
P2X4 receptors control the fate and survival of activated microglia
14
RESEARCH ARTICLE P2X4 Receptors Control the Fate and Survival of Activated Microglia Nuria V azquez-Villoldo, 1 Mar ıa Domercq, 1 Abraham Mart ın, 2 Jordi Llop, 3 Vanessa G omez-Vallejo, 3 and Carlos Matute 1 Microglia, the resident immune cells of the central nervous system, responds to brain disarrangements by becoming activated to contend with brain damage. Here we show that the expression of P2X4 receptors is upregulated in inflammatory foci and in activated microglia in the spinal cord of rats with experimental autoimmune encephalomyelitis (EAE) as well as in the optic nerve of multiple sclerosis patients. To study the role of P2X4 receptors in microgliosis, we activated microglia with LPS in vitro and in vivo. We observed that P2X4 receptor activity in vitro was increased in LPS-activated microglia as assessed by patch-clamp recordings. In addition, P2X4 receptor blockade significantly reduced microglial membrane ruffling, TNFa secre- tion and morphological changes, as well as LPS-induced microglial cell death. Accordingly, neuroinflammation provoked by LPS injection in vivo induced a rapid microglial loss in the spinal cord that was totally prevented or potentiated by P2X4 receptor blockade or facilitation, respectively. Within the brain, microglia in the hippocampal dentate gyrus showed particular vulnerability to LPS-induced neuroinflammation. Thus, microglia processes in this region retracted as early as 2 h after injec- tion of LPS and died around 24 h later, two features which were prevented by blocking P2X4 receptors. Together, these data suggest that P2X4 receptors contribute to controlling the fate of activated microglia and its survival. GLIA 2014;62:171–184 Key words: microglia, P2X4 purinergic receptor, EAE and multiple sclerosis, cell death Introduction M icroglial cells are the main effectors of the innate immune response after CNS injury (Kreutzberg, 1996). The precursors of adult microglia are yolk sac progenitors, which enter the embryo around E8 and colonize the CNS at E10 (Ginhoux et al., 2010). Microglia can be found postna- tally in all regions of the CNS, in the parenchyma and in perivascular spaces in a nonoverlapping territorial fashion, and comprise a large proportion of the total cellular makeup of the CNS, estimated to be as high as 12% of all cells. Per- haps owing to this origin, microglia share many commonal- ties with resident tissue macrophages in peripheral organ systems (Ransohoff and Perry, 2009). Like their peripherals macrophage counterparts, microglia displays a remarkable range in both morphology and activity, depending in part on the state of the surrounding tissue (Ransohoff and Perry, 2009). In their resting state, microglia continuously monitor the tissue for injury or pathological changes by extending and retracting highly motile processes on a time scale of minutes (Davalos et al., 2005; Nimmerjahn et al., 2005; reviewed in Kettenmann et al., 2013). Exogenous factors (infectious agents) as well as endogenous factors or neurotransmitters released by damaged cells or tissues induce progressive changes in microglia including altered expression of cell sur- face markers and inflammation related genes, process retrac- tion and acquisition of an ameboid morphology, migration of cell bodies, proliferation and increased phagocytic rate (Ket- tenmann et al., 2011). The rapidly diffusible neurostransmitter ATP is one of the main regulators of microglial functions. Low ATP concen- trations almost exclusively activate chemotaxis, through P2Y12 receptor activation, in order to recruit cells at the site View this article online at wileyonlinelibrary.com. DOI: 10.1002/glia.22596 Published online November 19, 2013 in Wiley Online Library (wileyonlinelibrary.com). Received May 23, 2013, Accepted for publication Oct 16, 2013. Address correspondence to Carlos Matute, Departamento de Neurociencias, Universidad del Pa ıs Vasco, E-48940 Leioa, Spain. E-mail: [email protected] or Maria Domercq, Departamento de Neurociencias, Universidad del Pa ıs Vasco, E-48940 Leioa, Spain. E-mail: [email protected] From the 1 Departamento de Neurociencias, Universidad del Pa ıs Vasco-UPV/EHU, Barrio Sarriena s/n, 48940, Leioa, Spain, Achucarro Basque Center for Neuro- science-UPV/EHU, 48170, Zamudio, Spain, Instituto de Salud Carlos III, Centro de Investigaci on Biom edica en Red de Enfermedades Neurodegenerativas (CIBERNED), 48940, Leioa, Spain; 2 Molecular Imaging Unit, CIC biomaGUNE, Paseo Miramon 182, 20009, San Sebasti an, Spain; 3 Radiochemistry Department, Molecular Imaging Unit, CIC biomaGUNE, Paseo Miramon 182, 20009, San Sebasti an, Spain. Additional Supporting Information may be found in the online version of this article. V C 2013 Wiley Periodicals, Inc. 171
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of P2X4 receptors control the fate and survival of activated microglia

RESEARCH ARTICLE
P2X4 Receptors Control the Fate andSurvival of Activated Microglia
Nuria V�azquez-Villoldo,1 Mar�ıa Domercq,1 Abraham Mart�ın,2 Jordi Llop,3
Vanessa G�omez-Vallejo,3 and Carlos Matute 1
Microglia, the resident immune cells of the central nervous system, responds to brain disarrangements by becoming activatedto contend with brain damage. Here we show that the expression of P2X4 receptors is upregulated in inflammatory foci andin activated microglia in the spinal cord of rats with experimental autoimmune encephalomyelitis (EAE) as well as in the opticnerve of multiple sclerosis patients. To study the role of P2X4 receptors in microgliosis, we activated microglia with LPS invitro and in vivo. We observed that P2X4 receptor activity in vitro was increased in LPS-activated microglia as assessed bypatch-clamp recordings. In addition, P2X4 receptor blockade significantly reduced microglial membrane ruffling, TNFa secre-tion and morphological changes, as well as LPS-induced microglial cell death. Accordingly, neuroinflammation provoked byLPS injection in vivo induced a rapid microglial loss in the spinal cord that was totally prevented or potentiated by P2X4receptor blockade or facilitation, respectively. Within the brain, microglia in the hippocampal dentate gyrus showed particularvulnerability to LPS-induced neuroinflammation. Thus, microglia processes in this region retracted as early as 2 h after injec-tion of LPS and died around 24 h later, two features which were prevented by blocking P2X4 receptors. Together, these datasuggest that P2X4 receptors contribute to controlling the fate of activated microglia and its survival.
GLIA 2014;62:171–184Key words: microglia, P2X4 purinergic receptor, EAE and multiple sclerosis, cell death
Introduction
Microglial cells are the main effectors of the innate
immune response after CNS injury (Kreutzberg, 1996).
The precursors of adult microglia are yolk sac progenitors,
which enter the embryo around E8 and colonize the CNS at
E10 (Ginhoux et al., 2010). Microglia can be found postna-
tally in all regions of the CNS, in the parenchyma and in
perivascular spaces in a nonoverlapping territorial fashion,
and comprise a large proportion of the total cellular makeup
of the CNS, estimated to be as high as 12% of all cells. Per-
haps owing to this origin, microglia share many commonal-
ties with resident tissue macrophages in peripheral organ
systems (Ransohoff and Perry, 2009). Like their peripherals
macrophage counterparts, microglia displays a remarkable
range in both morphology and activity, depending in part on
the state of the surrounding tissue (Ransohoff and Perry,
2009). In their resting state, microglia continuously monitor
the tissue for injury or pathological changes by extending and
retracting highly motile processes on a time scale of minutes
(Davalos et al., 2005; Nimmerjahn et al., 2005; reviewed in
Kettenmann et al., 2013). Exogenous factors (infectious
agents) as well as endogenous factors or neurotransmitters
released by damaged cells or tissues induce progressive
changes in microglia including altered expression of cell sur-
face markers and inflammation related genes, process retrac-
tion and acquisition of an ameboid morphology, migration of
cell bodies, proliferation and increased phagocytic rate (Ket-
tenmann et al., 2011).
The rapidly diffusible neurostransmitter ATP is one of
the main regulators of microglial functions. Low ATP concen-
trations almost exclusively activate chemotaxis, through
P2Y12 receptor activation, in order to recruit cells at the site
View this article online at wileyonlinelibrary.com. DOI: 10.1002/glia.22596
Published online November 19, 2013 in Wiley Online Library (wileyonlinelibrary.com). Received May 23, 2013, Accepted for publication Oct 16, 2013.
Address correspondence to Carlos Matute, Departamento de Neurociencias, Universidad del Pa�ıs Vasco, E-48940 Leioa, Spain. E-mail: [email protected] or
Maria Domercq, Departamento de Neurociencias, Universidad del Pa�ıs Vasco, E-48940 Leioa, Spain. E-mail: [email protected]
From the 1Departamento de Neurociencias, Universidad del Pa�ıs Vasco-UPV/EHU, Barrio Sarriena s/n, 48940, Leioa, Spain, Achucarro Basque Center for Neuro-
science-UPV/EHU, 48170, Zamudio, Spain, Instituto de Salud Carlos III, Centro de Investigaci�on Biom�edica en Red de Enfermedades Neurodegenerativas
(CIBERNED), 48940, Leioa, Spain; 2Molecular Imaging Unit, CIC biomaGUNE, Paseo Miramon 182, 20009, San Sebasti�an, Spain; 3Radiochemistry Department,
Molecular Imaging Unit, CIC biomaGUNE, Paseo Miramon 182, 20009, San Sebasti�an, Spain.
Additional Supporting Information may be found in the online version of this article.
VC 2013 Wiley Periodicals, Inc. 171

of injury or inflammation. When the ATP concentration
increases, additional effector functions, such as phagocytosis
and cytokine secretion, are also triggered. The switch on
microglial ATP responses are sometimes linked to an acute
remodeling of purinoceptor expression, i.e., upregulation of
the Gs-coupled A2A receptor and P2X4 receptor (Beggs et al.,
2012; Orr et al., 2009) and downregulation of P2Y12 (Hay-
nes et al., 2006).
While still a lot has to be learned about the events
affecting microglia throughout the activation process, even
less is known about the period after. Acute activation of
microglia after neural injury rapidly leads to reactive micro-
gliosis, a cardinal feature of expansion of microglia in the
affected CNS region (Wirenfeldt et al., 2005). The state of
reactive microgliosis resolves days to weeks later, according to
an inherently tightly regulated schedule, which has been sug-
gested to involve microglial apoptosis (Shin et al., 2004; Soria
et al., 2011). Like in the immune system, microglial cell
death secondary to its activation (called AICD, activation-
induced cell death) may be a mechanism to resolve brain
inflammation. However, little is known about the underlying
mechanisms responsible for cell death of activated microglia.
Moreover, in addition to apoptosis, other forms of cell death
such as autophagy (Arroyo et al., 2013), pyroptosis, a pro-
grammed cell death triggered by caspase-1 activation by the
inflammasome (Miao et al., 2011), and necroptosis (Fricker
et al., 2013) could contributed to microglia AICD. In this
study, we first showed that P2X4 receptors are upregulated in
multiple sclerosis (MS) samples and in one of its animal
models, encephalitis autoimmune experimental (EAE). To
gain information on the role of P2X4 signaling, we used the
most classical model of neuroinflammation, LPS treatment or
injection. We report here that a microglia can die rapidly
(<24 h) both in vitro and in vivo by a mechanism dependent
on P2X4 activation and independent of signaling by the
inflammasome.
Materials and Methods
Human Tissue SamplesPostmortem optic nerve samples from 13 long-standing MS patients
and 12 control subjects (who died from non-neurological diseases)
were obtained at autopsy under the management of the Netherlands
Brain Bank. All patients and controls had previously given written
approval for the use of their tissue, according to the guidelines of
the Netherlands Brain Bank. Clinical characteristics for control and
patient groups have been previously described (see Vallejo-
Illarramendi et al., 2006; Pampliega et al., 2011). We used clinical
data together with macroscopic tissue analysis to classify MS samples
as normal appearing (NAON) or damaged optic nerves (DON),
when showing macroscopic plaques, atrophy and/or optic neuritis.
For comparisons, MS samples were matched with control samples
for age, sex, and postmortem delay.
Induction of Acute EAE and NeuroinflammationIn vivo experiments in rats were performed in accordance with the
approved protocols and guidelines of the University of the Basque
Country Ethics Review Committee for animal experiments. Animals
were handled in accordance with the European Communities Coun-
cil Directive and all possible efforts were made to minimize animal
suffering and the number of animals used. Young adult male Lewis
rats, (200–220 g weight) were each injected subcutaneously in both
hind feet with inoculum containing 100 lg of guinea pig myelin
basic protein (Sigma) diluted in water, emulsified in equal volumes
of Freund’s incomplete adjuvant (Sigma), supplemented with 500 lg
of heat-inactivated M. tuberculosis H37Ra (DIFCO Laboratories).
The neurological deficits started 10 days postimmunization (dpi),
peaked after 14 days and remitted by 20 dpi. Neurological impair-
ment was monitored and scored daily for clinical signs of EAE
according to the following scale: 0, normal; 1, flaccid tail; 2, tail
paralysis; 3, loss of muscle tone in hind limbs; 4, hind limbs hemi-
paralysis; 5, complete hind limbs paralysis; 6, moderate paraparesis;
7, tetraparalysis; 8, death. The tissues and samples examined in this
study were obtained at 14 days after immunization when rats devel-
oped peak EAE (score 3–6).
Lipopolysaccharide (LPS) or phosphate-buffered saline (PBS)
was injected intraperitoneally (ip; 3 mg kg21) and stereotaxically in
the lateral ventricule (icv; 1 mg kg21, 10 lL). Sprague–Dawley rats
(200 g weight) were anesthetized with a mixture of ketamine (80 mg
kg21) and xylazine (10 mg kg21), both ip. The stereotaxic injection
(21 mm from Bregma; 2 mm lateral; 4 mm deep) was performed
at a rate of 1 lL min21, with the use of a microinjection autopump
and a 25-lL Hamilton microsyringe. TNP-ATP (10 mg kg21) was
administered intraperitoneally immediately and at 12 h after LPS
injection.
RNA Isolation and Quantitative Real Time PCRTotal RNA was extracted from rat and human samples using TRIzol
reagent kit (Invitrogen) and purified using the RNeasy MniKit (Qia-
gen). No DNase I treatment was performed in RNA obtained from
rat and human samples. The integrity of RNA was analyzed by ethi-
dium bromide agarose gel or using a Bioanalyzer (Agilent). For
cDNA synthesis, 2 lg of total RNA was used using random hexamer
primers and Super Script II reverse transcriptase (Invitrogen). Mock
reverse transcription (without enzyme) for each sample served as neg-
ative controls.
The relative expression level of genes was measured according
to the manufacturer’s protocol with quantitative real time PCR as
previously described (Vallejo-Illarramendi et al., 2006). Briefly,
qPCR reactions were carried out with 20–80 ng of reverse tran-
scribed RNA and 50–300 nM of forward and reverse primers (Gen-
otek, Sabadell, Spain) diluted in SYBRGreen PCR universal master
mix reagent (Applied Biosystems) in an ABI PRISM 7000 Sequence
Detection System instrument (Applied Biosystems). The expression
level was normalized with a normalization factor obtained using the
Genorm software (Vandesompele et al., 2002) based on the expres-
sion of three to six housekeeping genes (see Supp. Info. Table 1).
Primers were designed in exon–exon junction regions to avoid
gDNA amplification with the PrimerExpress software (Applied
172 Volume 62, No. 2

Biosystems; see Supp. Info. Table 1). We verified that generated fluo-
rescence was not overestimated by contamination resulting from
residual genomic DNA amplification (using RT negative controls)
and from primer dimer formation or external DNA contamination
(no template controls). qPCR products were also subjected to a dis-
sociation protocol to ensure that a single amplicon of the expected
melting temperature was indeed obtained.
Western BlotTotal protein was extracted from microglia cell cultures by scraping
the cells in SDS/sample buffer. Samples (10 lg of protein per lane)
of microglia cell cultures and rat spinal cord were boiled for 5 min,
loaded and size separated in 12.5% SDS/PAGE polyacrylamide gels.
After electroblotting on nitrocellulose membranes, proteins were
visualized using primary antibodies to P2X4 (1 lg mL21; Alomone
Labs, Jerusalem, Israel) and b-actin (1:1,000; Sigma, Madrid, Spain),
followed by secondary peroxidase-coupled goat anti-rabbit antibodies
(1:5,000; Sigma). After washing, the blots were developed using an
enhanced chemiluminescence detection kit according to the manu-
facturer’s instructions (Super Signal ULTRA, Pierce). Images were
acquired with a ChemiDoc MP system (BioRad) and quantified
using Scion Image Software.
ImmunochemistryMicroglial expression of purinergic receptors was examined by
immunofluorescence applied to cultured microglia, BV-2 cells and
tissue from rat spinal cord and cerebrum as well as human spinal
cord. Cells were fixed in 4% p-formaldehyde in PBS and processed
for immunocytochemistry as previously described (Domercq et al.,
1999). For tissue, adult Sprague–Dawley and Lewis rats were deeply
anesthetized with chloral hydrate (500 mg kg21, i.p) and transcar-
dially perfused with 0.1 M sodium phosphate buffer, pH 7.4, fol-
lowed by 4% p-formaldehyde in the same buffer. Human spinal
cord samples from MS and control subjects were obtained at autopsy
from the Netherlands Brain Bank. Characteristics of spinal cord sam-
ples are described previously (Pampliega et al., 2011). Frozen tissue
was kept at 280�C until use.
Primary antibodies were as follows to: P2X1, P2X2, P2X4,
and P2X7 (1–2 lg mL21; Alomone Labs, Jerusalem, Israel), P2X3
(1 lg mL21; Chemicon, Temecula, CA), CD11b (OX42, 1:50,
Serotec), Iba-1 (1:1,000; Wako) and CD68 (1:50; DakoCytomation)
For secondary antibodies, we used Alexa 488-conjugated goat anti-
bodies to rat IgG (Molecular Probes), and Alexa546-conjugated goat
antibody to mouse IgG (Molecular Probes). Controls were carried
out by omitting the primary antibody. Images were acquired using a
laser scanning confocal Olympus Fluoview FW500 microscopy or a
bright field Axiovision microscopy (Zeiss). Images were acquired
with the same settings for all samples within one experimental
group. P2X4 immunoreactivity per cell was calculated with the
ImageJ software (NIH, http://rsb.info.nih.gov/ij/) (n 5 40–50 cells
from at least three different experiments or samples). Morphology
analysis of microglia was performed with ImageJ software as
described before (Fontainhas et al., 2011). Microglial cells were out-
lined with the CD11b immunostaining as the defining parameter
for the region of interest. Dendritic tree area as a morphological
parameter was determined by circumscribing the area outlined by
the ends of dendritic processes using a smooth polygon tool in NIH
ImageJ. The 3D reconstructions were performed with ImageSurfer
(http://www.imagesurfer.org). CD11b and Iba11 cells were counted
blindly in longitudinal sections of spinal cord using a 403 objective
in an Axiovision microscopy (Zeiss). At least 40 different fields from
three slices per animal were counted and each treatment was per-
formed in duplicate in three different experiments.
Microglia Cell Cultures and In Vitro AssaysMicroglial cultures derived from cortical tissue from postnatal P0
rats. Briefly, cortices were dissociated and plated with serum-
supplemented Iscove’s modified Dulbecco’s medium (Gibco). After 2
weeks, confluent monolayer of cultured astrocytes was depleted from
microglia by mechanical shaking, according to standard protocols.
Free-floating microglia were collected from shaken astrocyte flasks
and purified by plating on noncoated plastic Petri dishes (Sterilin).
After 24 h, nonadhered cells (progenitors cells) were eliminated and
microglial cells were re-plated on PDL-coated coverslips or wells.
The purity of cultured microglia was higher than 99% pure under
these conditions. The immortalised murine microglial cell line BV-2
was also used as an alternative model to primary microglia. BV-2
cells were grown in serum-supplemented Dulbecco’s modified Eagle
medium (Gibco). Cells were treated with lipopolysaccharide (LPS,
E. coli O11:B4) for 24 h unless otherwise indicated.
Membrane ruffling was determined on the basis of phalloidin
immunostaining. Briefly, after LPS treatment, cells were fixed in 4%
PFA in PBS for 10 min, permeabilized for 20 min with PBS con-
taining 0.1% Triton X-100 and stained for 1 h with 2 U mL21
Texas-Red-conjugated phallodin (Invitrogen) diluted in PBS contain-
ing 1% bovine serum albumin. Phalloidin staining was observed
under the fluorescence microscopy (Axioplan, Zeiss) and blind-eye
quantified in at least 15 different fields from n 5 4 experiments per-
formed in triplicate. Cell viability was determined by measuring
LDH release to the extracellular medium after 24-h treatment with
LPS in the presence or absence of P2XR receptor antagonists. Cell-
free supernatants from microglia cultures were collected after incuba-
tion with LPS and release of rat TNFa and mature IL-1b was moni-
tored using highly sensitive ELISA kits (Invitrogen) according to the
manufacturer instructions.
Patch-Clamp ElectrophysiologyStandard whole-cell recordings of microglia (Vhold 5 270 mV)
were performed at room temperature on an inverted microscope
(CK-40, Olympus) using the EPC-7 patch-clamp amplifier (HEKA,
Lambrecht, Germany). Recordings were low-pass filtered at 2 kHz,
digitized at 5 kHz and stored as data files on a computer using the
pClamp 8.2 program (Axon Instruments, CA) for later analysis. The
extracellular bath solution contained (in mM): NaCl 140, KCl 5.4,
CaCl2 2, MgCl2 1, Hepes 10, pH 7.3. Divalent cation-free extracel-
lular solutions were obtained by omitting Ca21 and Mg21. Patch
clamp pipettes (3–5 MX) were filled with a solution containing (in
mM): CsCl 140, CaCl2 2, MgCl2 2, Hepes 10, EGTA 11, Na-ATP
2, pH 7.3. Local microperfusion of agonist was performed with a
V�azquez-Villoldo et al.: P2X4 Receptor Activation Kills Microglia
February 2014 173

multibarreled apparatus connected to an electronically driven rota-
tory motor (RSC-100, Biologic, France).
Radiochemistry[11C]CH4 was directly generated in an IBA Cyclone 18/9 cyclotron
and transferred to a TRACERlab FXC Pro synthesis module (GE
Healthcare) where [11C]CH3I was generated. [11C]PK11195 was
synthesized with the captive solvent method (Wilson et al., 2000)
using DMSO as the solvent and KOH as the base (reaction time 5
5 min). Average decay corrected radiochemical yield and specific
activity were 40% and 125 GBq lmol21, respectively. Radiochemi-
cal purity was higher than 99%.
Positron Emission Tomography Scans, DataAcquisition, and Image AnalysisPositron emission tomography was performed in rats treated with
LPS (n 5 5) in the absence or in the presence of TNP-ATP (n 5
5) and vehicle (control, n 5 5) rats at 6 h after administration to
assess the peripheral benzodiazepine receptor/translocator protein (18
kDa) TSPO (PBR/TSPO) binding. Animals were placed into a rat
holder compatible with PET acquisition systems and PET images
acquired on a General Electric eXplore Vista CT camera (GE
Healthcare) during 63 min after intravenous injection of 20 MBq of
[11C]PK11195. The attenuation correction factors were calculated
from the CT acquisition. Dynamic acquisitions were reconstructed
(decay and CT-based attenuation corrected) with filtered back pro-
jection (FBP) using a Ramp filter with a cutoff frequency of 1 Hz.
PET images were analyzed using PMOD image analysis soft-
ware (PMOD Technologies, Z€urich, Switzerland). To verify the ana-
tomical location of the signal, PET images were coregistered to the
anatomical data of the CT of the same animal for the analysis of the
cervical spinal cord and the MRI rat brain template for the analysis
of the brain and cerebellum. Likewise, three type of VOIs were
established as follows: (i) A first set of volumes of interest (VOIs)
was defined in the inner part of the spine using the CT scan of the
same rat defined to study the [11C]PK11195 binding in the cervical
cord. A VOI was manually defined in the inner part of the spine
using the CT scan of the same rat. (ii) A second set of VOIs was
automatically generated in both (ii) the brain and (iii) the cerebel-
lum, by using the regions proposed by the PMOD rat brain tem-
plate. For quantification of both VOIs, summed uptake during the
last 15 minutes of acquisition was expressed as percent of injected
dose per cc of tissue (ID/cc).
Statistical AnalysisData are presented as mean 6 SEM. Comparisons between two
groups were analyzed using Student’s two tailed t test. Comparisons
among multiple groups were analyzed by one-way ANOVA followed
by Bonferroni post hoc test.
Results
P2X4 is Overexpressed in EAE Rats and in MSSamplesWe previously demonstrated that purinergic signaling is
altered in chronic EAE, an animal model of MS, and that it
is involved in oligodendroglial cell death and axonal damage
(Matute et al., 2007). We have analyzed here the expression
of purinergic ionotropic receptors in the spinal cord of rats
after induction of acute EAE, which reproduces the neuroin-
flammatory component of the disease. At 14 days postimmu-
nization, when animals showed maximal motor deficits, we
detected a high increase in the expression of P2X4 mRNA in
spinal cord samples from rats with EAE versus control ani-
mals (Fig. 1A). Accordingly, P2X4 protein levels were signifi-
cantly increased in EAE rats (Fig. 1B). We also analyzed the
expression of P2X4 in the spinal cord by double immunoflu-
orescence using antibodies to OX-42, a marker of microglia
lineage. P2X4 was expressed at low levels in OX421 micro-
glial cells from control rats (Fig. 1C) but the expression was
increased after EAE induction in parenchymal and perivascu-
lar microglia (Fig. 1C, P2X4 grey level units in control and
EAE rats were 32.8 6 3.2 and 74.8 6 3.74, respectively, n
5 6 per group, P < 0.01).
We next studied the expression of P2X4 in post-
mortem optic nerve samples of MS patients. This cohort of
samples has been previously characterized and showed astro-
glial reactivity, inflammation and oligodendroglial cell death
(Vallejo-Illarramendi et al., 2006). We observed that the levels
of P2X4 mRNA were significantly higher in MS than in con-
trol optic nerves (1.34 average fold increase; Fig. 1D). Sur-
prisingly, we found that the increased expression in P2X4 was
higher in the normal appearing optic nerves (NAON)
(�1.56-fold; Fig. 1D) than in the damaged optic nerves
(DON) characterized by the presence of macroscopic plaques,
atrophy and/or optic neuritis (Vallejo-Illarramendi et al.,
2006; Fig. 1D). Double immunofluorescence analysis revealed
a higher expression of P2X4 in CD681 microglia/macro-
phages in the spinal cord of MS patients versus controls (Fig.
1E, P2X4 gray level units in control and MS samples were
30.2 6 1.9 and 60.1 6 3.6, respectively, n 5 4, P < 0.01).
These results are consistent with the increase of P2X4 expres-
sion in EAE animals, and suggest that altered P2X4 expres-
sion may have a pathophysiological role in MS.
P2X4 Expression and Function is Increased inActivated MicrogliaWe next investigated P2X receptor expression using in vitroas well as in vivo systemic administration of bacterial lipo-
polysaccharide, an endotoxin that signals through Toll-like
receptor 4 (Lehnardt et al., 2003), as a model of neuroinflam-
mation. In BV-2 cells, all receptors analyzed (P2X1, 2, 3, 4,
and 7) were expressed in control conditions (data not shown).
After LPS treatment (1 lg mL21, 24 h) we found an increase
in the expression of P2X4 receptors (Fig. 2A). Similar
changes after LPS treatment were observed in primary cul-
tures of microglial cells, as revealed by immunocytochemistry
174 Volume 62, No. 2

(data not shown) and Western blot (Fig. 2B), in accordance
with previous reports (Raouf et al., 2007). In control spinal
cord, microglial cells express low levels of P2X3 and P2X4
receptors and high levels of P2X7 receptors (Fig. 2C). The
other P2X receptors analyzed, P2X1 and P2X2, were located
exclusively in axons and oligodendrocytes, respectively (Fig.
2C). We did not detect any change in P2X3 and P2X7
expression after injection of LPS (3 mg kg21, i.p.; 24 h). In
contrast, there was a robust increase in P2X4 in LPS-
activated microglia (Fig. 2D; P2X4 gray level units in control
and in LPS-treated animals were 68.3 6 3.6 and 126.3 6
5.5, respectively, P < 0.001).
We next used electrophysiological recordings to analyze
P2X4 function in control and LPS-stimulated microglial cells.
ATP at low concentrations (30–100 lM), to avoid activation
of low–affinity P2X7 receptor, induced an inward current in
almost all microglial cells (n 5 18; Fig. 3A). ATP-induced
currents were greatly reduced by TNP-ATP (10 lM; n 5 12;
Fig. 3B), an antagonist of P2X1-4 receptors, by paroxetine
(10 lM, n 5 11; Fig. 3B), an antidepressant that specifically
inhibits P2X4 receptor function (Nagata et al., 2009) and by
the selective P2X4 receptor antagonist 5-BDBD (25 lM; n5 10; *P < 0.05; Fig. 3C; Bal�azs et al., 2013). Moreover,
ivermectin (IVM, 2 lM), an allosteric modulator and endo-
cytosis inhibitor of P2X4 receptor (Khakh et al., 1999;
Toulme et al., 2006), potentiated ATP (30 lM)-induced
inward currents in microglia (n 5 12; *P < 0.05; Fig. 3C).
Treatment with LPS (1 lg mL21, 24 h; n 5 18; Fig. 3A)
FIGURE 1: Increased expression of P2X4 receptors in microglia of EAE rats and of MS patients. (A–B) P2X4 mRNA (A) and protein (B) isincreased in the spinal cord of EAE rats at the peak of neurological disability (14 days after immunization). n 5 6–7 per group; *P <0.05, **P < 0.01. (C) Double immunofluorescence of P2X4 (green) and CD11b (red) in the spinal cord of EAE rats reveal an increasedexpression of P2X4 in perivascular (arrowheads) as well as parenchymal (arrows) microglial cells. Scale bar 5 25 lm (top) and 10 lm(middle and bottom). (D) P2X4 mRNA expression is increased in human optic nerves from MS patients as compared with matched con-trols. Normal appearing optic nerves (NAON), lacking macroscopic plaques, atrophy and/or optic neuritis have a significantly higherincrease in P2X4 mRNA. Controls, n 5 13; Total MS, n 5 13; MS with normal appearing optic nerves (NAON), n 5 6; MS with damagedoptic nerves (DON), n 5 7. *P < 0.05. (E) Representative images showing an increase in the level of P2X4 receptors in CD681 microglia/monocytic cells from human MS spinal cord. Scale bar 5 10 lm.
V�azquez-Villoldo et al.: P2X4 Receptor Activation Kills Microglia
February 2014 175

induced a significant increased in P2X4-mediated inward cur-
rents. In addition, currents in activated microglia cells were
similarly blocked by paroxetine and TNP-ATP (Fig. 3B).
Therefore, the pharmacological profile of ATP responses in
microglia under the conditions examined indicates that they
are mainly mediated by P2X4 receptors, and that microglial
activation leads to enhanced P2X4 receptor function.
Role of P2X4 in Microglial ActivationTo investigate the role of P2X4 receptors in microglial activa-
tion, we stimulated microglial cells with LPS in the presence
or absence of TNP-ATP. LPS treatment induced membrane
ruffling, increased in TNFa release, and morphological
changes associated with microglial activation (Fig. 4A–C). All
these LPS-induced effects were significantly inhibited by the
P2X4 receptor antagonist TNP-ATP (Fig. 4A–C). In contrast,
TNP-ATP treatment did not change the phagocytic ability of
activated microglia which was reduced by the wide spectrum
P2X antagonist PPADS (data not shown), suggesting that
other P2X receptors are responsible for regulating this func-
tion. All this data suggest that ATP release in response to
LPS treatment contributes to the phenotypic responses of
activated microglia through P2X4 receptor activation.
Activated microglia eventually undergo apoptosis by a
process known as activated-induced cell death (Mayo et al.,
2008; Takeuchi et al., 2006; Yun et al., 2011). Because of
that, we next analyzed microglia cell death after activation
with LPS. Using BV-2 cells, we observed that addition of
LPS (24 h) kill BV-2 cells in dose-dependent manner
(Fig. 4D). Cell death was blocked by TNP-ATP (10 lM),
but not in the presence of PPADS (100 lM), antagonist of
P2X1, 2, 3, 5, and 7, or BBG (50 nM), at a concentration
that preferentially blocked P2X7 receptors (Anderson and
Nerdergaard, 2006) (Fig. 4D). This data suggested that LPS-
activated microglia release ATP that induces cell death pre-
dominantly by acting at P2X4 receptor, but not at the cytoly-
tic P2X7 receptor. Accordingly addition of ATP was toxic to
BV-2 cells, an effect inhibited in the presence of TNP-ATP
(10 lM; Fig. 4E). Cell death was similarly induced in pri-
mary microglial cells after 24 h exposure to LPS and signifi-
cantly reduced in the presence of TNP-ATP (Fig. 4F) and
the P2X4 specific antagonist 5-BDBD (25 lM; Fig. 4H).
Previous reports have shown that P2X4 receptors in
neurons activate the inflammasome by promoting caspase-1
cleavage as well as the processing and maturation of IL-1b(de Ribero Vaccari et al., 2012). Caspase-1 activation by the
inflammasome leads to a form of programmed cell death
called pyroptosis, which is present in macrophages and den-
dritic cells (Miao et al., 2011). Therefore, we next analyzed
whether microglia cell death was caused by inflammasome
activation. Microglial activation with LPS showed an increase
in IL-1b production. However, IL-1b production was not
FIGURE 2: Increased expression of P2X4 receptors in microglia activated with LPS. (A) P2X4 receptors in control and LPS-treated (1 lgmL21, 24 h) BV-2 cells. Original pixel intensities from 0 to 255 are represented as a gradient (right). Scale bar 5 50 lm. (B) Western blotanalysis shows an increase in P2X4 expression in microglial cells after LPS stimulation (1 lg mL21, 24 h). n 5 3, *P < 0.05.(C,D) Immuno-staining of P2X receptors in spinal cord sections from control rats. P2X3 (C), P2X4 (D) and P2X7 (C) (green) receptors are located in thesurface of CD11b1 (red)-microglial cells, whereas no expression of P2X1 and P2X2 receptors is detected in microglia. Note the presenceof P2X4 receptors also in spinal cord motor neurons (arrowheads in D). Representative images in D of vehicle (PBS)- and LPS-injectedrats show an increased expression of P2X4 receptors (green) in microglia (red, CD11b) 24 h after LPS injection. Scale bar 5 10 lm.Insets, higher magnification of microglia showing P2X4 receptor as viewed by original pixel intensity shown in A.
176 Volume 62, No. 2

altered in the presence of TNP-ATP (Fig. 4G), excluding any
role of P2X4 receptors in inflammasome signaling. In turn,
LPS (100 ng mL21)-induced microglial cell death was
blocked by inhibiting caspase-1 with YVAD, a protective
effect that added to that of TNP-ATP, suggesting that down-
stream signaling by P2X4 did not converge with caspase-1
activation pathways (Fig. 4H). Together, these data indicates
that P2X4 receptor-mediated toxicity in microglia does not
activate the inflammosome.
P2X4 Receptors Contribute to Microglial Cell DeathIn VivoWe next analyzed the behavior of microglia in vivo 24 h after
injecting LPS (3 mg kg21, i.p.) in the presence or in the
absence of TNP-ATP (10 mg kg21, i.p.). Spinal cord sections
were stained with antibodies to CD11b (OX42), a marker of
microglia and infiltrating monocytes. CD11b1 microglia in
LPS-treated rats showed thicker and shorter processes, a hall-
mark of activated microglia and of CNS inflammation, a fact
which was more evident in grey matter microglia (Fig. 5A,B).
In addition, LPS induced a significant reduction in CD11b1
cells both in white and grey matter (Fig. 5A). Administration
of TNP-ATP completely blocked the retraction of microglial
processes and cell death (Fig. 5A,B). To further assess micro-
glial cell loss, we imaged microglia in vivo by PET using
[11C]PK11195, a radioligand that binds to peripheral benzo-
diazepine receptor PBR/TSPO, a mitochondrial transmem-
brane protein that is mainly expressed in the cerebral
monocytic lineage (microglia and monocyte). We compared
[11C]PK11195 PET signal in control and LPS-injected rats.
LPS induced a reduction in [11C]PK11195 specific binding
in spinal cord and cerebellum whereas no significant change
was detected in total brain (Fig. 5C,D). Moreover, TNP-ATP
completely prevented LPS-induced reduction in
[11C]PK11195 PET signal in the spinal cord (P < 0.05) and
cerebellum (P 5 0.06) (Fig. 5C,D).
To exclude any role of peripheral immune system, we
evaluated microglial cell number in the spinal cord at 2, 6,
and 24 h after intracerebroventricular (icv) LPS injection.
Cells were identified using two markers of microglia, CD11b
FIGURE 3: P2X4 receptor-mediated responses increase in LPS-activated microglia. (A) ATP (100 lM, 1 mM) evokes inward, nondesensi-tizing currents in microglia, which are potentiated in the presence of LPS (1 lg mL21, 24 h). (B) ATP (100 lM)-induced currents areblocked by TNP-ATP (10 lM) and paroxetine (10 lM), two antagonists of P2X4 receptors, in control and LPS-activated microglia. (C)Ivermectin (IVM, 3 lM), an allosteric modulator of microglia, potentiates ATP (30 lM)-induced inward currents whereas 5-BDBD (25 lM)another antagonist of P2X4 receptors, blocks ATP-inward currents.
V�azquez-Villoldo et al.: P2X4 Receptor Activation Kills Microglia
February 2014 177

(OX42) and Iba-1 (ionized calcium-binding adapter molecule
1), which is not expressed in infiltrating monocytes (Ajami
et al., 2011). A reduction of microglial cells (CD11b1,
Iba11) was similarly detected in spinal cord (white and gray
matter) as early as 6 h after icv injection of LPS, an effect
that was increased at 24 h, and P2X4 receptor blockage com-
pletely prevented microglial cell loss (Fig. 6A). Microglial loss
at 24 h was further increased by potentiating P2X4 receptor
function with ivermectin (IVM, 1 mg kg21, i.p.; Fig. 6B)
and inhibited by the P2X4 receptor antagonist 5-BDBD (0.7
mg kg21; i.c.v.; Fig. 6B), thus confirming the role of P2X4
receptors in microglia cell death in vivo.
Surprisingly, when we analyzed the cerebrum after icv
LPS administration, we observed clear region heterogeneity
FIGURE 4: P2X4 receptors regulate LPS-induced microglial activation and cell death. (A–C) P2X4 receptor blockade with TNP-ATP (10lM) prevents LPS-induced microglia ruffling (A, 24 h), attenuates secretion of inflammatory mediators such as TNFa (B, 24 h) and themorphological changes accompanying microglial activation (C; analyzed at 4 h). *P < 0.05 vs. LPS-treated cultures from at least three dif-ferent experiments performed in triplicate. Scale bar 5 20 lm. (D–F) LPS (24 h) induces dose-dependent cell death of BV-2 cells (D) andmicroglia (F). BV-2 cell death is inhibited by TNP-ATP (10 lM), but not by PPADS, antagonist of P2X1, 2, 3, 5, and 7, or BBG (50 nM), aselective blocker of P2X7 at this concentration (D). ATP (1 mM, 24 h) induces BV-2 cell death that is significantly inhibited in the pres-ence of TNP-ATP (E). Microglia cell death is significantly reduced in the presence of TNP-ATP and YVAD, an inhibitor of caspase-1 (F).(G) Mature IL-1b secretion, a measure of inflammasome activation, induced by LPS (24 h) is not modulated by P2X4 blockade. (H) Simul-taneous blocking of P2X4 receptors and caspase-1 with YVAD (50 lM) further reduces LPS-induced microglia cell death currents, indicat-ing that both signaling pathways in microglia are independent. Cell death is also reduced by the P2X4 antagonist 5-BDBD (25 lM).Curves and histograms in D–H represents mean 6 SEM from at least three different experiments performed in triplicate. *P < 0.05.[Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
178 Volume 62, No. 2

on the microglial response with the hippocampal dentate
gyrus being most sensitive. Thus, at 2 h after LPS injection,
microglia showed retracted processes in this area, a hallmark of
activated microglia, as quantified by measuring microglia area
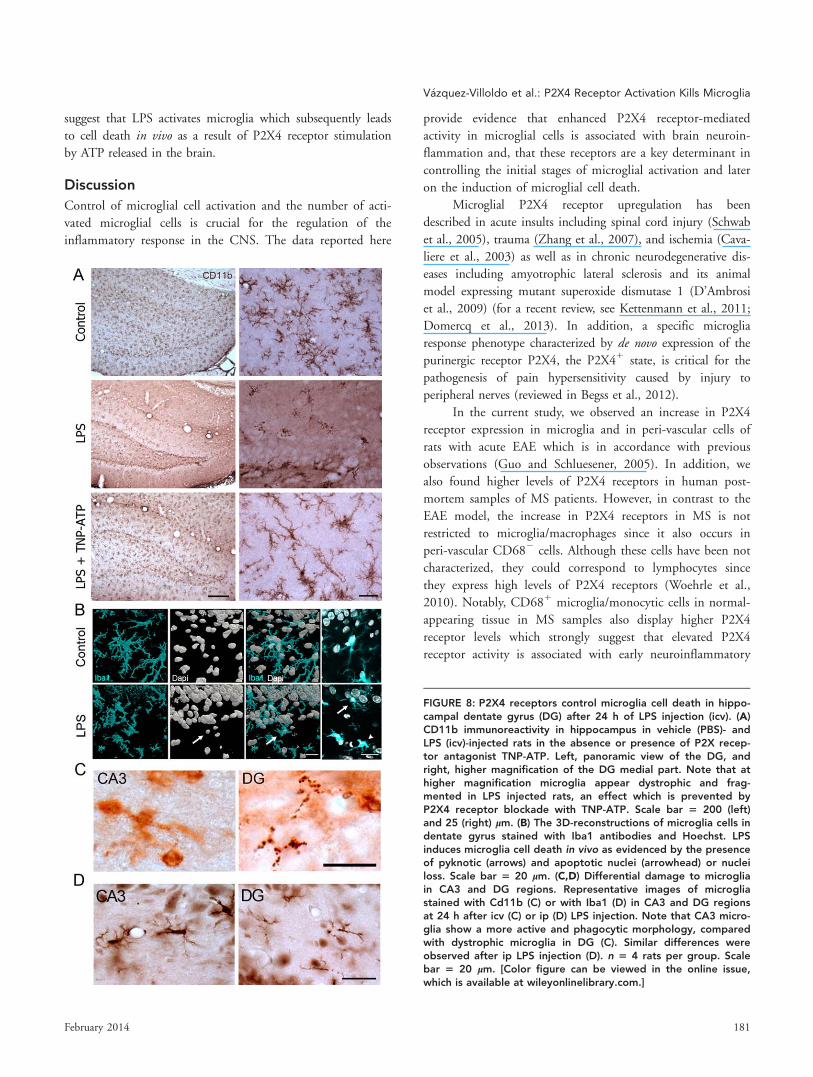
and process length (Fig. 7A,B). At 24 h, not at 6 h, microglia
in dentate gyrus appeared dystrophic and showed fragmented
processes, a phenomenon called cytorrhexis and which associ-
ated to aging in human brains (Streit et al., 2004) and to neu-
rodegenerative diseases (Fendrick et al., 2007; Streit et al.,
2009; Xue et al., 2011) (Fig. 8A). Double immunostaining
with Hoechst showed nuclei loss, piknosis and DNA fragmen-
tation in microglia (Fig. 8B), further confirming the degenera-
tion of microglia. In contrast to dentate gyrus, microglia in
CA3 at 24 h showed ameboid and phagocytic morphology
(Fig. 8C). In other regions like cortex, no clear signs of activa-
tion were detected at the earlier stages examined and rare
degenerating microglial cells were observed close to the cortical
surface. The different response of microglia in dentate gyrus
could reflect differences in the vulnerability of cells in this area
and/or differences in LPS exposure. However, we also observed
microglia cell death in the dentate gyrus 24 h after i.p. LPS
injection (Fig. 8D), suggesting that microglia in this region
FIGURE 5: P2X4 receptor blockade prevents microglial loss in the spinal cord after i.p. injection of LPS. (A) Top, Photographs of spi-nal cord sections of vehicle (saline)-, LPS-, and LPS 1 TNP-ATP-injected rats stained for CD11b. Scale bar 5 50 lm. Bottom, Histo-gram illustrates that rats receiving LPS show a significant decrease in the number of CD11b1 cells compared with vehicle-treatedcontrols, and that TNP-ATP administration (i.p.) reverts this effect. Similar results were obtained in three independent experimentsperformed in duplicate. **, ##P < 0.01 vs. control (*) or vs. LPS-treated rats (#). (B) Higher magnification images showing representa-tive microglia morphology in the different experimental conditions. Note the change in cell shape after LPS injection, which isreverted by TNP-ATP treatment. Scale bar 5 20 lm. (C,D) Representative images of [11C]PK11195 PET signal in the spinal cord (C)and cerebrum (D) of vehicle (PBS)-, LPS-, and LPS 1 TNP-ATP-injected rats. PET images were coregistered with an MRI (T2W) rat tem-plate to localize anatomically the PET signal. The percentage of injected dose per cubic centimeter (%ID/cc; mean 6 s.d.) was quanti-fied in ROI areas defined in the cervical spinal cord, in the cerebellum and in total brain. [11C]PK11195 PET signal decreasessignificantly in the spinal cord (C) and cerebellum (D) of LPS-injected rats versus control rats. Administration of TNP-ATP significantlyblocks the LPS-induced decrease in [11C]PK11195 PET signal. n 5 5 per group. *, #P < 0.05 and **, ##P < 0.01 vs. control (*) or vs.LPS-treated rats (#). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
V�azquez-Villoldo et al.: P2X4 Receptor Activation Kills Microglia
February 2014 179

shows higher vulnerability. As described above for the spinal
cord, TNP-ATP treatment prevented microglial activation and
cell death in the hippocampus of icv LPS-injected rats. These
observations are in accordance with the in vitro data and
FIGURE 6: P2X4 receptors control microglial loss in the spinalcord following intracerebroventricular LPS injection. (A–C) Histo-grams showing the time course of microglial loss in the spinalcord after intracerebroventricular injection of LPS. Spinal cord sec-tions of vehicle (saline)- and LPS-injected rats were stained withIba1 and positive cells counted at 2 h (A), 6 h (B), and 24 h (C)after LPS injection. TNP-ATP, administered (i.p.) immediately afterLPS injection, significantly blocks microglial loss induced by LPS.Similar effects were found both in grey and white matter. (D) Iver-mectin, a positive modulator of P2X4 receptor function, signifi-cantly increases microglial loss induced by LPS whereas the P2X4antagonist 5-BDBD has the opposite effect. Values in A–D repre-sents mean 6 SEM from n 5 4 rats per group. *P < 0.05. *, #P <0.05 and **, ##P < 0.01 vs. control (*) or vs. LPS-treated rats (#).
FIGURE 7: P2X4 receptors determine early microglial activationin hippocampal dentate gryus at 2 h after LPS injection (icv). (A)CD11b immunoreactivity in hippocampus in vehicle (saline)-, LPS(icv)-injected rats in the absence or presence of the P2X4 recep-tor antagonist TNP-ATP. Scale bar 5 200 (left) and 50 (right) lm.(B) Reconstruction of microglia morphology in hippocampal den-tate gyrus in two-dimensional image projections of z-stacks con-focal images. Scale bar 5 20 lm. Two parameters were analyzedusing Image J software (see methods): (1) arborization area (areacircumscribed by the polygonal object defined by connecting theouter points of the ramified arbor), (2) Length of the segmentsidentified in a skeletonization of the arbor. Blockage of P2X4receptors with TNP-ATP abolishes microglial process retractioninduced by LPS. **, ##P < 0.01 vs. control (**) or vs. icv LPS-treated rats (##) (n 5 4 rats per group). [Color figure can beviewed in the online issue, which is available atwileyonlinelibrary.com.]
180 Volume 62, No. 2
suggest that LPS activates microglia which subsequently leads
to cell death in vivo as a result of P2X4 receptor stimulation
by ATP released in the brain.
Discussion
Control of microglial cell activation and the number of acti-
vated microglial cells is crucial for the regulation of the
inflammatory response in the CNS. The data reported here
provide evidence that enhanced P2X4 receptor-mediated
activity in microglial cells is associated with brain neuroin-
flammation and, that these receptors are a key determinant in
controlling the initial stages of microglial activation and later
on the induction of microglial cell death.
Microglial P2X4 receptor upregulation has been
described in acute insults including spinal cord injury (Schwab
et al., 2005), trauma (Zhang et al., 2007), and ischemia (Cava-
liere et al., 2003) as well as in chronic neurodegenerative dis-
eases including amyotrophic lateral sclerosis and its animal
model expressing mutant superoxide dismutase 1 (D’Ambrosi
et al., 2009) (for a recent review, see Kettenmann et al., 2011;
Domercq et al., 2013). In addition, a specific microglia
response phenotype characterized by de novo expression of the
purinergic receptor P2X4, the P2X41 state, is critical for the
pathogenesis of pain hypersensitivity caused by injury to
peripheral nerves (reviewed in Begss et al., 2012).
In the current study, we observed an increase in P2X4
receptor expression in microglia and in peri-vascular cells of
rats with acute EAE which is in accordance with previous
observations (Guo and Schluesener, 2005). In addition, we
also found higher levels of P2X4 receptors in human post-
mortem samples of MS patients. However, in contrast to the
EAE model, the increase in P2X4 receptors in MS is not
restricted to microglia/macrophages since it also occurs in
peri-vascular CD682 cells. Although these cells have been not
characterized, they could correspond to lymphocytes since
they express high levels of P2X4 receptors (Woehrle et al.,
2010). Notably, CD681 microglia/monocytic cells in normal-
appearing tissue in MS samples also display higher P2X4
receptor levels which strongly suggest that elevated P2X4
receptor activity is associated with early neuroinflammatory
FIGURE 8.
FIGURE 8: P2X4 receptors control microglia cell death in hippo-campal dentate gyrus (DG) after 24 h of LPS injection (icv). (A)CD11b immunoreactivity in hippocampus in vehicle (PBS)- andLPS (icv)-injected rats in the absence or presence of P2X recep-tor antagonist TNP-ATP. Left, panoramic view of the DG, andright, higher magnification of the DG medial part. Note that athigher magnification microglia appear dystrophic and frag-mented in LPS injected rats, an effect which is prevented byP2X4 receptor blockade with TNP-ATP. Scale bar 5 200 (left)and 25 (right) lm. (B) The 3D-reconstructions of microglia cells indentate gyrus stained with Iba1 antibodies and Hoechst. LPSinduces microglia cell death in vivo as evidenced by the presenceof pyknotic (arrows) and apoptotic nuclei (arrowhead) or nucleiloss. Scale bar 5 20 lm. (C,D) Differential damage to microgliain CA3 and DG regions. Representative images of microgliastained with Cd11b (C) or with Iba1 (D) in CA3 and DG regionsat 24 h after icv (C) or ip (D) LPS injection. Note that CA3 micro-glia show a more active and phagocytic morphology, comparedwith dystrophic microglia in DG (C). Similar differences wereobserved after ip LPS injection (D). n 5 4 rats per group. Scalebar 5 20 lm. [Color figure can be viewed in the online issue,which is available at wileyonlinelibrary.com.]
V�azquez-Villoldo et al.: P2X4 Receptor Activation Kills Microglia
February 2014 181

events in this disorder. Determination of these early molecular
alterations in normal appearing tissue in MS can be useful to
elucidate the mechanisms of lesion formation and to develop
new treatments.
Purinergic receptors control several microglial functions,
including the motility of their fine processes, migration,
cytokine release, and phagocytosis (Domercq et al., 2013;
Kettenmann et al., 2013). Thus, P2X4 receptors control
microglial chemotaxis in conjunction with P2Y12 receptors
(Ohsawa et al., 2007), and stimulating P2X4 receptors ini-
tiates a core signaling pathway that mediates the synthesis
and release of brain-derived neurotrophic factor (Trang et al.,
2009), which produces a disinhibition of nociceptive trans-
mission (Tsuda et al., 2003; reviewed in Beggs et al., 2012).
In turn, our data indicates that P2X4 receptor activity con-
tributes to LPS-induced microglial activation in vitro since
P2X4 antagonists reduce microglia ruffling, TNFa secretion,
morphological changes and cell death. Indeed, the effect of
blocking P2X4 receptors is striking in vivo since it prevents
morphological changes of microglia associated to their activa-
tion and microglia cell death in the spinal cord and hippo-
campus. Differences between microglia in culture and in vivocould be due to differences in P2X4 receptor expression in
microglia from newborn rats versus microglia from adult rats
since purinergic receptors are developmentally regulated
(Xiang and Burnstock, 2005).
Consistent with our findings, microglial cell death sec-
ondary to microglial activation by LPS or tissue injury has
been reported both in vitro and in vivo (Arroyo et al., 2013;
Fricker et al., 2013; Jeong et al., 2010; Ji et al., 2007; Jun
et al., 2011; Min et al., 2012; Shin et al., 2004). However, to
our knowledge the data described in the current study pro-
vides the first evidence that activation of P2X4 receptors, sec-
ondary to ATP released by LPS stimulation, induces
microglia cell death. In contrast to our results, a recent study
in cultured microglia and BV-2 cells demonstrated that pro-
longed activation of P2X4 receptors in microglia by ATP
induces opening of a pore permeable to large molecules that
does not lead to cell death (Bernier et al., 2012). This appa-
rent discrepancy may be due to differences on the age (2- to
4-day old vs. newborn) and species (mice vs. rats) of the ani-
mals used for culturing microglia, and to the time window
examined for monitoring microglia cell death after ATP or
LPS stimulation in vitro (90–180 min vs. 24 h).
We observed that, within the brain, microglia in the
dentate gyrus were particularly vulnerable to LPS stimulation.
Microglia in dentate gyrus are more densely populated, acti-
vated and proliferative because they are located in a neuro-
genic niche and neural progenitor cells have a secretory
protein profile distinct from other brain cells that regulates
microglial activation state (Mosher et al., 2012). Thus, it is
conceivable that this higher level of basal activation renders
microglia more vulnerable to LPS stimulation and the ensu-
ing activation of P2X4 receptors. However, we did not detect
higher P2X4 receptor expression in dentate gyrus microglia
by immunohistochemistry (data not shown) though electro-
physiological recordings will be needed to analyze whether
these cells have enhanced responses to ATP.
ATP homeostasis is compromised in most CNS pathol-
ogies. Immediately after acute CNS injury, damaged cells
release ATP as a “find-me” signal of death to recruit phago-
cytes. The main mechanism of ATP release is the plasma
membrane channel pannexin 1, which opens in most patho-
logical conditions (Domercq et al., 2010; MacVicar and
Thompson, 2010; Pelegrin and Surprenant, 2006; Thompson
et al., 2006; for a review see Domercq et al., 2013). Our
results suggest that microglia releases ATP in response to LPS
stimuli. Indeed, microglia releases ATP after activation with
LPS, leading to an increase in excitatory neurotransmission
(Pascual et al., 2012). However, the expression of pannexins
in microglia and their possible influence on ATP release
under normal conditions and after microglial activation has
not been characterized. Other mechanisms of ATP release in
microglia have been proposed, including ATP release via zinc
uptake by zinc transporters (Higashi et al., 2011) and Ca21-
dependent exocytosis from lysosomes (Dou et al., 2012).
The physiological significance of microglia cell death
remains to be determined. It has been proposed that the
extent and duration of inflammation in the CNS may be
controlled by removal of activated microglia. According to
this hypothesis, microglia proliferates and then undergo apo-
ptosis to achieve tissue homeostasis, as described in chronic
EAE (Ajami et al., 2011). However, microglial cell death
occurs within hours after the insult in the acute neuroinflam-
mation experimental paradigms used in our study (6 and 24
h after ip and icv LPS injection, respectively). Similarly, early
microglial cell death was also observed in the penumbra of
spinal cord lesions where ramified Iba-11 cells (resident
microglia) die earlier (3–6 h) than neurons (12–24 h) (Min
et al., 2012). Moreover, microglia die as early as neurons
within the core region after ATP injection into the substantia
nigra pars compacta, indicating that microglia are as vulnera-
ble to this type of insult as neurons (Jeong et al., 2010). Con-
sequently, early microglia cell death appears to be a direct
consequence of some insults and determines a previously
unnoticed susceptibility of microglia to CNS damage, infec-
tion or altered homeostasis. Early microglia cell death is fol-
lowed by the infiltration and proliferation of circulating
monocytes with stronger phagocytic activity that could con-
tribute to resolve and repair the damage tissue by removing
apoptotic cells and facilitating angiogenesis (Ji et al., 2007;
Jeong et al., 2010).
182 Volume 62, No. 2

Classically, microglia are considered as the immune cells
of the brain and key players in brain injury and disease.
However, in the last decade our view of microglia has dramat-
ically changed after discovering that resting microglia are
extremely dynamic in vivo, perpetually changing their mor-
phology by extending and retracting highly motile processes,
a mechanism that allows microglia to regulate important
functions such as postnatal development, adult neuronal
plasticity, and synaptic excitability (Kettenmann et al., 2013;
Li et al., 2012; Tremblay et al., 2011). Therefore, primary
microglial loss at the initial stages of disease may contribute
to impairing brain function. This may indeed be the case of
microglial apoptosis in perilesional white matter in X-linked
adrenoleukodistrophy (Eichler et al., 2008) as well as of dys-
trophic (fragmented) microglia associated with tau pathology
which precedes the spread of tau pathology and the neuro-
degeneration in Alzheimer’s disease (Streit et al., 2009; Xue
and Streit, 2011). The findings reported here illustrating
that P2X4 receptors mediate early microglial cell death dur-
ing neuroinflammation provide new avenues to control the
fate of activated microglia, and possibly to manipulate
microgliosis towards a beneficial phenotype in CNS injury
and disease.
Acknowledgment
Grant sponsor: CIBERNED; the Spanish Ministry of Educa-
tion and Science; Eranet-Neuron (Nanostroke); the Education
Department and the Industry Department of the Basque
Government.
We would like to thank to M. Gonzalez, A. Leukona and
M. Errasti for technical support in the radiosynthesis, and to
A. Cano and B. Szczupak for technical assistance in the the
PET studies.
References
Ajami B, Bennett JL, Krieger C, McNagny KM, Rossi FM. 2011. Infiltratingmonocytes trigger EAE progression, but do not contribute to the residentmicroglia pool. Nat Neurosci 14:1142–1149.
Anderson CM, Nedergaard M. 2006. Emerging challenges of assigning P2X7receptor function and immunoreactivity in neurons. Trends Neurosci 29:257–262.
Arroyo DS, Soria JA, Gaviglio EA, Garcia-Keller C, Cancela LM, Rodriguez-Galan MC, Wang JM, Iribarren P. 2013. Toll-like receptor 2 ligands promotemicroglial cell death by inducing autophagy. FASEB J 27:299–312.
Bal�azs B, Dank�o T, Kov�acs G, K€oles L, Hediger MA, Zsembery A. 2013. Inves-tigation of the inhibitory effects of the benzodiazepine derivative, 5-BDBD onP2X4 purinergic receptors by two complementary methods. Cell Physiol Bio-chem 32:11–24.
Beggs S, Trang T, Salter MW. 2012. P2X4R1 microglia drive neuropathicpain. Nat Neurosci 15:1068–1073.
Bernier LP, Ase AR, Bou�e-Grabot E, S�egu�ela P. 2012. P2X4 receptor channelsform large noncytolytic pores in resting and activated microglia. Glia 60:728–737.
Cavaliere F, Florenzano F, Amadio S, Fusco FR, Viscomi MT, D’Ambrosi N,Vacca F, Sancesario G, Bernardi G, Molinari M, Volonte C. 2003. Up-regula-tion of P2X2, P2X4 receptor and ischemic cell death: Prevention by P2 antag-onists. Neuroscience 120:85–98.
D’Ambrosi N, Finocchi P, Apolloni S, Cozzolino M, Ferri A, Padovano V,Pietrini G, Carr�ı MT, Volont�e C. 2009. The proinflammatory action of micro-glial P2 receptors is enhanced in SOD1 models for amyotrophic lateral sclero-sis. J Immunol 183:4648–4656.
Davalos D, Grutzendler J, Yang G, Kim JV, Zuo Y, Jung S, Littman DR, DustinML, Gan WB. 2005. ATP mediates rapid microglial response to local braininjury in vivo. Nat Neurosci 8:752–758.
de Rivero Vaccari JP, Bastien D, Yurcisin G, Pineau I, Dietrich WD, DeKoninck Y, Keane RW, Lacroix S. 2012. P2X4 receptors influence inflamma-some activation after spinal cord injury. J Neurosci 32:3058–3066.
Domercq M, Perez-Samartin A, Aparicio D, Alberdi E, Pampliega O, MatuteC. 2010. P2X7 receptors mediate ischemic damage to oligodendrocytes. Glia58:730–740.
Domercq M, S�anchez-G�omez MV, Areso P, Matute C. 1999. Expression ofglutamate transporters in rat optic nerve oligodendrocytes. Eur J Neurosci11:2226–2236.
Domercq M, V�azquez-Villoldo N, Matute C. 2013. Neurotransmitter signalingin the pathophysiology of microglia. Front Neurosci 7:49.
Dou Y, Wu HJ, Li HQ, Qin S, Wang YE, Li J, Lou HF, Chen Z, Li XM, LuoQM, Duan S. 2012. Microglial migration mediated by ATP-induced ATPrelease from lysosomes. Cell Res 22:1022–1033.
Eichler FS, Ren JQ, Cossoy M, Rietsch AM, Nagpal S, Moser AB, Frosch MP,Ransohoff RM. 2008. Is microglial apoptosis an early pathogenic change incerebral X-linked adrenoleukodystrophy? Ann Neurol 63:729–742.
Fendrick SE, Xue QS, Streit WJ. 2007. Formation of multinucleated giant cellsand microglial degeneration in rats expressing a mutant Cu/Zn superoxidedismutase gene. J Neuroinflammation 4:9.
Fontainhas AM, Wang M, Liang KJ, Chen S, Mettu P, Damani M, Fariss RN,Li W, Wong WT. 2011. Microglial morphology and dynamic behavior is regu-lated by ionotropic glutamatergic and GABAergic neurotransmission. PLoSOne 6:e15973.
Fricker M, Vilalta A, Tolkovsky AM, Brown GC. 2013. Caspase inhibitors pro-tect neurons by enabling selective necroptosis of inflamed microglia. J BiolChem 288:9145–9152.
Ginhoux F, Greter M, Leboeuf M, Nandi S, See P, Gokhan S, Mehler MF,Conway SJ, Ng LG, Stanley ER, Samokhvalov IM, Merad M. 2010. Fate map-ping analysis reveals that adult microglia derive from primitive macrophages.Science 330:841–845.
Guo LH, Schluesener HJ. 2005. Lesional accumulation of P2X(4) receptor(1)macrophages in rat CNS during experimental autoimmune encephalomyelitis.Neuroscience 134:199–205.
Haynes SE, Hollopeter G, Yang G, Kurpius D, Dailey ME, Gan WB, Julius D.2006. The P2Y12 receptor regulates microglial activation by extracellularnucleotides. Nat Neurosci 9:1512–1519.
Higashi Y, Segawa S, Matsuo T, Nakamura S, Kikkawa Y, Nishida K,Nagasawa K. 2011. Microglial zinc uptake via zinc transporters induces ATPrelease and the activation of microglia. Glia 59:1933–1945.
Jeong HK, Ji KM, Kim B, Kim J, Jou I, Joe EH. 2010. Inflammatory responsesare not sufficient to cause delayed neuronal death in ATP-induced acutebrain injury. PLoS One 5:e13756.
Ji KA, Yang MS, Jeong HK, Min KJ, Kang SH, Jou I, Joe EH. 2007. Residentmicroglia die and infiltrated neutrophils and monocytes become majorinflammatory cells in lipopolysaccharide-injected brain. Glia 55:1577–1588.
Kahkh BS, Proctor WR, Dunwiddie TV, Labarca C, Lester HA. 1999. Allostericcontrol of gating and kinetics at P2X(4) receptor channels. J Neurosci 19:7289–7299.
Kettenmann H, Hanisch UK, Noda M, Verkhratsky A. 2011. Physiology ofmicroglia. Physiol Rev 91:461–553.
V�azquez-Villoldo et al.: P2X4 Receptor Activation Kills Microglia
February 2014 183

Kettenmann H, Kirchhoff F, Verkhratsky A. 2013. Microglia: New roles for thesynaptic stripper. Neuron 77:10–18.
Kreutzberg GW. 1996. Microglia: A sensor for pathological events in theCNS. Trends Neurosci 19:312–318.
Lehnardt S, Massillon L, Follett P, Jensen FE, Ratan R, Rosenberg PA, VolpeJJ, Vartanian T. 2003. Activation of innate immunity in the CNS triggers neu-rodegeneration through a Toll-like receptor 4-dependent pathway. Proc NatlAcad Sci USA 100:8514–8519.
Li Y, Du XF, Liu CS, Wen ZL, Du JL. 2012. Reciprocal regulation betweenresting microglial dynamics and neuronal activity in vivo. Dev Cell 23:1189–1202.
MacVicar BA, Thompson RJ. 2010. Non-junction functions of pannexin-1channels. Trends Neurosci 33:93–102.
Matute C, Torre I, P�erez-Cerd�a F, P�erez-Samart�ın A, Alberdi E, Etxebarria E,Arranz AM, Ravid R, Rodr�ıguez-Antig€uedad A, S�anchez-G�omez M, DomercqM. 2007. P2X(7) receptor blockade prevents ATP excitotoxicity in oligoden-drocytes and ameliorates experimental autoimmune encephalomyelitis.J Neurosci 27:9525–9533.
Mayo L, Jacob-Hirsch J, Amariglio N, Rechavi G, Moutin MJ, Lund FE, SteinR. 2008. Dual role of CD38 in microglial activation and activation-inducedcell death. J Immunol 181:92–103.
Miao EA, Rajan JV, Aderem A. 2011. Caspase-1-induced pyroptotic celldeath. Immunol Rev 243:206–214.
Min KJ, Jeong HK, Kim B, Hwang DH, Shin HY, Nguyen AT, Kim JH, Jou I,Kim BG, Joe EH. 2012. Spatial and temporal correlation in progressivedegeneration of neurons and astrocytes in contusion-induced spinal cordinjury. J Neuroinflammation 9:100.
Mosher KI, Andres RH, Fukuhara T, Bieri G, Hasegawa-Moriyama M, He Y,Guzman R, Wyss-Coray T. 2012. Neural progenitor cells regulate microgliafunctions and activity. Nat Neurosci 15:1485–1487.
Nagata K, Imai T, Yamashita T, Tsuda M, Tozaki-Saitoh H, Inoue K. 2009.Antidepressants inhibit P2X4 receptor function: A possible involvement inneuropathic pain relief. Mol Pain 5:20.
Nimmerjahn A, Kirchhoff F, Helmchen F. 2005. Resting microglial cells arehighly dynamic surveillants of brain parenchyma in vivo. Science 308:1314–1318.
Ohsawa K, Irino Y, Nakamura Y, Akazawa C, Inoue K, Kohsaka S. 2007.Involvement of P2X4 and P2Y12 receptors in ATP-induced microglial chemo-taxis. Glia 55:604–616.
Orr AG, Orr AL, Li XJ, Gross RE, Traynelis SF. 2009. Adenosine A(2A) recep-tor mediates microglial process retraction. Nat Neurosci 12:872–878.
Pampliega O, Domercq M, Soria FN, Villoslada P, Rodr�ıguez-Antig€uedad A,Matute C. 2011. Increased expression of cystine/glutamate antiporter in mul-tiple sclerosis. J Neuroinflammation 8:63.
Pascual O, Ben Achour S, Rostaing P, Triller A, Bessis A. 2012. Microglia acti-vation triggers astrocyte-mediated modulation of excitatory neurotransmis-sion. Proc Natl Acad Sci USA 109:197–205.
Pelegrin P, Surprenant A. 2006. Pannexin-1 mediates large pore formationand interleukin-1beta release by the ATP-gated P2X7 receptor. EMBO J 25:5071–5082.
Ransohoff RM, Perry VH. 2009. Microglial physiology: Unique stimuli, special-ized responses. Annu Rev Immunol 27:119–145.
Raouf R, Chabot-Dor�e AJ, Ase AR, Blais D, S�egu�ela P. 2007. Differential reg-ulation of microglial P2X4 and P2X7 ATP receptors following LPS-inducedactivation. Neuropharmacology 53:496–504.
Schwab JM, Guo L, Schluesener HJ. 2005. Spinal cord injury induces earlyand persistent lesional P2X4 receptor expression. J Neuroimmunol 163:185–189.
Shin WH, Lee DY, Park KW, Kim SU, Yang MS, Joe EH, Jin BK. 2004. Micro-glia expressing interleukin-13 undergo cell death and contribute to neuronalsurvival in vivo. Glia 46:142–152.
Soria JA, Arroyo DS, Gaviglio EA, Rodriguez-Galan MC, Wang JM, IribarrenP. 2011. Interleukin 4 induces the apoptosis of mouse microglial cells by acaspase-dependent mechanism. Neurobiol Dis 43:616–624.
Streit WJ, Braak H, Xue QS, Bechmann I. 2009. Dystrophic (senescent) ratherthan activated microglial cells are associated with tau pathology and likelyprecede neurodegeneration in Alzheimer’s disease. Acta Neuropathol 118:475–485.
Streit WJ, Sammons NW, Kuhns AJ, Sparks DL. 2004. Dystrophic microglia inthe aging human brain. Glia 45:208–212.
Takeuchi H, Wang J, Kawanokuchi J, Mitsuma N, Mizuno T, Suzumura A.2006. Interferon-gamma induces microglial-activation-induced cell death: Ahypothetical mechanism of relapse and remission in multiple sclerosis. Neuro-biol Dis 22:33–39.
Thompson RJ, Zhou N, Mac Vicar BA. 2006. Ischemia opens neuronal gapjunction hemichannels. Science 312:924–927.
Toulm�e E, Soto F, Garret M, Bou�e-Grabot EC. 2006. Functional properties ofinternalization-deficient P2X4 receptors reveal a novel mechanism of ligand-gated channel facilitation by ivermectin. Mol Pharmacol 69:576–587.
Trang T, Beggs S, Wan X, Salter MW. 2009. P2X4-receptor-mediated synthe-sis and release of brain-derived neurotrophic factor in microglia is dependenton calcium and p38-mitogen-activated protein kinase activation. J Neurosci29:3518–3528.
Tremblay M�E, Stevens B, Sierra A, Wake H, Bessis A, Nimmerjahn A. 2011.The role of microglia in the healthy brain. J Neurosci 31:16064–16069.
Tsuda M, Shigemoto-Mogami Y, Koizumi S, Mizokoshi A, Kohsaka S, SalterMW, Inoue K. 2003. P2X4 receptors induced in spinal microglia gate tactileallodynia after nerve injury. Nature 424:778–783.
Vallejo-Illarramendi A, Domercq M, P�erez-Cerd�a F, Ravid R, Matute C. 2006.Increased expression and function of glutamate transporters in multiple scle-rosis. Neurobiol Dis 21:154–164.
Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A,Speleman F. 2002. Accurate normalization of real-time quantitative RT-PCRdata by geometric averaging of multiple internal control genes. Genome Biol3:RESEARCH0034.
Wilson AA, Garcia A, Jin L, Houle S. 2000. Radiotracer synthesis from [11C]-iodomethane: A remarkably simple captive solvent method. Nucl Med Biol27:529–532.
Wirenfeldt M, Babcock AA, Ladeby R, Lambertsen KL, Dagnaes-Hansen F,Leslie RG, Owens T, Finsen B. 2005. Reactive microgliosis engages distinctresponses by microglial subpopulations after minor central nervous systeminjury. J Neurosci Res 82:507–514.
Woehrle T, Yip L, Elkhal A, Sumi Y, Chen Y, Yao Y, Insel PA, Junger WG.2010. Pannexin-1 hemichannel-mediated ATP release together with P2X1 andP2X4 receptors regulate T-cell activation at the immune synapse. Blood 116:3475–3484.
Xiang Z, Burnstock G. 2005. Expression of P2X receptors on rat microglialcells during early development. Glia 52:119–126.
Xue QS, Streit WJ. 2011. Microglial pathology in Down syndrome. Acta Neu-ropathol 122:455–466.
Yun HJ, Yoon JH, Lee JK, Noh KT, Yoon KW, Oh SP, Oh HJ, Chae JS,Hwang SG, Kim EH, Maul GG, Lim DS, Choi EJ. 2011. Daxx mediatesactivation-induced cell death in microglia by triggering MST1 signalling.EMBO J 30:2465–2476.
Zhang Z, Zhang Z, Artelt M, Burnet M, Schluesener HJ. 2007. Dexametha-sone attenuates early expression of three molecules associated with micro-glia/macrophages activation following rat traumatic brain injury. ActaNeuropathol 113:675–682.
184 Volume 62, No. 2




















