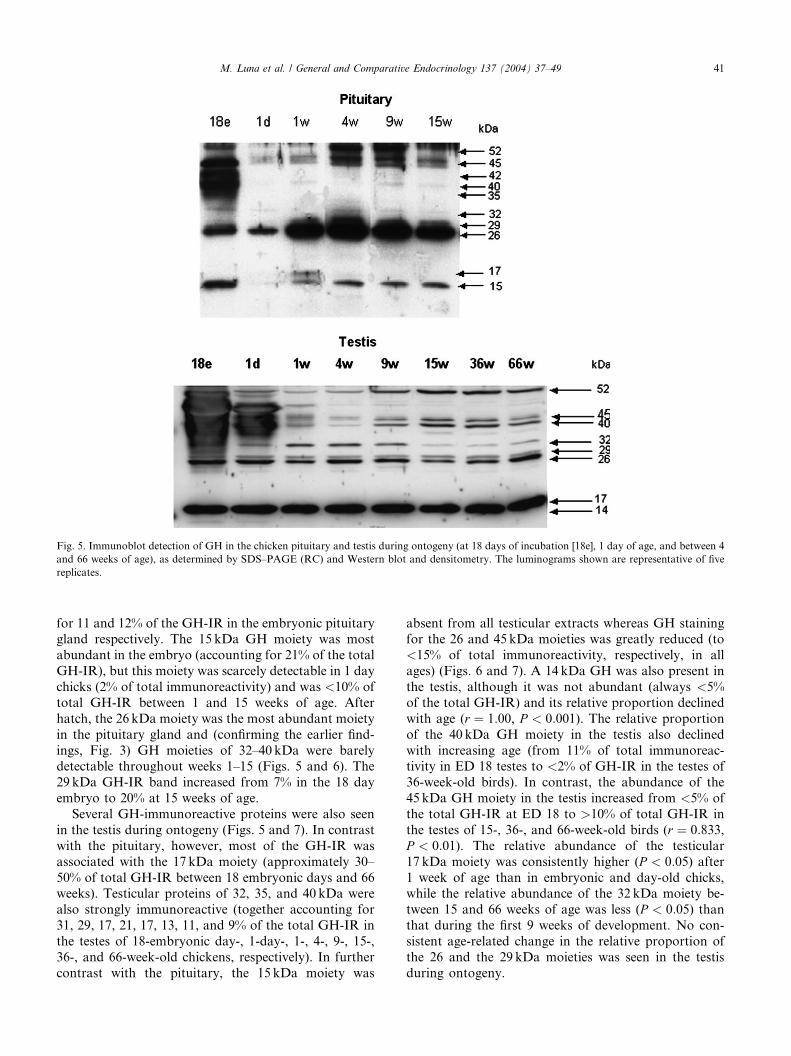
Fig. 5. Immunoblot detection of GH in the chicken pituitary and testis during ontogeny (at 18 days of incubation [18e], 1 day of age, and between 4
and 66 weeks of age), as determined by SDS–PAGE (RC) and Western blot and densitometry. The luminograms shown are representative of five
replicates.
M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49 41
for 11 and 12% of the GH-IR in the embryonic pituitarygland respectively. The 15 kDa GH moiety was most
abundant in the embryo (accounting for 21% of the total
GH-IR), but this moiety was scarcely detectable in 1 day
chicks (2% of total immunoreactivity) and was <10% of
total GH-IR between 1 and 15 weeks of age. After
hatch, the 26 kDa moiety was the most abundant moiety
in the pituitary gland and (confirming the earlier find-
ings, Fig. 3) GH moieties of 32–40 kDa were barelydetectable throughout weeks 1–15 (Figs. 5 and 6). The
29 kDa GH-IR band increased from 7% in the 18 day
embryo to 20% at 15 weeks of age.
Several GH-immunoreactive proteins were also seen
in the testis during ontogeny (Figs. 5 and 7). In contrast
with the pituitary, however, most of the GH-IR was
associated with the 17 kDa moiety (approximately 30–
50% of total GH-IR between 18 embryonic days and 66weeks). Testicular proteins of 32, 35, and 40 kDa were
also strongly immunoreactive (together accounting for
31, 29, 17, 21, 17, 13, 11, and 9% of the total GH-IR in
the testes of 18-embryonic day-, 1-day-, 1-, 4-, 9-, 15-,
36-, and 66-week-old chickens, respectively). In further
contrast with the pituitary, the 15 kDa moiety was
absent from all testicular extracts whereas GH stainingfor the 26 and 45 kDa moieties was greatly reduced (to
<15% of total immunoreactivity, respectively, in all
ages) (Figs. 6 and 7). A 14 kDa GH was also present in
the testis, although it was not abundant (always <5%
of the total GH-IR) and its relative proportion declined
with age (r ¼ 1:00, P < 0:001). The relative proportion
of the 40 kDa GH moiety in the testis also declined
with increasing age (from 11% of total immunoreac-tivity in ED 18 testes to <2% of GH-IR in the testes of
36-week-old birds). In contrast, the abundance of the
45 kDa GH moiety in the testis increased from <5% of
the total GH-IR at ED 18 to >10% of total GH-IR in
the testes of 15-, 36-, and 66-week-old birds (r ¼ 0:833,P < 0:01). The relative abundance of the testicular
17 kDa moiety was consistently higher (P < 0:05) after1 week of age than in embryonic and day-old chicks,while the relative abundance of the 32 kDa moiety be-
tween 15 and 66 weeks of age was less (P < 0:05) thanthat during the first 9 weeks of development. No con-
sistent age-related change in the relative proportion of
the 26 and the 29 kDa moieties was seen in the testis
during ontogeny.
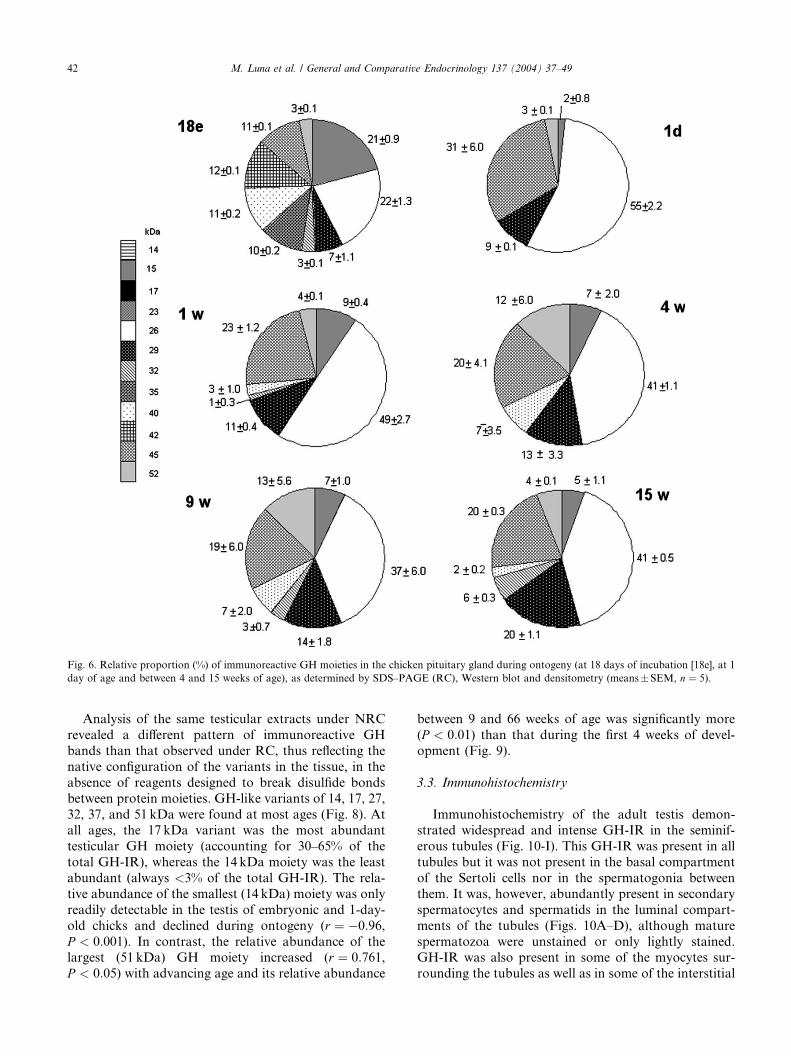
Fig. 6. Relative proportion (%) of immunoreactive GH moieties in the chicken pituitary gland during ontogeny (at 18 days of incubation [18e], at 1
day of age and between 4 and 15 weeks of age), as determined by SDS–PAGE (RC), Western blot and densitometry (means� SEM, n ¼ 5).
42 M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49
Analysis of the same testicular extracts under NRC
revealed a different pattern of immunoreactive GH
bands than that observed under RC, thus reflecting the
native configuration of the variants in the tissue, in the
absence of reagents designed to break disulfide bonds
between protein moieties. GH-like variants of 14, 17, 27,
32, 37, and 51 kDa were found at most ages (Fig. 8). Atall ages, the 17 kDa variant was the most abundant
testicular GH moiety (accounting for 30–65% of the
total GH-IR), whereas the 14 kDa moiety was the least
abundant (always <3% of the total GH-IR). The rela-
tive abundance of the smallest (14 kDa) moiety was only
readily detectable in the testis of embryonic and 1-day-
old chicks and declined during ontogeny (r ¼ �0:96,P < 0:001). In contrast, the relative abundance of thelargest (51 kDa) GH moiety increased (r ¼ 0:761,P < 0:05) with advancing age and its relative abundance
between 9 and 66 weeks of age was significantly more
(P < 0:01) than that during the first 4 weeks of devel-
opment (Fig. 9).
3.3. Immunohistochemistry
Immunohistochemistry of the adult testis demon-strated widespread and intense GH-IR in the seminif-
erous tubules (Fig. 10-I). This GH-IR was present in all
tubules but it was not present in the basal compartment
of the Sertoli cells nor in the spermatogonia between
them. It was, however, abundantly present in secondary
spermatocytes and spermatids in the luminal compart-
ments of the tubules (Figs. 10A–D), although mature
spermatozoa were unstained or only lightly stained.GH-IR was also present in some of the myocytes sur-
rounding the tubules as well as in some of the interstitial

Fig. 7. Relative proportion (%) of immunoreactive GH moieties in the chicken testis during ontogeny (at 18 days of incubation [18e], 1 day of age,
and between 4 and 66 weeks of age), as determined by SDS–PAGE (RC) and Western blotting (means� SEM, n ¼ 5).
Fig. 8. Immunoblot detection of GH moieties in the testis of chickens during ontogeny (at 18 days of incubation [18e], 1 day of age, and between 4
and 66 weeks of age), as determined by SDS–PAGE under non-reducing conditions (NRC), followed by Western blotting and densitometry. The
GH-immunoreactivity of recombinant chicken GH (rcGH), proteolytically cleaved fragment (frag) and the pituitary gland (of adult chickens) is
shown for comparison. The luminograms shown are representative of at least five replicates.
M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49 43
(Leydig) cells between them (Fig. 10B). The specificity of
this staining is further shown by its absence following
the preabsorbtion of the primary antibody with excess
GH (Fig. 10-II) and the complete lack of staining in the
absence of the primary antibody (Fig. 10-III). As an-
other control for the specificity of the antibody em-
ployed, sections of chicken pituitaries were also analyzed
as shown in Fig. 11, where GH-immunoreactive cells

Fig. 9. Relative proportion (%) of immunoreactive GH moieties in the chicken testis during ontogeny (at 18 days of incubation [18e], 1 day of age,
and between 4 and 66 weeks of age), as determined by non-reducing SDS–PAGE, Western blotting, and densitometry (means�SEM, n ¼ 5).
44 M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49
were intensely stained only in the caudal lobe (Fig. 11A)
as expected for the known distribution of somatotropes.
No staining was observed in the caudal lobe after
preadsorption with excess rcGH (Fig. 11C), nor in the
cephalic lobe (Fig. 11B) or in the negative control(Fig. 11D).
4. Discussion
These results clearly demonstrate the presence of GH-
like proteins in the testis and the male reproductive tract
and suggest local roles for GH in male reproductivefunction.
Although GH gene expression has been demonstrated
in the human testis, it is primarily of the placental GH
variant rather than the pituitary GH gene (Untergasser
et al., 1997, 1998). The placental GH variant is not
present in chickens, in which the testicular GH proteins
are thus likely to reflect translation of the pituitary GH
gene. The recent demonstration of a GH mRNA in thechicken testis, with a nucleotide sequence >99.5% ho-
mologous to that in the pituitary (Harvey et al., 2002),
supports this possibility. Furthermore, immunoreactive
GH in the thymus, bursa, spleen, and brain of chickens
(Render et al., 1995a,b) is similarly thought to reflect the
extrapituitary expression of the pituitary GH gene
Fig. 11. GH-immunoreactivity in 4lm sections of 4-week-old chicken pi
recombinant cGH (1:3000 dilution): (A) caudal lobe; (B) cephalic lobe; (C) G
with excess rcGH; and (D) negative control without primary antibody.
rather than a variant GH gene, since GH cDNAs in
these tissues are identical to pituitary GH cDNA.
This is the first report on the presence of GH in the
testis, although GH has previously been demonstrated
in ovarian tissue (Mendoza et al., 2002; Schw€arzleret al., 1997; Zhao et al., 2002). The GH-IR in the testis
was clearly associated with proteins of multiple size.
This molecular heterogeneity is also characteristic of
pituitary GH and is thought to partly result from post-
translational modifications of the protein (Ar�amburo
et al., 2000, 2001b). Indeed, in the chicken several GH
variants have been described in the pituitary gland:
phosphorylated GH, glycosylated GH; oligomeric GH(dimer, trimer, tetramer, and pentamer), cleaved GH
and various charge variants (Ar�amburo et al., 1989a,
1990, 1991, 2001b). In the present study all the immu-
noreactive GH moieties in the testis appeared to be
present in the pituitary gland, although the relative
proportion of these moieties clearly differed in different
tissues. The most abundant GH moiety in the pituitary
gland, for instance, has a molecular mass of 26 kDawhen electrophoresed under RC, whereas the 17 kDa
variant is the most abundant GH variant in the testis
and vas deferens. Similarly the GH moieties of 32, 35,
and 40 kDa are much more abundant in the testis and
vas deferens than in the pituitary gland. This suggests
the translation of the GH transcript or the processing of
tuitary. 100� amplification of sections incubated with antibody to
H-immunoreactivity is lost in the caudal lobe following preabsorption
c

Fig. 10. GH-immunoreactivity in 6lm sections of 15-week-old chicken testes. Upper micrographs: (I) 20� amplification of sections incubated with
antibody to recombinant cGH (1:500 dilution). (II) GH immunoreactivity is lost following preabsorption of the antibody with excess rcGH. (III)
negative control without primary antibody. Lower micrographs: 40� and 100� amplification of sections showing positive GH-like immunoreac-
tivity. Arrowheads indicate immunopositive cell types: (A), (C), and (D) germ cells (primary/secondary spermatocytes and spermatids); (B) Leydig
(interstitial) cells and myoid cells.
M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49 45

46 M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49
the mature hormone may be tissue-specific. It may alsobe age-related.
In the pituitary, the relative abundance of the 26 and
45 kDa GH moieties was relatively constant following
hatch, although the proportions of both were less in
embryonic birds, in which the 15 kDa variant was rela-
tively more abundant than in hatched birds. These on-
togenic differences confirm previous findings on the
relative abundance of these moieties in the pituitarygland (Ar�amburo et al., 2000). Ontogenic changes in the
relative abundance of testicular GH moieties were also
evident. These included marked decreases in the abun-
dance of the 14 and 40 kDa forms (detected under RC)
between ED 18 and 66 weeks of age. Similarly, while the
relative abundance of the 17 kDa form increased be-
tween 1 and 7 days of age, it remained constant there-
after. The relative abundance of the 45 kDa moiety alsoincreased steadily between ED 18 and 15 weeks of age.
When analyzed under NRC, the testicular immunore-
active GH bands reflect their in situ conditions in the
tissue, maintaining the integrity of the disulfide bonds.
Interestingly, the monomeric GH isoform (22 kDa) was
only present in the embryonic testis (ED 18) but not
later in development. The profile observed under this
conditions after hatching revealed mainly the presenceof submonomeric forms (14 and 17 kDa) and oligomeric
forms (32,37,51 kDa) thus suggesting that at these stages
the hormone may be preferentially processed to sub-
monomeric forms (e.g., by proteolytic cleavage) or
complexed through disulfide bonding. The relative
proportion of 14 kDa GH in the testicular extracts again
decreased with age, whereas the 17 and 51 kDa GH
moieties increased (between ED 18 and 7 days and be-tween 4 and 9 weeks of age, respectively). Since the
production of GH moieties in the pituitary gland can be
differentially regulated by GH-secretagogues (Martinez-
Coria et al., 2002) it is possible that these ontogenic
changes reflect age-related differences in the control of
GH secretion (Harvey, 1995), although the factors reg-
ulating extrapituitary GH secretion are still largely un-
known (Harvey et al., 2001). It may, however, bepertinent that many of the major factors that regulate
the release of pituitary GH (e.g., GHRH, Matsubara
et al., 1995, Olchovsky et al., 1996; ghrelin, Tanaka
et al., 2001, Tena-Sempere et al., 2002; TRH, Wilber
and Xu, 1998, Li et al., 2002; somatostatin, Pekary et al.,
1984, Zhu et al., 1998; IGF-I, Morera et al., 1987, Baker
et al., 1996) are also present in the testis.
The GH variants in the chicken pituitary gland havebeen shown to differ in biological activity and potency
(Ar�amburo et al., 1989c, 1990, 2001b). Tissue and age-
related changes in the relative abundance of testicular
GH variants may therefore also reflect changes in GH
activity and reproductive function. Indeed, it is now well
established that the testis and the male reproductive
tract are target sites for GH action (Hull and Harvey,
2000a,b). GH receptor (GHR) immunoreactivity andGHR mRNA are present in the testes of fetal and adult
rats (Garcia-Aragon et al., 1992; Lobie et al., 1990,
1992) and GHR mRNA is present in the testes of adult
chickens (Harvey et al., 2002; Lobie et al., 1990). High-
affinity GH-binding sites are also present in the trout
testis (Gomez et al., 1998; Le Gac et al., 1992, 1996) and
rat testis (Bonifacino and Dufau, 1985). These receptors
are thus likely to mediate endocrine actions of GHwithin the testis, since small testis and male infertility are
associated with pituitary GH deficiency (Shimonovitz
et al., 1993; Shoham et al., 1992; Spiteri-Grech and
Nieschlog, 1992). Exogenous (monomer) GH has also
been shown to induce testicular steroidogenesis (Kan-
zaki and Morris, 1999) and gametogenesis (Gravance
et al., 1997; Ovesen et al., 1998), but biological roles for
other GH moieties have yet to be determined, and theseactions could be autocrine or paracrine, in view of the
distribution of GH and GH receptors in the testis.
Within the testis, GH-IR was largely associated with
the developing germ cells, although it was not present in
spermatogonia. It is therefore of interest that GH has
mitotic activity in germ cells (Loir, 1999) and regulates
the longevity of spermatozoa in vitro (Champion et al.,
2002). It may also be responsible for the production ofIGF-I and IGF mRNA in germ cells (Dombrowicz
et al., 1992) and in Leydig and Sertoli cells, since GHRs
are present in these cell types (Gomez et al., 1998; Lobie
et al., 1990). On the other hand, some results suggest
that in the chicken testis GH seems to inhibit IGF-I
mRNA expression (Tanaka et al., 1996). It is also of
interest that GHRH and GHRH mRNA are present in
testicular germ cells (Fabbri et al., 1995; Pescovitz et al.,1990; Srivastava et al., 1993), in which the promoter is
spermatogenic specific (Srivastava et al., 1995). Testic-
ular GHRH in the rat, like testicular GH in chicken also
appears to be stage related in germ cells and is not
present in mature sperm (Fabbri et al., 1995). Testicular
GHRH in the rat (Fabbri et al., 1995), like testicular
GH in the chicken, also appears to be absent in Sertoli
cells. GH-IR is, in contrast, present in some interstitialcells, in which exogenous monomer GH has been shown
to activate steroidogenesis (Kanzaki and Morris, 1999).
The finding of GH in some myocytes surrounding the
seminiferous tubules is consistent with the presence of
GH mRNA in the smooth muscle surrounding blood
vessels (Recher et al., 2001) and may indicate an auto-
crine role for GH in tubule contraction.
The GH concentration detected in the testis is muchlower (1%) than that in the pituitary and hence it is
unlikely that contributes significantly to circulating GH,
and is probably for local actions in the testis. This
suggestion is substantiated by the fact that the immu-
nohistochemistry results indicate that the GH-IR is in-
side the blood–testis barrier which would support the
notion that it is not secreted into the plasma. On the

M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49 47
other hand, it is of interest to consider that the testismust be 5–1000 times the size of the pituitary, so the
total testicular GH content at some ages, particularly
after sexual maturation, can be significant and maybe
comparable to that in the pituitary.
In addition to the testis, GH was also detected (at
comparable concentrations) in the vas deferens of ju-
venile cockerels. It is therefore of interest that the GH
and GHR genes are also expressed in the vas deferens(Harvey et al., 2002). The differentiation of the vas
deferens from the embryonic Wolffian duct also appears
to be GH-dependent, since it is blocked in fetal mice by
GH immunoneutralization and restored by exogenous
GH (Nguyen et al., 1996). GH and GHRs have similarly
been shown, by immunocytochemistry, to be present in
the Wolffian ducts of embryonic chicks (Harvey et al.,
2000; Wang, 1989). The cellular location of the GH-IRin the vas deferens is, however, unknown and its mo-
lecular characteristics are uncertain.
In summary, these results demonstrate abundant
GH-IR in the testis of embryonic, neonatal, juvenile,
pubertal, and adult chickens. This immunoreactivity
increases during development and is associated with
numerous molecular GH moieties, the relative propor-
tions of which generally differ from those in the pituitarygland. Tissue and age-related changes in the relative
abundance of testicular GH variants may be related to
changes in reproductive function, and may reflect local
(autocrine/paracrine) actions of testicular GH.
Acknowledgments
We thank Martha Carranza (lab technician), Dr.Alfonso C�arabez and Lourdes Palma (histochemistry
advice), Pilar Galarza-Barrios (librarian); Martin
Garc�ia-Serv�in (vivarium), Alberto Lara, Omar
Gonz�alez-Hern�andez, Fernando L�opez-Barrera (com-
puting and figure editing), Leopoldo Gonz�alez-Santos(digital analysis), and Gerardo Courtois for excellent
technical assistance. This work was supported by grants
from DGAPA-UNAM (PAPIIT IN227399 andIN226002-3), CONACyT (31817-N and 41218-Q), Pil-
grim�s Pride of Mexico and NSERC of Canada.
References
Allen, J.T., Bloor, C.A., Kedia, R.K., Knight, R.A., Spiteri, M.A.,
2000. Expression of growth hormone-releasing factor, growth
hormone, insulin-like growth factor-1 and its binding proteins in
human lung. Neuropeptides 34, 98–107.
Ar�amburo, C., Carranza, M., S�anchez, R., Perera, G., 1989a. Partial
biochemical and biological characterization of purified chicken
growth hormone (cGH). Isolation of cGH charge variants and
evidence that cGH is phosphorylated. Gen. Comp. Endocrinol. 76,
330–339.
Ar�amburo, C., S�anchez, R., Fenton, B., Perera, G., Valverde-Rodri-
guez, C., 1989b. Desarrollo de un radioinmunoensayo homologo y
especifico para la determinacion de hormona de crecimiento de
pollo (cGH). Vet. Mex. 20, 397–405.
Ar�amburo, C., Campbell, R.M., Scanes, C.G., 1989c. Heterogeneity of
chicken growth hormone (cGH). Identification of lipolytic and
non-lipolytic variants. Life Sci. 45, 2201–2207.
Ar�amburo, C., Montiel, J.L., Perera, G., Navarrete, S., Sanchez, R.,
1990. Molecular isoforms of chicken growth hormone (cGH):
different bioactivities of cGH charge variants. Gen. Comp.
Endocrinol. 80, 59–67.
Ar�amburo, C., Navarrete, S., Montiel, J.L., Sanchez, R., Berghman,
L.R., 1991. Purification and electrophoretic analysis of glycosy-
lated chicken growth hormone (G-cGH): evidence of G-cGH
isoforms. Gen. Comp. Endocrinol. 84, 135–146.
Ar�amburo, C., Luna, M., Carranza, M., Reyes, M., Mart�inez-Coria,H., Scanes, C.G., 2000. Growth hormone size variants: changes in
the pituitary during development of the chicken. Proc. Soc. Exp.
Biol. Med. 223, 67–74.
Ar�amburo, C., Carranza, M., Mart�inez-Coria, H., Reyes, M., Beru-
men, L., L�opez, J., Pascacio, H., Huerta, L., Luna, M., 2001a.
Molecular and functional heterogeneity of growth hormone. In:
Dawson, A., Chaturvedi, C.M. (Eds.), Avian Endocrinology.
Narosa Publishing House, New Delhi, pp. 273–286.
Ar�amburo, C., Carranza, M., Reyes, M., Luna, M., Martinez-Coria,
H., Berumen, L., Scanes, C.G., 2001b. Characterization of a
bioactive 15 kDa fragment produced by proteolytic cleavage of
chicken growth hormone. Endocrine 15, 231–240.
Baker, J., Hardy, M.P., Zhou, J., Bondy, C., Lupu, F., Bellve, A.R.,
Efstratiadis, A., 1996. Effects of an IGF1 gene null mutation on
mouse reproduction. Mol. Endocrinol. 10, 903–918.
Baudet, M.L., Sanders, E.J., Harvey, S., 2003. Retinal growth
hormone in chicken embryo. Endocrinology 144, 5459–5468.
Berger, P., Untergasser, G., Hermann, M., Hittmair, A., Madersb-
acher, S., Dirnhofer, S., 1999. The testis-specific expression pattern
of the growth hormone/placental lactogen (GH/PL) gene cluster
changes with malignancy. Human Pathol. 30, 1201–1206.
Boguszewski, C.L., Svensson, P.-A., Jansson, T., Clark, R., Carlsson,
L.M.S., Carlsson, B., 1998. Cloning of two novel growth hormone
transcripts expressed in human placenta. Clin. Endocrinol. Metab.
83, 2878–2885.
Bonifacino, J.S., Dufau, M.L., 1985. Lactogen receptors in rat Leydig
cells: analysis of their structure with bifunctional cross-linking
reagents. Endocrinology 116, 1610–1614.
Costa, A., Zoppetti, G., Benedetto, C., Bertino, E., Marozio, L.,
Fabris, C., Arisio, R., Giraudi, G.F., Testori, O., Ariano, M., et al.,
1993. Immunelike growth hormone substance in tissues from
human embryos/fetuses and adults. J. Endocrinol. Invest. 16,
625–633.
Champion, Z.J., Vickers, M.H., Gravance, C.G., Breier, B.H., Casey,
P.J., 2002. Growth hormone or insulin-like growth factor-I extends
longevity of equine spermatozoa in vitro. Theriogenology 57, 1793–
1800.
Dombrowicz, D., Hooghe-Peters, E.L., Gothot, A., Sente, B., Vanha-
elst, L., Closset, J., Hennen, G., 1992. Cellular localization of IGF-
I and IGF-II mRNAs in immature hypophysectomized rat testis
and epididymis after in vivo hormonal treatment. Arch. Int.
Physiol. Biochim. Biophys. 100, 303–308.
Fabbri, A., Ciocca, D.R., Ciampani, T., Wang, J., Dufau, M.L., 1995.
Growth hormone-releasing hormone in testicular interstitial and
germ cells: potential paracrine modulation of follicle-stimulating
hormone action on Sertoli cell function. Endocrinology 136, 2303–
2308.
Garcia-Aragon, J., Lobie, P.E., Muscat, G.E.O., Gobius, K.S.,
Norstedt, G., Waters, M.J., 1992. Prenatal expression of growth
hormone (GH) receptor/binding protein in the rat: a role for GH in
embryonic and fetal development? Development 114, 869–876.

48 M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49
Gomez, J.M., Loir, M., Le Gac, F., 1998. Growth hormone receptors
in testis and liver during the spermatogenetic cycle in rainbow trout
(Oncorhynchus mykiss). Biol. Reprod. 58, 483–491.
Gravance, C.G., Breier, B.H., Vickers, M.H., Casey, P.J., 1997.
Impaired sperm characteristics in postpubertal growth-hormone-
deficient dwarf (dw/dw) rats. Anim. Reprod. Sci. 49, 71–76.
Harvey, S., 1995. Growth hormone release: integrative hypothalamic
control. In: Harvey, S., Scanes, C.G., Daughaday, D.H. (Eds.),
Growth Hormone. CRC Press, Boca Raton, pp. 131–162.
Harvey, S., Johnson, C.D.M., Sanders, E.J., 2000. Extrapituitary
growth hormone in peripheral tissues of early chick embryos. J.
Endocrinol. 166, 489–502.
Harvey, S., Johnson, C.D.M., Sanders, E.J., 2001. Growth hormone in
neural tissues of the chick embryo. J. Endocrinol. 169, 487–
498.
Harvey, S., Murphy, A.E., Hull, K.L., Luna, M., Ar�amburo, C., 2002.
Testicular growth hormone (GH) and GH mRNA. GH IGF Res.
12, 262.
Hu, L., Lytras, A., Bock, M.E., Yuen, C.K., Dodd, J.G., Cattini, P.A.,
1999. Detection of placental growth hormone variant and chori-
onic somatomammotropin-L RNA expression in normal and
diabetic pregnancy by reverse transcriptase-polymerase chain
reaction. Mol. Cell. Endocrinol. 157, 131–142.
Hull, K.L., Harvey, S., 2000a. Growth hormone: roles in male
reproduction. Endocrine 13, 243–250.
Hull, K.L., Harvey, S., 2000b. Growth hormone: a reproductive
endocrine–paracrine regulator? Rev. Reprod. 5, 175–182.
Hull, K.L., Thiagarajah, A., Harvey, S., 1996. Cellular localization of
growth hormone receptors/binding proteins in immune tissues. Cell
Tissue Res. 286, 69–80.
Hull, K.L., Nette, F., Harvey, S., 2001. Bidirectional communication
between the immune and neuroendocrine system: role of growth
hormone. In: Dawson, A., Chaturvedi, C.M. (Eds.), Avian
Endocrinology. Narosa Publishing House, New Delhi, pp. 437–
443.
Kanzaki, M., Morris, P.L., 1999. Growth hormone regulates steroi-
dogenic acute regulatory protein expression and steroidogenesis in
Leydig cell progenitors. Endocrinology 140, 1681–1686.
Kyle, C.V., Evans, M.C., Odell, W.D., 1981. Growth hormone-like
material in normal human tissues. Clin. Endocrinol. Metab. 53,
1138–1144.
Laemmli, U.K., 1970. Cleavage of structural proteins during assembly
of the head of bacteriophage T4. Nature 227, 680–685.
Le Gac, F., Ollitrault, M., Loir, M., Le Bail, P.Y., 1992. Evidence for
binding and action of growth hormone in trout testis. Biol. Reprod.
46, 949–957.
Le Gac, F., Loir, M., Le Bail, P.Y., Ollitrault, M., 1996. Insulin-like
growth factor (IGF-1) mRNA and IGF-1 receptor in trout testis
and in isolated spermatogenic and Sertoli cells. Mol. Reprod. Dev.
44, 23–35.
Li, Z., Zhang, Y., Liu, X., Xu, R., 2002. Correlation of expression of
preprothyrotropin-releasing hormone and receptor with rat testis
development. Chin. Med. 115, 6–12.
Lobie, P.E., Breiphol, W., Aragon, J.G., Waters, M.J., 1990. Cellular
localization of the growth hormone receptor/binding protein in the
male and female reproductive systems. Endocrinology 126, 2214–
2221.
Lobie, P.E., Garcia-Aragon, J., Wang, B.S., Baumbach, W.R., Waters,
M.J., 1992. Cellular localization of the growth hormone binding
protein in the rat. Endocrinology 130, 3057–3065.
Loir, M., 1999. Spermatogonia of rainbow trout: II. In vitro study of
the influence of pituitary hormones, growth factors and steroids on
mitotic activity. Mol. Reprod. Dev. 53, 434–442.
Malamed, S., Deaver, D., Perez, F., Radecki, S., Gibney, J., Scanes,
C.G., 1997. Quantitative studies of chicken somatotrophs during
growth and development by morphometry, immunocytochemistry,
and flow cytometry. Gen. Comp. Endocrinol. 108, 25–34.
Martinez-Coria, H., Lopez-Rosales, L.J., Carranza, M., Berumen, L.,
Luna, M., Ar�amburo, C., 2002. Differential secretion of chicken
growth hormone variants after growth hormone-releasing hormone
stimulation in vitro. Endocrine 17, 91–102.
Matsubara, S., Sato, M., Mizobuchi, M., Niimi, M., Takahara, J.,
1995. Differential gene expression of growth hormone (GH)-
releasing hormone (GRH) and GRH receptor in various rat tissues.
Endocrinology 136, 4147–4150.
Mendoza, C., Ruiz-Requena, E., Ortega, E., Cremades, N., Martinez,
F., Bernabeu, R., Greco, E., Tesarik, J., 2002. Follicular fluid
markers of oocyte developmental potential. Human Reprod. 17,
1017–1022.
Morera, A.M., Chauvin, M.A., de Peretti, E., Binoux, M., Benahmed,
M., 1987. Somatomedin C/insulin-like growth factor 1: an
intratesticular differentiative factor of Leydig cells? Horm. Res.
28, 50–57.
Nguyen, A.P., Chandorkar, A., Gupta, C., 1996. The role of growth
hormone in fetal mouse reproductive tract differentiation. Endo-
crinology 137, 3659–3666.
Olchovsky, D., Bruno, J.F., Berelowitz, M., 1996. Growth hormone-
releasing factor expression is discordantly regulated in the hypo-
thalamus and testis of streptozotocin-diabetic rats. Endocrinology
148, 189–192.
Ovesen, P., Jorgensen, J.O., Ingerslev, J., Orskov, H., Christiansen,
J.S., 1998. Growth hormone treatment of men with reduced sperm
quality. Ugeskr Laeger 160, 176–180.
Palmetshofer, A., Zechner, D., Luger, T.A., Barta, A., 1995. Splicing
variants of the human growth hormone mRNA: detection in
pituitary, mononuclear cells and dermal fibroblasts. Mol. Cell.
Endocrinol. 113, 225–234.
Pekary, A.E., Yamada, T., Sharp, B., Bhasin, S., Swerdloff, R.S.,
Hershman, J.M., 1984. Somatostatin-14 and -28 in the male rat
reproductive system. Life Sci. 5, 939–945.
Pescovitz, O.H., Berry, S.A., Laudon, M., Ben-Jonathan, N., Martin-
Myers, A., Hsu, S.M., Lambros, T.J., Felix, A.M., 1990. Locali-
zation and growth hormone (GH)-releasing activity of rat testicular
GH-releasing hormone-like peptide. Endocrinology 127, 2336–
2342.
Recher, S., Raccurt, M., Lambert, A., Lobie, P.E., Mertani, H.C.,
Morel, G., 2001. Prenatal and adult growth hormone gene
expression in rat lymphoid organs. Histochem. Cytochem. 49,
347–354.
Render, C.L., Hull, K.L., Harvey, S., 1995a. Neural expression of the
pituitary GH gene. J. Endocrinol. 147, 413–422.
Render, C.L., Hull, K.L., Harvey, S., 1995b. Expression of the growth
hormone gene in immune tissues. Endocrine 3, 729–735.
Schw€arzler, P., Untergasser, G., Hermann, M., Dirnhofer, S., Abend-
stein, B., Madersbacher, S., Berger, P., 1997. Selective growth
hormone/placental lactogen gene transcription and hormone pro-
duction in pre- and postmenopausal human ovaries. Clin. Endo-
crinol. Metab. 82, 3337–3341.
Shimonovitz, S., Zacut, D., Benchetrit, A., Ron, M., 1993. Growth
hormone status in patients with maturation arrest of spermato-
genesis. Human Reprod. 8, 919–921.
Shoham, Z., Homburg, R., Owen, E.J., Conway, S.S., Ostergaard, H.,
Jacobs, H.S., 1992. The role of treatment with growth hormone in
infertile patients. Baillieres Clin. Obst. Gynecol. 6, 267.
Slominski, A., Malarkey, W.B., Wortsman, J., Asa, S.I., Carlson, A.,
2000. Human skin expresses growth hormone but not the prolactin
gene. Lab. Clin. Med. 136, 476–481.
Spiteri-Grech, J., Nieschlog, E., 1992. The role of growth hormone and
insulin-like growth factor I in the regulation of male reproductive
function. Horm. Res. 38 (Suppl. 1), 22–27.
Srivastava, C.H., Collard, M.W., Rothrock, J.K., Peredo, M.J., Berry,
S.A., Pescovitz, O.H., 1993. Germ cell localization of a testicular
growth hormone-releasing hormone-like factor. Endocrinology
133, 83–89.

M. Luna et al. / General and Comparative Endocrinology 137 (2004) 37–49 49
Srivastava, C.H., Monts, B.S., Rothrock, J.K., Peredo, M.J., Pesco-
vitz, O.H., 1995. Presence of a spermatogenic-specific promoter in
the rat growth hormone-releasing hormone gene. Endocrinology
136, 1502–1508.
Tanaka, M., Hayashida, Y., Nakao, N., Nakai, N., Nakashima, K.,
2001. Testis-specific and developmentally induced expression of a
ghrelin gene-derived transcript that encodes a novel polypeptide in
the mouse. Biochem. Biophys. Acta 1522, 62–65.
Tanaka, M., Hayashida, Y., Sakaguchi, K., Ohkubo, T., Wakita, M.,
Hoshino, S., Nakashima, K., 1996. Growth hormone-independent
expression of insulin-like growth factor I messenger ribonucleic
acid in extrahepatic tissues of the chicken. Endocrinology 137, 30–
34.
Tena-Sempere, M., Barreiro, M.L., Gonzalez, L.C., Gaytan, F.,
Zhang, F.P., Caminos, J.E., Pinilla, L., Casanueva, F.F., Dieguez,
C., Aguilar, E., 2002. Novel expression and functional role of
ghrelin in rat testis. Endocrinology 143, 717–725.
Tresguerres, J.A., Ariznavarreta, C., Granados, B., Costoy, J.A.,
P�erez-Romero, A., Salam�e, F., Hermanussen, M., 1999. Salivary
gland is capable of GH synthesis under GHRH stimulation. J.
Endocrinol. 160, 217–222.
Untergasser, G., Kranewitter, W., Schwarzler, P., Berger, P., 1997.
Organ-specific expression pattern of the human growth hormone/
placental lactogen gene-cluster in the testis. Mol. Cell. Endocrinol.
130, 53–60.
Untergasser, G., Hermann, M., Rumpold, H., Berger, P., 1998.
Complex alternative splicing of the GH-V gene in the human testis.
Eur. J. Endocrinol. 39, 424–427.
Wang, J.J., 1989. Immunocytochemical demonstration of the binding
of growth-related polypeptide hormones on chick embryonic
tissues. Histochemistry 93, 133–141.
Wilber, J.F., Xu, A.H., 1998. The thyrotropin-releasing hormone gene:
cloning, characterization, and transcriptional regulation in the
central nervous system, heart, and testis. Thyroid 8, 897–
901.
Wu, H., Devi, R., Malarkey, W.B., 1996. Localization of growth
hormone messenger ribonucleic acid in the human immune
system—a clinical research study. Clin. Endocrinol. Metab. 81,
1278–1282.
Yoshizato, H., Fujikawa, T., Soya, H., Tanaka, M., Nakashima, K.,
1998. The growth hormone (GH) gene is expressed in the lateral
hypothalamus: enhancement by GH-releasing hormone and re-
pression by restraint stress. Endocrinology 139, 2545–2551.
Zhang, C.Z., Li, H., Young, W.G., Bartold, P.M., Chen, C., Waters,
M.J., 1997. Evidence for local action of growth hormone in
embryonic tooth development in the rat. Growth Factors 14, 131–
143.
Zhao, J., Taverne, M.A., Van, W.G., Bevers, M.M., Van, H.R., 2002.
Immunohistochemical localization of growth hormone (GH), GH
receptor (GHR), insulin-like growth factor I (IGF-I) and type I
IGF-I receptor, and gene expression of GH and GHR in rat pre-
antral follicles. Zygote 10, 85–94.
Zhu, L.J., Krempels, K., Bardin, C.W., O�Carroll, A.M., Mezey, E.,
1998. The localization of messenger ribonucleic acids for somato-
statin receptors 1, 2, and 3 in rat testis. Endocrinology 139,
350–357.




















