
2. CORRELATION OF QUATERNARY TEPHRAS THROUGHOUT THE IZU-BONIN AREAS1
Upload
independentCategory
view
0download
0
Journal of Applied Phycology 9: 137–146, 1997. 137c 1997 Kluwer Academic Publishers. Printed in Belgium.
Changes in atrazine toxicity throughout succession of stream periphytoncommunities
Helena Guasch�, Isabel Munoz, Nuria Roses & Sergi SabaterDepartament d’Ecologia, Facultat de Biologia, Universitat de Barcelona, Avinguda Diagonal 645, 08028Barcelona, Spain(�Author for correspondence)
Received 5 February 1997; revised 25 April 1997; accepted 4 May 1997
Key words: atrazine toxicity, periphyton, stream, succession
Abstract
A study was made to describe atrazine toxicity and its changes throughout succession of periphyton communities ofan undisturbed Mediterranean stream. Toxicity was assessed by short-term physiological tests (concentration-effectcurves of photosynthesis to atrazine) in the laboratory using artificial substrates colonized in one stream site duringwinter, and two stream sites (one open and the other shaded) during summer. In the winter experiment, whenenvironmental conditions were relatively steady and chlorophyll content was low, toxicity increased accordingto the increases in cell density and chlorophyll content throughout colonization. EC50 (concentration inhibitingphotosynthesis by 50%) was above 0.8 �M atrazine until day 16 and below 0.4 �M atrazine after three weeks. Inthe summer experiment, under more variable environmental conditions, the differences between the EC50 at thebeginning and the end of the colonization experiments were not significant (one factor ANOVA) at the two sites.EC50 was on average 0.89 �M atrazine in the shaded site and 0.29 �M atrazine in the open site. A significantnegative correlation between irradiance and EC50 was observed when all the experiments were considered together(r = 0.464, n = 20, p<0.05), suggesting that light history may have an important role in the response to atrazine. Thisinvestigation reveals that the response of stream periphyton to atrazine is likely to be influenced by colonizationtime and the corresponding changes in algal density and community composition as well as by environmentalconditions (e.g. light regime) throughout succession.
Introduction
Increased run-off of natural and man-made substancesfrom catchments has marked effects on river commu-nities. Natural periphyton communities are among thecomponents of lotic ecosystems which may be usedto monitor impacts (Whitton & Kelly, 1995). With-in the methods applied to assess the effect of toxicantsusing freshwater algae, most research has been focusedon single-species tests, conducted in the laboratory.Batch-culture toxicity methods have been reviewedcritically by Nyholm and Kallqvist (1989). In the lastdecade, several methods using complete periphytoncommunities have been used to assess the toxicity ofchemicals on the structure and function of aquatic sys-
tems (e.g. Niederlehner & Cairns, 1993; Lewis et al.,1993; Blanck et al., 1988).
The herbicide atrazine (6-chloro-N-ethil-N0-(1-methylethyl)-1,3,5-triazine-2,4-diamine) is found inmany surface and groundwaters and its ecologicaleffects are a possible concern for aquatic ecosys-tems. Such concerns have generated much researchon its toxicity to aquatic organisms (e.g. Huber, 1993;Solomon et al., 1996). Concentrations above 100 �gL�1 cause very marked effects on the photosynthe-sis, growth, chlorophyll content and biomass of mostaquatic primary producers (e.g. Plumley & Davis,1980; Brockway et al., 1984; Kosinski & Merkle,1984). Long-term exposure to these high atrazineconcentrations performed in lentic mesocosms canchange the algal community composition towards a
Article: japh 477 GSB: Pips nr 140602 BIO2KAP
*140602 japh477.tex; 9/07/1997; 12:22; v.7; p.1

138
diatom-dominated community (Hamilton et al., 1988;Hoagland et al., 1993). The effect of atrazine on aquaticecosystems when applied at environmental concentra-tions (15–25�g L�1) has been reported in a few studies(e.g. Krieger et al., 1988; Detenbeck et al., 1996).
The effects of atrazine on lotic periphyton com-munities are even less clear. Lotic environments areusually highly dynamic systems. Linked to this naturalvariability, changes in the physiology of periphytonwhich may in turn affect or even obscure the response totoxic substances, may occur. Jurgensen and Hoagland(1990) introduced short-term pulses of atrazine (at 0–100�g L�1) in a low order stream. In this study, runoff-related sediment deposition and associated fluctuationsin stream flow had a much more important impact onstream periphyton than short-term pulses of atrazine.Lynch et al. (1985) found that effects associated withseasonal changes had a greater influence on modelstream periphyton communities than 30-day treatmentswith atrazine. Therefore, toxicity tests performed withperiphyton communities of lotic ecosystems are nec-essary in order to assess the natural variability in theresponse to chemicals.
In order to reduce the heterogeneity occurring onnatural substrata (Townsend, 1989) and to obtain highnumbers of replicates, samples with periphyton may beobtained from colonized artificial substrates. It is notsufficiently known whether changes associated withcolonization might influence the sensitivity of the com-munity to toxicants. Throughout colonization, succes-sional processes might be accompanied by changes inthe physiology, species composition, as well as bio-mass accrual of the community (Hill & Boston, 1991;Sabater & Romanı, 1996), which may also influencethe response to toxic substances.
The current study was performed in an undisturbedstream without traces of herbicides in its waters, andthe toxicity of the herbicide atrazine was followed withshort-term tests. Toxicity tests were performed in thelaboratory with intact periphyton communities sam-pled during two periods (winter and summer) of con-trasting environmental characteristics. During sum-mer, two sites of differing ambient light conditions(one open and the other shaded) were simultaneous-ly sampled. Artificial substrates (etched glass) wereused for colonization and atrazine toxicity was fol-lowed throughout succession.
The objectives of this study were twofold: 1. todescribe the response of periphyton communities toatrazine; 2. to follow variations in tolerance throughout
succession, under different environmental conditionsof light and temperature.
Materials and methods
Description of study site
The study was carried out in L’Avenco, an undisturbedMediterranean stream. It is located at 41�460N 2�190E,45 km N of Barcelona (NE Spain) and drains a wellforested watershed in the Montseny Natural Reserve,virtually free of human disturbance. The Avenco is anundisturbed 3rd order stream characterized by highalkalinity and nutrient-poor waters (Table 1). Thestream has a well-preserved riparian vegetation mainlymade up of alder (Alnus glutinosa).
Water velocity was measured at each sampling dateby performing short-term injections of sodium chloride(Martı & Sabater, 1996). Incident light at the streambottom was measured with a LiCor quantum sensorwith an underwater cell (Li-192SB). Dissolved oxygen,pH, water temperature and conductivity were measuredwith selective electrodes. Total alkalinity was deter-mined by potentiometric (pH) titration followed byantilogarithmic Gran (Kramer, 1982). Dissolved nutri-ents (phosphorus as orthophosphate and nitrate) wereanalyzed following APHA (1989).
Analysis of stream water residues of triazineherbicides
Analysis of stream water residues of triazine herbicideswere carried out at two occasions during this period.This investigation was focused on atrazine toxicity,but simazine was also analyzed. The motor action ofsimazine is similar to atrazine and its application asherbicide is common in the area of study. Both her-bicides were analyzed in order to know whether theperiphyton community was preexposed to this groupof herbicides or not. Samples were filtered with What-man GF/F filters and processed by solid-phase extrac-tion applying 500 mg C18 cartridges (J & W ScientificFisons) and determined using high performance liquidchromatography (Durand & Barcelo, 1990). Atrazineand simazine were identified by comparing their spe-cific retention times with known reference standards(pure atrazine and simazine). The detection level forboth herbicides was 0.1 �g L�1.
japh477.tex; 9/07/1997; 12:22; v.7; p.2

139
Table 1. Environmental conditions in the Avenco stream during the experimental period. Mean values and standarddeviation (in parentheses), minimum and maximum recorded values of irradiance, water temperature (�C), velocity,dissolved oxygen (DO)(% sat), conductivity, alkalinity, pH (at 25�C) and nutrient concentration during winter (20February–24 March) and summer (26 June–16 August).
Variable Unit Winter Summer
mean (SD) min. max. mean (SD) min. max.
Irradiance �mol m�2s�1 345 (242)1 39 650 1257 (216)2 1000 1600
103 (89)�;2 16� 271�
Temperature �C 5.1 (1.1)1 4.0 6.2 21.9 (4.6)2 16.2 30.2
Velocity cm s�1 5.6 (3.2)1 4.1 10.5 2.6 (1.9)2 0 5.8
DO % sat 99.7 (6.1)1 93 105 127 (65)2 66 240
Conductivity �S cm�1 236 (44)1 203 286 357 (91)2 217 505
Alkalinity meq L�1 2.05 (0.25)1 1.85 2.30 3.83 (1.75)2 2.46 6.90
pH 25�C 8.2 (0.1)1 8.1 8.2 8.0 (0.3)2 7.3 8.4
NO�3 -N � M 14.7 (9.2)3 4.7 16.8 19.54
SRP � M 0.134 0.15 0.06 0.27
� Values of irradiance in a nearby shaded site. 1n = 5; 2
n = 6; 3n = 2; 4
n = 1.
Experimental set-up and sampling
Colonization experiments were carried out during win-ter and summer 1995. During these two periods smalletched glass substrates (1.2 cm2 surface area) wereglued to stream boulders and placed in the stream bed.Colonization sequences are summarized in Figure 1.During winter, glass substrates were allowed to colo-nize in an open site. Colonization was followed fromearly February and glass substrates were collected ondays 9, 16, 23, 30 and 41. The summer experimentstarted on July in the same site and simultaneously ina nearby shaded site. Sampling at the two sites wasperformed on days 6, 14, 29 and 36. In order to obtainreplicate experiments, other sets of etched glass sub-strates were allowed to colonize and were sampled atsimilar colonization dates. Two sets of substrates wereleft to colonize at the end of February and in mid-Marchand collected after 20 and 11 days, respectively, in thewinter experiment, and another was left to colonizein late June and sampled after 13 days in the summerexperiment (Figure 1).
Colonized substrates were sampled in early morn-ing and transported to the laboratory in cool-boxesfilled with site water, thereby maintaining in-situ tem-perature. Incubations for the measurement of photo-synthetic activity and the effect of atrazine on photo-synthesis were carried in the laboratory within 3–6 hof sampling. Samples for the determination of chloro-phyll content were frozen and those for the quantifica-tion of algal density and species composition determi-nation were preserved in 4% formaldehyde.
Figure 1. Colonization series during the winter (top) and summer(bottom) experiments in the open site (straight line) and shaded site(broken line). Arrows indicate the time when substrates were leftcolonize (left limit) and the time when colonized substrates weresampled (right end). The length of each colonization period is givento the right of each arrow.
Photosynthesis measurement
Photosynthesis was measured using a 14C technique ina similar way to that described in Blanck and Wangberg
japh477.tex; 9/07/1997; 12:22; v.7; p.3

140
(1988). Intact glass substrates were incubated in glassvials, each containing 4 mL filtered stream water(Whatman GF/F). 50�L aliquots of NaH14CO3 (2�Ci,1.9 �g L�1) were added at 10-s intervals. Incubationswere performedat light intensity which was consideredto be near saturation: 150 �mol photon m�2 s�1 whenenvironmental light was above 100 �mol photon m�2
s�1 or 50 �mol photon m�2 s�1 when environmen-tal light was below this value (for the shaded site insummer). Previous investigations in which the pho-tosynthesis irradiance relationships of intact streamperiphyton were studied (Guasch & Sabater, 1995),showed that irradiance of saturation was 30–100 and100–380 �mol photon m�2 s�1 for open and shadedsites, respectively. Water temperature was set at therange of 2 �C of that in the stream at the time the sam-ples were collected. Samples were continuously shak-en to avoid the formation of diffusion gradients. After60 min for incorporation of the 14C, photosynthesiswas stopped by adding 100 �L formaldehyde. Previ-ous incubations indicated linearity in the 14C incorpo-ration between 30–120 min, the minimum variabilitybetween replicates being at 60 min. The non incorpo-rated carbon dioxide was eliminated by rinsing eachsubstrate in clean stream water for 10–20 s. The glasssubstrates were then transferred to scintillation vialscontaining 1 mL stream water and sonicated until allperiphyton was removed. The photosynthetic activityper surface area and per chlorophyll concentration werecalculated throughout the colonization experiments.
Tolerance quantification
Periphyton photosynthetic activity (carbon uptake persample) was measured in a concentration series ofatrazine. Test solutions were prepared by dilutinga stock solution of atrazine in acetone with fil-tered stream water. Intact glass substrates were pre-incubated for 120 min in glass vials, each containing4 mL test solution in filtered stream water (What-man GF/F). A series of eight concentrations from0 to 20 �mol L�1 atrazine was used in each testwith five replicates for each concentration. From theconcentration-effect curves generated, the atrazine tol-erance could be estimated, i.e. as the effective con-centrations inhibiting 50% (EC50) of the photosyn-thetic activity. EC50 values were quantified by log-linear interpolation giving the photosynthetic activityin atrazine-treated samples as a percentage of the aver-age activity of the controls which was set to 100%.
Figure 2. Relationship between atrazine concentration (�M) andphotosynthesis (% control) for periphyton after 13 days of colo-nization during summer in the open site (#) and shaded site ( ).Effective atrazine concentration (EC50) is calculated by log-linearinterpolation.
Figure 2 shows an example of two concentration-effectcurves.
Chlorophyll analysis.
Chlorophyll a was measured in triplicate after extrac-tion in 90% acetone and sonication for 4 min. Chloro-phyll a concentration was measured spectrophotomet-rically following Jeffrey and Humphrey (1975). Thepercentage of pheo-pigments was calculated after theaddition of HCl up to a concentration of 10�3 M(Lorenzen, 1967).
Abundance of algal species
Algal density was estimated by triplicate after sonicat-ing the colonized glass substrates. For each colonizedglass, successive sonications (20 s each) were per-formed until the algae had been removed completelyand become suspended. After each sonication, the son-icated material was removed and 2 mL distilled wateradded. The success of the procedure was checked bymicroscopy observations of the glass. The suspendedmaterial was collected in a known volume (e.g. 10 mL)of 4% formaldehyde. Subsequently, an aliquot wassedimented using a phytoplankton cuvette followingthe technique of Utermohl (1958) and counted usingan inverted microscope at 600� magnification. Cellnumbers are normalized per surface area (1.2 cm2).
japh477.tex; 9/07/1997; 12:22; v.7; p.4

141
Species composition was determined under a lightmicroscope at 1000� magnification by permanentslides of diatom frustules complemented with theobservation on aliquots of the suspended material forthe determination and quantification of non-diatomcells. Diatom frustules were obtained after the diges-tion of the samples following the procedure of Barberand Haworth (1981). Permanent slides were mountedwith a high refractive resin (Naphrax). 200–300 cellswere counted per sample. Counted taxa were referredas percentage of the total number counted. The diversi-ty of the algal community (i.e. including all the groups)was calculated with the Shannon-Wiener index usinglogarithms to base 2.
Data treatment
Differences in chlorophyll content and algal densitiesbetween sites were analyzed by one-factor analysis ofvariance (ANOVA). Relationships between the com-munity response to atrazine (EC50 values) and colo-nization time (in days), biological variables (photosyn-thetic activity, chlorophyll content, algal densities, per-centage of algal groups and species diversity) and envi-ronmental conditions (water temperature, light, pH,alkalinity and conductivity) were examined using Pear-son correlation analysis. Differences between the effectof atrazine on the photosynthesis of different commu-nities incubated simultaneously were established bytwo-factor analysis of variance (ANOVA). The analy-sis compared percentages of the control (five replicatesper concentration) for five concentrations of atrazine(from 0.2–20 �M) between the two communities.
Results
No traces of atrazine or simazine were detected in thestream water of the Avenco.
Winter experiment
Water quality characteristics during winter are given inTable 1. The hydrological (velocity), physical (watertemperature) and chemical (conductivity and alkalin-ity) variables remained relatively steady during thisperiod. Irradiance showed a progressive increase from100 to 650 �mol photon m�2 s�1.
Diatoms usually accounted for half of the totalcell number (40–48%). Green algae and cyanobacte-ria shared the remaining percentage (52–66%). Early
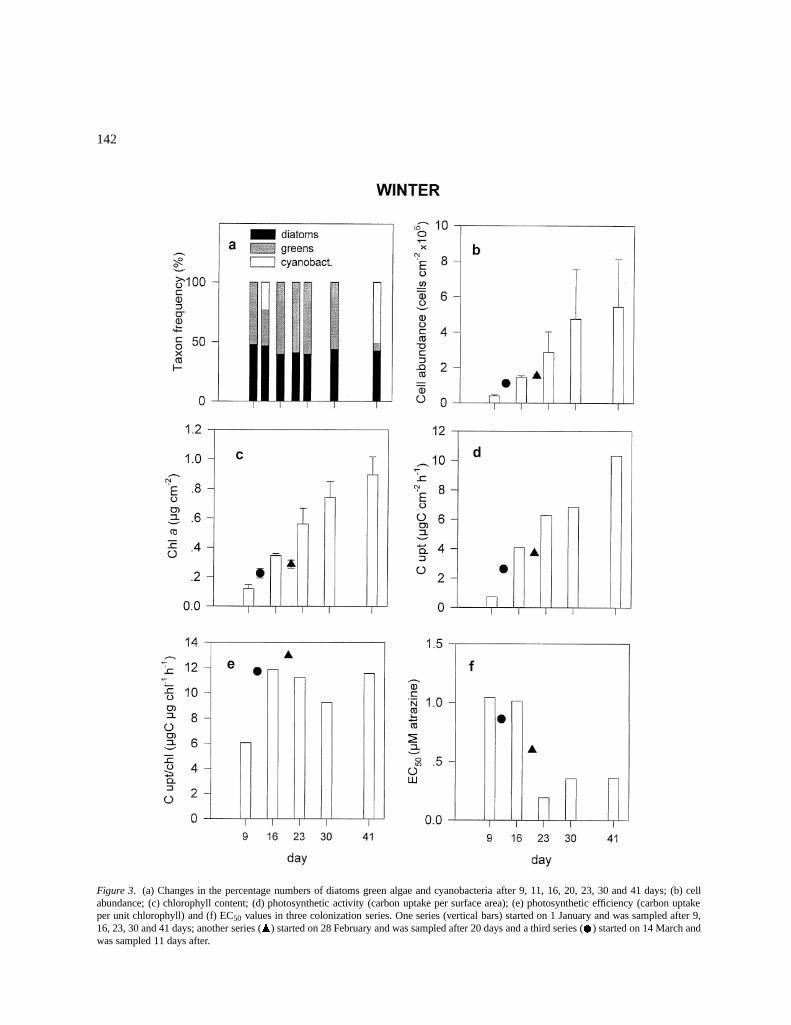
colonizers on the glass substrates were the diatomsAchnanthes spp. and Cymbella spp. and the encrust-ing green alga Gongrosira sp. Cyanobacteria becameabundant (51%) after 40 days of colonization (Fig-ure 1a). Shannon-Wiener’s index of diversity was 3.2in the first days of colonization indicating the existenceof an initial inoculum of colonizers, and declined to1.5–1.8 up to day 41.
There was a linear increase in cell abundance,chlorophyll a content and photosynthetic activi-ty throughout the colonization experiment (Fig-ures 3b, 3d). This pattern was also observed for the twosets of colonization experiments started later. This wasshown by the significant correlation between coloniza-tion day and algal density and between colonizationday and chlorophyll content when all winter data werereported together (r = 0.91, r = 0.97, n = 7, p<0.01,respectively). Photosynthetic activity per chlorophyllunit had slightly increased at day 16 and continued todo so to the end of the experiment (Figure 3e).
EC50 values for atrazine ranged between 0.19 and1.04 �M. Values above 0.8 �M were determined inthe first days of colonization (until day 16) and below0.4 �M after three weeks of colonization (Figure 3f).The lowest value was reached on day 23. There was asignificant negative correlation between the time ofcolonization and EC50 (r =�0.774, n = 7, p<0.05).EC50 was also negatively correlated with the chloro-phyll a content (r =�0.815, p<0.05) and algal density(r =�0.759, p<0.05). However, the EC50 was not cor-related with any of the environmental variables (watertemperature and light), or with any of the variablesrelated with the algal community composition (speciesdiversity and percentage of different algal groups).
Summer experiment
Environmental conditions during the summer exper-iment were highly variable (Table 1). Reduction inwater flow was accompanied by changes in waterchemistry. The main physical and chemical differencebetween the open and shaded sites was ambient irradi-ance (Table 1).
In the open site, diatoms accounted for the 54%of the cell abundance in the first days of colonization.A slight increase in the percentage of green algae wasreported in later stages (Figure 4a). Several cyanobac-teria appeared after four weeks of colonization. Thediatom community was dominated by Cocconeis spp.during the first days of colonization, and subsequent-ly by rosette-like colonies of Synedra sp. Navicula
japh477.tex; 9/07/1997; 12:22; v.7; p.5
142
Figure 3. (a) Changes in the percentage numbers of diatoms green algae and cyanobacteria after 9, 11, 16, 20, 23, 30 and 41 days; (b) cellabundance; (c) chlorophyll content; (d) photosynthetic activity (carbon uptake per surface area); (e) photosynthetic efficiency (carbon uptakeper unit chlorophyll) and (f) EC50 values in three colonization series. One series (vertical bars) started on 1 January and was sampled after 9,16, 23, 30 and 41 days; another series (N) started on 28 February and was sampled after 20 days and a third series ( ) started on 14 March andwas sampled 11 days after.
japh477.tex; 9/07/1997; 12:22; v.7; p.6

143
spp., Nitzschia spp. and Gomphonema spp. were alsocommon throughout the colonization. The green algacommunity was composed of crusts of Gongrosira sp.,small mats of Spirogyra sp., Stigeoclonium tenue Kutz.and Oedogonium sp., as well as Scenedesmus coloniesthat were common during some periods of low flow.In the shaded site, diatoms accounted for 52% of totalalgal cells on day 6, while green algae (mostly Stigeo-clonium tenue and Oedogonium) became dominant upto the end of the colonization experiment (Figure 4).
The Shannon-Wiener index of diversity showed aninitial high value (2.6 and 3.4 in the open and shad-ed sites respectively), then declined (2.4 and 2.8 onday 13) and subsequently increased again in both sites(3.4 and 3.8 on day 36).
Chlorophyll a content maintained values below1 �g cm�2 in early days of colonization (up to day 15)in both sites (Figure 4c). Values up to 2.8 �g cm�2
were recorded after 29 and 36 days of colonization inthe open site, while it was between 0.9–1.4 �g cm�2
in the shaded site. The photosynthetic activity per unitarea ranged between 2–7�gC cm�2 h�1 in the open siteand between 1–3 �gC cm�2 h�1 in the shaded site. Itexperienced a continued rise up to day 29, and later ondecreased in both sites (Figure 4d). The photosynthet-ic activity per chlorophyll unit reached ca. 5–8 �gC�g chl a�1 h�1 at the beginning of the colonizationsequence, but then declined after day 36 at both sites(Figure 4e).
In the open site, EC50 was below 0.3 �M atrazineuntil day 29 and showed a slight increase at the endof the experiment (Figure 4f). In the shaded site, EC50
values above 1.5 �M were observed in two occasionsin the early days of colonization (days 13 and 14),while EC50 values below 0.4 �M were obtained at lat-er stages of the colonization sequence. Comparisonbetween sites showed that after one week (day 6) andtwo weeks (days 13 and 14), atrazine toxicity wasslightly higher in the open than in the shaded site(ANOVA, F = 4.27, p<0.05; F = 4.15, p<0.01, andF = 4.26, p<0.005, respectively). Differences were notsignificant on day 29, and EC50 was slightly higherin the open than the shaded site on day 36 (ANOVA,F = 4.0, p<0.05). No correlation was obtained betweenEC50 and the environmental or biological variables.
Discussion
This investigation shows that changes in the physiolo-gy and taxonomic structure of periphyton communities
throughout succession may influence their response toatrazine. A negative relationship between the age ofthe community (colonization time) and tolerance toatrazine (EC50 values) was observed in winter. Thelowest EC50 value was reached after 23 days of col-onization and remained below 0.5 �M up to the endof the experiment. During the experiment the algalcommunity (made up of early colonizers-diatoms) wasreplaced by less diverse and more mature communityin which green algae and cyanobacteria had a higherpercentage. Differences in tolerance to atrazine amongdifferent groups of periphytic algae may be inferredfrom long-term effects on the species composition. Ingeneral, exposure of periphyton to atrazine tends toshift community composition from dominance by fil-amentous green-algae or cyanobacteria to dominanceby diatoms (e.g. Hamala & Kollig, 1985; Juttner et al.,1995). These results indicate that tolerance profiles ofearly colonizers probably do not reflect the response ofthe mature community. Studies aiming to describe theresponse of the natural community to toxicants shouldinclude colonization times of sufficient length to devel-op a community similar to the natural one.
Microbenthic communities in L’Avenco had notbeen pre-exposed to atrazine (no traces of atrazineor simazine were detected in the stream water) buttheir EC50 values for atrazine ranged from 0.19 �M(31 �g L�1) for the old winter community, to 2.08 �M(447�g L�1) for the shade-adapted summer communi-ty. Similar ranges of effective atrazine concentrationshave been described for phytoplankton and periphytoncommunities (Solomon et al., 1996). A low level effectof atrazine (15–25 �g L�1) on periphyton productivityand biomass have been reported in some field stud-ies (e.g. Krieger et al., 1988; Detenbeck et al., 1996).However, other experiments performed in mesocosmsand artificial streams have shown no effect at 25�g L�1
(Lynch et al., 1985) and the first effect at 130–180 �gL�1 (Juttner et al., 1995; Krieger et al., 1988).
Differences in tolerance could be explained partial-ly by differences in environmental conditions duringgrowth. During the summer experiment, environmen-tal conditions in the stream (water temperature, dis-solved oxygen and alkalinity) fluctuated according tovariations in the water flow. These variations proba-bly affected the physiological behaviour of the algalcommunities and hence contributed to mask their eco-toxicological responses to atrazine. This could partial-ly explain why differences in atrazine toxicity betweenyoung and more mature communities were not detectedat that time. However, it is difficult to distinguish which
japh477.tex; 9/07/1997; 12:22; v.7; p.7

144
Figure 4. Changes in (a) percentage numbers of diatoms and green algae after 6, 13, 14, 29 and 36 days of colonization in the open (leftbar) and shaded (right bar) sites; (b) cell abundance; (c) chlorophyll content; (d) photosynthetic activity (carbon uptake per surface area);(e) photosynthetic efficiency (carbon uptake per unit chlorophyll) and (f) EC50 values of two colonization series. One series was started inmid-June and sampled on days 6, 14, 29 and 36 (vertical bars) and the other started in late June and was sampled on day 13 in the open (#) andshaded ( ) sites.
japh477.tex; 9/07/1997; 12:22; v.7; p.8

145
variable or variables may explain the temporal varia-tion in atrazine toxicity reported here. Atrazine toxicitywas higher in the open than in the shaded sites of thesummer experiment (averages of 0.29 and 0.89 �Mrespectively), and also it was lower in winter than insummer when the open sites were compared (averagesof 0.63 and 0.29�M respectively). When the EC50 datafrom the winter and summer experiments were relat-ed with irradiance a negative correlation was detect-ed (r =�0.464, n = 20, p<0.05). This suggests thatlight history may have an important role in influenc-ing atrazine toxicity in periphyton communities. Therelationship between the toxicity of triazine herbicidesand light conditions during growth has been reportedin algal cultures. Mayasich et al. (1986) reported dif-ferences in atrazine-induced inhibition of growth fortwo algal cultures grown under different light condi-tions. Millie et al. (1992) found a negative relationshipbetween EC50 values of simazine (a triazine-herbicide)for oxygen evolution and growth rate and light condi-tions during growth of populations of Anabaena circi-nalis. They concluded that differences in sensitivityto photosystem II herbicides among photoacclimatedpopulations were related to differences in pigment con-tents. These results indicate that environmental condi-tions may affect the ecotoxicological response of nat-ural communities, specifically periphytic algae grow-ing in lotic systems. A range in sensitivity to an inducedstress is perhaps characteristic of natural assemblagesof micro- organisms and emphasizes the importance ofusing natural communities to assess the effect of toxicchemicals in real-world environments.
Acknowledgements
This work was supported financially by the EC project(EN AA 13496). The ‘Serveis cientıfico tecnics de laUniversitat de Barcelona’ provided facilities and tech-nical help with HPLC analysis. We especially thankProf. W. Admiraal and V. Lehmann for reviewing anearlier draft of the manuscript. An anonymous refereeprovided very useful comments to the manuscript.
References
American Public Health Association (APHA) (1989) Standard Meth-ods for the Examination of Water and Wastewater. 17th edn,Washington DC.
Barber HG, Haworth EY (1981) A Guide to the Morphology of theDiatom Frstule. Sci. Publs Freshwat. Biol. Ass. U.K. 44, 112 pp.
Blanck H, Wangberg SA (1988) Induced community tolerance inmarine periphyton established under arsenate stress. Can. J. Fish.aquat. Sci. 45: 1816–1819.
Blanck H, Wangberg SA, Molander S (1988) Pollution-InducedCommunity Tolerance – A New Ecotoxicological Tool. In Func-tional testing of aquatic biota for estimating hazards of chemicals.ASTM STP 988. Cairns J, Jr, Pratt JR (eds), American Societyfor Testing and Materials. Philadelphia, 219–230.
Brockway DL, Smith PD, Stancil FE (1984) Fate and effects ofatrazine in small aquatic microcosms. Bull. envir. Contam. Toxi-col. 32: 345–353.
Detenbeck EN, Hermanutz R, Allen K, Schwift MC (1996) Fateand effects of the herbicide atrazine in flow-through wetlandmesocosms. Envir. Toxicol. Chem. 15: 937–946.
Durand D, Barcelo D (1990) Determination of chlorotriazinesand their photolysis products by liquid chromatography withphotodiode-array and thermospray mass spectrometric detection.J. Chromat. 502: 275–286.
Guasch H, Sabater S (1995) Seasonal variation in photosynthesis-irradiance responses by biofilms in Mediterranean streams. J.Phycol. 31: 725–735.
Hamala JA, Kollig HP (1985) The effects of atrazine on periphytoncommunities in controlled laboratory ecosystems. Chemosphere14: 1391–1408.
Hamilton PB, Jackson GS, Kaushik NK, Solomon KR, Stephen-son GL (1988) The impact of two applications of atrazine on theplankton communities of in situ enclosures. Aquat. Toxicol. 13:123–140.
Hill WR, Boston HL (1991) Effects of community development onphotosynthesis irradiance relations in stream periphyton. Limnol.Oceanogr. 36: 1375–1389.
Hoagland KD, Drenner RW, Smith JD, Cross DR (1993) Freshwa-ter community responses to mixtures of agricultural pesticides:Effects of atrazine and bifenthrin. Envir. Toxicol. Chem. 12: 627–637.
Huber W (1993) Ecotoxicological relevance of atrazine in aquaticsystems. Envir. Toxicol. Chem. 12: 1865–1881.
Jeffrey S, Humphrey GF (1975) New spectrophotometric equationsfor determining chorophylls a, b, c1 and c2 in higher plants, algaeand natural phytoplankton. Biochem. Physiol. Pflanzen 167: 191–194.
Jurgensen TA, Hoagland KD (1990) Effects of short-term pulses ofatrazine on attached algal communities in a small stream. Arch.envir. Contam. Toxicol. 19: 617–623.
Juttner I, Peither A, Lay JP, Kettrup A, Ormerod SJ (1995) Anoutdoor mesocosm study to assess ecotoxicological effects ofatrazine on a natural plankton community. Arch. envir. Contam.Toxicol. 29: 435–441.
Kosinski R, Merkle MG (1984) The effect of four terrestrial herbi-cides on the productivity of artificial stream algal communities.J. envir. Qual. 13: 75–82.
Kramer JR (1982) Alkalinity and acidity. In Water Analysis, Acad-emic Press, 131–134.
Krieger KA, Baker DB, Kramer JW (1988) Effects of herbicides onstream Aufwuchs productivity and nutrient uptake. Arch. envir.Contam. Toxicol. 17: 299–306.
Lewis MA, Pittinges CA, Davidson DH, Ritchie CJ (1993) In situresponse of natural periphyton to an anionic surfactant and anenvironmental risk assessment for phytotoxic effects. Envir. Tox-icol. Chem. 12:1803–1812.
Lorenzen C (1967) Determination of chlorophyll and pheo-pigments: spectrophotometric equations. Limnol. Oceanogr. 12:343–346.
japh477.tex; 9/07/1997; 12:22; v.7; p.9

146
Lynch T, Johnson HE, Adams WJ (1985) Impact of atrazine andhexaclorobiphenyl on the structure and function of model streamecosystems. Envir. Toxicol. Chem. 4: 399–413.
Martı E, Sabater F (1996) High variability in temporal and spatialnutrient retention in Mediterranean streams. Ecology 77: 854–869.
Mayasich JM, Karlander EP, Terlizzi DE, Jr (1986) Growth responsesof Nannochloris oculata Droop and Phaeodactylum tricornutumBohlin to the herbicide atrazine as influenced by light intensityand temperature. Aquat. Toxicol. 8: 175–184.
Millie DF, Harrsh CM, Dionigri CP (1992) Simazine-induced inhi-bition in photoacclimated populations of Anabaena circinalis(Cyanophyta). J. Phycol. 28: 19–26.
Niederlehner BR, Cairns J, Jr (1993) Effects of previous zinc expo-sure on pH tolerance of periphyton communities. Envir. Toxicol.Chem. 12: 743–753.
Nyholm N, Kallqvist T (1989) Methods for growth inhibition toxicitytests with freshwater algae. Envir. Toxicol. Chem. 8: 689–703.
Plumley F, Davis DE (1980) The effect of a photosynthesis inhibitor,atrazine, on salt marsh edaphic algae, in culture, microecosys-tems, and in the field. Estuaries 3: 217–223.
Sabater S, Romanı A (1996) Metabolic changes associated withbiofilm formation in an undisturbed Mediterranean stream.Hydrobiologia 335: 107–113.
Solomon KR, Baker DB, Richards RP, Dixon KR, Klaine SJ, LaPoint TW, Kendall RJ, Weiskopf CP, Giddings JM, Giesy JP,Hall LW, Marty Williams W (1996) Ecological risk assessmentof atrazine in North American surface waters. Envir. Toxicol.Chem. 15: 31–76.
Townsend CR (1989) The patch dynamics concept of stream com-munity ecology. J. North am. Benth. Soc. 8: 36–60.
Utermohl H (1958) Zur Vervollkommung der quantitativenPhytoplankton-Methodik. Mitt. int. Ver. Limnol. 9: 1–39.
Whitton BA, Kelly MG (1995) Use of algae and other plants formonitoring rivers. Austral. J. Ecol. 20: 45–56.
japh477.tex; 9/07/1997; 12:22; v.7; p.10
Copyright © 2022 FDOKUMEN




















