
Ontogeny of form and function: Locomotor morphology and drag in zebrafish (Danio rerio)

Anim. Behav., 1989,37,415-428
The ontogeny of foraging in squirrel monkeys, Saimiri oerstedi
S. BOINSKI* & D. M. FRAGASZYT * Laboratory of Comparative Ethology, National Institute of Child Health and Human Development,
NIH Animal Center, P.O. Box 289, Bldg 112, Poolesville, MD 20837, U.S.A. t Department of Psychology, Washington State University, PuIlman. WA 99164, U.S.A.
Abstract. The development of foraging and locomotor independence by wild infant squirrel monkeys is described. Infants began to locomote independently during the fourth week of life and successful foraging appeared within the next 2 weeks. Infants were weaned by 16 weeks. The frequency and targets of foraging activities in infants became indistinguishable from those of older animals within 4.5 months after the onset of foraging, with four exceptions. Compared with older animals, infants exhibited (1) greater reliance on methods that did not involve prehension of small prey, (2) less visual inspection of potential foraging sites, (3) less skilled processing of certain prey to remove potentially toxic gut contents and to prevent escape, and (4) less avoidance of urticaceous caterpillars. Observations of infants watching other infants experiencing difficulties with noxious prey and prevention of approach to noxious prey by adults suggest that avoidance of certain species could be transmitted socially. Otherwise, no direct social influence on foraging was shown. Only a few features of foraging require extensive individually acquired motor skills, and weanlings can feed themselves adequately before these skills are developed. Infants in the wild population were weaned much earlier than has been observed in captive groups, and at about the age at which abilities are sufficiently mature for foraging to reach minimal efficiency. The age of weaning in this population probably represents the lower limit for this taxon.
This report describes the time-course of weaning and the ontogeny of foraging in infants born into a troop of wild Central American squirrel monkeys. Because successful foraging is nearly always a prerequisite to successful reproduction, and because provision of nourishment to offspring is costly to care-givers, stringent selective constraints on the duration of lactation and the assumption of nutritional independence by offspring can be assumed.
An evolutionary and developmental perspective leads to consideration of how selective constraints on foraging are translated into species-typical patterns of behavioural development. We consider three questions here. (1) How and when does weaning occur in relation to the appearance of efficient foraging by the young individual? (2) How does foraging in the young animal differ from foraging in its older group-mates, and how does it become more like that of older animals? (3) How does this pattern reflect selection for, or constraints on, timing of weaning?
The development of foraging competence by young primates apparently requires greater time, and presumably learning, than the development of foraging competence in other orders; this feature of
primate biology is intimately associated with other aspects of life history peculiar to primates (Alt- mann & Alberts 1987). In primates, more so than in other orders, foraging behaviour involves extensive eye-hand and hand-mouth coordination (Gibson 1986). This coordination either is not present at birth or is compromised by the ascendance of reflexes associated with clinging (Hines 1942; Ehr- lich 1974; Fragaszy, in press). Specific skills involved in capturing and processing particular foods, such as gleaning arthropods from leaf cavities (Boinski & Timm 1985, for squirrel mon- keys) or opening up enclosed foods (Fragaszy 1986, for capuchin monkeys, Cebus spp.), require further refinements of eye-hand coordination and timing acquired only through experience, perhaps in com- bination with social learning. As in other taxa, many other skills must also be acquired, including perceptual discrimination of prey from non-prey (cf. Chai 1986) and appropriate sites and tech- niques to search for food (cf. Greenberg 1987; Werner & Sherry 1987). Taken together, the tasks to be accomplished by young primates prior to nutritional independence require a minimum amount of physical maturation and individual practice. Weaning must fit into the general time-
OOO3-3472/89/030415+ 14$03.OO/O 0 1989 The Association for the Study of Animal Behaviour 415

416 Animal Behaviour, 37, 3
frame of acquisition of foraging and locomotor skills. Aside from the studies by Altmann and colleagues (Ahmann 1980; Altmann & Alberts 1987) on baboons, Papio cynocephalus, the precise intercalations of weaning and the assumption of independent foraging by young primates are not well-documented.
Social transmission of information about ac- ceptable and non-acceptable foods is one means widely available across vertebrate orders for reduc- ing the length of time it takes new individuals, particularly young animals, to acquire preference for certain foods and avoidance of others (Galef 1976). The effectiveness of social observation for transmission of avoidance of noxious foods has been demonstrated to vary considerably in adults of some primate species (e.g. Cambefort 1981) and we know of only one report of social facilitation of food selection in young monkeys relevant to their acquisition of preferences shown by other group members (Whitehead 1986). Certainly species with wide dietary variation across seasons and habitats would be likely to benefit from social transmission of food preferences. Squirrel monkeys fit this pattern of wide dietary variation (Boinski, in press a, b), and thus the extent of social influence on selection of foods in young squirrel monkeys is an interesting question.
The influence of social partners on the acqui- sition of instrumental behaviour is less well-docu- mented than the effect of perceptual discrimina- tions (Galef 1988). We were interested to see the extent to which proximity to others occurred during foraging, to gauge the opportunity for young squirrel monkeys to learn from observation of others about places to forage, what to eat, and how to capture or process foods.
METHODS
Site This report is based on the first long-term study
of a wild population of squirrel monkeys in which troop members were individually recognized (Boinski 1986). The study site was a Costa Rican preserve, Parque National Corcovado. Parque National Corcovado is located on the Peninsula de Osa of southeastern Costa Rica. The 2.5-h km2 research site is at Estacion Sirena (8”30’N, 83”35’W) and includes ‘primary’, high-graded (selectively logged), and second-growth forest
(Boinski & Scott 1988; Boinski, in press a; Boinski & Fowler, in press).
In Corcovado, squirrel monkeys eat a wide array of plant and animal material, including arthro- pods, berry-like fruits, nectar, frogs, anole lizards, and even bats. Most foods are typically small, unpredictable in location, and found and eaten by one individual (Boinski & Timm 1985; Boinski & Scott 1988). The relative abundance of food, especially arthropods, changed markedly over the study (Boinski, in press a, b; Boinski & Fowler, in press). Arthropod abundance in sweep samples of live foliage was low in November-December 1983, moderate in January-March, greatest in April- May, and declined from June-September 1984 to moderate levels. Fruits and flowers were available throughout the year, but like arthropods, were least abundant in November-December 1984.
Animals The data reported here were collected by Boinski
from November 1983 through September 1984. She had conducted 16 months of field work with Corcovado squirrel monkeys prior to this period. An animal dye designed for field use (Nyanza brand, Medford, Massachusetts, U.S.A.) was applied to the members of the study troop. All animals in this troop were individually recognized by December 1983. Troop size varied from 38 to 45 animals during the study. As a consequence of discontinuous birth distributions, ages in females could be distinguished up to 4 years, and those of males up to 5 or 6 years. The birth dates of the 14 infants born in 1984 were determined precisely and six survived until September 1984. Yearlings dur- ing the study were estimated to have been born in 1983, juveniles in 1982, and young adults and full adults, respectively, in 198 1 and 1980 birth seasons and earlier.
Data Collection Measures of behaviour on individuals in the
study troop were collected on at least 17, and usually more than 20, days each month using a focal-animal technique. The time of samples was biased toward the morning hours. The sequence of individuals selected closely approximated a ran- dom sample within periods of troop contact. No significant age or sex differences were found in the temporal distribution of foraging activities (Boinski, unpublished data). Infants were included

Boinski & Fragaszy: Foraging ontogeny 417
Table 1. Definitions of behaviour categories
Behaviour Definition
Rest Describes instances in which the animal does not change location and is not engaged in any food-related activity. This behaviour includes instances when infants are being carried by mothers
Locomotion Progressive movement, both in food-related and non-food-related activities. This behaviour and rest do not sum to 100% of an individual’s activities because rest does not include those instances in which an individual was stationary and engaged in food-related activities
Wander This is a subcategory of locomotion seen in infants only. Apparently non- purposive quadrupedal locomotion not associated with social play. The focus of attention and direction of movement change frequently and are unpredic- table to an observer. Limb movements often appear clumsy and/or exagger- ated. Head-cocking (Menzel 1980) often occurs
Food-related activity
Eat arthropods
Foliage-scraping
Eat fruits and flowers
Manual forage
Live foliage forage
Dead foliage
Fruit and flower forage
Visual search
Scored when the animal is concerned with food, such as eating, searching for, or processing food, whether the forager is immobile or not
Animal protein is ingested, including instances of foliage-scraping. Rare instances of eating non-arthropod prey are also included
An arthropod is gnawed, chewed, or licked from a substrate. A technique used for arthropods too small to grasp readily. A subset of ‘eat arthropods’
The ingestion of fruit pulp or chewing and licking flowers to obtain nectar. Petals, fruit skins and peels, and other associated plant structures were rarely ingested
Instances in which an individual manipulates a substrate in search of food or processes a food item. This is further subdivided by target: live foliage, dead foliage, and fruits and flowers
Live foliage is searched manually for arthropods
Dead leaves, vines, twigs and bark are searched manually for arthropods
Gleaning, holding and processing fruit and flowers
Look at a substrate, usually foliage, from less than 30 cm away, often systematically shifting the head. Visual search is restricted to scanning surfaces or materials not being held. If the material is held, it is called manual forage
as focal subjects after 3 1 March 1984, at which time surviving infants ranged in age from 14 to 57 days. One infant was born on 21 April 1984.
A focal sample was comprised of 58 instanta- neous individual samples (Altmann 1974) each hour at I-min intervals. An average of 72 I-min interval instantaneous samples were obtained from each member of the troop each month. Each instantaneous sample included, when applicable, the following data categories: (1) type of food eaten or held, (2) type of substrate investigated, (3) foraging method employed, (4) type of locomotor activity, (5) the identity of the nearest neighbour and its distance from the focal animal within a 5-m radius at l-m intervals, and (6) the distance
between a mother and her infant within a 5-m radius at l-m intervals. Visibility of a nearest neighbour or infant was not a function of distance from the focal animal because (1) the individuals in close proximity to a focal animal seldom changed positions markedly, and (2) the focal animal was monitored for a period of sufficient length to detect the entrance or exit of another within a 5-m radius. The behaviour categories were not all mutually exclusive (Table I). For example, an individual could simultaneously eat an arthropod and hold a flower while scanning vegetation for more arthro- pods. The resulting data set allows conclusions as to the proportional allocation of behaviour over time but not duration or rate.

418 Animal Behaviour, 37, 3
Analytical Techniques
The relationship of infants to mothers was examined by considering nursing and spacing. The behavioural samples of each mother were pooled into l-month (30 day) periods beginning from the birthdate of her 1984 infant and continuing until the end of the study or the death of the infant. These monthly samples were further subdivided by (1) the mother’s activity (whether she was engaged in a food-related activity or not) and (2) the infant’s distance from the mother. Within each behavioural category the distribution of samples as a function of the infant’s distance from the mother was calculated. The percentage of samples per month spent nursing by infants was calculated from the behavioural samples of both the mothers and infants.
Activities of the infant pertinent to the acqui- sition of foraging behaviour are described based on instantaneous samples and ad libitum observa- tions. The raw frequency counts of each behaviour were first transformed into a proportion of the total number of instantaneous samples for each indi- vidual during each time period. The relationships among relative frequency of a foraging behaviour (independent factor), age cohort (between-subject factor), and time period (a within-subject factor that reflects increasing infant age and seasonal differences in food availability) were analysed by repeated measures analyses of variance (ANOVAs) on the arcsine transformations of proportional measures. April-September 1984 was divided into three 2-month-long time periods. Individuals which were not represented by behavioural samples in all three time periods were excluded from this analysis. Initially, three age cohorts were employed: six infants, five yearlings, and three female and four male juveniles. Average ages of infants during the periods of behavioural sampling were: April-May, 78 days (SD = 2 1.3); June-July, 125 days (SD= 33.4); and August-September, 189 days (SD = 21.6). The yearling and juvenile cohorts were pooled into a subadult cohort because no significant differences between them were found by difference of means tests (Tukey’s studentized range test, Duncan’s multiple range test) in one- way ANOVAs on each behaviour in each time period. The data for subadults are presented graphically for the late wet (November-December 1983) and dry season (January-March 1984) to document more clearly the presence of seasonal
fluctuations in the frequencies of foraging beha- viour, but these data were not included in the analyses.
The data on the spacing of infants 1 m or less from others was analysed using a repeated measure ANOVA in a similar model. The distance of 1 m or less was chosen to reflect the opportunity for close observation by an infant of any food-related behaviour by the focal animal. This analysis differs from the previous ones in two ways. First, four age cohorts were employed: infants (N= 6). yearlings (N= 5) juvenile and adult females (N= 13) juve- niles and adult males (N = 11). Juveniles and adults were pooled by sex because analyses did not indicate significant differences between these age classes within sex in foraging behaviour. Second, instantaneous samples were assigned to one of two activity categories, food-related or not. The pro- portion of samples for which the individual had an infant as a nearest neighbour was determined separately for each category of activity.
RESULTS
Timing of Independence from Mother The spatial affiliation between mothers and
infants declined sharply with increasing infant age (Fig. 1). Infants were in nearly constant physical contact during the first month of life; from the fourth through the seventh month physical contact was negligible (less than 3% of the samples). During the breeding season, when the infants were about 8 months old, physical contact increased to nearly 5% of the mothers’ sample in non-food- related contexts because infants were apparently attracted to the mothers during sexual interactions. On average, an infant was within a 5-m radius of its mother during less than 10% of the mothers’ behavioural samples after the fourth month. Proxi- mity of the infant to its mother, especially in the contact category, was far less likely when the mother was engaged in food-related activities compared to other activities.
No food-sharing was ever observed by mothers or other troop members with infants, nor did infants obtain other forms of nurturance from troop members. Allomothering was a negligible contribution to infant care. During the first 2 months after the birth of their infants, mothers strongly resisted the persistent attempts of other females, especially juvenile females, to carry and
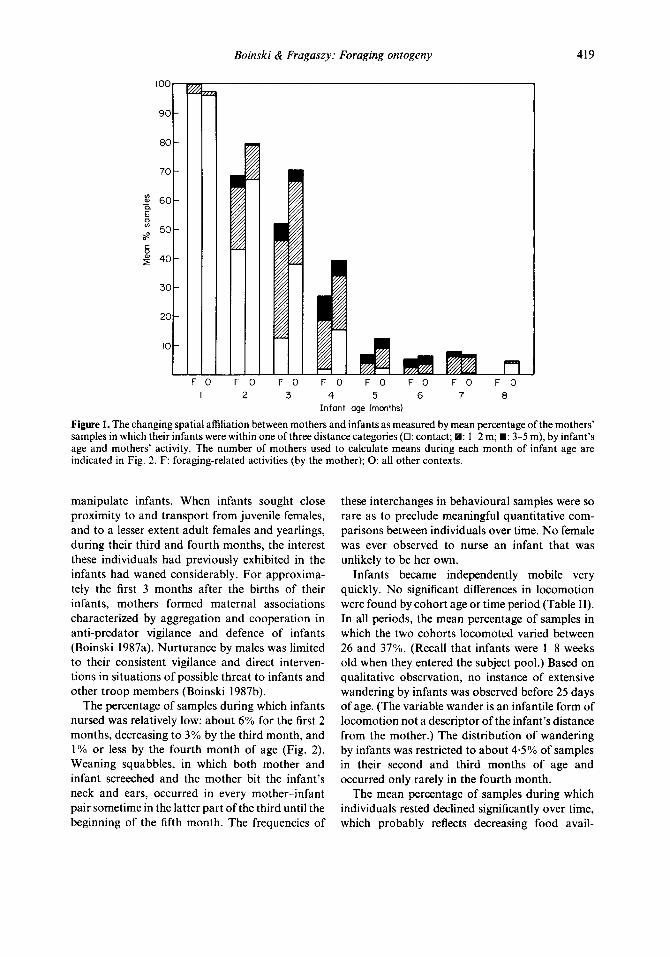
Boinski & Fragaszy: Foraging ontogeny 419
80-
2 60- E :: z 50-
6 g 40-
30-
20-
10 -
F 0 2
F 0
3 F 0 F 0 F 0 F 0
4 5 6 7 8 Infant age (months)
Figure 1. The changing spatial affiliation between mothers and infants as measured by mean percentage of the mothers’ samples in which their infants were within one of three distance categories (0: contact; : 1-2 m; n : 3-5 m), by infant’s age and mothers’ activity. The number of mothers used to calculate means during each month of infant age are indicated in Fig. 2. F: foraging-related activities (by the mother); 0: all other contexts.
manipulate infants. When infants sought close proximity to and transport from juvenile females, and to a lesser extent adult females and yearlings, during their third and fourth months, the interest these individuals had previously exhibited in the infants had waned considerably. For approxima- tely the first 3 months after the births of their infants, mothers formed maternal associations characterized by aggregation and cooperation in anti-predator vigilance and defence of infants (Boinski 1987a). Nurturance by males was limited to their consistent vigilance and direct interven- tions in situations of possible threat to infants and other troop members (Boinski 1987b).
The percentage of samples during which infants nursed was relatively low: about 6% for the first 2 months, decreasing to 3% by the third month, and 1% or less by the fourth month of age (Fig. 2). Weaning squabbles, in which both mother and infant screeched and the mother bit the infant’s neck and ears, occurred in every mother-infant pair sometime in the latter part of the third until the beginning of the fifth month. The frequencies of
these interchanges in behavioural samples were so rare as to preclude meaningful quantitative com- parisons between individuals over time. No female was ever observed to nurse an infant that was unlikely to be her own.
Infants became independently mobile very quickly. No significant differences in locomotion were found by cohort age or time period (Table II). In all periods, the mean percentage of samples in which the two cohorts locomoted varied between 26 and 37%. (Recall that infants were l-8 weeks old when they entered the subject pool.) Based on qualitative observation, no instance of extensive wandering by infants was observed before 25 days of age. (The variable wander is an infantile form of locomotion not a descriptor of the infant’s distance from the mother.) The distribution of wandering by infants was restricted to about 4.5% of samples in their second and third months of age and occurred only rarely in the fourth month.
The mean percentage of samples during which individuals rested declined significantly over time, which probably reflects decreasing food avail-
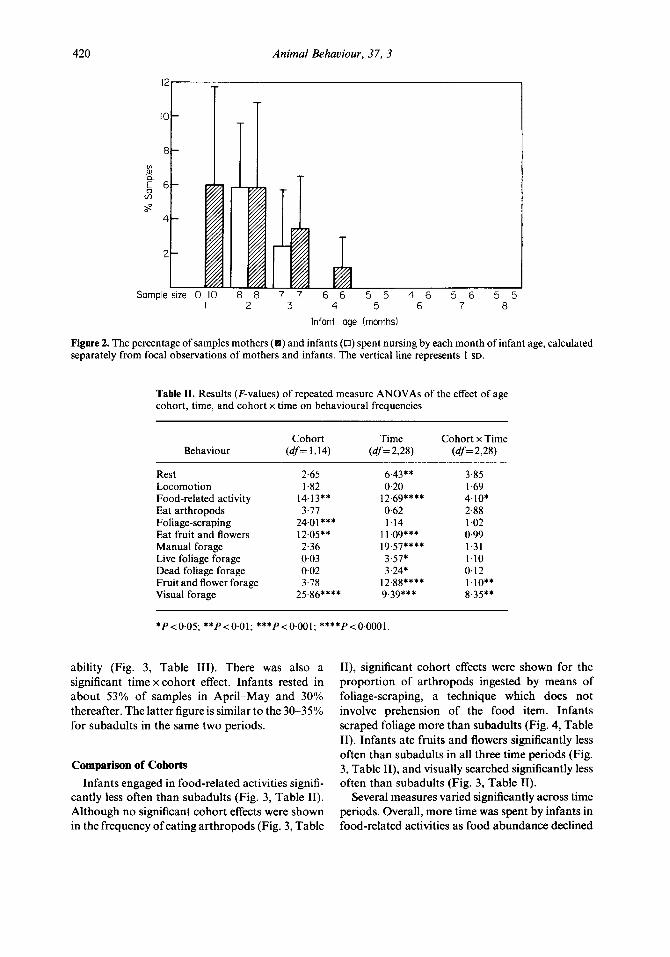
420 Animal Behaviour. 37, 3
Sample we 0 IO 8 8 7 7 6 6 5 5 4 6 5 6 5 5 I 2 3 4 5 6 7 8
Infant age (months)
Figure 2. The percentage of samples mothers ( ) and infants (0) spent nursing by each month of infant age, calculated separately from focal observations of mothers and infants. The vertical line represents 1 SD.
Table II. Results (F-values) of repeated measure ANOVAs of the effect of age cohort, time, and cohort x time on behavioural frequencies
Behaviour Cohort Time
(df= 1,14) (liy= 2,28) Cohort x Time
(df=2,28)
Rest 265 Locomotion 1.82 Food-related activity 14.13** Eat arthropods 3.77 Foliage-scraping 24.01*** Eat fruit and flowers 12.05** Manual forage 2.36 Live foliage forage 0.03 Dead foliage forage 0.02 Fruit and flower forage 3.78 Visual forage 25,86****
6.43** 3.85 0.20 1.69
12.69**** 4.10* 0.62 2.88 I.14 1.02
1 I .09*** 0.99 19,57**** 1.31
3,57* 1.10 3,24* 0.12
12,88**** 1.10** 9.39*** 8.35**
ability (Fig. 3, Table III). There was also a II), significant cohort effects were shown for the significant time x cohort effect. Infants rested in proportion of arthropods ingested by means of about 53% of samples in April-May and 30% foliage-scraping, a technique which does not thereafter. The latter figure is similar to the 30-35% involve prehension of the food item. Infants for subadults in the same two periods. scraped foliage more than subadults (Fig. 4, Table
II). Infants ate fruits and flowers significantly less
Comparison of Cohorts often than subadults in all three time periods (Fig. 3, Table II), and visually searched significantly less
Infants engaged in food-related activities signifi- often than subadults (Fig. 3, Table II). cantly less often than subadults (Fig. 3, Table II). Several measures varied significantly across time Although no significant cohort effects were shown periods. Overall, more time was spent by infants in in the frequency of eating arthropods (Fig. 3, Table food-related activities as food abundance declined
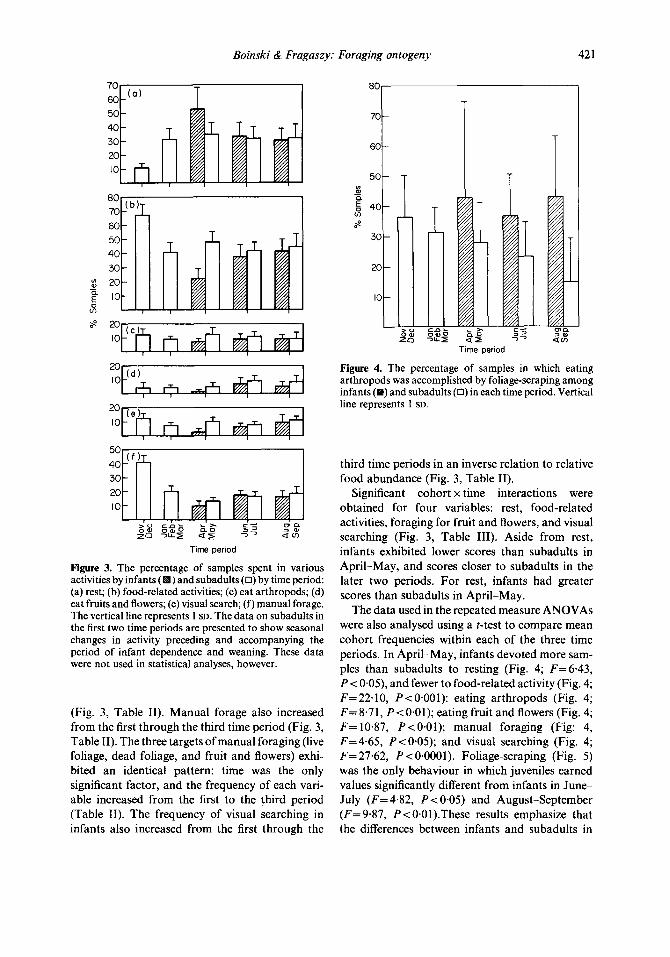
Boinski & Fragaszy: Foraging ontogeny 421
Time period
Figure 3. The percentage of samples spent in various ) and subadults (0) by time period:
(a) rest; (b) food-related activities; (c) eat arthropods; (d) eat fruits and flowers; (e) visual search; (f) manual forage. The vertical line represents 1 SD. The data on subadults in the first two time periods are presented to show seasonal changes in activity preceding and accompanying the period of infant dependence and weaning. These data were not used in statistical analyses, however.
(Fig. 3, Table II). Manual forage also increased from the first through the third time period (Fig. 3, Table II). The three targets ofmanual foraging (live foliage, dead foliage, and fruit and flowers) exhi- bited an identical pattern: time was the only significant factor, and the frequency of each vari- able increased from the first to the third period (Table II). The frequency of visual searching in infants also increased from the first through the
70-
60-
50- t
z g 40-
* .\”
30-
20-
IO -
p&i
Time period
Figure 4. The percentage of samples in which eating arthropods was accomplished by foliage-scraping among infants(S) and subadults (0) in each time period. Vertical line represents 1 SD.
third time periods in an inverse relation to relative food abundance (Fig. 3, Table II).
Significant cohort x time interactions were obtained for four variables: rest, food-related activities, foraging for fruit and flowers, and visual searching (Fig. 3, Table III). Aside from rest, infants exhibited lower scores than subadults in April-May, and scores closer to subadults in the later two periods. For rest, infants had greater scores than subadults in April-May.
The data used in the repeated measure ANOVAs were also analysed using a t-test to compare mean cohort frequencies within each of the three time periods. In April-May, infants devoted more sam- ples than subadults to resting (Fig. 4; F=6.43, P-C 0.05), and fewer to food-related activity (Fig. 4; F=22.10, PC 0.001): eating arthropods (Fig. 4; F= 8.71, P < 0.01); eating fruit and flowers (Fig. 4; F= 10.87, P~0.01); manual foraging (Fig: 4, F=4.65, PC O-05); and visual searching (Fig. 4; F= 27.62, P-C 0.0001). Foliage-scraping (Fig. 5) was the only behaviour in which juveniles earned values significantly different from infants in June- July (F= 4.82, P < 0.05) and August-September (F=9%7, P<O.Ol).These results emphasize that the differences between infants and subadults in
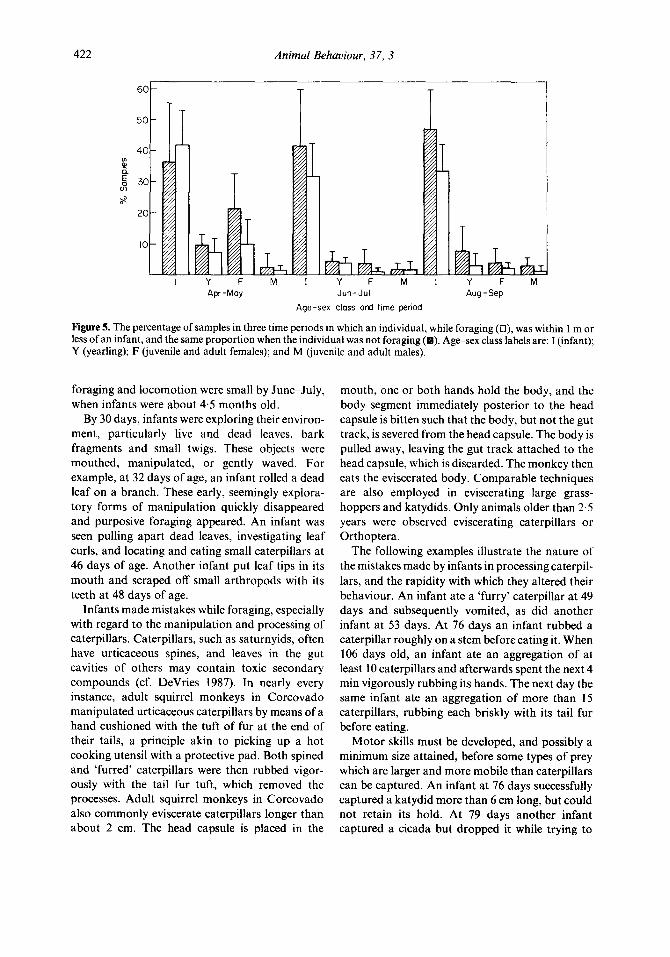
422 Animal Behaviour, 37, 3
60
I Y F M I Y F M I Y F M Apr-May Jun- Jul
Age-sex class and time period
Aug - Sep
Figure 5. The percentage of samples in three time periods in which an individual, while foraging (o), was within I m or less of an infant, and the same proportion when the individual was not foraging ( ). Age-sex class lab& are: 1 (infant); Y (yearling); F (juvenile and adult females); and M (juvenile and adult males).
foraging and locomotion were small by June-July, when infants were about 4.5 months old.
By 30 days, infants were exploring their environ- ment, particularly live and dead leaves, bark fragments and small twigs. These objects were mouthed, manipulated, or gently waved. For example, at 32 days of age, an infant rolled a dead leaf on a branch. These early, seemingly explora- tory forms of manipulation quickly disappeared and purposive foraging appeared. An infant was seen pulling apart dead leaves, investigating leaf curls, and locating and eating small caterpillars at 46 days of age. Another infant put leaf tips in its mouth and scraped off small arthropods with its teeth at 48 days of age.
Infants made mistakes while foraging, especially with regard to the manipulation and processing of caterpillars. Caterpillars, such as saturnyids, often have urticaceous spines, and leaves in the gut cavities of others may contain toxic secondary compounds (cf. DeVries 1987). In nearly every instance, adult squirrel monkeys in Corcovado manipulated urticaceous caterpillars by means of a hand cushioned with the tuft of fur at the end of their tails, a principle akin to picking up a hot cooking utensil with a protective pad. Both spined and ‘furred’ caterpillars were then rubbed vigor- ously with the tail fur tuft, which removed the processes. Adult squirrel monkeys in Corcovado also commonly eviscerate caterpillars longer than about 2 cm. The head capsule is placed in the
mouth, one or both hands hold the body, and the body segment immediately posterior to the head capsule is bitten such that the body, but not the gut track, is severed from the head capsule. The body is pulled away, leaving the gut track attached to the head capsule, which is discarded. The monkey then eats the eviscerated body. Comparable techniques are also employed in eviscerating large grass- hoppers and katydids. Only animals older than 2.5 years were observed eviscerating caterpillars or Orthoptera.
The following examples illustrate the nature of the mistakes made by infants in processing caterpil- lars, and the rapidity with which they altered their behaviour. An infant ate a ‘furry’ caterpillar at 49 days and subsequently vomited, as did another infant at 53 days. At 76 days an infant rubbed a caterpillar roughly on a stem before eating it. When 106 days old, an infant ate an aggregation of at least 10 caterpillars and afterwards spent the next 4 min vigorously rubbing its hands. The next day the same infant ate an aggregation of more than 15 caterpillars, rubbing each briskly with its tail fur before eating.
Motor skills must be developed, and possibly a minimum size attained, before some types of prey which are larger and more mobile than caterpillars can be captured. An infant at 76 days successfully captured a katydid more than 6 cm long, but could not retain its hold. At 79 days another infant captured a cicada but dropped it while trying to

Boinski & Fragaszy: Foraging ontogeny 423
bite its head. Afrer running directly to a 5-cm-long grasshopper on the ground from a position 7 m away, an infant at 126 days pounced more than six times, always missing its prey. An infant at 143 days captured a 6-cm Agalychnis callidryas tree frog, and recaptured it after the frog struggled free, only to lose hold again.
Social Context of Foraging
Infants were far more likely to have another infant as a neighbour at a radius of 1 m or less than to be near members of other age-sex classes (F=6&27, df=3,31, P<O.OOOl; Fig. 5). Only in April-May did another age-sex class, females, have infants as neighbours in more than 10% of samples. Infants were significantly less likely to be close to another individual when the other indi- vidual was engaged in food-related activities than if it was engaged in other activities (F=35.75, df = 1,3 1, P < 0.0001). During their third month of life, infants were within a l-m radius of their mothers in less than 15% of the mothers’ beha- vioural samples (Fig. 1). In short, there is little evidence that infants are spending much time monitoring experienced foragers, even their mothers, at close proximity. Infants and subadults occasionally approached adults eating large items (e.g. when eating large butterflies or bats; Boinski & Timm 1985) but they approached other infants at least as often under the same circumstances. For example, at 124 days of age an infant ran over 7 m to watch another infant eat a large grasshopper, eventually observing from less than 15 cm away.
There is, however, some anecdotal evidence that opportunities occur for social transmission of selective avoidance of hazardous caterpillars. At 153 days of age an infant touched a 6-cm-long saturnyid caterpillar, and subsequently shrieked and rubbed its hands vigorously. Another infant, 121 days old, thereupon approached the infant which had shrieked, stared at the caterpillar (its nose was less than 5 cm away), but did not touch the apparently toxic caterpillar. Other examples involve adults apparently warning infants to avoid noxious animals. A striking example occurred on 12 June 1984, when the majority of the troop used the same branch to cross a forest gap. An 8-cm- long black saturnyid caterpillar lay unobscured on a leaf adjacent to the travel path. At least eight adults came to a full stop in front of the leaf bearing the caterpillar before continuing. When a group of
four infants also paused by the caterpillar an adult male returned, gave several bark vocalizations (Newman 1985), and placed himself between the infants and the caterpillar. The infants then looked closely at the caterpillar again before continuing. Three other similar episodes of adults of both sexes apparently warning infants about urticaceous caterpillars were also observed. Adults used the same intervening behaviour and bark vocalizations with regard to other potentially hazardous animals encountered by infants and juveniles, including Didelphis and Marmosa opposums, snakes Boa constrictor and Bothrops asper and owls, Lophos- trix cristata.
DISCUSSION
The data from this study increase the scarce information on the acquisition of nutritional inde- pendence by a social mammal in its natural habitat, in this case an insectivorous and frugivorous primate, the squirrel monkey. We first compare physical and motor development of captive squirrel monkeys with the data reported here, and discuss its relation to weaning. We then consider the development of foraging in Saimiri oerstedi, and make inferences about learning and other processes supporting skill development. It must be stressed that the comparisons with captive squirrel mon- keys involve species of Saimiri other than S. oerstedi (Hershkovitz 1984; but see Thorington 1985). In captivity, the body weights of Saimiri species differ, but growth trajectories are parallel (Kaack et al. 1979; Kaplan 1979). Thus, we feel that the comparisons are a better indicator of the influence of the environment on behaviour than of species differences. Until S. oerstedi is studied in captivity this remains a conjecture.
Infant squirrel monkeys in Corcovado loco- moted independently by the end of 4 weeks of age, foraged successfully at 7 weeks, and were weaned by 16 weeks. Weaning occurred over the course of only 6 weeks. By 4.5 months of age most of the foraging patterns of infants were indistinguishable from those of older individuals. The early timing of weaning would not be predicted from laboratory studies of squirrel monkeys. In captivity, although independent locomotion begins by the second month of life, weaning is not terminated until 6 or 7 months, and eating solid food only becomes com- monplace in the fourth month (Rosenblum 1968)

424 Animal Behaviour, 37, 3
an age when Corcovado squirrel monkeys were already weaned. The Corcovado data are also surprising when considered in the context of the usual pattern of weaning in primates. In general, the period between the age the infant initially tries solid food and the termination of weaning is long and the transition to solid food gradual (Pereira & Altmann 1985). Infants often continue to nurse at least occasionally until the latter part of the mothers’ next pregnancy (Pereira & Altmann 1985).
The match in behavioural development between the infant squirrel monkeys at Corcovado and in several laboratory studies is consistently close. In captive settings, squirrel monkeys, Saimirisciureus, dismount from the mother and walk in the third or fourth week (Rosenblum 1968; Elias 1977; Fra- gaszy, unpublished data), in good agreement with the field data on early locomotion (called ‘wander’ here). In both wild and captive infants, prior episodes of visually fixating the hand while on the mother’s back precede dismounting and accom- pany early exploratory contacts of the surfaces in the environment (Boinski, personal observation; Fragaszy, personal observation). Captive and wild infants both display the classic hand-to-mouth pattern, placing the fist in the mouth after moving the arm and regarding the hand. In captivity, the earliest bouts of mouthing solid food occur in the second month; in Corcovado, episodes of eating solid foods were first observed in the seventh week. Nursery-reared infants first walked to and grasped a small object at 7 weeks, and uncovered a hidden object at 12 weeks (displaying memory skills; Elias 1977). Exaggerated visual inspection during early bouts of independent locomotion occurred in the second and third months, exactly overlapping with the peak in the curve in Menzel’s (1980) data on the occurrence of head-cocking in two captive infant squirrel monkeys. Head-cocking is a behaviour which probably aids visual acuity in small monkeys (Menzel 1980). Its sharp increase at about 6-8 weeks and subsequent decline seem well matched with the neural maturation reflected by the comple- tion by 8 weeks of the most rapid phase of post- natal growth of the brain (Bauchot & Manocha 1981). Episodes of capturing, or attempting to capture, mobile prey, were first observed in Corco- vado infants at 11 weeks. Weaning fights began at this age and continued to about 16 weeks. This matches Rosenblum’s (1968) observation that cap- tive infants routinely eat solids by the fourth
month, soon after dentition is complete (Long & Cooper 1968). However, captive infants are not weaned until about 6 months, or even older.
In sum, we see that Corcovado infants ended nursing sooner than infants in captivity by about 2 months, reducing the overlap in nursing and independent locomotion from about 5 months (the second through sixth month) to less than 3 months (6 weeks through 4 months). Infants in the wild were weaned after deciduous dentition was com- plete, brain growth slowed, locomotor skills were well developed, visual acuity had reached greater maturity (as reflected in reduced head-cocking), and memory for hidden objects was in place. Thus, it is also likely that infants in captivity were also capable, physically and behaviourally, of foraging for themselves by 16 weeks of age.
Weaning is probably delayed or extended in captive monkeys compared with Corcovado squir- rel monkeys because of different energetic and time-budget constraints in the two contexts. In an allometric plot of weaning age versus maternal weight in 23 primate taxa (Schwartz & Rosenblum 1983) the calculated point for S. sciureus was above the line. When we calculated the point for S. oerstedi (using 16 weeks as weaning age and body weights obtained from captive animals in Costa Rica, Boinski, unpublished data), the point fell directly on the regression line. This, together with the capacity for independent foraging indicated at 16 weeks, strongly suggests that mothers in cap- tivity are delaying weaning. For these mothers, with access to food ad libitum, the relative energetic costs of lactation are certainly minimized.
In baboons, early nursing conflicts are not instigated to limit nursing directly, but instead to schedule nursing to accommodate better the female’s own foraging and maintenance activities (Altmann 1980). Izard (1987) reported that Galago nurse their infants at night when daytime nursing has been eliminated. However, we do not believe that S. oerstedi mothers are merely restricting the timing of nursing to night for infants older than 16 weeks. The positions of troop members in their usual sleeping site, the fronds of an Astrocarvum alutum palm, were periodically surveyed at night with powerful lights. By 2 June 1984 (when infants ranged from 42 to 118 days in age), nearly all infants were consistently observed closely huddled in sleeping groups separate from adult females (Boinski, personal observation). In any case, the squirrel monkey data from both captivity and the

Boinski & Fragaszy: Foraging ontogeny
field are consistent with a model of behavioural development positing a relatively invariant begin- ning, but a flexible endpoint (Hines 1942; Pereira & Altmann 1985).
Young squirrel monkeys at Corcovado assumed self-sufficient foraging within 8-10 weeks after the onset of independent locomotion. This finding suggests that the essential skills used by squirrel monkeys do not require individual practice over a long period prior to adequate peformance. This in itself is an interesting result, as it contrasts with the general pattern in primates noted by Altmann & Alberts (1987) of slow acquisition of foraging skills and gradual weaning. However, certain aspects of foraging do appear to require specific experience for refinement. It also seems probable that continu- ing physical maturation through the first 6 or 7 months of life influences fine motor coordination and perhaps visual acuity. We consider below what young squirrel monkeys may be learning and how such learning is mediated.
as it seems to vary with species (Cambefort 1981). In any case, young squirrel monkeys were clearly more overtly attentive to the foraging activities of others, especially with larger items, than were older monkeys.
Specific experience contributes to foraging beha- viour in several domains of knowledge, which can be paraphrased as knowing which, knowing where, and knowing how. The first two involve perceptual discrimination, and are reflected in recognition of which species and parts of items to eat and which to avoid, and selection of appropriate places to search for food. The last type of knowledge involves motor skills, such as specialized methods of hand- ling urticaceous caterpillars or eviscerating inverte- brates. Observation of others appears to facilitate acquisition of perceptual knowledge, most clearly concerning what to avoid. Infant squirrel monkeys received potentially meaningful ‘warnings’ from adults concerning noxious prey, as they did con- cerning potentially dangerous animals. Although we do not have data to evaluate the influence of such ‘warnings’ on subsequent behaviour towards these objects, some influence is probable. Mineka & Cook (1988) have shown that negative affective responses (e:.g. fear) are readily transmitted socially in rhesus monkeys Macaca mulatta; expression of affect by the demonstrator facilitates observational learning in humans, too (Bandura 1986). In general, most of the literature on social learning in many orders (reviewed in Galef 1988) is consistent with the conclusion that observation of the affec- tive response of others can lead readily to shared interest in, or avoidance of, specific places or objects. It is unknown if sensitivity to the ex- pression of affective state by others varies with age,
Differential selectivity of foraging sites or fruits (a form of knowing which and where) is probably not directly influenced by observation of others, but rather indirectly through proximity to others. Infants were exposed to areas in which others foraged as a consequence of movement with the troop. Nearly all foods exploited by these monkeys, from both animal and plant sources, are indivi- dually located, harvested and processed. The mon- keys rarely foraged as a group in large fruit trees for many minutes at a time (Boinski, in press a), as do howler monkeys (Alouatta; Glander 1981; White- head, in press). Social transmission of preferences for leaves of certain plant species and avoidance of others (Whitehead 1986) is both more likely, as a result of exposure, and more relevant to efficient foraging in howler monkeys than it is in squirrel monkeys. Leaves and unripe fruit are more likely to contain toxins than ripe fruits (Hladik & Hladik 1969; Oates et al. 1980), and squirrel monkeys clearly prefer ripe fruit over less-ripe fruit (Boinski 1986). Additionally, during the weaning period fruit consumption is relatively low, and inverte- brate consumption at an annual high (Boinski, in press b). Thus, social influences on selection of fruits, if they occur, are more probable sometime after weaning.
In contrast to perceptual knowledge, ‘knowing how’ must be learned individually. Social facili- tation of motor acts or learning by imitation are much less frequent (indeed rare) in non-humans than enhancement of interest or avoidance (Galef 1988). There was no evidence of direct social influence on motor acts used in foraging in squirrel monkeys. Specialized handling techniques for both plant and animal foods appeared to be learned through direct experience: irritation from urtica- ceous caterpillars, nausea and loss of mobile prey.
The social context of foraging appears to be quite different in squirrel monkeys than in the Callitrichidae, Callicebus, Cebus, or Alouatta. In all these taxa, either overt sharing of food between infant and adult occurs (Callitrichidae, Callicebus; Brown & Mack 1978; Starin 1978; Fragaszy et al. 1982) or adult tolerance of infants and interest by infants in adult foraging is marked and sustained (Alouatta, Cebus; Whitehead 1986; Fragaszy &
425

426 Animal Behaviour, 37,3
Adams-Curtis, unpublished data). In the case of callitrichids and Callicebus, a spatially cohesive and intensively affiliative family unit supports social influence on activity in many forms (Menzel & Juno 1982; Fragaszy & Mason 1983). In Cebus, the behaviour of individual members of the group is generally less synchronized than in Alouatta, Callicebus, or Callitrichidae. However, individuals engage in long bouts of exploitation of single items which require extensive processing (e.g. palm pith or snails), and they also exploit large fruiting trees in subgroups (Robinson 1981; Fragaszy, unpub- lished data). In contrast, in Corcovado, a squirrel monkey troop with 40 members is typically dis- persed over l-2 ha, exhibits little coordination among troop members in foods harvested or foraging methods employed, and spatial distances between animals are exacerbated in foraging com- pared with non-foraging contexts, probably because of indirect food competition within troops (Boinski, in press a, b). If an experienced forager tolerated an infant closely observing its foraging behaviour or sharing foraging sites, its foraging efficiency would most probably be compromised. Even in the closer confines of captive settings, adult Saimiri exhibit little to no coordination among members of a group or pair (Fragaszy & Mason 1978, 1983; Fragaszy 1979). All these character- istics would lead to the reduced importance of social influence on foraging activities and the acquisition of foraging behaviour.
Certain sensorimotor limitations may have influenced the foraging activities of infants relative to juveniles. Infants devoted the same proportion of samples to foraging for invertebrates as did older monkeys, but infants used the less efficient method of scraping leaves more than did juveniles. Arthro- pods obtained in this way are generally very small (Boinski 1986). However, this technique requires neither prehension of the prey nor visual fixation of small prey. Thus, it is the easiest technique, in terms of sensorimotor requirements, of obtaining inver- tebrate prey, other than simply picking up large, essentially sessile, surface arthropods, such as caterpillars.
Subadults exhibited greater proportions of visual investigation than did infants, as well as lesser proportions of scraping. This difference in the distribution of foraging activities suggests that juveniles are probably more efficient at selecting sites in which to forage manually. Nearly all bouts of scraping by infants and juveniles probably
resulted in the ingestion of food, as indicated by either direct identification of the food taken, or by the observation that the subject chewed and swal- lowed material. However, the volume and quality of foods ingested by scraping were unknown, and infants may not have obtained more food by scraping than did subadults, even though they devoted more effort to it.
Young squirrel monkeys displayed naivety about foraging in various ways besides inefficient searching and inept handling of certain prey. For example, the open-mouthed displays of the lizard Corytophanes cristatus apparently intimidated infant squirrel monkeys attempting to capture them, although the same display was ignored by adults (Boinski, personal observation). Nearly all skins, peels and seeds larger than I.5 mm are removed from fruit by mature squirrel monkeys prior to ingestion (Boinski 1986), which probably results in more protein and other nutrients per mass ingested than unprocessed fruits (Clark 1980). Judging by faecal samples, infants were much less thorough in separating fruit pulp from skins than were adults (Boinski, personal observation), and thus less efficient at foraging these fruits.
To summarize, infant squirrel monkeys in Cor- covado were weaned earlier and more abruptly than would be predicted from captive studies. However, in all other ways, behavioural develop- ment in wild and captive infants was similar. lnvariance in the initial stages presumably reflects constraints imposed by neuromuscular and skeletal development of the infant. Variation in the timing of weaning, however, probably reflects the relative cost of lactation for the mother. For the 2 months after weaning, the infants in Corcovado still lagged behind older animals in those skills requiring advanced motor and visual coordination. It appeared that individual experience was important in the acquisition of the finer points of foraging, and that social transmission was relatively unim- portant except (perhaps) for learned avoidance and the exposure to potential foods and foraging sites because of group movement patterns.
ACKNOWLEDGMENTS
We wish to thank the Servicio de Parques Nacio- nales of Costa Rica for making research in Corco- vado possible. The data were collected while SB was a graduate student in the Department of

Boinski 8 Fragaszy: Foraging ontogeny
Zoology at The University of Texas at Austin. The field work was supported by Sigma Xi, Theodore Roosevelt Fund, American Museum of Natural History, the World Worldlife Fund-U.S.-Primate Action Fund, a NSF Dissertation Improvement Grant, and a National Science Foundation Grant to L. E. Gilbert. During the period that the data were analysed and the manuscript prepared, SB was supported by a National Research Service Award. DMF was supported by Research Scientist Development Award MH-00694 from the Natio- nal Institutes of Mental Health during the prep- aration of this manuscript. We thank M. Biben, J. D. Newman, S. Suomi, E. Visalberghi and J. M. Whitehead for helpful comments on an earlier draft of this manuscript.
REFERENCES
Altmann, J. 1974. Observational study of behaviour: samnling methods, Behaviour, 69.2277267.
Altmann, j. 1980. Baboon Mothers and Infants. Cam- bridge, Massachusetts: Harvard University Press.
Altmann, J. & Alberts, S. 1987. Body mass and growth rates in a wild primate population. Oecologia (Berl.), 72, 15-20.
Bandura, A. 1986. The Social Foundation of Thought and Action. New York: Prentice-Hall.
Bauchot, R. & Manocha, S. 1981. La croissance encepha- lique chez Saimiri sciureus (Simiens, Cebides). Mam- malia, 45, 251-255.
Boinski, S. X986. The ecology of squirrel monkeys in Costa Rica. Ph.D. thesis, The University of Texas at Austin.
Boinski, S. 1987a. Birth synchrony in squirrel monkeys (Saimiri oerstedi): a strategy to reduce neonatal preda- tion. Behaa. Ecol. Sociobiol., 21. 393-400.
Boinski, S. 1987b. Mating patterns in squirrel monkeys: implications for seasonal sexual dimorphism. Behav. Ecol. Sociobiol., 21, 13-21.
Boinski, S. In press a. Habitat use by Costa Rican squirrel monkeys (Saimiri oerstedi). Folia primatol.
Boinski, S. In press b. Sex differences in the foraging behavior of squirrel monkeys in a seasonal habitat. Behav. Ecol. Sociobiol.
Boinski, S. & Fowler, N. L. In press. Seasonal patterns in a tropical lowland forest. Biotropica.
Boinski. S. &Scott. P. E. 1988. Association between birds and monkeys in Costa Rica. Biotropica, 20, 136-143.
Boinski, S. 6r Timm, R. M. 1985. Predation by squirrel monkeys <and double-toothed kites on tent-making bats. Am. J. Primatol., 9, 121-128.
Brown, K. & Mack, D. S. 1978. Food sharing among captive Leontopithecus rosalia. Foliaprimatol., 29,268- 290.
Cambefort, 3. P. 1981. A comparative study of culturally transmitted patterns of feeding in the chacma baboon (Papio ursinus) and the vervet monkey (Cercopithecus aethiops). Folia primatol., 36, 243-263.
427
Chai, P. 1986. Field observations and feeding experi- ments on the responses of rufous-tailed jacamars (Galbula rujicauda) to free-flying butterflies in a tropi- cal rainforest. Biol. J. Linn. Sot., 29, 161-189.
Clark, D. A. 1980. Age- and sex-dependent foraging strategies of a small mammalian omnivore. J. Anim. Ecol., 49, 549-564.
DeVries, P. J. 1987. Butterflies of Costa Rica and their Nafural History. Princeton: Princeton University Press.
Ehrlich, A. 1974. Infant development in two prosimian species, greater galago and slow loris. Devl Psychobiol., 7,439-454.
Elias, M. 1977. Relative maturity of cebus and squirrel monkeys at birth and during infancy. Devl Psychobiol., 10,5 19-528.
Fragaszy, D. M. 1979. Squirrel and titi monkeys in a novel environment. In: Captivity and Behavior (Ed. by J. Erwin, T. Maple & G. Mitchell), pp. 172-216. New York: Holt, Rhinehart & Winston.
Fragaszy, D. M. 1986. Activity budgets and foraging behavior in wedge-capped capuchins (Cebus oliva- ceous): age and sex differences. In: Current Perspectives in Primate Social Dynamics (Ed. by D. Taub & F. King), pp. 1599174. New York: Van Nostrand.
Fragaszy, D M. In press. Activity states and motor activity in an infant capuchin monkey (Cebus apella) from birth through eleven weeks. Devl Psychol.
Fragaszy, D M. & Mason, W. 1978. Response to novelty in Saimiri and Callicebus: influence of social context. Primates, 19, 31 t-331.
Fragaszy, D. M. &Mason, W. 1983. Comparative studies of feeding behavior in squirrel and titi monkeys. J. camp. Psychol., 97, 310-326.
Fragaszy, D. M., Schwartz, S. & Shimosaka, D. 1982. Longitudinal observations of infant care and develop- ment in Callicebus moloch. Am. J. Primatol., 2, 191- 200.
Galef, B. 1976. Social transmission of acquired behavior: a discussion of tradition and social learning in verte- brates In: Advances in the Study of Behavior. Vol. 6 (Ed. by J. S. Rosenblatt, R. Hinde, E. Shaw & C. Beer), pp. 77-100. New York: Academic Press.
Galef, B. 1988. Imitation in animals: history, definition, and interpretation of data from the psychological laboratory. In: Social Learning: Psychological and Biological Perspectives (Ed. by T. Zentall & B. Galef), pp. 3328. Hillsdale: Erlbaum.
Gibson, K. 1986. Cognition, brain size, and theextraction of embedded food resources. In: Primate Ontogeny, Cognition, andSocial Behavior (Ed. by J. Else & P. Lee), pp. 93-103. Cambridge: Cambridge University Press.
Glander, K. E. 1981. Feeding patterns in mantled howling monkeys. In: Foraging Behavior (Ed. by A. Kamil & T. Sargent), pp. 231-257. New York: Gar- land.
Greenberg, R. 1987. Development of dead leaf foraging in a tropical migrant warbler. Ecology, 68, 130-141.
Hershkovitz, P. 1984. Taxonomy of squirrel monkeys genus Saimiri (Cebidae, Platrryhini): a preliminary report with a description of a hitherto unnamed form. Am. J. Primatol., 7, 1555210.
Hines, M. 1942. The development and regression of

428 Animal Beh, aviour, 37, 3
reflexes, postures and progression in the young maca- que. Contr. Embryol. Carneg. Inst., l%, 155-209.
Hladik, A. & Hladik, C. M. 1969. Rapports trophiques entre vegetation et primates dans la foret de Barro Colorado (Panama). Terre Vie, 1, 255117.
Izard, M. K. 1987. Lactation length in three species of Galago. Am. J. Primatol., 13, 73-76.
Kaack. B.. Walker. L. St Brizzee. K. R. 1979. The growth and ‘development of (Saimiri sciureus). GrowTh, 43, 116-135.
Kaplan, .I. N. 1979. Growth and development of infant squirrel monkeys during the first six months of life. In: Nursery Care of Nonhuman Primates (Ed. by G. C. Ruppenthal), pp. 153-164. New York: Plenum Press.
Long, J. 0. & Cooper, R. W. 1968. Physical growth and dental eruption in captive-bred squirrel monkeys, Saimiri sciureus (Leticia, Colombia). In: The Squirrel Monkey (Ed. by L. A. Rosenblum & R. W. Cooper), pp. 193-205. New York: Academic Press.
Menzel, C. R. 1980. Head-cocking and visual perception in primates. Anim. Behav., 28, 151-159.
Menzel, E. W. &Juno, C. 1982. Marmosets: are learning sets learned? Science, N. Y., 217, 750-752.
Mineka, S. & Cook, M. 1988. Social conditioning of fear in rhesus monkeys. In: Social Learning: Psychological and Biological Perspectives (Ed. by T. Zentall & B. Galef), pp. 51-73. Hillsdale: Erlbaum.
Newman, J. D. 1985. Squirrel monkey communication. In: Handbook of Squirrel Monkey Research (Ed. by L. A. Rosenblum & C. L. Coe), pp. 99-126. New York: Plenum Press.
Oates, J. F., Waterman, P. G. & Choo, G. M. 1980. Food selection by the South Indian leaf-monkey, Presbytis johnii, in relation to leaf chemistry. Oecologia (Berl.) ., 45, 56.
Pereira, M. E. & Altmann, J. 1985. Development of social behavior in free-living nonhuman primates. In: Nonhu- man Primate Models for Human Growth and Develop-
ment (Ed. by E. S. Watts), pp. 217-309. New York: Alan R. Liss.
Robinson, J. G. 198 1. Spatial structure in foraging groups of wedge-capped capuchin monkeys Cebus nigrivatta- tus. Anim. Behav., 29, 1036-1056.
Rosenblum, L. A. 1968. Mother-infant relations and early behavioral development in the squirrel monkey. In: The SquirrelMonkey (Ed. by L. A. Rosenblum & R. W. Cooper), pp. 2077233. New York: Academic Press.
Schwartz, G. G. & Rosenblum, L. A. 1983. Allometric influences on primate mothers and infants. In: Symbio- sis in Parent-Ofspring Interactions (Ed. by L. A. Rosenblum & H. Moltz), pp. 215-248. New York: Plenum Press.
Starin, E. D. 1978. Food transfer by wild titi monkeys (Callicebus torquatus torquatus). Folia primatol., 30, 1455151.
Thorington, R. W., Jr. 1985. The taxonomy and distribu- tion of squirrel monkeys (Saimiri). In: The Squirrel Monkey (Ed. by L. A. Rosenblum & C. L. Coe), pp. l- 34. New York: Plenum Press.
Werner, T. K. & Sherry, T. W. 1987. Behavioral feeding specializations in Pinaroloxias inornata, the ‘Darwin’s Finch’ of Cocos Island, Costa Rica. Proc. natn. Acad. Sri. U.S.A.,84, 5506-5510.
Whitehead, J. M. 1986. Development of feeding selecti- vity in mantled howling monkeys, Alouattapalliata. In: Primate Ontogeny, Cognition and Social Behaviour (Ed. by J. G. Else & P. C. Lee), pp. 1055117. Cambridge: Cambridge University Press..
Whitehead. J. M. In press. The consequences of the location of a simulated intruder for responses to long distance vocalizations of mantled howling monkeys, Alouatta palliata palliata. Behaviour.
(Received 7 January 1988: revised 25 April 1988; MS. number: A 5207)
Copyright © 2022 FDOKUMEN




















