
Auditory stimuli elicit hippocampal neuronal responses during sleep

Ž .Brain Research Reviews 32 2000 57–71www.elsevier.comrlocaterbres
Short review
Temporal expression of neuronal connexins during hippocampalontogeny
R. Rozental a,b,f,), M. Srinivas a, S. Gokhan c, M. Urban a, R. Dermietzel d, J.A. Kessler a,c,¨D.C. Spray a,e,f, M.F. Mehler a,c,f,g
a Department of Neuroscience, Albert Einstein College of Medicine, 1300 Morris Park AÕenue, Bronx, NY 10461, USAb Department of Anesthesiology, Albert Einstein College of Medicine, 1300 Morris Park AÕenue, Bronx, NY 10461, USA
c Department of Neurology, Albert Einstein College of Medicine, 1300 Morris Park AÕenue, Bronx, NY 10461, USAd Ruhr UniÕersity Bochum, D-44780 Bochum, Germany
e Department of Medicine, Albert Einstein College of Medicine, 1300 Morris Park AÕenue, Bronx, NY 10461, USAf Departments of Internal Medicine, Vision Institute and IPTESP, Federal UniÕersity of Goias, Goiania 74000, Brazil´ ˆg Department of Psychiatry, Albert Einstein College of Medicine, 1300 Morris Park AÕenue, Bronx, NY 10461, USA
Abstract
Communication through gap junction channels provides a major signaling mechanism during early brain histogenesis, a developmentaltime during which neural progenitor cells are inexcitable and do not express ligand-gated channel responses to the major CNSneurotransmitters. Expression of different gap junction types during neurogenesis may therefore define intercellular pathways for
Ž .transmission of developmentally relevant molecules. To better understand the molecular mechanism s by which growth and differentia-tion of neurons are modulated by gap junction channels, we have been examining the developmental effects of a specific set of cytokineson differentiation and gap junction expression in a conditionally immortalized mouse embryonic hippocampal neuronal progenitor cell
Ž .line MK31 . When multipotent MK31 cells are in an uncommitted state, they uniformly express the neuroepithelial intermediate filamentŽ . Ž .class VI marker, nestin, are strongly coupled by gap junctions composed of connexin43 Cx43 and express connexin45 Cx45 at the
mRNA level. As these cells undergo neuronal lineage commitment and exit from cell cycle, they begin to express the early neurofilamentmarker, NF66, and coupling strength and expression of Cx43 begin to decline with concurrent expression of other connexin proteins,including Cx26, Cx33, Cx36, Cx40 and Cx45. Terminal neuronal differentiation is heralded by the expression of more advancedneurofilament proteins, increased morphologic maturation, the elaboration of inward currents and action potentials that possess maturephysiological properties, and changing profiles of expression of connexin subtypes, including upregulation of Cx36 expression. Theseimportant developmental transitions are regulated by a complex network of cell cycle checkpoints. To begin to examine the precise rolesof gap junction proteins in traversing these developmental checkpoints and in thus regulating neurogenesis, we have focused on individual
Ž . Žmembers of two classes of genes involved in these seminal events: ID inhibitor of differentiation -1 and GAS growth arrest-specific.gene 5. When MK31 cells were maintained in an uncommitted state, levels of ID-1 mRNA were high and GAS5 transcripts were
essentially undetectable. Application of cytokines that promote neuronal lineage commitment and cell cycle exit resulted in down-regu-lation of ID-1 and upregulation of GAS5 transcripts, whereas additional cytokine paradigms that promoted terminal neuronaldifferentiation resulted in the delayed down-regulation of GAS5 mRNA. Stable MK31 transfectants were generated for ID-1 and GAS5.In basal conditions, cellular proliferation was enhanced in the ID-1 transfectants and inhibited in the GAS5 transfectants when comparedwith control MK31 cells. When cytokine-mediated neurogenesis was examined in these transfected cell lines, constitutive expression ofID-1 inhibited and constitutive expression of GAS5 enhanced initial and terminal stages of neuronal differentiation, with evidence thatterminal neuronal maturation in both transfectant lines was associated with decreased cellular viability, possibly due to the presence ofconflicting cell cycle-associated developmental signals. These experimental reagents will prove to be valuable experimental tools to helpdefine the functional interrelationships between changing profiles of connexin protein expression and cell cycle regulation duringneuronal ontogeny in the mammalian brain. The present review summarizes the current state of research involving the temporal
AbbreÕiations: Cx, connexin; CNS, central nervous system; Steady-state junctional conductance, g ; Half-inactivation voltage, V ; Ratio of minimal toj 0Ž .maximal junctional conductance, g rg ; Lucifer yellow dye , LY; Serum-free media, SFM; Nestin, N; Vimentin, V; Growth arrest-specific gene,min max
GAS; Inhibitor of differentiation protein, ID) Corresponding author. Department of Neuroscience RFK a724, Albert Einstein College of Medicine, 1300 Morris Park Avenue, Bronx, NY 10461,
USA. Fax: q1-718-430-8682; e-mail: [email protected]
0165-0173r00r$ - see front matter q 2000 Elsevier Science B.V. All rights reserved.Ž .PII: S0165-0173 99 00096-X

( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–7158
expression of such connexin types in differentiating hippocampal neurons and speculates on the possible role of these intercellularchannels in the development and plasticity of the nervous system. In addition, we describe the functional properties and expression patternof the newly discovered neuronal-specific gap junctional protein, Cx36, in the developing mouse fetal hippocampus and in the rat retinaand brain. q 2000 Elsevier Science B.V. All rights reserved.
Keywords: Cx26; Cx33; Cx36; Cx37; Cx40; Cx43; Cx45; Neuronal differentiation; RT–PCR; Gap junction; Electrotonic synapse; Hippocampus
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
Ž .2. Generation of an immortalized neuronal precursor cell line MK31 and environmental regulation of early neurogenesis . . . . . . . . . . . . . . 60
3. Differential expression of ID-1 and GAS5 genes in MK31 cells undergoing neuronal differentiation . . . . . . . . . . . . . . . . . . . . . . . . . 613.1. Roles of ID-1 and GAS5 in neuronal differentiation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
4. Cx36, a newly discovered connexin type. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 634.1. Cx36 forms functional channels. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 634.2. Functional coupling in Cx36-transfected N2A and PC12 cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 644.3. Junctional currents in Cx36-transfected cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 644.4. Brain and retinal distribution of the neuronal Cx36 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
5. Expression of neuronal connexins during hippocampal ontogeny . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 655.1. Expression of Cx36 in the developing hippocampus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68
6. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
Acknowledgements. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
1. Introduction
Ž .Neurons throughout the central nervous system CNSundergo proliferation, migration, differentiation and pheno-typic specification including synaptogenesis during theirdevelopmental cycle. Each of the major stages of neuronaldifferentiation has been associated with distinct regulatorymechanisms. However, how specific molecular mecha-nisms modulate progressive stages of terminal differentia-tion of neuronal cells remains unclear, as do the timing andthe individual roles of gap junction channels in the acquisi-tion of the neuronal phenotype.
Gap junction channels mediate intercellular communica-w xtions in most tissues and organ systems 11 . These chan-
nels allow bidirectional exchange of ions and smallmolecules between cells, coupling the cells both electro-tonically and metabolically. Gap junction-mediated inter-cellular communication appears to be required for normalcellular development as well as for tissue differentiationw x11,14,25 . The critical role of gap junctions during em-bryogenesis may be to provide intercellular pathways forthe diffusion of morphogens and other developmentallyrelevant factors, including Ca2q and a range of additionalsecond messenger molecules. Dynamic changes in thecellular localization of such cytoplasmic factors may initi-
ate concurrent programs required for subsequent cellulardifferentiation.
In the brain, a number of different connexins are ex-pressed, including a variety in astrocytes and both Cx32
w xand Cx45 in oligodendrocytes 3,10 . In contrast, the iden-tity of connexins that participate in the formation of gapjunctions that are expressed in developing and adult neu-rons has not until recently been clearly defined, despitecompelling functional evidence that gap junctions are ex-pressed in various regions.
Electrotonic coupling mediated by gap junctions hasbeen proposed to be responsible for synchronization of
w xsignals in the inferior olive 23 and among hippocampalw x w xCA3 neurons 27,54 , in the retina 57 and during neural
w xdevelopment 19,42,45,60 . In addition, metabolic couplingmediated by gap junctions has been proposed to play animportant role synchronizing electrical relations amongadult neurons as well as in pattern formation during neu-
w xronal development and differentiation 19,42,45 .Among neurons, the predominant connexin type has
appeared to be Cx32, for which immunostaining has beendetected in certain neuronal populations of the brainstem,in cerebral cortical layers, in the basal ganglia, and in
w xsubstantia nigra 12 . Interestingly, Cx32 staining has notbeen demonstrated between hippocampal pyramidal cells,

( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–71 59
which in the case of CA3 neurons have been shown to beelectrically coupled, raising the possibility that gap junc-tions are too small to be detected by this technique or thatadditional connexin types might contribute to neuronal gapjunctions. In fact, Cx26, Cx30, Cx32, Cx33, Cx36, Cx37,Cx40, Cx43, Cx45, Cx46 and Cx50 have all been reportedto be present in certain neuronal populations in vivo or inculture and at certain developmental stages or under cer-
w xtain culture conditions 10,46,49,52 .Numerous studies have demonstrated that the incidence
Žof interneuronal coupling and, by inference, the expres-.sion of gap junction proteins decreases dramatically dur-
ing the processes of brain embryogenesis and neuronalw xmaturation 24,42,44–46,52 . In addition, during both early
and late stages of neural differentiation, expression shiftsfrom one type of connexin to another. For instance, Cx26is highly expressed in fetal as compared to adult brainwhile Cx32 shows developmentally regulated increases inabundance that only reach maximal levels after birth.Because these changes in coupling strength and patterns ofconnexin expression coincide with the progressive differ-entiation and commitment of cells and of cell groups to theneuronal lineage, it has been hypothesized that the pres-ence of gap junctions provides diffusion of signaling
w xmolecules among the requisite cell types 52 .Even after neuronal circuits are formed, however, cou-
pling among neurons persists to various degrees in differ-w xent brain regions 3,24,42,52 . In some regions such as the
neocortex, coupling gradually becomes more and morerestricted during early neonatal life, as the necessity forsynchrony in the establishment of projections onto adja-cent cells declines. In this way, the individual identity ofneurons within functional subcircuits is optimized. A ma-jor recent discovery regarding circuitry formation in thecortex and in the hippocampus is that this developmentalprocess coincides with the peak of expression of theneuronal-specific gap junction channels formed of Cx36.
Studies of neural development have been fostered bythe introduction of immortalized precursor cells that can bemanipulated in vitro towards graded stages of cellular
w xdifferentiation 29,32,44,45 . Our approach has been toutilize a conditionally immortalized murine hippocampal
Ž .cell line MK31 that is developmentally responsive to aŽ .specific set of cytokines, particularly interleukin 7 IL-7 ,
in concert with two other growth factors found in theŽ .developing brain, basic fibroblast growth factor bFGF
Ž . w xand transforming growth factor alpha TGFa 32,44,45 .IL-7 has been detected in embryonic and adult murinebrain and has been shown to possess neuronotrophic ac-tions on primary hippocampal cultures examined in vitrow x2,33 . The use of immortalized neuronal cell lines hasproven to be invaluable as an experimental cellular modelin which the progressive modulation of functional expres-sion of a specific set of connexins can be followed duringneurogenesis and circuit formation; the different develop-mental profile of expression of these different connexin
types in neuronal precursor cells has helped us to under-stand their distinct functions during hippocampal on-togeny. However, this leads to the following question:How is the expression of different connexin types regu-lated in differentiating neurons?
The developmental transitions associated with changingprofiles of connexin transcript and protein expression dur-ing progressive stages of neurogenesis coincides with dif-ferent cell cycle checkpoints. Two classes of genes that areinvolved in different phases of cell cycle control and havebeen implicated directly in mammmalian neurogenesis are
Ž .the ID inhibitors of neurogenesis and growth arrest-Ž . w xspecific GAS families 16,17 . ID genes code for domi-
Ž .nant-negative regulators of basic helix–loop–helix bHLHproteins, and are known to interact with ubiquitous class AbHLH proteins to sequester them and prevent them frominteracting with tissue-restricted class B bHLH proteins,including those that represent sequential sets of neural
w xdetermination and differentiation factors 26 . There areŽ .four known ID genes ID-1–4 that are expressed in
complex and partially overlapping patterns during neuroge-w xnesis 17 . ID-1 and -3 appear to be involved in preventing
uncommitted neural precursor species from initiating aprogram of premature neuronal lineage commitment and
w xcellular differentiation 26 . Neural precursor cells in micewith knockouts of both ID-1 and -3 genes exhibit prema-ture cell cycle withdrawal, precocious expression of neuraldetermination and differentiation genes and upregulation
w xof cyclin-dependent kinase inhibitors 26 . ID-2 and -4exhibit different profiles of CNS developmental expressionand environmental regulation, and may be involved in later
w xevents in neural lineage specification 1,43,55 . There areseveral classes of GAS genes that code for a wide varietyof cellular proteins of diverse functions. GAS7 is ex-pressed in mature neurons of the cerebral cortex, cerebel-lum and hippocampus, and appears to positively regulate
w xneurite outgrowth 18 . GAS3 codes for the peripheralŽ .myelin protein22 PMP22 present in Schwann cells; this
gene is mutated in the neuropathic disorder, Charcot–w x ŽMarie–Tooth type 1A disease 37 see also Ressot et al..and Abrams et al. in this edition . GAS6 is the ligand for
the AxlrRse and Sky family of proteins and is known tow xpromote growth of Schwann cells 22,28 . GAS5 is a
multi-small-nucleolar-RNA host gene that is a member ofX w xthe 5 -terminal oligopyrimidine gene family 50 . Spliced
GAS5 mRNAs are normally associated with ribosomesand are rapidly degraded. During growth arrest, GAS5 has
w xbeen shown to accumulate in mRNP particles 50 . C-myc,a positive regulator of cell cycle progression in uncommit-ted progenitors, has recently been shown to repress the
w xexpression of transcripts for GAS1 21 .In this article, we focus on the temporal expression of
gap junction channels during neuronal differentiation. Wereview the interrelationship between expression of gapjunctions and voltage- and ligand-gated responses in differ-entiating hippocampal neurons, discuss the properties of

( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–7160
the newly discovered specific gap junction protein Cx36and discuss two classes of genes involved in the regulationof neurogenesis. Studies such as this, made possible by thegeneration of an immortalized hippocampal neuronal pro-genitor cell line, are critical for understanding the molecu-lar physiology and functional roles of gap junctions duringneuronal ontogeny.
2. Generation of an immortalized neuronal precursor( )cell line MK31 and environmental regulation of early
neurogenesis
A variety of primary cell culture techniques have beendeveloped to provide reliable, genetically homogeneoussystems for developmental studies. In general, these tech-niques are limited by the heterogeneity of the neuralprecursor cells. To circumvent this problem, we haveutilized clonal cell lines established by retroviral transduc-
Ž .tion of embryonic E16.5–17.5 murine hippocampal cellsŽ .using temperature-sensitive alleles A58rU19 of the
Ž . Ž .simian virus SV 40 large tumor T antigen. Single copySouthern blot analysis using probes for the neomycinresistance gene and for the SV40 large T antigen haveverified the clonal nature of selected experimental celllines. Among them, we have concentrated in one clonalcell line, termed MK31, that exhibited pronounced re-
w x Žsponses to different pharmacological treatments 32 Fig..1 . Under conditions permissive for T antigen expressionŽ .338C , individual neural progenitor cells exhibited self-re-newal, clonal expansion, both symmetric and asymmetricmodes of cell division and the potential to generate neu-ronal, astroglial and oligodendroglial progeny, all of whichare characteristic of stem cells. By contrast, at the non-per-
Ž .missive temperature for T antigen expression 398C , spe-cific sets of cytokines potentiated the progressive elabora-tion of intermediate and progressively more mature neurallineage species.
When propagated at the non-permissive temperature inSFM in the absence of additional growth factors, neurallineage species derived from MK31 stem cells exhibitedimmunoreactivity for early and intermediate progenitor
Ž .cell markers vimentin, nestin, GD3, A2B5 , but were notreactive for markers of committed neural lineage speciesŽneuronal class III btubulin, NF66, O4 and glial fibrillary
.acidic protein, GFAP . Cultures were initially composed ofprimordial epithelioid cells that were immunoreactive for
Ž .vimentin, but not for nestin VqrNy or for additionallineage-independent or -specific protein species. Whenthese early neural cells were replated at 338C in SFM as
Ž .single cells limiting dilution protocols , they displayedŽ .rapid doubling times 6–8 h , exhibited the property of
self-renewal and were capable of generating mature neu-rons, oligodendrocytes and astrocytes when examined un-der differentiating culture conditions. Propagation of thesecells in clonal density conditions at 398C in SFM resulted
Fig. 1. Morphological and immunocytochemical characteristics of neu-Ž . Ž .ronal progenitor cells MK31 treated with different cytokines. A Plated
Ž . Ž .in fetal calf serum at 338C and immunostained for nestin. B As in A ,Ž . Ž .but with high-power view. C Serum-free media SFM at 398C; im-
Ž . Ž .munostained for NF66. D As in C , but with IL-11 as well for 6 days;Ž . Ž .immunostained for NFH-P. E As in C , but with IL-7 and TGFa for 6
Ž .days after pretreatment with bFGF; immunostained for NFH-P. FŽ .Primary dissociated murine hippocampal neurons E17 propagated for 6
days; immunostained for NFHP. Note progressive neurite extension andexpression of more mature neurofilaments in response to differentiating
Ž . Ž . Ž .conditions. Scale bars: A 50 mm; D 20 mm; B, C, E and F 10 mm.w xReproduced with permission from Ref. 32 .
Ž .in rapid 4–6 h loss of cellular viability. By contrast,Žpropagation of these neural progenitors on substrate poly-
. ŽD-lysine, PDL in media containing fetal calf serum FCS,.1–10% resulted in preferential survival and delayed cellu-
Ž .lar expansion 3 DIV , with the formation of uniformVqrNy clones that were growth-arrested at the 6–10 cellstage of development. When these early progenitor cellclones were transferred to SFM, a subset of small, round,phase-bright progenitor species were generated that nowexhibited immunoreactivity for both vimentin and nestinŽ .VqrNq .
Under differentiating conditions, the VqrNq progeni-tor cells were capable of generating post-mitotic neurons.
Ž .bFGF 10 ngrml enhanced the generation of uniformŽ .clones VqrNq from individual progenitor species. In
addition, members of a subset of intermediate-actingw xhemopoietins 40 , that allow the constitutive proliferation
of early, activated multipotent hematopoietic progenitorcells, potentiated the bFGF-mediated expansion of uniformVqrNq clones and enhanced the size of individual VqrNq clones. These experimental observations lend supportto the findings that early CNS stem cells are comprised ofnestin-negative precursors that give rise to a subset of
w xnestin-immunoreactive neuronal cells 20 . The clonal ex-pansion and survival of these distinct early progenitorspecies are regulated by different sets of early-acting CNS
Ž .factors e.g., epidermal growth factor, EGF or bFGF andby two classes of early and intermediate-acting hemopoi-
( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–71 61
w xetins 40 that exhibit analogous developmental actionsduring neurogenesis within the mammalian brain.
When the early neural progenitor species were propa-gated under clonal density conditions at 398C, distinctcytokine paradigms were required to selectively programthe elaboration of individual neural lineages: neurons,oligodendrocytes and astrocytes. Exposure to bFGF and to
Žindividual members of an interleukin subset IL-5, -7, or.-9 promoted the elaboration of VqrNq progenitors
Ž .within a primordial VqrNy cellular network, with thesubsequent generation of bipolar phase-bright progenitorsthat were immunoreactive for early neuronal lineage mark-
Ž . Žers NF66 or neuronal class III-btubulin . Molecular RT–. ŽPCR and immunological Western blotrimmunocyto-
.chemistry studies have shown that the neural stemrprogenitor species express the appropriate profiles ofcytokines, receptor subunits and selected intracellular sig-naling molecules necessary to mediate the range of cellularand developmental actions observed in the MK31 cell linew x15 . In addition, immunocytochemical and Western blotanalyses employing antibodies to selected cytokine recep-tors have verified the close correlation between transcriptand protein expression within these early neural progenitorpopulations. Transcripts and proteins for these cytokines,their cognate receptors and for selected proximal intra-cellular signaling components have been shown to bepresent at the requisite developmental stages within themammalian brain to mediate these manifold cellular ac-
w xtions on the neural stemrprogenitor species 29,30 .
3. Differential expression of ID-1 and GAS5 genes inMK31 cells undergoing neuronal differentiation
To begin to investigate the common mechanisms in-volved in neural progenitor cell cycle regulation and neu-ronal differentiation, we focused on two gene families thatare essential for mediating transcriptional repression of
Žearly neural differentiation ID, inhibitory class of bHLH. Žfactors and subsequent cell cycle exit GAS; growth
. w xarrest-specific genes 5,13,31,48 . We concentrated onID-1 because this gene represents the prototype of mem-bers of this family that are involved in early inhibition ofpremature neural differentiation. We also elected to focuson GAS5 because of its positive modulation in associationwith growth arrest and its regulated expression in responseto experimental paradigms that promote graded stages ofneuronal differentiation in the MK31 cell line, and alsobecause of the presence of GAS5 transcripts in mammalian
w xbrain 15 .Northern analysis using a cDNA probe for ID-1 re-
vealed high levels of transcript expression during theinitial expansion of uncommitted progenitors derived fromMK31 stem cells. The generation of post-mitotic neuralspecies using treatment paradigms that promote cellulardifferentiation resulted in progressive reductions in the
levels of expression of ID-1 transcripts at 12 and 36 h.Thus, ID-1 transcripts are expressed at high levels inuncommitted neural progenitors and are promptly down-regulated with the onset of cellular differentiation andincipient cell cycle exit.
By contrast, GAS5 transcripts are not expressed duringthe expansion phase of uncommitted MK31 progenitor
Ž .species 338C, FCS, non-confluent cultures . However,following growth arrest and mandatory cell cycle exitŽ .398C, SFM, cellular confluence , GAS5 transcripts were
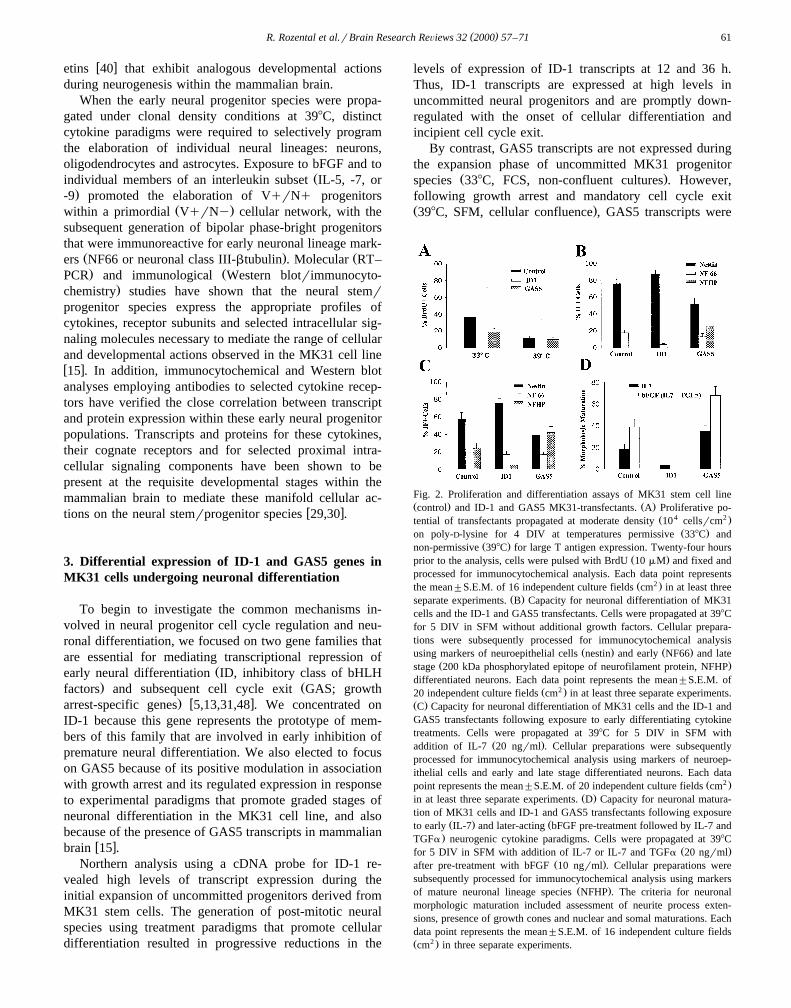
Fig. 2. Proliferation and differentiation assays of MK31 stem cell lineŽ . Ž .control and ID-1 and GAS5 MK31-transfectants. A Proliferative po-
Ž 4 2 .tential of transfectants propagated at moderate density 10 cellsrcmŽ .on poly-D-lysine for 4 DIV at temperatures permissive 338C and
Ž .non-permissive 398C for large T antigen expression. Twenty-four hoursŽ .prior to the analysis, cells were pulsed with BrdU 10 mM and fixed and
processed for immunocytochemical analysis. Each data point representsŽ 2 .the mean"S.E.M. of 16 independent culture fields cm in at least three
Ž .separate experiments. B Capacity for neuronal differentiation of MK31cells and the ID-1 and GAS5 transfectants. Cells were propagated at 398Cfor 5 DIV in SFM without additional growth factors. Cellular prepara-tions were subsequently processed for immunocytochemical analysis
Ž . Ž .using markers of neuroepithelial cells nestin and early NF66 and lateŽ .stage 200 kDa phosphorylated epitope of neurofilament protein, NFHP
differentiated neurons. Each data point represents the mean"S.E.M. ofŽ 2 .20 independent culture fields cm in at least three separate experiments.
Ž .C Capacity for neuronal differentiation of MK31 cells and the ID-1 andGAS5 transfectants following exposure to early differentiating cytokinetreatments. Cells were propagated at 398C for 5 DIV in SFM with
Ž .addition of IL-7 20 ngrml . Cellular preparations were subsequentlyprocessed for immunocytochemical analysis using markers of neuroep-ithelial cells and early and late stage differentiated neurons. Each data
Ž 2 .point represents the mean"S.E.M. of 20 independent culture fields cmŽ .in at least three separate experiments. D Capacity for neuronal matura-
tion of MK31 cells and ID-1 and GAS5 transfectants following exposureŽ . Žto early IL-7 and later-acting bFGF pre-treatment followed by IL-7 and
.TGFa neurogenic cytokine paradigms. Cells were propagated at 398CŽ .for 5 DIV in SFM with addition of IL-7 or IL-7 and TGFa 20 ngrml
Ž .after pre-treatment with bFGF 10 ngrml . Cellular preparations weresubsequently processed for immunocytochemical analysis using markers
Ž .of mature neuronal lineage species NFHP . The criteria for neuronalmorphologic maturation included assessment of neurite process exten-sions, presence of growth cones and nuclear and somal maturations. Eachdata point represents the mean"S.E.M. of 16 independent culture fieldsŽ 2 .cm in three separate experiments.
( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–7162
expressed at high levels within 24 h. During limited prolif-Ž .eration 398C, FCS, 30% confluent cultures , GAS5 tran-
scripts were expressed at low levels. Further, duringŽearly-phase growth arrest 398C, SFM, 80% confluent
.cultures , GAS5 transcripts increased one-to-two fold,Žwhereas in late-phase growth arrest 338C, SFM, conflu-
.ent , they increased three- to four-fold. Treatment of MK31cells for 24 h with agents that induce cell cycle exit andincipient neural differentiation resulted in GAS5 mRNAupregulation that was accentuated by application of inter-mediate-acting neural differentiating and survival factors.Finally, application of later-acting cytokines that promoteterminal neural differentiation resulted in further transcriptupregulation at 36 h, followed by precipitous downregula-tion by 96 h. These overall observations suggest thatGAS5 participates in important neural regulatory functionsintimately linked to exit from cell cycle. This developmen-tal transition may be essential for establishing the cellularconditions necessary to ensure the precisely orchestrated
w xelaboration of the mature neural phenotype 35 .
3.1. Roles of ID-1 and GAS5 in neuronal differentiation
To further define the roles of ID-1 and GAS5 in neuraldevelopment, MK31 cells were separately co-transfectedwith ID-1 or GAS5 and a plasmid containing a hy-gromycin-resistance gene. A series of stable transfectantsand mock-transfected clones were established. Southernand Northern blot analyses using the relevant molecular
and immunological probes have established the constitu-tive expression of the appropriate gene transcripts andprotein species, and the clonality of the established celllines. These experiments have verified that the use ofdifferent sites of genomic integration does not alter thephysiological properties of the transfected cell lines.
In the control MK31 cell line at the permissive tempera-ture, a large proportion of neural progenitors were under-
Ž .going cellular proliferation BrdU-incorporation ; these pa-rameters were significantly enhanced in the ID-1 transfec-
Ž .tants and inhibited in the GAS5 transfectants Fig. 2A . Atthe non-permissive temperature, more limited cellular ex-pansion occurred in the control cell line, and proliferationwas modestly increased in ID-1 transfectants but not in the
Ž .GAS5 transfectants Fig. 2A . In the basal state withoutcytokine additions, the control cell line displayed veryminimal elaboration of early neuronal lineage speciesŽ . Ž .NF66-immunoreactive Fig. 2B . This profile of neuronaldifferentiation was inhibited in the ID-1 transfectants anddramatically potentiated in the GAS5 transfectants, withthe elaboration of a significant pool of more mature neu-
Ž .ronal lineage species Fig. 2B . Application of IL-7 re-sulted in a profile of differentiated neuronal species in thecontrol cell line that was similar to that seen in the GAS5
Ž .transfectants in the absence of added cytokines Fig. 2C .Similarly, the degree of neuronal differentiation was signif-icantly reduced in the ID-1 transfectants and dramatically
Ž .enhanced in the GAS5 transfectants Fig. 2C . Analogousbut more advanced profiles of differentiated neuronal lin-
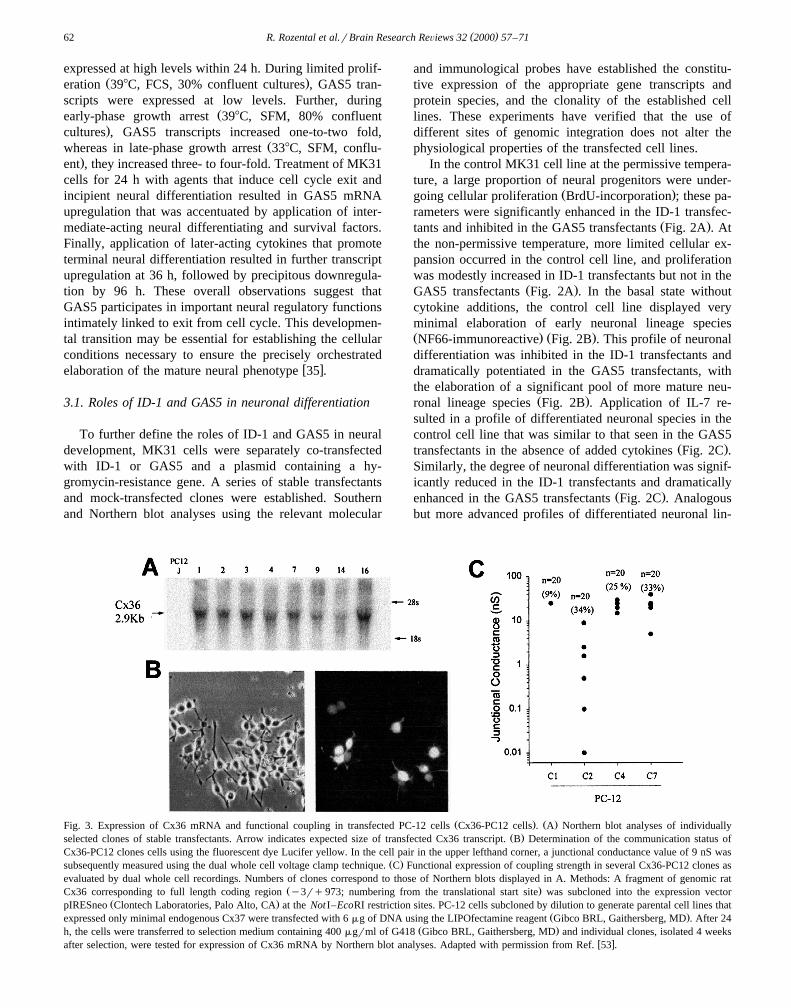
Ž . Ž .Fig. 3. Expression of Cx36 mRNA and functional coupling in transfected PC-12 cells Cx36-PC12 cells . A Northern blot analyses of individuallyŽ .selected clones of stable transfectants. Arrow indicates expected size of transfected Cx36 transcript. B Determination of the communication status of
Cx36-PC12 clones cells using the fluorescent dye Lucifer yellow. In the cell pair in the upper lefthand corner, a junctional conductance value of 9 nS wasŽ .subsequently measured using the dual whole cell voltage clamp technique. C Functional expression of coupling strength in several Cx36-PC12 clones as
evaluated by dual whole cell recordings. Numbers of clones correspond to those of Northern blots displayed in A. Methods: A fragment of genomic ratŽ .Cx36 corresponding to full length coding region y3rq973; numbering from the translational start site was subcloned into the expression vector
Ž .pIRESneo Clontech Laboratories, Palo Alto, CA at the NotI–EcoRI restriction sites. PC-12 cells subcloned by dilution to generate parental cell lines thatŽ .expressed only minimal endogenous Cx37 were transfected with 6 mg of DNA using the LIPOfectamine reagent Gibco BRL, Gaithersberg, MD . After 24
Ž .h, the cells were transferred to selection medium containing 400 mgrml of G418 Gibco BRL, Gaithersberg, MD and individual clones, isolated 4 weeksw xafter selection, were tested for expression of Cx36 mRNA by Northern blot analyses. Adapted with permission from Ref. 53 .
( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–71 63
eage species were seen with specific cytokine co-inductionŽparadigms bFGF pre-treatment followed by application of
. Ž .IL-7 and TGFa Fig. 2D . However, the more advancedcytokine induction paradigm resulted in a moderate loss ofcellular viability in both the ID-1 and the GAS5 transfec-
w xtant lines 15 . This latter observation suggests that incom-patible cellular signals may change the cellular rheostat forviability. These cumulative observations suggest that ID-1and GAS5 may be important for regulating key cell cyclecheckpoints in ways that significantly modulate neurogene-sis in the mammalian hippocampus.
4. Cx36, a newly discovered connexin type
w xRecently, O’Brien et al. 38 cloned a novel connexinŽ .Cx35 from the skate that is expressed at high levels inthe retina, and subsequently reported a highly homologousCx34.7 from perch that is expressed in both retina and
w xbrain 39 . Cx36, the mammalian homologue of skate andŽ .teleost Cx35 see also White and Bruzzone in this edition ,
is preferentially expressed in the mouse retina and inw x Žvarious neuronal cell populations 8,51 see also
.Condorelli et al. in this edition . In situ hybridizationtechniques have shown high levels of Cx36 mRNA inolfactory bulbs, pineal gland, inferior olive, CA3rCA4
w xhippocampal neurons and in the retina 8 , implying thatthis connexin may participate in neuronal gap junctionformation. In addition, Cx36 expression has been shown to
w xbe developmentally regulated 51 : highest levels of thisŽ .transcript in rat brain were detected at postnatal day 7 P7
with subsequent decline to lower levels in adult neurons.The spatial and temporal characteristics of Cx36, togetherwith the lack of expression of this connexin in othertissues, make it the first neuron-specific connexin thus faridentified. Further, we have recently shown that gap junc-
w xtions composed of Cx36 form functional channels 53 .
4.1. Cx36 forms functional channels
Expression of connexins in communication-deficient celllines through transfection with vectors containing cDNAs
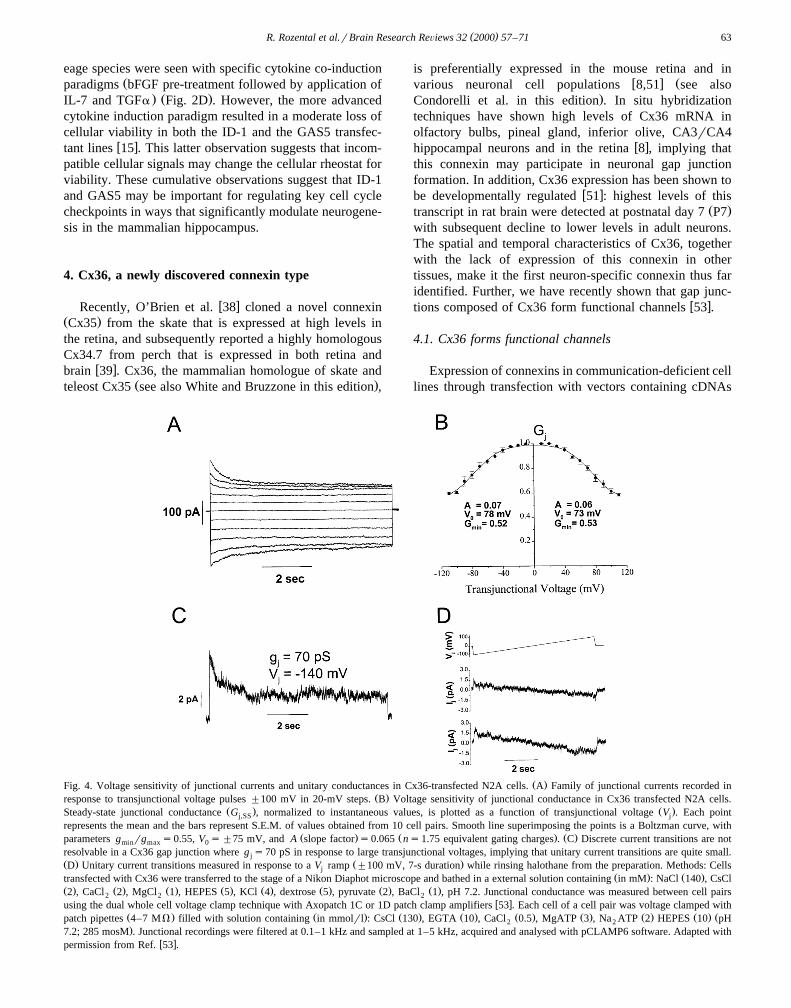
Ž .Fig. 4. Voltage sensitivity of junctional currents and unitary conductances in Cx36-transfected N2A cells. A Family of junctional currents recorded inŽ .response to transjunctional voltage pulses "100 mV in 20-mV steps. B Voltage sensitivity of junctional conductance in Cx36 transfected N2A cells.
Ž . Ž .Steady-state junctional conductance G , normalized to instantaneous values, is plotted as a function of transjunctional voltage V . Each pointj,SS j
represents the mean and the bars represent S.E.M. of values obtained from 10 cell pairs. Smooth line superimposing the points is a Boltzman curve, withŽ . Ž . Ž .parameters g rg s0.55, V s"75 mV, and A slope factor s0.065 ns1.75 equivalent gating charges . C Discrete current transitions are notmin max 0
resolvable in a Cx36 gap junction where g s70 pS in response to large transjunctional voltages, implying that unitary current transitions are quite small.jŽ . Ž .D Unitary current transitions measured in response to a V ramp "100 mV, 7-s duration while rinsing halothane from the preparation. Methods: Cellsj
Ž . Ž .transfected with Cx36 were transferred to the stage of a Nikon Diaphot microscope and bathed in a external solution containing in mM : NaCl 140 , CsClŽ . Ž . Ž . Ž . Ž . Ž . Ž . Ž .2 , CaCl 2 , MgCl 1 , HEPES 5 , KCl 4 , dextrose 5 , pyruvate 2 , BaCl 1 , pH 7.2. Junctional conductance was measured between cell pairs2 2 2
w xusing the dual whole cell voltage clamp technique with Axopatch 1C or 1D patch clamp amplifiers 53 . Each cell of a cell pair was voltage clamped withŽ . Ž . Ž . Ž . Ž . Ž . Ž . Ž . Žpatch pipettes 4–7 MV filled with solution containing in mmolrl : CsCl 130 , EGTA 10 , CaCl 0.5 , MgATP 3 , Na ATP 2 HEPES 10 pH2 2.7.2; 285 mosM . Junctional recordings were filtered at 0.1–1 kHz and sampled at 1–5 kHz, acquired and analysed with pCLAMP6 software. Adapted with
w xpermission from Ref. 53 .

( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–7164
encoding connexins of interest yields clonal cellular popu-lations in which exogenous channel properties can beevaluated against a very low endogenous background of
w xcoupling 58,59 . We stably transfected N2A and PC-12cells with a vector containing a full-length coding region
Ž .of rat Cx36 cDNA Fig. 3A and have characterized Cx36properties in multiple clones of transfectants obtained fromboth N2A neuroblastoma and PC12 pheochromocytoma
Ž .parental cells Fig. 3B,CFig. 4A–D . Our results indicatethat Cx36 gap junction channels have an extremely small
Ž .single channel conductance main state g ;10–15 pS ,j
are only weakly sensitive to transjunctional voltage and areŽ . Ž .permeable to the dye Lucifer yellow LY M 454 Dar
Ž .Figs. 3 and 4 . These observations demonstrate that Cx36forms gap junction channels that have the smallest unitaryconductance and the weakest sensitivity to transjunctionalvoltage of any mammalian connexin studied to date, yetthese channels are permeable to anions in the size range ofsecond messengers such as cAMP and IP .3
4.2. Functional coupling in Cx36-transfected N2A andPC12 cells
Functional coupling can be measured in several ways,and each type of measurement has a different threshold fordetection, may depend on different sets of variables and
w xhas different limitations 47 . We have characterized theexpression and the functional properties of Cx36 using acombination of several different methodologies, as de-
scribed below. First, we have compared dye coupling inCx36 stable-transfectants using LY microinjections. Undercontrol conditions, dye coupling was not observed amongN2A cells that were not transfected with Cx36, whereas innon-transfected PC-12 cells dye coupling was observed
Ž .only very infrequently 6% of the cell clusters . In con-trast, dye coupling was frequently observed both amongN2A or PC-12 transfectants; these results indicate that
ŽCx36 can form functional, anion permeant channels Fig..3B Fig. 4 .
4.3. Junctional currents in Cx36-transfected cells
Gating properties of Cx36 in transfectants were exam-ined by evaluating voltage sensitivity of macroscopic junc-tional currents and amplitudes of single channel currentsŽ . Ž .Fig. 4 . The steady-state junctional conductance g atss
each voltage was normalized relative to the instantaneouscurrent and these G values were plotted as a function ofj,SS
V . The relationship between G and V was determinedj j,SS j
assuming a two-state Boltzmann equation:
G s G yG r 1qexp A V yV qGŽ . Ž Ž .� 4j ,SS max min j 0 min
where V is the voltage at which the conductance is0
half-maximal, G is the maximum normalized conduc-max
tance, G is the normalized voltage-insensitive residualmin
conductance and A is a parameter defining the steepnessŽof voltage sensitivity AsnqF, where n is the equivalent
number of gating charges of valence q and F is the
Ž . Ž .Fig. 5. Postnatal brain and retinal distribution of the neuronal Cx36. A–C In situ hybridization of Cx36 mRNA expression in the P7 brain. A Note theabundant expression in the superficial layers of the developing cortex and in the olfactory bulb. Also note that expression is virtually absent in the
Ž . Ž .subcortical white matter. B Cx36 mRNA in the cerebellum is abundant in the Purkinje cell and the molecular layer. C Sense control shows no specificŽ . Ž .labeling. D Nuclease protection assay for Cx36 mRNA expression in the P7 brain. Expression is moderately abundant in the hippocampus Hipp and
Ž . Ž .olfactory bulb OB but undetectable in subcortical white matter SCW . Twenty micrograms of total mRNA were loaded onto each lane. Adapted withw xpermission from Ref. 53 .

( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–71 65
.Faraday constant . To determine the sensitivity of junc-Ž . Ž .tional conductance g to transjunctional voltage V inj j
Cx36-transfected cells, pulses of 7–10 s duration from aholding potential of 0 mV to voltages ranging from y110to 110 mV were applied to one cell of a pair and junctional
Ž .currents I traces, Fig. 4A,C,D were measured. ThejŽ . Žinstantaneous I junctional currents peak values at thej,0
.start of the pulses were found to vary linearly withtransjunctional voltage; in contrast, I showed rectifica-j,ss
tion.The dependence of the normalized steady-state g j
Ž .G on V is shown in Fig. 4B. G was obtained byj,SS j j,SS
dividing the steady-state I by I for each V . G dataj,ss j,0 j j,SS
for V ’s of each polarity were well fit to a two-statej
Boltzmann relationship. The best fit for the Boltzmannequation for the average G values was obtained: G sj,SS min
Ž .0.52, V sy78 mV and As0.07 gating charges1.750
for pulses of negative polarity, and G s0.53, V s73min 0Ž .mV and As0.06 gating charges1.5 for positive polar-
ity. These results indicate that Cx36 forms gap junctionchannels that are only weakly sensitive to transjunctionalvoltage.
One way to examine the amplitude of the voltage-insen-sitive channels has involved the use of cell pairs whereonly a few channels were expressed. Under these condi-tions, the unitary conductances of Cx36 channels wereshown to be ;10–15 pS; application of halothane com-pletely abolished this junctional current. In response tovoltage ramps from y100 to 100 mV, openings of a
Ž .channel to the 1.5 pA level 15 pS as well as partialclosure of the channel could be detected.
4.4. Brain and retinal distribution of the neuronal Cx36
In order to explore in more detail the expression ofCx36 in brain tissue, in situ hybridization and RNaseprotection assays were performed on P7 rat brains and ratretina. As is illustrated in Fig. 5, in situ hybridizationsignals were strongly localized to the superficial layers of
Ž .the neocortex, the olfactory bulb Fig. 5A and the hip-Ž .pocampus see Section 5.1 . In the cerebellum, a strong
signal was detected in the Purkinje cell layer and in theŽ .molecular layer Fig. 5B . Signals were virtually absent in
Ž .cortical white matter Fig. 5A and in brain sections hy-Ž .bridized with sense constructs Fig. 5C . In situ hybridiza-
tion signals were much lower in adult brain. Nucleaseprotection assays performed on material from these regions
Ž .in newborn P7 rat confirmed the expression of Cx36 inolfactory cortex and hippocampus and its absence in corti-
Ž .cal white matter Fig. 5D .Although Cx36 specific antibodies are not yet available,
a polyclonal antibody has been raised against the intra-w xcellular loop domain of the closely related skate Cx35 39 .
Staining of retinal sections with this antibody revealedimmunoreactivity in the inner nuclear and ganglion cell
Ž .layers Fig. 6A . Immunostaining of adult rat hippocampus
Fig. 6. Immuno-identification of Cx36 expressed in retinal neurons andŽ .hippocampal neurons using antibodies prepared against skate Cx35. A
Retina. Low-power micrograph of a transverse retinal section showslabeling of neurons in the inner nuclear and ganglion cell layers; IPL,inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform
Ž .layer; ONL, outer nuclear layer; P, photoreceptor layer. B Hippocam-pus. Note the positive reaction product in pyramidal cells and the reducedreactivity in the granular cells of the dentate gyrus. Methods: The ABCmethod was use for labeling of sections of the hippocampal and retinal
Ž .sections Vector Labs . In brief, unfixed samples of were cut at y308C,collected on poly-L-lysine coated coverslips and stored at y208C untiluse. Before immunoincubation, slices were fixed in 100% ethanol for 20min, rinse in PBS and blocked with 0.1% BSA, followed by incubationfor 46 h with primary anti-Cx35 antibodies at 48C. Sections were thenincubated with biotinylated goat anti-rabbit secondary antibody for 24 h.After washing and blocking, the sections were finally incubated for 4 hwith the avidin–biotinylated peroxidase complex. Peroxidase activity wasvisualized with 3, 3X diamonobenzidine followed by enhancement with0.1% osmium tetroxide.
sections showed positive reaction product in pyramidalŽ .cells Fig. 6B and led to the question of precisely at what
developmental time point is Cx36 expressed in differentiat-ing neurons and how does this expression correlate withmaturation of neurons?
5. Expression of neuronal connexins during hippocam-pal ontogeny
Intercellular coupling is quite strong among MK31embryonic cells destined to become hippocampal neurons
( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–7166
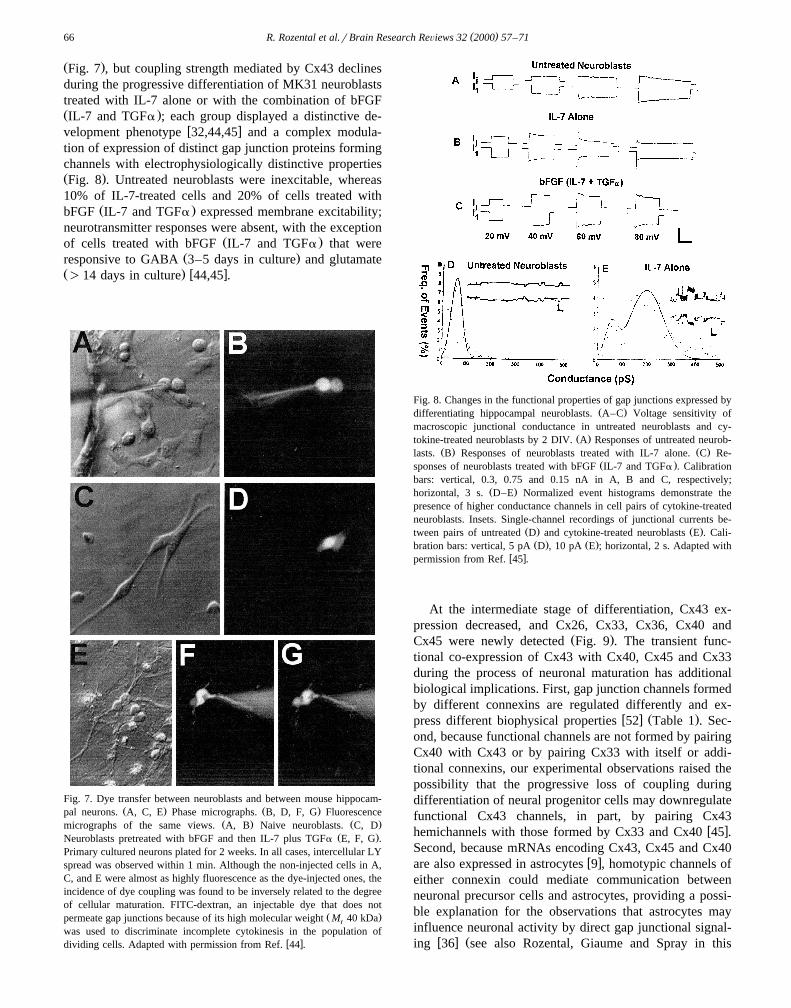
Ž .Fig. 7 , but coupling strength mediated by Cx43 declinesduring the progressive differentiation of MK31 neuroblaststreated with IL-7 alone or with the combination of bFGFŽ .IL-7 and TGFa ; each group displayed a distinctive de-
w xvelopment phenotype 32,44,45 and a complex modula-tion of expression of distinct gap junction proteins formingchannels with electrophysiologically distinctive propertiesŽ .Fig. 8 . Untreated neuroblasts were inexcitable, whereas10% of IL-7-treated cells and 20% of cells treated with
Ž .bFGF IL-7 and TGFa expressed membrane excitability;neurotransmitter responses were absent, with the exception
Ž .of cells treated with bFGF IL-7 and TGFa that wereŽ .responsive to GABA 3–5 days in culture and glutamate
Ž . w x)14 days in culture 44,45 .
Fig. 7. Dye transfer between neuroblasts and between mouse hippocam-Ž . Ž .pal neurons. A, C, E Phase micrographs. B, D, F, G Fluorescence
Ž . Ž .micrographs of the same views. A, B Naive neuroblasts. C, DŽ .Neuroblasts pretreated with bFGF and then IL-7 plus TGFa E, F, G .
Primary cultured neurons plated for 2 weeks. In all cases, intercellular LYspread was observed within 1 min. Although the non-injected cells in A,C, and E were almost as highly fluorescence as the dye-injected ones, theincidence of dye coupling was found to be inversely related to the degreeof cellular maturation. FITC-dextran, an injectable dye that does not
Ž .permeate gap junctions because of its high molecular weight M 40 kDar
was used to discriminate incomplete cytokinesis in the population ofw xdividing cells. Adapted with permission from Ref. 44 .
Fig. 8. Changes in the functional properties of gap junctions expressed byŽ .differentiating hippocampal neuroblasts. A–C Voltage sensitivity of
macroscopic junctional conductance in untreated neuroblasts and cy-Ž .tokine-treated neuroblasts by 2 DIV. A Responses of untreated neurob-
Ž . Ž .lasts. B Responses of neuroblasts treated with IL-7 alone. C Re-Ž .sponses of neuroblasts treated with bFGF IL-7 and TGFa . Calibration
bars: vertical, 0.3, 0.75 and 0.15 nA in A, B and C, respectively;Ž .horizontal, 3 s. D–E Normalized event histograms demonstrate the
presence of higher conductance channels in cell pairs of cytokine-treatedneuroblasts. Insets. Single-channel recordings of junctional currents be-
Ž . Ž .tween pairs of untreated D and cytokine-treated neuroblasts E . Cali-Ž . Ž .bration bars: vertical, 5 pA D , 10 pA E ; horizontal, 2 s. Adapted with
w xpermission from Ref. 45 .
At the intermediate stage of differentiation, Cx43 ex-pression decreased, and Cx26, Cx33, Cx36, Cx40 and
Ž .Cx45 were newly detected Fig. 9 . The transient func-tional co-expression of Cx43 with Cx40, Cx45 and Cx33during the process of neuronal maturation has additionalbiological implications. First, gap junction channels formedby different connexins are regulated differently and ex-
w x Ž .press different biophysical properties 52 Table 1 . Sec-ond, because functional channels are not formed by pairingCx40 with Cx43 or by pairing Cx33 with itself or addi-tional connexins, our experimental observations raised thepossibility that the progressive loss of coupling duringdifferentiation of neural progenitor cells may downregulatefunctional Cx43 channels, in part, by pairing Cx43
w xhemichannels with those formed by Cx33 and Cx40 45 .Second, because mRNAs encoding Cx43, Cx45 and Cx40
w xare also expressed in astrocytes 9 , homotypic channels ofeither connexin could mediate communication betweenneuronal precursor cells and astrocytes, providing a possi-ble explanation for the observations that astrocytes mayinfluence neuronal activity by direct gap junctional signal-
w x Žing 36 see also Rozental, Giaume and Spray in this

( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–71 67
Ž . w xFig. 9. Determination by PCR of the connexins expressed by differentiating hippocampal neuroblasts MK31 45 . A composite of gels fromw xfirst-generation cDNA obtained by RT–PCR, run on 2% agarose gel, and visualized by ethidium bromide staining 45,56 . RNA was obtained from cells at
Ž . w x338C, from cells kept at 398C and treated with IL-7 alone or from mouse hippocampal primary cultured neurons kept for 2 weeks in culture E18 45,56 .w x Ž . Ž . Ž . Ž .Specific primers were used as previously described 56 . Mouse liver Cx26 , testis Cx33 , eye ball Cx36 and heart Cx40, Cx43 and Cx45 provide
w xpositive controls. Methods: RT–PCR assays on total RNA isolated from primary cultured cells were obtained as described 56 and were performed usingŽ . Žthe Thermoscript RT–PCR System Gibco BRL, Life Technologies, Grand Island, NY . RNA was treated with DNase I Boehringer Mannheim,
.Indianapolis, IN to eliminate contamination with residual genomic DNA. Oligonucleotides corresponding to the carboxyl terminal of Cx36 wereŽ . Žsynthesized by Gene Link Thornwood, NY . The sense and antisense primers used and their predicted fragments are described below for more details, see
w x.Ref. 56 . PCR reactions contained 1–2 mg of first strand cDNA, 50 mM of sense and antisense primers, 5 ml of 10= PCR Buffer, 1.5 mM MgCl , 1 ml2Ž .10 mM dNTP, and 2.5 U Taq Polymerase Gibco BRL, Life Technologies, Grand Island, NY in a final volume of 50 ml. Thirty cycles were performed onŽ . Ž . Ž .the samples using a PTC-100 Thermocycler M.J. Research, Watertown, MA as follows: 1 denaturation at 948C for 30 s; 2 annealing at 558C for 30 s;
Ž .3 extension at 728C for 30 s. This was followed by a final extension cycle at 728C for 8 min and soak cycle at 48C. Reaction products were analyzed byelectrophoresis on 2% agarose gels.
Ž .mCx26 AGATGGAGGGAGAGGATGAG TCAGAGGAAGAGAAACAATGTG 312 bpŽ .rCx33 AAACCATCTTCATCCTCTTC GCTTTTCTGTCTACCTAAAACC 386 bp
Ž .mCx36 GAGCAAACGAGAAGATAAGAAG TGGATGATGTAGAAGCGG 195 bpŽ .mCx40 TTTGGCAAGTCACGGCAGGG TTGTCACTGTGGTAGCCCTGAGG 311 bp
Ž .mCx43 TACCACGCCACCACTGGC AATCTCCAGGTCATCAGG 407 bpŽ .mCx45 AAAGAGCAGAGCCAACCAAA GTCCCAAACCCTAAGTGAAGC 313 bp
.edition . Such a cellular mechanism might underlie suchdiverse physiological and pathological processes as Leao˜
Žspreading depression see Martin-Ferreira, Nedeergard and
.Nicholson in this edition or specific forms of epilepsy inŽwhich levels of Cx43 mRNA are increased see Carlen in
.this edition .
Table 1Functional properties of exogenously expressed gap junctions formed by different connexin types
Ž . Ž . Ž .V mV is transjunctional voltage V at which junctional conductance g is reduced by 50%; G is residual junctional conductance at high V ; n is the0 j j min j
equivalent gating charges of valence q; Pq and Py indicate relative cationic and anionic permeabilities, respectively. Ion selectivity was determined bymeasuring reversal potentials for current through gap junction channels with asymmetric salt solutions; NF: non-functional channels. ND; not determined.
Ž .Connexin Voltage dependence Unitary conductance pS PqrPy
Ž .V mV n g rg Main state Substate0 min max
aCx26 q82, y89 2 q0.17, y0.36 140 15, 20, 30 NDCx32 "48 2 0.27 70 10, 19, 26 0.77Cx33 NF NF NF NF NF NF
bCx36 "75 1.5 0.53 -15 ND NDaCx37 "29 3 0.10 280 60 ND
Cx40 "48 3 0.28 180 35 5.2Cx43 "60 2.5 0.37 60, 90 30 1.2Cx45 "13 2.7 0.06 32 ND ND
a Low LY permeability implies PqrPy41.b w xLY permeable. Table compiled from Refs. 4,6,7,34,41,53,58,59 .
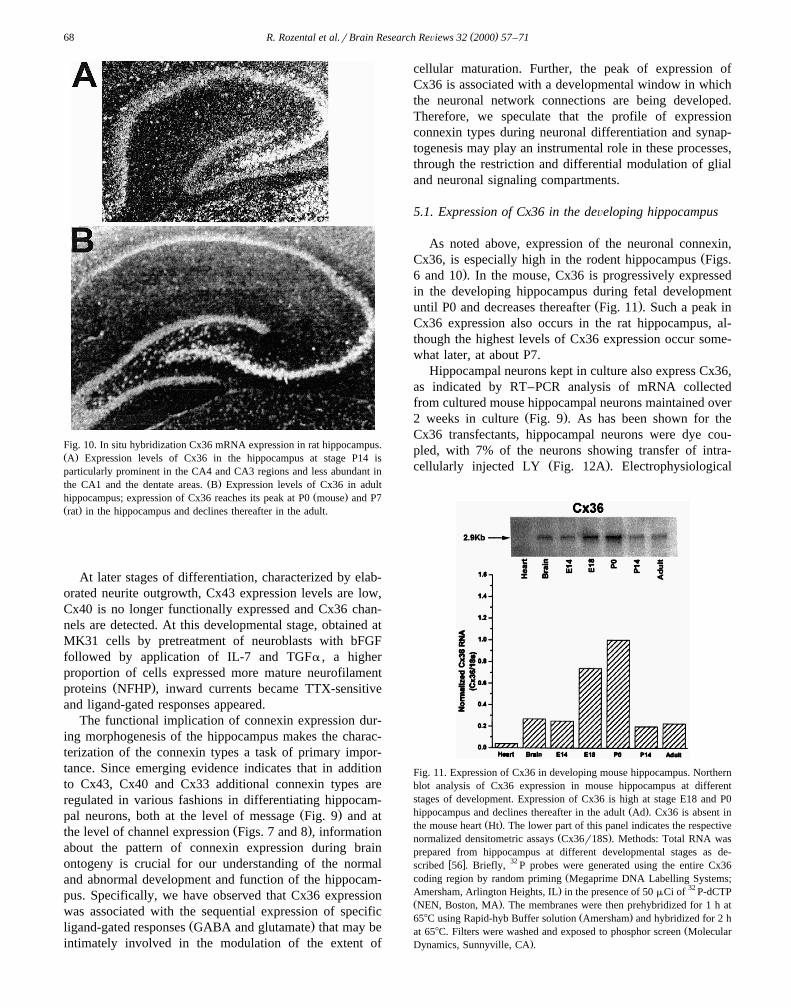
( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–7168
Fig. 10. In situ hybridization Cx36 mRNA expression in rat hippocampus.Ž .A Expression levels of Cx36 in the hippocampus at stage P14 isparticularly prominent in the CA4 and CA3 regions and less abundant in
Ž .the CA1 and the dentate areas. B Expression levels of Cx36 in adultŽ .hippocampus; expression of Cx36 reaches its peak at P0 mouse and P7
Ž .rat in the hippocampus and declines thereafter in the adult.
At later stages of differentiation, characterized by elab-orated neurite outgrowth, Cx43 expression levels are low,Cx40 is no longer functionally expressed and Cx36 chan-nels are detected. At this developmental stage, obtained atMK31 cells by pretreatment of neuroblasts with bFGFfollowed by application of IL-7 and TGFa , a higherproportion of cells expressed more mature neurofilament
Ž .proteins NFHP , inward currents became TTX-sensitiveand ligand-gated responses appeared.
The functional implication of connexin expression dur-ing morphogenesis of the hippocampus makes the charac-terization of the connexin types a task of primary impor-tance. Since emerging evidence indicates that in additionto Cx43, Cx40 and Cx33 additional connexin types areregulated in various fashions in differentiating hippocam-
Ž .pal neurons, both at the level of message Fig. 9 and atŽ .the level of channel expression Figs. 7 and 8 , information
about the pattern of connexin expression during brainontogeny is crucial for our understanding of the normaland abnormal development and function of the hippocam-pus. Specifically, we have observed that Cx36 expressionwas associated with the sequential expression of specific
Ž .ligand-gated responses GABA and glutamate that may beintimately involved in the modulation of the extent of
cellular maturation. Further, the peak of expression ofCx36 is associated with a developmental window in whichthe neuronal network connections are being developed.Therefore, we speculate that the profile of expressionconnexin types during neuronal differentiation and synap-togenesis may play an instrumental role in these processes,through the restriction and differential modulation of glialand neuronal signaling compartments.
5.1. Expression of Cx36 in the deÕeloping hippocampus
As noted above, expression of the neuronal connexin,ŽCx36, is especially high in the rodent hippocampus Figs.
.6 and 10 . In the mouse, Cx36 is progressively expressedin the developing hippocampus during fetal development
Ž .until P0 and decreases thereafter Fig. 11 . Such a peak inCx36 expression also occurs in the rat hippocampus, al-though the highest levels of Cx36 expression occur some-what later, at about P7.
Hippocampal neurons kept in culture also express Cx36,as indicated by RT–PCR analysis of mRNA collectedfrom cultured mouse hippocampal neurons maintained over
Ž .2 weeks in culture Fig. 9 . As has been shown for theCx36 transfectants, hippocampal neurons were dye cou-pled, with 7% of the neurons showing transfer of intra-
Ž .cellularly injected LY Fig. 12A . Electrophysiological
Fig. 11. Expression of Cx36 in developing mouse hippocampus. Northernblot analysis of Cx36 expression in mouse hippocampus at differentstages of development. Expression of Cx36 is high at stage E18 and P0
Ž .hippocampus and declines thereafter in the adult Ad . Cx36 is absent inŽ .the mouse heart Ht . The lower part of this panel indicates the respective
Ž .normalized densitometric assays Cx36r18S . Methods: Total RNA wasprepared from hippocampus at different developmental stages as de-
w x 32scribed 56 . Briefly, P probes were generated using the entire Cx36Žcoding region by random priming Megaprime DNA Labelling Systems;
. 32Amersham, Arlington Heights, IL in the presence of 50 mCi of P-dCTPŽ .NEN, Boston, MA . The membranes were then prehybridized for 1 h at
Ž .658C using Rapid-hyb Buffer solution Amersham and hybridized for 2 hŽat 658C. Filters were washed and exposed to phosphor screen Molecular
.Dynamics, Sunnyville, CA .
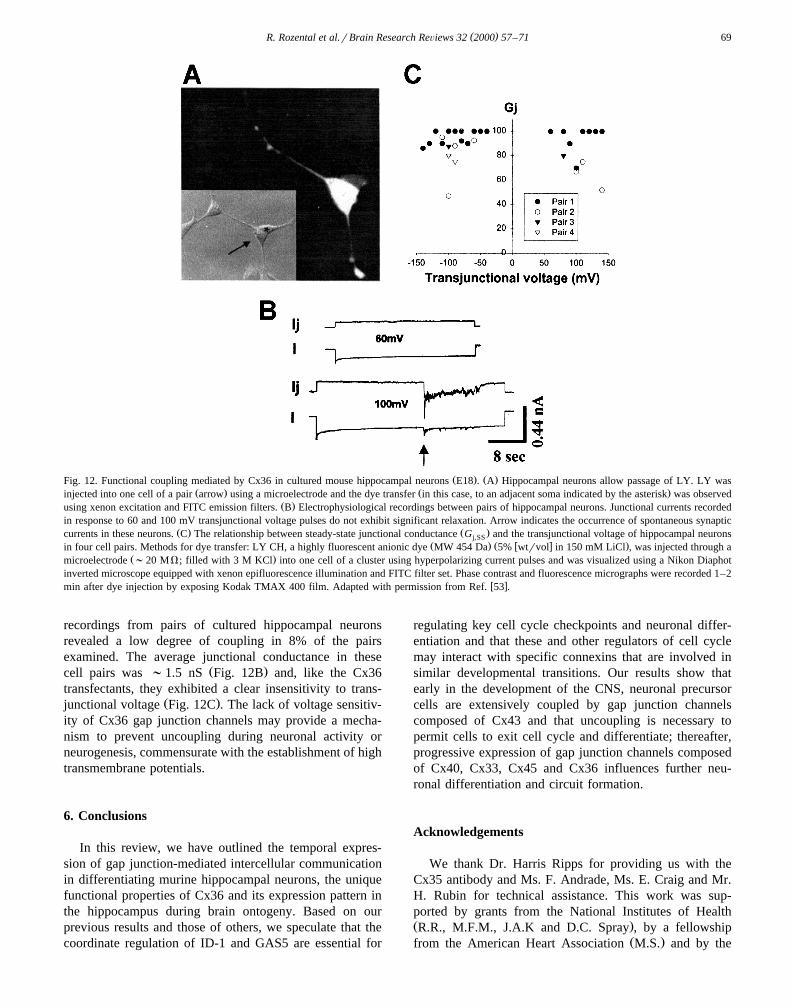
( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–71 69
Ž . Ž .Fig. 12. Functional coupling mediated by Cx36 in cultured mouse hippocampal neurons E18 . A Hippocampal neurons allow passage of LY. LY wasŽ . Ž .injected into one cell of a pair arrow using a microelectrode and the dye transfer in this case, to an adjacent soma indicated by the asterisk was observed
Ž .using xenon excitation and FITC emission filters. B Electrophysiological recordings between pairs of hippocampal neurons. Junctional currents recordedin response to 60 and 100 mV transjunctional voltage pulses do not exhibit significant relaxation. Arrow indicates the occurrence of spontaneous synaptic
Ž . Ž .currents in these neurons. C The relationship between steady-state junctional conductance G and the transjunctional voltage of hippocampal neuronsj,SSŽ . Ž w x .in four cell pairs. Methods for dye transfer: LY CH, a highly fluorescent anionic dye MW 454 Da 5% wtrvol in 150 mM LiCl , was injected through a
Ž .microelectrode ;20 MV; filled with 3 M KCl into one cell of a cluster using hyperpolarizing current pulses and was visualized using a Nikon Diaphotinverted microscope equipped with xenon epifluorescence illumination and FITC filter set. Phase contrast and fluorescence micrographs were recorded 1–2
w xmin after dye injection by exposing Kodak TMAX 400 film. Adapted with permission from Ref. 53 .
recordings from pairs of cultured hippocampal neuronsrevealed a low degree of coupling in 8% of the pairsexamined. The average junctional conductance in these
Ž .cell pairs was ;1.5 nS Fig. 12B and, like the Cx36transfectants, they exhibited a clear insensitivity to trans-
Ž .junctional voltage Fig. 12C . The lack of voltage sensitiv-ity of Cx36 gap junction channels may provide a mecha-nism to prevent uncoupling during neuronal activity orneurogenesis, commensurate with the establishment of hightransmembrane potentials.
6. Conclusions
In this review, we have outlined the temporal expres-sion of gap junction-mediated intercellular communicationin differentiating murine hippocampal neurons, the uniquefunctional properties of Cx36 and its expression pattern inthe hippocampus during brain ontogeny. Based on ourprevious results and those of others, we speculate that thecoordinate regulation of ID-1 and GAS5 are essential for
regulating key cell cycle checkpoints and neuronal differ-entiation and that these and other regulators of cell cyclemay interact with specific connexins that are involved insimilar developmental transitions. Our results show thatearly in the development of the CNS, neuronal precursorcells are extensively coupled by gap junction channelscomposed of Cx43 and that uncoupling is necessary topermit cells to exit cell cycle and differentiate; thereafter,progressive expression of gap junction channels composedof Cx40, Cx33, Cx45 and Cx36 influences further neu-ronal differentiation and circuit formation.
Acknowledgements
We thank Dr. Harris Ripps for providing us with theCx35 antibody and Ms. F. Andrade, Ms. E. Craig and Mr.H. Rubin for technical assistance. This work was sup-ported by grants from the National Institutes of HealthŽ .R.R., M.F.M., J.A.K and D.C. Spray , by a fellowship
Ž .from the American Heart Association M.S. and by the

( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–7170
ŽKirby Foundation through a generous grant to the Kennedy.Center, AECOM .
References
w x1 P.J. Andres-Barquin, M.C. Hernandex, M.A. Israel, Id4 expressioninduces apoptosis in astrocytic cultures and is down-regulated byactivation of the cAMP-dependent signal transduction pathway, Exp.
Ž .Cell Res. 247 1999 347–355.w x2 D.M. Araujo, C.W. Cotman, Trophic effects of interleukin-4, -7, and
-8 on hippocampal neuronal cultures: potential involvement of glial-Ž .derived factors, Brain Res. 600 1993 49–55.
w x3 D.K. Batter, R.A. Corpina, C. Roy, D.C. Spray, E. Hertzberg, J.A.Kessler, Heterogeneity gap junction expression in astrocytes cultured
Ž .from different brain regions, Glia 6 1992 213–221.w x4 L.C. Barrio, T. Suchyna, T. Bargiello, L.X. Xu, R.S. Roginski,
M.V.L. Bennett, B.J. Nicholson, Gap junctions formed by connexins26 and 32 alone and in combination are differently affected by
Ž .applied voltage, Proc. Natl. Acad. Sci. U.S.A. 88 1991 8410–8414.w x5 R. Benezra, R.L. Davis, D. Lockshon, D.L. Turner, H. Weintraub,
The protein Id: a negative regulator of helix–loop–helix DNAŽ .binding proteins, Cell 6 1990 49–59.
w x6 D.A. Beblo, R.D. Veenstra, Monovalent cation permeation throughthe connexin40 gap junction channel Cs, Rb, K, Na, Li, TEA, TMA,TBA and effects of anions Br, Cl, F, acetate, aspartate, glutamate,
Ž .and NO , J. Gen. Physiol. 109 1997 509–522.3w x7 F.F. Bukauskas, C. Elfgang, K. Willecke, R. Weingart, Biophysical
properties of gap junction channels formed by mouse connexin40 ininduced pairs of transfected human HeLa cells, Biophys. J. 68Ž .1995 2289–2298.
w x8 D.F. Condorelli, R. Parenti, F. Spinella, A. Trovato Salinaro, N.Belluardo, V. Cardile, F. Ciciata, Cloning of a new gap junction
Ž .gene Cx36 highly expressed in mammalian brain neurons, Eur. J.Ž .Neurosci. 10 1998 1202–1208.
w x9 R. Dermietzel, Molecular diversity of gap junction expression inŽ .brain tissues, in: D.C. Spray, R. Dermietzel Eds. , Gap Junctions in
The Nervous System, Landes Bioscience Publishers, TX, 1996, pp.13–38.
w x Ž .10 R. Dermietzel, D.C. Spray, From neuro-glue ‘‘Nervenkitt’’ to glia:Ž .a prologue, Glia 24 1998 1–7.
w x11 R. Dermietzel, D.C. Spray, Gap junctions in the brain: where, whatŽ .type, how many and why?, TINS 16 1993 186–192.
w x12 R. Dermietzel, O. Traub, T.K. Hwang, E. Beyer, M.V.L. Bennett,D.C. Spray, K. Willecke, Differential expression of three gap junc-tion proteins in developing and mature brain tissues, Proc. Natl.
Ž .Acad. Sci. U.S.A. 86 1989 10148–10152.w x13 M. Duncan, E.M. DiCicco-Bloom, X. Xiang, R. Benezra, K. Chada,
The gene for the helix–loop–helix protein, Id, is specifically ex-Ž .pressed in neural precursors, Dev. Biol. 154 1992 1–10.
w x14 E.J. Furshpan, D.D. Potter, Low-resistance junctions between cellsŽ .in embryos and tissue culture, In: A. Moscona, A. Monroy Eds. ,
Current Topics in Developmental Biology, Vol. 3, Academic Press,New York, 1968, pp. 95–127.
w x15 S. Gokhan, Q. Song, M.F. Mehler, Generation and regulation ofdeveloping immortalized neural cell lines, Methods: A Companion
Ž .to Methods in Enzymology 16 1998 345–358.w x16 E.S. Gonos, Expression of the growth arrest specific genes in rat
embryonic fibroblasts undergoing senescence, Ann. N. Y. Acad. Sci.Ž .851 1998 466–469.
w x17 Y. Jen, K. Manova, R. Beneza, Each member of the Id gene familyexhibits a unique expression pattern in mouse gastrulation and
Ž .neurogenesis, Dev. Dyn. 208 1997 92–106.w x18 Y.T. Ju, A.C. Chang, B.R. She, M.L. Tsaur, H.M. Hwang, C.C.
Chao, S.N. Cohen, S. Lin-Chao, gas7: a gene expressed preferen-tially in growth-arrested fibroblasts and terminally differentiated
Purkinje neurons affects neurite formation, Proc. Natl. Acad. Sci.Ž .U.S.A. 95 1998 11423–11428.
w x19 K. Kandler, L.C. Katz, Relationship between dye coupling andspontaneous activity in developing ferret visual cortex, Dev. Neu-
Ž .rosci. 20 1998 59–64.w x20 V.G. Kukekov, E.D. Laywell, L.B. Thomas, D.A. Steindler, A
nestin-negative precursor cell from the adult mouse brain gives riseŽ .to neurons and glia, Glia 21 1997 399–407.
w x21 T.C. Lee, L. Li, L. Philipson, E.B. Ziff, Myc represses transcriptionof the growth arrest gene gas1, Proc. Natl. Acad. Sci. U.S.A. 94Ž .1997 12886–12891.
w x22 R. Li, J. Chen, G. Hammonds, H. Phillips, M. Armanini, P. Wood,R. Bunge, P.J. Godowski, M.X. Sliwkowski, J.P. Mather, Identifica-tion of Gas6 as a growth factor for human Schwann cells, J.
Ž .Neurosci. 16 1996 2012–2019.w x23 R. Llinas, R. Baker, C. Sotelo, Electrotonic coupling between
Ž .neurons in cat inferior olive, J. Neurophysiol. 37 1974 560–571.w x24 J.J. Lo Turco, A.R. Kriegstein, Clusters of coupled neuroblasts in
Ž .embryonic neocortex, Science 252 1991 563–566.w x25 W.R. Lowenstein, Junctional intercellular communication and the
Ž .control of growth, Biochem. Biophys. Acta 560 1979 1–65.w x26 D. Lyden, A.Z. Young, D. Zagzag, W. Yan, W. Gerald, R. O’Reilly,
B.L. Bader, R.O. Hynes, Y. Zhuang, K. Manova, R. Benezra, Id1and Id3 are required for neurogenesis, angiogenesis and vasculariza-
Ž .tion of tumor xenografts, Nature 401 1999 670–677.w x27 B. MacVicar, E.F. Dudek, Electrotonic coupling between pyramidal
cells: a direct demonstration in rat hippocampal slices, Science 213Ž .1981 782–784.
w x28 M.R. Mark, J. Chen, R.G. Hammonds, M. Sadick, P.J. Godowsk,Characterization of Gas6, a member of the superfamily of G do-main-containing proteins, as a ligand or Rse and Axl, J. Biol. Chem.
Ž .271 1996 9785–9789.w x29 M.F. Mehler, J.A. Kessler, Hematolymphopoietic and inflammatory
Ž .cytokines in neural development, Trends Neurosci. 20 1997 357–365.
w x30 M.F. Mehler, P.C. Mabie, D. Zhang, J.A. Kessler, Bone morpho-Ž .genetic proteins in the nervous system, Trends Neurosci. 20 1997
309–317.w x31 M.F. Mehler, R. Marmur, R. Gross, P.C. Mabie, A. Zang, A.
Papavasiliou, J.A. Kessler, Cytokines regulate the cellular phenotypeof developing neural lineage species, Int. J. Dev. Neurosci. 13Ž .1995 213–240.
w x32 M.F. Mehler, R. Rozental, M. Dougherty, D.C. Spray, J.A. Kessler,Cytokine regulation of neuronal differentiation of hippocampal pro-
Ž .genitor cells, Nature 362 1993 62–65.w x33 M.D. Michaelson, M.F. Mehler, H. Xu, R.E. Gross, J.A. Kessler,
Interleukin-7 is trophic for embryonic neurons and is expressed inŽ .developing brain, Dev. Biol. 179 1996 251–263.
w x34 A.P. Moreno, G.I. Fishman, E.C. Beyer, D.C. Spray, Voltage depen-dent gating and single channel analysis of heterotypic gap junctionchannels formed of Cx45 and Cx43, in: Y. Kanno, K. Kataoka, Y.
Ž .Shiba, Y. Shibata, T. Shimazu Eds. , Intercellular CommunicationŽ .Through Gap Junctions, Prog. Cell Res. 4 1995 405–408.
w x35 S.J. Morrison, N.M. Shah, D.J. Anderson, Regulatory mechanismsŽ .in stem cell biology, Cell 88 1997 278–298.
w x36 M. Nedergaard, Direct signaling from astrocytes to neurons inŽ .cultures of mammalian brain cells, Science 263 1996 1768–1771.
w x37 G.A. Nicholson, L. Yeung, A. Corbett, Efficient neurophysiologicselection of X-linked Charcot–Marie–Tooth families: ten novel
Ž .mutations, Neurology 511 1998 1412–1416.w x38 J. O’Brien, M.R. Al-Ubaidi, H. Ripps, Connexin 35: a gap-junc-
tional protein expressed preferentially in the skate retina, Mol. Biol.Ž .Cell 7 1996 233–243.
w x39 J. O’Brien, R. Bruzzone, T.W. White, M.R. Al-Ubaidi, H. Ripps,Cloning and expression of two related connexins from the perchretina define a distinct subgroup of the connexin family, J. Neurosci.
Ž .18 1998 7625–7637.

( )R. Rozental et al.rBrain Research ReÕiews 32 2000 57–71 71
w x40 M. Ogawa, Differentiation and proliferation of hematopoietic stemŽ .cells, Blood 81 1993 2844–2853.
w x41 S. Oh, J.B. Rubin, M.V. Bennett, V.K. Verselis, T.A. Bargiello,Molecular determinants of electrical rectification of single channelconductance in gap junctions formed by connexins 26 and 32, J.
Ž .Gen. Physiol. 114 1999 339–364.w x42 A. Peinado, R. Yuste, L.C. Katz, Extensive dye coupling between
rat neocortical neurons during the period of circuit formation, Neu-Ž .ron 10 1993 103–114.
w x43 V. Riechmann, F. Sablitzky, Mutually exclusive expression of twoŽ .dominant-negative helix–loop–helix dnHLH genes, Id4 and Id3, in
the developing brain of the mouse suggests distinct regulatory rolesof these dnHLH proteins during cellular proliferation and differentia-
Ž .tion of the nervous system, Cell Growth Differ. 6 1995 837–843.w x44 R. Rozental, M.F. Mehler, M. Morales, A.F. Andrade-Rozental, J.A.
Kessler, D.C. Spray, Differentiation of hippocampal progenitor cellsin vitro: temporal expression of intercellular coupling and voltage-
Ž .and ligand-gated responses, Dev. Biol. 167 1995 350–352.w x45 R. Rozental, M. Morales, M. Urban, M.F. Mehler, M. Kremer, J.A.
Kessler, R. Dermietzel, D.C. Spray, Changes in the properties of gapjunctions during neuronal differentiation of hippocampal progenitor
Ž .cells, J. Neurosci. 18 1998 1753–1762.w x46 R. Rozental, D.C. Spray, Temporal expression of gap junctions
Ž .during neuronal ontogeny, in: D.C. Spray, R. Dermietzel Eds. , GapJunctions in The Nervous System, R.G. Landes, Austin, TX, 1996,pp. 261–277.
w x47 R. Rozental, M. Srinivas, D.C. Spray, How to close a gap junctionchannel: efficacies and potencies of uncoupling agents, in: R. Bruz-
Ž .zone, C. Giaume Eds. , Connexin Methods and Protocols, HumanaPress, 1999, in press.
w x48 C. Schneider, R.M. King, L. Philipson, Genes specifically expressedŽ .at growth arrest of mammalian cells, Cell 54 1988 787–793.
w x49 M. Schutte, S. Chen, A. Buku, J.M. Wolosin, Connexin50, a gapjunction protein of macrogliaP6n the mamalian retina and visual
Ž .pathway, Exp. Eye Res. 66 1998 605–613.
w x50 C.M. Smith, J.A. Steitz, Classification of gas5 as a multi-small-Ž . Xnucleolar-RNA snoRNA host gene and a member of the 5 -termi-
nal oligopyrimidine gene family reveals common features of snoRNAŽ .host genes, Mol. Cell. Biol. 18 1998 6897–6909.
w x51 G. Sohl, J. Degen, B. Teubner, K. Willecke, The murine gapjunction gene connexin36 is highly expressed in mouse retina and
Ž .regulated during brain development, FEBS Lett. 428 1998 27–31.w x52 D.C. Spray, E. Scemes, R. Rozental, Cell–cell communication via
Ž .gap junctions, in: Zigmond, Bloom, Landis, Roberts, Squire Eds. ,Fundamental Neuroscience, Academic Press, New York, NY, 1998,pp. 317–343.
w x53 M. Srinivas, R. Rozental, T. Kojima, R. Dermietzel, M. Mehler,D.F. Condorelli, J.A. Kessler, D.C. Spray, Functional properties ofchannels formed by the neuronal gap junction protein connexin36, J.
Ž .Neurosci. 19 1999 9848–9855.w x54 C.P. Taylor, F.E. Dudek, Synchronous neural afterdischarges in rat
hippocampal slices without active chemical synapses, Science 218Ž .1982 810–812.
w x55 S.F. Tzeng, J. deVellis, Id1, Id2 and Id3 gene expression in neuralŽ .cells during development, Glia 24 1998 372–381.
w x56 M. Urban, R. Rozental, D.C. Spray, Simple strategy for screeningconnexin-identity using RT–PCR assays, Braz. J. Med. Biol. Res.
Ž .32 1999 1029–1037.w x57 D.I. Vaney, Cell coupling in the retina, in: D.C. Spray, R. Dermiet-
Ž .zel Eds. , Gap Junctions in the Nervous System, R.G. Landes,Austin, TX, 1996, pp. 79–102.
w x58 R.D. Veenstra, H.Z. Wang, E.C. Beyer, P.R. Brink, Selective dyeand ionic permeability of gap junctions formed by connexin45, Circ.
Ž .Res. 75 1994 483–490.w x59 M.N. Waltzman, D.C. Spray, Exogenous expression of connexins
for physiological characterization of channel properties: comparisonŽ .of methods and results, Prog. Cell Res. 4 1995 9–17.
w x60 W.T. Wong, J.R. Sanes, R.O. Wong, Developmentally regulatedspontaneous activity in the embryonic chick retina, J. Neurosci. 18Ž .1998 8839–8852.
Copyright © 2022 FDOKUMEN




















![Role of desensitization of AMPA receptors on the neuronal viability and on the [Ca2+]i changes in cultured rat hippocampal neurons](https://static.fdokumen.com/doc/165x107/63230bd664690856e1096f90/role-of-desensitization-of-ampa-receptors-on-the-neuronal-viability-and-on-the-ca2i.jpg)