
Paleohistology of the bones of pterosaurs (Reptilia: Archosauria): anatomy, ontogeny, and...
37
Zoological Journal of the Linnean Society (2000), 129: 349–385. With 12 figures doi:10.1006/zjls 1999.0239, available online at http://www.idealibrary.com on Palaeohistology of the bones of pterosaurs (Reptilia: Archosauria): anatomy, ontogeny, and biomechanical implications ARMAND J. de RICQLE ` S E ´ quipe Formations Squelettiques, URA CNRS 11 37, Universite ´ Paris VII, case postale 70 77, 75251 Paris cedex 05, and Colle `ge de France, Paris, France KEVIN PADIAN∗ Department of Integrative Biology and Museum of Paleontology, University of California, Berkeley, CA 94720-3140, U.S.A. JOHN R. HORNER Museum of the Rockies, Montana State University, Bozeman, MT 59717, U.S.A. HE ´ LE ` NE FRANCILLON-VIEILLOT E ´ quipe Formations Squelettiques, URA CNRS 11 37, Universite ´ Paris VII, case postale 70 77, 75251 Paris cedex 05, France Received October 1998; accepted for publication March 1999 Thin sections from long bones of specimens representing pterosaurs ranging from the Early Jurassic to the latest Cretaceous provide a profile of bone histology across a range of sizes, skeletal elements, growth stages, and phylogenetic positions. Most pterosaur bone is fibro- lamellar, organized in an unusual way that suggests high growth rates through ontogeny. Fibro-lamellar deposits are finished by a relatively abrupt deceleration or cessation of growth represented by lamellar, poorly vascularized subperiosteal bone in what appear to be adults. Pterosaurs had the thinnest bone walls of any tetrapods; they complemented high rates of periosteal deposition with almost equally high rates of endosteal erosion. Pterosaurs show a great variety of histologic features that include articular calcified cartilage, sub-chondral bone plates, trabecular bone struts and related internal supports, and secondary deposition and remodeling of bone. They remodeled their bones internally by (1) depositing endosteal bone coatings on the inner cortex and over struts of pre-existing internal bone, (2) secondarily filling bone spaces, and (3) Haversian reworking. The construction of these struts reflects ∗ Corresponding author. Email: [email protected] 349 0024–4082/00/070349+37 $35.00/0 2000 The Linnean Society of London
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Paleohistology of the bones of pterosaurs (Reptilia: Archosauria): anatomy, ontogeny, and...

Zoological Journal of the Linnean Society (2000), 129: 349–385. With 12 figures
doi:10.1006/zjls 1999.0239, available online at http://www.idealibrary.com on
Palaeohistology of the bones of pterosaurs(Reptilia: Archosauria): anatomy, ontogeny,and biomechanical implications
ARMAND J. de RICQLES
Equipe Formations Squelettiques, URA CNRS 11 37, Universite Paris VII,case postale 70 77, 75251 Paris cedex 05, and College de France, Paris, France
KEVIN PADIAN∗
Department of Integrative Biology and Museum of Paleontology, University of California,Berkeley, CA 94720-3140, U.S.A.
JOHN R. HORNER
Museum of the Rockies, Montana State University, Bozeman, MT 59717, U.S.A.
HELENE FRANCILLON-VIEILLOT
Equipe Formations Squelettiques, URA CNRS 11 37, Universite Paris VII,case postale 70 77, 75251 Paris cedex 05, France
Received October 1998; accepted for publication March 1999
Thin sections from long bones of specimens representing pterosaurs ranging from the EarlyJurassic to the latest Cretaceous provide a profile of bone histology across a range of sizes,skeletal elements, growth stages, and phylogenetic positions. Most pterosaur bone is fibro-lamellar, organized in an unusual way that suggests high growth rates through ontogeny.Fibro-lamellar deposits are finished by a relatively abrupt deceleration or cessation of growthrepresented by lamellar, poorly vascularized subperiosteal bone in what appear to be adults.Pterosaurs had the thinnest bone walls of any tetrapods; they complemented high rates ofperiosteal deposition with almost equally high rates of endosteal erosion. Pterosaurs show agreat variety of histologic features that include articular calcified cartilage, sub-chondral boneplates, trabecular bone struts and related internal supports, and secondary deposition andremodeling of bone. They remodeled their bones internally by (1) depositing endosteal bonecoatings on the inner cortex and over struts of pre-existing internal bone, (2) secondarilyfilling bone spaces, and (3) Haversian reworking. The construction of these struts reflects
∗Corresponding author. Email: [email protected]
3490024–4082/00/070349+37 $35.00/0 2000 The Linnean Society of London

A. J. DE RICQLES ET AL.350
both developmental patterns of bone construction and biomechanical function. Alternatingplywood-like layers of bone, heretofore undescribed in tetrapods, provided strength, as didthe obliquely oriented system of reticular blood vessels in the bones. The distribution andontogenetic features of pterosaur bone tissues, when combined with other evidence, suggestgenerally high growth rates, high metabolic levels, altricial birth, and extended parental care.
2000 The Linnean Society of London
ADDITIONAL KEY WORDS:—Pterosauria – Archosauria – palaeohistology –palaeobiology.
CONTENTS
Introduction . . . . . . . . . . . . . . . . . . . . . . . 350Historical background . . . . . . . . . . . . . . . . . . . 351Material and methods . . . . . . . . . . . . . . . . . . . 353Abbreviations . . . . . . . . . . . . . . . . . . . . . . 355Descriptive material . . . . . . . . . . . . . . . . . . . . 355
Primary bone tissues of the cortex . . . . . . . . . . . . . . 355Secondary bone tissues of the cortex . . . . . . . . . . . . . 363Secondary bone tissues: cancellous bone and epiphyses . . . . . . . 366Summary: general histologic characters of pterosaur bone . . . . . . 370
Discussion . . . . . . . . . . . . . . . . . . . . . . . 372Functional significance of tissue types and their microanatomical
organization . . . . . . . . . . . . . . . . . . . . 372Integration of histologic and other evidence for growth rates and ontogeny . 375Implications for pterosaur palaeobiology . . . . . . . . . . . . 378
Conclusions . . . . . . . . . . . . . . . . . . . . . . . 381Acknowledgements . . . . . . . . . . . . . . . . . . . . 381References . . . . . . . . . . . . . . . . . . . . . . . 382
INTRODUCTION
Pterosaurs have always been perceived as unusual members of the Reptiliabecause, as flying animals, they did not fit the ‘creeping’ mode of locomotion forwhich the reptiles were named (Cuvier, 1809; Padian, 1987). As pterosaur anatomyand flight mechanics became better known, the extremely thin walls of their boneswere often noted as adaptations for flight, but the inferences drawn about thephysiology necessary for their flight were often conflicting (for example, contrastOwen, 1870 and Seeley, 1870; Padian, 1987, 1995). Nevertheless, the thinness ofpterosaur bone walls has traditionally been acknowledged by many authors (e.g.von Meyer, 1859–60; Owen, 1870; Seeley, 1870; Bramwell & Whitfield, 1974;Currey, 1984; Padian, 1985; Wellnhofer, 1991). Generally speaking, among in-dividuals of comparable sizes, the walls of pterosaur bones are even thinner thanthose of birds. Typical values of half a millimeter or less are given for long boneswith lengths of well over 100 mm (e.g. Bramwell & Whitfield, 1974), and bones ofsmaller specimens can be much thinner, ranging to less than 0.2 mm (Padian, unpub.data).
The purpose of this study is to examine some of the factors that contribute tothe thinness of pterosaur bone walls, and to explain them in histological, mechanical,ontogenetic, and functional terms. To do this, we first describe the various kinds ofbone tissues and related structures that we have examined in a variety of pterosaurbones. We describe these tissues in detail because up to now only scattered

PTEROSAUR BONE HISTOLOGY 351
observations of pterosaur bone, usually based on single thin-sections, have beenreported in the literature, and these were not generally framed in contemporaryterminology. Second, we interpret these tissues in the context of bone formationand growth. Third, we relate the structures of these bone tissues to their sources inthe skeleton and discuss their local function and mechanical construction. Finally,we compare these tissues in pterosaurs to those of other animals, both phylogeneticallyrelated (e.g. dinosaurs and crocodiles) and similar because of the functional demandsof flight (e.g. birds).
Studies of the bones of both extinct and extant tetrapods allow us to hypothesize thatthere are four principal determinants of osteohistologic form: ontogeny, phylogeny,biomechanics, and environment (Ricqles et al., 1997, 1998). Bone tissues throughoutthe skeleton should be described at as many stages of growth as can be madeavailable in taxa whose phylogenetic relationships are established on the basis ofother characters. Once variation in histology can be understood skeletally andontogenetically for a range of taxa placed in phylogenetic context, questions maybe asked more informatively about the influences of biomechanics and environmenton the histological form of bone.
The present study cannot be optimally comprehensive for several reasons. Ptero-saur skeletons are seldom complete, and even incomplete ones are prized by museumsand collectors. Complete ontogenetic sequences are not known for pterosaurs; thosefor which sequences are most complete are probably Rhamphorhychus and Pterodactylus,from the Solnhofen limestones of Bavaria (Bennett, 1995, 1996a,b), but thesespecimens are scattered among museums and are generally considered prohibitivelyvaluable for this kind of study. Instead, we have sampled a range of tissues throughthe skeleton of pterosaurs including Early Jurassic and Late Cretaceous forms. Thedata reported here are intended to provide baseline information on pterosaurhistology and to note features that may bear further investigation on functional,developmental, and biomechanical grounds.
HISTORICAL BACKGROUND
Investigations into the histological structures of pterosaur bone started quite early.Quekett (1849b, Pl. VIII: 2) illustrated a thin-section of bone tissue from the humeralshaft of a pterosaur as an example of the condition “in the higher orders of Reptilia”.His section ran through two successive layers of the characteristic “plywood-like”bone structure that we describe below (see also Quekett, 1855). Seitz (1907: 289–291)described in detail the poorly preserved bone tissues in a phalanx of the Jurassicpterosaur Rhamphocephalus bucklandi, and he drew attention to the detailed histologicand microanatomical convergences in bone organization between pterosaurs andbirds. He noticed the extreme thinness of the bone cortex and the great developmentof secondary endosteal bone, laid down unconformably under the thin periostealcortex. Seitz (1907: 350–352) found similar features in a Pteranodon phalanx fromthe Kansas chalk. He noted details that were generally similar to the histology oflarge flying birds. Although his material was badly crushed, he recognized thedevelopment and importance of the network of bony struts in the marrow cavityand their secondary endosteal structure.
Gross (1934: 759) clearly described the histology of a wing bone of Dorygnathus

A. J. DE RICQLES ET AL.352
banthensis (Early Jurassic). He recognized in the periosteal cortex an obliquely orientedreticulum of primary osteons that lack a truly lamellar organization, and that areembedded in a woven tissue of periosteal origin. Gross did not describe lamellar-zonal tissues as such at the periphery of the cortex, but he noted that blood vesselsare almost entirely absent in this layer, and that the bone cells take on a morecircular orientation. It is thus likely that Gross’ material came from a nearly adultindividual, as we will discuss later.
Enlow & Brown (1957: 199–200, Pl. XXI: 9; 1958: 226–227) figured the tissueof a Pteranodon rib and provided a short description of other skeletal elements andgeneral discussion. Like previous workers, they found a reticular network of primaryvascular canals in the periosteal cortex, similar to those in many birds (e.g. Numida:Enlow & Brown, 1957, Pl. XXVI: 7). Those tissues would clearly fall into thereticular category of the “fibro-lamellar complex”, according to Ricqles’ (1975)terminology (see e.g. Ricqles et al., 1991). However, Enlow and Brown also mentionedtissues in Pteranodon that would instead fit into the “lamellar-zonal” category ofRicqles (1975) or “Zonaren periostknochen” of Gross (1934). If these tissues werewidespread in their specimen, they would suggest that the specimen was an adult,as we will discuss below.
The general observations of pterosaurian bone histology developed in the worksquoted above have been summarized by Ricqles on several occasions (1976: 135;1977: 38; 1978: 90, 93; 1979: 21–22; Ricqles et al., 1991: 57), notably in the contextof the evolution of thermo-metabolic physiology among archosaurs. Wellnhofer(1991: 149, 162–163, 164) also provided a detailed overview of bone structures atthe microanatomical level and discussed related problems such as growth, epiphysealmechanisms, and pneumaticity.
Among more recent original work, Bennett (1993) published an excellent paperon the ontogeny of Pteranodon. Using several criteria such as the surface ornamentationof bones by vascular canals, the structure of epiphyseal surfaces, and bone histology,he was able to show that sub-adult Pteranodon individuals reached almost adultdimensions, and that their growth was probably fast but limited (determinate). Sub-adults have a bone cortex entirely made of fibro-lamellar tissue with a reticularpattern of vascularization, whereas adults have a poorly vascularized tissue thatforms both the external (subperiosteal) and internal (endosteal) surfaces of the cortex(Fig. 2C,D in Bennett, 1993). Bennett also studied the biometrics of size distributionin the Late Jurassic pterosaurs from the Solnhofen limestones of Germany, includingRhamphorhynchus (1995, 1996a), and Pterodactylus (1996a) and its allies. He concludedthat most Rhamphorhynchus and Pterodactylus specimens were of juveniles; all nominalRhamphorhynchus species fit a single growth trajectory and probably belonged toa single species; and though monospecificity was not found for the Solnhofenpterodactyloids, he was able to group their nominal species into four or five coherentgroupings, of which one (including Pterodactylus kochi and P. antiquus) showed a bimodaldistribution, which he also found in the Rhamphorhynchus data set. Bennett (1995,1996a) concluded that there were not several separate species within each genus,but age distributions (year classes) of probably a single valid species for each genus.This would imply growth that is not as rapid as in birds. Our present study supportsBennett’s (1993) view that growth in Pteranodon and other large pterodactyloids wasrapid and determinate. There is indeed, as Bennett (1995) notes, no direct histologicalevidence for Rhamphorhynchus or Pterodactylus to date. We have limited informationon pre-Cretaceous forms; what we do have suggests generally similar patterns of

PTEROSAUR BONE HISTOLOGY 353
growth among pterosaurs, but more data are needed to test these patterns. Forexample, the bone histology of Montanazhdarcho, a small azhdarchid pterosaur, revealsthat although it was relatively small for this lineage (wingspan about 3 meters, stilltwice as large as most pre-Cretaceous forms), it was almost certainly fully grown(Padian, de Ricqles & Horner, 1995), and in other respects its histological featuresare like those of other Cretaceous pterodactyloids. In general respects these patternsof apparently rapid growth are seen as far back as the Early Jurassic Dorygnathusdescribed by Gross (1934), but Dimorphodon appears to have a less specialized histology(see below).
More recently, several short works have demonstrated the utility of histology instudies of pterosaurs. Chiappe & Chinsamy (1996) used histologic analysis to concludethat the bristle-like teeth of Pterodaustro were true teeth, not cornified baleine-liketissue as had sometimes been suggested. Unwin et al. (1996) used similar techniquesto conclude that the pteroid bone of the wrist did not show features of ossifiedtendons, with which it had often been identified, but of true bone. However, it stillcannot be determined whether the pteroid is homologous to any particular carpalor to part of the first digit, or is a neomorph.
Finally, new collaborations among palaeontologists and biomechanicians of boneare carrying forward some of the insights pioneered by Currey (e.g. 1984) and otherson what factors determine the expression of bone structure. Consensus is developingthat mechanical factors alone do not determine the shape of bone, but also factorsrelated to phylogenetic legacy, genetic and developmental program, and environmentof the individual as well as the lineage (e.g. Carter, 1987; Carter, Wong & Orr,1991; van der Meulen, Padian & Carter, 1992; van der Meulen & Carter, 1995;van der Meulen, Beaupre & Carter, 1993; de Ricqles, Padian & Horner, 1997; Mikic,Carter & Padian, 1998). These factors are extremely relevant to the interpretation ofspecialized histologic structures observed among pterosaurs, but their details arebeyond the scope of the present study, in which we limit ourselves to suggestingbiomechanical hypotheses that analyses of further available materials may test.
MATERIAL AND METHODS
Long bone shafts and epiphyses were sectioned in a variety of pterosaurs includingthe Early Jurassic Dimorphodon, the early Late Cretaceous Ornithocheirus, the LateCretaceous Pteranodon and Montanazhdarcho, the latest Cretaceous Quetzalcoatlus, andseveral other Late Cretaceous elements that are of indeterminate (almost certainlypterodactyloid) taxonomic status (Table 1). Specimens were embedded in epoxyresin, ground and polished, and examined under ordinary and polarized light.Whenever possible, several sections from the same anatomical region were processed(e.g. transverse, longitudinal, and oblique), and similar bones from individuals ofdifferent sizes (and presumed ages) from the same taxon were compared. Standardsectioning, grinding, and polishing techniques, such as those described by Chinsamyand Raath (1992), were used, with modifications as appropriate. Our histologicaldescriptions are based on all these samples, though some are emphasized for clarityand brevity.
Our descriptions of pterosaur bone tissues are organized not by taxon but bytheir primary and secondary histologic features. For a review of these tissues, the
A. J. DE RICQLES ET AL.354
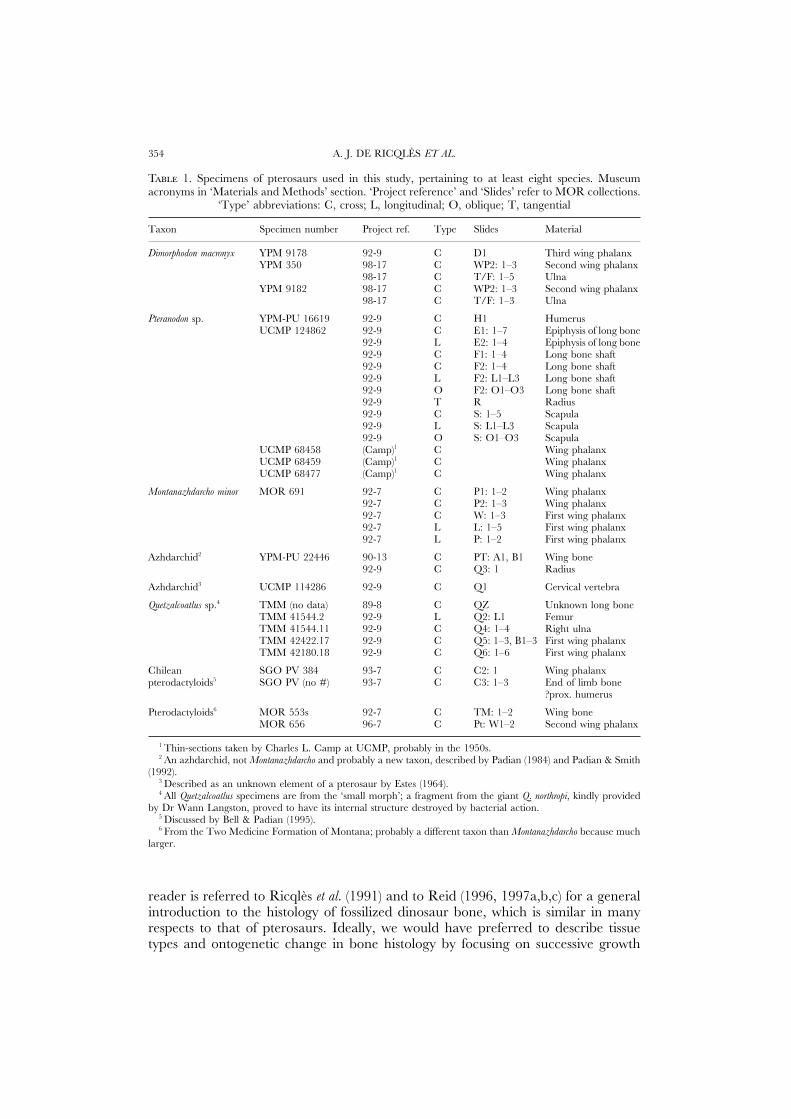
T 1. Specimens of pterosaurs used in this study, pertaining to at least eight species. Museumacronyms in ‘Materials and Methods’ section. ‘Project reference’ and ‘Slides’ refer to MOR collections.
‘Type’ abbreviations: C, cross; L, longitudinal; O, oblique; T, tangential
Taxon Specimen number Project ref. Type Slides Material
Dimorphodon macronyx YPM 9178 92-9 C D1 Third wing phalanxYPM 350 98-17 C WP2: 1–3 Second wing phalanx
98-17 C T/F: 1–5 UlnaYPM 9182 98-17 C WP2: 1–3 Second wing phalanx
98-17 C T/F: 1–3 Ulna
Pteranodon sp. YPM-PU 16619 92-9 C H1 HumerusUCMP 124862 92-9 C E1: 1–7 Epiphysis of long bone
92-9 L E2: 1–4 Epiphysis of long bone92-9 C F1: 1–4 Long bone shaft92-9 C F2: 1–4 Long bone shaft92-9 L F2: L1–L3 Long bone shaft92-9 O F2: O1–O3 Long bone shaft92-9 T R Radius92-9 C S: 1–5 Scapula92-9 L S: L1–L3 Scapula92-9 O S: O1–O3 Scapula
UCMP 68458 (Camp)1 C Wing phalanxUCMP 68459 (Camp)1 C Wing phalanxUCMP 68477 (Camp)1 C Wing phalanx
Montanazhdarcho minor MOR 691 92-7 C P1: 1–2 Wing phalanx92-7 C P2: 1–3 Wing phalanx92-7 C W: 1–3 First wing phalanx92-7 L L: 1–5 First wing phalanx92-7 L P: 1–2 First wing phalanx
Azhdarchid2 YPM-PU 22446 90-13 C PT: A1, B1 Wing bone92-9 C Q3: 1 Radius
Azhdarchid3 UCMP 114286 92-9 C Q1 Cervical vertebra
Quetzalcoatlus sp.4 TMM (no data) 89-8 C QZ Unknown long boneTMM 41544.2 92-9 L Q2: L1 FemurTMM 41544.11 92-9 C Q4: 1–4 Right ulnaTMM 42422.17 92-9 C Q5: 1–3, B1–3 First wing phalanxTMM 42180.18 92-9 C Q6: 1–6 First wing phalanx
Chilean SGO PV 384 93-7 C C2: 1 Wing phalanxpterodactyloids5 SGO PV (no #) 93-7 C C3: 1–3 End of limb bone
?prox. humerus
Pterodactyloids6 MOR 553s 92-7 C TM: 1–2 Wing boneMOR 656 96-7 C Pt: W1–2 Second wing phalanx
1 Thin-sections taken by Charles L. Camp at UCMP, probably in the 1950s.2 An azhdarchid, not Montanazhdarcho and probably a new taxon, described by Padian (1984) and Padian & Smith
(1992).3 Described as an unknown element of a pterosaur by Estes (1964).4 All Quetzalcoatlus specimens are from the ‘small morph’; a fragment from the giant Q. northropi, kindly provided
by Dr Wann Langston, proved to have its internal structure destroyed by bacterial action.5 Discussed by Bell & Padian (1995).6 From the Two Medicine Formation of Montana; probably a different taxon than Montanazhdarcho because much
larger.
reader is referred to Ricqles et al. (1991) and to Reid (1996, 1997a,b,c) for a generalintroduction to the histology of fossilized dinosaur bone, which is similar in manyrespects to that of pterosaurs. Ideally, we would have preferred to describe tissuetypes and ontogenetic change in bone histology by focusing on successive growth

PTEROSAUR BONE HISTOLOGY 355
stages (Horner, Ricqles & Padian, in prep.). Unfortunately, entire growth stages ofsufficiently complete pterosaur skeletons are not currently available, and so we havesampled a broad range of pterosaurs of different sizes, taxa, and geologic ages. Asa result, the determination of ontogenetic stage is based on several lines of histological(Castanet et al., 1993) and gross morphological (Bennett, 1993) criteria. As we explainbelow, these methods are quite powerful for determining both ontogenetic stageand developmental dynamics (see also Padian et al., 1995). Our observations areconsistent enough from section to section that we have been able to make and testgeneralizations about structure, growth, and possible biomechanical role.
ABBREVIATIONS
Figuresbl bright line m matrix (terrigenous, non-skeletal)cc calcified cartilage obl oblique anastomosis of vascular canalsccb coarse cancellous bone ost osteonefs external fundamental system pc peripheral cortexend endosteal bone ply ‘plywood’ layerser erosion room pn pneumatic channelfi fibrous (woven) bone rew reworked endosteal bonef-l fibro-lamellar bone s strut of mostly endosteal boneh secondary (Haversian) osteon sc, sub subchondral bone plateLAG line of arrested growth sh Sharpey’s fibreslam lamellar bone vc vascular canals
Institutional abbreviationsMCZ, Museum of Comparative Zoology, Harvard University, Cambridge; MOR,
Museum of the Rockies, Montana State University, Bozeman; SGO-PV, ServicioNacional de Geologia y Mineria, Santiago, Chile; TMM, Texas Memorial Museum,University of Texas, Austin; TMP, Tyrrell Museum of Palaeontology, Drumheller,Alberta, Canada; UCMP, University of California Museum of Paleontology, Berke-ley; YPM, Yale Peabody Museum, New Haven; YPM-PU, former Princeton Uni-versity vertebrate palaeontological collections now at YPM. Thin-sections listed inTable 1 are curated in the paleohistologic collection of the Museum of the Rockies.Duplicates of most of these, as well as other listed specimens, are housed at UCMP.
DESCRIPTIVE MATERIAL
Primary bone tissues of the cortex
Fibro-lamellar tissues in juvenilesExamples from two Late Cretaceous taxa, Azhdarchidae and Pteranodontidae,
show the basic features of bone histology at the juvenile stage. One, a small fragmentof ?azhdarchid pterosaur wing bone from the Two Medicine Formation of Montana

A. J. DE RICQLES ET AL.356
(Figs 1A–C, 5A), shows a rare example among pterosaurs of primary bone in theprocess of deposition, probably in a quite immature individual. The thin cortex isentirely of primary bone. The innermost cortex is progressively resorbed so as tobalance the outward growth in diameter of the marrow cavity, and no endostealbone is deposited. Most of this bone, up to its external, subperiosteal surface, ismade of densely vascularized fibro-lamellar tissue with a peculiar organization. Thebone can be described as a mixture of the laminar and reticular types, falling withinthe general fibro-lamellar tissue pattern (e.g. Ricqles et al., 1991). The vascular canalsare numerous and small in diameter. Their orientation is either longitudinal orradial but distinctly oblique and sinuous, with irregular anastomoses. The core ofthe laminae (equivalent to the “helle Mittelinie” described by Gross [1934] and toCurrey’s [1960, 1962] “bright line”; see Ricqles, 1975) have radial, oblique orirregular sinuous orientations and are formed of parallel-fibreed tissue, slightlyanisotropic under crossed nicols. Most of the bone tissue deposited around theseinitial cores of the laminae is woven and organized as poorly defined primary osteonslaid down centripetally around the vascular canals. This latter bone tissue is isotropicunder crossed nicols, which is characteristic of young, fast-growing bone in whichthe collagenous matrix is poorly organized spatially and has a more or less meshlikethree-dimensional organization. Numerous cell lacunae are exposed transversely incross section; they have an extensively developed system of radially oriented,branching canaliculi that is typical of pterosaur bone. Most of the primary osteonsare composed of a fibrous, woven material, oriented longitudinally, and not by aplywood of lamellae. Only the very thin coating of tissue that forms the walls of thevascular canals of the primary osteons has a more finely bundled structure, but itis more translucent and almost deprived of cell lacunae, and is still isotropic undercrossed nicols. On most parts of the bone surface, the vascular canals open directlyat the external (subperiosteal) surface of the bone; they form oblique, broad cleftsthat evidently had not yet acquired their full complement of osteonal deposition.The outermost cortex has a very thin layer of lamellar subperiosteal bone tissue,which forms a “fundamental external system” (Cormack, 1987), only in a small,localized region. This tissue is grossly lamellated; it contains some Sharpey’s fibresand almost no vascular canals. It originally may have covered a larger portion ofthe well-vascularized primary cortex, but the preservation is too poor to tell. Thereis absolutely no evidence of any kind of secondary bone erosion-reconstruction in
Figure 1. A–C, ?azhdarchid pterodactyloid, MOR PT-C, wing bone; cross-section of diaphysis, showingreticular fibro-lamellar tissue of the primary cortex in the process of deposition. Note vascular canalsopening at the surface in A. D, Pteranodon, UCMP 68459, manual phalanx; cross-section of diaphysis,showing primary periosteal cortex with numerous longitudinally oriented primary osteons and largeresorption spaces. E–H, Quetzalcoatlus, TMM 43433-17, first wing-phalanx; cross-section of diaphysis,showing primary cortical bone with fibro-lamellar reticular bone of periosteal origin. F, magnificationof E; note Currey’s ‘bright lines’ and anastomosing vascular canals. This part of the cortex has wellorganized laminae. A line of arrested growth can be seen near the periphery, and the bone is lessvascularized outside it; a transition to this more lamellar bone can be seen just below the LAG. H,magnification of G; the bone in this thicker part of the cortex is more typically reticular, suggesting ahigher growth rate. I & J, cross-section of a turkey vulture bone for comparison; note the well-developed laminar fibro-lamellar bone (compare with E and F), and endosteal bone in I. Scale bar is250 �m in A, D, E, G, and I, 100 �m in B, F, H and J, and 25 �m in C.
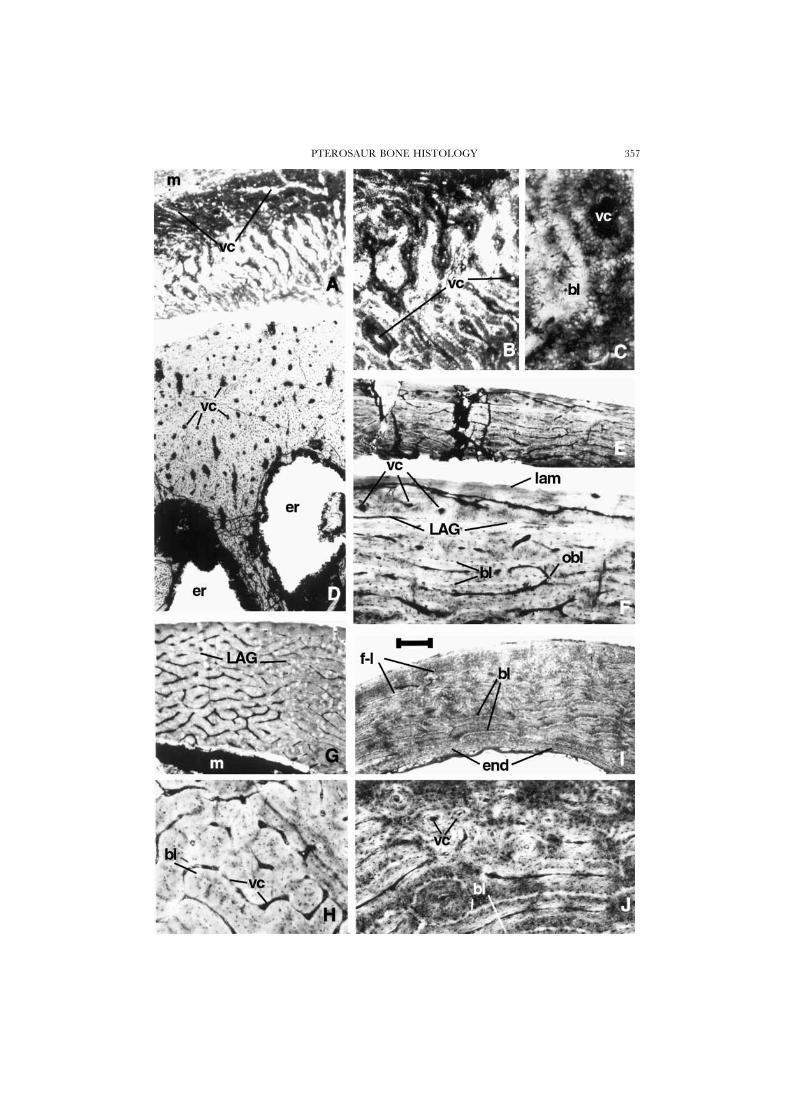
PTEROSAUR BONE HISTOLOGY 357

A. J. DE RICQLES ET AL.358
this section. These features clearly suggest a primary fast-growing tissue involved inthe formation of the cortex, presumably in a young individual, or at least in a fast-growing part of a bone.
A second sample that we analysed comprises thin sections made many years agoby Charles L. Camp of a Pteranodon finger bone (UCMP 68459). A cross sectionthrough the proximal portion of the shaft (Fig. 1D), although badly crushed, offersanother example of an almost complete circumferential section of a pterosaur bonewhose diameter is actively growing. One side of the bone is entirely formed by aprimary, compact, well-vascularized cortex of the general fibro-lamellar tissuepattern. The tissue is almost 2 mm thick and contains about ten superimposedprimary osteons counted radially. Each osteon is oriented longitudinally. Most ofthe tissue in which they are embedded is woven and isotropic under crossed nicols,as is most of the material that forms the osteons themselves. Hence a majority ofits bone fibres were longitudinally oriented. However, an important contingent ofcollagenous fibres is oriented radially in the section. These fibres cross and highlighttwo thin, circularly oriented regions within the cortex that may record two temporarychanges in the local speed of the radial deposition of bone.
From its inner side, the cortex is eroded by five large, regular, oval, outwardlygrowing expansions of the medullary cavity, of which two are visible at the base ofFigure 1D. The erosion of the deep cortex has a scalloped outline; no endostealbone is laid down locally, and there is no trace of Haversian erosion and redeposition.What is left of the deep primary cortex between the erosion cavities becomes thebasis of large, radially oriented struts of bone, which are overlain by new endostealbone along their lengths. In the marrow cavity, these struts are entirely secondarilyendosteal, with extensive erosion-reconstruction cycles taking place. Some of thestruts are locally hollow, and have central vascular canals that are surrounded bycircular, centripetal deposits of endosteal tissue. These struts vary in bone wallthickness along their lengths, from as much as one-third of the diameter of the strutnear its base (Fig. 4C) or greater (Fig. 5E) down to a very thin wall, much like theaspect of the long bones themselves. These struts evidently faced a substantial riskof local buckling, and this may explain their presence and distribution throughoutthe bone (Karam & Gibson, 1994). On the other surface of the bone (Fig. 4A,B),however, the struts meet the primary cortex with a very different structure. Thewide bases of the struts still contain several primary osteons, but the cortex itself isformed of a plywood-like lamellar tissue with a less dense vascularization, as seenin sections taken from the scapula of another Pteranodon (UCMP 124862; Fig. 3A,B; see below).
Figure 2. A, C–E, Pteranodon, UCMP 124862, bone fragment; laminar bone showing Currey’s ‘brightlines’. C, magnification of A; E, magnification of D, showing three distinct layers of deposition: innerendosteal bone, a cortex of poor reticular organization with irregular anastomoses, and an outer layerof lamellar bone. B, Pteranodon, UCMP 68458, a tangential section showing network of anastomosingcanals in reticular tissue. F, Quetzalcoatlus sp., TMM, cross-section of wing bone, showing irregularlyanastomosing vascular canals, and primary osteons. G, azhdarchid, MOR 553s, bone fragment showingwell formed laminar bone. H, Dimorphodon, YPM 9178, cross-section of wing-phalanx; outer lamellarbone overlies a thick fibro-lamellar cortex and beneath this is a thin region of coarse compact cancellousbone tissue; endosteal bone rims the medullar cavity. Scale bar is 250 �m in A, B, and D, and 100 �min C and E–H.
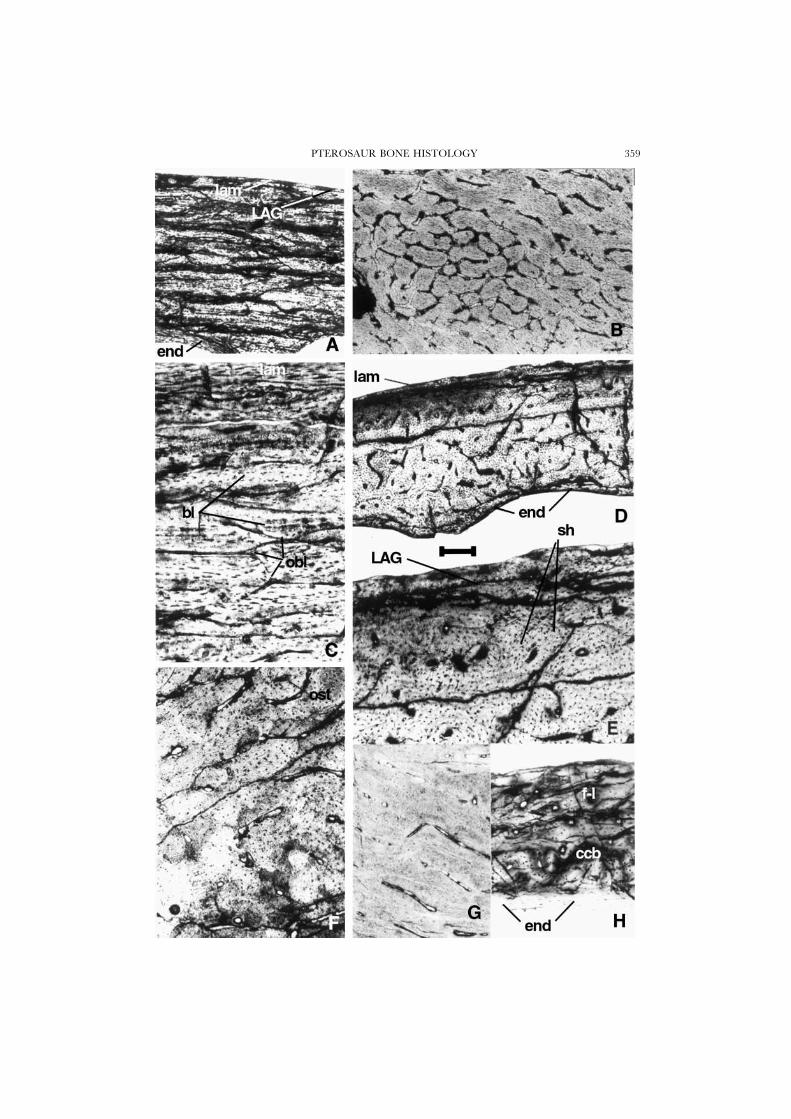
PTEROSAUR BONE HISTOLOGY 359

A. J. DE RICQLES ET AL.360
Fibro-lamellar tissues in sub-adults and adultsThese tissues come from the regular cortex, matching previously published
descriptions (Gross, 1934; Enlow & Brown, 1957). The long bone shafts of Pteranodon(Fig. 2C) and Quetzalcoatlus (Fig. 1E–H) appear to have roughly similar structures.The cortex is thin and compact, formed of two or three distinct sheets of bone (Fig.2A,D,E). Endosteally, a thin coating of avascular, lamellar or (most often) parallel-fibred endosteal bone tissue is separated from the periosteal bone tissue by a line ofresorption. This endosteal bone occasionally intrudes into the marrow cavity, asdoes the periosteal bone above it, and these structures form the bases of the birdlikestruts that cross the medullary cavity.
A primary periosteal tissue forms the bulk of the cortex thickness (Fig. 2A,D,E);a comparison to similar structures in the turkey vulture (Fig. 1I, J) shows that thisbone fits the general fibro-lamellar pattern seen in birds. Up to ten laminae can besuperimposed. The ‘bright line’ (Currey, 1960, 1962) that forms the core of eachlamina is isotropic under crossed nicols, as is most of the tissue (compare Figs 1Jand 2C). In cross-sections the vascular canals run mostly circularly as well aslongitudinally, with numerous oblique anastomoses among the circular canals ofsuperposed laminae (Fig. 2D). Hence, the vascularization varies locally from rathertypically laminar to reticular (Fig. 1E–H). However, there seems to be a distincttendency for the vascular canals to take a somewhat oblique course, both laterallyand vertically, relative to tangential and cross (vertical) sections, respectively. Thismay account for the diagonally oriented (geodetic) pattern described in the compactbone of some pterosaurs (Gross, 1934), and also observed in tangential (horizontal)sections in the cortex (e.g. Pteranodon: UCMP 68477, 68458, 68459: Fig. 2B). Incross-section, most of the cell lacunae of the primary bone tissue are cut transversely(Fig. 2F). But the lacunae are mostly cut longitudinally in the parallel-fibred materialthat forms the innermost deposit of the endosteal bone lining the marrow cavity.In oblique cross-sections, set at 45° from the vertical, all the cells from the periostealtissue appear to be oriented longitudinally, or slightly obliquely, and parallel to eachother. This is confirmed by longitudinal vertical sections of the cortex, in which allthe cell lacunae of the periosteal tissue appear elongated and spindle-shaped, whilethose of the endosteal bone tissue are now cut transversely.
The laminar cortex just described may be divided by one or a few lines of arrestedgrowth (LAGs) parallel to the external surface of the cortex. They appear as thin
Figure 3. A & B, Pteranodon, UCMP 124862, scapula; A, cross-section showing plywood constructionof the bone, in which a few simple primary vascular canals can be seen. Bone fibres alternately runtransversely and longitudinally; B, oblique section. Notice how the dashlike small osteons, cut inoblique sections, change orientation from one plywood layer to the next (compare to Fig. 5G, inpolarized light). C & D, Montanazhdarcho, MOR 691, cross-section of ?ulna; alternating plywood layers,overlying endosteal bone (lower right corner in C, lowest layer in D). E, Ornithocheirus, YPM 537A,cross-section of wing bone shaft. F, Montanazhdarcho, MOR 691, cross-section of wing-phalanx; theouter primary cortex has been reworked internally by secondary Haversian cells, and erosion has theneaten away some of this tissue, finally depositing an inner layer of endosteal bone. G–J, Quetzalcoatlussp., TMM, cross-section of wing bone, a transition zone showing both primary and secondary osteons(less maturely formed in lower left corner of photo in G). H shows mostly compacted coarse cancellousbone. I and J show erosion rooms with fragments of original trabecular bone struts separating them;these rooms are being filled in by primary bone, but may be remodelled by secondary (Haversian)osteons later on. Scale bar is 250 �m in J, 100 �m in A–I.
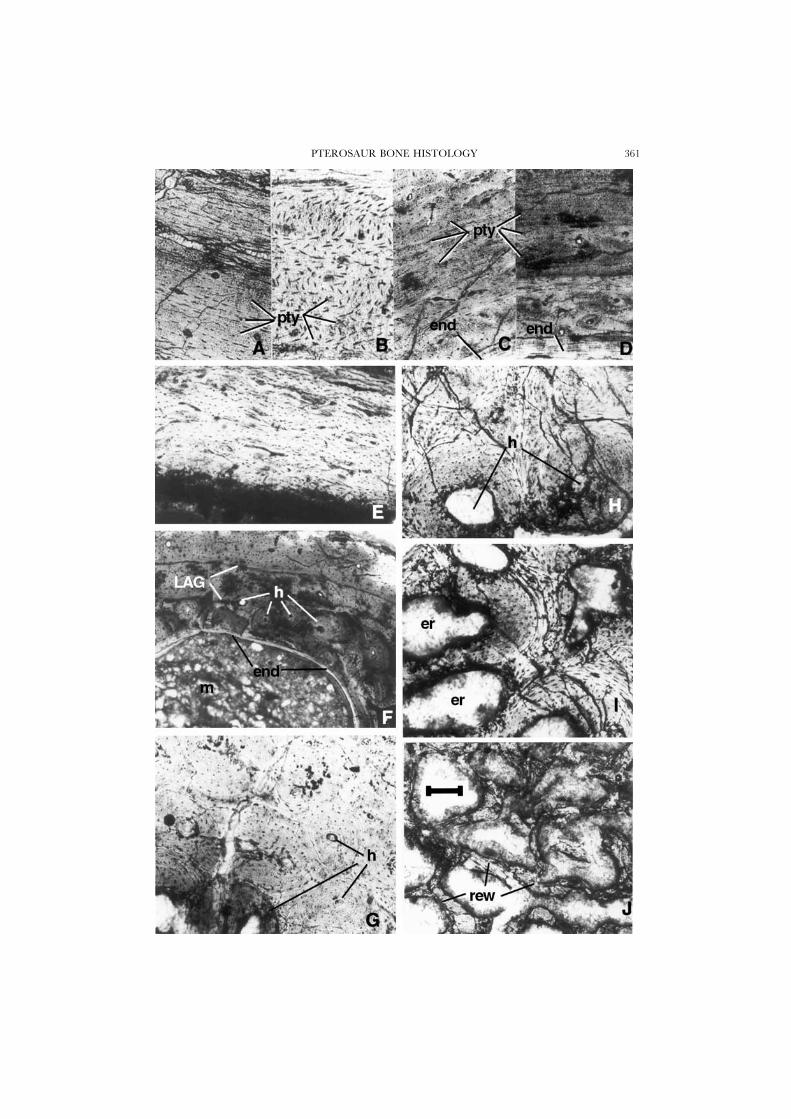
PTEROSAUR BONE HISTOLOGY 361

A. J. DE RICQLES ET AL.362
discontinuities, undulating close to the periphery of the local primary osteons. Theprimary periosteal cortex may contain Sharpey’s fibres.
In some regions, the fibro-lamellar cortex just described is covered by a thincoating of almost avascular lamellar or parallel-fibred bone tissue, deposited sub-periosteally (Fig. 2D,E). This tissue is separated from the subjacent cortex by aresting line. The cell spaces and fibrillar orientation of this external coating of bonegenerally agree with those of the subjacent bone, and are orthogonal to those ofthe endosteal bone deposited around the marrow cavity.
Other regions of compact bone in Pteranodon (Fig. 2A,C–E) and Ornithocheirus (YPM537 B,C; MCZ 2437) have somewhat different structures, especially in smallerbones. In the fibro-lamellar complex that forms the bulk of the cortex, no laminarorganization is apparent. Most of the primary osteons are oriented longitudinally,and are united by irregular, oblique anastomoses that give the tissue a distinctlyreticular pattern. Both the woven tissue between the osteons and the tissue that formsthe osteons themselves are coarsely woven (Fig. 2F). Most fibres run longitudinally andin cross section the tissues appear isotropic between crossed nicols. A few LAGsmay be present and the tissue is coated periosteally and endosteally by thin sheetsof non-vascular, parallel-fibred bone tissue. The external part of the periosteal cortexof a third wing-phalanx of Dimorphodon (Fig. 2H) is similar to this, but most primaryosteons are longitudinal. The external (periosteal) cortex accounts for one-half totwo-thirds of the total thickness of the cortex. It is formed by poorly-defined parallel-fibred bone tissue that is organized longitudinally, with numerous, very small,longitudinal primary osteons forming three to five radial rows. The most externalwall of the cortex is locally formed by an extremely thin coating of lamellar tissue.Endosteal bone is well developed around the marrow cavity and its fibres runcircularly. There are no struts of bone in this part of the cavity of this wing phalanx.Between the periosteal cortex and the endosteal coating of the marrow cavity justdescribed, the remains of a former osteogenetic stage has been partially resorbedboth outwardly by sub-periosteal osteoclasts, and inwardly by endosteal osteoclasts.This older bone is a compacted coarse cancellous tissue that contains a few secondaryosteons. This structure suggests remodeling that occurred when regions formerly inthe metaphyseal regions were relocated into what became the diaphysis as the bonegrew (Fig. 2H: ccb; Enlow, 1963).
Plywood-like and lamellar-zonal primary bone tissuesExamples of these bone tissues were taken from a scapula of Pteranodon (Fig. 3A,
B: UCMP 124862). Similar structures are also developed in sections representinglimb bones and a vertebra from three different azhdarchids (Table 1) that also showreticular vascularization in the ‘plywood’ layers. These structures are also presentin Montanazhdarcho (Padian, Ricqles & Horner, 1995).
Some parts of fairly large bones (Pteranodon, UCMP 68459) and smaller boneshave a cortex formed of peculiar primary periosteal tissue. This tissue fits the generallamellar-zonal pattern (e.g. Ricqles et al., 1991) and can be described as a biologicalplywood (e.g. Meunier & Castanet, 1982), heretofore not reported in tetrapods.Each ply is relatively thick and is formed by homogeneous, dense, parallel-fibredbone (Fig. 3A–C; compare to Figure 5G in polarized light for emphasis). The cellsare spindle-shaped and oriented according to the orientation of the fibres in theply. Locally, vascular canals of small diameter cross the ply. They are primary

PTEROSAUR BONE HISTOLOGY 363
simple vascular canals because they do not have rings of bony material depositedcentripetally around them (Enlow, 1966). From one ply to the next, the orientationof the bone fibres changes about 90°, as also seen by the change in orientation ofthe bone cells (Fig. 5G). This tissue qualifies as an orthogonal plywood as definedby Meunier & Castanet (1982; see Fig. 3C, Montanazhdarcho). In transverse cross-section, the cell lacunae of the plies are roughly circular, and have an extensivesystem of canaliculi. In a cross-section examined between crossed nicols, the plieswith longitudinally oriented fibres are highly anisotropic, whereas those with thefibres cut transversely are isotropic. Oblique sections at 45° in the structure confirmthe alternate, perpendicular orientations of the fibres and cells from one ply to thenext, and hence the orthogonal construction of the plywood (see e.g. Girauld et al.,1978). This bone tissue cannot merely be described as simple lamellar bone becauseeach ply is considerably thicker than the lamellae of ordinary lamellar bone. Theplywood tissue just described is sometimes completed by the deposition of a thincoat of finely fibred endosteal tissue around the medullary cavity.
Very thin bones (e.g. Montanazhdarcho) have a simpler structure (Fig. 3C,D). Thecortex is composed of parallel-fibred tissue, with scattered vascularization of thereticular type consisting of simple primary vascular canals. Groups of circularlyoriented bone fibres scattered throughout the cortical thickness give the overallcortex a poorly defined, plywood-like structure. LAGs may also be present in thiskind of tissue.
The separation between the various tissue types is not always distinct in pterosaurs.Several sections from shafts of various small long bones (diameter about 1 cm)referred to Ornithocheirus from the Cambridge Greensand (Fig. 3E) show combinationsof the various tissue types so far described. The periosteal cortex contains vascularcanals that are longitudinally and circularly oriented, with an oblique, reticularpattern. These canals form small primary osteons embedded in a woven matrix, asshown by the orientation of the osteocytes. This tissue is interrupted by up to threebeds of fibres that are oriented circularly, and are anisotropic under crossed nicols,giving the cortex an overall plywood structure. Locally, this cortex takes on a typicallamellar-zonal pattern. Primary osteons are longitudinal and a great number ofradially oriented Sharpey’s fibres pervade the cortex. Two or three LAGs arepresent. This slow-growing lamellar-zonal tissue may be older than the fibro-lamellartissue laid down above it. Cross-sections of the shaft also show that the bone tissuemay differ from one side of the bone to the other: for example, the regions of fast-growing (fibro-lamellar) cortex are roughly diametrically opposed to those withLAGs and many Sharpey’s fibres. Similarly, the slower-growing regions of the cortexseem to be associated with many more endosteal spongiosa struts in the marrowcavity. Finally, in some regions, the fibres of cortical bone are circularly orientednot only in the most external part of the cortex (Gross, 1934), but almost throughoutthe entire periosteal cortex.
Secondary bone tissues of the cortex
Secondary bone tissue replaces, repairs, or remodels primary tissue, and pterosaurstended to deposit secondary bone in many different bones and at many stages ofgrowth.

A. J. DE RICQLES ET AL.364
Dense Haversian tissueWe studied several small cross-sections of cylindrical bones from azhdarchid
pterosaur remains from the Late Cretaceous Two Medicine Formation (Padian etal., 1995; Fig. 3F) and the Judith River Formation (Padian, 1984; Padian & Smith,1992; Fig. 3G). The sections are through large marrow cavities (cancellae) lined bywalls of endosteal bone. In several places (Fig. 3G), there are remains of periostealbone tissue that have either a plywood-like structure or a structure with numerousprimary osteons cut longitudinally, with little anastomosing among them. In eachregion where cortical bone becomes thick, notably where the endosteal struts meetthe deep cortex, the cortex is reworked by a group of typical secondary osteons.Each group is formed by two to eight secondary osteons, with variable amounts ofcentripetal deposition. In some osteons, the central Haversian canal is almostobliterated by secondary deposition. The cell lacunae are cut transversely and areorganized in the osteon at random, not as concentric rows of cells. There is clearlyno lamellar organization of the osteon because it is entirely isotropic under crossednicols. Hence, the bone fibres would all be oriented longitudinally, apart from themost external region of each secondary osteon, with few circularly oriented fibres.In Montanazhdarcho (MOR 691; Padian et al., 1995), a somewhat similar developmentof Haversian bone tissue is observed (Fig. 3F). The secondary osteons are orientedlongitudinally and form clusters that are especially well developed in the deep cortex,at the border between its deep periosteal and endosteal components. The secondaryosteons appear especially numerous where the endosteal struts that form themedullary spongiosa meet the cortex.
Compacted coarse cancellous tissueThis tissue type was studied in a wing-phalanx of the ‘small morph’ of the giant
azhdarchid pterodactyloid Quetzalcoatlus (Fig. 3H,I; a similar compacted coarse-cancellous tissue also forms the inner cortex in the metaphyses of Dimorphodon [Fig.2H]). The cortex is composed of dense bone tissues. The innermost cortex mergesprogressively with a well-developed ‘traditional-looking’ spongiosa (Fig. 3J) througha transitional endosteal margin, with erosion rooms that appear as large holes inthe deep cortex; some of them are reconstructed by incipient secondary osteons. Insome places, the deep cortex is formed by dense Haversian bone built over formertissues (Fig. 3G). However, a large part of the cortex is formed of compacted coarsecancellous tissue, with a false overall appearance of Haversian bone (Fig. 3H,I),although some true secondary osteons are indeed present in this tissue. A primary(periosteal) cortex is well-developed only in one region, where numerous primary
Figure 4. A & B, Montanazhdarcho, MOR 691, cross-section of wing-phalanx; the relatively poorlyvascularized mature cortex is seen at the top of A and at the left of B; secondary osteons, partly erodedtoward the medullar cavity, are covered by an internal layer of endosteal bone that will form theinternal struts of the cavity. C, Pteranodon, UCMP 124862, oblique section of epiphyseal fragmentshowing a section through a bone strut, formed of layers of secondary reworked endosteal bonesurrounding a single secondary osteon-like structure. D, Ornithocheirus, MCZ 2437, transverse sectionof bone shaft showing similar features. E–H, Pteranodon, UCMP 124862, longitudinal section ofepiphyseal fragment in progressively higher magnification, showing calcified cartilage, sub-chondralbone plate, erosion rooms and endosteal struts. Scale bar is 250 �m in C–E, 100 �m in A, B, and F,50 �m in G, and 25 �m in H.

PTEROSAUR BONE HISTOLOGY 365

A. J. DE RICQLES ET AL.366
osteons are cut transversely (Fig. 2F). Most fibrous material runs longitudinally andthe cortex is thus isotropic under crossed nicols. This external primary cortex hasits primary osteons connected by radially oriented or oblique anastomoses, andshows no lamellar tissue. Canaliculi are well developed around all cell lacunae.
Secondary bone tissues: cancellous bone and epiphyses
Cancellous boneIn all sections in which spongy bone is developed, there is a great prevalence of
endosteal tissues in the cancellous structures. In juveniles or in fast-growing regionsof new bone, some trabeculae include a core of primary periosteal bone covered byendosteal tissues, as already noted. Some regions (e.g. in Quetzalcoatlus, Fig. 3J) arecomposed of regular and numerous bone cancellae that form a spongy bone pattern;this structure does not differ from those found in other tetrapods. There is also awell-developed endosteal margin toward the compact cortex.
Other cancellous bone is much more typical of pterosaurs: in this type of bone,the endosteal trabeculae are not very numerous, but are highly individualized asstrong, well-separated struts (Fig. 4A,B: Montanazhdarcho; Fig. 4C: Pteranodon) that arecircular, elongated, or somewhat irregular in cross-section. The thicker struts include
Figure 5. Cross sections of pterosaur bones. Outer edge is up unless otherwise noted. A, azhdarchid,Late Cretaceous, Montana (MOR PT-C), cross-section of wing bone diaphysis. This is rapidly growingjuvenile bone, as evidenced by its dense, reticular vascular supply (compare to B). Dark brown vascularcanals run both obliquely and longitudinally. The light yellow line in the middle of each pink wovenperiosteal bone tissue trabecula is the ‘bright line’ of Currey. B, Quetzalcoatlus sp, TMM 42422-17,Late Cretaceous, Texas, cross-section of diaphysis of first wing-phalanx. This is well vascularized,rapidly growing fibrolamellar bone, comparable to that of a living bird (Figs 8,9), with a thin cortexthat is being eroded endosteally as rapidly as it is being deposited periosteally (Fig. 7B). In this laminartissue type, the ‘bright lines’ are visible equidistant between the dark vascular canals. C, same taxon,wing bone cross-section showing compacted coarse cancellous bone, heavily reworked by Haversianosteons. A triangle of primary cortical bone can be seen in the upper left quadrant. D, same taxon,showing tissue transition from the endosteal margin (lower right corner) to the deep cortex (centre)and finally to the outer primary (periosteal) bone cortex (ending at the upper left corner). Right sideof photo shows erosion bays, frequently infilled by secondary osteons and endosteal bone. E, Pteranodon(UCMP 124862), oblique section of bone cutting through an endosteal bone strut, which is formedby secondary endosteal bone (tan-coloured) with numerous obliquely sectioned osteocytic lacunae(bone cell spaces). Abrupt changes of bone fibres and cell orientations on each side of the cementinglines (at right) demonstrate the secondary origin of the tissue. F, Montanazhdarcho (MOR 691), showingthe base of a transversely cross-sectioned strut. Although the direction of primary growth has beenfrom bottom to top, erosion has removed early-deposited bone (lower corners), leaving only a base ofprimary bone that is then invaded by secondary osteons, eroded further endosteally, and finally coveredwith a coating of endosteal bone (see Fig. 7C,D). The strut grows further by new deposition of endostealbone. G, Pteranodon, an oblique section through part of the scapula, showing alternating ‘plywood’layers in different colours revealed by polarized light using a quartz wedge. Note the streaklike profileof the tiny cell lacunae (black) changing their orientation, alternating in conformity with the bonefibres. H, same taxon, longitudinal section through an epiphysis, with outer edge to the upper right.Blebs of calcified cartilage appear on the surface of the bone; the sub-chondral bone plate beneath itis supported by struts of endosteal bone surrounding large erosion rooms. Trabecular bone can beseen in the deep cortex in the lower left. Scale for photos: B, D, and H, 250 �m; for A, C, E, F, andG, 100 �m.
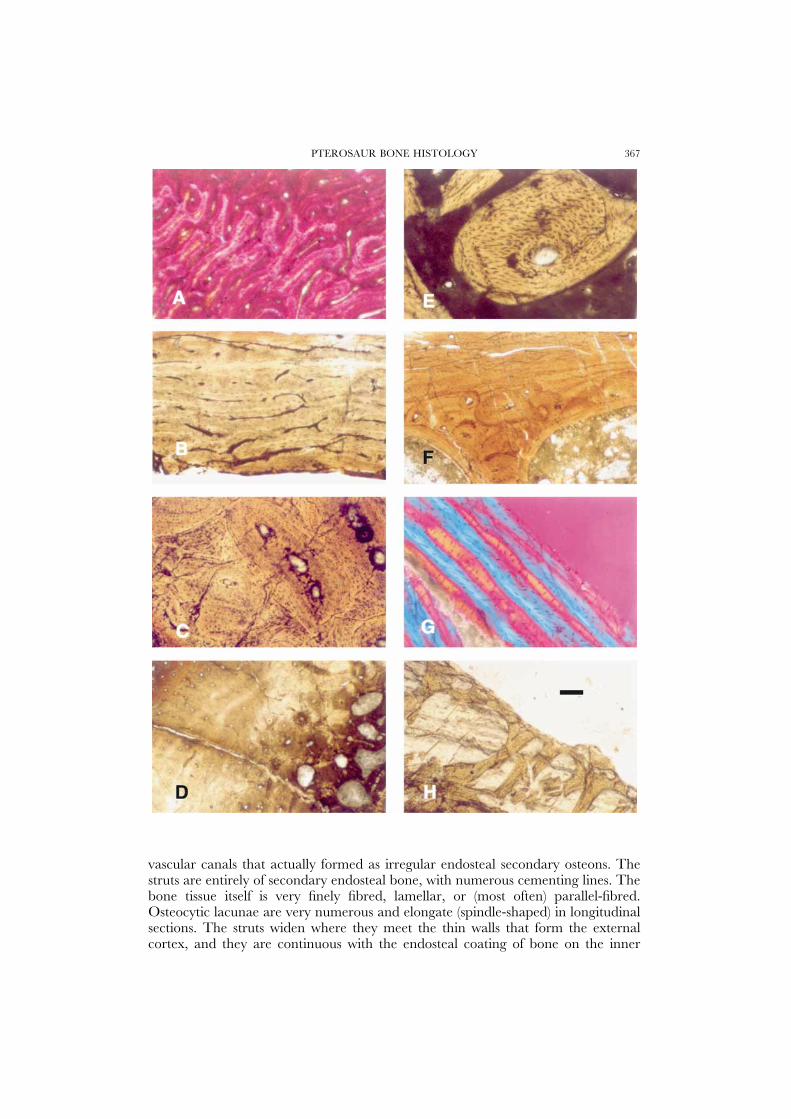
PTEROSAUR BONE HISTOLOGY 367
vascular canals that actually formed as irregular endosteal secondary osteons. Thestruts are entirely of secondary endosteal bone, with numerous cementing lines. Thebone tissue itself is very finely fibred, lamellar, or (most often) parallel-fibred.Osteocytic lacunae are very numerous and elongate (spindle-shaped) in longitudinalsections. The struts widen where they meet the thin walls that form the externalcortex, and they are continuous with the endosteal coating of bone on the inner

A. J. DE RICQLES ET AL.368
Figure 6. Schematic partial cross section of a mature pterosaur long bone. Note the coating of lamellarperiosteal bone and endosteal bone lining the marrow cavity. The cortex is primarily composed offibrolamellar bone; the endosteal tissue has been invaded by some secondary (Haversian) osteons,particularly at the bases of struts and pneumatic canals.
face of the cortex, when present. Apart from these struts, additional endostealmaterial may form a well defined spongiosa in the internal cavity of long bones. InOrnithocheirus, endosteal trabeculae may form bony arches, observed in cross-sections(Fig. 4D). These arches are somewhat irregular: they can unite two more or lessadjacent regions of the cortex and thus outline elongated spaces or cavities lined bybone and set in the ‘marrow’ cavity of long bones. Some of these arches mayrepresent the pathways of former intra-osseous air canals or air sacs.
EpiphysesIn epiphyseal regions (e.g. of Pteranodon: Fig. 4E–G) the endosteal struts become
more numerous and many seem to be somewhat oriented in a fan-shaped patternbelow the epiphyseal plates that form the articular surfaces. They retain a strongindividuality as struts.
The epiphyseal surfaces have a remarkable structure. Histological details show athin, well-finished adult-like structure very similar to the ‘finished’ anatomical aspectof epiphyses among adults, which have tight, well-formed articular surfaces at thejoints (Padian & Rayner, 1993; Bennett, 1993). The epiphyseal surfaces are formedby a thin (three to six cells thick) coating of hypertrophied calcified cartilage. Below
Figure 7. Schematic transverse sections of a pterosaur long bone in various stages of growth (A to D).Circled plus signs indicate areas of bone growth; circled minus signs indicate areas of bone erosion.The bracketed ‘x’ tracks a single layer in the ontogeny of the bone. A, juvenile stage, showing primarywoven bone and ‘bright lines’; compare to Figs 1A and 5A. Oblique vascular canals are wide openon the external (subperiosteal) surface as large clefts. Rounded expansions at the ends of transverselyoriented canals represents their junctions with longitudinal canals. B, immature stage, in which laminarbone growth has settled in; compare to Fig. 1F. Deposition of primary osteons diminishes the diametersof vascular canals. In C, a remnant of primary bone at the internal margin is coated with endostealbone that will begin to build a new internal bone strut. In D, the primary vascular canals have reachedtheir ultimate diameters by centripetal deposition of the primary osteons. Secondary (Haversian)osteons have invaded the internal (older) cortex; endosteal bone has further eroded and rebuilt thebase of the strut and also coats the internal surface of the cortex; and a thin, nearly avascular periostealcoating has been applied to the outer surface of the bone.

PTEROSAUR BONE HISTOLOGY 369

A. J. DE RICQLES ET AL.370
the cartilage is a continuous wall of endosteal bone, thicker than the cartilage. Thisbone forms a sub-chondral plate as described in adult mammals (e.g. Lempberg,1971). The bone tissue of the plate is structurally continuous with the endostealstruts of the spongiosa. Between the struts and below the plate are numerous large,well-delineated cancellae or cavities that form a spongiosa with the struts.
Almost no remnants of calcified cartilage can be observed in the struts or in theplate, because both are secondarily endosteal. This epiphyseal structure appears toreflect an adult condition with very little, if any, opportunity for further growth inlength provided by the epiphysis (Ricqles, 1972). But it does not theoretically preventfurther growth. The uncalcified articular cartilage could resume growth in its deepregion, while erosion bays could destroy the sub-chondral bone plate and againinvade the cartilage outwardly. However, no evidence of such processes has beenfound because all the cancellae of the external spongiosa seem to be separated fromthe calcified cartilage by the sub-chondral bone plate.
No evidence of transphyseal canals, or specialized canals linked with air sacs,could be unequivocally observed. Although the presence of transphyseal canalsseems unlikely, we cannot exclude the possibility that some endosteal trabeculae didindeed form the walls of once-present bony canals to air sacs. Unfortunately, theepiphyseal regions are almost always badly crushed in Pteranodon.
In Montanazhdarcho (Fig. 4H), an uncrushed epiphyseal surface and other structuresand surfaces similar to those described above in Pteranodon suggest an adult condition(Padian et al., 1995). In this well-preserved material, there is a tentative indicationthat some elongated, regularly developed, longitudinally oriented endosteal tra-beculae could have formed the walls of canals to air sacs. Moreover, on the lateralsurface of the epiphysis, it is still possible to observe a peculiar thin coat of tissue,the so-called metaplastic bone of Haines & Mohuiddin (1967), linked to a strongtendinous or ligamentous insertion. The ossified material would have been formedby the most proximal part of the dense connective tissue itself, where it is anchoredto the lateral surface of the epiphysis (Haines, 1969: 92; Ricqles, 1975: 114).
Summary: general histologic characters of pterosaur bone
As might be expected for a large and unusual group of vertebrates spanning awide range of body sizes, bone tissue structures are highly varied in pterosaurs.Among the primary tissues, very simple lamellar-zonal tissues are seen in the externalcortex of larger bones (Fig. 6), and they may form most of the cortex in the smallerbones of adults. But fibro-lamellar tissues form the bulk of the innermost periostealcortex in most larger bones and in smaller ones (Fig. 7). These tissues vary extensively,from a pattern with few longitudinal and circular vascular canals embedded in awoven matrix to a dense, regularly organized reticular pattern of vascularizationwithin well-marked laminae. The vascular canals are occasionally of the simpleprimary type (i.e. there is no centripetal deposition of osteonal bone around them),but more often, they form the canals of primary osteons. Sharpey’s fibres arecommon in periosteal bone, especially in association with simple lamellar-zonaltissues. These fibres seem to be exceptionally long and fine. The material that formsthe primary osteons is often characterized by longitudinally oriented fibres, andhence a ‘lamellar’ structure of the osteons is not obvious (Fig. 7A,B). This alsoappears to hold for secondary osteons (=Haversian systems), which can be common

PTEROSAUR BONE HISTOLOGY 371
Figure 8. Schematic cross sections of a pterosaur long bone. A is the distal end of the bone inlongitudinal section, showing the internal struts of endosteal bone. B is an enlargement of the rectanglein the epiphysis in A; the sub-chondral bone plate underlies the calcified cartilage of the epiphysis. Cis a bird’s-eye view (transverse section) of the metaphysis indicated by the line in A.
and numerous (Fig. 7D). Other types of secondary (i.e. substitution) tissues form (a)compacted coarse cancellous bone (often in association with secondary osteons), (b)secondary endosteal cancellous tissue of the spongiosa in epiphyseal regions (Fig.8A), and (c) more or less well-individualized endosteal struts in the metaphyseal andsometimes diaphyseal regions (Fig. 7D). In addition, secondary endosteal bone formsa well-developed and significant component of the cortex, as a coat of bonytissue around the ‘marrow’ cavity (Fig. 6). This endosteal bone is almost alwaysunconformably deposited over the deep periosteal cortex. Finally, highly remodeledendosteal tissue forms the sub-chondral bone plates that develop under the articularfacets of the epiphyses (Fig. 8A). Thin calcified cartilages are developed at well-ossified articular surfaces, in association with the sub-chondral bone plates. It isobvious that a thin coating of non-mineralized articular cartilage overlay in vivo thecalcified cartilage that has been fossilized (Ricqles, 1972). Specialized tissues for theinsertion of tendons and ligaments (metaplastic bone of Haines & Mohuiddin, 1967)are well-developed on the lateral surfaces of epiphyses.
Lines of discontinuity are routinely observed in all bones. While some of themare distinct resorption lines (e.g. in the deep cortex between endosteal and periostealtissues), others may be true LAGs in the periosteal cortex. The number of theselines in the cortex of pterosaurs always appears to be very low, and restricted to theoutermost cortex. These are not associated with the annulus that is typical of poorlyvascularized tissues when found in the matrix of fibro-lamellar bone.
It thus seems, so far, that pterosaurs share the main categories of bony tissuesgenerally observed in other tetrapods. But some tissue types, as well as the relativedevelopment or associations of tissues, seem to be peculiar to pterosaurs. What has

A. J. DE RICQLES ET AL.372
Figure 9. Schematic cross section of the ‘plywood’ type of pterosaur bone, showing alternating layersof obliquely aligned bone matrices containing vascular canals (direction of arrows).
been described above as ‘plywood-like’ bone (Fig. 9) appears to be unique to the group.As far as we know, such structures have not been reported in the long bones of birdsand other dinosaurs, crocodiles, or lepidosaurs. A closer analogue seems to be theisopedine family of bony tissues that have exceptionally well developed plywood-likestructures. These have been described in detail from the basal region of scales inactinopterygian fishes, notably teleosteans (e.g. Meunier & Castanet, 1982; Meunier,1984). However, in pterosaurs the plywood can be routinely well-vascularized and itcan show intermediate conditions that allow a smooth structural transition to the morefamiliar lamellar-zonal tissue types. It is tempting to hypothesize that the evolution ofthis tissue in pterosaurs is linked to the peculiar biomechanical properties of plywood-like materials, in the special adaptive context of pterosaur growth and flight (see below).
The great development of reticular vascularization in the fibro-lamellar tissuesappears to be another specialized feature of pterosaur bone, a situation paralleledamong large soaring birds. The fact that the vascular canals in this fibro-lamellarcortex are obliquely organized, as Gross (1934) first emphasized, may also bebiomechanically important. The great development of endosteal bony struts (Figs 6–8;Wellnhofer, 1985) parallels the avian condition, probably for similar biomechanicalreasons (e.g. Currey, 1968, 1984). As mentioned above, since the observations ofthe earliest workers (Quekett, 1849b), the bone cells of pterosaurs have appeared tobe unique by virtue of their small size, number, elongate shape, and canaliculardevelopment. The differentiation of sub-chondral bone plates, which is most similarto those of adult mammals, raises the issue of growth patterns among pterosaurs.Comparisons with birds appear natural for issues such as growth rates, metabolism,and biomechanical adaptation.
DISCUSSION
Functional significance of tissue types and their microanatomical organization
There are no tissue types peculiar to pterosaurs as a group, except possibly theplywood-like bone described above, which has not as yet been reported in any other

PTEROSAUR BONE HISTOLOGY 373
tetrapod. However, the forms of even common bone tissues are often highly modifiedfrom typical patterns in other taxa, and this suggests that they had a precisefunctional significance. Moreover, the ways in which these tissues are integratedinto the bones as individual organs and as a skeletal system is peculiar to pterosaurs.These lines of evidence can help to elucidate questions of growth dynamics andbiomechanical adaptation.
Growth dynamicsThe great prevalence of fibro-lamellar bone tissues, especially of the reticular
type, strongly suggests that growth was rapid in pterosaurs. This is at least clear forlarger pterodactyloids, for which we have been able to sample some ostensiblyjuvenile material. We did not have juvenile material for more basal pterosaurs, soour inference that their growth was similarly rapid is based on comparisons of tissuestructures at the sub-adult level, which do not substantially differ. Fibro-lamellartissues are well-developed among large flying birds (Amprino & Godina, 1947;Enlow & Brown, 1957; Ricqles, 1978), which are known to grow very quickly. Thesetissues do not persist through ontogeny in living animals that are known to growslowly, although they can appear ephemerally in young animals, and can also beenhanced under optimal captive conditions. In most principal skeletal bones (i.e.the larger long bones) of pterosaurs, the bulk of the cortex would be thus built up,during early life, by very quickly deposited fibro-lamellar tissues (e.g. in Mon-tanazhdarcho). Smaller bones (e.g. scapula, some long bones such as the short wingmetacarpal of non-pterodactyloids) may have accommodated an overall lower rateof growth, as suggested by the variations of fibro-lamellar tissues observed in thedeep cortex of these bones in (e.g.) Dimorphodon and Pteranodon. As Bennett (1995:577) notes, growth in these bones could take place by remodeling the entire bone,even after fusion of girdle and vertebral elements. Even smaller bones (e.g. metatarsalsand phalanges) could rely on various modulations of still more slowly growing bonetissues such as lamellar-zonal tissues and perhaps plywood-like bone.
When (what we interpret as) an almost adult condition is reached, there arehistologic indications of a dramatic change in growth rate. These are manifested bythe structural changes in the external cortex, including one or more lines of arrestedgrowth and further deposition of a thin coat of lamellar, poorly vascularized bone.This condition corresponds to the so-called ‘external fundamental system’ classicallydescribed at the periphery of the cortex among adult mammals (Figs 6, 7D; Cormack,1987). It would suggest that growth was determinate in pterosaurs (Bennett, 1993,1995, 1996a), so maximum size was reached at the onset of adult life, as amongextant endotherms (mammals and birds). Pterosaurs have an extremely low cortico-medullary index; i.e. the bony cortex is extremely thin compared to the overall shaftdiameter. This pattern persists throughout ontogeny because the deeper cortex iscontinually and almost completely resorbed on the perimedullary side. Thus only asmall portion (generally the most recent) of the individual ontogeny is recorded inthe structure of its shafts (Figs 6, 7). In this respect, pterosaurs differ markedly fromliving reptiles such as lepidosaurs, chelonians, and crocodiles, in which this recordcan be fairly complete (e.g. Castanet et al., 1993), and are more similar to birds.Because the cortex of adult pterosaur bones may only record the latest stages ofontogeny, the presence of slowly growing tissues (e.g. lamellar-zonal and plywood-like) in adult bones may be misleading in the sense that it could suggest an overall

A. J. DE RICQLES ET AL.374
slow growth for pterosaurs; however, only the latest phases of growth, which indeedwere slower than in early life, are in fact recorded.
Along the same lines, Haversian bone can be observed in the cortex of pterosaurlong bones (Fig. 7D), even in small bones whose structure otherwise suggests thatgrowth in diameter was almost negligible and hence that the adult condition hadbeen reached. But Haversian bone by itself does not necessarily mean that the adultstage has been reached, except in the case of dense Haversian bone. Haversiantissue may form for several reasons; it can be found in the fibro-lamellar cortex ofsome bones that show no other adult features. For example, we have identified itat the bases of some endosteal struts (Figs 4A–D, 5E), where it may function in oneor more of several ways, including inhibiting the spread of microfractures, andincreasing fatigue resistance (Carter et al., 1976); however, the exact functions ofHaversian replacement remain debatable.
Independent histologic evidence pointing to the same inference is provided bythe structure of epiphyseal surfaces (see also Bennett, 1993). The differentiation ofsub-chondral bone plates, as among adult mammals, provides a mature epiphysealsurface that can be finely tuned to its biomechanical functions in articulation, butthat would probably prevent further growth during adulthood.
In summary, the histologic structures of cortical bone in the shafts and of theepiphyseal surfaces both suggest rapid growth during early life and a cessation ofgrowth in adults (Bennett, 1993). Such a finite growth curve is best seen in mammalsand birds today, although it appears that some living ectotherms (e.g. anurans:Francillon, 1981) have also independently devised epiphyseal mechanisms adaptedto finite growth. In concluding that all pterosaurs grew rapidly, we do not implythat they all grew at identical rates. Large and small birds do not grow at identicalrates; they differ both in the kinds of tissues they lay down and in the time takento reach adult size, which is partly a function of size itself. We mean to imply thatpterosaurs grew faster than typical living reptiles, and resembled birds much moreclosely in this respect.
Possible biomechanical implicationsAdaptations of flight, and the means by which pterosaurs achieved it, impose
general and particular constraints on bone structures (Padian, 1985; Rayner, 1989a,1989b; Padian & Rayner, 1993). Among general constraints, weight reduction is anobvious one for the bones of most flying animals, but the extreme elongation of theforelimb bones, particularly the wing metacarpal and phalanges, is unique topterosaurs. The thin-walled, tubular long bones have to cope with all the strains ofaerodynamic and terrestrial locomotory and other forces, so they must have anacceptable safety factor (Bramwell & Whitfield, 1974). Hence it is likely thatarchitectural details that could enhance the biomechanical properties of these boneswould be incorporated as far as genetic and developmental factors allowed.
One aspect of the apparent fine-tuning of pterosaurian cortical bone to copewith mechanical strain using minimal weight may be found in the plywood-likeorganization of the bone (Figs 3A–D, 9). In the bones of larger adults, the thincortex comprises at least three plies: the endosteal bone of the deep cortex, withfibres and cells mostly circularly oriented; the periosteal cortex, with most fibres andcells longitudinally and obliquely oriented; and the most external, lamellar periostealcortex, with fibres sometimes oriented circularly and cells oriented longitudinally.

PTEROSAUR BONE HISTOLOGY 375
In smaller bones, or flat bones with a relatively poor vascularization of the cortex,the bone tissue may be entirely formed by relatively thick plies oriented orthogonallyto adjacent plies (“orthogonal plywood” of Meunier & Castanet, 1982). As many aseight plies may form the periosteal component of the cortex.
Furthermore, the osteonal material itself is unusually organized. Rather thanbeing twisted at a low angle, and hence with a ‘lamellar’ aspect in cross section(Girauld-Guille, 1988), the collagenous fibres of the osteons seem longitudinal, orat most twisted at a very high angle.
In the spongiosa, there is a clear differentiation of a system of struts and endostealbony ‘pipes’ in the diaphyseal and metaphyseal regions. Their presence and orientationmay be explained in two ways: first, as the results of local developmental stresses thatcaused the apposition of bone at certain times during ontogeny (Carter et al., 1991),and second, as a functionally advantageous structural system (Barlee, 1953; Rogers &LaBarbera, 1993) spatially organized to transmit and distribute local forces of strain(Fig. 8). Their morphology and distribution in pterosaur long bones seem to convergeclosely upon the struts found in the long bones of many (especially larger) birds(Wellnhofer, 1985; Currey, 1984: 155–156). Internal struts seem to be more developedin metaphyseal cross-sections with an irregular shape than in circular diaphyseal cross-sections. In general, many of the characteristics of pterosaur bone histology seem tobe manifestly similar to those of large birds (Seitz, 1907; Enlow & Brown, 1957; Gross,1934; Bennett, 1993), probably because of similar biomechanical requirements of flight(Padian, 1985; Padian & Rayner, 1993).
As noted earlier, Haversian substitution begins in juvenile pterosaurs and continuesthroughout growth. However, we have not observed the dense Haversian tissuefound only in adults of other taxa. Haversian osteons may be found throughout thecortex but are frequently concentrated at the bases of endosteal struts. Their presencehere may have partly developmental and partly biomechanical causes. Secondaryosteons run longitudinally, and their accompanying longitudinally oriented collagenfibres increase resistance of the bone to tensile forces (in contrast, oblique andtransverse fibres confer resistance to compression: Riggs et al., 1993). This propertymay have been important in the growth and development of these wing bones.
Finally, in well preserved long bones there are frequently remnants of bothseparating walls and septae, as noted above, and endosteal struts. These remnantscan often be seen connected to the cortical wall of the marrow cavity, and oftenrun oblique to the direction of the bone shaft (Fig. 10; Currie & Russell, 1982).They appear to be vestiges of formerly complete structures that have been partlydestroyed. We offer the hypothesis that they may have served a biomechanicalfunction of helping the thin-walled bone shaft to resist torsion and other stress,much like earthquake bracing does to walls. The addition of such centripetallyoriented structures on the internal sides of the walls of hollow support structureshas been shown to lend them resistance to local buckling (e.g. Karam & Gibson,1994). Buckling is probably the most important biomechanical ‘hazard’ that pterosaurbones had to face, in view of their inordinately thin walls.
Integration of histologic and other evidence for growth rates and ontogeny
Wingspans for pterosaurs range from 15 to 20 cm in juveniles of Pterodactylus(Soemmerring, 1817; Wellnhofer, 1970) up to 11–12 m in Quetzalcoatlus (Langston,

A. J. DE RICQLES ET AL.376
Figure 10. Cf. Quetzalcoatlus sp., femur in anterior view (above) and an enlargement of the internalsurface of its proximal end, showing the development of struts and buttresses. These internal featuresappear to result from the erosion of primary bone into remnants that are then covered by new layersof endosteal bone. The hypothesis that these features promote resistance to torsion or other stressesrequires mechanical testing. From Currie & Russell (1982).
1981). The smallest individual that we surveyed in our study, represented by a wingphalanx of Dimorphodon (YPM 9178), would have had a wingspan of approximately1.5 m (Padian, 1983); the largest, represented by specimens of the ‘small morph’ ofQuetzalcoatlus and other azhdarchids, had wing spans of 6–9 m (Langston, 1981;Padian & Smith, 1992). (A sample of bone from the 11–12 m wingspan Q. northropiproved difficult to assess because much of its internal structure had been destroyedby decomposing organisms.) Hence there is a sixfold range of size in the taxa wesurveyed. However, it is not possible on the basis of size alone to determine whetheran individual is a juvenile. Bennett (1993) provided several criteria related to bonesurface finish and fusion, and these have already been discussed. The internalstructure of the bone, particularly as revealed by histology, provides additionalinformation (Padian et al., 1995).
Our study determined that fibro-lamellar tissue is the most prevalent in pterosaursof all size classes, and this finding is supported by the work of other authors (e.g.Gross [1934] for Dorygnathus; Enlow & Brown [1957] and Bennett [1993] forPteranodon). Fibro-lamellar tissue is most generally associated in extant tetrapods withhigh growth rates, and it has also been found to predominate in the long bones ofdinosaurs (Ricqles, 1974, 1976, 1980). It has sometimes been postulated that thelarge size of dinosaurs was possible only because of high growth rates, and high

PTEROSAUR BONE HISTOLOGY 377
growth rates apparently also characterized pterosaurs. The basal members of bothgroups were small (Weishampel, Dodson & Osmolska, 1990; Wellnhofer, 1991), aswere those of their immediate outgroups, such as Scleromochlus, Lagosuchus, andLagerpeton (Gauthier, 1984; Sereno, 1991; but see Bennett, 1996b). It is probablethat growth rates were high in the common ornithodiran ancestor. High growthrates permitted large size, but did not evolve initially in the context of evolutionarysize increase; other metabolic and functional considerations were almost certainlyat work (Ricqles, 1980).
Of equal interest in this context is the presence of outer lamellar bone in thesubperiosteal cortex of some specimens (e.g. Montanazhdarcho; Fig. 6). This bone ispoorly vascularized, and when found in living animals is a general indicator of slowbone growth. Because it occurs only on the outer perimeter of some pterosaurbones, and is separated from the underlying fibro-lamellar bone by an unconformityof deposition, it clearly represents a dramatic deceleration of growth rate, such asone finds in fully mature individuals of birds and mammals (Currey, 1984). Syn-thesizing these observations, we conclude that pterosaur bone grew rapidly throughontogeny up to a determinate adult stage, at which point growth effectively ceased(Padian et al., 1995). At this time, it is important to note, erosion of the boneendosteally also effectively seems to have ceased, and the outside of the marrowcavity receives a coat of endosteal bone; even the deep primary cortex can beinvaded by secondary osteons (Haversian systems) (Fig. 6).
D.R. Carter and his associates (e.g. Carter, 1987; Carter et al., 1991; Carter,Beaupre & Padian, 1992; Carter, van der Meulen & Padian, 1992; van der Meulen,Beaupre & Carter, 1993; van der Meulen & Carter, 1995) have modeled the formof bone as the result of interactions between apposition and erosion, in which thewidth of the ‘lazy zone’ represents a range of strains that induce apposition anderosion of the bone. This range is much narrower for pterosaurs than for mammalsof similar size. Furthermore, the width of the ‘lazy zone’ is directly proportional tobone wall thickness, and the span and distance of this zone from the ordinal axisrepresent the radius of the bone itself. This model suggests that pterosaur bone grewrapidly, but adds the further dimension that it may have done so in part as aresponse to stress, which could readily be interpreted from the demands of activeflight. The fact that the hindlimb bones show the same profile of thin bone wallssuggests that they were also capable of absorbing similar stresses, and van derMeulen, Padian & Carter (1992; see also Carter, van der Meulen & Padian, 1992;Padian, van der Meulen & Carter, 1992) calculated that pterosaur leg bones arenearly as strong as mammalian bones of the same length, despite their much thinnerwalls.
Although growth rates of long bones comprising fibrolamellar tissues can varyamong bones in a single skeleton, these rates are uniformly higher than for typical‘reptilian’ bone. Castanet et al. (1996) found that a mallard’s tibiotarsus, femur, andhumerus grew at 4 �m, 7.5 �m, and 25 to 40 �m per day, respectively, at 7 weeksold, decreasing proportionally as the animal reached maturity (about 20 weeks). Itis normal for similar parts of different long bones in the same individual not todisplay the same histological structures at the same growth stage. Carrier & Leon(1990) found that very rapid growth was characteristic of altricial birds such as theCalifornia gull, whose hindlimbs are functional long before its wings; however, theydid not study the comparative histology of the limb bones. We did not have availableto us sufficient hindlimb material from an ontogenetic series of pterosaurs to be able

A. J. DE RICQLES ET AL.378
Figure 11. Comparison between the skeletons of a typical ‘juvenile’ (possibly hatchling) Pterodactylus, atypical juvenile Pterodactylus kochi, and the rostrum of an adult P. longicollum (the last after Bennett,1996a). Scale bar=1 cm.
to determine at this time whether bone wall thickness is initially greater in thehindlimbs, as it is in the gull (Carrier & Leon, 1990). It is not so in adults or near-adults of Dimorphodon macronyx: the second wing-phalanx is 97 mm long and the tibia104 mm in YPM 350; the mid-shaft thickness of the former bone ranges from 0.5to 0.8 mm, and from 0.4 to 0.7 mm for the latter. In YPM 9182, the second wing-phalanx is 74 mm long and the tibia 85 mm; the mid-shaft thickness of the formerbone ranges from 0.3 to 0.4 mm, and from 0.2 to 0.3 mm for the latter.
Implications for pterosaurian palaeobiology
Growth ratesComparison of the proportions of typical and small (presumably juvenile) Ptero-
dactylus (Fig. 11) indicates that growth was allometric, with the long bones and snoutmore positively allometric than other parts of the skeleton (allometric rank variesfor individual long bones among taxa: Padian & Warheit, 1988). Bennett (1993)discussed the ontogeny of Pteranodon, and found that size alone was not a reliable

PTEROSAUR BONE HISTOLOGY 379
indicator of maturity, because even though only 14.5% of the animals he studiedsatisfied one or more of his three criteria of sub-adulthood, even the smallestindividuals were 70% as large as the largest individuals. Bennett applied his criteriato re-evaluate the conclusions of Wellnhofer (1970) and Mateer (1976) on Pterodactylus,and of Wellnhofer (1975) on Rhamphorhynchus, and he concluded that size-dependenttaxonomic criteria used to separate these genera into species frequently conflictedwith criteria based on maturity of bone. He inferred that growth rates in smallpterosaurs were typically crocodilian, but must have been rapid in large ptero-dactyloids. Our study of the bone histology, summarized above, supports Bennett’scontention that growth was rapid in large pterosaurs, because the histologicalstructures that predominate in pterosaur long bones are commensurate with thoseobserved in the rapidly growing bone of extant birds and mammals.
As to the growth rates of early (basal) and small pterosaurs, the few histologicaldata so far available do not allow us to test fully Bennett’s (1995, 1996a) conclusionsthat the traditionally identified species of Late Jurassic Rhamphorhynchus and Pterodactylusrepresent multiple year-classes, indicating slow growth and several years beforeadulthood is reached. The fibrolamellar bone observed in Dimorphodon is somewhatless specialized than in large, derived pterodactyloids, and may indicate a slowergrowth rate, although no histological details (such as lines of arrested growth) suggesta growth protracted over several years. Comparison with histology of the mallard(Castanet et al., 1996) suggests that the cortical bone seen in the specimens ofDimorphodon that we studied may have grown at about 2.5 to 6 � per day. Thehistology of several bone elements (see Table 1) from three different individuals isvery consistent. As a simple calculation, a bone 5 mm in diameter (2.5 mm radius)would take 500 days to grow at a speed of 5 � per day. However, it is likely, basedon growth rates of all known archosaurs, that growth was more rapid at early stages(some mallard bones grow at 40 � per day during early ontogeny). Even with theseslower rates it is unlikely that Dimorphodon took more than a year to reach whatappears to be nearly adult size. Based simply on the evidence from bone histology,if Rhamphorhynchus and Pterodactylus grew at rates commensurate with either Dimorphodonor derived Cretaceous pterodactyloids, it is difficult to see how adult size would takemuch more than a year to attain.
Life-history strategiesPadian & Rayner (1993), in a study of the wings of pterosaurs, inferred from
several lines of evidence that pterosaurs must have been born in a highly altricialstate. The long wing bones of pterosaurs must have been folded in the egg, andcould not have achieved flying proportions without rapid growth after hatching.During this time the wing membrane and its supporting stiffened fibres also had todevelop, along with a musculo-skeletal system capable of withstanding the demandsof flight. This hypothesis appears to be consistent with the fast-growing tissues thatpredominate in the bones of juvenile pterodactyloids described here.
Carrier & Leon (1990) studied the allometry of juveniles of the California gull.They found that rates of long bone growth were very high after hatching, particularlyin the wing bones, which are not used for flight until the animals are nearly fullgrown. The leg bones, by contrast, grow more slowly, but are functional at a muchearlier stage in life. Carrier and Auriemma (1992) found that in birds, fledging timeis strongly correlated with wing bone length; this may be important for pterosaurs,

A. J. DE RICQLES ET AL.380
Figure 12. Thin sections from Gross (1934: 759) of a long bone of the Early Jurassic basal pterosaurDorygnathus in longitudinal (left) and transverse (right) views. Although some tissue has been destroyednear the outer cortex (right), both sections show a matrix of well vascularized fibro-lamellar bone (seetext). The secondary endosteal layer of bone on the inner cortex and the thin avascular layer of outerperiosteal bone suggest that the individual has reached or will soon reach full size. Scale=1 cm.
which had more long bones in the wing than birds do, and a longer, thinner leadingedge spar.
Based on the growth rates reflected in pterosaurian long bones, we hypothesizethat many if not all pterosaurian taxa had similar ontogenetic histories, partlyaffected (as in birds and mammals) by adult size and ecological factors. It may beobjected that we had only a few mature or nearly mature specimens of Dimorphodonto represent all non-pterodactyloid, pre-Cretaceous taxa, and so could not assessthe growth rates of basal pterosaurs with precision. This objection is strictly true.On the other hand, many of the tissues that we see in the adult Dimorphodon specimenscorrespond to those stages of the growth trajectories of other pterosaurs and dinosaursthat grew rapidly and whose cortex through ontogeny was primarily composed offibro-lamellar bone. Gross (1934), in describing thin-sections of the basal pterosaurDorygnathus, provided observations that corroborate this conclusion (Fig. 12). Hedescribed reticular, fast-growing primary fibro-lamellar bone with no Haversianreworking. Vascular canals were numerous, mostly longitudinal, and anastomosing;‘bright lines’ were commonly visible. Endosteal bone was secondarily overlain onthe eroded inner cortex, and growth had clearly slowed at the outer cortex (a featureof sub-adults or near-adults). Gross compared these tissues in several respects tothose of birds. As we noted above, we do not imply that all pterosaurs grew at thesame rates, only that they grew at rates that were generally very high, and muchmore like those of birds than of typical reptiles.
To test this hypothesis fully would require (a) complete histologic assessment ofthe bones from a range of ontogenetic stages, or (b) embryonic and young nestlingskeletons, neither of which is abundantly available for study. However, in our viewthe available lines of evidence point to generally high growth rates, altricial conditionat birth, and, by implication, parental care in feeding the young (Horner & Makela,1979) until they were old enough to forage for terrestrial prey by running downfood items and eventually to take wing and feed as adults did. In the Jurassic depositsof Solnhofen and Holzmaden, juveniles and sub-adults of adult size constitute mostof the preserved specimens (Bennett [1995, 1996a] on Rhamphorhynchus, Pterodactylusand its allies; Padian & Wild unpub. data on Dorygnathus and Campylognathoides).Hence, a mortality sample would be expected to preserve younger juveniles ingreater proportion than sub-adults and mature animals, a feature reflected in theSolnhofen and Holzmaden beds as in mortality samples of many living birds today.Many of the other tetrapod taxa preserved in the Solnhofen limestones, suchas crocodiles, sphenodontids, lizards, and birds, include predominantly juvenile

PTEROSAUR BONE HISTOLOGY 381
individuals (Bennett, 1996a; Houck et al., 1990; Wellnhofer, 1971) with unfusedsutures and carpals and tarsals of simple morphology. We infer that pterosaurgrowth was rapid at first and then tailed off, following a convex, asymptotic curve,and that the relatively higher sub-adult morphology has yielded this curve forpreserved animals.
CONCLUSIONS
As reported in many previous studies (e.g. Quekett, 1849a,b, 1855; Seitz, 1907;Gross, 1934; Enlow & Brown, 1957, 1958; Ricqles, 1976, 1979, etc.), in their generalfeatures pterosaur bone tissues resemble more closely those of birds than those ofcrocodiles or lizards. To date, most analyses of pterosaur bone have been opportunisticin the sense of sampling only a single section of a single bone, often withoutcontextual information of its place in the skeleton, or of the size or ontogenetic stageof the individual. Our study, though not based on an ideal sample, shows additionalfeatures and variability in structures that have not been previously reported, includingarticular calcified cartilage, sub-chondral bone plates, trabecular bone struts andrelated internal supports, and secondary deposition and remodeling of bone. Ingeneral, our analyses indicate that the thinness of pterosaur cortical bone can betraced to quite high growth rates from early ontogeny. During this time, diametralexpansion of the long bone shafts was rapid, and periosteal bone was laid downexternally as quickly as osteoclasts eroded the cortex from inside the expandingmarrow cavity. High growth rates seem to have prevailed until adult size wasreached, at which point growth often slowed in the periosteal deposition of denselamellar bone externally. Occasionally secondary osteons were formed by erosionand redeposition within the pre-existing internal bone. Erosion of most internal(perimedullary) bone surfaces was followed by extensive secondary deposition ofendosteal parallel-fibred bone, which forms the innermost cortex in adults. Aninternal network of trabecular struts of endosteal origin also developed, and thismay have had a function in biomechanical support (Rogers & LaBarbera, 1993).The rapid growth rates of pterosaur bones, together with considerations of theirunusual morphology and flight adaptations, suggest that pterosaurs hatched in analtricial state, and were cared for by the parents until they were able to fly.
ACKNOWLEDGEMENTS
We thank Drs Wann Langston, John Ostrom, and Jacques Gauthier for allowingus to section specimens in their care. Allison Gentry and Ellen Lamm (MOR)prepared the thin-sections. Photographs were taken in the facilities of the MOR,the Universite Paris VII, and Prof. Marvalee Wake at UCB. We thank Drs DennisCarter and Marjolein van der Meulen for helpful discussions. We are particularlygrateful to Dr S.C. Bennett and two reviewers for extremely helpful comments ona previous draft of the manuscript, and to Adele Blanc-Sec for inspiration. Thiswork was supported by grants to JRH (Merck Family Fund; Ruth and VernonTaylor Foundation and The Charlotte and Walter Kohler Charitable Trust), by the

A. J. DE RICQLES ET AL.382
CNRS, and by the UCB Committee on Research and the UCMP. This is UCMPcontribution No. 1679.
REFERENCES
Amprino R, Godina G. 1947. La struttura della ossa nei vertebrate. Ricerche comparative negliamfibi e negli amnioti. Commentationes Pontificae Academiae Scientiarum 11: 329–467.
Barlee J. 1953. Le vol des oiseaux. Aviation Magazine 74: 16–19.Bell CM, Padian K. 1995. Pterosaur fossils from the Cretaceous of Chile: evidence for a pterosaur
colony on an inland desert plain. Geological Magazine 132: 31–38.Bennett SC. 1993. The ontogeny of Pteranodon and other pterosaurs. Paleobiology 19: 92–106.Bennett SC. 1995. A statistical study of Rhamphorhynchus from the Solnhofen Limestone of Germany:
year-classes of a single large species. Journal of Paleontology 69: 569–580.Bennett SC. 1996a. Year-classes of pterosaurs from the Solnhofen Limestone of Germany: taxonomic
and systematic implications. Journal of Vertebrate Paleontology 16: 432–444.Bennett SC. 1996b. The phylogenetic position of the Pterosauria within the Archosauromorpha.
Zoological Journal of the Linnean Society 118: 261–308.Bramwell CD, Whitfield GR. 1974. Biomechanics of Pteranodon. Philosophical Transactions of the Royal
Society of London B 267: 503–581.Carrier DR, Auriemma J. 1992. A developmental constraint on the fledging time of birds. Biological
Journal of the Linnean Society 47: 61–77.Carrier DR, Leon L. 1990. Skeletal growth and function in the California gull (Larus californicus).
Journal of Zoology (London) 222: 375–389.Carter DR. 1987. Mechanical loading history and skeletal biology. Journal of Biomechanics 20:
1095–1109.Carter DR, Beaupre GS, Padian K. 1992. Evolutionary constraints imposed by skeletal de-
velopmental mechanics. Transactions of the Orthopedic Research Society, 38th Annual Meeting 17(1): 95.Carter DR, Hayes WC, Schurman DJ. 1976. Fatigue life of compact bone – II. Effects of
microstructure and density. Journal of Biomechanics 9: 211–218.Carter DR, Wong M, Orr TE. 1991. Musculoskeletal ontogeny, phylogeny, and functional adaptation.
Journal of Biomechanics 24 (suppl. 1): 3–16.Carter DR, Van Der Meulen MCH, Padian K. 1992. Historical and functional factors in the
construction of pterosaur bones: a preliminary analysis. Journal of Vertebrate Paleontology 12 (suppl. 3):21A–22A.
Castanet J, Francillon-Viellot F, Meunier FJ, de Ricqles A. 1993. Bone and individual aging.In: Hall BK, ed. Bone, Volume 7: Bone Growth – B. Boca Raton, Florida: CRC Press, 245–283.
Castanet J, Grandin A, Abourachid A, Ricqles A. 1996. Expression de la dynamique de croissancedans la structure de l’os periostique chez Anas platyrhynchos. Comptes Rendus de l’Academie des Sciences,Paris: Sciences de la Vie 319: 301–308.
Chiappe LM, Chinsamy A. 1996. Pterodaustro’s true teeth. Nature 379: 211–212.Chinsamy A, Raath MA. 1992. Preparation of bone for histological study. Palaeontologia africana 29:
39–44.Cormack D. 1987. Ham’s Histology. New York: Lippincott.Currey JD. 1960. Differences in the blood supply of bone of different histological types. Quarterly
Journal of the Microscopical Society 101: 351–370.Currey JD. 1962. The histology of the bone of a prosauropod dinosaur. Palaeontology 5: 238–246.Currey JD. 1968. The adaptation of bones to stress. Journal of Theoretical Biology 20: 91–106.Currey JD. 1984. The Mechanical Adaptations of Bones. Princeton, New Jersey: Princeton University
Press.Currie PJ, Russell DA. 1982. A giant pterosaur (Reptilia: Archosauria) from the Judith River
(Oldman) Formation of Alberta. Canadian Journal of Earth Sciences 19: 894–897.Cuvier G. 1809. Memoire sur le squelette fossile d’un Reptile volant des environs d’Aichstedt. Annales
du Musee d’Histoire Naturelle, Paris 13: 424–437.Enlow DH. 1963. Principles of Bone Remodeling. Springfield, Illinois: Charles C. Thomas.Enlow DH. 1966. An evaluation of the use of bone histology in forensic medicine and anthropology.

PTEROSAUR BONE HISTOLOGY 383
In: Evans G, ed. Third International Congress of Anatomists: Symposium on joints and bones. Berlin: Springer-Verlag, 92–112.
Enlow DH, Brown SO. 1957. A comparative histological study of fossil and recent bone tissues.Part II. Texas Journal of Sciences 9: 185–214.
Enlow DH, Brown SO. 1958. A comparative histological study of fossil and recent bone tissues.Part III. Texas Journal of Sciences 10: 187–230.
Estes R. 1964. Fossil vertebrates from the Late Cretaceous Lance Formation, Eastern Wyoming.University of California Publications in the Geological Sciences 49: 1–180.
Francillon H. 1981. Contribution a l’etude des structures et du fonctionnement des epiphysesfemorales chez les Amphibiens Anoures. Archives de Zoologie, Experimentales et Generales 122: 289–310.
Gauthier JA. 1984. A phylogenetic analysis of the higher categories of Diapsida (Reptilia). Ph.D.Thesis, Department of Paleontology, University of California, Berkeley.
Girauld-Guille MM. 1988. Twisted plywood architecture of collagen fibrils in human compact boneosteons. Calcified Tissue International 42: 167–180.
Girauld MM, Castanet J, Meunier FJ, Bouligand Y. 1978. The fibrous structure of coelacanthscales: a twisted “plywood”. Tissue and Cell 10: 671–686.
Gross W. 1934. Die Typen des mikroskopischen Knochenbaues bei fossilen Stegocephalen undReptilien. Zeitschrift fur Anatomie 103: 731–764.
Haines RW. 1969. Epiphyses and sesamoids. In: Gans C, Bellairs Ad’A, Parsons TS, eds. Biology ofthe Reptilia: Morphology A, Volume 1. London: Academic Press, 81–115.
Haines RW, Mohuiddin A. 1967. Metaplastic bone. Journal of Anatomy 103: 527–538.Horner JR, Makela R. 1979. Nest of juveniles provides evidence of family structure among dinosaurs.
Nature 282: 296–298.Horner JR, Weishampel DB. 1988. A comparative embryological study of two ornithischian
dinosaurs. Nature 332: 256–257.Houck MA, Gauthier JA, Strauss RE. 1990. Allometric scaling in the earliest fossil bird, Archaeopteryx
lithographica. Science 247: 195–198.Karam GN, Gibson LJ. 1994. Biomimicking of animal quills and plant stems: natural cylindrical
shells with foam cores. Materials Science and Engineering C2: 113–132.Langston W. 1981. Pterosaurs. Scientific American 247: 122–136.Lempberg R. 1971. The subchondral bone plate of the femoral head in adult rabbits. Virchows Archiv,
Abt. A: Pathologisches Anatomie 352: 1–13, 16–25.Mateer NJ. 1976. A statistical study of the genus Pterodactylus. Bulletin of the Geological Institution of the
University of Uppsala, N.S. 6: 97–105.Meunier FJ. 1984. Spatial organization and mineralization of the basal plate of elasmoid scales in
osteichthyans. American Zoologist 24: 953–964.Meunier FJ, Castanet J. 1982. Organisation spaciale des fibres de collagene de la plaque basale
des ecailles des Teleosteens. Zoologica Scripta 11: 141–153.Meyer H von. 1859–60. Zur Fauna der Vorwelt, Vierte Abteilung. Frankfurt am Main: H. Keller.Mikic B, Carter DR, Padian K. 1998. Epigenetic mechanical factors in the evolution of long bone
epiphyses. Zoological Journal of the Linnean Society, London 123: 163–178.Owen R. 1870. Monograph on the Fossil Reptilia of the Liassic Formations. I. Part III. Plesiosaurs, Dimorphodon,
and Ichthyosaurs. London: Palaeontographical Society.Padian K. 1983. Osteology and functional morphology of Dimorphodon macronyx (Buckland) (Pterosauria:
Rhamphorhynchoidea) based on new material in the Yale Peabody Museum. Postilla 189: 1–44.Padian K. 1984. A large pterodactyloid from the Two Medicine Formation (Campanian) of Montana.
Journal of Vertebrate Paleontology 4: 516–524.Padian K. 1985. The origins and aerodynamics of flight in extinct vertebrates. Palaeontology 28:
413–433.Padian K. 1984. A large pterodactyloid pterosaur from the Two Medicine Formation (Campanian)
of Montana. Journal of Vertebrate Paleontology 4: 516–524.Padian K. 1987. The case of the bat-winged pterosaur: typological taxonomy and the influence of
pictorial representation on scientific presentation. In: Czerkas SM, Olson EC, eds. Dinosaurs Pastand Present, vol. 2. Seattle: University of Washington Press, 65–81.
Padian K. 1995. Pterosaurs and typology: archetypal physiology in the Owen-Seeley dispute of 1870.In: Sarjeant WAS, ed. Vertebrate Fossils and the Evolution of Scientific Concepts. Yverdon, Switzerland:Gordon and Breach, 285–298.
Padian K, Clemens WA. 1985. Terrestrial vertebrate diversity: episodes and insights. In: ValentineJW, ed. Phanerozoic Diversity Patterns. Princeton, New Jersey: Princeton University Press, 41–96.

A. J. DE RICQLES ET AL.384
Padian K, Smith M. 1992. New light on Late Cretaceous pterosaur material from Montana. Journalof Vertebrate Paleontology 12: 87–92.
Padian K, Rayner JMV. 1993. The wings of pterosaurs. American Journal of Science 293-A: 91–166.Padian K, de Ricqles A, Horner JR. 1995. Bone histology determines identification of a new fossil
taxon of pterosaur (Reptilia: Archosauria). Comptes Rendus de l’Academie des Sciences, Paris: Sciences dela Vie 320, IIA (1): 77–84.
Padian K, Smith M. 1992. New light on Late Cretaceous pterosaur material from Montana. Journalof Vertebrate Paleontology 12: 87–92.
Padian K, Van Der Meulen MCH, Carter DR. 1992. Pterosaurs as optical illusions: allometry,ontogeny, and the mechanics of bone. PaleoBios 14 (suppl. 1): 5.
Padian K, Warheit KI. 1989. Morphometrics of the pterosaur wing: one sharp division, few trends.Journal of Vertebrate Paleontology 9 (3, suppl.): 35A.
Quekett JT. 1849a. On the intimate structure of bone as composing the skeleton in the four greatclasses of animals, etc. Transactions of the Microscopical Society of London 2: 46–58.
Quekett JT. 1849b. Additional observations on the intimate structure of bone. Transactions of theMicroscopical Society of London 2: 59–64.
Quekett JT. 1855. Descriptive and illustrative catalogue of the histological series contained in the Museum of theRoyal College of Surgeons of England, vol. 2. London.
Rayner JMV. 1989a. Mechanics and physiology of flight in fossil and recent vertebrates. Transactionsof the Royal Society of Edinburgh, Earth Sciences 80: 311–320.
Rayner JMV. 1989b. Vertebrate flight and the origins of flying vertebrates. In: Allen KC, BriggsDEG, eds. Evolution and the Fossil Record. London: Belhaven Press, 188–217.
Reid REH. 1996. Bone histology of the Cleveland-Lloyd dinosaurs and of dinosaurs in general, PartI: Introduction: Introduction to bone tissues. Brigham Young University Geology Studies 41: 25–72.
Reid REH. 1997a. Histology of bones and teeth. In: Currie PJ, Padian K, eds. Encyclopedia of Dinosaurs.San Diego: Academic Press, 329–339.
Reid REH. 1997b. How dinosaurs grew. In: Farlow JO, Brett-Surman MK, eds. The Complete Dinosaur.Bloomington and Indianapolis: Indiana University Press, 403–413.
Reid REH. 1997c. Dinosaurian physiology: the case for “intermediate” dinosaurs. In: Farlow JO,Brett-Surman MK, eds. The Complete Dinosaur. Bloomington and Indianapolis: Indiana UniversityPress, 449–473.
Ricqles A de. 1972. Nature et signification des “surface epiphysaires” chez les tetrapodes fossiles.Comptes Rendus de l’Academie des Sciences, Paris D 274: 3527–3530.
Ricqles A de. 1974. Evolution of endothermy: histological evidence. Evolutionary Theory 1: 51–80.Ricqles A de. 1975. Sur la classification, la signification fonctionnelle et l’histoire des tissus osseux
des tetrapodes. Premiere partie: structures. Annales de Paleontologie (Vertebres) 61(1): 51–129.Ricqles A de. 1976. On bone histology of fossil and living reptiles, with comments on its functional
and evolutionary significance. In: Bellairs Ad’A, Cox CB, eds. Morphology and Biology of Reptiles.London: Academic Press, 123–150.
Ricqles A de. 1976–77. Sur la classification, la signification fonctionnelle et l’histoire des tissus osseuxdes tetrapodes. Deuxieme partie: fonctions. Annales de Paleontologie (Vertebres) 62(1): 71–126 [1976];63(1): 33–56 [1977]; 63(2): 133–160 [1977].
Ricqles A de. 1978. Sur la classification, la signification fonctionnelle et l’histoire des tissus osseuxdes tetrapodes. Troisieme partie: evolution. Annales de Paleontologie (Vertebres) 64(1): 85–111; 64(2):153–184.
Ricqles A de. 1979. Quelques remarques sur l’histoire evolutive des tissues squelettiques chez lesVertebres et plus particulierement chez les Tetrapodes. Annee Biologique 18: 1–35.
Ricqles A de. 1980. Tissue structures of dinosaur bone: functional significance and possible relationto dinosaur physiology. In: Thomas RDK, Olson EC, eds. A Cold Look at the Warm-Blooded Dinosaurs.Boulder: Westview Press, 103–139.
Ricqles A de, Horner JR, Padian K. 1998. Growth dynamics of the hadrosaurid dinosaur Maiasaurapeeblesorum. Journal of Vertebrate Paleontology 18 (3, suppl.): 72A.
Ricqles A de, Meunier FJ, Castanet J, Francillon-Viellot H. 1991. Comparative microstructureof bone. In: Hall BK, ed. Bone, Volume 3: Bone Matrix and Bone-Specific Products. Boca Raton, Florida:CRC Press, 1–78.
Ricqles A de, Padian K, Horner JR. 1997. Comparative biology and the bone histology of extincttetrapods: what does it tell us? Proceedings, Fifth International Congress of Vertebrate Morphology. Journal ofMorphology 232(3): 246 (abs.).

PTEROSAUR BONE HISTOLOGY 385
Riggs CM, Vaughan LC, Evans GP, Lanyon LE, Boyde A. 1993. Mechanical implications ofcollagen fibre orientation in cortical bone of the equine radius. Anatomy and Embryology 187: 239–248.
Rogers RR, LaBarbera M. 1993. Contribution of internal bony trabeculae to the mechanicalproperties of the humerus of the pigeon (Columba livia). Journal of Zoology (London) 230: 433–441.
Seeley HG. 1870. Remarks on Prof. Owen’s monograph on Dimorphodon. Annals and Magazine ofNatural History (4)6: 129–152.
Seitz AL. 1907. Vergleichende Studien uber den mikroskopischen Knochenbau fossiler und rezenterReptilien. Nova Acta Abhandlungen der Kaiserliches Leop.-Carol. Deutschen Akademie der Naturforschung 37:230–370.
Sereno PC. 1991. Basal archosaurs: phylogenetic relationships and functional implications. Memoir ofthe Society of Vertebrate Paleontology 2: 1–53.
Soemmerring ST von. 1817. Uber einen Ornithocephalus brevirostris der Vorwelt. Denkschriften derKoniglichen Bayerische Akademie der Wissenschaften (math.-phys. Klasse) 6: 89–104.
Unwin DM, Frey E, Martill DM, Clarke JB, Riess J. 1996. On the nature of the pteroid inpterosaurs. Proceedings of the Royal Society of London B 263: 45–52.
Van der Meulen MCH, Beaupre GS, Carter DR. 1993. Mechanobiologic influences in long bonecross-sectional growth. Bone 14: 635–642.
Van der Meulen MCH, Carter DR. 1995. Developmental mechanics determine long bone allometry.Journal of Theoretical Biology 172: 323–327.
Van der Meulen MCH, Padian K, Carter DR. 1992. Taxon-specific and functional adaptationcharacteristics of pterosaur bones. Transactions of the Orthopedic Research Society 17(2): 537.
Weishampel DB, Dodson P, Osmolska H (eds). 1990. The Dinosauria. Berkeley: University ofCalifornia Press.
Wellnhofer P. 1970. Die Pterodactyloidea (Pterosauria) der Oberjura-Plattenkalke Suddeutschlands.Bayerische Akademie der Wissenschaften, Mathematisch-Wissenschaftlichen Klasse, Abhandlungen 141: 1–113.
Wellnhofer P. 1971. Die Atoposauridae (Crocodylia, Mesosuchia) der Oberjura-Plattenkalke Bayerns.Palaeontographica A 138: 133–165.
Wellnhofer P. 1975. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Sud-deutschlands. Palaeontographica A 148: 1–33, 132–186; A 149: 1–30.
Wellnhofer P. 1985. Neue Pterosaurier aus der Santana-Formation der Chapada do Araripe, Brasilien.Palaeontographica A 187: 105–182.
Wellnhofer P. 1991. The Illustrated Encyclopedia of Pterosaurs. New York: Crescent Books.




















