
Development of models to predict Pinus radiata productivity throughout New Zealand

© 2002 Blackwell Science Ltd
Prokaryotic genetic diversity throughout the salinitygradient of a coastal solar saltern
alpha- and gamma-Proteobacteria, cyanobacteria andCFB were the only groups found, and most of themwere related to specialized halophilic bacteria. Fromthe 32% salt pond, only CFB were found, and most ofthe sequences retrieved clustered with Salinibacterruber, an extremely halophilic bacterium. A decreasein the richness of bacterial genera was thereforeapparent along the gradient. Archaea behaved quitesimilarly. In the lowest salinity ponds, sequenceswere related to environmental clones of MarineArchaea Group III (Thermoplasmales relatives) and tounclassified branches of Euryarchaeaota. In the 8%,22% and 32% ponds, most of the clones were relatedto different cultured strains of Halobacteriaceae.Finally, most sequences from the crystallizers clus-tered with the uncultured square archaeon SPhT.Crenarchaeaota were not detected. Despite the factthat higher prokaryotic richness was apparent in thelower salinity ponds than in the crystallizers, thediversity index from clone libraries calculated accord-ing to Shannon and Weaver did not show this trend.This was because diversity in the crystallizers can be considered as ‘microdiversity’, the co-existence ofseveral closely related clones of Bacteria (the S. rubercluster) and Archaea (the SPhT cluster). Regardlessof the changes in abundance, both Bacteria andArchaea showed the same pattern; as salinity in-creased, the number of different clusters decreased,and only one cluster became dominant. Both clusters,however, showed a considerable degree of microdi-versity. The meaning of such microdiversity remainsto be determined.
Introduction
Multipond salterns are semi-artificial coastal systemsdesigned to harvest common salt (NaCl) from sea water.To achieve this purpose, sea water is pumped through aset of shallow ponds where water evaporates and saltsconcentrate. Sequential precipitation of CaCO3 andCaSO4 occurs during the first stages. Finally, when seawater has been reduced to about 1/10th the originalvolume, NaCl precipitates and can be collected. Thesesystems often operate as continuous or semi-continuoussystems, so that each set of ponds maintains a similarrange of salinity throughout the year. Important ecologicalchanges occur through this gradient, with decreasing
Environmental Microbiology (2002) 4(6), 349–360
Susana Benlloch,1‡ Arantxa López-López,1‡ Emilio O. Casamayor,2*† Lise Øvreås,3 Victoria Goddard,4
Frida Lise Daae,3 Gary Smerdon,4 Ramón Massana,2
Ian Joint,4 Frede Thingstad,3 Carlos Pedrós-Alió2
and Francisco Rodríguez-Valera1
1División de Microbiología, Universidad MiguelHernández, Alicante, Spain.2Departament de Biologia Marina i Oceanografia,Institut de Ciències del Mar, CMIMA-CSIC, Barcelona,Spain.3Department of Microbiology, University of Bergen,Bergen, Norway.4NERC Plymouth Marine Laboratory, Prospect Place,The Hoe, Plymouth, UK.
Summary
Bacterial and archaeal assemblages have beenstudied in a multipond solar saltern using a range ofmicrobial ecology techniques by four laboratoriessimultaneously. These include 16S rDNA sequencingfrom both denaturing gradient gel electrophoresis(DGGE) and clone libraries, and culturing methods.Water samples from eight ponds were analysed, cov-ering a salinity range from near sea water (4% salt) to saturated sodium chloride (37% salt; ponds calledcrystallizers). Clone libraries focused on ponds withsalinity of 8%, 22% and 32%. Although differentcloning strategies were able to retrieve the same typeof dominant sequences, there were differing degreesof success with less abundant sequences. Thus, theuse of two sets of primers recovered a higher numberof phylotypes. Bacterial and archaeal isolates were,however, different from any of the retrieved environ-mental sequences. For Bacteria, most sequences inthe 8% salt pond were related to organisms of marineorigin. Thus, representatives of the alpha-, beta-,gamma- and epsilon-subdivisions of Proteobacteria,the Cytophaga–Flavobacterium–Bacteroides group(CFB), high-G+C Gram-positive bacteria andcyanobacteria were found. In the 22% salt pond,
Received 4 April, 2002; accepted 4 April, 2002. *For correspondence.E-mail [email protected]; Tel. (+33) 468 88 73 42; Fax (+33) 468 88 73 98. †Present address: Observatoire Océanologique deBanyuls–CNRS, BP 44, F-66651 Banyuls-sur-Mer, France. ‡Theseauthors have contributed equally to this work.

biodiversity with increasing salinity and the dominance of microbes (Rodríguez-Valera, 1988; 1993; Javor, 1989).There are a large number of traditional studies on theecology of multipond salterns, to the point where they canbe considered model systems for the variations inducedby one environmental factor (Pedrós-Alió et al., 2000).These ecosystems are nice and very interesting modelsystems to investigate the influence of salinity on micro-bial diversity and productivity (Joint et al., 2002).
The multipond solar salterns in Santa Pola (Alicante,Spain) have been studied extensively for several years.Studies based on culturing showed a decrease in prokary-otic diversity recovered on plates as salt concentrationincreased (Rodríguez-Valera et al., 1981; 1985). At salin-ity up to 15%, most isolates were those commonly foundin sea water. Above 15% salt, most strains were special-ized halophilic. However, when a clone library from a crys-tallizer pond (salinity around NaCl saturation) was carriedout, the most frequent 16S rDNA archaeal sequence (theSPhT phylotype) did not correspond to any previouslyknown microorganism (Benlloch et al., 1995a). In addi-tion, fluorescence in situ hybridization (FISH) indicatedthat such a phylotype together with a new extremelyhalophilic bacterium, Salinibacter ruber (Antón et al.,2002), accounted for most of the microbial biomass in thecrystallizers (Antón et al., 1999; 2000). To date, the bestcharacterization of prokaryotic assemblages from SantaPola salterns has been done in the crystallizers. Pondswith lower salinity have been explored occasionally, but a detailed view of changes in prokaryotic populations inthe transition from sea water to salt-saturated watermasses has been not carried out. A general view usingfingerprinting techniques has previously indicated majorchanges at salinities of 10% and 22% and in those pondsclose to salt saturation (Martínez-Murcia et al., 1995;Casamayor et al., 2000a).
The present study represents a major effort to comparedifferent approaches carried out by four laboratories in thestudy of microbial ecology in Santa Pola salterns, and wasundertaken within the European Union project MIDAS.Cultivation and two major 16S rDNA-based molecularmethods were used in order to expand our knowledge ofthe changes in biodiversity along an increasing salt gra-dient. Two laboratories made clone libraries for both Bac-teria and Archaea: the Plymouth Marine Laboratory (PML)and the University Miguel Hernández of Alicante (UMH).Two different combinations of primers were used by eachlaboratory in three selected ponds (8%, 22% and 32%salinity). In parallel, two laboratories used denaturing gradient gel electrophoresis (DGGE) and sequencing ofexcised bands along the whole salinity gradient (4–37%salinity): the University of Bergen (UiB) and the MarineScience Institute of Barcelona (ICM). The study allowedcomparison of analytical approaches among them and
with cultivation, and expanded the range of known diver-sity in the salterns with new clusters of hitherto unculturedmicroorganisms. In addition, 16S rDNA genetic diversityshifted from ‘macrodiverse’ to ‘microdiverse’ microbialassemblages along the salinity gradient.
Results
Analysis of bacterial assemblages – DGGE fingerprints
A bacterial DGGE gel covering the salinity gradient from4% to 37% is shown in Fig. 1. Up to 63 bands were
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360
350 S. Benlloch et al.
Fig. 1. Bacterial 16S rDNA DGGE profile obtained by the UiBlaboratory along the salinity gradient. Bands marked with anumbered dot were excised from the gel, reamplified andsequenced. The closest relatives for these sequences are reportedin Tables 1–3 except for bands 18 and 21 (94.9% similarity tomarine cyanobacteria HstpI4), bands 58 and 60 (97.9% similarity toSalinibacter ruber), band 40 (74.8% similarity to Psychroflexustorquis) and band 43 (76.2% similarity to Bacillus halophilus).

Prokaryotes in a saltern 351
excised from the gel, but only 27 bands producedsequences that could be used satisfactorily in the BLAST
analysis (bands marked with numbered dots in Fig. 1).The identity of such bands is shown in Tables 1–3 (wherethe code for the bands includes the number shown in Fig. 1 followed by UiB). We observed progressivechanges in band patterns from one pond to the next thatwere related to different bacterial populations with over-lapping salinity ranges. Thus, a relative of the unculturedmarine gamma-Proteobacterium CHAB-III-7 (98.5% similarity in 16S rDNA sequence) was detected from 4%to 15% salinity ponds (bands 42, 46 and 52), and a rela-tive of the uncultured gamma-Proteobacterium BCP022(88% similarity) was detected between 11% and 37%ponds (bands 2–53). Finally, most of the bands between22% and 37% salinity were related to Salinibacter ruber.We found only one band present throughout the salt gra-dient (band 38, present in all ponds; 79.9% similarity toFlexibacter sp.). Conversely, several bands were found inone or two ponds. Such bands were related to the
gamma-Proteobacterium Thiomicrospira sp. in the 8%pond, to Nitrococcus mobilis in the 22% pond, to the high-G+C, Gram-positive Agrococcus jenensis in the 8–11%ponds, and to Bacillus halophilus in the 5% pond. Thecase of the 22% pond seems to be peculiar, as most ofthe bands formed a cluster whose closest cultured rela-tive was Nitrococcus mobilis (89% similarity, see Table 2).Such sequences had similarities among themselves in therange 94.8–98.7% and might represent a hitherto undis-covered new group of halophilic gamma-Proteobacteria.In parallel, we observed a similar trend in the 32% pond:most of the bands clustered with the recently discoveredS. ruber, a new group of extremely halophilic Cytopha-gales. Such sequences had similarities in the range97–99.8% among themselves. As shown below, severalsequences closely related to Nitrococcus and Salinibac-ter were also obtained from the same ponds by cloningand after using different primer combinations.
On the basis of DGGE results, three ponds wereselected for clone libraries. We chose first the 8% pond
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360
Table 1. Bacterial and archaeal 16S rDNA sequences from the 8% salinity pond.
Group Sequences Closest relative % similarity range
Alpha-Proteobacteria 5-PML Antarctobacter heliothermus, hypersaline lake, Y11552 96.924-UMH Roseobacter denitrificans, M59063 97.61/3-PML; 26-UMH Uncultured ZD0410 from algal bloom, AJ400351 88.2–90.0A72 Paracoccus denitrificans, Y16927 95.0
Beta-Proteobacteria 39-UMH Alcaligenes sp., AJ002808 95.5Gamma-Proteobacteria 17-UMH Nitrococcus mobilis, L35510 87.0
32/70-UMH Clone CHAB-III-7, Mediterranean Sea, AJ240921 93.5–96.223/33/40-UMH; 42/46/52-UiB Clone CHAB-III-7, Mediterranean Sea, AJ240921 97.1–98.545-UiB Thiomicrospira sp. strain SL-1, AJ011066 92.1A1 Halomonas halmophila, M59153 95.1A6/A12 Salinivibrio costicola ssp. vallismortis, AF057016 94.8–95.3A16 Pseudoalteromonas piscicida, AF297662 95.0A45 Alteromonas macleodii, X82145 95.0A58 Marinomonas vaga, X67025 95.6A74 Marinobacter hydrocarbonoclasticus, Y16735 95.5
Epsilon-Proteobacteria 10-UMH Sulphurospirillum arcachonense, marine sediment, Y11561 91.0CFB phylum 7-PML Cytophaga sp., AB007009 92.2
38-UiB Flexibacter sp., AF034938 79.9A4 Psychroflexus torquis, AF001365 97.2
High-GC, Gram-positive 4-PML Microthrix parvicella, X82546 88.728-UMH; 3/54-UiB Agrococcus jenensis, soil and sandstone surface, X92492 91.5–94.3
Cyanobacteria and plasts 35-UMH Dactylococcopsis sp. plankton Solar Lake, AJ000711 99.062-UiB Spyrogyra maxima chloroplast, U24596 90.1
Unclassified Euryarchaeota 2/3/5-PML Clone pISA35 deep-sea hydrothermal vent, AB019748 74.0–90.010-PML Clone JTA69 sediment, AB015280 86.011-PML Clone S15-6 anoxic rice field soil, AJ236487 85.06/7-PML DGGE band CIARC-3 sulphide-rich lake, AJ239990 81.0–83.042/52-UMH Clone J4.75-24, hypersaline lake, AF199377 80.6–82.0
Rice cluster V 4/8-PML Clone Rot13 sediment freshwater lake, Y18089 83.0–84.0Group III Archaea 9-PML Clone PMC2A24 hydrothermal vent, AB019736 84
12-PML Clone HA11 landfill site, AB046195 9550/60-UMH Clone TA1f2, marine sediment, AF134390 80.0–94.0
Halobacteriaceae 63-UMH Natronomonas pharaonis, D87971 86.944-UMH Halorubrum sodomense, D13379 98.051-UMH Haloarcula mukohatei, D50850 90.011-UMH Clone organic1-A, saline lake, AF142985 90.0
PML, Plymouth clone libraries; UMH, Alicante clone libraries; UiB, DGGE bands from the Bergen group; A, isolates grown on plates.Haloarchaeal sequences are included in the Halobacteriaceae family.
as an example of the lower salinity ponds (Casamayoret al., 2002a). Here, most sequences retrieved fromDGGE were related to marine microorganisms and tomoderately halophilic representatives such as the Thiomicrospira sp. strain SL-1, isolated from Solar Lake(Brinkhoff and Muyzer, 1997). Secondly, we chose the32% pond as a representative of the higher salinity ponds,in which most DGGE bands corresponded to S. ruber.Finally, we selected the 22% salinity pond as a transitionalpond between the two main salinity groups. Here, mostsequences from DGGE were distantly related to both cultured strains (e.g. 82% similarity in 16S rDNA to
Psychroflexus torquis or 89% to N. mobilis) and environmental clones (e.g. 88% to uncultured gamma-Proteobacterium BCP022).
Analysis of bacterial assemblages – clone libraries
Clone libraries were constructed by UMH and PMLlaboratories and screened by restriction fragment length polymorphism (RFLP) and DGGE respectively. In total,105 bacterial clones were obtained (63 by UMH and 42 by PML), corresponding to 42 different patterns, i.e.operational taxonomic units (OTUs) (Table 4). In addition,
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360
352 S. Benlloch et al.
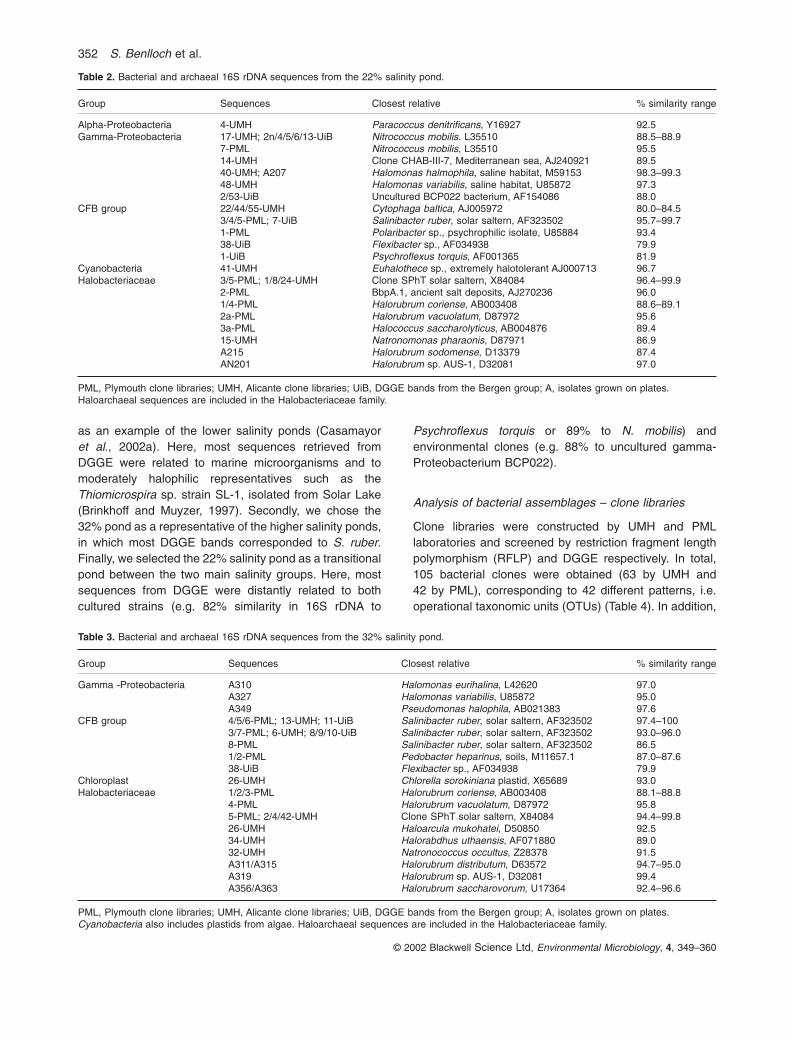
Table 2. Bacterial and archaeal 16S rDNA sequences from the 22% salinity pond.
Group Sequences Closest relative % similarity range
Alpha-Proteobacteria 4-UMH Paracoccus denitrificans, Y16927 92.5Gamma-Proteobacteria 17-UMH; 2n/4/5/6/13-UiB Nitrococcus mobilis. L35510 88.5–88.9
7-PML Nitrococcus mobilis, L35510 95.514-UMH Clone CHAB-III-7, Mediterranean sea, AJ240921 89.540-UMH; A207 Halomonas halmophila, saline habitat, M59153 98.3–99.348-UMH Halomonas variabilis, saline habitat, U85872 97.32/53-UiB Uncultured BCP022 bacterium, AF154086 88.0
CFB group 22/44/55-UMH Cytophaga baltica, AJ005972 80.0–84.53/4/5-PML; 7-UiB Salinibacter ruber, solar saltern, AF323502 95.7–99.71-PML Polaribacter sp., psychrophilic isolate, U85884 93.438-UiB Flexibacter sp., AF034938 79.91-UiB Psychroflexus torquis, AF001365 81.9
Cyanobacteria 41-UMH Euhalothece sp., extremely halotolerant AJ000713 96.7Halobacteriaceae 3/5-PML; 1/8/24-UMH Clone SPhT solar saltern, X84084 96.4–99.9
2-PML BbpA.1, ancient salt deposits, AJ270236 96.01/4-PML Halorubrum coriense, AB003408 88.6–89.12a-PML Halorubrum vacuolatum, D87972 95.63a-PML Halococcus saccharolyticus, AB004876 89.415-UMH Natronomonas pharaonis, D87971 86.9A215 Halorubrum sodomense, D13379 87.4AN201 Halorubrum sp. AUS-1, D32081 97.0
PML, Plymouth clone libraries; UMH, Alicante clone libraries; UiB, DGGE bands from the Bergen group; A, isolates grown on plates.Haloarchaeal sequences are included in the Halobacteriaceae family.
Table 3. Bacterial and archaeal 16S rDNA sequences from the 32% salinity pond.
Group Sequences Closest relative % similarity range
Gamma -Proteobacteria A310 Halomonas eurihalina, L42620 97.0A327 Halomonas variabilis, U85872 95.0A349 Pseudomonas halophila, AB021383 97.6
CFB group 4/5/6-PML; 13-UMH; 11-UiB Salinibacter ruber, solar saltern, AF323502 97.4–1003/7-PML; 6-UMH; 8/9/10-UiB Salinibacter ruber, solar saltern, AF323502 93.0–96.08-PML Salinibacter ruber, solar saltern, AF323502 86.51/2-PML Pedobacter heparinus, soils, M11657.1 87.0–87.638-UiB Flexibacter sp., AF034938 79.9
Chloroplast 26-UMH Chlorella sorokiniana plastid, X65689 93.0Halobacteriaceae 1/2/3-PML Halorubrum coriense, AB003408 88.1–88.8
4-PML Halorubrum vacuolatum, D87972 95.85-PML; 2/4/42-UMH Clone SPhT solar saltern, X84084 94.4–99.826-UMH Haloarcula mukohatei, D50850 92.534-UMH Halorabdhus uthaensis, AF071880 89.032-UMH Natronococcus occultus, Z28378 91.5A311/A315 Halorubrum distributum, D63572 94.7–95.0A319 Halorubrum sp. AUS-1, D32081 99.4A356/A363 Halorubrum saccharovorum, U17364 92.4–96.6
PML, Plymouth clone libraries; UMH, Alicante clone libraries; UiB, DGGE bands from the Bergen group; A, isolates grown on plates.Cyanobacteria also includes plastids from algae. Haloarchaeal sequences are included in the Halobacteriaceae family.

Prokaryotes in a saltern 353
51 pure bacterial cultures were isolated from three ponds,corresponding to 13 different RFLP patterns. One repre-sentative of each OTU was sequenced independently(Escherichia coli positions 150–600 by UMH and 1055–1401 by PML). Results for each of the three ponds aredisplayed in Tables 1–3 respectively.
Sequences retrieved from the 8% pond were distributedin several phylogenetic groups: alpha, beta, gamma and epsilon subdivisions of Proteobacteria, Cytophaga–Flavobacterium–Bacteroides (CFB) phylum, high-G+CGram-positive and cyanobacteria (Fig. 2). Most se-quences were related to organisms of marine origin. Of 17 sequences from the 8% pond, six clustered withmembers of gamma-Proteobacteria, five with members of
alpha-Proteobacteria, two sequences with high-G+CGram-positive and one sequence with each of the remain-ing groups (Table 1). New clusters of alpha- and gamma-Proteobacteria distantly related to previous sequences inthe database were found (Fig. 3). However, cultures (represented by A- followed by a number) were not representative of such diversity and were mostly relatedto well-known gamma-Proteobacteria (up to 90% of the cultures obtained) and to well-known alpha-Proteobacteria and CFB.
Sequences retrieved from the 22% pond showed lower phylogenetic diversity: alpha- and gamma-Proteobacteria, cyanobacteria and CFB were the groupsfound (Fig. 2). Thus, of 14 sequences obtained, sevenclustered with CFB, five with gamma-Proteobacteria, andonly one representative of each of the other two groupscould be retrieved (Table 2). Most of the sequences wererelated to specialized halophilic bacteria. As noted aboveusing DGGE, we found by cloning a cluster of sequencesdistantly related to N. mobilis (Fig. 3). Unfortunately, allbacterial cultures obtained from the 22% pond wereclosely related to Halomonas halmophila and, therefore,a representative of such an unidentified halophilicgamma-Proteobacteria cluster could not be grown in theconditions tested. The same happened in the case of the CFB phylum; a cluster distantly related to marinecytophagas (similarity in 16S < 85%) was detected, butno isolate was recovered from such a halophilic CFBcluster (Fig. 3).
Finally, clones from the 32% pond affiliated only withthe CFB group and with plastids from algae, probablyDunaliella salina (Fig. 2, Table 3). From 11 sequencesretrieved in this pond, eight clustered with S. ruber. Theplastid sequence probably corresponded to D. salina, the only alga present in the crystallizers. All bacterial cultures here were closely related to well-known gamma-Proteobacteria, but S. ruber could not be isolated fromany pond. In recent studies, this bacterium has been
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360
Table 4. Summary information for the clone libraries constructed in the present study.
Library Salinity (%) OTUs Clones Coverage (%) Shannon’s index
PML Bacteria 8 5 9 78 1.52322 5 14 100 1.53232 8 19 74 1.704
UMH Bacteria 8 12 22 82 2.41322 9 29 86 1.89132 3 12 83 0.566
PML Archaea 8 11 17 59 2.28222 7 31 87 1.49432 5 10 70 1.418
UMH Archaea 8 8 18 72 1.78522 4 9 67 1.00332 6 11 73 1.673
Different OTUs correspond to different RFLPs (UMH library) or positions in a DGGE gel (PML library) for the clones analysed. Coverage valuesof clone libraries calculated according to Mullins et al. (1995).
Fig. 2. Distribution of 16S rDNA clones derived from the watercolumn of the 8%, 22% and 32% salinity ponds across bacterialand archaeal phylogenetic groups. Values are expressed aspercentages for the sum of UMH and PML clone libraries.

isolated easily from salinities between 22% and 37%(Antón et al., 2002). Obviously, the growth conditionsused in our work did not allow its growth.
Analysis of archaeal assemblages – DGGE fingerprint
Two DGGE were run, one from a direct polymerase chainreaction (PCR) amplification and the other from a nestedPCR amplification. Direct PCR yielded suitable productsfor DGGE analysis for most samples. However, theamount and quality of the PCR product from the 4%, 5%and 8% ponds were insufficient either to obtain a goodDGGE profile or to extract and reamplify bands for a sub-sequent sequencing study. Therefore, nested PCR wasperformed for such initial ponds (see Fig. 4). AlthoughPCR-independent quantitative data are not available, thelow yield of PCR products is consistent with low archaealbiomass in the low-salinity ponds. After direct PCR, thevisual analysis of DGGE gels showed two groups of rela-tively similar band patterns: 11% and 15% salinity ponds,on the one hand, and 22%, 32% and 37% ponds on theother (Fig. 4, right). One very intense band was obtainedfor each group: band 506-ICM (99% similarity to archaeon
JDS, a haloarchaeal clone detected in Solar Lake) andband 513-ICM (99% similarity to the uncultured squarearchaeon SPhT). Other minor bands also excised fromthe gel were also related to these closest relatives (datanot shown). After nested PCR, 10 sequences wereretrieved from the DGGE bands from the 4%, 5% and 8%ponds (Fig. 4, left). All 10 sequences showed the highestsimilarity to environmental 16S rDNA sequences of Thermoplasmales-relative Marine Archaea Group III.
Analysis of archaeal assemblages – clone libraries
The same samples selected for clone libraries of Bacte-ria were used for Archaea. Two clone libraries were con-structed, and 96 archaeal clones were obtained, including38 clones from UMH and 58 from PML libraries (Table 4).Thirty archaeal strains were isolated from the sameponds. The screening procedure was as for Bacteria, andwe found 19 different OTUs at 8% salinity, 11 OTUs at22% and 11 OTUs at 32% salinity for both clone libraries.Among the cultures, archaeal isolates were not retrievedfrom the 8% pond, two OTUs were found at 22% salinityand five OTUs at 32% salinity. One representative of each
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360
354 S. Benlloch et al.
Fig. 3. Phylogenetic associations for the most representative bacterial uncultured groups belonging to alpha- and gamma-Proteobacteria andthe CFB phylum found in the three ponds studied by clone libraries. One bacterial isolate (A-72) obtained from the 8% pond and distantlyrelated to the genus Paracoccus is included. The dendrogram was built by the neighbour-joining method using the Jukes–Cantor distanceestimation from partial 16S rDNA sequences (E. coli positions 150–600) of the UMH clone library. Numbers at nodes are bootstrap confidencevalues expressed as percentages of 500 bootstrap replications. Bootstrap values higher than 50% are indicated in the main nodes.

Prokaryotes in a saltern 355
OTU was partially sequenced (E. coli positions 85–555 byUMH and 344–915 by PML). Results for each of the threeponds are displayed in Tables 1–3 respectively. In anycase, members of the kingdom Crenarchaeaota weredetected in the water column from the ponds studied.
Sequences retrieved from the 8% pond were distantlyrelated to sequences deposited in the database (mostwithin the range 80–90% similarity in 16S rDNA). Thisresult was consistent for both libraries (Table 1). As in thecase of sequences recovered from DGGE, we observedclones related to uncultured Thermoplasmales relativeArchaea group III and clones related to other unclassifiedEuryarchaeota. Only one sequence was closely related to a cultured haloarchaea (98% similarity to Halorubrumsodomense) and, unfortunately, no archaeal isolates grew on the plates from the 8% pond. Moreover, otherclones distantly related to Halobacteriaceae were foundand could represent a new group of Archaea betteradapted to growth at lower salinity (Table 1). Thus, thispond can be considered as a reservoir of new groups of unidentified Archaea (Fig. 5) that deserves furtherresearch.
In the 22% and 32% ponds, most of the sequences thataffiliated with environmental clones did so with SPhT(Fig. 5, using PML data). The remaining clones related to
haloarchaea from the family Halobacteriaceae availablein culture (86.9–95.8% similarities in 16S rDNA; Tables 2and 3). However, none of the strains isolated in this workwas related to the SPhT phylotype. In fact, all the isolatesclustered with the Halorubrum genus, but some were not closely related and could represent novel groupswithin the Halobacteriaceae (e.g. the haloarchaeal isolate A-215, 87.4% similarity in 16S rDNA to Halorubrumsodomense).
Discussion
Comparison of methods
Changes in prokaryotic assemblages throughout thesalinity gradient were approached by two complementaryviews: DGGE followed by sequencing and clone librariesof 16S rRNA genes. DGGE is particularly good for fol-lowing changes in the community structure along the gradient, but it is only able to retrieve sequences that arepresent in at least 0.5–1% of the total cells in the sample(Muyzer et al., 1997; Casamayor et al., 2000b). Clonelibraries, on the other hand, allow the retrieval of less pre-dominant sequences. However, cloning is time consum-ing and expensive and, therefore, it is not practical to use
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360
Fig. 4. Archaeal DGGE profiles carried out atthe ICM laboratory (direct PCR) and UiBlaboratory (nested PCR) along the salinitygradient. Nested PCR was applied in theinitial ponds that yielded low-amplificationproduct by direct PCR. Bands marked with anumbered dot were excised from the gel,reamplified and sequenced. The closestrelatives for the sequences up to 8% salinitywere related to the Marine Group IIIThermoplasmales-relatives, whereas thesequences from the ponds with higher than11% salinity were related to haloarchaea.

with a large number of samples. We circumvented thisproblem to some extent by partial sequencing of theclones (500 bp). We were aware that we lost phylogeneticresolution with this study design. However, our strategyanalysed a large number of clones and, in addition, useddifferent sets of primers. Thus, some patterns of geneticdiversity and community structure arose along the salin-ity gradient. Similarities and discrepancies were alsofound between the results from PML and UMH librariesthat used different primer combinations. In both cases,there was a tendency for fewer different prokaryoticsequences as salinity increased. Also in both cases,sequences and clones from the 32% pond were over-whelmingly related to Salinibacter (i.e. 16 clones out of 19in PML and 11 out of 12 in UMH). This result is in agree-ment with FISH counts, in which over 90% of the prokary-otes stained by the eubacterial probe were also stained
with the Salinibacter probe (Antón et al., 2000). TheDGGE bands from this pond also belonged to Salinibac-ter. Another point of similarity was the retrieval of uncul-tured CFB and uncultured gamma-Proteobacteria as themost abundant sequences recovered from the bacterialbiota of the 22% pond. Finally, alpha-Proteobacteria werenumerous in both libraries and, moreover, relatives of theuncultured ZD0410 clone were well represented in both.However, we saw many more different groups retrievedin the UMH library than in the PML one, which probablyrelates to the different coverage of the primers and otherartifacts (Wintzingerode et al., 1997), but also to thelimited number of clones examined (see clone libraries’coverage values in Table 4). Apparently, both cloningstrategies were able to retrieve the most abundantsequences, but they had different degrees of success withthe less frequent sequences. The main DGGE bands
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360
356 S. Benlloch et al.
Fig. 5. Phylogenetic association for the most representative 16S rDNA archaeal fragments obtained from the PML laboratory and for thosethat were distantly related to previous sequences in the database. The dendrogram was built by the neighbour-joining method using theJukes–Cantor distance estimation from partial 16S rDNA sequences (E. coli positions 344–915). The numbers at the nodes are bootstrapconfidence values expressed as percentages of 500 bootstrap replications. Bootstrap values higher than 50% are indicated at the main nodes.

Prokaryotes in a saltern 357
coincided with the most frequently retrieved clones in thelibraries, but some groups were missed (especially thepresence of any alpha-Proteobacteria at 8% salinity). Itmust be taken into account that many of the bands in theDGGE were not excised and sequenced. These bandscould correspond to the missing sequences.
Microbial assemblages in the lower salinity ponds
Up to 11% salinity, the bacterial assemblage was remi-niscent of those found in brackish coastal lagoons andeven in coastal marine waters (Benlloch et al., 1995b;Acinas et al., 1999; Giovannoni and Rappé, 2000).Although 11% is nearly three and a half times the salinityof Mediterranean sea water, it is still not high enough toprevent the presence of microorganisms that are alsofound in marine environments. Marine bacteria have beenoften shown to be remarkably halotolerant (e.g. Forsythet al., 1971), but our approach does not give any indica-tion as to what extent such groups are active componentsof the assemblage. A significant difference when com-paring these three environments is the absence ofsequences belonging to the SAR clusters in salterns.These are very abundant in open waters and are presentin coastal lagoons as well, but they are probably not welladapted to grow in nutrient-rich environments.
Archaea seemed to be scarce at low salinity, as judgedfrom the band intensities in the DGGE gels and the needto use nested PCR in order to detect them. This is inagreement with the abundance reported in surface marinewaters (Massana et al., 1997; Karner et al., 2001). Mostsequences retrieved at low salinity showed the closestrelation to clones amplified directly by PCR from marineand/or fresh-water environments including the associatedsediments (DeLong, 1998 and references therein; Cytrynet al., 2000; Casamayor et al., 2001 and referencestherein). In this sense, Thermoplasmales-relatives MarineArchaea Group III have been detected in diverse locations(Massana et al., 2000 and references therein), includinghighly saline deep-sea brines from Kebrit Deep (Ederet al., 1999) and non-saline environments (Ferris et al.,1996; Dojka et al., 1998).
Microbial assemblages in the intermediate- and high-salinity ponds
In the 22% salinity pond, we found bacterial sequencesclosely related to previously reported marine isolates/clones representative of alpha- and gamma-Proteobac-teria, CFB and cyanobacteria. An increase in relatives of halophilic microorganisms such as Halomonas andSalinibacter (Ventosa et al., 1998; Antón et al., 2002) werefound. In a recent study (Antón et al., 2000), high numbers(14–18% of the total prokaryotic assemblage according to
FISH counts) of a new group of Bacteria were counted insalinities from 30% to 37%. This new group of Bacteriawas affiliated with the CFB phylum and has beendescribed as S. ruber (Antón et al., 2002). In our study,all DGGE bands from 32% and 37% salinity ponds cor-responded to S. ruber, and 27 out of 31 clones retrievedfrom the 32% salinity pond clustered with this extremelyhalophilic bacterium. However, this abundant bacterium isnot unique in being able to grow at extreme salinity, aswe also retrieved isolates of Pseudomonas halophila,Halomonas variabilis and H. halmophila from the 32%pond, and a relative to Pedobacter heparinus wasretrieved by cloning. Further research will yield newgroups of extremely halophilic bacteria.
In the case of Archaea, all the components belonged tothe Halobacteriaceae family. Several of these sequenceswere related to well-known members of the group suchas Halorubrum or Haloarcula. These genera have been isolated repeatedly from the crystallizers (Benllochet al., 1995a; 2001; Antón et al., 1999). The dominanthaloarchaeon, however, was the phylotype SPhT. Thisphylotype has not been obtained in pure culture, but itdominated all the previous molecular studies carried outin the crystallizers: it was the only clone retrieved inlibraries and was shown by FISH to make up a very largepercentage of the archaea. This was also reflected in ourclone libraries. Several attempts have been made to cul-tivate such square archaea but up to now they have failed(Benlloch et al., 2001).
Changes in diversity along the gradient
The increase in salinity along the gradient causes eachpond to be a more extreme environment than the previ-ous one. According to ecological theory, diversity shoulddecrease along this gradient. This decrease is very appar-ent for Eukarya (see Fig. 3 in Pedrós-Alió et al., 2000), inwhich the total number of taxons as well as all indices ofdiversity decreased very steeply from sea water to about15% salinity and more slowly thereafter. In the crystalliz-ers, Dunaliella salina was the only eukaryote found andonly at certain times. In the case of prokaryotes, however,such changes were more complex. When Shannon’sdiversity index (Magurran, 1988) was calculated for thelibraries, a decrease in the number of taxa (defined hereas all the clones with different DGGE or RFLP patterns)with increasing salinity was not apparent (Table 4). Thisindex takes into account not only taxon richness but alsoequitability. If we look at Fig. 2, however, it is apparentthat very different bacterial groups were found in the lowersalinity ponds, whereas those from the crystallizersbelonged essentially to one single group: the S. rubercluster. Archaea behaved quite similarly and, at thehighest salinity, most OTUs belonged to the SPhT cluster.
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360

Therefore, the diversity in the crystallizers can be con-sidered to be ‘microdiversity’, the co-existence of severalclosely related clones of microorganisms (Fuhrman andDavis, 1997; Moore et al., 1998; Casamayor et al.,2002b). In summary, and regardless of their changes inabundance, both Bacteria and Archaea showed the samepattern: as salinity increased, the number of different clus-ters decreased, and only one cluster become dominant.However, this cluster showed a considerable degree ofmicrodiversity. The physiological and ecological signifi-cance of this microdiversity remains to be studied.
Experimental procedures
Site description, sampling and DNA extraction fromenvironmental samples
During the MIDAS workshop in May 1999, samples were col-lected from eight shallow ponds with different salinities (4%,5.4%, 8%, 11%, 15%, 22.4%, 31.6% and 37%) from the mul-tipond solar saltern ‘Bras del Port’ located in Santa Pola (Ali-cante, Spain, 38∞12¢ N, 0∞36¢ W). Temperature ranged from23∞C in the 4% pond to 28∞C in the 37% pond. All sampleswere taken a few centimetres below the water surface, avoid-ing the corners of the ponds where wind accumulated organicmatter as scum and debris. Total salt concentrations weredetermined by a hand refractometer and by ash content of10 ml water samples dried at 110∞C for 24 h and then at400∞C for 4 h. Total DNA was extracted by different protocolsas described elsewhere (Casamayor et al., 2002a). Aliquotsof the DNA extracts from each pond were shared amonggroups, and DGGE was performed with all samples. TheDGGE gel consistently showed the same pattern independ-ent of the method of extraction used (Casamayor et al.,2002a).
Isolation of cultures and DNA extraction from cultures
Water (100 µl) from the 8%, 22% and 32% ponds was usedto inoculate plates containing 0.1% yeast extract (Difco) and
0.1% cocktail FRV [1:1:1 Artemia salina, Spirulina (Sigma)and fishmeal] in autoclaved water from the same ponds.Several dilutions (10-1, 10-2, 10-3 and 10-4) were inoculated,and the cultures were grown at two different incubation temperatures (25∞C and 45∞C). Half the cultures tested wereincubated under microaerophilic conditions. DNA wasextracted as described previously (Benlloch et al., 2001).
PCR amplification and DGGE analysis of PCR products
Two groups used DGGE and sequencing along the salinitygradient: the University of Bergen group (UiB) and the Insti-tut de Ciències del Mar of Barcelona (ICM) group. Protocolswere as described in Experimental procedures of the relatedpaper by Casamayor et al. (2002a). Primers used are shownin Table 5. DGGE bands to be sequenced were punched fromthe gel and processed as described previously (Øvreås et al.,1997; Casamayor et al., 2000b). PCR amplifications carriedout by UiB were performed with the 8f(GC)-518r primers forBacteria, whereas for Archaea, the primers 21f and 958r wereused first to produce a 955 bp fragment, which was then usedas a template for the nested PCR in combination with theprimers PARCH340f(GC) and PARCH519r (Øvreås et al.,1997). PCR amplifications carried out by ICM were per-formed with the 344f(GC)-915r primers for Archaea.
Analysis of clone libraries
Two laboratories made clone libraries: the Plymouth MarineLaboratory (PML) and the Universidad Miguel Hernández(UMH). Each laboratory used one combination of primers.UMH used 8f-Sr for Bacteria and 21f-Sr for Archaea (Table 5).PCR amplification was carried out as described previously(Benlloch et al., 2001). PCR products were cloned with the original TOPO TA cloning kit (Invitrogen) according to the manufacturer’s recommendations. The clones with thecorrect insert were reamplified, purified with the QIA-quickPCR purification kit (Qiagen) and digested with HinfI (Gibco).The generated fragments were run on a 2% agarose gel, andclones with different RFLP band patterns were chosen forpartial sequencing (E. coli positions 150–600 for bacterial
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360
358 S. Benlloch et al.
Table 5. Primers used in the present study.
Primera Sequences (5¢ to 3¢) Target siteb Domain Reference
344f ACGGGGYGCAGCAGGCGCGA 344–363 Archaea Raskin et al. (1994)915r GTGCTCCCCCGCCAATTCCT 915–934 Archaea Stahl and Amann (1991)21f TTCCGGTTGATCCTGCCGGA 7–26 Archaea DeLong (1992)Sr GGTTACCTTGTTACGACTT 1509–1491 Universal Lane (1991)958r (C/T)CCGGCGTTGA(A/C)TCCAATT 958–976 Archaea DeLong (1992)PARCH519r TTACCGCGGC(G/T)GCTG 519–533 Archaea Øvreås et al. (1997)PARCH340f CCCTACGGGG(C/T)GCA(G/C)CAG 340–357 Archaea Øvreås et al. (1997)8f AGAGTTTGATCCTGGCTCAG 8–27 Bacteria Lane (1991)518r ATTACCGCGGCTGCTGG 518–534 Bacteria Muyzer et al. (1993)1055f AATGGCTGTCGTCAGCTCGT 1055–1074 Bacteria Amann et al. (1995)1392r GYACACACCTCCCGT 1406–1390 Universal Amann et al. (1995)GC clampc CGCCCGCCGCGCCCCGCGCCCGTCCCGCCGCCCCCGCCCG Muyzer et al. (1993)
a. f, forward primer; r, reverse primer.b. The numbering of positions is based on E. coli 16S rDNA.c. The GC clamp was attached to the 5¢ end of the primers 344f, 1392r, 915r, 8f and 340f.

Prokaryotes in a saltern 359
clones and positions 85–555 for archaeal clones). PML used1055f-1392r for Bacteria and 344f-915r for Archaea (Table 5).PCR amplification was carried out at 52∞C annealing tem-perature. The PCR product was run in agarose gel, cut outand purified with a QIAEX gel extraction kit (Qiagen), andligated into the plasmid pBluescript SK-. Clones contain-ing the correct insert were reamplified and examined on a DGGE gel.
Sequence analysis and accession numbers
16S rDNA fragments were sequenced with the ABI Prism Ter-minator cycle sequencing ready reaction kit (Perkin-Elmer)and an ABI Prism 377 automated sequencer (Perkin-Elmer)according to the manufacturer’s instructions. The sameprimers used for PCR amplification were used for sequenc-ing except that the GC clamp was omitted. Sequences werecompared with 16S rRNA sequences available in both theRibosomal Database Project (Maidak et al., 2001) and theGenBank and EMBL databases obtained from the NationalCenter for Biotechnology Information (NCBI) Database by the advanced BLAST search (Altschul et al., 1997). Thesequences were aligned with CLUSTALW program (GCGpackage), and their similarity matrix was calculated by theJukes–Cantor method using the program MEGA (molecularevolutionary genetics analysis). We have deposited 125nucleotide sequences in GenBank under the accessionnumbers AF477831 to AF477955.
Acknowledgements
This work was financed by the European Union through con-tract MAS3-CT97-0154 ‘MIDAS’. We thank Miguel Cuervo-Arango, owner of the Bras-del-Port salterns, for his kind helpand permission to sample the hypersaline ponds.
References
Acinas, S.G., Antón, J., and Rodríguez-Valera, F. (1999)Diversity of free-living and attached bacteria in offshorewestern Mediterranean waters as depicted by analysis ofgenes encoding 16S rRNA. Appl Environ Microbiol 65:514–522.
Altschul, S.F., Madden, T.L., Schäffer, A.A., Zhang, J., Zhang,Z., Miller, W., and Lipman, D.J. (1997) Gapped BLAST andPSI-BLAST: a new protein of protein database searchprogram. Nucleic Acids Res 25: 3389–3402.
Amann, R.I., Ludwig, W., and Schleifer, K.H. (1995) Phylo-genetic identification and in situ detection of individualmicrobial cells without cultivation. Microbiol Rev 59: 143–169.
Antón, J., Llobet-Brossa, E., Rodríguez-Valera, F., andAmann, R.I. (1999) Fluorescence in situ hybridizationanalysis of the prokaryotic community inhabiting crystal-lizer ponds. Environ Microbiol 1: 517–525.
Antón, J., Rosselló-Mora, R., Rodríguez-Valera, F., andAmann, R.I. (2000) Extremely halophilic Bacteria in crys-tallizer ponds from solar salterns. Appl Environ Microbiol66: 3052–3057.
Antón, J., Oren, A., Benlloch, S., Rodríguez-Valera, F.,
Amann, R., and Rosselló-Mora, R. (2002) Salinibacterruber gen. nov., sp. nov., a new species of extremelyhalophilic Bacteria from saltern crystallizer ponds. Int JSyst Evol Microbiol 52: 485–491.
Benlloch, S., Martínez-Murcia, A.J., and Rodríguez-Valera, F.(1995a) Sequencing of bacterial and archaeal 16S rRNAgenes directly amplified from a hypersaline environment.Syst Appl Microbiol 18: 574–581.
Benlloch, S., Rodríguez-Valera, F., Acinas, S.G., andMartínez-Murcia, A.J. (1995b) Bacterial diversity in twocoastal lagoons deduced from 16S rDNA PCR amplifica-tion and partial sequencing. FEMS Microbiol Ecol 18:267–280.
Benlloch, S., Acinas, S.G., Antón, J., Lopez-Lopez, A., Luz,S.P., and Rodríguez-Valera, F. (2001) Archaeal Biodiver-sity in crystallizer ponds from a solar saltern: culture versusPCR. Microb Ecol 41: 12–19.
Brinkhoff, T., and Muyzer, G. (1997) Increased species diver-sity and extended habitat range of sulfur-oxidizing Thiomi-crospira spp. Appl Environ Microbiol 63: 3789–3796.
Casamayor, E.O., Calderón-Paz, J.I., and Pedrós-Alió, C.(2000a) 5S rRNA fingerprints of marine bacteria, halophilicarchaea and natural prokaryotic assemblages along asalinity gradient. FEMS Microbiol Ecol 34: 113–119.
Casamayor, E.O., Schäfer, H., Bañeras, L., Pedrós-Alió, C.,and Muyzer, G. (2000b) Identification of and spatio-temporal differences between microbial assemblages fromtwo neighboring sulfurous lakes: comparison by micro-scopy and denaturing gradient gel electrophoresis. ApplEnviron Microbiol 66: 499–508.
Casamayor, E.O., Muyzer, G., and Pedrós-Alió, C. (2001)Composition and temporal dynamics of planktonic archaealassemblages from anaerobic sulfurous environmentsstudied by 16S rDNA denaturing gradient gel electro-phoresis and sequencing. Aquat Microb Ecol 25: 237–246.
Casamayor, E.O., Massana, R., Benlloch, S., Øvreås, L.,Díez, B., Goddard, V., et al. (2002a) Changes in archaeal,bacterial and eukaryal assemblages along a salinity gra-dient by comparison of genetic fingerprinting methods in amultipond solar saltern. Environ Microbiol 4: 338–348.
Casamayor, E.O., Pedrós-Alió, C., Muyzer, G., and Amann,R. (2002b) Microheterogeneity in 16S rDNA-defined bac-terial populations from a stratified planktonic environmentis related to temporal changes and to ecological adapta-tions. Appl Environ Microbiol 68: 1706–1714.
Cytryn, E., Dror, M., Oremland, R.S., and Cohen, Y. (2000)Distribution and diversity of Archaea corresponding to thelimnological cycle of a hypersaline stratified lake (SolarLake, Sinai, Egypt). Appl Environ Microbiol 66: 3269–3276.
DeLong, E.F. (1992) Archaea in coastal marine environ-ments. Proc Natl Acad Sci USA 89: 5685–5689.
DeLong, E.F. (1998) Everything in moderation: Archaea as‘non-extremophiles’. Curr Opin Genet Dev 8: 649–654.
Dojka, M.A., Jr, Hugenholz, P., Haack, S., and Pace, N.R.(1998) Microbial diversity in a hydrocarbon and chlori-nated-solvent-contaminated aquifer undergoing intrinsicbioremediation. Appl Environ Microbiol 172: 213–218.
Eder, W., Ludwig, W., and Huber, R. (1999) Novel 16S rRNAgene sequences retrieved from highly saline brine sediments of Kebrit Deep, Red Sea. Arch Microbiol 172:213–218.
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360

Ferris, M.J., Muyzer, G., and Ward, D.M. (1996) Denaturinggradient gel electrophoresis profile of 16S rRNA-definedpopulations inhabiting a hot spring microbial mat commu-nity. Appl Environ Microbiol 62: 340–346.
Forsyth, M.P., Shindler, D.B., Gouchnauer, M.B., andKushner, D.J. (1971) Salt tolerance of intertidal marinebacteria. Can J Microbiol 17: 825–828.
Fuhrman, J.A., and Davis, A.A. (1997) Widespread Archaeaand novel Bacteria from the deep sea as shown by 16SrRNA gene sequences. Mar Ecol Prog Series 150: 275–285.
Giovannoni, S.J., and Rappé, M.S. (2000) Evolution, diversity and molecular ecology of marine prokaryotes. InMicrobial Ecology of the Ocean. Kirchman, D.L. (eds). New York: Wiley Interscience, pp. 47–84.
Javor, B. (1989) Solar salterns. In Hypersaline Environments:Microbiology and Biochemistry. Javor, B. (eds). Brock/Springer Series in Contemporary Bioscience. Berlin:Springer-Verlag, pp. 189–204.
Joint, I., Henriksen, P., Garde, K., and Riemann, B. (2002)Primary production, nutrient assimilation and microzoo-plankton grazing along a hypersaline gradient. FEMSMicrobiol Ecol 39: 245–257.
Karner, M.B., DeLong, E.F., and Karl, D.M. (2001) Archaealdominance in the mesopelagic zone of the Pacific Ocean.Nature 409: 507–510.
Lane, D.J. (1991) 16S/23S rRNA sequencing. In Nucleic AcidTechniques in Bacterial Systematics. Stackebrandt, E., andGoodfellow, M. (eds). New York: John Wiley and Sons, ,pp. 115–175.
Magurran, A.E. (1988) Ecological Diversity and its Measure-ment. Princeton, NJ: Princeton University Press.
Maidak, B.L., Cole, J.R., Lilburn, T.G., Parker, C.T., Jr,Saxman, P.R., Farris, R.J., et al. (2001) The RDP-II (Ribosomal Database Project). Nucleic Acids Res 29:173–174.
Martínez-Murcia, A.J., Acinas, S.G., and Rodríguez-Valera,F. (1995) Evaluation of prokaryotic diversity by restrictasedigestion of 16S rDNA directly amplified from hypersalineenvironments. FEMS Microbiol Ecol 17: 247–256.
Massana, R., Murray, A.E., Preston, C.M., and DeLong, E.F.(1997) Vertical distribution and phylogenetic characteriza-tion of marine planktonic archaea in the Santa Barbarachannel. Appl Environ Microbiol 63: 50–56.
Massana, R., DeLong, E.F., and Pedrós-Alió, C. (2000) A fewcosmopolitan phylotypes dominate planktonic archaealassemblages in widely different oceanic provinces. ApplEnviron Microbiol 66: 1777–1787.
Moore, L.R., Rocap, G., and Chisholm, S.W. (1998) Physiol-ogy and molecular phylogeny of coexisting Prochlorococ-cus ecotypes. Nature 393: 464–467.
Mullins, T.D., Britschgi, T.B., Krest, R.L., and Giovannoni, S.J.(1995) Genetic comparisons reveal the same unknown
bacterial lineages in Atlantic and Pacific bacterioplanktoncommunities. Limnol Oceanol 40: 148–158.
Muyzer, G., de-Waal, E.C., and Uitterlinden, A.G. (1993) Profiling of complex microbial populations by denaturinggradient gel electrophoresis analysis of polymerase chain reaction amplified genes coding for 16S rRNA. ApplEnviron Microbiol 59: 695–700.
Muyzer, G., Brinkhoff, T., Nübel, U., Santagoeds, C., Schäfer,H., and Wawer, C. (1997) Denaturing gradient gel elec-trophoresis (DGGE) in microbial ecology. In MolecularMicrobial Ecology Manual. Vol. 3.4.4. van Elsas, J.D., andde Bruijn, F.J. (eds). Dordrecht: Kluwer Academic Publish-ers, pp. 1–27.
Øvreås, L., Forney, L., Daae, F.L., and Torsvik, V. (1997) Distribution of bacterioplankton in meromictic lake Sælenvannet, as determined by denaturing gradient gelelectrophoresis of PCR amplified gene fragments codingfor 16S rRNA. Appl Environ Microbiol 63: 3367–3373.
Pedrós-Alió, C., Calderón-Paz, J.I., and Gasol, J.M. (2000)Comparative analysis shows that bacterivory, and not virallysis, controls the abundance of heterotrophic prokaryoticplankton. FEMS Microbiol Ecol 32: 157–165.
Raskin, L., Stromley, J.M., Rittmann, B.E., and Stahl, D.A.(1994) Group specific 16S rRNA hybridization probes todescribe natural communities of methanogens. ApplEnviron Microbiol 60: 1232–1240.
Rodríguez-Valera, F. (1988) Characteristics and microbialecology of hypersaline environments. In Halophilic Bacte-ria. Rodríguez-Valera, F. (eds). Boca Raton, FL: CRCPress.
Rodríguez-Valera, F. (1993) Introduction to saline environ-ments. In The Biology of Halophilic Bacteria. Vreeland, R.,and Hochstein, L.I. (eds). Boca Raton, FL: CRC Press, pp. 1–23.
Rodríguez-Valera, F., Ruíz-Berraquero, F., and Ramos-Cormenzana, A. (1981) Characteristics of the heterotrophicbacterial populations in hypersaline environments of differ-ent salt concentrations. Microb Ecol 7: 235–243.
Rodríguez-Valera, F., Ventosa, A., Juez, G., and Imhoff, J.F.(1985) Variation of environmental features and microbialpopulations with salt concentrations in a multi-pond saltern.Microb Ecol 11: 107–115.
Stahl, D.A., and Amann, R.I. (1991) Development and ampli-fication of nucleic acid probes. In Nucleic Acid Techniquesin Bacterial Systematics. Stackebrandt, E., and Goodfel-low, M. (eds). New York: Wiley, pp. 205–248.
Ventosa, A., Nieto, J.J., and Oren, A. (1998) Biology of mod-erately halophilic aerobic bacteria. Microbiol Mol Biol Rev62: 504–544.
Wintzingerode, F.V., Goebel, U.B., and Stackebrandt, E.(1997) Determination of microbial diversity in environmen-tal samples: pitfalls of PCR-based rRNA analysis. FEMSMicrobiol Rev 21: 213–229.
© 2002 Blackwell Science Ltd, Environmental Microbiology, 4, 349–360
360 S. Benlloch et al.
Copyright © 2022 FDOKUMEN




















