
Newly discovered breast cancer susceptibility loci on 3p24 and 17q23.2
Upload
independentCategory
view
1download
0
Predictive Biomarkers and Personalized Medicine
A Genome-Wide Association Study Identifies Novel Loci forPaclitaxel-InducedSensory Peripheral Neuropathy inCALGB40101
R. Michael Baldwin1, Kouros Owzar5,6, Hitoshi Zembutsu11, Aparna Chhibber1, Michiaki Kubo11, Chen Jiang6,DorothyWatson6, Rachel J. Eclov1, JoelMefford2, Howard L.McLeod7, Paula N. Friedman8, Clifford A. Hudis9,Eric P. Winer10, Eric M. Jorgenson3,4, John S. Witte2,4, Lawrence N. Shulman10, Yusuke Nakamura11,Mark J. Ratain8, and Deanna L. Kroetz1,4
AbstractPurpose: Sensory peripheral neuropathy is a common and sometimes debilitating toxicity associated
with paclitaxel therapy. This study aims to identify genetic risk factors for the development of this toxicity.
Experimental Design: A prospective pharmacogenetic analysis of patients with primary breast cancer,
randomized to the paclitaxel arm of CALGB 40101, was used to identify genetic predictors of the onset and
severity of sensory peripheral neuropathy. A genome-wide association study in 855 subjects of European
ancestry was conducted and findings were replicated in additional European (n ¼ 154) and African
American (n ¼ 117) subjects.
Results: A single nucleotide polymorphism in FGD4was associated with the onset of sensory peripheral
neuropathy in the discovery cohort [rs10771973; HR, 1.57; 95% confidence interval (CI), 1.30–1.91; P ¼2.6� 10�6] and in a European (HR, 1.72; 95% CI, 1.06–2.80; P¼ 0.013) and African American (HR, 1.93;
95% CI, 1.13–3.28; P ¼ 6.7 � 10�3) replication cohort. There is also evidence that markers in additional
genes, including EPHA5 (rs7349683) and FZD3 (rs10771973), were associated with the onset or severity of
paclitaxel-induced sensory peripheral neuropathy.
Conclusions: A genome-wide association study has identified novel genetic markers of paclitaxel-
induced sensory peripheral neuropathy, including a common polymorphism in FGD4, a congenital
peripheral neuropathy gene. These findings suggest that genetic variation may contribute to variation in
development of this toxicity. Validation of these findings may allow for the identification of patients at
increased risk of peripheral neuropathy and inform the use of an alternative to paclitaxel and/or the clinical
management of this toxicity. Clin Cancer Res; 18(18); 5099–109. �2012 AACR.
IntroductionPaclitaxel is a useful microtubule-stabilizing agent with
efficacy in the treatment of many cancers. It is effective forthe treatment of breast cancer in the metastatic, adjuvant,and neoadjuvant settings (1, 2). Sensory peripheral neu-ropathy remains a significant issue in the clinical use of thisagent.More than 50%of patients experience some degree ofsensory peripheral neuropathy during their course of pac-litaxel treatment, with 5% to 30% experiencing grade 3 or4 toxicity (3, 4). Paclitaxel-induced sensory peripheralneuropathy is dose-, treatment schedule-, and infusiontime–dependent (3). Cumulative dose is a significant pre-dictor of sensory peripheral neuropathy, as is underlyingdiabetes and concurrent or previous administrationof otherdrugs associated with this toxicity. A recent study suggeststhat mild to moderate symptoms of sensory peripheralneuropathy can persist for up to 2 years following comple-tion of paclitaxel treatment (5). Long-term neuropathy is
Authors' Affiliations: Departments of 1Bioengineering and TherapeuticSciences,2Epidemiology and Biostatistics, 3Neurology, and 4Institute forHuman Genetics, University of California San Francisco, San Francisco,California; 5Department of Biostatistics and Bioinformatics and 6CALGBStatistical Center, Duke University, Durham; 7Department of Pharmaco-therapy and Experimental Therapeutics, University of North Carolina,Chapel Hill, North Carolina; 8Department of Medicine, University of Chi-cago, Chicago, Illinois; 9Memorial Sloan-Kettering Cancer Center, NewYork, New York; 10Dana-Farber Cancer Institute, Boston, Massachusetts;and 11University of Tokyo and Riken Center for Genomic Medicine, Yoko-hama, Japan
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
R.M. Baldwin and K. Owzar contributed equally to this work.
Corresponding Author: Deanna L. Kroetz, Department of Bioengineeringand Therapeutic Sciences, University of California San Francisco, 1550 4thStreet RH584E, San Francisco, CA 94158-2911. Phone: 415-476-1159;Fax: 415-514-4361; E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-12-1590
�2012 American Association for Cancer Research.
ClinicalCancer
Research
www.aacrjournals.org 5099
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590

particularly concerning for patients with primary breastcancer, such as those evaluated in the current study, asmorethan 80% will be long-term survivors whose quality of lifewill be compromised. Significant sensory peripheral neu-ropathy during paclitaxel treatment can lead to dose reduc-tions and treatment suspension, possibly resulting in sub-optimal disease treatment and the potential for an increasedlikelihood of relapse. A predictive marker for this dose-limiting toxicity would enable studies to identify whetheran individualized assessment of adverse event risk could beuseful in the clinical decision making process. It could alsoprovide a possible target for therapeutic interventions.
Substantial interindividual differences in the prevalence,reported and objective severity, and onset of peripheralneuropathy is consistent with an underlying genetic sus-ceptibility to this toxicity. CALGB 40101 is a phase IIIrandomized study comparing cyclophosphamide anddoxorubicin versus single-agent paclitaxel as adjuvant ther-apy for patients with breast cancer at relatively low risk forrelapse. In addition, the study compared short versus longertherapy of each regimen as a 2 � 2 factorial design. Apharmacogenetic companion study (CALGB 60202) wasincluded in this trial to prospectively evaluate germlinedeterminants of interindividual differences in response andtoxicity. An initial analysis of treatment outcome in CALGB40101 has shown no difference in response between the 4and 6 cycle treatment arms (6); additional analyses ofresponse await complete follow-up data. The goal of thispresent study was to identify genetic markers predictive ofsensory peripheral neuropathies in the paclitaxel treatmentarm of CALGB 40101 and to further our understanding ofthe underlying mechanism of injury and repair. Herein, wereport the results of a genome-wide association study
(GWAS) of 1,040 paclitaxel-treated women to identifynovel germline susceptibility loci associated with the devel-opment of sensory peripheral neuropathies. This representsthe largest prospective breast cancer pharmacogenetic studyof paclitaxel treatment toxicities to date and provides aparadigm for the identification of genetic markers withpotential clinical application in personalized medicine.
Materials and MethodsParticipants
All study participants were enrolled in CALGB 40101 andgave their additional consent to participate in the pharma-cogenetic companion study (CALGB 60202). CALGB40101 was open from May 15, 2002 until July 30, 2010.The final total accrual was 3,873 patients. Patients eligiblefor the treatment protocol were females with histologicallyconfirmed invasive carcinoma of the breast and 0 to 3axillary nodes positive for cancer. Eastern CooperativeOncology Group (ECOG) performance status of 0–1, ade-quate organ function, and absence of congestive heartfailure or myocardial infarction in the previous 6 monthswere required. Enrollment was required within 84 days ofbreast surgery (either modified radical mastectomy orlumpectomy) and the treatment began within 7 days ofregistration. Patients with locally advanced, inflammatory,or metastatic breast cancer or involvement of dermal lym-phatics were ineligible. Patients were disease-free from anyprior malignancies for at least 5 years. Previous trastuzu-mab, chemotherapy, or hormonal therapy, with the excep-tion of tamoxifen, for the current malignancy was notpermitted nor was anthracycline treatment for any previousdisease. Patients who received tamoxifen or any otherselective estrogen receptor modulators (SERM) for preven-tion or other indications (e.g., osteoporosis) were eligible.Treatment with tamoxifen, other SERMs, or exogenoushormones (e.g., hormone replacement therapy, oral contra-ceptives, raloxifene) was discontinued before enrollment.Trastuzumab was recommended for patients with HER2-positive disease. Patients could also enroll in adjuvantstudies of bisphosphonates or hormonal therapies (e.g.,ovarian suppression concurrent with chemotherapy). Allpatients provided written informed consent for both thetreatment and companion protocols that met state, federal,and institutional guidelines.
TreatmentPatients were randomly assigned with equal probability
to 4 or 6 cycles of cyclophosphamide/doxorubicin (AC) orpaclitaxel. A full description of the study design is includedin a recent publication describing the initial analysis oftreatment response (6). The first 570 patients were treatedwith AC every 3 weeks, or paclitaxel weekly for 12 or 18weeks. Thereafter, both regimens were administered every 2weeks for 4 or 6 cycles. Pharmacogenetic samples werecollected only from patients enrolled on every 2-week regi-mens, who received dose-dense paclitaxel for 4 or 6 cycles.Paclitaxel was given for more than 3 hours at 175 mg/m2
when given every 2 weeks. The 6 cycle treatment arms for
Translational RelevancePaclitaxel is widely used in the treatment of many
cancers, including breast cancer. Treatment with pacli-taxel is often limited by the development of peripheralneuropathies that can significantly impact a patient’squality of life. Biomarkers for the prediction of paclitax-el-induced peripheral neuropathy could be used to opti-mize the use of paclitaxel. A genome-wide genotypingapproach in women receiving single-agent paclitaxel asadjuvant therapy for breast cancer identified severalnovel genetic loci implicated in paclitaxel-induced sen-sory peripheral neuropathy. In particular, a commongenetic variant in FGD4, a causal gene for the congenitalperipheral neuropathy Charcot–Marie–Tooth disease,was associated with increased onset of neuropathy inboth Europeans and African Americans. This variant andothers identified in these studies could be validated asgenetic predictors of paclitaxel-induced sensory periph-eral neuropathy. The genetic variants identified in thesestudies will also lead to investigations into novel path-ways for this common chemotherapy-induced toxicity.
Baldwin et al.
Clin Cancer Res; 18(18) September 15, 2012 Clinical Cancer Research5100
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590
both drugs were closed after enrolling 3,172 patients.Arms were stratified by menopausal, estrogen receptor(ER), progesterone receptor (PR), and HER2 status.Patient demographics are shown in Table 1. Premedica-tion recommendations for the initial dose were 12.5 to 50mg diphenhydramine and 50 mg ranitidine, 300 mgcimetidine, or 20 mg famotidine administered i.v. 30 to60 minutes before paclitaxel. Dexamethasone was givenas a 10 mg i.v. dose within 60 minutes of paclitaxel oralternatively, as a 10 mg or 20 mg oral dose more thanone hour before paclitaxel. To facilitate the 14-day dosing
schedule, filgrastim was recommended on days 3 to 10 ofeach cycle (5 mg/kg rounded to either 300 or 480 mg).Sargramostim (250–500 mg/m2, days 3–10) or pegfilgras-tim (6 mg s.c., 24–36 hours after paclitaxel) could be usedin place of filgrastim. The treating physician could omitgranulocyte colony-stimulating factor (G-CSF) treatmentwhen confident neutrophils would recover within 14days; however, if treatment could not be delivered onschedule, then a G-CSF was required in subsequent cycles.Erythropoetin was permitted at the discretion of thetreating physician. Patients positive for HER2 by either
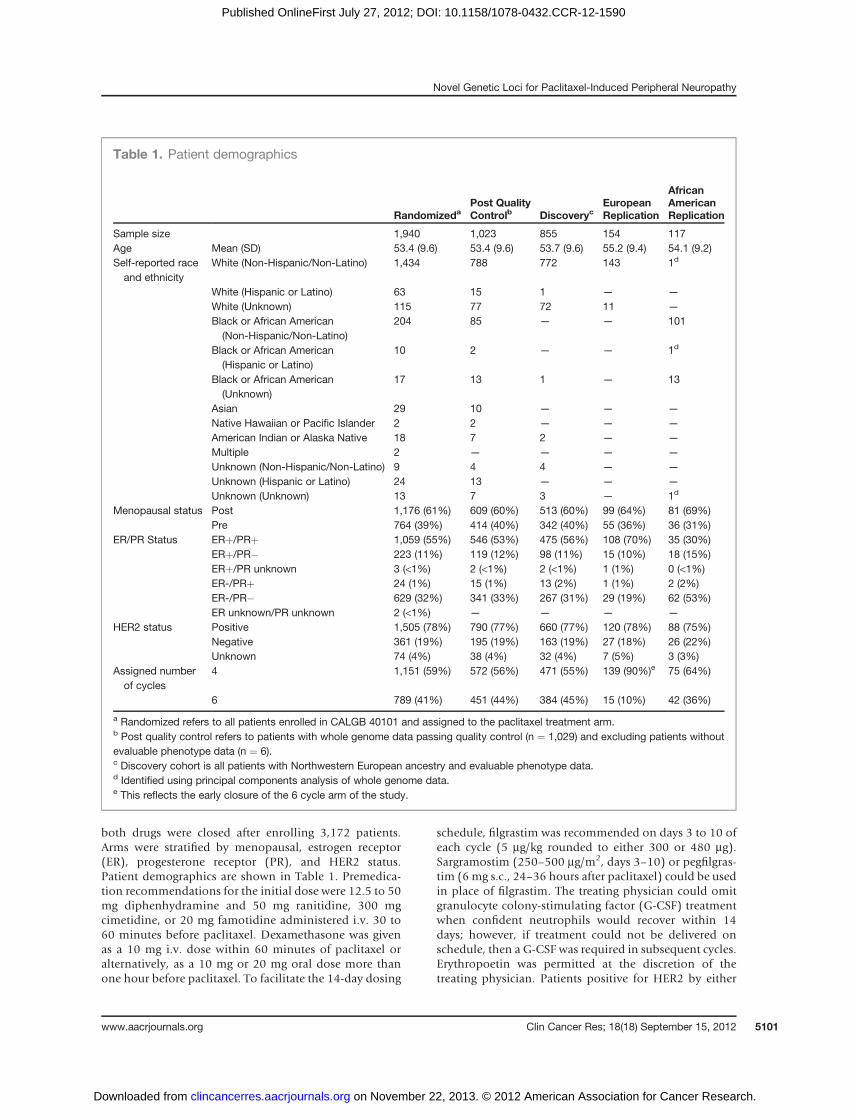
Table 1. Patient demographics
RandomizedaPost QualityControlb Discoveryc
EuropeanReplication
AfricanAmericanReplication
Sample size 1,940 1,023 855 154 117Age Mean (SD) 53.4 (9.6) 53.4 (9.6) 53.7 (9.6) 55.2 (9.4) 54.1 (9.2)Self-reported raceand ethnicity
White (Non-Hispanic/Non-Latino) 1,434 788 772 143 1d
White (Hispanic or Latino) 63 15 1 — —
White (Unknown) 115 77 72 11 —
Black or African American(Non-Hispanic/Non-Latino)
204 85 — — 101
Black or African American(Hispanic or Latino)
10 2 — — 1d
Black or African American(Unknown)
17 13 1 — 13
Asian 29 10 — — —
Native Hawaiian or Pacific Islander 2 2 — — —
American Indian or Alaska Native 18 7 2 — —
Multiple 2 — — — —
Unknown (Non-Hispanic/Non-Latino) 9 4 4 — —
Unknown (Hispanic or Latino) 24 13 — — —
Unknown (Unknown) 13 7 3 — 1d
Menopausal status Post 1,176 (61%) 609 (60%) 513 (60%) 99 (64%) 81 (69%)Pre 764 (39%) 414 (40%) 342 (40%) 55 (36%) 36 (31%)
ER/PR Status ERþ/PRþ 1,059 (55%) 546 (53%) 475 (56%) 108 (70%) 35 (30%)ERþ/PR� 223 (11%) 119 (12%) 98 (11%) 15 (10%) 18 (15%)ERþ/PR unknown 3 (<1%) 2 (<1%) 2 (<1%) 1 (1%) 0 (<1%)ER-/PRþ 24 (1%) 15 (1%) 13 (2%) 1 (1%) 2 (2%)ER-/PR� 629 (32%) 341 (33%) 267 (31%) 29 (19%) 62 (53%)ER unknown/PR unknown 2 (<1%) — — — —
HER2 status Positive 1,505 (78%) 790 (77%) 660 (77%) 120 (78%) 88 (75%)Negative 361 (19%) 195 (19%) 163 (19%) 27 (18%) 26 (22%)Unknown 74 (4%) 38 (4%) 32 (4%) 7 (5%) 3 (3%)
Assigned numberof cycles
4 1,151 (59%) 572 (56%) 471 (55%) 139 (90%)e 75 (64%)
6 789 (41%) 451 (44%) 384 (45%) 15 (10%) 42 (36%)
a Randomized refers to all patients enrolled in CALGB 40101 and assigned to the paclitaxel treatment arm.b Post quality control refers to patients with whole genome data passing quality control (n ¼ 1,029) and excluding patients withoutevaluable phenotype data (n ¼ 6).c Discovery cohort is all patients with Northwestern European ancestry and evaluable phenotype data.d Identified using principal components analysis of whole genome data.e This reflects the early closure of the 6 cycle arm of the study.
Novel Genetic Loci for Paclitaxel-Induced Peripheral Neuropathy
www.aacrjournals.org Clin Cancer Res; 18(18) September 15, 2012 5101
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590

immunohistochemical 3þ staining or gene amplificationby FISH could initiate adjuvant trastuzumab concurrentwith paclitaxel (weekly administration) or at the com-pletion of paclitaxel (weekly or every 3 weeks). Weeklytrastuzumab consisted of a 4 mg/kg i.v. loading dosefollowed by weekly doses of 2 mg/kg and the 3-weekschedule of a loading dose of 8 mg/kg and 6 mg/kg every3 weeks for a total duration of one year.
Genotyping and quality controlA summary of the steps included in sample and single
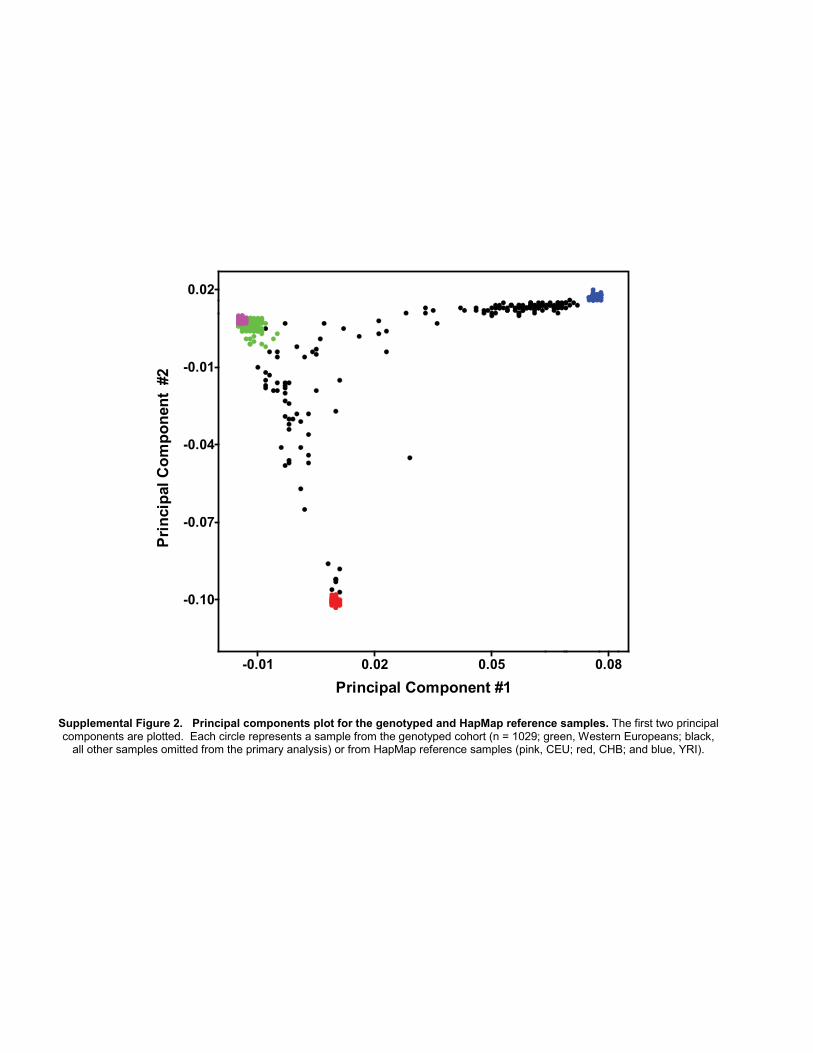
nucleotide polymorphism (SNP) quality control and inprincipal components analysis (PCA) is illustrated in Sup-plementary Fig. S1. A total of 1,040 paclitaxel-treatedpatients with informed consent and a DNA sample(obtained from peripheral blood) available as of July 1,2009 were included in the primary study. Genomic DNAwas genotyped using the HumanHap610-Quad Genotyp-ing BeadChip (Illumina), which interrogated 592,532SNPs. Subjects with call rates less than 0.98 (n¼ 5) or withsuboptimal genotype clustering performance (n ¼ 1) wereexcluded followed by reassessment of genotypes within theremaining subjects. SNPs with call rates less than 0.95, poorgenotype clustering performance, more than 1 replicate orMendelian discordance, relative minor allele frequency(MAF) less than 0.005, nondiploid (e.g., Y ormitochondrialchromosomes), or deemed unreliable by Illumina (n ¼4,106; Tech Note: Infinium Genotyping Data Analysis,2007) were excluded, leaving 572,745 SNPs. Identity-by-descent (IBD) analysis verified the absence of closely relatedindividuals (proportion IBD > 0.15) and identified oneunintended duplicate pair, which was removed and laterconfirmed to be due to a DNA plating error (PLINK version1.07; ref. 7). Evaluation of X-chromosome heterozygosityidentified 3 genetic males that were also removed andsimilarly confirmed to be due to a DNA plating error (8).PCA, as implemented by EIGENSOFT version 3.0, was usedto visualize the genetic ancestry of the 1,029 individualspassing quality control (9). PCA was conducted usinggenotypes from study subjects combined with genotypesof unrelated individuals from the HapMap Project repre-senting Northwest European (CEU, n ¼ 73), African (YRI,n ¼ 77), and Chinese (CHB, n ¼ 75) ancestries and geno-typed using the same platform by Illumina (SupplementaryFig. S2; ref. 10). To address the potential bias arising frompopulation stratification, we chose to focus our primaryanalysis on individuals of Northern European descent. Asecond PCAwas conducted using only 1,029 study subjects.Mean values for the first 3 eigenvectors within all patientsself-declaring "White" race and "Non-Hispanic" ethnicitywere determined. "Genetic Northwest Europeans" (hereincalled Europeans) were defined as individuals with each oftheir first 3 eigenvectors within 2 SDs of each mean valueirrespective of self-declared race and ethnicity. A total of 859individuals were identified and identical results wereobtained when repeated with the inclusion of HapMapindividuals (data not shown). These 859 individuals werethe focus of the primary analysis (Supplementary Fig. S2).
Imputation of genotypes was conducted within the 859Europeans using MACH 1.0 (11) and reference haplotypesfrom unrelated CEU individuals from either HapMap (r22)or the 1000 Genomes Project (June 2010 release). Beforeimputation, study genotypes were more stringently filteredand limited to autosomal SNPs withMAF 0.01 ormore andexact Hardy–Weinberg P values� 0.001 in control subjects.To address any potential stranding inconsistencies betweenstudy genotypes and the reference haplotypes, all symmetricSNPs (A/T or C/G) withMAFmore than 0.40, and thereforedifficult to resolve, were removed leaving 548,596 and547,465 SNPs for imputation using the HapMap and1000 Genomes reference haplotypes, respectively. ImputedSNPs with MAF less than 0.01 or R2 less than 0.5 wereexcluded. Genotyping within the replication cohorts(described below) was conducted using TaqMan AllelicDiscrimination assays (Applied Biosystems), and individ-ual assays are shown in Supplementary Tables S1 and S2.
One hundred fifty nine self-declared "White" individualswith either "Non-Hispanic" or "Unknown" ethnicity, whoenrolled in theCALGB40101pharmacogenetic companionstudy subsequent to the genotyping of the original 1,040subjects, were used as a replication cohort. Within thediscovery set, these criteria accurately identified 98.7% ofthe 859 Europeans with a false-positive rate of 2.4%. Anadditional 100 individuals of African ancestry were alsoidentified from within the group of 1,029 individualspassing sample quality control. African ancestrywas definedusing individuals who self-declared "Black/African Ameri-can" race with either "Non-Hispanic" or "Unknown" eth-nicity. Any individual with their first 3 eigenvectors within 3SDsof each eigenvectormeanvaluewere considered tobeofAfrican descent. This self-declared race/ethnicity criteriaidentified 94.2% of individuals with African ancestry andincorrectly identified 2.0%. The final African Americanreplication cohort consisted of the 100 patients of Africandescent with genome-wide data and an additional 20 self-declared "Black/African American" individuals with either"Non-Hispanic" or "Unknown" ethnicity, who enrolledafter the original genotyping.
Statistical analysisThe primary objective was the identification of SNPs
associated with the occurrence of sensory peripheral neu-ropathy. The analyses were carried out using 2 complemen-tary endpoints: (i) the cumulative dose level triggering thefirst grade II or higher treatment related sensory peripheralneuropathy episode and (ii) the maximum observed treat-ment-related sensory peripheral neuropathy grade. Theadverse events were graded according to the National Can-cer Institute Common Toxicity Criteria for Adverse Events(NCI-CTCAE) version 2.0. The timing of sensory peripheralneuropathy was assessed with a time-to-event approach inwhich an eventwas defined as the first incidence of a grade IIor higher neuropathy and the time as cumulative paclitaxelexposure (mg/m2). For patients not experiencing any event,the total study paclitaxel drug exposure was used. Thesepatients are effectively right-censored at the cumulative dose
Baldwin et al.
Clin Cancer Res; 18(18) September 15, 2012 Clinical Cancer Research5102
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590

level. The marginal associations were tested using the Coxscore test (12). The severity of sensory peripheral neurop-athy, defined as the maximum grade neuropathy observedduring paclitaxel treatment or within 30 days following thelast dose, was evaluated using ordinal logistic regression.Cumulative dose (mg/m2) was log-transformed and incor-porated into the ordinal regression model. For both cases,the marginal null sampling distribution was approximatedusing asymptotics. These analyses were powered for anadditive genetic model. To minimize type I error due tosparseness, SNPs within the European discovery set wereconstrained to relative MAF of 1% or more and the obser-vation of a minimum of 2 minor allele homozygous geno-types leaving 521,600 evaluable SNPs. Imputed genotypeswere represented as allele dosages bound between 0.0 and2.0. All analyses were conducted using the R statisticalenvironment version 2.12 with the cumulative dose-to-event and ordinal analyses implemented using functionsfrom the survival and MASS extension packages (13–16).Quantile-quantile plots of themarginal asymptotic P valueswere evaluated for potential remaining population stratifi-cation or inflation of significance levels. Each SNP with amarginal P value� 10�5 was evaluated further for potentialerrors by checking its MAF (vs. HapMap), Hardy–WeinbergEquilibrium within unaffected subjects, and potentiallyinformative missing rates; they were also visually inspectedfor genotype clustering performance.On the basis of the combined results of the time-to-event
and ordinal regression analyses of the 859 Europeanpatients, a replication plan delineating SNPs, regressionmodel, genetic model (the most plausible model suggestedfromKaplan–Meier estimates), and effect direction for one-sided testing was drafted a priori to any data collectionwithin the replication cohorts. Three SNPs from the genesFZD3, EPHA5, and FGD4 (rs7001034, rs7349683, andrs10771973) were selected for replication based on mar-ginal significance levels, biologic plausibility, and estimatedeffect size (as detailed in theResults). An additional 10 SNPswith P values < 10�5 and/or previously implicated incongenital sensory peripheral neuropathies (NDRG1) werealso evaluated with the specified limitation of being con-strained to exploratory analyses. Genotypes for the FZD3SNP rs7001034 were captured indirectly using a proxy SNP(rs7833751;R2¼ 1.0 CEUHapMap r27) due to the absenceof acceptable TaqMan assays to evaluate the locus directly.Because of the impracticality of capturing the FZD3 linkagedisequilibrium (LD) block to the same extent as the Euro-pean group, this locus was not evaluated in the Africanancestry replication group. Direct sequencing was used tocapture the FGD4 rs10771973 genotypes within the repli-cation cohorts. To limit the overall type I error rate for thevalidation study at the one-sided 0.05 level, we tested eachof the 3 SNPs at the marginal 0.01 level. Because the FGD4locus replicated in both populations and there are signif-icant differences in LD structure between the European andAfrican American populations, an additional 4 codingregion SNPs were chosen from the approximately 30 kbLD block containing rs10771973 to further extend this
finding. In addition, to evaluate the independence of theidentified association in rs10771973, the time-to-eventanalysis was repeated with rs10771973 as a covariate. Thisanalysis was conducted using the R extension packageGenABEL (13). A haplotype-based association test was alsoconducted for the 3 genes containing the top hits (EPHA5,FZD3, and FGD4), using all genotyped SNPs within 100 kbof the transcription start and stop sites for each gene. Phasefor each SNP set was estimated using fastPHASE v 1.1 in allsamples combined (17). Haplotype block boundaries usingthe method of Gabriel and colleagues were generated inHaploview v4.2 usingHapMap v3 r2CEU samples (18). Foreach haplotype block that included an allele with a per SNPassociation signal of less than 10�3, individual haplotypeswere extracted from fastPHASE output, andhaplotypeswithfrequency less than 5% were combined. Association withoutcome was analyzed on a per haplotype basis using time-to-event or maximum grade as described above.
ResultsOf the 859 individuals with European ancestry randomly
assigned to paclitaxel treatment, 4 withdrew before studytreatment and were therefore excluded (Supplementary Fig.S1). Patient characteristics of the CALGB 40101 paclitaxeltreatment arm, the genotyped samples, and the discoveryand replication cohorts are listed in Table 1. The meno-pausal, ER, PR andHER2 status, and the assigned number ofcycles were not different between the genotyped paclitaxelcohort and the European discovery cohort. The genotypedsamplewas also representative of all patients randomized topaclitaxel treatment in CALGB 40101. One exception is afewer number of samples from the 6 cycle paclitaxel armin the European replication cohort, which reflects theearly closure of the 6 cycle arm and the later study enroll-ment of this group of patients. Peripheral sensory neurop-athy was the major dose limiting toxicity in the paclitaxelarm, and the distribution of toxicity grades within the 855patients in the primary analysis, stratified for number oftreatment cycles assigned, is shown in Table 2. Sensoryperipheral neuropathy was dose dependent with 17% ofthe patients randomized to 4 cycles of paclitaxel experienc-ing a grade II or greater event as comparedwith 33%of thoserandomized to 6 cycles of treatment. The cumulative inci-dence of sensory peripheral neuropathy was similarbetween the entire cohort randomized to paclitaxel treat-ment and the discovery set (Supplementary Fig. S3), andbetween the discovery set and both replication groups (datanot shown). There was no effect of age on cumulative dosetriggering a grade II or greater peripheral neuropathy event(data not shown).
Among the SNPs analyzed in the GWAS for associationwith the initial onset of sensory peripheral neuropathy,none reached genome-wide significance although 7 had amarginal significance level of P < 10�5 (Table 3 and Sup-plementary Fig. S4). Inspection of the quantile-quantileplot of the marginal P values (Supplementary Fig. S5A)indicates the absence of any remaining population sub-structure (l ¼ 1.01). Of these top SNPs, biologic relevance
Novel Genetic Loci for Paclitaxel-Induced Peripheral Neuropathy
www.aacrjournals.org Clin Cancer Res; 18(18) September 15, 2012 5103
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590

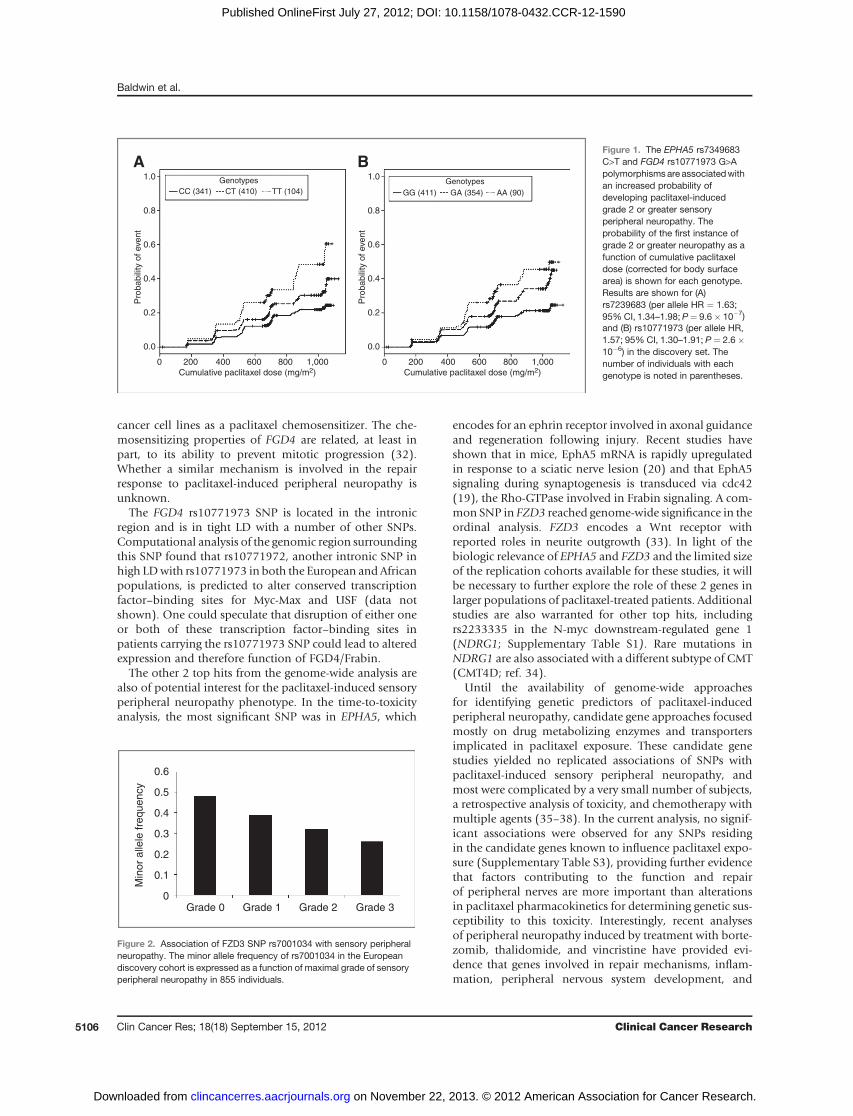
was apparent for polymorphisms in EPHA5 (rs7349683;per allele HR, 1.63; 95%CI, 1.34–1.98; P¼ 9.6� 10�7; Fig.1A) and FGD4 (rs10771973; per allele HR, 1.57; 95% CI,1.30–1.91; P ¼ 2.6 � 10�6; Fig. 1B). EPHA5 encodes anephrin receptor gene implicated in the process of neuronalregeneration following nerve injury and FGD4 encodes aRho-GTPase guanine nucleotide exchange factor previouslyimplicated in congenital peripheral neuropathies (19–22).The FGD4 (Table 3; Supplementary Fig. S6A and S6B) andEPHA5 (Table 3; Supplementary Fig. S7A and S7B) SNPswere tested in replication cohorts, and association for theformer was confirmed in both the European and AfricanAmerican samples (Europeans: rs10771973; per allele HR,1.72; 95% CI, 1.06–2.80; P ¼ 0.013; African Americans:rs10771973; per alleleHR, 1.93; 95%CI, 1.13–3.28;P¼6.7� 10�3). Considering the high minor allele frequency ofthis risk allele in Europeans, 42%of patients are expected tohave a 1.6-fold increased risk and 9% a 2.6-fold increasedrisk of peripheral neuropathy; in African Americans (MAF17%), the increased risk is 1.9- and 3.7-fold, respectively.Inspection of the Kaplan–Meier genotype stratified time toneuropathy distributions suggests that an allele dose–effectassumption for FGD4 rs10771973 is appropriate (Fig. 1B).
No haplotypes in FGD4 or EPHA5 showed stronger asso-ciation with time to sensory peripheral neuropathy than thesingle SNP analyses in these regions (data not shown). Afterconditioning the time-to-event analysis on rs10771973, noother genotyped markers at the FGD4 locus showed asso-ciation with time to peripheral neuropathy (data notshown). Using imputation to infer additional untypedmarkers andvisualizing theLDstructurewithin theHapMapCEU population revealed an approximately 30 kb region ofhigh LD within the FGD4 locus showing a strong andreproducible association with the onset of sensory periph-eral neuropathy (Supplementary Fig. S8). Approximately,16 SNPs are strongly linked (R2� 0.80) with rs10771973, 5of which are synonymous variants within the coding region.
Ordinal logistic regression analyses were used to identifySNPs associated with the severity of sensory peripheralneuropathy. Four SNPs were associated with toxicity gradewith a significance level of P < 1 � 10�5 (Table 4 andSupplementary Fig. S4). As with the Cox analysis, a quan-tile-quantile plot of the normalized marginal P values(Supplementary Fig. S5B) suggests the absence of anyremaining population substructure (l ¼ 0.986). A SNPwithin the Frizzled 3 homologWNT signaling receptor gene(FZD3) met the threshold of genome-wide significance(rs7001034; P ¼ 3.1 � 10�9; OR, 0.57; 95% CI, 0.48–0.69) and showed a clear relationship between allele dosageand sensory peripheral neuropathy grade (Fig. 2). However,none of these top SNPs from the ordinal regression analysisreplicated in either the European or African Americanpopulations (Table 4).
DiscussionA small subset of patients exposed to paclitaxel have
significant and occasionally protracted neuropathy thathas a major impact on quality of life. If we could prospec-tively identify these patients before administration ofpaclitaxel, they might be otherwise equally well servedwith alternative nonpaclitaxel containing regimens. Usinga genome-wide association study of CALGB 40101, we haveidentified several genetic loci associated with the onset orseverity of paclitaxel-induced sensory peripheral neuropa-thy. One of these novelmarkers associatedwith early-onset,paclitaxel-induced sensory peripheral neuropathy (FGD4,rs10771973) was replicated in both Europeans and AfricanAmericans and resides within a gene with a clearly estab-lished role in the hereditary peripheral neuropathy Char-cot–Marie–Tooth disease (CMT). These findings willinform studies to test the application of genetic markersfor optimization of paclitaxel selection, dosing, and adverseevent management. Several features of the study design andanalysis support the robustness of our findings, including
Table 2. Incidence of sensory peripheral neuropathies in study groups
Sensory peripheral neuropathy grade
0 1 2 3 4 Event rateb
Discovery set4 Cycles 181 (38%)a 209 (44%) 66 (14%) 15 (3%) 0 (0%) 17%6 Cycles 99 (26%) 160 (42%) 81 (21%) 44 (11%) 0 (0%) 33%European replication4 Cycles 44 (32%) 67 (48%) 24 (17%) 4 (3%) 0 (0%) 20%6 Cycles 6 (40%) 4 (27%) 3 (20%) 2 (13%) 0 (0%) 33%African American Replication4 Cycles 23 (31%) 33 (44%) 12 (16%) 7 (9%) 0 (0%) 25%6 Cycles 9 (21%) 19 (45%) 7 (17%) 6 (14%) 1 (2%) 33%
a Number of patients and percentage (in parentheses) of all patients in the discovery or replication cohort assigned to 4 or 6 cycles ofdose dense paclitaxel.b Event rate is the incidence of a grade 2 or greater sensory peripheral neuropathy.
Baldwin et al.
Clin Cancer Res; 18(18) September 15, 2012 Clinical Cancer Research5104
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590

the prospective design, a large cohort of patients withprimary breast cancer who are chemotherapy naive andtreated with single-agent paclitaxel, careful collection ofsensory peripheral neuropathy and covariate data, strictcensoring for dose and cycle reductions for other adversereactions and preexisting neuropathy, and the use of cumu-lative dose to the initial incidence of grade 2 toxicity toaccount for the established effect of total drug exposure onsensory peripheral neuropathy.
The current finding that FGD4 plays a role in the devel-opment of paclitaxel-induced sensory peripheral neuropa-thy and/or the repair response of peripheral nerves follow-ing paclitaxel injury is consistent with the known functionsof the gene. FGD4 encodes for the protein FGD1-related F-actin binding protein (Frabin), and previous studies haveshown specific point mutations in FGD4 can cause thecongenital peripheral neuropathy CMT (CMT4H; refs. 21–24). The disease is characterized by a slow progressivedemyelination of peripheral sensory and motor neuronsaccompanied by distal muscle weakness and atrophy, sen-sory loss, hyporeflexia, and skeletal deformity (25). Pacli-taxel-induced peripheral neuropathy shares some of thesecharacteristics, including sensory loss and secondary demy-elination (26–28). Frabin is a guanine nucleotide exchangefactor for cdc42, a Rho-GTPase that regulates cellular mor-phogenesis, including myelination. Several hypotheseshave been proposed to explain how mutations in FGD4might lead to demyelinating CMT4H disease, includingdisruption of the actin/microtubule cytoskeleton, loss ofc-Jun-NH-terminal kinase (JNK) activation signals, anddisruption of phosphoinositide signaling pathways, all ofwhich could affect Schwann cell myelination and/or thebidirectional communication between Schwann cells andaxons (21).
The observed association between the FGD4 SNPrs10771973 and paclitaxel-induced sensory peripheral neu-ropathy is consistent with the hypothesis that commonFGD4 polymorphisms subtly affect the development and/or maintenance of Schwann cell function. In this case,carriers of common FGD4 polymorphisms would havepreexisting subclinical abnormalities and a predispositionfor toxicity. This is supportedby increased risk for paclitaxel-induced sensory peripheral neuropathy in asymptomaticpatients with diabetes, previous platinum drug exposuresand alcohol use (3), and early Schwann cell activation inresponse to paclitaxel administration (29). Alternatively,FGD4 polymorphisms could lead to impaired repair pro-cesses such as Schwann cell remyelination and/or axonalregeneration after paclitaxel exposure. Genetic variation inFGD4 could also directly affect the response of Schwanncells to axonal injury via its ability to activate JNK (30). Aneuronal protective role for activated JNK in cultured dorsalroot ganglion cells exposed to oxaliplatin has been reported(31). Whether changes in frabin activity or expression leadto a decreased neuronal regenerative capacity and/or anincreased sensitivity to paclitaxel-induced sensory periph-eral neuropathy requires further study. Interestingly, FGD4was identified through a genome-wide siRNA screen in lung
Tab
le3.
TopSNPsfrom
cumulativedos
eto
even
tan
alysis
Disco
very
(n¼
855)
Europea
nreplic
ation
(n¼
154)
dAfrican
American
replic
ation
(n¼
117)
d
SNP
Chr
Gen
eaAlle
les
MAFb
HR
(95%
CI)
Pc
MAF
HR
(95%
CI)
PMAF
HR
(95%
CI)
P
rs73
4968
34
EPHA5
C/T
0.36
1.63
(1.34–
1.98
)9.6�
10�7
0.32
0.96
(0.57–
1.60
)0.43
0.13
1.16
(0.55–
2.42
)0.35
rs47
3726
48
XKR4
A/C
0.22
1.68
(1.36–
2.09
)1.9�
10�6
0.24
1.84
(1.02–
3.33
)0.02
10.18
1.23
(0.69–
2.21
)0.24
rs10
7719
7312
FGD4
G/A
0.31
1.57
(1.30–
1.91
)2.6�
10�6
0.33
1.72
(1.06–
2.80
)0.01
30.17
1.93
(1.13–
3.28
)6.7�
10�3
rs16
9487
4817
PITPNA
T/G
0.04
2.37
(1.63–
3.44
)2.7�
10�6
0.02
2.65
e(0.63–
11.1)
0.08
30.07
1.07
e(0.41–
2.77
)0.45
rs16
9169
3210
CACNB2
C/T
0.06
2.08
(1.51–
2.87
)4.3�
10�6
0.06
0.38
e(0.09–
1.58
)0.08
20.08
1.13
e(0.47–
2.74
)0.39
rs17
7810
8212
GRIP1/CAND1
C/T
0.42
1.60
(1.31–
1.96
)4.3�
10�6
0.43
1.22
(0.74–
1.99
)0.22
0.21
1.32
(0.76–
2.30
)0.16
rs19
0321
63
BCL6
/G/A
0.48
1.59
(1.30–
1.95
)5.6�
10�6
0.41
2.08
f(0.99–
4.37
)0.02
40.03
3.02
f(1.04–
8.73
)0.01
6rs22
3333
58
NDRG1
T/G
0.38
0.65
(0.52–
0.80
)5.2�
10�5
0.39
0.94
(0.58–
1.53
)0.41
0.23
1.40
(075
–2.60
)0.14
aIntergen
icSNPsareden
oted
bytheclos
estflan
king
anno
tatedge
ne(s).
bMAFwas
calculated
with
intheindicated
coho
rt.
cPva
lues
are2-sided
fordisco
very
analysis
andon
e-sided
forreplication.
dAsstated
inthereplicationplan,
analys
eswereex
ploratory
foralle
xcep
ttheEPHA5an
dFG
D4SNPs.
eAna
lysisas
sumed
ado
minan
tmod
el.
fAna
lysisas
sumed
arece
ssivemod
el.
Novel Genetic Loci for Paclitaxel-Induced Peripheral Neuropathy
www.aacrjournals.org Clin Cancer Res; 18(18) September 15, 2012 5105
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590
cancer cell lines as a paclitaxel chemosensitizer. The che-mosensitizing properties of FGD4 are related, at least inpart, to its ability to prevent mitotic progression (32).Whether a similar mechanism is involved in the repairresponse to paclitaxel-induced peripheral neuropathy isunknown.
The FGD4 rs10771973 SNP is located in the intronicregion and is in tight LD with a number of other SNPs.Computational analysis of the genomic region surroundingthis SNP found that rs10771972, another intronic SNP inhigh LDwith rs10771973 in both the European andAfricanpopulations, is predicted to alter conserved transcriptionfactor–binding sites for Myc-Max and USF (data notshown). One could speculate that disruption of either oneor both of these transcription factor–binding sites inpatients carrying the rs10771973 SNP could lead to alteredexpression and therefore function of FGD4/Frabin.
The other 2 top hits from the genome-wide analysis arealso of potential interest for the paclitaxel-induced sensoryperipheral neuropathy phenotype. In the time-to-toxicityanalysis, the most significant SNP was in EPHA5, which
encodes for an ephrin receptor involved in axonal guidanceand regeneration following injury. Recent studies haveshown that in mice, EphA5 mRNA is rapidly upregulatedin response to a sciatic nerve lesion (20) and that EphA5signaling during synaptogenesis is transduced via cdc42(19), the Rho-GTPase involved in Frabin signaling. A com-mon SNP in FZD3 reached genome-wide significance in theordinal analysis. FZD3 encodes a Wnt receptor withreported roles in neurite outgrowth (33). In light of thebiologic relevance of EPHA5 and FZD3 and the limited sizeof the replication cohorts available for these studies, it willbe necessary to further explore the role of these 2 genes inlarger populations of paclitaxel-treated patients. Additionalstudies are also warranted for other top hits, includingrs2233335 in the N-myc downstream-regulated gene 1(NDRG1; Supplementary Table S1). Rare mutations inNDRG1 are also associated with a different subtype of CMT(CMT4D; ref. 34).
Until the availability of genome-wide approachesfor identifying genetic predictors of paclitaxel-inducedperipheral neuropathy, candidate gene approaches focusedmostly on drug metabolizing enzymes and transportersimplicated in paclitaxel exposure. These candidate genestudies yielded no replicated associations of SNPs withpaclitaxel-induced sensory peripheral neuropathy, andmost were complicated by a very small number of subjects,a retrospective analysis of toxicity, and chemotherapy withmultiple agents (35–38). In the current analysis, no signif-icant associations were observed for any SNPs residingin the candidate genes known to influence paclitaxel expo-sure (Supplementary Table S3), providing further evidencethat factors contributing to the function and repairof peripheral nerves are more important than alterationsin paclitaxel pharmacokinetics for determining genetic sus-ceptibility to this toxicity. Interestingly, recent analysesof peripheral neuropathy induced by treatment with borte-zomib, thalidomide, and vincristine have provided evi-dence that genes involved in repair mechanisms, inflam-mation, peripheral nervous system development, and
0.6
0.5
0.4
0.3
0.2
0.1
0Grade 0 Grade 1 Grade 2 Grade 3
Min
or
alle
le fre
quency
Figure 2. Association of FZD3 SNP rs7001034 with sensory peripheralneuropathy. The minor allele frequency of rs7001034 in the Europeandiscovery cohort is expressed as a function of maximal grade of sensoryperipheral neuropathy in 855 individuals.
1.0
0.8
0.6
0.4
0.2
0.0
CC (341) CT (410) TT (104)
Genotypes
A B
GG (411) GA (354) AA (90)
Genotypes
Pro
babili
ty o
f eve
nt
1.0
0.8
0.6
0.4
0.2
0.0
0 200 400 600 800 1,000
Cumulative paclitaxel dose (mg/m2)
0 200 400 600 800 1,000
Cumulative paclitaxel dose (mg/m2)
Pro
babili
ty o
f eve
nt
Figure 1. The EPHA5 rs7349683C>T and FGD4 rs10771973 G>Apolymorphismsare associatedwithan increased probability ofdeveloping paclitaxel-inducedgrade 2 or greater sensoryperipheral neuropathy. Theprobability of the first instance ofgrade 2 or greater neuropathy as afunction of cumulative paclitaxeldose (corrected for body surfacearea) is shown for each genotype.Results are shown for (A)rs7239683 (per allele HR ¼ 1.63;95%CI, 1.34–1.98; P¼ 9.6� 10�7)and (B) rs10771973 (per allele HR,1.57; 95% CI, 1.30–1.91; P ¼ 2.6�10�6) in the discovery set. Thenumber of individuals with eachgenotype is noted in parentheses.
Baldwin et al.
Clin Cancer Res; 18(18) September 15, 2012 Clinical Cancer Research5106
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590

mitochondrial dysfunction could influence an individualpatient’s riskof developing toxicity (39–42).However, therewas no overlap of implicated genes with the current study(Supplementary Table S3), suggesting that the mechanismsunderlying this common toxicity might be drug specific.
To assess the potential translational implications of thisfinding to clinical practice, we estimated the cumulativedose level triggering an event for each FGD4 rs10771973genotype. Considering the data in Fig. 1B, to control theprobability of experiencing a neuropathy event at a criticalthreshold of 33%, the tolerated cumulative dose level forpatients with 2 copies of the risk allele is 710 mg/m2. Thecorresponding expected critical dose level for patients withone copy of the risk allele is increased to 877 mg/m2.Patients with no copies of the risk allele are expected totolerate more than 1047 mg/m2, corresponding to the fulldose of paclitaxel for 6 cycles. If these thresholds areprospectively validated and further refined in follow-upstudies, they may be used to estimate tolerable dose levelsbased on genotype and to tailor the treatment regimen.
While this pharmacogenetic study has several advantagesover previous studies on paclitaxel pharmacogenetics,including a large cohort of treatment-naive patients receiv-ing single-agent paclitaxel and a genome-wide approach todiscovery, several limitations also exist. Themost significantlimitation is the sole use of the NCI-CTC for assessment ofsensory peripheral neuropathy. It is widely recognized thatdetailed patient-reported symptomdata and a quality of lifeassessment more accurately describes this phenotype andthat physician-reported NCI-CTC grading underreportsperipheral neuropathy (43–45). However, it remains diffi-cult to apply these techniques across the multiple sites andlarge sample sizes required for the sufficient power forpharmacogenetic analyses. In a recent phase III study of1,060 women treated with taxanes, the Patient Neurotox-icity Questionnaire and the Functional Assessment of Can-cer Therapy-General were administered to only the first 300patients in the study (46). The only use of patient-reportedtoxicity data and symptom measurements for pharmaco-genetic analysis of taxane peripheral neuropathy is limitedby the very small sample size of the study (38). While it willbe important in follow-up studies to validate these findingsusing additional instruments, it should be noted thatdespite its limitations, the NCI-CTC scores are widelyaccepted for primary evaluation of treatment toxicity inlarge phase III studies such as CALGB 40101. A secondlimitation of the current study is the small sample size of thereplication cohorts, a common issue confronting almost allpharmacogenetic studies (47).
In summary, our findings support the use of prospectivepharmacogenetic analyses of well-phenotyped data setscollected under controlled clinical trial settings and unbi-ased genome-wide genetic approaches for the identificationof novel genes involved in drug efficacy and toxicity. Using aprospective design for validation and replication and awell-controlled single-agent clinical study, we have identified anSNP in FGD4 associated with increased risk of developingpaclitaxel-induced sensory peripheral neuropathy. The
Tab
le4.
TopSNPsfrom
ordinal
analysis
Disco
very
(n¼
855)
Europea
nRep
lication
(n¼
154)
dAfrican
American
Rep
lication(n
¼11
7)d
SNP
Chr
Gen
eaAlle
les
MAFb
OR
(95%
CI)
Pc
MAF
OR
(95%
CI)
PMAF
OR
(95%
CI)
P
rs70
0103
4e8
FZD3
G/T
0.40
0.57
(0.48–
0.69
)3.1�
10�9
rs78
3375
1f8
FZD3
G/T
0.40
0.58
(0.49–
0.70
)7.5�
10�9
0.41
1.67
f(0.88–
3.19
)0.05
8rs59
3468
3X
/SHROOM2
C/T
0.36
1.61
(1.33–
1.93
)6.0�
10�7
0.33
0.99
g(0.54–
1.82
)0.49
0.30
1.96
g(0.49–
7.89
)0.17
rs29
4162
78
/ZFP
M2
A/G
0.13
1.91
(1.40-2.51)
3.5�
10�6
0.15
1.37
g(0.68–
2.73
)0.19
0.25
0.49
g(0.24–
0.97
)0.02
1rs79
7353
312
/BCAT1
T/G
0.46
0.66
(0.55–
0.79
)8.4�
10�6
0.44
0.75
(0.47–
1.19
)0.11
0.27
1.13
(0.65–
1.94
)0.33
aIntergen
icSNPsareden
oted
bytheclos
estflan
king
anno
tatedge
ne.
bMAFwas
calculated
with
intheindicated
coho
rt.
cPva
lues
are2-sided
fordisco
very
analysis
andon
e-sided
forreplication.
dAsstated
inthereplicationplan,
alla
nalyse
swereex
ploratory
exce
ptfortheFZ
D3SNP.
eTh
erewas
noav
ailable
TaqMan
assa
yforrs70
0103
4to
usein
replicationstud
ies.
fTh
isSNPtags
rs70
0103
4in
theEurop
eanpop
ulationbu
tno
tin
theAfrican
American
pop
ulation.
gAna
lysisas
sumed
adom
inan
tmod
el.
Novel Genetic Loci for Paclitaxel-Induced Peripheral Neuropathy
www.aacrjournals.org Clin Cancer Res; 18(18) September 15, 2012 5107
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590

involvement of FGD4 in CMT disease, a congenital periph-eral neuropathy, provides strong evidence for the biologicsignificance of this finding. The fact that a common FGD4SNP is associated with an increased risk of paclitaxel-induced sensory peripheral neuropathy in patients withboth European and African ancestry makes it of potentiallybroad clinical significance. Additional SNPs in EPHA5 andFZD3 were also identified as potential risk factors for theonset and severity of sensory peripheral neuropathy. Addi-tional samples for extension and validation of these find-ings are currently being collected inongoingCALGB clinicaltrials of paclitaxel in the setting of metastatic breast cancer.
Disclosure of Potential Conflicts of InterestThe content of this manuscript is solely the responsibility of the authors
and does not necessarily represent the official views of the National CancerInstitute. No potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: R.M. Baldwin, K. Owzar, H.L. McLeod, P.N.Friedman, C.A. Hudis, E.P. Winer, L. Shulman, M.J. Ratain, D. KroetzDevelopment of methodology: R.M. Baldwin, K. Owzar, H. Zembutsu, M.Kubo, E. Jorgenson, J.S. Witte, D. KroetzAcquisitionofdata (provided animals, acquired andmanagedpatients,provided facilities, etc.): R.M. Baldwin, H. Zembutsu, R.J. Eclov, C.A.Hudis, E.P. Winer, L. Shulman, Y. Nakamura, D. Kroetz
Analysis and interpretation of data (e.g., statistical analysis, biosta-tistics, computational analysis): R.M. Baldwin, K. Owzar, H. Zembutsu,A. Chhibber, M. Kubo, C. Jiang, R.J. Eclov, J. Mefford, H.L. McLeod, C.A.Hudis, E. Jorgenson, J.S. Witte, L. Shulman, D. KroetzWriting, review, and/or revision of the manuscript: R.M. Baldwin, K.Owzar, A. Chhibber, C. Jiang, H.L. McLeod, P.N. Friedman, C.A. Hudis, E.P.Winer, E. Jorgenson, J.S. Witte, L. Shulman, M.J. RatainAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): R.M. Baldwin, K. Owzar, M. Kubo,D. Watson, H.L. McLeod, P.N. Friedman, C.A. HudisStudy supervision: C.A. Hudis, L. Shulman, Y. Nakamura, M.J. Ratain, D.Kroetz
Grant SupportThe research for CALGB 60202 and 40101 was supported, in part, by
grants from the National Cancer Institute (CA31946) to the Cancer andLeukemiaGroup B (toM.M. Bertagnolli) and to the CALGB Statistical Center(to D.J. Sargent, CA33601). This work was also supported in part by NIHgrants GM61390 and GM61393, and the Biobank Japan Project funded bythe Japanese Ministry of Education, Culture, Sports, and Science and Tech-nology. This work is part of the NIH Pharmacogenomics Research Network-RIKEN Center for Genomic Medicine Global Alliance. A. Chhibber and R.J.Eclov were supported in part by NIH Training Grant T32 GM007175.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received May 15, 2012; revised June 27, 2012; accepted July 17, 2012;published OnlineFirst July 27, 2012.
References1. Bishop JF, Dewar J, Toner GC, Smith J, Tattersall MH, Olver IN, et al.
Initial paclitaxel improves outcome compared with cmfp combinationchemotherapy as front-line therapy in untreated metastatic breastcancer. J Clin Oncol 1999;17:2355–64.
2. BuzdarAU,SingletarySE, Theriault RL,BooserDJ, ValeroV, IbrahimN,et al. Prospective evaluation of paclitaxel versus combination chemo-therapy with fluorouracil, doxorubicin, and cyclophosphamide asneoadjuvant therapy in patients with operable breast cancer. J ClinOncol 1999;17:3412–7.
3. Lee JJ, Swain SM. Peripheral neuropathy induced by microtubule-stabilizing agents. J Clin Oncol 2006;24:1633–42.
4. Rowinsky EK, Donehower RC. Paclitaxel (taxol). N Engl J Med 1995;332:1004–14.
5. HershmanDL,Weimer LH,Wang A, Kranwinkel G, Brafman L, FuentesD, et al. Association between patient reported outcomes and quan-titative sensory tests for measuring long-term neurotoxicity in breastcancer survivors treated with adjuvant paclitaxel chemotherapy.Breast Cancer Res Treat 2011;125:767–74.
6. Shulman LN, Cirrincione CT, Berry DA, Becker HP, Perez EA, O'ReganRO, et al. Six cycles of doxorubicin and cyclophosphamide or pacli-taxel are not superior to 4 cycles as adjuvant chemotherapy for breastcancer in women with 0–3 positive axillary nodes: Calgb 40101. J ClinOncol. 2012 Jul 23. [Epub ahead of print].
7. Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, BenderD, et al. Plink: A tool set for whole-genome association andpopulation-based linkage analyses. Am J Hum Genet 2007;81:559–75.
8. Weale ME. Quality control for genome-wide association studies.Methods Mol Biol 2010;628:341–72.
9. Price AL, Patterson NJ, Plenge RM, Weinblatt ME, Shadick NA, ReichD. Principal components analysis corrects for stratification in genome-wide association studies. Nat Genet 2006;38:904–9.
10. FrazerKA,BallingerDG,CoxDR,HindsDA,StuveLL,GibbsRA, et al. Asecond generation human haplotype map of over 3.1 million snps.Nature 2007;449:851–61.
11. Li Y, Willer CJ, Ding J, Scheet P, Abecasis GR. Mach: Using sequenceand genotype data to estimate haplotypes and unobserved geno-types. Genet Epidemiol 2010;34:816–34.
12. Cox DR. Regression models and life tables. J Royal Stat Soc Series B1972;34:187–220.
13. Aulchenko YS, Ripke S, Isaacs A, van Duijn CM. Genabel: An r libraryfor genome-wide association analysis. Bioinformatics 2007;23:1294–6.
14. Team RDC. R: A language and environment for statistical computing.Vienna, Austria: R Foundation for Statistical Computing; 2010.
15. Therneau TM, Grambsch PM. Modeling survival data: Extending thecox model. New York: Springer-Verlag; 2000.
16. VenablesWN,RipleyBD.Modern applied statisticswith S. 4th ed. NewYork: Springer; 2002.
17. Scheet P, Stephens M. A fast and flexible statistical model for large-scale population genotype data: Applications to inferring missinggenotypes and haplotypic phase. Am J Hum Genet 2006;78:629–44.
18. Gabriel SB, Schaffner SF, Nguyen H, Moore JM, Roy J, Blumenstiel B,et al. The structure of haplotype blocks in the human genome. Science2002;296:2225–9.
19. Akaneya Y, Sohya K, Kitamura A, Kimura F,Washburn C, Zhou R, et al.Ephrin-a5 and epha5 interaction induces synaptogenesis during earlyhippocampal development. PLoS One 2010;5:e12486.
20. Barrette B, Calvo E, Vallieres N, Lacroix S. Transcriptional profiling ofthe injured sciatic nerve of mice carrying the wld(s) mutant gene:Identification of genes involved in neuroprotection, neuroinflamma-tion, and nerve regeneration. Brain Behav Immun 2010;24:1254–67.
21. Delague V, Jacquier A, Hamadouche T, Poitelon Y, Baudot C, Boc-caccio I, et al. Mutations in fgd4 encoding the rho gdp/gtp exchangefactor frabin cause autosomal recessive charcot-marie-tooth type 4h.Am J Hum Genet 2007;81:1–16.
22. Stendel C, Roos A, Deconinck T, Pereira J, Castagner F, Niemann A,et al. Peripheral nerve demyelination caused by a mutant rho gtpaseguanine nucleotide exchange factor, frabin/fgd4. Am J Hum Genet2007;81:158–64.
23. Fabrizi GM, Taioli F, Cavallaro T, Ferrari S, Bertolasi L, Casarotto M,et al. Further evidence that mutations in fgd4/frabin cause charcot-marie-tooth disease type 4h. Neurology 2009;72:1160–4.
24. Houlden H, Hammans S, Katifi H, Reilly MM. A novel frabin (fgd4)nonsense mutation p.R275x associated with phenotypic variability incmt4h. Neurology 2009;72:617–20.
Baldwin et al.
Clin Cancer Res; 18(18) September 15, 2012 Clinical Cancer Research5108
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590

25. Patzko A, Shy ME. Update on charcot-marie-tooth disease. CurrNeurol Neurosci Rep 2011;11:78–88.
26. Lipton RB, Apfel SC, Dutcher JP, Rosenberg R, Kaplan J, Berger A,et al. Taxol produces a predominantly sensory neuropathy. Neurology1989;39:368–73.
27. Mielke S, Sparreboom A,Mross K. Peripheral neuropathy: A persistingchallenge in paclitaxel-based regimes. Eur J Cancer 2006;42:24–30.
28. Sahenk Z, Barohn R, New P, Mendell JR. Taxol neuropathy. Electro-diagnostic and sural nerve biopsy findings. Arch Neurol 1994;51:726–9.
29. Cavaletti G, Tredici G, Braga M, Tazzari S. Experimental peripheralneuropathy induced in adult rats by repeated intraperitoneal admin-istration of taxol. Exp Neurol 1995;133:64–72.
30. Parkinson DB, Bhaskaran A, Arthur-Farraj P, Noon LA, Woodhoo A,Lloyd AC, et al. C-jun is a negative regulator of myelination. J Cell Biol2008;181:625–37.
31. Scuteri A,Galimberti A,MaggioniD,RavasiM,Pasini S,Nicolini G, et al.Role of mapks in platinum-induced neuronal apoptosis. Neurotoxicol-ogy 2009;30:312–9.
32. Whitehurst AW, Bodemann BO, Cardenas J, Ferguson D, Girard L,Peyton M, et al. Synthetic lethal screen identification of chemosensi-tizer loci in cancer cells. Nature 2007;446:815–9.
33. EndoY,BeauchampE,WoodsD, TaylorWG,Toretsky JA,UrenA, et al.Wnt-3a and dickkopf-1 stimulate neurite outgrowth in ewing tumorcells via a frizzled3- and c-jun n-terminal kinase-dependent mecha-nism. Mol Cell Biol 2008;28:2368–79.
34. Kalaydjieva L, Gresham D, Gooding R, Heather L, Baas F, de JongeR, et al. N-myc downstream-regulated gene 1 is mutated in hered-itary motor and sensory neuropathy-lom. Am J Hum Genet 2000;67:47–58.
35. Bergmann TK, Green H, Brasch-Andersen C, Mirza MR, Herrstedt J,Holund B, et al. Retrospective study of the impact of pharmacogeneticvariants on paclitaxel toxicity and survival in patients with ovariancancer. Eur J Clin Pharmacol 2011;67:693–700.
36. Leskela S, Jara C, Leandro-Garcia LJ, Martinez A, Garcia-Donas J,Hernando S, et al. Polymorphisms in cytochromes p450 2c8 and 3a5are associated with paclitaxel neurotoxicity. Pharmacogenomics J2011;11:121–9.
37. Marsh S, Paul J, King CR, Gifford G, McLeod HL, Brown R. Pharma-cogenetic assessment of toxicity and outcome after platinum plustaxane chemotherapy in ovarian cancer: The scottish randomised trialin ovarian cancer. J Clin Oncol 2007;25:4528–35.
38. Sissung TM, Mross K, Steinberg SM, Behringer D, Figg WD, Sparre-boom A, et al. Association of abcb1 genotypes with paclitaxel-medi-ated peripheral neuropathy and neutropenia. Eur J Cancer 2006;42:2893–6.
39. Broyl A, Corthals SL, Jongen JL, van der Holt B, Kuiper R, de Knegt Y,et al. Mechanisms of peripheral neuropathy associated with bortezo-mib and vincristine in patientswith newly diagnosedmultiplemyeloma:a prospective analysis of data from the hovon-65/gmmg-hd4 trial.Lancet Oncol 2010;11:1057–65.
40. Cavaletti G, Alberti P, Marmiroli P. Chemotherapy-induced peripheralneurotoxicity in the era of pharmacogenomics. Lancet Oncol2011;12:1151–61.
41. Favis R, Sun Y, van de Velde H, Broderick E, Levey L, Meyers M, et al.Genetic variation associatedwith bortezomib-induced peripheral neu-ropathy. Pharmacogenet Genom 2011;21:121–9.
42. Johnson DC, Corthals SL,Walker BA, Ross FM, GregoryWM, DickensNJ, et al. Genetic factors underlying the risk of thalidomide-relatedneuropathy in patients with multiple myeloma. J Clin Oncol 2011;29:797–804.
43. Cleeland CS, Farrar JT, Hausheer FH. Assessment of cancer-relatedneuropathy and neuropathic pain. Oncologist 2010;15 Suppl 2:13–8.
44. Kuroi K, Shimozuma K. Neurotoxicity of taxanes: Symptoms andquality of life assessment. Breast Cancer 2004;11:92–9.
45. Postma TJ, Heimans JJ. Grading of chemotherapy-induced peripheralneuropathy. Ann Oncol 2000;11:509–13.
46. Shimozuma K, Ohashi Y, Takeuchi A, Aranishi T, Morita S, Kuroi K,et al. Taxane-induced peripheral neuropathy and health-related qualityof life in postoperative breast cancer patients undergoing adjuvantchemotherapy: N-sas bc 02, a randomized clinical trial. Support CareCancer. 2012 May 15. [Epub ahead of print].
47. Motsinger-Reif AA, JorgensonE,RellingMV,KroetzDL,WeinshilboumR, Cox NJ, et al. Genome-wide association studies in pharmacoge-nomics: successes and lessons. Pharmacogenet Genom 2010. 2010Jul 15. [Epub ahead of print].
Novel Genetic Loci for Paclitaxel-Induced Peripheral Neuropathy
www.aacrjournals.org Clin Cancer Res; 18(18) September 15, 2012 5109
on November 22, 2013. © 2012 American Association for Cancer Research.clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 27, 2012; DOI: 10.1158/1078-0432.CCR-12-1590

Supplemental Material
This supplemental material has been provided by the authors to give readers additional information about their work.
Supplement to: A Genome-Wide Association Study Identifies Novel Loci for Paclitaxel-Induced Peripheral Neuropathy in CALGB 40101

Acknowledgments
We acknowledge the women who took part in CALGB 40101 and provided DNA and
consent for its use in genetic studies.
The following institutions participated in this study:
Bay Area Tumor Institute CCOP, Oakland, CA
Board of Regents of the University of Oklahoma, Oklahoma City, OK, Shubham Pant,
M.D., supported by CA37447
Cancer Centers of the Carolinas, Greenville, SC, Jeffrey K. Giguere, M.D., supported by
CA29165
Christiana Care Health Services, Inc. CCOP, Wilmington, DE, Stephen Grubbs, M.D.,
supported by CA45418
Dana-Farber Cancer Institute, Boston, MA, Harold J. Burstein, M.D., Ph.D., supported
by CA32291
Dartmouth Medical School-Norris Cotton Cancer Center, Lebanon, NH, Konstantin
Dragnev, M.D., supported by CA04326
Duke University Medical Center, Durham, NC, Jeffrey Crawford, M.D., supported by
CA47577
Grand Rapids Clinical Oncology Program, Grand Rapids, MI, Martin J. Bury, M.D.,
Green Mountain Oncology Group CCOP, Bennington, VT, Herbert L. Maurer, M.D.,
supported by CA35091
Heartland Cancer Research CCOP, St. Louis, MO, Alan P. Lyss, M.D., supported by
CA114558
Hematology-Oncology Associates of Central New York CCOP, Syracuse, NY, Jeffrey
Kirshner, M.D., supported by CA45389

Illinois Oncology Research Association, Peoria, IL, John W. Kugler, M.D., supported by
CA35113
Kansas City Community Clinical Oncology Program CCOP, Kansas City, MO, Rakesh
Gaur, M.D.
Massachusetts General Hospital, Boston, MA, Jeffrey W. Clark, M.D., supported by
CA32291
Memorial Sloan-Kettering Cancer Center, New York, NY, Clifford A. Hudis, M.D.,
supported by CA77651
Missouri Baptist Medical Center, St. Louis, MO, Alan P. Lyss, M.D., supported by
CA114558-02
Missouri Valley Consortium-CCOP, Omaha, NE, Gamini S. Soori, M.D.
Moores University of California at San Diego, San Diego, CA, Barbara A. Parker, M.D.,
supported by CA11789
Mount Sinai Medical Center, Miami, FL, Michael A. Schwartz, M.D., supported by
CA45564
Mount Sinai School of Medicine, New York, NY, Lewis R. Silverman, M.D., supported by
CA04457
Nevada Cancer Research Foundation CCOP, Las Vegas, NV, John A. Ellerton, M.D.,
supported by CA35421
New Hampshire Oncology-Hematology PA, Concord, NH, Douglas J. Weckstein, M.D.
Northern Indiana Cancer Research Consortium CCOP, South Bend, IN, Rafat Ansari,
M.D., supported by CA86726
Rhode Island Hospital, Providence, RI, William Sikov, M.D., supported by CA08025
Roswell Park Cancer Institute, Buffalo, NY, Ellis Levine, M.D., supported by CA59518
Sibley Memorial Hospital, Washington, DC, Frederick Barr, M.D.
Southeast Cancer Control Consortium Inc. CCOP, Goldsboro, NC, James N. Atkins,
M.D., supported by CA45808

State University of New York Upstate Medical University, Syracuse, NY, Stephen L.
Graziano, M.D., supported by CA21060
The Ohio State University Medical Center, Columbus, OH, Clara D. Bloomfield, M.D.,
supported by CA77658
University of California at San Diego, San Diego, CA, Barbara A. Parker, M.D.,
supported by CA11789
University of Texas Southwestern Medical Center, Dallas, TX, supported by CA37347
University of Chicago, Chicago, IL, Hedy L. Kindler, M.D., supported by CA41287
University of Illinois MBCCOP, Chicago, IL, David J. Peace, M.D., supported by
CA74811
University of Iowa, Iowa City, IA, Daniel A. Vaena, M.D., supported by CA47642
University of Maryland Greenebaum Cancer Center, Baltimore, MD, Martin Edelman,
M.D., supported by CA31983
University of Massachusetts Medical School, Worcester, MA, William V. Walsh, M.D.,
supported by CA37135
University of Minnesota, Minneapolis, MN, Bruce A. Peterson, M.D., supported by
CA16450
University of Missouri/Ellis Fischel Cancer Center, Columbia, MO, Michael C. Perry,
M.D., supported by CA12046
University of Nebraska Medical Center, Omaha, NE, Anne Kessinger, M.D., supported
by CA77298
University of North Carolina at Chapel Hill, Chapel Hill, NC, Thomas C. Shea, M.D.,
supported by CA47559
University of Vermont, Burlington, VT, Steven M. Grunberg, M.D., supported by
CA77406
Wake Forest University School of Medicine, Winston-Salem, NC, David D. Hurd, M.D.,
supported by CA03927

Walter Reed Army Medical Center, Washington, DC, Brendan M. Weiss, M.D.,
supported by CA26806
Washington University School of Medicine, St. Louis, MO, Nancy Bartlett, M.D.,
supported by CA77440
Weill Medical College of Cornell University, New York, NY, John Leonard, M.D.,
supported by CA07968
Western Pennsylvania Cancer Institute, Pittsburgh, PA, John Lister, M.D.
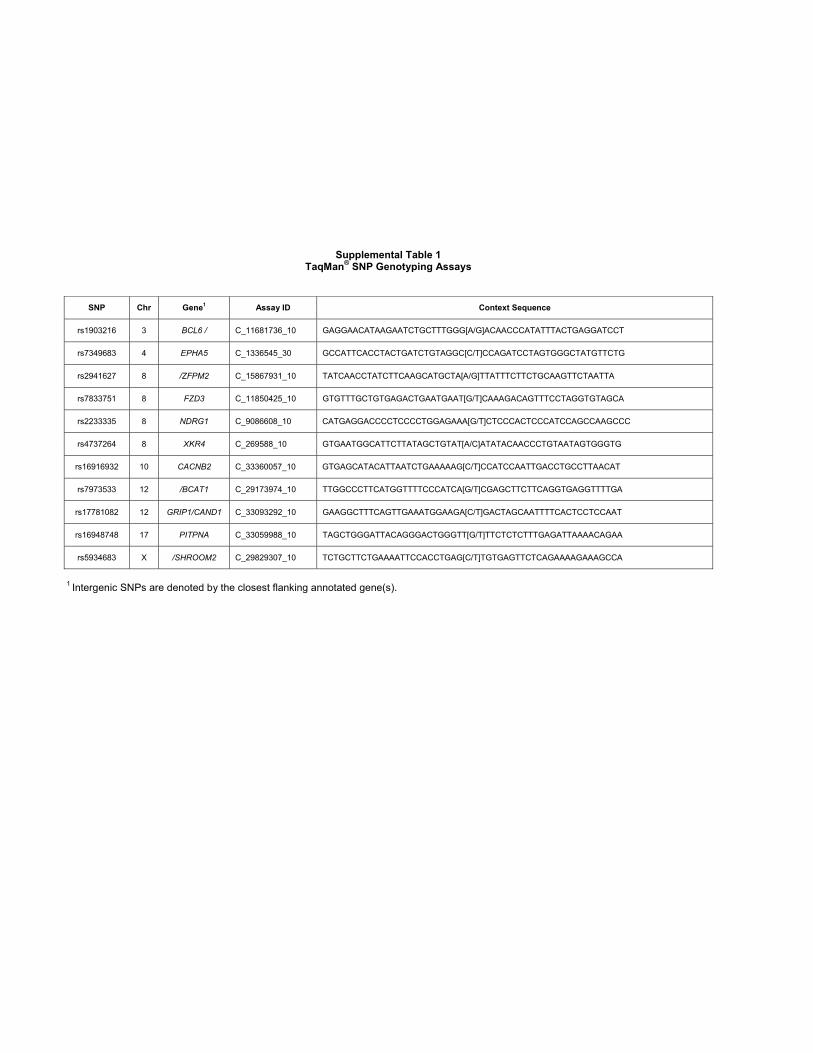
Supplemental Table 1TaqMan® SNP Genotyping Assays
SNP Chr Gene1 Assay ID Context Sequence
rs1903216 3 BCL6 / C_11681736_10 GAGGAACATAAGAATCTGCTTTGGG[A/G]ACAACCCATATTTACTGAGGATCCT
rs7349683 4 EPHA5 C_1336545_30 GCCATTCACCTACTGATCTGTAGGC[C/T]CCAGATCCTAGTGGGCTATGTTCTG
rs2941627 8 /ZFPM2 C_15867931_10 TATCAACCTATCTTCAAGCATGCTA[A/G]TTATTTCTTCTGCAAGTTCTAATTA
rs7833751 8 FZD3 C_11850425_10 GTGTTTGCTGTGAGACTGAATGAAT[G/T]CAAAGACAGTTTCCTAGGTGTAGCA
rs2233335 8 NDRG1 C_9086608_10 CATGAGGACCCCTCCCCTGGAGAAA[G/T]CTCCCACTCCCATCCAGCCAAGCCC
rs4737264 8 XKR4 C_269588_10 GTGAATGGCATTCTTATAGCTGTAT[A/C]ATATACAACCCTGTAATAGTGGGTG
rs16916932 10 CACNB2 C_33360057_10 GTGAGCATACATTAATCTGAAAAAG[C/T]CCATCCAATTGACCTGCCTTAACAT
rs7973533 12 /BCAT1 C_29173974_10 TTGGCCCTTCATGGTTTTCCCATCA[G/T]CGAGCTTCTTCAGGTGAGGTTTTGA
rs17781082 12 GRIP1/CAND1 C_33093292_10 GAAGGCTTTCAGTTGAAATGGAAGA[C/T]GACTAGCAATTTTCACTCCTCCAAT
rs16948748 17 PITPNA C_33059988_10 TAGCTGGGATTACAGGGACTGGGTT[G/T]TTCTCTCTTTGAGATTAAAACAGAA
rs5934683 X /SHROOM2 C_29829307_10 TCTGCTTCTGAAAATTCCACCTGAG[C/T]TGTGAGTTCTCAGAAAAGAAAGCCA
1 Intergenic SNPs are denoted by the closest flanking annotated gene(s).

Supplemental Table 2Custom TaqMan® SNP Genotyping Assays
SNP Chr Gene Forward Primer Reverse Primer Reporter 1 Sequence Reporter 2 Sequence
rs11052110 12 FGD4 TCACCCAGTGTGAATCATTAATATTA
TTTACATTCAAAAGGAAACAGAAATGAGGCTTACCT
5’-VIC-CCATTCTTCTTTGTCTTGTGC-NFQ-3’
5’-FAM-CATTCTTCTTTGTCTTGCGC-NFQ-3’
rs10844268 12 FGD4 GAGACACCTGACCTTTTACAGAACT GAGACCTTATTTAACCATGCTCGATT
ATCT5’-VIC-CCACATTTATCGTAATGTTT-NFQ-3’
5’-FAM-CACATTTATCGTCATGTTT-NFQ-3’
rs10844253 12 FGD4 GAACTAATAAAAGAAGGACAGATCC
TCAAACT GAAAAGGTAGCGTTCTTGTGCTG 5’-VIC-CTGCTCGGAACACTT-NFQ-3’ 5’-FAM-CTGCTCGAAACACTT-NFQ-3’
rs10771969 12 FGD4 AAGGTCCATTTGAGGCTAAGTGTAG TGTCCAATGAAGCATGTCTTTCCAT 5’-VIC-TCGTATGGCCTACCAAAC-
NFQ-3’5’-FAM-CGTATGGCCTTCCAAAC-NFQ-3’
rs7964947 12 FGD4 GTCATACCCCAAACATTGTCACTTTT AATGTGTGGCAGTCTCAAATTCTATTCT
5’-VIC-AAATCTACAAGAATGATCC-NFQ-3’
5’-FAM-ATCTACAAGGATGATCC-NFQ-3’

Supplemental Table 3Candidate Gene SNPs in Discovery Cohort
SNP Tag SNP1 R2 Gene/Allele2 SNP Location MAF3 Ordinal OR Ordinal P4 Cox HR Cox P
rs7909236 - - CYP2C8*1B Promoter 0.25 0.76 0.010 0.82 0.090
rs17110453 - - CYP2C8*1C Promoter 0.13 1.10 0.47 1.15 0.30
rs11572103 Not Tagged - CYP2C8*2 I269F - - - - -
rs11572080 rs10509681 1 CYP2C8*3 R139K 0.10 1.25 0.13 1.21 0.23
rs1058930 - - CYP2C8*4 I264M 0.06 1.20 0.36 1.07 0.75
rs1113129 rs10882520 1 CYP2C8_HapC Promoter 0.22 0.97 0.78 0.99 0.92
rs2740574 rs917152 0.59 CYP3A4*1B Promoter 0.04 1.56 0.056 1.37 0.14
rs4986910 Not Tagged - CYP3A4*3 M445T - - - - -
rs776746 - - CYP3A5*3 Splicing 0.07 1.35 0.10 1.28 0.16
rs1128503 - - ABCB1 G412G 0.43 1.14 0.15 1.08 0.42
rs2032582 rs4148738 1 ABCB1 A893S 0.45 1.15 0.12 1.06 0.57
rs1045642 - - ABCB1 I1145I 0.47 0.94 0.51 0.99 0.94
rs2306283 rs4149040 1 SLCO1B1 N130D 0.40 1.00 0.98 0.95 0.62
rs11045819 rs17329885 1 SLCO1B1 P155T 0.17 0.83 0.12 0.82 0.16
rs4149056 rs1871395 1 SLCO1B1 V174A 0.16 1.41 0.005 1.33 0.021
rs60140950 rs7306033 0.94 SLCO1B3 G256A 0.15 0.80 0.080 0.90 0.47
rs72559743 Not Tagged - SLCO1B3 G522C - - - - -
rs7311358 rs4149119 1 SLCO1B3 M233I 0.15 1.06 0.62 0.94 0.64
rs4149117 - - SLCO1B3 S112A 0.15 1.06 0.62 0.94 0.64
rs363717 rs2482430 0.945 ABCA1 3' UTR 0.17 1.21 0.11 1.14 0.27
rs246220 - - ABCC1 Intron 0.12 0.91 0.50 0.84 0.29
rs2644983 - - ABCC1 Intron 0.11 0.95 0.71 1.04 0.77
rs3887412 rs2644983 1 ABCC1 Intron 0.11 0.95 0.71 1.04 0.77
rs3740066 rs3740067 1 ABCC2 I1324I 0.37 0.93 0.46 0.84 0.12
rs2274407 rs4148477 1 ABCC4 K304K/K304N 0.07 0.94 0.72 0.89 0.54
rs3749442 - - ABCC5 L1208L 0.19 1.18 0.15 1.03 0.80
rs1042713 - - ADRB2 G16R 0.35 1.05 0.59 1.13 0.25
rs1042714 - - ADRB2 Q27E 0.44 1.02 0.79 0.99 0.93
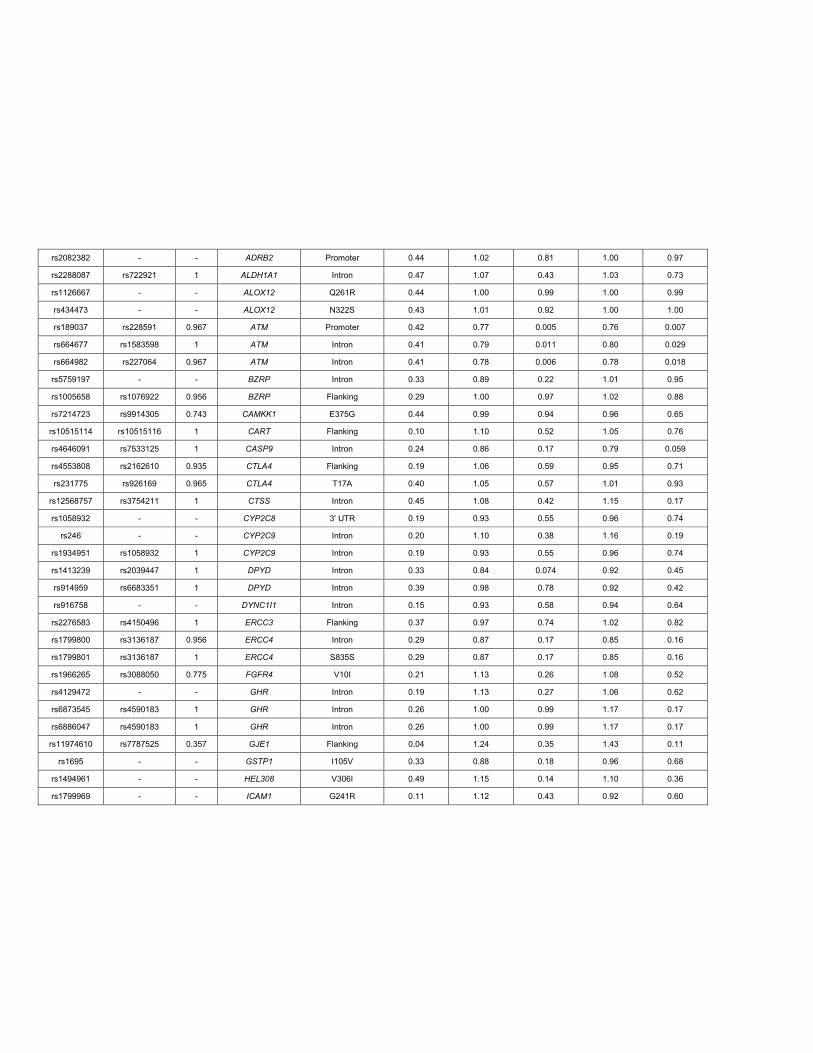
rs2082382 - - ADRB2 Promoter 0.44 1.02 0.81 1.00 0.97
rs2288087 rs722921 1 ALDH1A1 Intron 0.47 1.07 0.43 1.03 0.73
rs1126667 - - ALOX12 Q261R 0.44 1.00 0.99 1.00 0.99
rs434473 - - ALOX12 N322S 0.43 1.01 0.92 1.00 1.00
rs189037 rs228591 0.967 ATM Promoter 0.42 0.77 0.005 0.76 0.007
rs664677 rs1583598 1 ATM Intron 0.41 0.79 0.011 0.80 0.029
rs664982 rs227064 0.967 ATM Intron 0.41 0.78 0.006 0.78 0.018
rs5759197 - - BZRP Intron 0.33 0.89 0.22 1.01 0.95
rs1005658 rs1076922 0.956 BZRP Flanking 0.29 1.00 0.97 1.02 0.88
rs7214723 rs9914305 0.743 CAMKK1 E375G 0.44 0.99 0.94 0.96 0.65
rs10515114 rs10515116 1 CART Flanking 0.10 1.10 0.52 1.05 0.76
rs4646091 rs7533125 1 CASP9 Intron 0.24 0.86 0.17 0.79 0.059
rs4553808 rs2162610 0.935 CTLA4 Flanking 0.19 1.06 0.59 0.95 0.71
rs231775 rs926169 0.965 CTLA4 T17A 0.40 1.05 0.57 1.01 0.93
rs12568757 rs3754211 1 CTSS Intron 0.45 1.08 0.42 1.15 0.17
rs1058932 - - CYP2C8 3' UTR 0.19 0.93 0.55 0.96 0.74
rs246 - - CYP2C9 Intron 0.20 1.10 0.38 1.16 0.19
rs1934951 rs1058932 1 CYP2C9 Intron 0.19 0.93 0.55 0.96 0.74
rs1413239 rs2039447 1 DPYD Intron 0.33 0.84 0.074 0.92 0.45
rs914959 rs6683351 1 DPYD Intron 0.39 0.98 0.78 0.92 0.42
rs916758 - - DYNC1I1 Intron 0.15 0.93 0.58 0.94 0.64
rs2276583 rs4150496 1 ERCC3 Flanking 0.37 0.97 0.74 1.02 0.82
rs1799800 rs3136187 0.956 ERCC4 Intron 0.29 0.87 0.17 0.85 0.16
rs1799801 rs3136187 1 ERCC4 S835S 0.29 0.87 0.17 0.85 0.16
rs1966265 rs3088050 0.775 FGFR4 V10I 0.21 1.13 0.26 1.08 0.52
rs4129472 - - GHR Intron 0.19 1.13 0.27 1.06 0.62
rs6873545 rs4590183 1 GHR Intron 0.26 1.00 0.99 1.17 0.17
rs6886047 rs4590183 1 GHR Intron 0.26 1.00 0.99 1.17 0.17
rs11974610 rs7787525 0.357 GJE1 Flanking 0.04 1.24 0.35 1.43 0.11
rs1695 - - GSTP1 I105V 0.33 0.88 0.18 0.96 0.68
rs1494961 - - HEL308 V306I 0.49 1.15 0.14 1.10 0.36
rs1799969 - - ICAM1 G241R 0.11 1.12 0.43 0.92 0.60

rs1555026 - - ID3 Flanking 0.06 0.80 0.23 0.76 0.21
rs1059293 - - IFNGR2 3' UTR 0.43 1.08 0.39 1.12 0.26
rs1879612 - - IGF1R Intron 0.38 0.95 0.56 1.00 0.99
rs2272037 rs3784606 0.282 IGF1R Intron 0.26 0.95 0.63 0.86 0.18
rs11688 - - JUN Q250Q 0.05 1.09 0.69 1.02 0.92
rs9885672 - - KIAA0274 V654A 0.15 0.90 0.42 1.01 0.92
rs1805386 rs17498545 0.95 LIG4 D568D 0.17 1.07 0.55 1.09 0.47
rs7823144 - - LSM1 Intron 0.07 1.15 0.44 1.09 0.66
rs1041981 - - LTA T60N 0.32 1.07 0.49 0.99 0.90
rs2308327 rs2308321 1 MGMT K209R 0.13 0.99 0.93 0.84 0.25
rs10501815 rs10765682 1 MRE11A Intron 0.09 1.18 0.29 1.09 0.63
rs1029871 rs6976 1 NEK4 P225A 0.39 1.08 0.40 1.20 0.057
rs228832 rs228832 - NFATC2 Intron 0.23 1.00 0.98 1.02 0.86
rs3136794 rs2272733 1 POLB Intron 0.12 1.17 0.24 1.04 0.79
rs2076169 rs2076169 - PPARD Intron 0.12 1.15 0.33 1.14 0.41
rs2267668 rs2267668 - PPARD Intron 0.18 1.16 0.23 1.30 0.039
rs6457816 rs6457816 - PPARD Intron 0.07 0.85 0.37 0.91 0.65
rs6901410 rs6901410 - PPARD Intron 0.06 0.93 0.73 1.05 0.82
rs7739752 rs6919334 1 PPARD Intron 0.06 0.93 0.70 1.03 0.91
rs3735481 rs3801403 1 PPIA Intron 0.17 1.17 0.20 1.02 0.87
rs1474642 rs4710834 0.486 PSMB1 Intron 0.22 1.02 0.87 1.19 0.15
rs1236913 rs1236913 - PTGS1 W8R 0.07 0.99 0.95 0.98 0.93
rs4752904 rs10838852 0.935 PTPRJ E872D 0.46 1.00 0.98 0.93 0.49
rs1130499 rs1130500 1 PTPRN2 S325N 0.29 1.00 0.97 0.88 0.27
rs2251660 rs2251660 - RDM1 C127W 0.15 0.92 0.53 0.79 0.11
rs6131 rs6131 - SELP S331N 0.18 0.87 0.25 0.88 0.33
rs6103 rs1916659 1 SERPINB2 N404K 0.22 0.88 0.25 1.06 0.62
rs7242 rs7242 - SERPINE1 3' UTR 0.43 1.00 0.96 0.99 0.91
rs8192341 rs4715 1 SFTPC T138N 0.26 0.81 0.043 0.68 0.0014
rs3734354 rs3734354 - SIM1 P352T 0.16 1.00 0.99 0.94 0.65
rs2301157 rs2301157 - SLC10A2 3' UTR 0.43 1.12 0.22 1.30 0.009
rs3803258 rs3803258 - SLC10A2 3' UTR 0.20 0.91 0.40 0.95 0.68
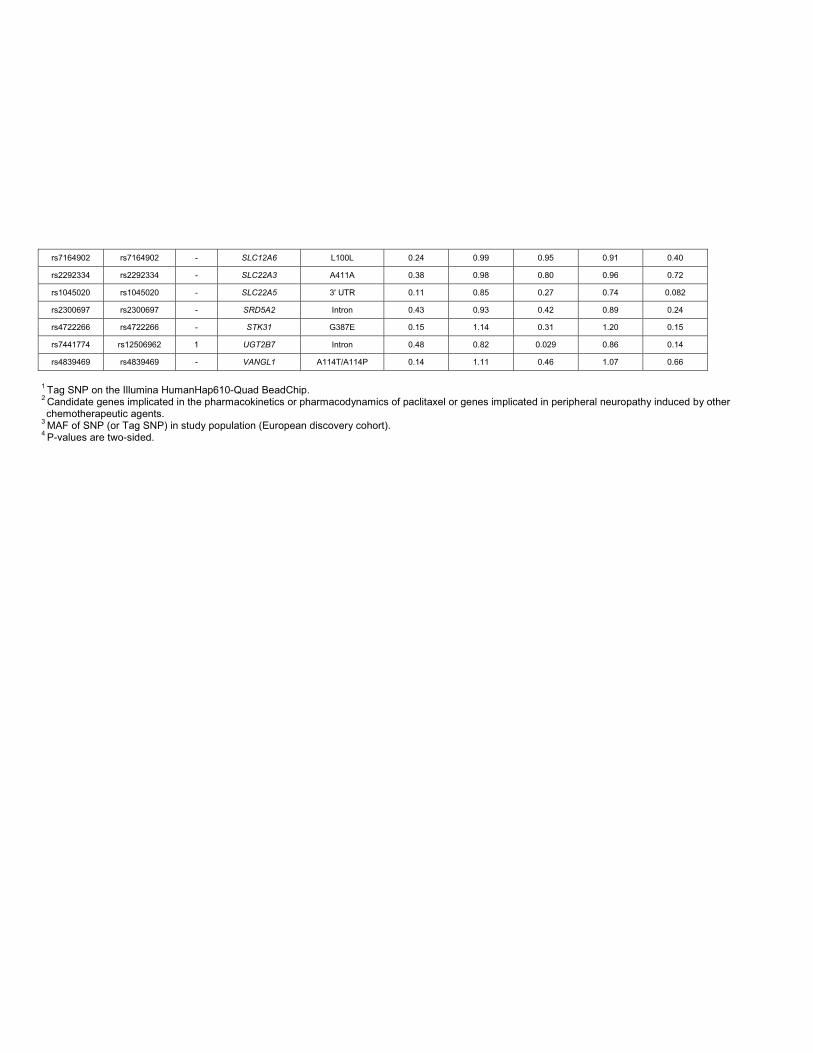
rs7164902 rs7164902 - SLC12A6 L100L 0.24 0.99 0.95 0.91 0.40
rs2292334 rs2292334 - SLC22A3 A411A 0.38 0.98 0.80 0.96 0.72
rs1045020 rs1045020 - SLC22A5 3' UTR 0.11 0.85 0.27 0.74 0.082
rs2300697 rs2300697 - SRD5A2 Intron 0.43 0.93 0.42 0.89 0.24
rs4722266 rs4722266 - STK31 G387E 0.15 1.14 0.31 1.20 0.15
rs7441774 rs12506962 1 UGT2B7 Intron 0.48 0.82 0.029 0.86 0.14
rs4839469 rs4839469 - VANGL1 A114T/A114P 0.14 1.11 0.46 1.07 0.66
1 Tag SNP on the Illumina HumanHap610-Quad BeadChip.2 Candidate genes implicated in the pharmacokinetics or pharmacodynamics of paclitaxel or genes implicated in peripheral neuropathy induced by other chemotherapeutic agents.
3 MAF of SNP (or Tag SNP) in study population (European discovery cohort).4 P-values are two-sided.
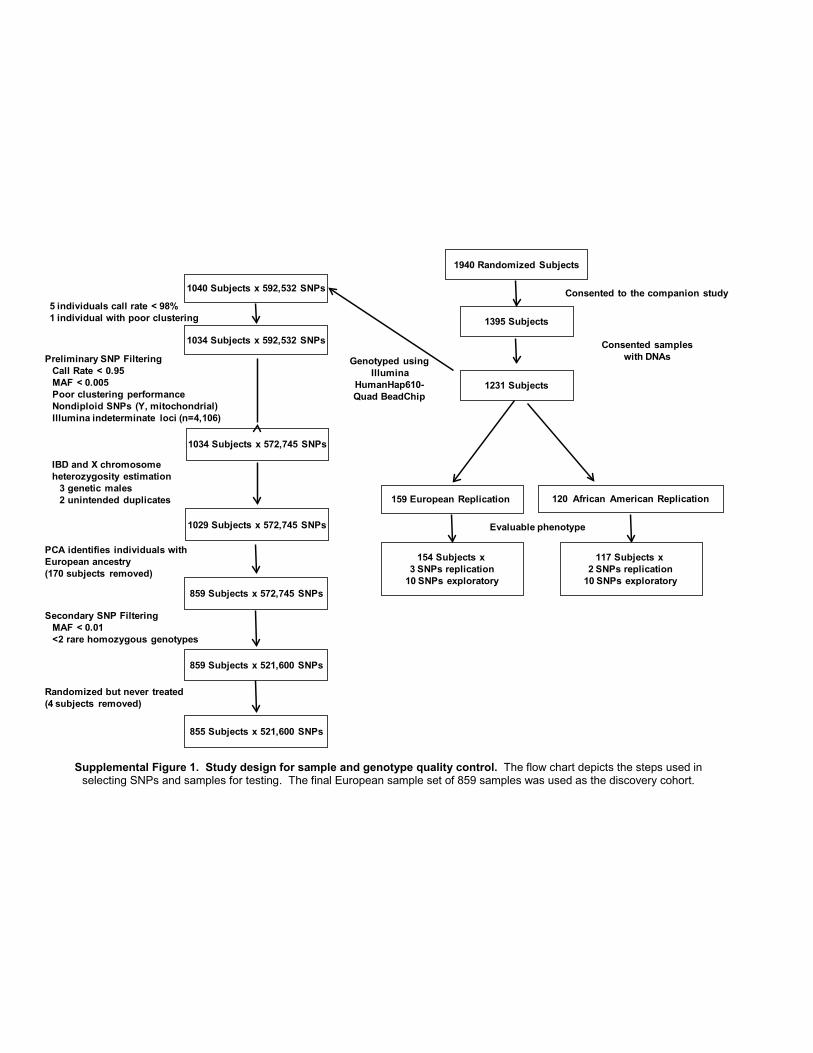
Supplemental Figure 1. Study design for sample and genotype quality control. The flow chart depicts the steps used in selecting SNPs and samples for testing. The final European sample set of 859 samples was used as the discovery cohort.
Preliminary SNP FilteringCall Rate < 0.95MAF < 0.005Poor clustering performanceNondiploid SNPs (Y, mitochondrial)Illumina indeterminate loci (n=4,106)
PCA identifies individuals with European ancestry(170 subjects removed)
1040 Subjects x 592,532 SNPs
Genotyped usingIllumina
HumanHap610-Quad BeadChip
1034 Subjects x 592,532 SNPs
1034 Subjects x 572,745 SNPs
1029 Subjects x 572,745 SNPs
859 Subjects x 572,745 SNPs
859 Subjects x 521,600 SNPs
5 individuals call rate < 98%1 individual with poor clustering
855 Subjects x 521,600 SNPs
Randomized but never treated(4 subjects removed)
1940 Randomized Subjects
Consented sampleswith DNAs
159 European Replication
1231 Subjects
154 Subjects x 3 SNPs replication
10 SNPs exploratory
Evaluable phenotype
120 African American Replication
117 Subjects x 2 SNPs replication
10 SNPs exploratory
1395 Subjects
Consented to the companion study
IBD and X chromosome heterozygosity estimation
3 genetic males2 unintended duplicates
Secondary SNP FilteringMAF < 0.01<2 rare homozygous genotypes
-0.01 0.02 0.05 0.08
-0.10
-0.07
-0.04
-0.01
0.02
Principal Component #1
Prin
cipa
l Com
pone
nt #
2
Supplemental Figure 2. Principal components plot for the genotyped and HapMap reference samples. The first two principal components are plotted. Each circle represents a sample from the genotyped cohort (n = 1029; green, Western Europeans; black,
all other samples omitted from the primary analysis) or from HapMap reference samples (pink, CEU; red, CHB; and blue, YRI).

Supplemental Figure 3. Cumulative probability plots for paclitaxel-induced sensory peripheral neuropathy. The probability of the first instance of grade 2 or higher sensory peripheral neuropathy is plotted as a function of the
cumulative paclitaxel dose for all subjects genotyped on the Illumina HumanHap610-Quad BeadChip passing QC and having complete phenotype data and for the subset of subjects with European ancestry used as the discovery set.
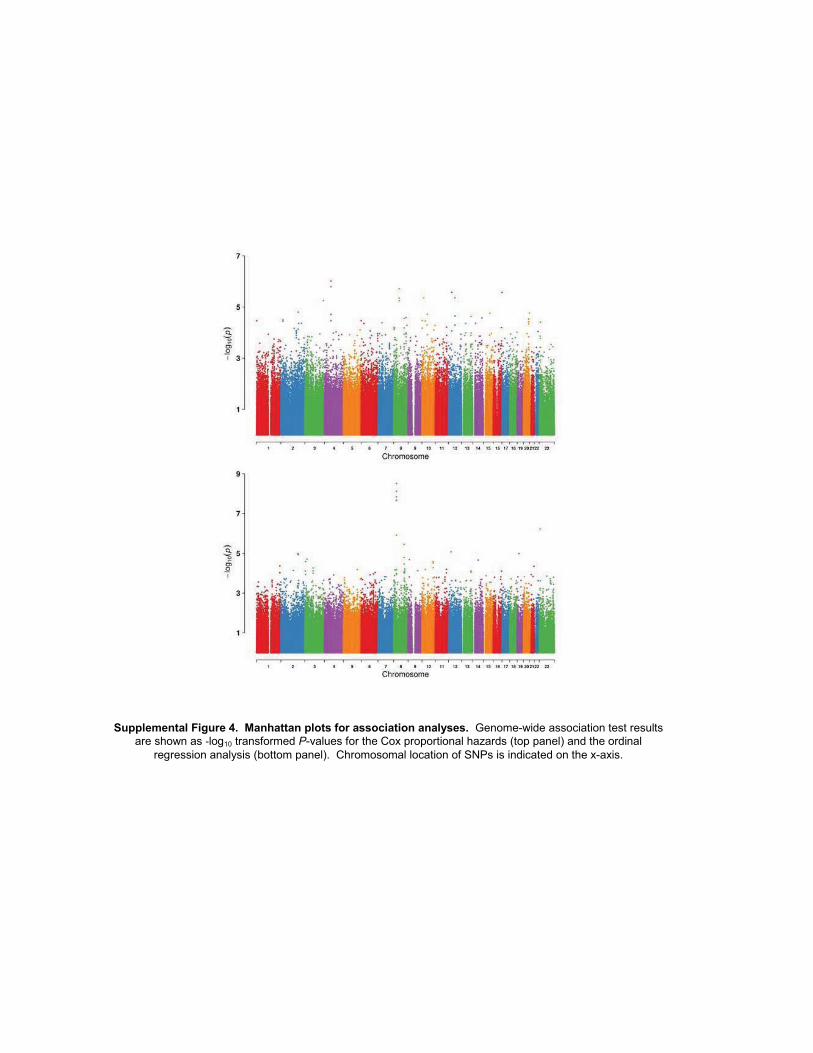
Supplemental Figure 4. Manhattan plots for association analyses. Genome-wide association test results are shown as -log10 transformed P-values for the Cox proportional hazards (top panel) and the ordinal
regression analysis (bottom panel). Chromosomal location of SNPs is indicated on the x-axis.

Supplemental Figure 5. Q-Q plots from association analyses. Observed and predicted P-value relationships are plotted for the A) Cox proportional hazards and the B) ordinal regression analyses. The solid line shows the expected distribution
assuming no inflation of statistics.
A. B.
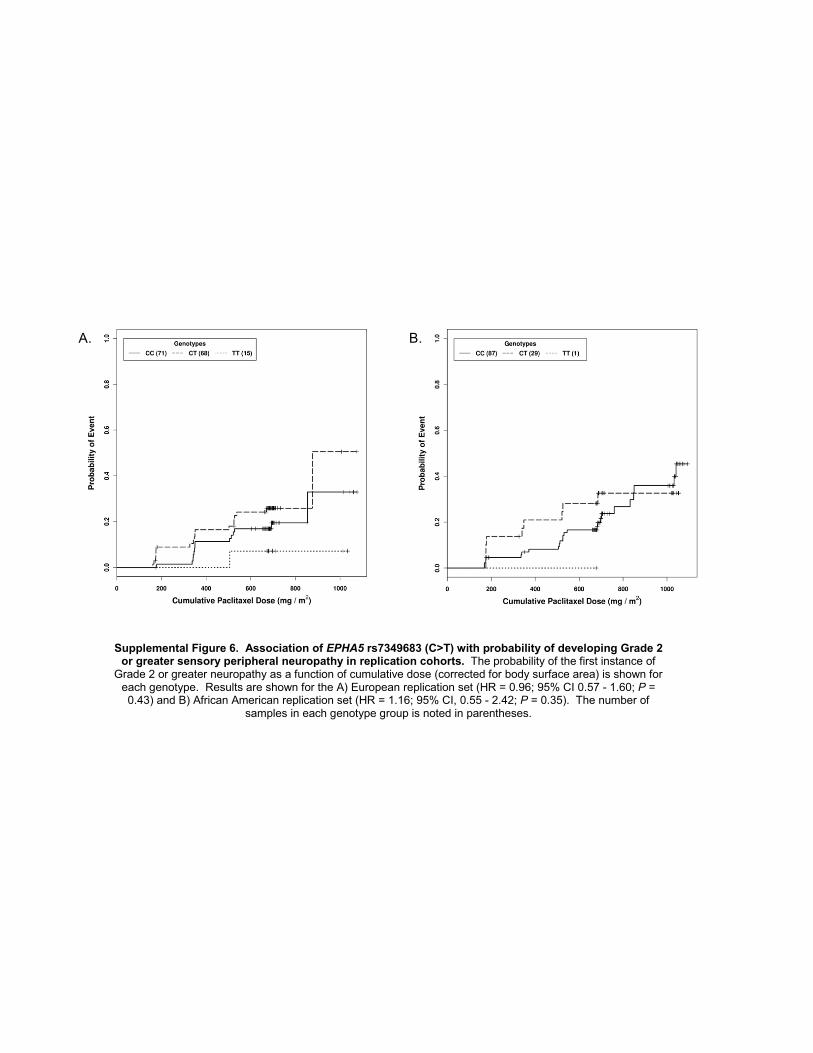
Supplemental Figure 6. Association of EPHA5 rs7349683 (C>T) with probability of developing Grade 2 or greater sensory peripheral neuropathy in replication cohorts. The probability of the first instance of
Grade 2 or greater neuropathy as a function of cumulative dose (corrected for body surface area) is shown for each genotype. Results are shown for the A) European replication set (HR = 0.96; 95% CI 0.57 - 1.60; P =0.43) and B) African American replication set (HR = 1.16; 95% CI, 0.55 - 2.42; P = 0.35). The number of
samples in each genotype group is noted in parentheses.
A. B.

Supplemental Figure 7. The FGD4 rs10771973 polymorphism is associated with an increased probability of developing paclitaxel-induced Grade 2 or greater sensory peripheral neuropathy in the replication cohorts. The probability of the first instance of Grade 2 or greater neuropathy as a function of
cumulative paclitaxel dose (corrected for body surface area) is shown for each genotype. Results are shown for the A) European replication set (HR, 1.72; 95% CI, 1.06 - 2.80; P = 0.013) and B) African American
replication set (HR, 1.93; 95% CI, 1.13 - 3.28; P = 6.7 x 10-3). The number of individuals with each rs10771973 genotype (G>A) is noted in parentheses.
A. B.

Supplemental Figure 8. Magnification of the FGD4 genomic region most strongly associated with the onset of sensory peripheral neuropathy. The P-values for genotyped and imputed SNPs within the ~150 kb
region surrounding rs10771973 highlights the positions and potential functional impact of SNPs in very high linkage disequilibrium with rs10771973. The genomic structure is noted below. SNPs genotyped on the HumanHap610-Quad BeadChip are red, SNPs from additional genotyping in blue, and imputed SNPs are unfilled. Circles represent SNPs in intronic or intergenic regions, squares indicate SNPs in the 3’-UTR and
diamonds are synonymous SNPs in the coding region.
Copyright © 2022 FDOKUMEN




















