
Expression of calcium-binding proteins in the mouse claustrum
10
Expression of calcium-binding proteins in the mouse claustrum M a A ´ ngeles Real, Jose ´ Carlos Da ´ vila, Salvador Guirado * Departamento de Biologı ´a Celular, Gene ´tica y Fisiologı ´a, Universidad de Ma ´laga, Campus de Teatinos, 29071 Ma ´laga, Spain Received 12 July 2002; received in revised form 29 October 2002; accepted 1 December 2002 Abstract The present paper describes the distribution of three calcium-binding proteins (calbindin D28k, calretinin, and parvalbumin) in the mouse dorsal claustrum and endopiriform nucleus. The three calcium-binding proteins were distinctly expressed in structures of both the claustrum and the endopiriform nucleus. Calbindin was the calcium-binding protein showing the highest expression in the claustrum and the endopiriform nucleus. In contrast, calretinin-immunoreactive structures, particularly cell bodies, were very scarce in these regions. Both calbindin-immunoreactive and parvalbumin-immunoreactive neurons were more abundant in the claustrum than in the endopiriform nucleus, and more in rostral than in caudal levels. Nevertheless, calcium-binding protein immunoreactive neurons constitute a minority population of claustral neurons. The colocalization study of calbindin and parvalbumin immunoreactivities has demonstrated that both calcium-binding proteins are mostly expressed by separate claustral neurons in the mouse. On the other hand, our results on parvalbumin and calretinin immunoreactivity match a novel subdivision of the mouse claustrum mostly based on the pattern of cadherin expression [Neuroscience 106 (2001) 505]. In this sense, we propose that a specific zone of the dorsal claustrum with cell bodies that strongly express Rcad and cadherin-8 would be the selective target for parvalbumin-expressing fibers, and that they would be mostly avoided by calretinin-expressing axons. # 2003 Elsevier Science B.V. All rights reserved. Keywords: Calbindin; Parvalbumin; Calretinin; Lateral pallium; Ventral pallium 1. Introduction The claustrum is a pallial subcortical region present in all mammals. Two parts of the claustrum can be distinguished: the dorsal part is usually called claustrum proper and it is located deep to the insular cortex (therefore it is also named dorsal or insular claustrum); the ventral part is called endopiriform nucleus and is located deep to the piriform cortex (Druga, 1966; Sherk, 1988; Dinopoulos et al., 1992). Krettek and Price (1977) considered a ventral division of the endopiriform nucleus located deep to the periamygdaloid cortex and adjoining ventral part of the middle region of piriform cortex. For the sake of clarity, from now on the dorsal claustrum will be referred to as claustrum whereas the ventral claustrum will be referred to as endopiriform nucleus. The claustrum has extensive connections with the neocortex (Pearson et al., 1982; Macchi et al., 1983; Markowitsch et al., 1984; Li et al., 1986; Sloniewski et al., 1986; Sherk, 1988; Sadowski et al., 1997; Kowianski et al., 1998; Majak et al., 2000), whereas the endopiri- form nucleus is hodologicaly related with the prepiri- form and entorhinal cortices (Druga, 1971; Markowitsch et al., 1984; Witter et al., 1988). Despite the evidence for hodologicaly different zones of the claustrum, previous cyto and chemoarchitectonic studies show an overall uniform structure (Sherk, 1988; Reynhout and Baizer, 1999). Recently, however, three subdivisions within the claustrum have been distin- guished on the basis of specific cadherin expression patterns (Obst-Pernberg et al., 2001). It is not known whether these claustral subdivisions have different connections or play different functions. In this context, the expression of calcium-binding proteins has probed to be very useful in revealing subdivisions in different regions of the central nervous system, notably the thalamus (Jones and Hendry, 1989; Da ´ vila et al., 2000). Therefore, here we studied the * Corresponding author. Tel.: /34-952-13-1961; fax: /34-952-13- 2000. E-mail address: [email protected] (S. Guirado). Journal of Chemical Neuroanatomy 25 (2003) 151 /160 www.elsevier.com/locate/jchemneu 0891-0618/03/$ - see front matter # 2003 Elsevier Science B.V. All rights reserved. doi:10.1016/S0891-0618(02)00104-7
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Expression of calcium-binding proteins in the mouse claustrum

Expression of calcium-binding proteins in the mouse claustrum
Ma Angeles Real, Jose Carlos Davila, Salvador Guirado *
Departamento de Biologıa Celular, Genetica y Fisiologıa, Universidad de Malaga, Campus de Teatinos, 29071 Malaga, Spain
Received 12 July 2002; received in revised form 29 October 2002; accepted 1 December 2002
Abstract
The present paper describes the distribution of three calcium-binding proteins (calbindin D28k, calretinin, and parvalbumin) in
the mouse dorsal claustrum and endopiriform nucleus. The three calcium-binding proteins were distinctly expressed in structures of
both the claustrum and the endopiriform nucleus. Calbindin was the calcium-binding protein showing the highest expression in the
claustrum and the endopiriform nucleus. In contrast, calretinin-immunoreactive structures, particularly cell bodies, were very scarce
in these regions. Both calbindin-immunoreactive and parvalbumin-immunoreactive neurons were more abundant in the claustrum
than in the endopiriform nucleus, and more in rostral than in caudal levels. Nevertheless, calcium-binding protein immunoreactive
neurons constitute a minority population of claustral neurons. The colocalization study of calbindin and parvalbumin
immunoreactivities has demonstrated that both calcium-binding proteins are mostly expressed by separate claustral neurons in
the mouse. On the other hand, our results on parvalbumin and calretinin immunoreactivity match a novel subdivision of the mouse
claustrum mostly based on the pattern of cadherin expression [Neuroscience 106 (2001) 505]. In this sense, we propose that a specific
zone of the dorsal claustrum with cell bodies that strongly express Rcad and cadherin-8 would be the selective target for
parvalbumin-expressing fibers, and that they would be mostly avoided by calretinin-expressing axons.
# 2003 Elsevier Science B.V. All rights reserved.
Keywords: Calbindin; Parvalbumin; Calretinin; Lateral pallium; Ventral pallium
1. Introduction
The claustrum is a pallial subcortical region present in
all mammals. Two parts of the claustrum can be
distinguished: the dorsal part is usually called claustrum
proper and it is located deep to the insular cortex
(therefore it is also named dorsal or insular claustrum);
the ventral part is called endopiriform nucleus and is
located deep to the piriform cortex (Druga, 1966; Sherk,
1988; Dinopoulos et al., 1992). Krettek and Price (1977)
considered a ventral division of the endopiriform
nucleus located deep to the periamygdaloid cortex and
adjoining ventral part of the middle region of piriform
cortex. For the sake of clarity, from now on the dorsal
claustrum will be referred to as claustrum whereas the
ventral claustrum will be referred to as endopiriform
nucleus.
The claustrum has extensive connections with the
neocortex (Pearson et al., 1982; Macchi et al., 1983;
Markowitsch et al., 1984; Li et al., 1986; Sloniewski et
al., 1986; Sherk, 1988; Sadowski et al., 1997; Kowianski
et al., 1998; Majak et al., 2000), whereas the endopiri-
form nucleus is hodologicaly related with the prepiri-
form and entorhinal cortices (Druga, 1971;
Markowitsch et al., 1984; Witter et al., 1988).
Despite the evidence for hodologicaly different zones
of the claustrum, previous cyto and chemoarchitectonic
studies show an overall uniform structure (Sherk, 1988;
Reynhout and Baizer, 1999). Recently, however, three
subdivisions within the claustrum have been distin-
guished on the basis of specific cadherin expression
patterns (Obst-Pernberg et al., 2001). It is not known
whether these claustral subdivisions have different
connections or play different functions.
In this context, the expression of calcium-binding
proteins has probed to be very useful in revealing
subdivisions in different regions of the central nervous
system, notably the thalamus (Jones and Hendry, 1989;
Davila et al., 2000). Therefore, here we studied the
* Corresponding author. Tel.: �/34-952-13-1961; fax: �/34-952-13-
2000.
E-mail address: [email protected] (S. Guirado).
Journal of Chemical Neuroanatomy 25 (2003) 151�/160
www.elsevier.com/locate/jchemneu
0891-0618/03/$ - see front matter # 2003 Elsevier Science B.V. All rights reserved.
doi:10.1016/S0891-0618(02)00104-7

distribution of three calcium-binding proteins (calbindin
D28k, calretinin, and parvalbumin) in the mouse
claustrum searching for specific distribution patterns
putatively related either to hodologicaly-based subdivi-sions or cadherin expression patterns.
2. Material and methods
Fifteen adult OF1 mice (42�/46 g body weight) were
used in the present study. Throughout the experimental
work animals were treated according to the EuropeanCommunities Council directive (86/609/EEC) on treat-
ment of experimental animals.
Mice were deeply anesthetized with sodium pentothal
(65 mg/kg; Abbott Laboratories) and transcardially
perfused with 0.1 M phosphate-buffered saline (PBS),
pH 7.4, followed by 4% paraformaldehyde, 0.1%
glutaraldehyde and 0.2% picric acid in PBS at room
temperature for 30 min. The brains were then removedand stored in 4% paraformaldehyde and 0.2% picric acid
in PBS at 4 8C overnight; afterwards they were
embedded in 4% agar and cut into 50-mm-thick frontal
sections, using a vibratome. The sections were washed
extensively in PBS prior to immunocytochemical stain-
ing with the peroxidase�/antiperoxidase method.
Free-floating sections were first incubated in 2%
normal goat serum and 0.3% Triton X-100 in PBS atroom temperature for 1 h, to block nonspecific binding
of the antibodies and permeate the tissues, respectively,
and then were transferred to the primary antibody. The
three polyclonal antibodies, anti-parvalbumin, anti-
calretinin, and anti-calbindin, were raised in rabbits
(SWant, Bellinzona, Switzerland) and used at a dilution
of 1:2000 for 18 h. After three washes in PBS for 45 min,
the sections were incubated in goat anti-rabbit IgGdiluted 1:35 for 1 h, washed again in PBS for 45 min,
and incubated in peroxidase�/antiperoxidase diluted
1:100 for 1 h. The immunolabeling was revealed with
0.05% diaminobenzidine (DAB; Sigma Chemical Co, St.
Louis, MO), 0.05% nickel ammonium sulfate and 0.03%
hydrogen peroxide (H2O2) in PBS. All steps were carried
out at room temperature with gentle agitation. After a
thorough wash in PBS, the sections were mounted on
gelatinized slides, air dried, dehydrated in ethanol,
cleared in xylene and coverslipped with DPX (BDH,
Poole, UK).
In order to provide some quantitative data about the
populations of calcium-binding protein immunoreactive
neurons as well as the putative colocalization of these
proteins in the same neuron, we carried out post-
embedding immunocytochemistry on semithin sections
of the claustrum in one mouse. After fixation, brain
slices 500-mm-thick, obtained on a vibratome, were
collected in PBS, dehydrated in acetone, and flat
embedded in Araldite 502 (Sigma). Resin-embedded
slices containing the claustrum were glued onto poly-
merized blocks, trimmed and then sectioned on an
ultramicrotome. Series of three sections of 1.5 mm were
made. The first series was stained with toluidine blue,
whereas the second and third series were stained with
anti-calbindin and anti-parvalbumin, respectively. After
removing the resin with sodium ethoxide, sections were
processed according to an immunohistochemical proce-
dure similar to that described above, except for that the
primary antibodies were used at a higher concentration
(anti-CB: 1/1000; anti-PV: 1/500). To determine coex-
istence, we took into consideration only those cells
clearly positive to calbindin or parvalbumin and dis-
playing a wide nuclear profile.
Controls: as controls of the immunohistochemical
method used in the present study, sections were pro-
cessed as indicated but the corresponding primary
antiserum was replaced by rabbit nonimmune serum
(1:500). No immunostaining could be detected under
these conditions. In addition, as a control of the
specificity of the different primary antisera under the
experimental conditions used in this work, we incubated
control sections in the primary antibody preadsorbed
with the corresponding protein (1 mg/ml of the diluted
antibody). As a result, specific immunostaining was
completely abolished.
Fig. 1. Calbindin-immunoreactivity. (A) Panoramic view of the claustrum (Cl) and dorsal endopiriform nucleus (DEn) at a rostral level (see Fig. 17
of Paxinos and Franklin, 2001). Transverse section. Scale bar: 1 mm. (B) Detail of the claustrum and dorsal endopiriform nucleus at an intermediate
rostrocaudal level. The claustrum is characterized by the presence of a number of immunoreactive cells within a moderately immunostained neuropil,
similarly to the adjacent cortical areas and dorsal endopiriform nucleus. Scale bar: 500 mm. (C) Most neuronal cell bodies in the claustrum are devoid
of calbindin immunoreactivity and appear as white profiles surrounded by an immunostained neuropil. Scale bar: 100 mm. (D) Morphological
features of calbindin immunoreactive neurons. Most stained neurons are multipolar cells with aspiny dendrites. Scale bar: 100 mm. (E) Pairs of
calbindin immunostained neurons with apposed cell bodies are sometimes observed in the claustrum (arrow). A number of lightly stained somata can
also be observed. Scale bar: 100 mm. (F) Panoramic view of the endopiriform region at a caudal level (see Fig. 41 of Paxinos and Franklin, 2001).
Both the number of immunoreactive neurons and the intensity of neuropil staining decrease at these levels. Scale bar: 1 mm. aca, Anterior
commissure, anterior; AID, agranular insular cortex, dorsal; AIP, agranular insular cortex, posterior; AIV, agranular insular cortex, ventral; BLA,
basolateral amygdaloid nucleus, anterior; Cl, claustrum; CPu, caudate putamen; DEn, dorsal endopiriform nucleus; DI, dysgranular insular cortex;
ec, external capsule; La, lateral amygdaloid nucleus; lo, lateral olfactory tract; Pir, piriform cortex; Ven, ventral endopiriform nucleus.
M.A. Real et al. / Journal of Chemical Neuroanatomy 25 (2003) 151�/160152

Fig. 1
M.A. Real et al. / Journal of Chemical Neuroanatomy 25 (2003) 151�/160 153

Light microscopic images were photographed by
using a Leica microscope equipped with a Nikon
DXM1200 digital camera. Digital images were loaded
into ADOBE PHOTOSHOP software and converted tograyscale images. Brightness and contrast were adjusted
for the final images. No additional filtering or manip-
ulation of the images was performed. The final figures
were composed and labeled with ADOBE PAGEMAKER
software and printed with an Epson Photo 750 printer.
3. Results
The three calcium-binding proteins studied in this
work were distinctly expressed in structures of both theclaustrum and the endopiriform nucleus. We will next
describe separately calbindin-immunoreactivity in the
claustrum and in the endopiriform nucleus, and then
calretinin- and parvalbumin-immunoreactivities, respec-
tively.
3.1. Calbindin-immunoreactivity
3.1.1. Claustrum
Sections immunostained for calbindin showed an
overall stronger staining of both neurons and neuropil
than those stained for either of the other antibodies;
accordingly numerous calbindin-immunoreactive (ir)neurons embedded in a moderately immunostained
neuropil were found from rostral to caudal levels of
the claustrum (Fig. 1A and B). Nevertheless, the number
of immunostained cells decreased slightly at caudal
levels. As in other parts of the telencephalon, two cell-
staining patterns for calbindin were observed through-
out the claustrum: darkly stained neurons with a Golgi-
like appearance, and lightly stained cell bodies withbarely visible processes (Fig. 1D and E). Cell morphol-
ogies varied from small, round or elongated neurons to
medium-sized multipolar neurons (Fig. 1C and D).
These distinct cell types appeared intermingled through-
out the claustrum. Multipolar calbindin-ir neurons
displayed typically three to five processes splitting into
a few secondary aspiny dendrites. Sometimes, pairs of
calbindin-ir multipolar neurons with apposed somatawere observed within the claustrum (Fig. 1E).
The uniform background staining of the claustrum
and adjacent cortical areas made it difficult to delineate
the borders of the claustrum in calbindin immunos-
tained sections (Fig. 1B). A moderately stained neuropil
extended over the whole anterior�/posterior extent of the
claustrum. Neuropil staining consisted of varicose axons
oriented in all directions and puncta surroundingnumerous immunonegative cell profiles (Fig. 1C).
Although many perisomatic calbindin-positive terminals
were observed, they did not form pericellular baskets.
3.1.2. Endopiriform nucleus
As in the claustrum, calbindin-ir neurons in the
endopiriform nucleus exhibited a variety of staining
intensities, sizes, and morphologies. Small or medium-sized multipolar cells were the most common types,
although pyramidal-shaped cell bodies were also found
at intermediate levels of the dorsal endopiriform nu-
cleus. In the dorsal endopiriform nucleus, the number of
calbindin immunostained neurons was highest at rostral
levels (Fig. 1A), decreasing at intermediate and caudal
levels (Fig. 1F). On the other hand, the number of
calbindin immunoreactive neurons in the ventral en-dopiriform nucleus was similar to that found in the
caudal part of the dorsal endopiriform nucleus. Pairs of
calbindin immunostained cells with apposed somata
were also found in the endopiriform nucleus.
A uniform, moderately stained, neuropil extended
throughout both the dorsal and ventral regions of the
endopiriform nucleus (Fig. 1F). Calbindin-positive ax-
ons bearing varicosities were observed in this neuropil.
3.2. Calretinin-immunoreactivity
3.2.1. Claustrum
Calretinin immunostaining in the claustrum consistedof a few scattered positive cells against a background of
stained fibers and boutons. Most calretinin-ir neurons
were densely stained and exhibited small, round or
elongated cell bodies (Fig. 2C), from which two or three
aspiny dendrites arose. Some immunostained bipolar
neurons located near the external capsule had their cell
bodies and dendrites oriented parallel to it whereas
other calretinin-ir neurons extended their dendrites,perpendicular to the external capsule and to the cortical
surface.
In contrast to the piriform cortex and to the subjacent
striatum, the claustrum is characterized by the presence
of a dense calretinin-ir neuropil consisting of both
stained fibers and puncta (Fig. 2A and B). Nevertheless,
the neuropil immunostaining was unevenly distributed
within the claustrum. A calretinin-negative neuropilzone, virtually devoid of immunoreactive fibers or
puncta, appeared as an oval region in the core of the
claustrum (Fig. 2B and C). This oval region was also
largely devoid of calretinin-ir neurons, which, if present,
were found at the periphery of the region (Fig. 2C). The
calretinin-negative region was clearly visible from inter-
mediate to caudal levels of the claustrum since it was
surrounded by immunopositive structures, including athin deep layer of neuropil that separates it from the
fibers of the external capsule (Fig. 2C), and a superficial
layer of neuropil continuous with the deep layers of the
neighboring insular cortex.
M.A. Real et al. / Journal of Chemical Neuroanatomy 25 (2003) 151�/160154

3.2.2. Endopiriform nucleus
The overall calretinin immunostaining in the endo-
piriform nucleus was similar to that of the claustrum
and the insular cortex. Few scattered neurons embedded
in a moderate-to-dense immunoreactive neuropil were
found in the endopiriform nucleus. The most abundant
calretinin-ir cell type consisted of small bipolar cells with
the elongated cell body and processes oriented parallel
to the external capsule. Bipolar neurons were observed
in both the dorsal and ventral endopiriform nuclei.
Neuropil staining was highest at intermediate levels in
the region of the dorsal endopiriform nucleus adjacent
to the claustrum (Fig. 2B). For the rest of the dorsal
endopiriform nucleus and the whole ventral endopiri-
Fig. 2. Calretinin-immunoreactivity. (A) Transverse section at a rostral level of the mouse telencephalon (see Fig. 16 of Paxinos and Franklin, 2001)
showing the claustrum (Cl) and the dorsal endopiriform nucleus (DEn). Scale bar: 1 mm. (B) At intermediate rostrocaudal levels (see Fig. 31 of
Paxinos and Franklin, 2001), the claustrum, the dorsal endopiriform nucleus, and the insular cortex are characterized by a moderately
immunoreactive neuropil. An oval immunonegative area can be recognized within the claustrum. Scale bar: 1 mm. (C) Detail of the boxed area in B.
This oval area is virtually devoid of immunoreactive fibers. A small stained neuron can be observed in the periphery of the negative region (arrow).
Scale bar: 100 mm. (D) Panoramic view of the endopiriform region at a caudal level (see Fig. 41 of Paxinos and Franklin, 2001). Scale bar: 1 mm. aca,
anterior commissure, anterior; acp, anterior commissure, posterior; AID, agranular insular cortex, dorsal; AIP, agranular insular cortex, posterior;
AIV, agranular insular cortex, ventral; BLA, basolateral amygdaloid nucleus, anterior; Cl, claustrum; CPu, caudate putamen; DEn, dorsal
endopiriform nucleus; DI, dysgranular insular cortex; ec, external capsule; GI, granular insular cortex; La, lateral amygdaloid nucleus; Pir, piriform
cortex; VEn, ventral endopiriform nucleus.
M.A. Real et al. / Journal of Chemical Neuroanatomy 25 (2003) 151�/160 155

form nucleus, the neuropil was moderately immunos-
tained for calretinin (Fig. 2D). Calretinin immunoreac-
tive axons were fine and displayed small varicosities.
3.3. Parvalbumin-immunoreactivity
3.3.1. Claustrum
Parvalbumin immunostaining in the claustrum con-
sisted of a number of immunoreactive neurons sur-
rounded by a dense meshwork of stained fibers andpuncta (Fig. 3C and D). Both darkly and lightly stained
neurons were found throughout the claustrum but they
were less abundant at the caudal part of the claustrum.
Most parvalbumin-ir cells were medium in size and had
multipolar cell bodies (Fig. 3C) with several thin, beaded
dendrites that extended for long distances.
Parvalbumin-ir varicose axons were oriented in all
directions within the claustrum (Fig. 3C and D).However, it was unclear whether these axons formed
perisomatic contacts.
In sections immunostained for parvalbumin, the
claustrum was characterized by the presence of a sharply
defined, moderately stained patch of neuropil particu-
larly evident at intermediate and caudal levels of the
claustrum (Fig. 3B). The deep layers of the adjacent
agranular insular cortex presented also a patch ofmoderately stained neuropil, and between these two
neuropil patches there was an intervening immunone-
gative cell-poor zone that was most apparent at inter-
mediate levels of the claustrum (Fig. 3B).
The patch of parvalbumin immunopositive neuropil
in the claustrum roughly corresponds to the calretinin-
negative oval area described above.
3.3.2. Endopiriform nucleus
A number of parvalbumin-ir neurons embedded in aplexus of immunostained varicose axons and processes
were found at rostral levels of the dorsal endopiriform
nucleus (Fig. 3A). At intermediate and caudal levels of
the dorsal endopiriform nucleus, the number of positive
neurons and axons decreased dramatically: very few
immunoreactive neurons were observed, and the neuro-
pil was almost negative at these levels (Fig. 3E). The
parvalbumin immunoreactivity pattern in the ventral
endopiriform nucleus was similar to that of the caudal
part of the dorsal endopiriform nucleus.
Most parvalbumin stained neurons located in the
rostral part of the dorsal endopiriform nucleus were
medium in size and exhibited multipolar morphologies,
without a specific orientation of their cell processes. In
the ventral endopiriform nucleus, some multipolar
immunoreactive neurons with the cell body and main
dendrites oriented parallel to the pial surface were
observed (Fig. 3F).
3.4. Colocalization study
We carried out a colocalization study for the two
major calcium-binding proteins present in neurons of
the mouse claustrum, i.e. calbindin and parvalbumin,
using post-embedding immunocytochemistry on adja-
cent semithin sections. The analysis was made in the
dorsal claustrum, since it was the claustral region with
more positive neurons for either calcium-binding pro-
tein.
In addition, by comparing immunostained semithin
sections with the adjacent Nissl-stained semithin section,
we obtained quantitative data about the relative density
of calbindin- and parvalbumin-immunoreactive popula-
tions.
Calbindin- and parvalbumin-immunoreactive neu-
rons constituted a minority of the claustral neurons,
accounting for only 5.4% (n�/122) and 7.9% (n�/178)
of the total number of counted cells (n�/2260), respec-
tively. Neurons expressing either of the two proteins
represent 12.3% of the claustral cells. In addition,
calbindin- and parvalbumin-immunoreactive cells are
mostly segregated populations (Fig. 4): more than 87%
of parvalbumin-ir cells do not express calbindin (156 out
of 178) and, conversely, 82% of calbindin-ir cells do not
express parvalbumin (100 out of 122), whereas only 22
out of 2260 cells coexpress both proteins.
Fig. 3. Parvalbumin-immunoreactivity. (A) Low power photomicrograph of a transverse section of the rostral telencephalon (see Fig. 17 of Paxinos
and Franklin, 2001). At this level, the dorsal endopiriform nucleus (DEn) presents its highest number of immunoreactive neurons. Scale bar: 1 mm.
(B) Detail of the claustrum at an intermediate level. A central region with a moderately stained neuropil, just deep to the ventral part of the agranular
insular cortex can be easily recognized (arrows). Scale bar: 500 mm. (C) Two multipolar medium-size neurons can be observed in the central region of
the claustrum. Scale bar: 100 mm. (D) Neuropil immunostaining consists of a dense meshwork of axons and terminals, although no pericellular
baskets can be distinguished. Scale bar: 50 mm. (E) Detail of the endopiriform region at a caudal level (see Fig. 38 of Paxinos and Franklin, 2001).
Both the dorsal and ventral parts of the endopiriform nucleus are characterized by an immunonegative neuropil. Scale bar: 1 mm. (F) Multipolar
stained neurons with their elongated cell bodies and dendrites oriented parallel to the pial surface are observed in the ventral endopiriform nucleus
(VEn). Scale bar: 100 mm. aca, Anterior commissure, anterior; AID, agranular insular cortex, dorsal; AIP, agranular insular cortex, posterior; AIV,
agranular insular cortex, ventral; BLA, basolateral amygdaloid nucleus, anterior; Cl, claustrum; CPu, caudate putamen; DEn, dorsal endopiriform
nucleus; DI, dysgranular insular cortex; ec, external capsule; La, lateral amygdaloid nucleus; lo, lateral olfactory tract; Pir, piriform cortex; VEn,
ventral endopiriform nucleus.
M.A. Real et al. / Journal of Chemical Neuroanatomy 25 (2003) 151�/160156
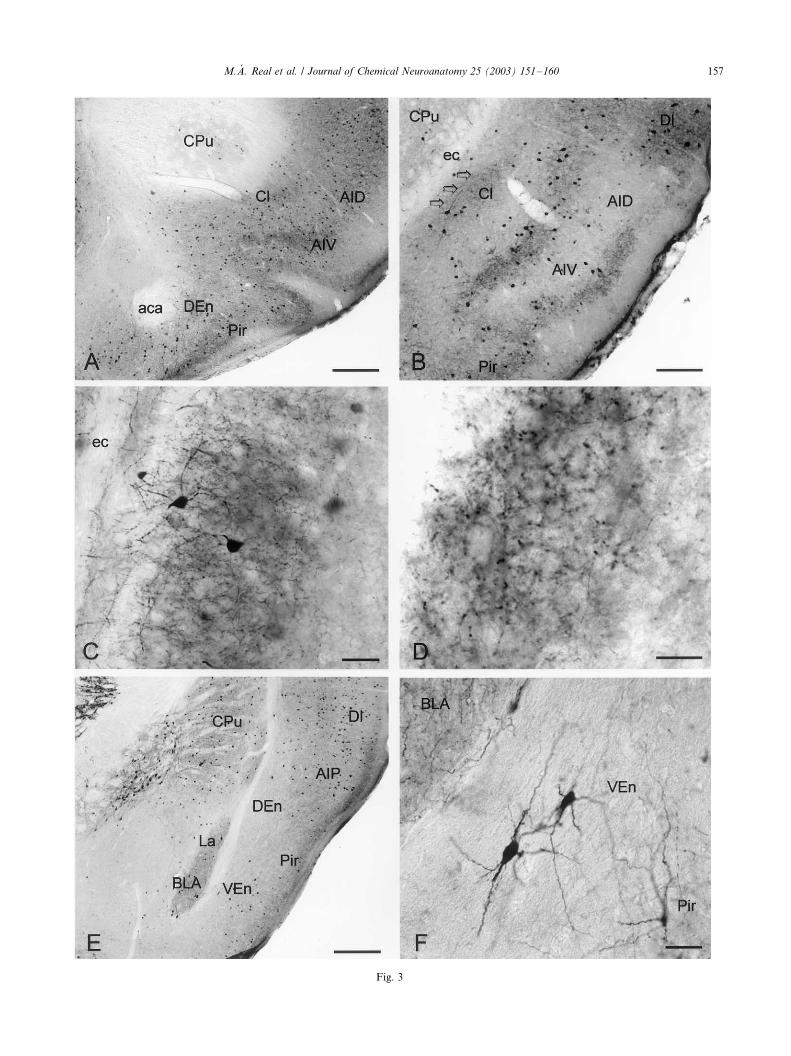
Fig. 3
M.A. Real et al. / Journal of Chemical Neuroanatomy 25 (2003) 151�/160 157

4. Discussion
Although little is known about the exact functions of
the calcium-binding proteins parvalbumin, calbindinand calretinin, they have provided useful markers of
specific neuronal subpopulations in studies of the
neuronal circuitry of the cerebral cortex and other brain
regions, and they reveal chemoarchitectonic subdivi-
sions in different regions of the central nervous system.
Results of the present study indicate the existence of
distinct immunoreactivity patterns for each of the three
calcium-binding proteins in both the claustrum and theendopiriform nucleus as it has been reported in other
mammals, and also reveal details not described pre-
viously. We will next discuss our results in comparison
with other mammals and then analyze the calcium-
binding protein expression patterns in the light of
recently described subdivisions within the mouse claus-
trum.
Calbindin and parvalbumin immunoreactivities havebeen reported in the claustrum and endopiriform
nucleus in other mammals but inter-specific differences
exist regarding the expression pattern of each protein. In
the rat, parvalbumin and calbindin exhibit a largely
complementary distribution pattern: parvalbumin-im-
munoreactivity is concentrated in the claustrum,
whereas calbindin-immunoreactivity prevails in the en-
dopiriform nucleus (Celio, 1990; Druga et al., 1993).From our results in mouse, we cannot demonstrate this
complementary distribution pattern between calbindin
and parvalbumin; instead, we showed that for either of
these calcium-binding proteins, more immunostained
neurons were observed in the claustrum than in the
endopiriform nucleus, and more in rostral than in
caudal levels.
Our colocalization analysis has demonstrated that, inspite of a similar distribution pattern, parvalbumin and
calbindin are mostly expressed by separate claustral
neurons in the mouse. In this context, is to be noted that
in the rat cortex parvalbumin- and calbindin-ir cells
represent two classes of GABAergic interneurons dis-
playing morphological and neurochemical specificity
(Celio, 1986; Hendry et al., 1989; Kosaka and Heiz-
mann, 1989), and indirect evidence suggests that cal-cium-binding protein immunoreactive neurons may also
represent, at least in part, inhibitory local circuit
neurons in the rat claustrum (Druga et al., 1993).
With regards to calretinin, it has been claimed that the
rat claustrum is demarcated by the absence of calretinin
staining (Paxinos et al., 1998). However, calretinin
immunoreactive structures have been reported in the
monkey claustrum (Reynhout and Baizer, 1999) and inthe endopiriform nucleus of the guinea pig (Frassoni et
al., 1998). Our results in the mouse showed that
calretinin staining consisted of a few positive cell bodies
embedded in a moderate neuropil both in the claustrum
and the endopiriform nucleus. Calretinin is present in
the claustral regions of monkeys, guinea pigs, and mice.
Thus, the reported absence of calretinin in the rat
claustrum could be due to inter-specific differences orto a differential sensitivity of the immunocytochemical
method.
Calcium-binding protein immunoreactive neurons
constitute together a small subset (12.3%) of the
claustral neurons in the mouse, as demonstrated by
our study on semithin sections. This low incidence of
calcium-binding protein immunoreactive neurons in the
claustrum was to be expected if they represent differentsubpopulations of GABAergic interneurons, as sug-
gested above, since GABAergic neurons in the rabbit
claustrum accounted only for 12% of all neurons in the
same claustral samples (Gomez-Urquijo et al., 2000).
These data on the low incidence of inhibitory claustral
neurons are also in agreement with the presence in the
rat claustrum of numerous latexin-immunoreactive
neurons (Arimatsu et al., 1992), most of which areglutamate positive (and GABA negative), suggesting
that they are excitatory projection neurons (Arimatsu et
al., 1999).
4.1. Different zones within the dorsal claustrum
Recently, three novel subdivisions within the mouse
claustrum based on the pattern of cadherin expression
and cytoarchitecture have been distinguished (Obst-Pernberg et al., 2001): a superior, an intermediate and
an inferior part. The superior and intermediate parts are
located deep to the insular cortex whereas the inferior
part is located deep to the dorsal part of the piriform
cortex. Cytoarchitectonically the intermediate part is
characterized by the presence of cell aggregates. The
superior and intermediate parts contain densely packed
cell bodies that strongly express R-cadherin (Rcad;Obst-Pernberg et al., 2001) and cadherin-8 mRNA
(Korematsu and Redies, 1997), whereas the inferior
part shows weak expression of Rcad, and only contain
scattered cell bodies expressing cadherin-8 mRNA. In
addition, the superior part shows a very strongly
immunoreactive neuropil for Rcad and moderately
labeled for cadherin-N (Ncad); the intermediate part is
strongly immunoreactive for Rcad and moderately tostrongly labeled for Ncad; and the inferior part is
weakly labeled for Rcad and moderately labeled for
Ncad (Obst-Pernberg et al., 2001).
Our results on parvalbumin and calretinin immunor-
eactivity match these novel subdivisions of the mouse
claustrum. Thus, in parvalbumin-immunostained sec-
tions a patch of moderately immunoreactive neuropil
occupies a position just deep to the ventral part of theagranular insular cortex. This parvalbumin positive
zone is bordered dorsal and ventrally by claustral zones
virtually devoid of immunostained neuropil. The dorsal
M.A. Real et al. / Journal of Chemical Neuroanatomy 25 (2003) 151�/160158

negative zone lies deep to the dorsal part of the
agranular insular cortex and to the dysgranular cortex,
whereas the ventral zone negative for parvalbumin is
continuous with the endopiriform nucleus. In calretinin
immunostained sections, on the other hand, a comple-
mentary pattern was observed: the intermediate region
was virtually devoid of calretinin immunoreactive fibers,
and dorsal and ventral calretinin-positive zones bor-
dered it. It is tempting to relate these three zones of the
dorsal claustrum showing a distinct calcium-binding
protein expression pattern to the three subdivisions
described by Obst-Pernberg and coworkers on the basis
of cadherin expression patterns (Obst-Pernberg et al.,
2001). These authors suggest that the selective adhesion
of neural structures that express the same types of
cadherin contribute to the formation of gray matter
areas, neural circuits and functional connections in the
postnatal forebrain of the mouse. In this sense, we
propose that the cell aggregates of the intermediate zone
of the dorsal claustrum with cell bodies that strongly
express Rcad and cadherin-8 would be the selective
target for parvalbumin-expressing fibers, and that they
would be mostly avoided by calretinin-expressing axons.
We cannot conclude whether these fibers have an
intrinsic or extrinsic origin.
While these novel subdivisions of the mouse dorsal
claustrum appear to be well documented from a cyto
and neurochemical point of view, it is difficult to relate
them to the different functional zones that have been
described in the mammalian claustrum on the basis of its
connections. In mammals with a well developed claus-
trum the claustro-neocortical connections are topogra-
phically organized, with an anterior part of the
claustrum linked mainly with motor and prefrontal
cortices, a central part linked with somatosensory
cortex, a posterior claustrum related to visual cortex,
and a ventral zone connected with auditory cortex
(Pearson et al., 1982; Macchi et al., 1983; Sherk, 1988;
Morys et al., 1996). In the rat claustrum, however, two
main cortico-related zones have been described, an
anterodorsal sensorimotor and a posteroventral vi-
suoauditory zones (Sadowski et al., 1997).
Our results indicate that the incidence of calcium-
binding protein-expressing neurons is higher in rostral
than caudal levels of the mouse dorsal claustrum. Since
all hodological studies confirm at least two different
functional zones in the claustrum, an anterior and a
posterior region, it is likely that calcium-binding pro-
tein-expressing neurons, putatively GABAergic cells, are
more represented in claustral intrinsic circuits related
with the limbic and motor cortices rather than with the
visuoauditory regions. As suggested for somatostatin-,
neuropeptide Y-, and vasoactive intestinal peptide-ir
neurons in the rat claustrum (Kowianski et al., 2001),
calcium-binding protein-expressing neurons do not
appear to play a significant role in the claustro-cortical
projection but are most probably involved in modula-
tion and information transfer in the claustrum.
It is to be noted that the complementary expression of
calretinin and parvalbumin in patches of neuropil in the
mouse claustrum is best seen at intermediate/posterior
levels in the rostrocaudal axis of the claustrum. It
Fig. 4. Photomicrographs of two adjacent semithin sections immunostained for calbindin (A) and parvalbumin (B). Only one neuron in this field is
immunoreactive for both calcium-binding proteins (arrow). Capillaries (c) were used as landmarks. Scale bar: 30 mm.
M.A. Real et al. / Journal of Chemical Neuroanatomy 25 (2003) 151�/160 159

cannot be determined, however, whether these claustral
zones correspond with claustral regions with specific
cortical connections. In this sense, it would be of interest
to perform combined immunohistochemical-fiber tra-cing experiments in order to study the specific connec-
tions of the chemoarchitectonic subdivisions of the
dorsal claustrum.
Acknowledgements
We would like to thank Luis Olmos for his excellent
technical assistance. This work was supported bySpanish DGI grant BFI2000-1359-C02-01, as well as
by Spanish FIS grant 01-0057-01.
References
Arimatsu, Y., Miyamoto, M., Nihonmatsu, I., Hirata, K., Uratani, Y.,
Hatanaka, Y., Takiguchi-Hayashi, K., 1992. Early regional speci-
fication for a molecular neuronal phenotype in the rat neocortex.
Proc. Natl. Acad. Sci. USA 89, 8879�/8883.
Arimatsu, Y., Ishida, M., Sato, M., Kojima, M., 1999. Corticocortical
associative neurons expressing latexin: specific cortical connectivity
formed in vivo and in vitro. Cereb. Cortex 9, 569�/576.
Celio, M.R., 1986. Parvalbumin in most gamma-aminobutyric acid-
containing neurons of the rat cerebral cortex. Science 231, 995�/
997.
Celio, M.R., 1990. Calbindin D-28k and parvalbumin in the rat
nervous system. Neuroscience 35, 375�/475.
Davila, J.C., Guirado, S., Puelles, L., 2000. Expression of calcium-
binding proteins in the diencephalon of the lizard Psammodromus
algirus . J. Comp. Neurol. 427, 67�/92.
Dinopoulos, A., Papadopoulos, G.C., Michaloudi, H., Parnavelas,
J.G., Uylings, H.B., Karamanlidis, A.N., 1992. Claustrum in the
hedgehog (Erinaceus europaeus ) brain: cytoarchitecture and con-
nections with cortical and subcortical structures. J. Comp. Neurol.
316, 187�/205.
Druga, R., 1966. The claustrum of the cat (Felis domestica ). Folia
Morphol. (Praha) 14, 7�/16.
Druga, R., 1971. Projection of prepyriform cortex into claustrum.
Folia Morphol. (Praha) 19, 405�/410.
Druga, R., Chen, S., Bentivoglio, M., 1993. Parvalbumin and
calbindin in the rat claustrum: an immunocytochemical study
combined with retrograde tracing frontoparietal cortex. J. Chem.
Neuroanat. 6, 399�/406.
Frassoni, C., Radici, C., Spreafico, R., de Curtis, M., 1998. Calcium-
binding protein immunoreactivity in the piriform cortex of the
guinea-pig: selective staining of subsets of non-GABAergic neurons
by calretinin. Neuroscience 83, 229�/237.
Gomez-Urquijo, S.M., Gutierrez-Ibarluzea, I., Bueno-Lopez, J.L.,
Reblet, C., 2000. Percentage incidence of gamma-aminobutyric
acid neurons in the claustrum of the rabbit and comparison with
the cortex and putamen. Neurosci. Lett. 282, 177�/180.
Hendry, S.H., Jones, E.G., Emson, P.C., Lawson, D.E., Heizmann,
C.W., Streit, P., 1989. Two classes of cortical GABA neurons
defined by differential calcium binding protein immunoreactivities.
Exp. Brain Res. 76, 467�/472.
Jones, E.G., Hendry, S.H.C., 1989. Differential calcium-binding
protein immunoreactivity distinguishes classes of relay neurons in
monkey thalamic nuclei. Eur. J. Neurosci. 1, 222�/245.
Korematsu, K., Redies, C., 1997. Restricted expression of cadherin-8
in segmental and functional subdivisions of the embryonic mouse
brain. Dev. Dynam. 208, 178�/189.
Kosaka, T., Heizmann, C.W., 1989. Selective staining of a population
of parvalbumin-containing GABAergic neurons in the rat cerebral
cortex by lectins with specific affinity for terminal N -acetylgalac-
tosamine. Brain Res. 483, 158�/163.
Kowianski, P., Morys, J., Karwacki, Z., Dziewiatkowski, J., Narkie-
wicz, O., 1998. The cortico-related zones of the rabbit claustrum-
study of the claustrocortical connections based on the retrograde
axonal transport of fluorescent tracers. Brain Res. 784, 199�/209.
Kowianski, P., Timmermans, J.P., Morys, J., 2001. Differentiation in
the immunocytochemical features of intrinsic and cortically
projecting neurons in the rat claustrum*/combined immunocyto-
chemical and axonal transport study. Brain Res. 905, 63�/71.
Krettek, J.E., Price, J.L., 1977. Projections from the amygdaloid
complex to the cerebral cortex and thalamus in the rat and cat. J.
Comp. Neurol. 172, 687�/722.
Li, Z.K., Takada, M., Hattori, T., 1986. Topographic organization
and collateralization of claustrocortical projections in the rat. Brain
Res. Bull. 17, 529�/532.
Macchi, G., Bentivoglio, M., Minciacchi, D., Molinari, M., 1983.
Claustroneocortical projections studied in the cat by means of
multiple retrograde fluorescent tracing. J. Comp. Neurol. 215,
121�/134.
Majak, K., Kowianski, P., Morys, J., Spodnik, J., Karwacki, Z.,
Wisniewski, H.M., 2000. The limbic zone of the rabbit and rat
claustrum: a study of the claustrocingulate connections based on
the retrograde axonal transport of fluorescent tracers. Anat.
Embryol. 201, 15�/25.
Markowitsch, H.J., Irle, E., Bang-Olsen, R., Flindt-Egebak, P., 1984.
Claustral efferents to the cat’s limbic cortex studied with retrograde
and anterograde tracing techniques. Neuroscience 12, 409�/425.
Morys, J., Berdel, B., Maciejewska, B., Sadowski, M., Sidorowicz, M.,
Kowianska, J., Narkiewicz, O., 1996. Division of the human
claustrum according to its architectonics, morphometric para-
meters and cortical connections. Folia Morphol. (Warsz) 55, 69�/
82.
Obst-Pernberg, K., Medina, L., Redies, C., 2001. Expression of R -
cadherin and N -cadherin by cell groups and fiber tracts in the
developing mouse forebrain: relation to the formation of functional
circuits. Neuroscience 106, 505�/533.
Paxinos, G., Franklin, K.B.J., 2001. The Mouse Brain in Stereotaxic
Coordinates. Academic Press, New York.
Paxinos, G., Kus, L., Ashwell, K.W.S., Watson, C., 1998. Chemoarch-
itectonic Atlas of the Rat Forebrain. Academic Press, New York.
Pearson, R.C., Brodal, P., Gatter, K.C., Powell, T.P., 1982. The
organization of the connections between the cortex and the
claustrum in the monkey. Brain Res. 234, 435�/441.
Reynhout, K., Baizer, J.S., 1999. Immunoreactivity for calcium-
binding proteins in the claustrum of the monkey. Anat. Embryol.
199, 75�/83.
Sadowski, M., Morys, J., Jakubowska-Sadowska, K., Narkiewicz, O.,
1997. Rat’s claustrum shows two main cortico-related zones. Brain
Res. 756, 147�/152.
Sherk, H., 1988. The claustrum and the cerebral cortex. In: Jones,
E.G., Peters, A. (Eds.), Sensory-Motor Areas and Aspects of
Cortical Connectivity, vol. 5. Plenum Press, New York, London,
pp. 467�/499.
Sloniewski, P., Usunoff, K.G., Pilgrim, C., 1986. Retrograde transport
of fluorescent tracers reveals extensive ipsi- and contralateral
claustrocortical connections in the rat. J. Comp. Neurol. 246,
467�/477.
Witter, M.P., Room, P., Groenewegen, H.J., Lohman, A.H., 1988.
Reciprocal connections of the insular and piriform claustrum with
limbic cortex: an anatomical study in the cat. Neuroscience 24,
519�/539.
M.A. Real et al. / Journal of Chemical Neuroanatomy 25 (2003) 151�/160160




















