
GDNF mediates glioblastoma-induced microglia attraction but not astrogliosis
14
1 23 Acta Neuropathologica Pathology and Mechanisms of Neurological Disease ISSN 0001-6322 Acta Neuropathol DOI 10.1007/s00401-013-1079-8 GDNF mediates glioblastoma-induced microglia attraction but not astrogliosis Min-Chi Ku, Susanne A. Wolf, Dorota Respondek, Vitali Matyash, Andreas Pohlmann, Sonia Waiczies, Helmar Waiczies, Thoralf Niendorf, et al.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of GDNF mediates glioblastoma-induced microglia attraction but not astrogliosis

1 23
Acta NeuropathologicaPathology and Mechanisms ofNeurological Disease ISSN 0001-6322 Acta NeuropatholDOI 10.1007/s00401-013-1079-8
GDNF mediates glioblastoma-inducedmicroglia attraction but not astrogliosis
Min-Chi Ku, Susanne A. Wolf, DorotaRespondek, Vitali Matyash, AndreasPohlmann, Sonia Waiczies, HelmarWaiczies, Thoralf Niendorf, et al.

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

ORIGINAL PAPER
GDNF mediates glioblastoma-induced microglia attractionbut not astrogliosis
Min-Chi Ku • Susanne A. Wolf • Dorota Respondek • Vitali Matyash •
Andreas Pohlmann • Sonia Waiczies • Helmar Waiczies • Thoralf Niendorf •
Michael Synowitz • Rainer Glass • Helmut Kettenmann
Received: 4 October 2012 / Accepted: 9 January 2013
� Springer-Verlag Berlin Heidelberg 2013
Abstract High-grade gliomas are the most common pri-
mary brain tumors. Their malignancy is promoted by the
complex crosstalk between different cell types in the cen-
tral nervous system. Microglia/brain macrophages infiltrate
high-grade gliomas and contribute to their progression. To
identify factors that mediate the attraction of microglia/
macrophages to malignant brain tumors, we established a
glioma cell encapsulation model that was applied in vivo.
Mouse GL261 glioma cell line and human high-grade
glioma cells were seeded into hollow fibers (HF) that allow
the passage of soluble molecules but not cells. The glioma
cell containing HF were implanted into one brain hemi-
sphere and simultaneously HF with non-transformed
fibroblasts (controls) were introduced into the contralateral
hemisphere. Implanted mouse and human glioma- but not
fibroblast-containing HF attracted microglia and up-regu-
lated immunoreactivity for GFAP, which is a marker of
astrogliosis. In this study, we identified GDNF as an
important factor for microglial attraction: (1) GL261 and
human glioma cells secret GDNF, (2) reduced GDNF
production by siRNA in GL261 in mouse glioma cells
diminished attraction of microglia, (3) over-expression of
GDNF in fibroblasts promoted microglia attraction in our
HF assay. In vitro migration assays also showed that
GDNF is a strong chemoattractant for microglia. While
GDNF release from human or mouse glioma had a pro-
found effect on microglial attraction, the glioma-induced
astrogliosis was not affected. Finally, we could show that
injection of GL261 mouse glioma cells with GDNF
knockdown by shRNA into mouse brains resulted in
reduced tumor expansion and improved survival as com-
pared to injection of control cells.
Keywords Glioblastoma � Microglia � GDNF �Astrocyte
Introduction
High-grade gliomas (HG-gliomas) are most aggressive
primary tumors of the central nervous system (CNS). HG-
gliomas are not a homogenous population of tumor cells,
but contain microglia which significantly contribute to the
actual tumor mass and are surrounded by activated astro-
cytes [4]. Microglia density in HG-gliomas can amount up
to 30 % of the tumor mass [25, 34]. However, unlike in an
inflamed tissue, immune functions in tumor-associated
microglia are often suppressed [10, 13, 32]. Therefore,
microglia do not attack glioma cells, but rather have been
shown to promote glioma growth. Altogether, the recent
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00401-013-1079-8) contains supplementarymaterial, which is available to authorized users.
M.-C. Ku � S. A. Wolf � D. Respondek � V. Matyash �H. Kettenmann (&)
Department of Cellular Neuroscience, Max Delbruck Center
for Molecular Medicine (MDC), Robert Rossle Str. 10,
13125 Berlin, Germany
e-mail: [email protected]
A. Pohlmann � S. Waiczies � H. Waiczies � T. Niendorf
Berlin Ultrahigh Field Facility (B.U.F.F.), Max Delbruck Center
for Molecular Medicine (MDC), 13125 Berlin, Germany
M. Synowitz
Department of Neurosurgery, Charite-Universitatsmedizin
Berlin, 13353 Berlin, Germany
R. Glass
Klinikum der Universitat Munchen (LMU), Klinik fur
Neurochirurgie, Neurochirurgische Forschung, Marchioninistr.
15, 81377 Munich, Germany
123
Acta Neuropathol
DOI 10.1007/s00401-013-1079-8
Author's personal copy

data by others and us indicate that blockade of microglia
infiltration into HG-gliomas could be a novel therapeutic
target in neuro-oncology [25, 26]. Earlier studies have
shown that reactive astrocytes are found in the glioma
environment and are associated with increased glioblas-
toma invasion [8]. Moreover, reactive astrocytes are highly
concentrated at the perivascular region of HG-gliomas and
may also contribute to glioma growth, e.g. mediated by the
release of TGF-b [6].
Recent studies focus on the factors that regulate the
cross talk between glioma cells and glial cells. There is a
positive correlation between the number of microglia and
the expression level of monocyte chemoattractant protein-3
(MCP-3) [28] or granulocyte/macrophage colony-stimu-
lating factor (GM-CSF) [18]. Hepatocyte growth factor [2]
and MCP-1 had also been identified as chemoattractant in
the glioma context [21, 29].
Glial-derived neurotrophic factor (GDNF) is a potent
trophic factor for a variety of neuronal cell populations in
the CNS. GDNF was first discovered in the conditioned
medium of the B49 rat glioma cell line [22]. Importantly,
GDNF is highly expressed in human HG-gliomas [35]. It is
not only a trophic factor, but also a chemoattractant for
various types of cells [5, 7, 17, 23, 33]. Although GDNF
has been studied for several years, and the correlation of
GDNF and glioma has been addressed, the role of its potent
chemoattractant factor in the crosstalk between glioma and
microglia has not been investigated.
In this study, we investigated the role of GDNF in
facilitating the attraction of microglia to the glioma. To
distinguish between the impact of soluble factors and cell
to cell contact, we have developed a method of implanting
encapsulated glioma cells into the mouse brains. Semi-
permeable hollow fibers (HF) allow the bi-directional
exchange of oxygen, nutrients, and growth factors, but
prevent physical contact between the encapsulated cells
and their environment [3, 37]. We show here that encap-
sulated glioma cells attract microglia and induce
astrogliosis. While GDNF does not regulate astrogliosis, it
strongly mediates microglia attraction to HG-gliomas and
promotes tumor growth.
Materials and methods
Cell culture
GL261 murine glioma cells (National Cancer Institute,
Bethesda, USA) and murine fibroblast cell (SCRC-1008,
ATCC) were cultured in DMEM with 200 mM glutamine,
50 units/ml penicillin, 50 lg/ml streptomycin, and 10 %
FBS (all from Invitrogen, Darmstadt, Germany). The
human glioblastoma cells (Tumor 1 and 2) were derived
from human tumor resections and cultured in RPMI 1640
with 20 lg/ml EGF, FGF (Cell Systems, Troisdorf, Ger-
many) and supplements. Microglia was prepared from
neonatal C57BL/6 mice as previously described [25]. All
cells were maintained in a 37 �C incubator with a 5 % CO2
humidified atmosphere.
Cell encapsulation
SCRC-1008, GL261, and human glioma cells (Tumor 1)
were infused into HF (Minntech, Minneapolis, USA) at a
density of 2 9 106 cells/ml. HFs were sealed at the ends in
5-cm segments. 1 day before measurements or transplan-
tation, the 5-cm fibers were subsequently cut into 5 mm
length and sealed at both ends. Those HFs were further
sectioned in a cryostat with 10 lm thickness. Hematoxylin
and Eosin (H&E) staining’s of fiber sections were
performed.
Cell proliferation
Cell proliferation and viability were monitored using the
Alamar blue assay following manufacturer’s recommended
procedures (Invitrogen). The SCRC-1008, GL261, and
human glioma cells (Tumor 1) were seeded in 96 well
plates with 5,000 cells/100 ll density. At day 1, 3, 5, and 7
of culture, 10 ll of Alamar blue solution was added to each
well to make a 1:10 dilution and incubated for 4 h at 37 �C
in a humidified atmosphere of 5 % CO2 and 95 % air.
Before transplanting the fibers into the mouse brains, HF
with SCRC-1008, GL261, and human glioma cells (Tumor
1) were cut into 5 mm of length and sealed at both ends.
The relative cell numbers in the fiber was also measured
and only fibers with the equal amount of cells were used in
a given experiment. Wells containing empty fibers were
used as control. Absorbance of Alamar blue was measured
in a spectrometer (M200, Tecan, Crailsheim, Germany) at
570 and 600 nm.
HF inoculation
Female C57BL/6 (8–10 weeks) mice were handled
according to governmental (LAGeSo) and internal (MDC)
rules and regulations. Briefly, after anesthetized, the mouse
head was placed onto a stereotactic frame (David Kopf
Instruments, Tujunga, USA). Through a midline incision, a
burr hole was made by drilling at 1 mm anterior to the
bregma and 1.5 mm both right and left side from the
midline. Canals were created by inserting a 26-gauge
Hamilton syringe 5 mm ventral from dura mater. Then the
HFs were inserted into both canals.
Acta Neuropathol
123
Author's personal copy

Immunofluorescent staining and image processing
Mouse brains were prepared as previously described [26].
Human glioblastoma biopsy specimens were obtained from
glioblastoma patients, according to governmental and
internal (Charite) rules and regulations. Cortical brain tissue
from epilepsy patients was performed according to the rules
laid down by the Ethical Committee (Charite, EA1/142/05),
and informed consent was obtained according to the Dec-
laration of Helsinki (BMJ, 1991; 302: 1194). Primary
antibodies were added overnight at a dilution of 1:750 for
Iba-1 (WAKO, Tokyo, Japan), 1:1,000 for GFAP (Sigma-
Aldrich, Munich, Germany), 1:100 for Vimentin (Abcam,
Cambridge, UK) and 1:100 for GDNF (Santa Cruz, Hei-
delberg, Germany) at 4 �C. Alexa 488, 594-conjugated goat
anti-rabbit IgG (1:200, Invitrogen) and Cy5-conjugated
donkey anti-rabbit IgG (1:200, Jackson Lab, Hamburg,
Germany) were subsequently applied. The nuclei were
counterstained with 40,6-diamidino-2-phenylindole (DAPI,
Sigma-Aldrich).
Images were taken using a confocal microscope (TCS
SP5, Leica, Wetzlar, Germany) with 209 or 409 oil
objectives. Iba-1 positive labeled cells were counted using
ImageJ software (NIH, Bethesda, USA). The intensity of
GFAP fluorescence was measured by ImageJ without
adjusting digital gain.
GDNF knockdown and over-expression
GDNF and scrambled non-targeted or GAPDH small inter-
fering RNA (siRNA) were transfected with Dharmafect 4
according to manufacturer’s instructions (Dharmacon,
Lafayette, USA). After 24 h, GDNF or non-targeted siRNA
treated GL261 cells were injected into HF and an aliquot was
used for quality control by RT-PCR and ELISA.
GDNF transient transfection of SCRC-1008 was per-
formed using the lipofectamine LTX (Invitrogen). Plasmid
cDNA encoding GDNF was kindly provided by Dr. Carlos F.
Ibanez at the Karolinska Institute, Sweden. PCDNA3 plasmid
DNA was used as negative control.
For stable GDNF knockdown, GDNF OmicsLinkTM short
hairpin RNA (shRNA) clone (GCCAGTGTTTATCTGAT
AC) and non-targeting scrambled shRNA (shNT) were
transfected into GL261 cells according to manufacturer’s
instructions (GeneCopoeia, Rockville, USA). Transfected
GL261 cells were selected by treating with 5 lg/ml puromy-
cin. After selection, GDNF secretion was measured by ELISA.
RT-PCR and ELISA
Sequences for PCR primer sets were as follow: for GDNF
forward 50-TATGGGATGTCGTGGCTGT-30, reverse 50-CGTCATCAAACTGGTCAGGA-30, GFRa-1 forward 50-TT
CCCACACACGTTTTACCAC-30, reverse 50-TTTGTGG
TTATGTGGCTGGAG-30, GFRa-2 forward 50-AAGGC
CAACAACTCCAAAGAG-30, reverse 50-CCAAGGTCAC
CATCAGGAGT-30, RET forward 50-CCTCCGTGACAG
CCGCAAGA-30, reverse 50-CGGAATCCGGCCCTTGCT
TT-30, and for actin forward 50-CCCTGAAGTACCCCAT
TGAA-30, reverse 50-GTGGACAGTGAGGCCAAGAT-30.Supernatant collected from SCRC-1008, GL261, mouse
primary microglia, astrocytes, NPCs and human glioma
cells (Tumor 1 and 2) were determined for GDNF by
ELISA kits according to the manufacturer’s protocols
(Promega, Madison, USA). Results are presented as pico-
grams of GDNF per 50,000 cells.
Chemotaxis assays
The agarose spot assay was described before in [36]. Two
spots containing GDNF and two containing PBS were
placed onto 35-mm glass dishes (MatTek, Ashland, USA).
After 3 and 7 h, microglial cells under the spot were
counted. For time-lapse microscopy, the dish was placed
into the BioStation IM (Nikon, Dusseldorf, Germany) and
the resulting stacks were analyzed by ImageJ.
The chemotaxis of microglia was also determined using
a transwell assay with an 8-lm pore size membrane (BD
Biosciences, Heidelberg, Germany). The upper chamber of
a transwell insert was filled with 2 9 105 microglia in
suspension, and the lower chamber (BD Biosciences) with
0, 500, or 1,000 ng/ml recombinant GDNF (Peprotech,
Hamburg, Germany) or GL261 conditioned medium. After
24 h, microglial migration was quantified by counting the
number of cells that migrated through the membrane using
an inverted bright-field microscope. Five fields were quanti-
fied for each well.
In vivo assessment of glioma growth by magnetic
resonance imaging (MRI)
A total number of 10-mice were imaged at day 14 after the
bilateral intracereberal inoculation of either control (shNT)
or shGDNF GL261 cells. MR imaging was performed on a
9.4 Tesla small animal MRI system (Biospec 94/20, Bruker
BioSpin, Ettlingen, Germany) equipped with CryoProbe
(400 MHz, Bruker BioSpin). T2-weighted images (RARE,
effective echo time = 60 ms; repetition time = 3,268 ms;
RARE factor = 12) were acquired with the same slice
geometry (FOV = 18 9 18 mm, matrix size = 350 9
350, slice thickness = 270 lm, in-plane spatial resolu-
tion = 51 lm, 21 coronal slices covering a brain region of
5.67 mm starting at the frontal end of the cerebral cortex
(approx. Bregma 3.56 to -2.11 mm) [9]. Tumor volume
was calculated in mipav (http://mipav.cit.nih.gov) by add-
ing up the voxel volumes.
Acta Neuropathol
123
Author's personal copy
Survival studies were performed as previously described
[11]. Two groups of wild-type C57BL/6 mice received
either shNT or shGDNF GL261 cells. Survival rate was
calculated by the Kaplan–Meier method (MedCalc, Mari-
akerke, Belgium) with log rank analysis.
Statistical analysis
All data represent the average of at least triplicate samples.
Error bars represent standard error of the mean. Data were
analyzed by Student’s t test and one way ANOVA in
Microsoft Office Excel 2007 and applied Bonferroni cor-
rection. The differences were considered statistically
significant at p \ 0.05.
Results
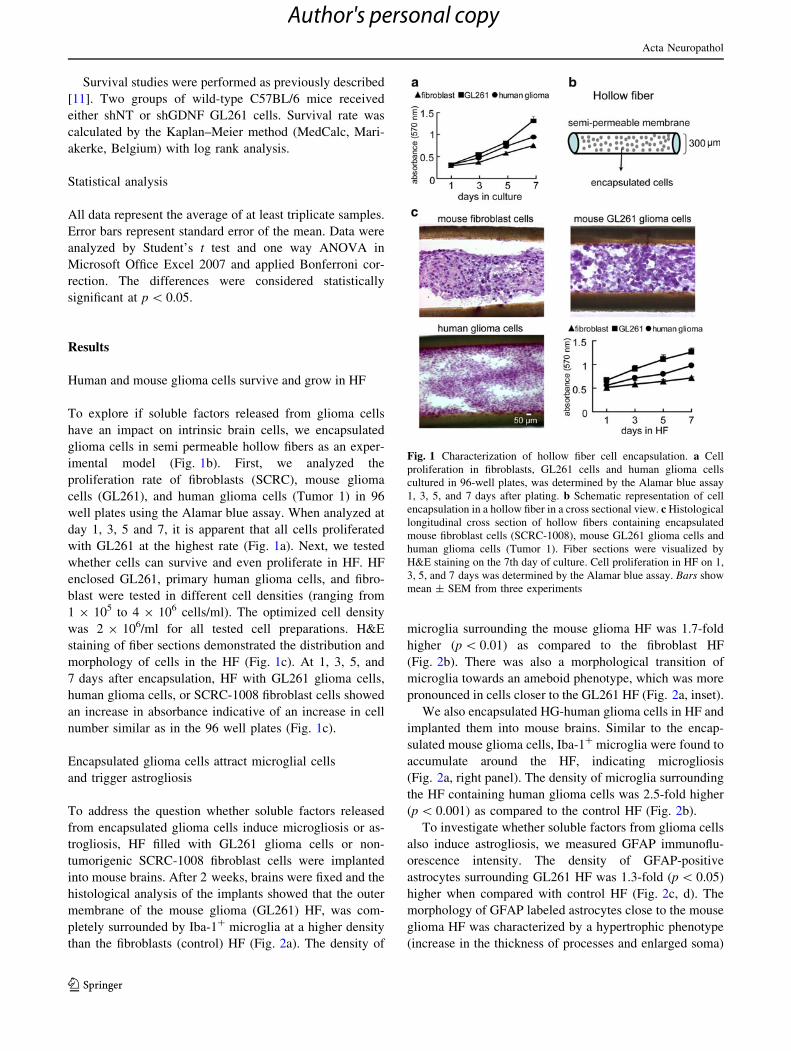
Human and mouse glioma cells survive and grow in HF
To explore if soluble factors released from glioma cells
have an impact on intrinsic brain cells, we encapsulated
glioma cells in semi permeable hollow fibers as an exper-
imental model (Fig. 1b). First, we analyzed the
proliferation rate of fibroblasts (SCRC), mouse glioma
cells (GL261), and human glioma cells (Tumor 1) in 96
well plates using the Alamar blue assay. When analyzed at
day 1, 3, 5 and 7, it is apparent that all cells proliferated
with GL261 at the highest rate (Fig. 1a). Next, we tested
whether cells can survive and even proliferate in HF. HF
enclosed GL261, primary human glioma cells, and fibro-
blast were tested in different cell densities (ranging from
1 9 105 to 4 9 106 cells/ml). The optimized cell density
was 2 9 106/ml for all tested cell preparations. H&E
staining of fiber sections demonstrated the distribution and
morphology of cells in the HF (Fig. 1c). At 1, 3, 5, and
7 days after encapsulation, HF with GL261 glioma cells,
human glioma cells, or SCRC-1008 fibroblast cells showed
an increase in absorbance indicative of an increase in cell
number similar as in the 96 well plates (Fig. 1c).
Encapsulated glioma cells attract microglial cells
and trigger astrogliosis
To address the question whether soluble factors released
from encapsulated glioma cells induce microgliosis or as-
trogliosis, HF filled with GL261 glioma cells or non-
tumorigenic SCRC-1008 fibroblast cells were implanted
into mouse brains. After 2 weeks, brains were fixed and the
histological analysis of the implants showed that the outer
membrane of the mouse glioma (GL261) HF, was com-
pletely surrounded by Iba-1? microglia at a higher density
than the fibroblasts (control) HF (Fig. 2a). The density of
microglia surrounding the mouse glioma HF was 1.7-fold
higher (p \ 0.01) as compared to the fibroblast HF
(Fig. 2b). There was also a morphological transition of
microglia towards an ameboid phenotype, which was more
pronounced in cells closer to the GL261 HF (Fig. 2a, inset).
We also encapsulated HG-human glioma cells in HF and
implanted them into mouse brains. Similar to the encap-
sulated mouse glioma cells, Iba-1? microglia were found to
accumulate around the HF, indicating microgliosis
(Fig. 2a, right panel). The density of microglia surrounding
the HF containing human glioma cells was 2.5-fold higher
(p \ 0.001) as compared to the control HF (Fig. 2b).
To investigate whether soluble factors from glioma cells
also induce astrogliosis, we measured GFAP immunoflu-
orescence intensity. The density of GFAP-positive
astrocytes surrounding GL261 HF was 1.3-fold (p \ 0.05)
higher when compared with control HF (Fig. 2c, d). The
morphology of GFAP labeled astrocytes close to the mouse
glioma HF was characterized by a hypertrophic phenotype
(increase in the thickness of processes and enlarged soma)
Fig. 1 Characterization of hollow fiber cell encapsulation. a Cell
proliferation in fibroblasts, GL261 cells and human glioma cells
cultured in 96-well plates, was determined by the Alamar blue assay
1, 3, 5, and 7 days after plating. b Schematic representation of cell
encapsulation in a hollow fiber in a cross sectional view. c Histological
longitudinal cross section of hollow fibers containing encapsulated
mouse fibroblast cells (SCRC-1008), mouse GL261 glioma cells and
human glioma cells (Tumor 1). Fiber sections were visualized by
H&E staining on the 7th day of culture. Cell proliferation in HF on 1,
3, 5, and 7 days was determined by the Alamar blue assay. Bars show
mean ± SEM from three experiments
Acta Neuropathol
123
Author's personal copy

that is typical of reactive astrocytes. Astrocytes in the
vicinity of control HF had a normal morphological
appearance. Human glioma HF in the mouse brains also
resulted in astrogliosis (Fig. 2c, d). These findings suggest
that glioma cells also influence astrocytes, although not as
potently as microglia, via soluble factors.
GDNF is highly expressed in glioma
To confirm that GDNF is produced and released from the
mouse and human glioma cell lines used in our study, we
further determined GDNF mRNA and protein levels. RT-
PCR shows that GL261 cells expressed much higher
mRNA levels than mouse astrocytes or microglia while we
detected no expression in neural precursor cells (NPCs) or
fibroblasts (Fig. 3a). We detected mRNA for GDNF
expression in human glioma cells from two different
patients (Tumor 1 and 2) (Fig. 3a). GDNF secretion was
then determined by ELISA and indicated that both HG-
human glioma cells (Tumor 1 and 2) and mouse glioma
cells (GL261) secret a higher amount of GDNF. GDNF
secretion levels were significantly lower in microglia and
astrocytes, and for SCRC-1008 fibroblasts even below the
level of detection (Tumor 1, 39.3 ± 4.9; Tumor 2,
47.9 ± 4.9; GL261, 42.3 ± 6.8; astrocyte, 13.7 ± 3.5;
microglia 7.6 ± 4.6 pg/50,000 cells/24 h) (Fig. 3b).
To determine if GDNF is expressed in the tumor envi-
ronment, we implanted mCherry expressing GL261 cells
into mouse brains. As shown in Fig. 3c, GDNF is highly
expressed by GL261 cells while we detected no expression
in non-tumor area. Human glioma biopsies also showed
high expression of GDNF while tissue from human cortex
(from an epilepsy surgery) did not express GDNF. These
results show a differential expression of GDNF between
glioma cells and the glioma-associated brain cells and
suggest GDNF as a candidate molecule mediating glioma-
glial cell interaction.
GDNF plays a key role in regulating microglia
attraction
To address whether GDNF mediates glioma-glial cell
interactions, we first analyzed GDNF receptor expression
on microglia. Figure 3a shows that microglia expressed
both GDNF receptors, GFRa-1 and GFRa-2, but not the
co-receptor RET. Astrocytes, NPCs, and fibroblasts
expressed GFRa-1 and GFRa-2, and glioma cells pre-
dominantly expressed GFRa-1 (Fig. 3a).
We next impaired GDNF expression in glioma cells by
an siRNA approach. GDNF gene expression in GL261 was
reduced by [80 % after 2 days (data not shown). GDNF
secretion was examined after siRNA treatment. Figure 4a
Fig. 2 Human and mouse glioma cells encapsulated in hollow fibers
induce microglia attraction and astrogliosis. a Fibroblast cells (SCRC-
1008), mouse glioma cells (GL261), or human glioma cells were
filled in HF and transplanted into the left or right hemisphere,
respectively. After 14 days, mice were killed and brain sections were
stained by immunofluorescence with Iba-1 (green). Cell nuclei were
counterstained with DAPI (blue). The yellow dashed line indicates
the border between HF and brain tissue. b Quantitative analysis of
Iba-1? cells surrounding GL261 HF (mean ± SEM, n = 9 mice;
**p \ 0.01) and human glioma HF (mean ± SEM, n = 13 mice;
***p \ 0.001) compared with fibroblast (control) HF. c Mouse brain
sections were stained with GFAP (red). d GFAP fluorescence
intensity measured from three random areas surrounding the GL261
HF (mean ± SEM, n = 8 mice; *p \ 0.05 compared with control)
and human glioma cells (mean ± SEM, n = 13 mice; *p \ 0.05
compared with control HF)
Acta Neuropathol
123
Author's personal copy

shows that the GDNF secretion was reduced by 66 %
(p \ 0.05) in GL261 cells (siGDNF) after 6 days of GNDF
siRNA treatment while non-targeted siRNA did not affect the
secretion of GDNF (Fig. 4a). On the next day of GDNF-
silencing, GL261 glioma cells were encapsulated into HF.
The depletion of GDNF by siRNA did not affect the glioma
cell proliferation in HF (supplemental Fig. 1). We then
implanted control (siNT) and siGDNF expressing GL261 cell
containing HF into mouse brains. 6 days after implantation,
we found that GDNF knockdown reduced microglia attrac-
tion (643 ± 47 cells/mm2) to 64 % (p \ 0.001) when
compared to control (siNT, 998 ± 48 cells/mm2) (Fig. 4b, c).
As a second approach, we used stable GDNF silencing by
shRNA approach to confirm the siRNA silencing results.
Either non-targeted shRNA (shNT) or GDNF shRNA
(shGDNF) treated GL261 cells were filled into HF. 6 days
after implantation, similar results show that GDNF knock-
down reduced microglia attraction (shGDNF, 734 ± 36
cells/mm2) to 65 % (p \ 0.01) when compared to control
(shNT, 1,124 ± 79 cells/mm2) (supplemental Fig. 2).
In addition, we have tested two lines of HG-human
glioma cells which secrete different levels of GDNF
(Fig. 3a, b). Tumor 1 and Tumor 2 HG-human glioma cells
were encapsulated in HF and implanted into mouse brains.
Similar to the encapsulated mouse glioma cells, Iba-1+
microglia were found to accumulate around both HF. The
density of microglia surrounding the Tumor 1 and Tumor 2
HF was not significantly different (data not shown).
To test whether over-expression of GDNF by non-gli-
oma cells could attract more microglial cells, SCRC-1008
fibroblast cells were transfected with either empty vector
(?vector) or GDNF vector (?GDNF). GDNF mRNA
expression in SCRC-1008 was increased by [80 % after
3 days of transfection (data not shown). GDNF secretion
was significantly increased in ?GDNF cells as compared to
control (?vector, 296.7 ± 16.8 pg/50,000 cells, p \ 0.01)
(Fig. 4d). Both ?vector and ?GDNF SCRC cells were
encapsulated into HF, implanted into mouse brains and
analyzed after 6 days. Iba-1 staining revealed that the mi-
croglial density increased by 1.7-fold (p \ 0.01) around
?GDNF HF, as compared to control (Fig. 4e, f). These
results showed that GDNF released from GL261 glioma
cells and GDNF overexpressing non-tumorigenic fibro-
blasts attract microglial cells and indicate the microglia
attraction is partly GDNF dependent.
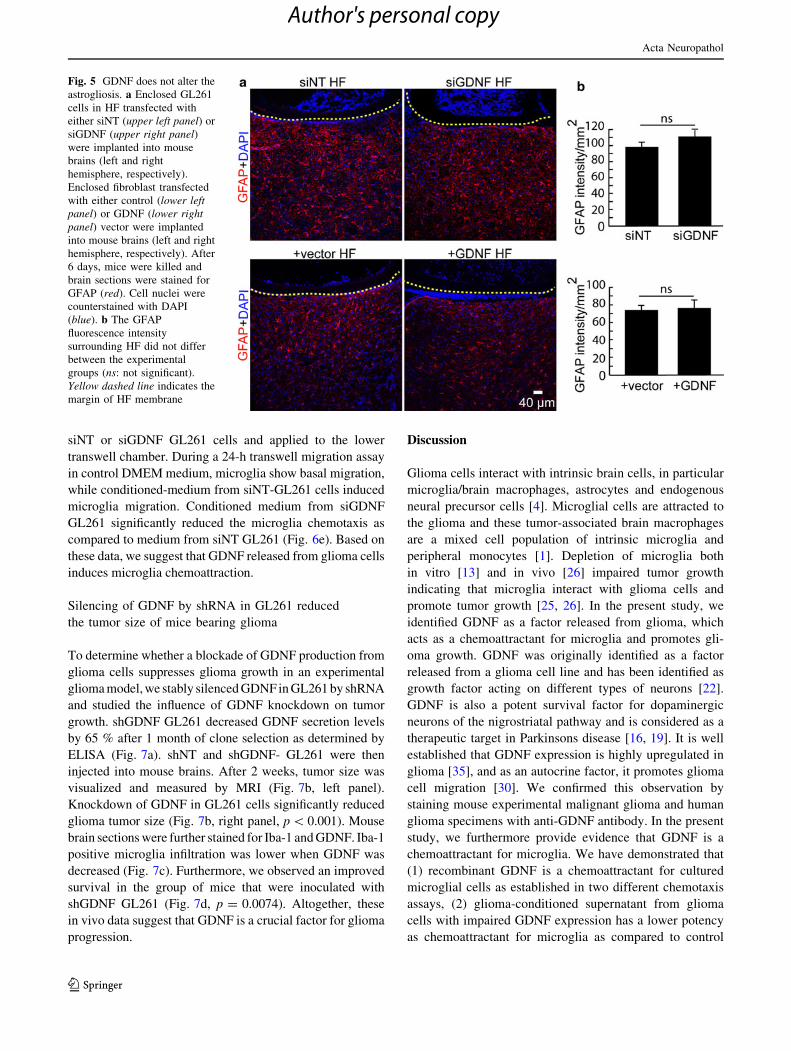
GDNF does not affect glioma-induced astrogliosis
We next evaluated the impact of GDNF on astrogliosis.
Control (siNT) versus GDNF knockdown (siGDNF)
Fig. 3 GDNF expression in glioma cells. a Semi-quantitative RT-
PCR of GDNF and its receptors GFRa-1, GFRa-2 and RET from
mouse and human cells: astrocytes, NPCs, microglia,fibroblast
(SCRC-1008), GL261 (mouse GL261 glioma cell line), human
glioma cells from two individual patients (Tumor 1 and 2). b-Actin
was used as an internal control for mRNA level. b Secreted GDNF
protein level was measured by ELISA. Bars represent the
mean ± SEM from three independent experiments (*p \ 0.05 com-
pared with fibroblast). c Tissue sections from a mouse inoculated with
mCherry GL261 glioma cells (red) and stained with GDNF (green).
Tissue sections from human cortex (from an epilepsy surgery) as a
control (left panel) and human glioma tissue from HG-glioma patient
(right panel) were stained with GDNF (red) and vimentin (green)
antibody and DAPI (blue)
Acta Neuropathol
123
Author's personal copy

GL261 glioma cells as well as SCRC-1008 fibroblasts
(?vector) versus GDNF overexpressing fibroblasts
(?GDNF) HF were implanted into mouse brains. There
was no significant difference in GFAP intensity and cell
morphology in cells surrounding both siNT and siGDNF
HF (Fig. 5a, b, upper panel) and in cells surrounding
?vector and ?GDNF HF (Fig. 5a, b, lower panel). This
result indicated that neither down-regulation of GDNF in
glioma cells nor over expression of GDNF in non-tumor-
igenic fibroblast cells influenced the glioma-induced
astrogliosis.
GDNF is a potent chemoattractant for microglia
Since we showed that GDNF only affected microgliosis but
not astrogliosis, we aimed to determine the effect of GDNF
on microglial chemotactic behavior. First we tested two
different concentrations (0, 500, and 1,000 ng/ml) of
GDNF on microglial chemotaxis by the transwell assay.
After 24 h incubation, mouse primary microglia migration
into the lower chamber triggered by GDNF increased dose
dependently (0 ng/ml: 35 ± 5 cells/counting field; 500 ng/
ml: 80 ± 5 cells/counting field, and 1,000 ng/ml:
142 ± 12 cells/counting field) (Fig. 6a).
As a second approach, we determined microglia che-
motaxis by analyzing migration into an agarose spot
containing either PBS (as control) or GDNF. The spots
were placed on glass-bottomed Petri dishes and microglial
cells in suspension were subsequently added (Fig. 6b).
Microglial invasion into the GDNF-containing agarose
spots was significantly higher as compared to the control
spots (at 3 h GDNF spot: 422 ± 17 cells/spot; PBS spot:
8 ± 1 cells/spot, p \ 0.001) (Fig. 6b, right panel). To
study the dynamics of microglial invasion, we analyzed
microglial migration by time-lapse recording over 2 h.
Microglia migrated faster and over longer distances into
the GDNF spots as compared to control (p \ 0.001,
Fig. 6c, d). Both velocity and migrated distances increased
in a dose-dependent fashion (p \ 0.001, Fig. 6d).
To verify whether GDNF secreted from GL261 cells
influences microglia migration, the transwell assay was
applied. Conditioned-medium was collected from either
Fig. 4 GDNF-mediated microglia attraction. a GDNF secretion from
non-transfected (GL261), non-targeted siRNA treated GL261 cells
(siNT) and GDNF siRNA treated cells (siGDNF) after 6 days of
transfection was analyzed by ELISA. b After transfection with either
siNT (left panel) or siGDNF (right panel), GL261 cells enclosed in
HF were implanted into the mouse brains (left and right hemisphere,
respectively). After 6 days of implantation, mice were killed and
brain sections were stained with Iba-1 (red) antibody. Cell nuclei
were counterstained with DAPI (blue). Yellow dashed line indicates
the border between HF and brain tissue. c Density of microglia
surrounding HF with siGDNF transfected GL261 cells is shown
(mean ± SEM, n = 11 mice; ***p \ 0.001) compared with control
siNT GL261 cells. d SCRC-1008 fibroblast cells were transfected
with control vector or GDNF. GDNF secretion was detected by
ELISA and shows increased secretion. e After transfection with either
control (left panel) or GDNF vector (right panel), enclosed fibroblast
cells in HF were implanted into mouse brains (left and right
hemisphere, respectively). After 6 days of transplantation, mice were
killed and brain sections were stained with Iba-1 antibody (red). Cell
nuclei were counterstained with DAPI (blue). Yellow dashed lineindicates the border between HF and brain tissue. f Density of
microglia surrounding HF with control (vector) and GDNF over-
expressing fibroblasts is shown (mean ± SEM, n = 5 mice;
***p \ 0.001)
Acta Neuropathol
123
Author's personal copy
siNT or siGDNF GL261 cells and applied to the lower
transwell chamber. During a 24-h transwell migration assay
in control DMEM medium, microglia show basal migration,
while conditioned-medium from siNT-GL261 cells induced
microglia migration. Conditioned medium from siGDNF
GL261 significantly reduced the microglia chemotaxis as
compared to medium from siNT GL261 (Fig. 6e). Based on
these data, we suggest that GDNF released from glioma cells
induces microglia chemoattraction.
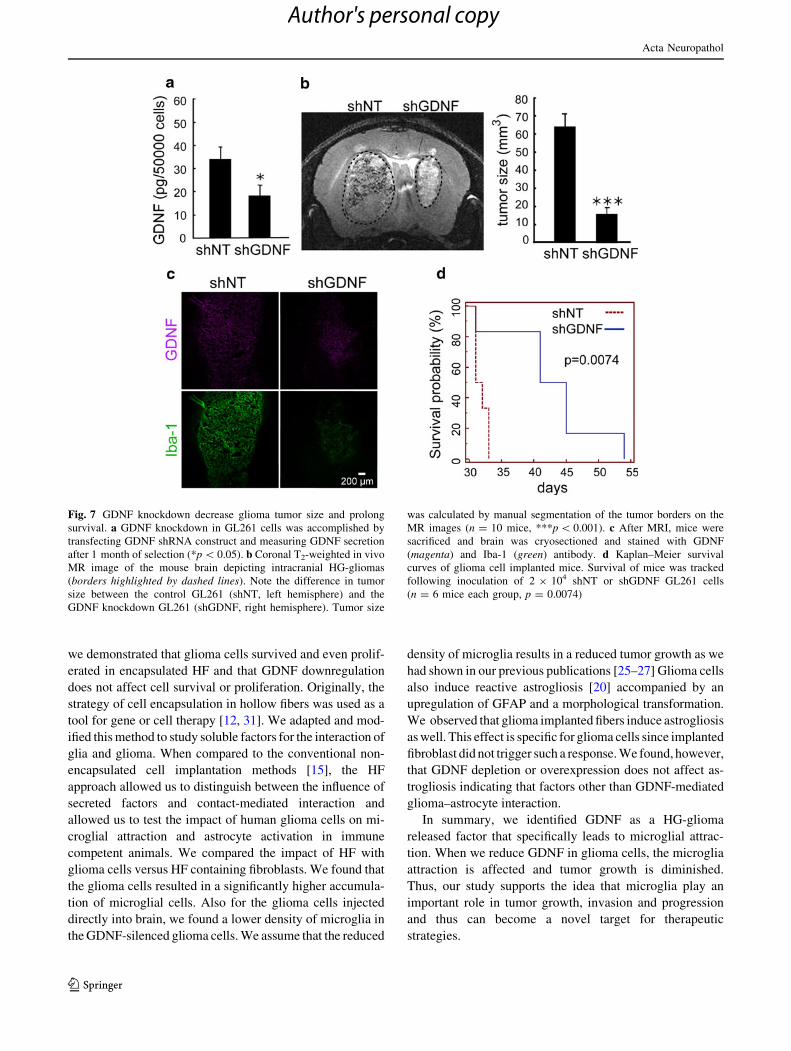
Silencing of GDNF by shRNA in GL261 reduced
the tumor size of mice bearing glioma
To determine whether a blockade of GDNF production from
glioma cells suppresses glioma growth in an experimental
glioma model, we stably silenced GDNF in GL261 by shRNA
and studied the influence of GDNF knockdown on tumor
growth. shGDNF GL261 decreased GDNF secretion levels
by 65 % after 1 month of clone selection as determined by
ELISA (Fig. 7a). shNT and shGDNF- GL261 were then
injected into mouse brains. After 2 weeks, tumor size was
visualized and measured by MRI (Fig. 7b, left panel).
Knockdown of GDNF in GL261 cells significantly reduced
glioma tumor size (Fig. 7b, right panel, p \ 0.001). Mouse
brain sections were further stained for Iba-1 and GDNF. Iba-1
positive microglia infiltration was lower when GDNF was
decreased (Fig. 7c). Furthermore, we observed an improved
survival in the group of mice that were inoculated with
shGDNF GL261 (Fig. 7d, p = 0.0074). Altogether, these
in vivo data suggest that GDNF is a crucial factor for glioma
progression.
Discussion
Glioma cells interact with intrinsic brain cells, in particular
microglia/brain macrophages, astrocytes and endogenous
neural precursor cells [4]. Microglial cells are attracted to
the glioma and these tumor-associated brain macrophages
are a mixed cell population of intrinsic microglia and
peripheral monocytes [1]. Depletion of microglia both
in vitro [13] and in vivo [26] impaired tumor growth
indicating that microglia interact with glioma cells and
promote tumor growth [25, 26]. In the present study, we
identified GDNF as a factor released from glioma, which
acts as a chemoattractant for microglia and promotes gli-
oma growth. GDNF was originally identified as a factor
released from a glioma cell line and has been identified as
growth factor acting on different types of neurons [22].
GDNF is also a potent survival factor for dopaminergic
neurons of the nigrostriatal pathway and is considered as a
therapeutic target in Parkinsons disease [16, 19]. It is well
established that GDNF expression is highly upregulated in
glioma [35], and as an autocrine factor, it promotes glioma
cell migration [30]. We confirmed this observation by
staining mouse experimental malignant glioma and human
glioma specimens with anti-GDNF antibody. In the present
study, we furthermore provide evidence that GDNF is a
chemoattractant for microglia. We have demonstrated that
(1) recombinant GDNF is a chemoattractant for cultured
microglial cells as established in two different chemotaxis
assays, (2) glioma-conditioned supernatant from glioma
cells with impaired GDNF expression has a lower potency
as chemoattractant for microglia as compared to control
Fig. 5 GDNF does not alter the
astrogliosis. a Enclosed GL261
cells in HF transfected with
either siNT (upper left panel) or
siGDNF (upper right panel)were implanted into mouse
brains (left and right
hemisphere, respectively).
Enclosed fibroblast transfected
with either control (lower leftpanel) or GDNF (lower rightpanel) vector were implanted
into mouse brains (left and right
hemisphere, respectively). After
6 days, mice were killed and
brain sections were stained for
GFAP (red). Cell nuclei were
counterstained with DAPI
(blue). b The GFAP
fluorescence intensity
surrounding HF did not differ
between the experimental
groups (ns: not significant).
Yellow dashed line indicates the
margin of HF membrane
Acta Neuropathol
123
Author's personal copy

conditioned medium, (3) glioma cells with downregulated
GNDF expression encapsulated into HF and implanted into
mouse brains attract less microglia as compared to control
glioma cells, (4) overexpression of GDNF in a fibroblast
cell line attracted more microglia as HF with control
fibroblasts in our in vivo model, (5) orthotopic implantation
of GDNF knockdown glioma cells into the mouse brain
generates smaller tumors as compared to controls. Impor-
tantly, reduced GDNF levels in gliomas improved survival
in our immunecompetent, orthotopic implantation model.
For our survival data, we also compared unilateral injec-
tions of glioma cells either control or silenced for GDNF
expression. In addition, the unilateral injections resulted in
a significant difference.
Our data also indicate that GDNF is a major but not the
sole chemoattractant for microglia/macrophages to HG-
gliomas. Other factors like MCP-1 (CCL-2), CX3CL1 or
SDF-1 (CXCL-12) may also contribute to microglial
attraction to primary brain tumors. However, the roles of
these factors on the recruitment of microglia towards gli-
oma have only been described in vitro [14, 24, 38].
As a tool to distinguish between cell contact and soluble
factor, we employed the HF approach where glioma cells
were encapsulated and inserted into the brain. In this study,
Fig. 6 GDNF is involved in glioma cell incuced microglia chemo-
taxis. a Microglia was seeded in a transwell plate in the presence of 0,
500, or 1,000 ng/ml of recombinant GDNF for 24 h. Migrated cells
were counted after the membranes were stained with H&E. Barsrepresent the mean ± SEM from three experiments, ***p \ 0.001
and **p \ 0.01 when compared with control. b Microglia was plated
on 35-mm cover slips containing both PBS and GDNF spots. 3 h after
plating, microglia migrated toward GDNF spots (dashed line depicts
border of agarose spot) and cells migrated under/into the spots were
counted (right panel). Bars represent the mean ± SEM from 12
experiments, ***p \ 0.001. c Microglia migration direction was
observed by time-lapse imaging and analyzed by ImageJ software.
Each line represents one cell (25 cells for PBS spot and 27 cells for
GDNF spot). d microglia migration velocity and distance was
calculated. Bars represent the mean ± SEM from 48 experiments,
***p \ 0.001. e Microglia was seeded in a transwell plate in the
presence of control medium (DMEM with 10 % FBS) or glioma-
conditioned medium from non-targeted siRNA transfected GL261
cells (siNT), or glioma-conditioned medium from GDNF-targeted
siRNA transfected GL261 cells (siGDNF). The transwell plate was
then incubated for 24 h. The migrating microglia in the opposite side
of the filter were fixed and stained. Migrated microglia was counted
from at least five fields. Bars represent the mean ± SEM from three
experiments (*p \ 0.05 and ***p \ 0.001)
Acta Neuropathol
123
Author's personal copy
we demonstrated that glioma cells survived and even prolif-
erated in encapsulated HF and that GDNF downregulation
does not affect cell survival or proliferation. Originally, the
strategy of cell encapsulation in hollow fibers was used as a
tool for gene or cell therapy [12, 31]. We adapted and mod-
ified this method to study soluble factors for the interaction of
glia and glioma. When compared to the conventional non-
encapsulated cell implantation methods [15], the HF
approach allowed us to distinguish between the influence of
secreted factors and contact-mediated interaction and
allowed us to test the impact of human glioma cells on mi-
croglial attraction and astrocyte activation in immune
competent animals. We compared the impact of HF with
glioma cells versus HF containing fibroblasts. We found that
the glioma cells resulted in a significantly higher accumula-
tion of microglial cells. Also for the glioma cells injected
directly into brain, we found a lower density of microglia in
the GDNF-silenced glioma cells. We assume that the reduced
density of microglia results in a reduced tumor growth as we
had shown in our previous publications [25–27] Glioma cells
also induce reactive astrogliosis [20] accompanied by an
upregulation of GFAP and a morphological transformation.
We observed that glioma implanted fibers induce astrogliosis
as well. This effect is specific for glioma cells since implanted
fibroblast did not trigger such a response. We found, however,
that GDNF depletion or overexpression does not affect as-
trogliosis indicating that factors other than GDNF-mediated
glioma–astrocyte interaction.
In summary, we identified GDNF as a HG-glioma
released factor that specifically leads to microglial attrac-
tion. When we reduce GDNF in glioma cells, the microglia
attraction is affected and tumor growth is diminished.
Thus, our study supports the idea that microglia play an
important role in tumor growth, invasion and progression
and thus can become a novel target for therapeutic
strategies.
Fig. 7 GDNF knockdown decrease glioma tumor size and prolong
survival. a GDNF knockdown in GL261 cells was accomplished by
transfecting GDNF shRNA construct and measuring GDNF secretion
after 1 month of selection (*p \ 0.05). b Coronal T2-weighted in vivo
MR image of the mouse brain depicting intracranial HG-gliomas
(borders highlighted by dashed lines). Note the difference in tumor
size between the control GL261 (shNT, left hemisphere) and the
GDNF knockdown GL261 (shGDNF, right hemisphere). Tumor size
was calculated by manual segmentation of the tumor borders on the
MR images (n = 10 mice, ***p \ 0.001). c After MRI, mice were
sacrificed and brain was cryosectioned and stained with GDNF
(magenta) and Iba-1 (green) antibody. d Kaplan–Meier survival
curves of glioma cell implanted mice. Survival of mice was tracked
following inoculation of 2 9 104 shNT or shGDNF GL261 cells
(n = 6 mice each group, p = 0.0074)
Acta Neuropathol
123
Author's personal copy

Acknowledgments This work was supported by the graduate school
of NeuroCure at the Charite, Berlin, (stipend to M. C. Ku) and by
Deutsche Forschungsgemeinschaft (TR 43). We are thankful to Prof.
Carlos Ibanez for providing GDNF cDNA construct for transfection
and Prof. Jochen Meier for providing human brain tissue. We thank
Dr. Zoltan Cseresnyes and Dr. Anje Sporbert for technical assistance
with confocal microscopy. We appreciate the support of Babette
Dieringer for MR imaging and Maria Pannell for manuscript proof
reading.
Conflict of interest The authors declare that they have no conflict
of interest.
References
1. Badie B, Schartner J (2001) Role of microglia in glioma biology.
Microsc Res Tech 54(2):106–113. doi:10.1002/jemt.112510.
1002/jemt.1125
2. Badie B, Schartner J, Klaver J, Vorpahl J (1999) In vitro mod-
ulation of microglia motility by glioma cells is mediated by
hepatocyte growth factor/scatter factor. Neurosurgery 44(5):
1077–1082 (discussion 1082–1073)
3. Broadhead KW, Biran R, Tresco PA (2002) Hollow fiber mem-
brane diffusive permeability regulates encapsulated cell line
biomass, proliferation, and small molecule release. Biomaterials
23(24):4689–4699. doi:10.1016/S0142-9612(02)00212-0
4. Charles NA, Holland EC, Gilbertson R, Glass R, Kettenmann H
(2011) The brain tumor microenvironment. Glia 59(8):1169–
1180. doi:10.1002/glia.21136
5. Cornejo M, Nambi D, Walheim C, Somerville M, Walker J, Kim
L, Ollison L, Diamante G, Vyawahare S, de Bellard ME (2010)
Effect of NRG1, GDNF, EGF and NGF in the migration of a
Schwann cell precursor line. Neurochem Res 35(10):1643–1651.
doi:10.1007/s11064-010-0225-0
6. Dhandapani KM, Khan MM, Wade FM, Wakade C, Mahesh VB,
Brann DW (2007) Induction of transforming growth factor-beta1
by basic fibroblast growth factor in rat C6 glioma cells and
astrocytes is mediated by MEK/ERK signaling and AP-1 acti-
vation. J Neurosci Res 85(5):1033–1045. doi:10.1002/jnr.21182
7. Dudanova I, Gatto G, Klein R (2010) GDNF acts as a chemo-
attractant to support ephrinA-induced repulsion of limb motor
axons. Curr Biol 20(23):2150–2156. doi:10.1016/j.cub.2010.11.
021
8. Edwards LA, Woolard K, Son MJ, Li A, Lee J, Ene C, Mantey
SA, Maric D, Song H, Belova G, Jensen RT, Zhang W, Fine HA
(2011) Effect of brain- and tumor-derived connective tissue
growth factor on glioma invasion. J Natl Cancer Inst
103(15):1162–1178. doi:djr22410.1093/jnci/djr224
9. Franklin K, Paxinos G (2007) The mouse brain in stereotaxic
coordinates, 3rd edn. Academic Press, San Diego
10. Geranmayeh F, Scheithauer BW, Spitzer C, Meyer FB, Svensson-
Engwall AC, Graeber MB (2007) Microglia in gemistocytic
astrocytomas. Neurosurgery 60(1):159–166 (discussion 166). doi:
10.1227/01.NEU.0000249192.30786.67
11. Glass R, Synowitz M, Kronenberg G, Walzlein JH, Markovic DS,
Wang LP, Gast D, Kiwit J, Kempermann G, Kettenmann H (2005)
Glioblastoma-induced attraction of endogenous neural precursor
cells is associated with improved survival. J Neurosci 25(10):2637–
2646. doi:25/10/263710.1523/JNEUROSCI.5118-04.2005
12. Han Q, Sun W, Lin H, Zhao W, Gao Y, Zhao Y, Chen B, Xiao Z,
Hu W, Li Y, Yang B, Dai J (2009) Linear ordered collagen
scaffolds loaded with collagen-binding brain-derived neurotro-
phic factor improve the recovery of spinal cord injury in rats.
Tissue Eng Part A 15(10):2927–2935. doi:10.1089/ten.TEA.2008.
0506
13. Hussain SF, Yang D, Suki D, Aldape K, Grimm E, Heimberger
AB (2006) The role of human glioma-infiltrating microglia/
macrophages in mediating antitumor immune responses. Neuro
Oncol 8(3):261–279. doi:10.1215/15228517-2006-008
14. Kenig S, Alonso MB, Mueller MM, Lah TT (2009) Glioblastoma
and endothelial cells cross-talk, mediated by SDF-1, enhances
tumour invasion and endothelial proliferation by increasing
expression of cathepsins B, S, and MMP-9. Cancer Lett 289(1):53–
61. doi:10.1016/j.canlet.2009.07.014
15. Kim YT, Hitchcock R, Broadhead KW, Messina DJ, Tresco PA
(2005) A cell encapsulation device for studying soluble factor
release from cells transplanted in the rat brain. J Control Release
102(1):101–111. doi:10.1016/j.jconrel.2004.10.003
16. Kirik D, Georgievska B, Bjorklund A (2004) Localized striatal
delivery of GDNF as a treatment for Parkinson disease. Nat
Neurosci 7(2):105–110. doi:10.1038/nn1175nn1175
17. Koelsch A, Feng Y, Fink DJ, Mata M (2010) Transgene-mediated
GDNF expression enhances synaptic connectivity and GABA
transmission to improve functional outcome after spinal cord
contusion. J Neurochem 113(1):143–152. doi:10.1111/j.1471-
4159.2010.06593.x
18. Komohara Y, Ohnishi K, Kuratsu J, Takeya M (2008) Possible
involvement of the M2 anti-inflammatory macrophage phenotype
in growth of human gliomas. J Pathol 216(1):15–24. doi:10.1002/
path.2370
19. Lang AE, Gill S, Patel NK, Lozano A, Nutt JG, Penn R, Brooks
DJ, Hotton G, Moro E, Heywood P, Brodsky MA, Burchiel K,
Kelly P, Dalvi A, Scott B, Stacy M, Turner D, Wooten VG, Elias
WJ, Laws ER, Dhawan V, Stoessl AJ, Matcham J, Coffey RJ,
Traub M (2006) Randomized controlled trial of intraputamenal
glial cell line-derived neurotrophic factor infusion in Parkinson
disease. Ann Neurol 59(3):459–466. doi:10.1002/ana.20737
20. Le DM, Besson A, Fogg DK, Choi KS, Waisman DM, Goodyer
CG, Rewcastle B, Yong VW (2003) Exploitation of astrocytes by
glioma cells to facilitate invasiveness: a mechanism involving
matrix metalloproteinase-2 and the urokinase-type plasminogen
activator-plasmin cascade. J Neurosci 23(10):4034–4043
21. Leung SY, Wong MP, Chung LP, Chan AS, Yuen ST (1997)
Monocyte chemoattractant protein-1 expression and macrophage
infiltration in gliomas. Acta Neuropathol 93(5):518–527
22. Lin LF, Doherty DH, Lile JD, Bektesh S, Collins F (1993)
GDNF: a glial cell line-derived neurotrophic factor for midbrain
dopaminergic neurons. Science 260(5111):1130–1132
23. Lu DY, Leung YM, Cheung CW, Chen YR, Wong KL (2010) Glial
cell line-derived neurotrophic factor induces cell migration and
matrix metalloproteinase-13 expression in glioma cells. Biochem
Pharmacol 80(8):1201–1209. doi:10.1016/j.bcp.2010.06.046
24. Magge SN, Malik SZ, Royo NC, Chen HI, Yu L, Snyder EY,
O’Rourke DM, Watson DJ (2009) Role of monocyte chemoattrac-
tant protein-1 (MCP-1/CCL2) in migration of neural progenitor
cells toward glial tumors. J Neurosci Res 87(7):1547–1555. doi:
10.1002/jnr.21983
25. Markovic DS, Glass R, Synowitz M, Rooijen N, Kettenmann H
(2005) Microglia stimulate the invasiveness of glioma cells by
increasing the activity of metalloprotease-2. J Neuropathol Exp
Neurol 64(9):754–762. doi:00005072-200509000-00002
26. Markovic DS, Vinnakota K, Chirasani S, Synowitz M, Raguet H,
Stock K, Sliwa M, Lehmann S, Kalin R, van Rooijen N, HolmbeckK, Heppner FL, Kiwit J, Matyash V, Lehnardt S, Kaminska B, Glass
R, Kettenmann H (2009) Gliomas induce and exploit microglial
MT1-MMP expression for tumor expansion. Proc Natl Acad Sci
USA 106(30):12530–12535. doi:10.1073/pnas.0804273106
27. Markovic DS, Vinnakota K, van Rooijen N, Kiwit J, Synowitz M,
Glass R, Kettenmann H (2011) Minocycline reduces glioma
Acta Neuropathol
123
Author's personal copy

expansion and invasion by attenuating microglial MT1-MMP
expression. Brain Behav Immun 25(4):624–628. doi:10.1016/j.bbi.
2011.01.015
28. Okada M, Saio M, Kito Y, Ohe N, Yano H, Yoshimura S, Iwama
T, Takami T (2009) Tumor-associated macrophage/microglia
infiltration in human gliomas is correlated with MCP-3, but not
MCP-1. Int J Oncol 34(6):1621–1627
29. Platten M, Kretz A, Naumann U, Aulwurm S, Egashira K, Is-
enmann S, Weller M (2003) Monocyte chemoattractant protein-1
increases microglial infiltration and aggressiveness of gliomas.
Ann Neurol 54(3):388–392. doi:10.1002/ana.10679
30. Song H, Moon A (2006) Glial cell-derived neurotrophic factor
(GDNF) promotes low-grade Hs683 glioma cell migration
through JNK, ERK-1/2 and p38 MAPK signaling pathways.
Neurosci Res 56(1):29–38. doi:10.1016/j.neures.2006.04.019
31. Visted T, Bjerkvig R, Enger PO (2001) Cell encapsulation
technology as a therapeutic strategy for CNS malignancies.
Neuro Oncol 3(3):201–210
32. Voisin P, Bouchaud V, Merle M, Diolez P, Duffy L, Flint K,
Franconi JM, Bouzier-Sore AK (2010) Microglia in close vicinity of
glioma cells: correlation between phenotype and metabolic altera-
tions. Front Neuroenergetics 2:131. doi:10.3389/fnene.2010.00131
33. Wan G, Too HP (2010) A specific isoform of glial cell line-
derived neurotrophic factor family receptor alpha 1 regulates
RhoA expression and glioma cell migration. J Neurochem
115(3):759–770. doi:10.1111/j.1471-4159.2010.06975.x
34. Watters JJ, Schartner JM, Badie B (2005) Microglia function in
brain tumors. J Neurosci Res 81(3):447–455. doi:10.1002/jnr.20485
35. Wiesenhofer B, Stockhammer G, Kostron H, Maier H, Hinterh-
uber H, Humpel C (2000) Glial cell line-derived neurotrophic
factor (GDNF) and its receptor (GFR-alpha 1) are strongly
expressed in human gliomas. Acta Neuropathol 99(2):131–137
36. Wiggins H, Rappoport J (2010) An agarose spot assay for chemotactic
invasion. Biotechniques 48(2):121–124. doi:10.2144/000113353
37. Zhang GJ, Chen TB, Hargreaves R, Sur C, Williams DL Jr (2008)
Bioluminescence imaging of hollow fibers in living animals: its
application in monitoring molecular pathways. Nat Protoc
3(5):891–899. doi:10.1038/nprot.2008.52
38. Zhao D, Najbauer J, Garcia E, Metz MZ, Gutova M, Glackin CA,
Kim SU, Aboody KS (2008) Neural stem cell tropism to glioma:
critical role of tumor hypoxia. Mol Cancer Res 6(12):1819–1829.
doi:10.1158/1541-7786.MCR-08-0146
Acta Neuropathol
123
Author's personal copy




















