
Effects of water quality and hydrologic drivers on periphyton colonization on Sparganium erectum in...
11
Effects of water quality and hydrologic drivers on periphyton colonization on Sparganium erectum in two Turkish lakes with different mixing regimes Meriç Albay & Reyhan Akçaalan Received: 27 July 2007 / Accepted: 6 November 2007 / Published online: 10 January 2008 # Springer Science + Business Media B.V. 2007 Abstract The aim of this paper is to determine and compare the environmental factors controlling vertical colonisation of periphyton on Sparganium erectum in a shallow eutrophic turbid lake, Manyas Lake, and an oligo-mesotrophic deep lake, Sapanca Lake, Turkey during the July 1997–November 1998. To investigate the effect of the environmental factors on periphyton colonization on S. erectum, the stem was cut above the rhizomes and subdivided into three equal sections. Multivariate statistical analyses have been applied to clarify relationships between environmental vari- ables and periphyton colonization on S. erectum. Results indicated that physical disturbance and trophic level of the lakes influenced the colonization of the periphyton. Among the measured parameters, low light intensity, total suspended solids, tempera- ture and water level fluctuation were observed as driving factors in Manyas Lake whereas nutrient deficiency was found as key factor in Sapanca Lake. The zonation of the periphyton, density, composition and dominant/subdominant taxa were significantly different in these lakes. However, Oedogonium sp., Mougeotia sp., Cylindrocapsa sp., Cladophora glom- erata (Linn.), Aulacoseira italica (Ehr.) Simonsen, Melosira varians C. Agardh, Navicula tripunctata (O.F. Müller) Bory and Fragilaria ulna (Nitzsch) Lange-Bertalot were found as dominant species at all sections of S. erectum both in Manyas Lake and Sapanca Lake. It can be thought that these species have a broad range of tolerance to several physical, chemical and hydrologic disturbances. This is the first study to introduce how much the water quality and hydrologic drivers have affected vertical coloni- zation of periphyton on S. erectum in two lakes with different mixing regimes. Keywords Periphyton colonization . Biomonitoring . Manyas lake . Sapanca lake . Sparganium erectum . Water quality Introduction The littoral zone is regarded very complex area both of deep and shallow lakes. Studies show that the littoral region provides almost entire production of the lakes (Meulemans 1998; Kiss et al. 2003), as an important food source for invertebrates and fish. The periphyton is the most important component with multiple interactions among the organisms in this system. It provides high level of primary productivity and a major regulator of nutrients fluxes. Besides lake trophic status, the quantity of the periphyton produc- tion depends on the multiple biotic and abiotic factors Environ Monit Assess (2008) 146:171–181 DOI 10.1007/s10661-007-0069-5 M. Albay (*) : R. Akçaalan Faculty of Fisheries, Istanbul University, Ordu Cad. No: 200 Laleli, 34470 Istanbul, Turkey e-mail: [email protected]
Transcript of Effects of water quality and hydrologic drivers on periphyton colonization on Sparganium erectum in...

Effects of water quality and hydrologic driverson periphyton colonization on Sparganium erectumin two Turkish lakes with different mixing regimes
Meriç Albay & Reyhan Akçaalan
Received: 27 July 2007 /Accepted: 6 November 2007 / Published online: 10 January 2008# Springer Science + Business Media B.V. 2007
Abstract The aim of this paper is to determine andcompare the environmental factors controlling verticalcolonisation of periphyton on Sparganium erectum ina shallow eutrophic turbid lake, Manyas Lake, and anoligo-mesotrophic deep lake, Sapanca Lake, Turkeyduring the July 1997–November 1998. To investigatethe effect of the environmental factors on periphytoncolonization on S. erectum, the stem was cut abovethe rhizomes and subdivided into three equal sections.Multivariate statistical analyses have been applied toclarify relationships between environmental vari-ables and periphyton colonization on S. erectum.Results indicated that physical disturbance andtrophic level of the lakes influenced the colonizationof the periphyton. Among the measured parameters,low light intensity, total suspended solids, tempera-ture and water level fluctuation were observed asdriving factors in Manyas Lake whereas nutrientdeficiency was found as key factor in Sapanca Lake.The zonation of the periphyton, density, compositionand dominant/subdominant taxa were significantlydifferent in these lakes. However, Oedogonium sp.,Mougeotia sp., Cylindrocapsa sp., Cladophora glom-erata (Linn.), Aulacoseira italica (Ehr.) Simonsen,
Melosira varians C. Agardh, Navicula tripunctata(O.F. Müller) Bory and Fragilaria ulna (Nitzsch)Lange-Bertalot were found as dominant species at allsections of S. erectum both in Manyas Lake andSapanca Lake. It can be thought that these specieshave a broad range of tolerance to several physical,chemical and hydrologic disturbances. This is thefirst study to introduce how much the water qualityand hydrologic drivers have affected vertical coloni-zation of periphyton on S. erectum in two lakes withdifferent mixing regimes.
Keywords Periphyton colonization . Biomonitoring .
Manyas lake . Sapanca lake . Sparganium erectum .
Water quality
Introduction
The littoral zone is regarded very complex area bothof deep and shallow lakes. Studies show that thelittoral region provides almost entire production of thelakes (Meulemans 1998; Kiss et al. 2003), as animportant food source for invertebrates and fish. Theperiphyton is the most important component withmultiple interactions among the organisms in thissystem. It provides high level of primary productivityand a major regulator of nutrients fluxes. Besides laketrophic status, the quantity of the periphyton produc-tion depends on the multiple biotic and abiotic factors
Environ Monit Assess (2008) 146:171–181DOI 10.1007/s10661-007-0069-5
M. Albay (*) : R. AkçaalanFaculty of Fisheries, Istanbul University,Ordu Cad. No: 200 Laleli,34470 Istanbul, Turkeye-mail: [email protected]

including light intensity, temperature, water levelfluctuation, type of the substratum, wave actions,biotic interaction, intra- and inter-specific competi-tion, grazing, host plant architecture etc. Quantity ofthe periphyton and species richness may closelyreflect the trophic state of lakes (Scheffer 1998;Meulemans 1998; Hill et al. 1995).
Light intensity is usually considered to be the mostderiving abiotic factor regulating the periphytoncolonization in lakes, particularly in shallow eutrophiclakes. Irradiance level can affect the epiphytic algalproductivity (Hill et al. 1995). The influence of thelight intensity depends upon the solar energy thatpenetrates the water column in the littoral zone aswell the self-shading within the periphyton (Müller1996). It is worthwhile to point out that most part ofthe light can be either absorbed or scattered into adifferent direction in shallow lakes. Abundance ofmacrophytes and periphyton decreases with turbidityin the shallow areas under the very turbid conditions(Scheffer 1998).
Nutrient availability is the most important factorcontrolling periphyton; several studies demonstratedthat when aquatic ecosystems are subjected toeutrophication, a subsequent loss of diversity can bedetermined (Cederwall and Elmgren 1990; Wormet al. 1999). However, there are controversial studiesthat nutrient enrichment can either increase ordecrease species diversity (Pringle 1990; Miller etal. 1992). Several workers reported that some domi-nant species can give a brief idea about the waterquality. For instance, Marker and Collett (1997) statedthat Cocconeis placentula Ehr. grows well under awide range of conditions and Niztschia amphibia,Niztschia palea and Gomphonema parvulum aregenerally recognised to be indicators of pollutedwaters. They also concluded that Achnanthes minu-tissima, Navicula minisculus, Nitzschia dissipata andFragilaria ulna are generally found in less pollutedbut eutrophic waters.
Periphyton is widely used as an indicator of waterquality studies since it is sensitive to both hydrologicand water quality changes, and responds rapidly tothese changes (Gaiser et al. 2004) and responses ofthis community to pollutants can be measured at avariety of time scales determining the physiological ofthe community-level changes (Strand and Weisner1996; Buczkó and Ács 1998; Ács et al. 2000).Therefore, structural and functional characteristics of
the periphyton have been used for several decades asindicators of water quality (Biggs 1989; Albay andAkçaalan 2003). Vis et al. (1998) stated that thecharacteristics of periphyton that respond to changesin water quality are not always the same. Theyemphasized that certain studies showed the correlationbetween indices of diversity or species autoecolgy andwater quality whereas other studies revealed strongerresponses of biomass indices (Biggs 1989). Accord-ingly, Albay and Akçaalan (2003) reported that somespecies (such as Aulacoseira italica, Melosira var-ians, and Planktolyngbya limnetica) have a broadrange of tolerance to physical disturbance includingwater level fluctuation, large amounts of suspendedsolids, and low Secchi disc depth.
This paper aimed to compare the environmentalfactors and hydrologic conditions controlling verticalcolonisation of periphyton on Sparganium erectum ina shallow eutrophic turbid lake (Manyas Lake) and anoligo-mesotrophic deep lake (Sapanca Lake). Thespecific goal was to clarify the possibility of the usageof the periphyton as bioindicators of anthropogeniccontamination in two lakes with different mixingregimes.
Materials and methods
Study sites
Descriptions of the studied lakes and complete data-sets are reported in Albay et al. (2003) and Albay andAkçaalan (2003). The main morphological andhydrological characteristics of the lakes are summa-rized in Table 1, Manyas Lake is a shallow-turbid lakewith a surface area of 159 km2. The investigated areaof the lake is covered with mostly emerged macro-phytes particularly, Phragmites australis, Typha sp. S.erectum and has a large littoral region where there islittle water movement because of dense macrophytestand, and is surrounded by agricultural land. ManyasLake is one of the most important world wide naturalreserve (Ramsar site) of Turkey because of itsecological values. The lake lies on a migratory waterbirds route and is rich in food for birds. The lakesupports 23 fish species, crayfish, otter, turtles, watersnakes etc. (Balik 1989; Erkakan 1997; Çelik 2006).
Sapanca is a moderately deep lake, located in thenortheast of the Marmara region of Turkey, has a
172 Environ Monit Assess (2008) 146:171–181

55 m maximum depth, 46.8 km2 surface area and ca.250 km2 catchment area. The lake is fed by severalcreeks and significant amount of ground water(Tuğrul and Morkoç 1991). The catchment area wasused for agricultural activities. The unexpecteddevelopment of tourism and recreation in the begin-ning of the 1990s has given rise to excessive nutrientinput to the lake. Then, first fish kill was observedafter the luxuriant growth of Planktothrix rubescensin 1997 (Albay et al. 2003). At present, the lakeprovides drinking water for the district of Sakarya. Itis also used for agriculture and abstracted for industrialuse.
Periphyton and chlorophyll-a
The samples of the periphyton were collected fromthe stems of the S. erectum from the Sapanca andManyas Lake between July 1997 and November 1998at monthly intervals. In a few cases, samples were notcollected for practical reasons during the studiedperiod. Each of the S. erectum stem was cut abovethe rhizome and subdivided into three equal sections(25 cm each) called “upper,” “middle” and “lower”ones. Periphyton matt was removed manually andcollected in 200 ml distilled water. A one hundredmillilitres of periphyton-containing water quantita-tively subsample for algal counting according toUtermöhl (1958). For each course, 600–800 cellswere counted. To determine the diatoms, watersamples were treated with H2O2. The remainingslurry (100 ml) filtered from the glass-fibre filters(Whatman GF/C) and chlorophyll-a measured accord-ing to Nusch (1980). The water level fluctuation
(WLF) data was kindly provided by the GeneralDirectorate of Electrical Power Resources Survey andDevelopment Administration. The methods for deter-minations of the epiphytic algal biomass, algaltaxonomy and water quality parameters have beendescribed by Albay and Akçaalan (2003).
Statistical analysis
The relationships between epiphytic algal groups, chl-a,and water quality parameters were explored byCanonical Correspondence Analysis (CCA) usingCANOCO 4.5 software (ter Braak and Smilauer2002). Relationship between environmental parametersand dominant species were evaluated using Spearmanrank correlation test. Changes in periphyton dynamicswere examined using Shannon’s diversity index, H′,calculated from the biovolume data as;
H' ¼Xs
i¼1
pi log2 pi
Where pi is the relative biovolume of species and i ands are number of species (Sommer 1993). Equitability(E) was calculated as H/Hmax.
Results
Manyas lake
The results presented in Table 2 shows the physicaland chemical conditions of Manyas Lake. The lakewater has showed eutrophic characters with lowphotic depth (zeu), high concentration of nutrients,conductivity and TSS because of intense sediment–water contact during the day. The CCA ordinationdemonstrated the importance of the different environ-mental factors in determining dominant epiphyticalgal species. The position of the dominant speciese.g. Rhoicosphaenia curvata (Kütz.) Grun. ex Rabh.,Nitzschia linearis (Agardh) W. Smith, Stigeocloniumsp., Cylindrocapsa sp., Limnothrix sp. were placedclosed to temperature and pH vectors; Melosiravarians C. Agardh and Nitzschia palea (Kütz.) W.Smith were closed to NO2+NO3, Secchi depth, o-PO4
and Chl-a vectors; Pediastrum boryanum (Turpin)Meneghini and Aulacoseira italica (Ehr.) Simonsenwere placed to near the TSS vector in the ordination
Table 1 Morphological characteristics and locations ofManyas and Sapanca lakes
Parameters Unit Manyas Lake Sapanca Lake
Coordinates 40°12′N–28°00′E 30°15′E–40°42′NAltitude M 18 30Maximum depth M 3.60 55Volume m3 265.3×106 1.2×109
Catchment area km2 3,352 250Surface area km2 159 45Retention time Year 0.71 UnknownMixing type Polymictic Warm-monomicticSpecial status National parka No special status
a Ramsar side
Environ Monit Assess (2008) 146:171–181 173

diagram of CCA (Fig. 1). Only 21 dominant algalspecies are shown in Fig. 1. The eigenvalues of CCAaxis 1 (0.35) and axis 2 (0.25), constrained to theeight variables account for 50% of the cumulativevariance in the species data. The species–environmentcorrelations of CCA axis 1 and 2 are high and the firsttwo axis account for 59.3% of the variance in theepiphytic algae–environment relationships (Table 3).
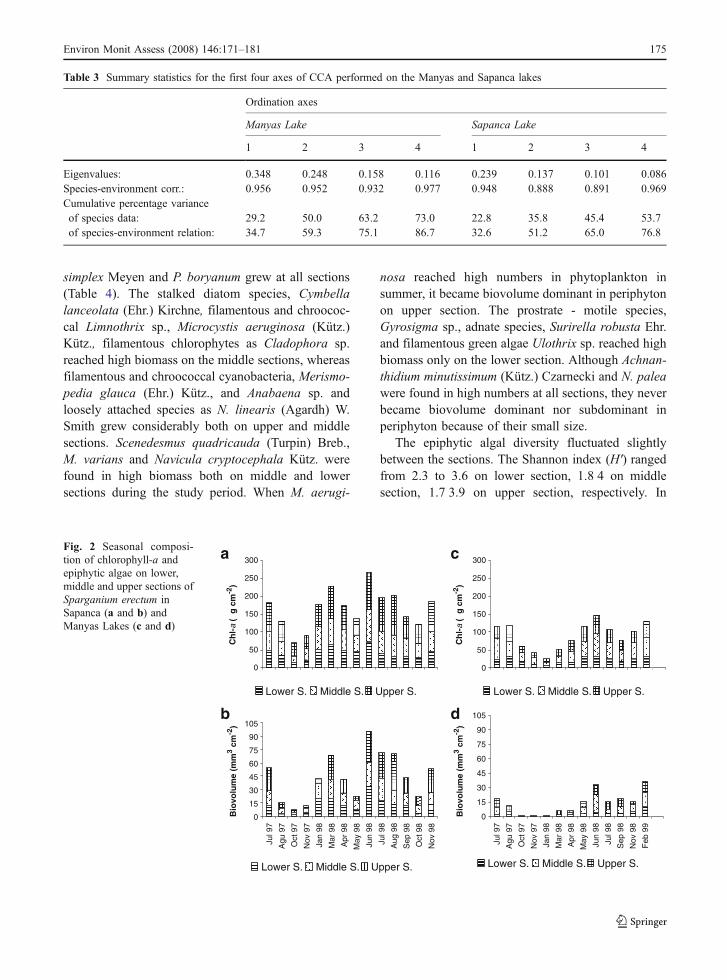
The seasonal variation of chlorophyll-a (chl-a)concentration showed considerable difference amongthe sections (p<0.05; Fig. 2). Mean chl-a concentra-tion was 21.4, 37.5 and 31.2 µg cm−2 on lower,middle and upper sections, respectively. It showedsimilar seasonal trend with algal biovolume at allsections (r=0.68, 0.72, 0.58; p<0.01 on lower,middle and upper sections). In the samples obtainedduring the 1997, maximum chl-a was recorded in Julyon the middle section as 59.4 µg cm−2 and minimumchl-a was measured in November on lower section as9 µg cm−2.
A total 123 species of periphyton were identified.Periphyton assemblages on S. erectum were identicalmostly in the eutrophic lakes. The predominantspecies at all sections were mostly filamentousmulticellular green algae, Oedogonium sp. and Sti-geoclonium sp., Cylindrocapsa sp., Spirogyra sp.,Mougeotia sp. In addition to these species, filamen-tous diatom, Aulacoseira italica, A. muzzanensis(Meist.) Krammer, filamentous blue-green algae, P.limnetica Lemm., loosely attached taxa F. ulna, theadnate species Navicula tripunctata, Surirella linearisW. Smith, and Chlorococcal green algae Pediastrum
Table 2 The physical, chemical and biological characters of Sapanca and Manyas lake between July 1997 and November 1998
Manyas Lake Sapanca Lake
Parameters Unit Mean (max–min) Mean (max–min)
Temperature °C 18,6 (7.1–27.1) 19.9 (8–27)pH 8.1 (7.4 –8.9) 8 (7.8–8.4)Conductivity µS cm−1 2,830 (980 –8,860) 205 (157–253)TSS mg l−1 94.4 (61–134) 3.89 (0.6 –6.8)SRP µg l−1 168 (40.4–324) 5.2 (2–16)TP µg l−1 628 (164 –922) 12.4 (4.5–38)NO2+NO3 µg l−1 614 (188–1,144) 27.1 (2–92)Secchi depth cm 21.6 (16–34) 276 (100 –380)Zeu cm 60.5 (44.8–95.2) 773 (280 –1,064)Total biomass mm3 cm−2 4.21 (0.17–11.35) 14.79 (2.29–31.86)Shannon diversity H′ 3.01 (2–3.7) 3.34 (2.4 –4.4)Number of species n 22 (13– 40) 27 (15–38)
-1.0 1.0
-1.0
1.0
achlin
cytum
melvar
nacry
cylinitlin oed1nipal
rhcur
sceco scqu
limno
aulit
syber
natri
pedsim
fraul
moug
plali
pedbor
stigeTemp.
pH
TSS
o-PO4
TP
NO2+NO3
WLF
Sec dep.
Fig. 1 Biplot of first and second CCA axis for periphyton inManyas Lake. Environmental variables: achlin, Achnantheslinearis; aulit, Aulacoseira italica; cyli, Cylindrocapsa; cymlan,Cymbella lanceolata; fraul, Fragilaria ulna; limno, Limnothrix;melvar, Melosira varians; natri, Navicula tripunctata; nacry,Navicula cryptocephala; nitlin, Nitzschia linearis; oed1, Oedo-gonium1; moug, Mougeotia; pedbor, Pediastrum boryanum;pedsim, Pediastrum simplex; plali, Planktolyngbya limnetica;rhcur, Rhoicosphaenia curvata; sceco, Scenedesmus ecornis;scqu, Scenedesmus quadricauda; stige, Stigeoclonium; syber,Synedra berolinensis; Temp, Temperature; Sec dep, SecchiDepth
174 Environ Monit Assess (2008) 146:171–181
simplex Meyen and P. boryanum grew at all sections(Table 4). The stalked diatom species, Cymbellalanceolata (Ehr.) Kirchne, filamentous and chroococ-cal Limnothrix sp., Microcystis aeruginosa (Kütz.)Kütz., filamentous chlorophytes as Cladophora sp.reached high biomass on the middle sections, whereasfilamentous and chroococcal cyanobacteria, Merismo-pedia glauca (Ehr.) Kütz., and Anabaena sp. andloosely attached species as N. linearis (Agardh) W.Smith grew considerably both on upper and middlesections. Scenedesmus quadricauda (Turpin) Breb.,M. varians and Navicula cryptocephala Kütz. werefound in high biomass both on middle and lowersections during the study period. When M. aerugi-
nosa reached high numbers in phytoplankton insummer, it became biovolume dominant in periphytonon upper section. The prostrate - motile species,Gyrosigma sp., adnate species, Surirella robusta Ehr.and filamentous green algae Ulothrix sp. reached highbiomass only on the lower section. Although Achnan-thidium minutissimum (Kütz.) Czarnecki and N. paleawere found in high numbers at all sections, they neverbecame biovolume dominant nor subdominant inperiphyton because of their small size.
The epiphytic algal diversity fluctuated slightlybetween the sections. The Shannon index (H′) rangedfrom 2.3 to 3.6 on lower section, 1.8 4 on middlesection, 1.7 3.9 on upper section, respectively. In
a
0
50
100
150
200
250
300
Ch
l-a
(µg
cm
-2)
Ch
l-a
( µg
cm
-2)
Lower S. Middle S. Upper S.
c
0
50
100
150
200
250
300
Lower S. Middle S. Upper S.
b
0
15
30
45
60
75
90
105
Jul 9
7
Agu
97
Oct
97
Nov
97
Jan
98
Mar
98
Apr
98
May
98
Jun
98
Jul 9
8
Aug
98
Sep
98
Oct
98
Nov
98
Jul 9
7
Agu
97
Oct
97
Nov
97
Jan
98
Mar
98
Apr
98
May
98
Jun
98
Jul 9
8
Sep
98
Nov
98
Feb
99
Bio
volu
me
(mm
3 cm
-2)
Bio
volu
me
(mm
3 cm
-2)
Lower S. Middle S. Upper S.
d
0
15
30
45
60
75
90
105
Lower S. Middle S. Upper S.
Fig. 2 Seasonal composi-tion of chlorophyll-a andepiphytic algae on lower,middle and upper sections ofSparganium erectum inSapanca (a and b) andManyas Lakes (c and d)
Table 3 Summary statistics for the first four axes of CCA performed on the Manyas and Sapanca lakes
Ordination axes
Manyas Lake Sapanca Lake
1 2 3 4 1 2 3 4
Eigenvalues: 0.348 0.248 0.158 0.116 0.239 0.137 0.101 0.086Species-environment corr.: 0.956 0.952 0.932 0.977 0.948 0.888 0.891 0.969Cumulative percentage varianceof species data: 29.2 50.0 63.2 73.0 22.8 35.8 45.4 53.7of species-environment relation: 34.7 59.3 75.1 86.7 32.6 51.2 65.0 76.8
Environ Monit Assess (2008) 146:171–181 175

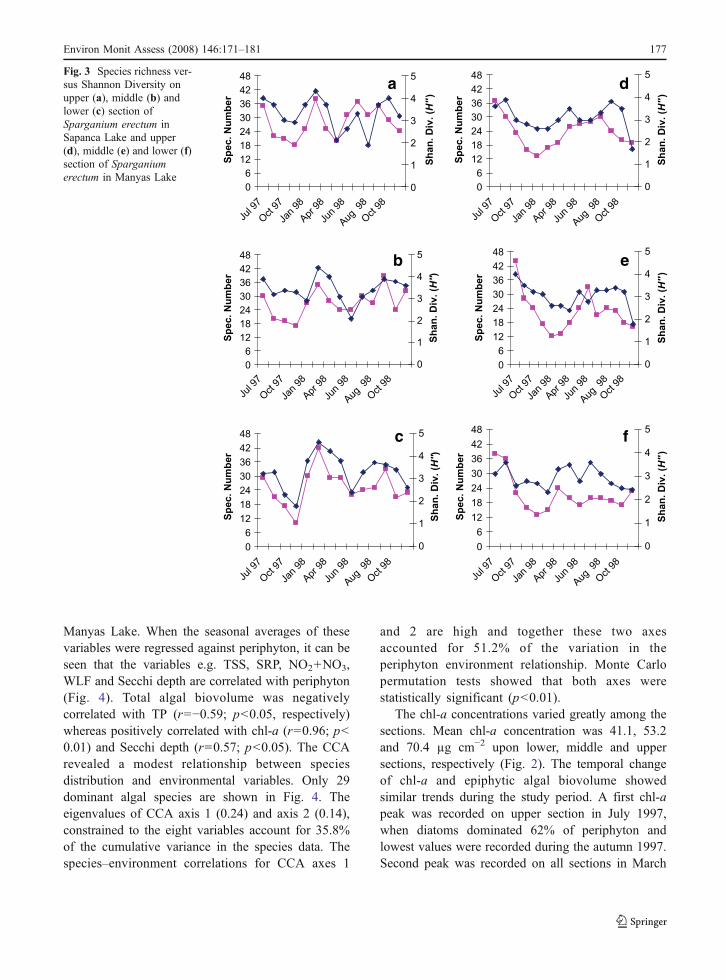
general, high values are found during mid spring tomid autumn, and low values were found during lateautumn to early spring months and maximum valueswere recoded in summer months (Fig. 3). Positivecorrelations were recorded between Shannon diversityand number of species on all sections; a modestpositive correlation was recorded on lower and uppersections (r=0.46 and r=0.62; p<0.01, respectively)and a strong positive correlation was found on middlesection (r=0.69; p<0.01). Temperature and Suspended
Solids were the factors which affect the ShannonDiversity profoundly during the study period.
Sapanca lake
Table (2) summarizes the measured physical andchemical characters of Sapanca Lake for the periodJuly 1997 November 1998. The seasonal dynamics ofnutrient concentrations and other water qualityparameters in Sapanca Lake differed notably from
Table 4 Dominance of most abundant species according to biovolume units on Sparganium erectum in Sapanca and Manyas Lake
Sapanca Lake Manyas Lake
Low. S. Mid. S. Upp. S. Low. S. Mid. S. Upp. S.
Aulacoseira italica * * * * * *Aulacoseira muzzanensis * * * * *Cocconeis placentula * * *Cymbella lanceolata * * * *Encyonema minuta * * *Fragilaria construens *Fragilaria crotonensis * * *Fragilaria ulna * * * * * *Gyrosigma sp. * * * *Melosira varians * * * * *Navicula cryptocephala * * * * *Navicula tripunctata * * * * * *Nitzschia linearis * *Nitzschia palea * *Rhoicosphaenia curvata *Surirella linearis * * * *Cladophora sp. * * * *Coleochaete sp. *Cosmarium formulosum * * *Cylindrocapsa sp. * * * * * *Draparnaldia sp. *Mougeotia sp. * * * * * *Oedogonium sp. (1) * * * * * *Oedogonium sp. (2) * * *Pediastrum boryanum * * *Pediastrum simplex * * *Scenedesmus quadricauda * *Stigeoclonium sp. * * *Ulothrix sp. * * *Merismopedia glauca * *Oscillatoria limnetica *Planktolyngbya limnetica * * *Pseudoanabaena sp. * * *
*Available
176 Environ Monit Assess (2008) 146:171–181
Manyas Lake. When the seasonal averages of thesevariables were regressed against periphyton, it can beseen that the variables e.g. TSS, SRP, NO2+NO3,WLF and Secchi depth are correlated with periphyton(Fig. 4). Total algal biovolume was negativelycorrelated with TP (r=−0.59; p<0.05, respectively)whereas positively correlated with chl-a (r=0.96; p<0.01) and Secchi depth (r=0.57; p<0.05). The CCArevealed a modest relationship between speciesdistribution and environmental variables. Only 29dominant algal species are shown in Fig. 4. Theeigenvalues of CCA axis 1 (0.24) and axis 2 (0.14),constrained to the eight variables account for 35.8%of the cumulative variance in the species data. Thespecies–environment correlations for CCA axes 1
and 2 are high and together these two axesaccounted for 51.2% of the variation in theperiphyton environment relationship. Monte Carlopermutation tests showed that both axes werestatistically significant (p<0.01).
The chl-a concentrations varied greatly among thesections. Mean chl-a concentration was 41.1, 53.2and 70.4 µg cm−2 upon lower, middle and uppersections, respectively (Fig. 2). The temporal changeof chl-a and epiphytic algal biovolume showedsimilar trends during the study period. A first chl-apeak was recorded on upper section in July 1997,when diatoms dominated 62% of periphyton andlowest values were recorded during the autumn 1997.Second peak was recorded on all sections in March
Fig. 3 Species richness ver-sus Shannon Diversity onupper (a), middle (b) andlower (c) section ofSparganium erectum inSapanca Lake and upper(d), middle (e) and lower (f)section of Sparganiumerectum in Manyas Lake
a
0
6
12
18
24
30
36
42
48
Jul 9
7
Oct 9
7
Jan 9
8
Apr 98
Jun 9
8
Aug 98
Oct 9
8
Jul 9
7
Oct 9
7
Jan 9
8
Apr 98
Jun 9
8
Aug 98
Oct 9
8
Jul 9
7
Oct 9
7
Jan 9
8
Apr 98
Jun 9
8
Aug 98
Oct 9
8
Jul 9
7
Oct 9
7
Jan 9
8
Apr 98
Jun 9
8
Aug 98
Oct 9
8
Jul 9
7
Oct 9
7
Jan 9
8
Apr 98
Jun 9
8
Aug 98
Oct 9
8
Jul 9
7
Oct 9
7
Jan 9
8
Apr 98
Jun 9
8
Aug 98
Oct 9
8
Sp
ec.
Nu
mb
er
Sp
ec
. N
um
ber
Sp
ec
. N
um
be
r
0
1
2
3
4
5
Sh
an
. D
iv. (H''
)
Sh
an
. D
iv. (H''
)S
han
. D
iv. (H''
)S
han
. D
iv. (H''
)
Sh
an
. D
iv. (H'')
Sh
an
. D
iv. (H'')
d
0
6
12
18
24
30
36
42
48
Sp
ec.
Nu
mb
er
0
1
2
3
4
5
b
0
6
12
18
24
30
36
42
48
0
1
2
3
4
5 e
0
6
12
18
24
30
36
42
48
Sp
ec
. N
um
ber
0
1
2
3
4
5
c
0
6
12
18
24
30
36
42
48
0
1
2
3
4
5 f
0
6
12
18
24
30
36
42
48
Sp
ec
. N
um
be
r
0
1
2
3
4
5
Environ Monit Assess (2008) 146:171–181 177
1998, when more than 95% of the periphytoncomprised of diatoms. A last and highest chl-a peakwas measured in June 1998 when more than 86% ofthe biovolume consisted of chlorophytes.
A total 109 species of periphyton were identifiedChlorophytes, diatoms, and cyanobacteria were foundas major algal groups in the periphyton on S. erectumin Sapanca Lake. However, the zonation of theperiphyton, density, composition and dominant/sub-
dominant taxa were significantly different in theselakes; mean algal biomass was considerably high inSapanca Lake whereas the number of the species washigh in Manyas Lake (Table 2). Mean diatom andcyanobacteria biomass showed a modest differenceamong the sections whereas mean chlorophyte bio-mass was found considerably high on upper section(p<0.01; Table 5). A. italica, F. crotonensis, C. lanceo-lata, N. cryptocephala, N. tripunctata, M. varians,Gyrosigma sp., Cylindrocapsa sp., F. ulna, C. placen-tula, Oedogonium spp., Pseudoanabaena sp., Mougeo-tia sp., and Cladophora glomerata (Linn.) Kütz. grewon all sections whereas N. palea, and Ulothrix sp. grewon lower and middle sections, Cosmarium formulosumW.E. Hoffmann ex Nordsted and Oedogonium sp.grew on middle and upper sections. Fragilariaconstruens var. venter (Ehr.) Grun., Draparnaldia sp.and Oscillatoria limosa (Dill.) C. Agardh wererecorded in high biomass only on the lower sectionand Cocconeis scutellum (Ehr), Surirella linearis W.Smith, Epithemia adnata (Kütz.) Breb., Coleochaetesp., R. curvata were found in high biomass only on theupper section (Table 4).
Figure 3 shows the seasonal differences in epi-phytic algal diversity on S. erectum in Sapanca Lake.In general, the diversity values were slightly higherthan Manyas Lake; They ranged from 1.8 to 4.6 onlower section, 2.1 to 4.4 on middle section and 1.9 to4.3 on upper sections, respectively. Contrary toManyas Lake, the highest diversity values were foundin early spring and the lowest values were found insummer months. Shannon diversity index and numberof species showed a very strong positive correlationon the lower section (r=0.87; p<0.01)) whereas amodest positive correlation was recorded on themiddle and upper sections (r=0.49 and r=0.42 p<0.01, respectively).
-1.0 1.0
-1.0
1.0
aulit
gomol
frcro
cymlan
encmin
nacry
achlin
fraul
surlin
gomancocpla nipal
epsor
epad
cosfor
oed1
oslim
rhcur
natri
gopar
nitlin
cyli
frconmougclad
oed2cocscu
cytum
melvarTemp.
Cond.
TSS
o-PO4
TP
NO2+NO3
SiO2
WLF
Sec dep.
Fig. 4 Biplot of first and second CCA axis for periphyton inSapanca Lake. Environmental variables: achlin, Achnantheslinearis; aulit, Aulacoseira italica; clad, Cladophora; cocpla,Cocconeis placentula; cocscu, C. scutellum; cosfor, Cosmariumformulosum; cyli, Cylindrocapsa; cymlan, Cymbella lanceolata;cytum, C. tumida; encmin, Encyonema minutum; epad, Epi-themia adnata; epsor, E. sorex; frcon, Fragilaria construens;frcro, F. crotonensis; fraul, F. ulna; gomol, Gomphonemaolivaceum; gopar, G. parvulum; melvar, Melosira varians;moug, Mougeotia; nacry, Navicula cryptocephala; natri, N.tripunctata; nipal, Nitzschia palea; oed1, Oedogonium1; oslim,Oscillatoria limosa; rhcur, Rhoicosphaenia curvata; surlin,Surirella linearis; Temp, Temperature; Sec dep, Secchi Depth
Table 5 Vertical colonization of major epiphytic algal groups on Sparganium erectum
Sapanca Lake Manyas Lake
Unit Low. S. Mid. S. Upp. S. Low. S. Mid. S. Upp. S.
Chlorophytes % 56.8 58 70.9 43.4 60.3 61.5Diatoms % 38.7 37.4 25.3 48.6 32.8 24.5Cyanobacteria % 4.4 4.4 3.8 8 6.9 14
Low S Lower Section, Mid S Middle Section, Upp S Upper Section
178 Environ Monit Assess (2008) 146:171–181

Discussion
Studies attempting to understand the complex eco-logical interrelationships existing in aquatic environ-ments have frequently overlooked the role of theperiphytic community (Sheldon and Boylen 1975).The factors affecting the colonization of epiphyticalgae on macrophytes and artificial substrates havelong been studied by several workers (Sheldon andBoylen 1975; Ács and Kiss 1991; Vis et al. 1998;Albay and Akçaalan 2003). Temporal and spatialvariability in physico-chemical parameters acts toorganize epiphytic algal assemblages, regulate speciesinteractions and, by providing conditions that supportdifferent taxa under various environmental conditionsmay be crucial for maintaining biodiversity (Vis et al.1998; Wellnitz and Ward 1998; Cardinale et al. 2000).
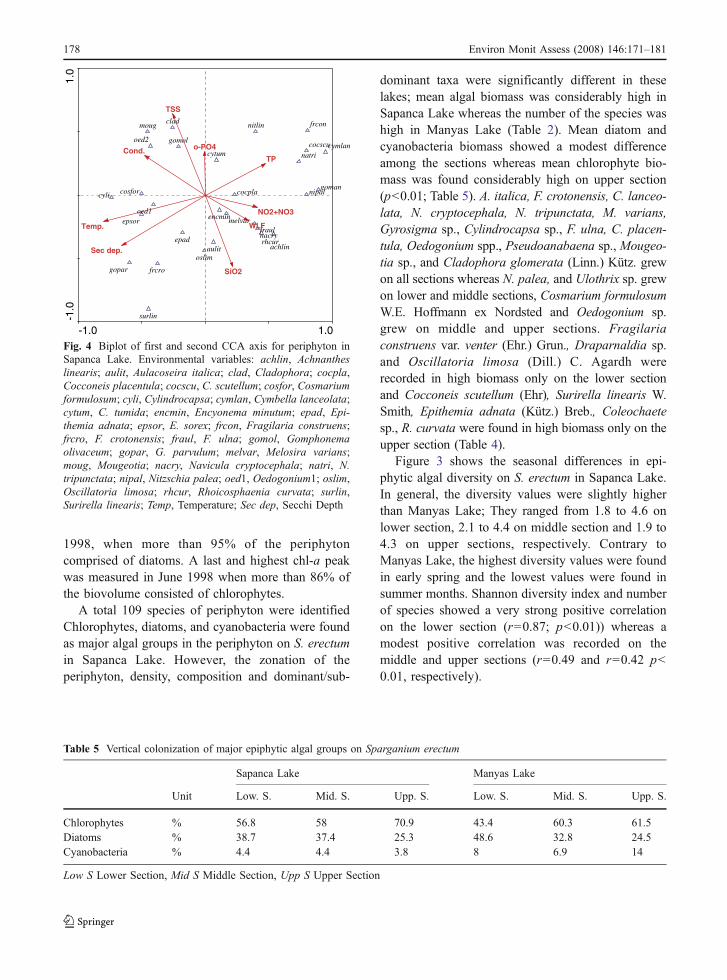
In the present study, we found distinct patterns ofperiphyton colonization related to water quality andhydrologic drivers. Our results indicate that three majorenvironmental gradients (high amount of TSS, lowlight climate and WLF) control the vertical coloniza-tion of periphyton assemblages for Manyas (Table 2).Mean periphyton biomass was 4.21 mm3 cm−2 inManyas lake whereas it was found considerably high(14.79 mm3 cm−2) in Sapanca lake. Scheffer (1998),stated that light is predominantly absorbed by par-ticulate matter such as phytoplankton cells, detritusand suspended sediment particles in many shallowlakes and shallow, slowly flowing side arms (inSoroksári Danube, Szabó et al. 2001). Accordingly,Müller (1994) stated that the amount of light reachingthe algal cells depends on external factors, such as thePAR and length of the macrophyte stand, and onfactors in the water such as Secchi depth andthickness of the periphytic layer. The high amountof Suspended Solids values (mean: 21.6 cm) limitedthe light penetration in the water in Manyas Lake.Therefore, it is likely to conclude that low lightintensity might have given rise to low amount ofperiphytic biomass on S. erectum in this lakecompared to Sapanca Lake (Table 2).
The nutrients level and other measured waterquality parameters in Manyas Lake are indicative ofthe lake’s present highly eutrophic status (Table 2)whereas Sapanca Lake is still in oligo-mesotrophicconditions according to OECD (1982). The commu-nity structure and vertical colonization of epiphyticalgae on S. erectum showed a distinct difference
between the two lakes. The diatoms were recorded asthe dominant group with 49% in Manyas Lake,whereas green algae accounted for 57% of theperiphyton in Sapanca Lake on the lower section(Table 5). The middle and upper sections weredominated by green algae both in Manyas Lake andSapanca Lake. The highest epiphytic algal biomasswas recorded on the middle section in Manyas Lakewhereas highest biomass values were found on uppersection in Sapanca Lake. The WLF might havebrought about this situation since it fluctuate ±31 cmin Manyas Lake during the study period. It can bethought that WLF has a most pronounced effect onperiphyton colonization in shallow lakes than deep lakesbecause of their morphometric character. Rychkova(1989) stated that the periphytic biomass is largelyeliminated by changes in the WLF and wave action.
Totally eight taxa (Oedogonium sp., Mougeotiasp., Cylindrocapsa sp., Cladophora sp., A. italica, M.varians, N. tripunctata, and F. ulna) were found asdominant species at all sections of S. erectum both inManyas Lake and Sapanca Lake (Table 4). Similarresults were found by Müller (1994) in a small,moderately-deep, hard-water German lake, LakeBelau. It can be thought that these species have abroad range of tolerance to several physical andchemical disturbances including nutrients, TSS, WLFand light intensity (Table 2). Oedogonium sp. was thepioneer species (mean 0.51 and 7.16 mm3 cm−2 inManyas Lake and Sapanca Lake, respectively) duringthe study period. The other filamentous chlorophytes,Stigeoclonium sp. and Spirogyra sp. reached highbiomass values in Manyas Lake, whereas Coleo-chaete sp., Draparnaldia sp., were found in highbiomass values in Sapanca Lake occasionally. Theexcessive growth of some filamentous chlorophytes,such as Oedogonium sp., Spirogyra sp., and Clado-phora glomerata, has long been associated witheutrophication (Cattaneo 1987; Albay and Akçaalan2003). Cattaneo (1987) stated that filamentous greenalgae were positively correlated with TP. Amongthese species Cladophora glomerata has primarilyresponded to eutrophication (Danilov and Ekelund2001) and to a lesser degree Ulothrix, Spirogyra andOedogonium. It can be thought that this species shouldbe used as a key species in lake monitoring studies.
The correlation between diversity (H′) and numberof species was positive however it was not highlysignificant at all sections except lower section of S.
Environ Monit Assess (2008) 146:171–181 179

erectum in Sapanca Lake. Seasonal variation inepiphytic algal diversity and number of speciesshowed highly different pattern between two lakes(Fig. 3 and Table 2). The highest diversity valueswere recorded in summer and lowest values wasrecorded in late autumn in Manyas Lake whereashighest and lowest diversity values were recorded inearly spring and summer months in Sapanca Lake. Itis worthwhile the point out that the highest diversityvalues were recorded when periphyton was dominat-ed by several epiphytic algal species and lowestvalues were recorded when periphyton was dominat-ed by few species in both lakes. Ács et al. (2000)stated that changes of number of species and evennessin periphyton depend on the immigration and repro-duction rate of algae. They concluded that immigra-tion causes an increase in the number of specieswhereas reproduction maintains species richness byreducing the losses deriving from death.
The periphyton community was dominated byspecies which is tolerant to eutrophic conditions inthe Manyas Lake whereas meso-oligotrophic taxadominated in the periphyton of Sapanca Lake. Ourresults showed that certain water quality and physicaldisturbances e.g. high amount of suspended solids,WLF and nutrients and low light intensity have had adistinctive effect on the vertical colonization ofperiphyton, number of species, diversity and compo-sition. The results also revealed that periphyton canbe used as bioindicators of anthropogenic contamina-tion in lakes. More information is needed to examinethe factors governing periphyton colonization in lakeswith different mixing and trophic level. This assess-ment is urgently required, given the need to beginimplementation of the Water Framework Directives(Flynn et al. 2002).
Acknowledgements We are thankful to Eva Acs and JorgenChristiansen for reading and comments on the draft of themanuscript. The present work was supported by the ResearchFund of Istanbul University. Project No: 993/090597 and UDP-156/04062003.
References
Ács, É., & Kiss, K. T. (1991). Investigation of periphytic algaein the Danube at Göd (1669 river km, Hungary).Algological Studies, 62, 47–67.
Ács, É., Kiss, K. T., Szabó, K., & Makk, J. (2000). Short-termcolonization sequence of periphyton on glass slides in a
large river (River Danube, near Budapest). AlgologicalStudies, 100, 135–156.
Albay, M., & Akçaalan, R. (2003). Comparative study ofperiphyton colonisation on common reed (Phragmitesaustralis) and artificial substrate in shallow lake, Manyas,Turkey. Hydrobiologia, 506–509, 531–540.
Albay, M., Akçaalan, R., Tüfekçi, H., Metcalf, J. S., Beattie, K. A.,& Codd, G. A. (2003). Depth profiles of cyanobacterialhepatotoxins (microcystins) in three Turkish freshwater lakes.Hydrobiologia, 505, 89–95.
Balik, S. (1989). Determination of present state of ManyasLake and Bird Paradise and look for the way of solution.Projet Number: 1987/050 139p (in Turkish).
Biggs, B. J. F. (1989). Biomonitoring of organic pollution usingperiphyton, South Branch, Canterbury, New Zealand.New-Zealand Journal Marine Freshwater Research, 23,263–274.
Buczkó, K., & Ács, E. (1998). Comparison of succession ofreed periphyton in a degraded and in an undisturbedpart of shallow lake (Lake Velencei, Hungary, CentralEuropa). Verh Internat Verein Theor Ang Limnol, 26,1674–1676.
Cardinale, B. J., Nelson, K., & Palmer, M. A. (2000). Linkingspecies diversity to the functioning of ecosystems; on theimportance of environmental context. Oixos, 91, 175–183.
Cattaneo, A. (1987). Periphyton in lakes of different Trophy.Canadian Journal of Fisheries and Aquatic Sciences, 44,296–303.
Cederwall, H., & Elmgren, R. (1990). Biological effects ofeutrophication in the Baltic Sea, particularly in the coastalzone. Ambio, 19, 109–112.
Çelik, K. (2006). Spatial and seasonal variations in chlorophyll-nutrient relationships in the shallow hypertrophic LakeManyas, Turkey. Environmental Monitoring and Assess-ment, 117(1–3), 261–269.
Danilov, R. A., & Ekelund, N. G. A. (2001). Comparison ofusefulness of three types of artificial substrata (glass, woodand plastic) when studying settlement patterns of periph-yton in lakes of different trophic status. Journal ofMicrobiological Methods, 45, 167–170.
Erkakan, F. (1997). Manyas Lake watery areas administrationplan project. Republic of Turkey, Ministry of Environ-ment, General Directorate of Environmental Protection,Ankara. TUBITAK Project Number: 94K100010.
Flynn, N. J., Snook, D. L., Wade, A. J., & Jarvie, H. P. (2002).Macrophyte and periphyton dynamics in a UK Cretaceouschalk stream: The River Kennet, a tributary of theThames.. The Science of the Total Environment, 282–283, 143–157.
Gaiser, E. E., Scinto, L. J., Richards, J. H., Jayachandran, K.,Childers, D. L., Trexler, J. C., et al. (2004). Phosphorus inperiphyton mats provides the best metric for detectinglow-depth P enrichment in an oligotrophic wetland. WaterResearch, 38, 507–516.
Hill, W. R., Ryon, M. G., & Schilling, E. M. (1995). Lightlimitation in a stream ecosystem: Response by primaryproducers and consumers. Ecology, 76, 1297–1309.
Kiss, M. K., Lakatos, G., Borics, G., Gido, Z., & Deak, C.(2003). Littoral macrophyte-periphyton complexes in twoHungarian shallow waters. Hydrobiologia, 506–509, 541–548.
180 Environ Monit Assess (2008) 146:171–181

Marker, A. F. H., & Collett, G. D. (1997). The spatial andtemporal characteristics of algae in the River Great Ouse.II. The epiphytic algal flora. Regulated Rivers, 13, 235–244.
Meulemans, J. T. (1998). Seasonal changes in biomass andproduction of periphyton growing upon reed in LakeMaarsseveen I. Archiv für Hydrobiologie, 112, 21–42.
Miller, M. C., De Oliveria, P., & Gibeau, G. (1992). Epilithicdiatom community response to years of PO4- fertilization:Kuparuk River, Alaska (68°N Latitude). Hydrobiologia,240, 103–119.
Müller, U. (1994). Seasonal development of epiphytic algae onPhragmites australis (CAV.) TRIN. Ex STEN. In aeutrophic lake. Archiv für Hydrobiologie, 129(3), 273–292.
Müller, U. (1996). Production rates of epiphytic algae in aeutrophic lake. Hydrobiologia, 330, 37–45.
Nusch, E. (1980). Comparison of different methods forchlorophyll and phaeopigment determination. Arch Hydro-biol Beih, 14, 14–36.
Organization for Economic Cooperation and Development(OECD) (1982). Eutrophication of waters: monitoring,assessment, and control. Technical Report, EnvironmentalDirectorate. OECD, Paris, France.
Pringle, C. M. (1990). Nutrient spatial heterogeneity: effects oncommunity structure, physiognomy, and diversity ofstream algae. Ecology, 71, 905–920.
Rychkova, M. A. (1989). Role of water mass dynamics information of epiphyte algae communities in a lake.Hydrobiological Journal, 25(3), 7–9.
Scheffer, M. (1998). Ecology of shallow lakes. p. 357. London:Chapman and Hall.
Sheldon, R. B., & Boylen, C. W. (1975). Factors affecting thecontribution by epiphytic algae to the primary productivity
of an oligotrophic freshwater lake. Applied Microbiology,30(4), 657–667.
Sommer, U. (1993). Disturbance–diversity relationship in twolakes of similar nutrient chemistry but contrasting distur-bance regimes. Hydrobiologia, 249, 59–65.
Strand, J. A., & Weisner, S. E. B. (1996). Wave exposurerelated growth of epiphyton: Implications for the distribu-tion of submerged macrophytes in eutrophic lakes. Hydro-biologia, 325(2), 113–119.
Szabó, K., Ács, É., Pápista, É., Kiss, K. T., Barreto, S., &Makk, J. (2001). Periphyton and phytoplankton in theSoroksár-Danube in Hungary. I. Periphytic algae on reedstems. Acta Botanica Hungarica, 43(1–2), 13–35.
ter Braak, C. J. F., & Smilauer, P. (2002). CANOCO referencemanual and CanoDraw for Windows user’s guide: Soft-ware for canonical community ordination (version 4.5).Microcomputer Power, Ithaca, New York, USA.
Tuğrul, S., & Morkoç, E. (1991). Determination of limnologicalcharacteristics of the Sapanca Lake. DOĞA, Tr. J.Engineering and Environmental Sciences, 15, 265–282.
Utermöhl, H. (1958). Zur Vervollkommung der quantitativenphytoplankton-methodik. Mitteil. Verh Internat VereinTheor Ang Limnol, 9, 39.
Vis, C., Hudon, C., Cattaneo, A., & Pinel-Alloul, B. (1998).Periphyton as an indicator of water quality in the StLawrence River (Quebec, Canada). Environmental Pollution,101, 13–24.
Wellnitz, T. A., & Ward, J. W. (1998). Does light intensitymodify the effect mayfly grazers have on periphyton?Freshwater Biology, 39, 135–149.
Worm, B., Lotze, H. K., Boström, C., Engkvist, R., Labanauskas,V., & Sommer, U. (1999). Marine diversity shift link tointeractions among grazers, nutrients and propagule banks.Marine Ecological Progress Series, 185, 309–314.
Environ Monit Assess (2008) 146:171–181 181




















