
Early Triassic Archosaur-Dominated Footprint Assemblage from the Argana Basin (Western High Atlas,...
14
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [Bergakadmei] On: 15 September 2010 Access details: Access Details: [subscription number 917157432] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Ichnos Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713643181 Early Triassic Archosaur-Dominated Footprint Assemblage from the Argana Basin (Western High Atlas, Morocco) Hendrik Klein a ; Sebastian Voigt b ; Abdelkbir Hminna c ; Hafid Saber c ; Jörg Schneider b ; Driss Hmich c a Saurierwelt Museum, Neumarkt, Germany b Geological Institute, TU Bergakademie Freiberg, Freiberg, Germany c Department of Geology, Chouaib Doukkali University, El Jadida, Morocco Online publication date: 15 September 2010 To cite this Article Klein, Hendrik , Voigt, Sebastian , Hminna, Abdelkbir , Saber, Hafid , Schneider, Jörg and Hmich, Driss(2010) 'Early Triassic Archosaur-Dominated Footprint Assemblage from the Argana Basin (Western High Atlas, Morocco)', Ichnos, 17: 3, 215 — 227 To link to this Article: DOI: 10.1080/10420940.2010.510030 URL: http://dx.doi.org/10.1080/10420940.2010.510030 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
chouaibdokkali -
Category
Documents
-
view
0 -
download
0
Transcript of Early Triassic Archosaur-Dominated Footprint Assemblage from the Argana Basin (Western High Atlas,...

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Bergakadmei]On: 15 September 2010Access details: Access Details: [subscription number 917157432]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
IchnosPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713643181
Early Triassic Archosaur-Dominated Footprint Assemblage from theArgana Basin (Western High Atlas, Morocco)Hendrik Kleina; Sebastian Voigtb; Abdelkbir Hminnac; Hafid Saberc; Jörg Schneiderb; Driss Hmichc
a Saurierwelt Museum, Neumarkt, Germany b Geological Institute, TU Bergakademie Freiberg,Freiberg, Germany c Department of Geology, Chouaib Doukkali University, El Jadida, Morocco
Online publication date: 15 September 2010
To cite this Article Klein, Hendrik , Voigt, Sebastian , Hminna, Abdelkbir , Saber, Hafid , Schneider, Jörg and Hmich,Driss(2010) 'Early Triassic Archosaur-Dominated Footprint Assemblage from the Argana Basin (Western High Atlas,Morocco)', Ichnos, 17: 3, 215 — 227To link to this Article: DOI: 10.1080/10420940.2010.510030URL: http://dx.doi.org/10.1080/10420940.2010.510030
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Ichnos, 17:215–227, 2010Copyright c© Taylor & Francis Group, LLCISSN: 1042-0940 print / 1563-5236 onlineDOI: 10.1080/10420940.2010.510030
Early Triassic Archosaur-Dominated Footprint Assemblagefrom the Argana Basin (Western High Atlas, Morocco)
Hendrik Klein,1 Sebastian Voigt,2 Abdelkbir Hminna,3 Hafid Saber,3 JorgSchneider,2 and Driss Hmich3
1Saurierwelt Museum, Neumarkt, Germany2Geological Institute, TU Bergakademie Freiberg, Freiberg, Germany3Department of Geology, Chouaib Doukkali University, El Jadida, Morocco
An assemblage of abundant and well-preserved tetrapodfootprints has been discovered in the Tanamert Member (T3)of the Triassic Timezgadiouine Formation (Argana basin, west-ern High Atlas, Morocco). It is the first fossil record fromT3. Surfaces from different localities show a uniform tetrapodichnofauna that consists of chirotherian and small lacertoidforms. The chirotherians are assigned to the plexus Protochi-rotherium—Synaptichnium, their trackmakers interpreted as basalarchosaurs. The lacertoid imprints show close affinities withRhynchosauroides and may reflect archosauromorphs or lepi-dosauromorphs. Protochirotherium—Synaptichnium assemblagesare characteristic of the Early Triassic and were known previouslyonly from units of this age in central Europe. Biostratigraphically,the European record implies a wide-spread pre-Anisian Protochi-rotherium—Synaptichnium dominated assemblage preceding thefirst appearance of Chirotherium barthii near the Olenekian-Anisian boundary. The stratigraphic position of T3 betweenLate Permian (uppermost T2) and Middle Triassic (T4) andthe European correlatives suggest an Early Triassic age of thisunit. It is the first record of Early Triassic continental depositsin Morocco. The surfaces from T3 open up perspectives forfurther contributions to ecology, biogeography and locomotion ofearly archosaurs. Furthermore, excellent outcrops and quality offootprint preservation in the Argana basin offer a potential forclarification of ichnotaxonomic and biostratigraphic issues.
Keywords Protochirotherium, Synaptichnium, tetrapod footprints,red-beds, Northern Africa
INTRODUCTIONThe Late Permian to Early Triassic is an important period
in the evolution of terrestrial tetrapods because it covers
Address correspondence to Hendrik Klein, Rubezahlstr. Sauri-erwelt Museum, 1 D-92318 Neumarkt, Germany. E-mail: [email protected]
a remarkable faunal change from Late Paleozoic synapsid-dominated communities to the predominance of eureptilesexpressed by the rise and early radiation of archosaurs (Bentonet al., 2004; Botha and Smith, 2006; Sahney and Benton, 2008).The term archosaurs is used here for a clade that includesthe crown-group Archosauria as well as some closely relatedoutgroups as the proterosuchids, erythrosuchids, euparkeri-ids, and proterochampsids (Archosauriformes; Gauthier, 1986;Sereno, 1991; Benton, 1999; Brusette et al., 2008; Dilkes andSues, 2009). Several large plant-eating tetrapods such as thepareiasaurs and many dicynodont synapsids disappeared as didcarnivorous forms such as the gorgonopsids. A few synapsids,procolophonids, and eureptiles such as the prolacertiformsand proterosuchid archosaurs crossed the Permian/Triassicboundary (Benton, 2008; Benton et al., 2004).
During the Early Triassic, archosaurs diversified into ad-ditional groups such as erythrosuchids, euparkeriids, prote-rochampsids, and rauisuchians. This first radiation was thebeginning of archosaur dominance continuing for the next180 million years. Cursorial archosaurs were among the mostsuccessful land-dwelling vertebrates during the Mesozoic era.Essential evolutionary developments concern the acquisitionof an upright limb posture and a parasagittal gait (Haubold,1967, 1984; Kubo and Benton, 2007; Padian et al., 2010).The position of the legs under the body enabled fast runningwith minor loss of energy compared with the gait of typicalsprawlers. This is reflected in narrow trackways of chirotherianswith occasional large strides. So far, little is known aboutlocomotory developments and foot morphology of some basalarchosaur groups as well as the earliest ancestry of dinosaursand crocodiles from the bone record (Gower, 1996; Gowerand Sennikov, 2000; Sereno, 1991). Fossil footprints couldbe an important source of data in this respect. Footprintsare widespread stratigraphically and geographically and oftenoccur in large numbers. In some sequences they are the onlydocuments of the presence of distinct osteological groups.
215
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010

216 H. KLEIN ET AL.
Triassic footprints have been described mainly from the middleand late part of the period (Peabody, 1948; Demathieu, 1970;Haubold, 1971a, b), whereas pre-Anisian ichnofaunas are lessknown. Representative surfaces with Early Triassic archosaur-dominated assemblages are documented only from a fewlocalities in Central Europe (Demathieu and Haubold, 1982;Fuglewicz et al., 1990; Ptaszynski, 2000; Fichter and Kunz,2004). Small therapsid footprints from the Early Triassic ofAntarctica and Australia (MacDonald et al., 1991; Retallack,1996) are poorly preserved.
Triassic footprints were described from three horizons inMorocco: (1) the base of the Irohalene Member (T5) of theCarnian Timezgadiouine Formation in the Argana Basin; (2)from Middle—Late Triassic strata of the Ourika basin (Bironand Dutuit, 1981); and (3) from the Late Triassic Oued Oum ErRbiaa Formation of the Sidi Saıd M’aachou Basin (Hminna etal., 2009a). Other adjoining occurrences in North Africa are theMiddle Triassic of Algeria (Kotanski et al., 2004) and possibleEarly Triassic deposits of Niger (Ginsburg et al., 1968; Taquet,1976).
Here we describe the discovery of tetrapod footprints inthe thus far unfossiliferous Tanamert Member (T3) of theTimezgadiouine Formation (Argana Basin, Morocco). These aremainly in situ-surfaces that display abundant and well preservedtrackways and footprints of archosaurs (chirotherians) as well asof small lacertoid forms (Klein et al., 2009). The assemblage isa rare document of early archosaur-dominated communities andmay contribute to our knowledge of evolutionary developmentsin the locomotor apparatus of these tetrapods. Of greatimportance are biostratigraphic implications that necessitate are-evaluation of the age of T3.
MATERIAL AND METHODSAll footprints are natural casts of fine to coarse clastics
on lower bed-surfaces (convex hyporeliefs), exposed in erodedrecent stream-beds. The material comes from 4 localities in T3(Figs. 1C-D; Appendix 1): CDUE localities 10 and 13 on thewestern bank of the Oued Issene river, south of Irerhi, east ofTikida and CDUE localities 11 and 12 east of Aguenza. CDUElocalities 10, 13 (levels 1–4) and 11 provided in-situ surfaces.They were discovered by some of the authors (SV, AH) duringprospection in 2008 and 2009. CDUE locality 12 demarcates anisolated large sandstone block in a seasonally dry stream bed. Itwas detected by F. Korner, Freiberg University, during fieldwork(with authors HS, JWS) in 2002. In a first description, the blockwas attributed to T2 (Hmich et al., 2006, fig. 12F, 13A). In themeanwhile, the source rock could be assigned to T3 strata bysedimentological and ichnological evidence.
The best preserved footprints were numbered, all otherswere recorded by outline drawings and photographs only. Thenumbered material consists of 2 trackways, 1 partial trackway,and several isolated footprints—19 imprints in total—along withseveral indistinct trackways of small forms. Five slabs with
original material were collected. Moulds in latex and orthopedicfoam were taken from the best preserved imprints, 14 in total.All collected specimens and molds are housed in the Departmentof Geology of the Chouaib Doukkali University El Jadida,Morocco. Mapping and drawing of outlines was carried outon transparency film with 50 cm and 20 cm square grids. Photoswere made in the field under natural light conditions. Outlinedrawings were worked out and digitalized with a vector-baseddrawing software. Measurements correspond to the commonstandard methods proposed by Haubold (1971b) and Leonardi(1987).
Institutional abbreviations. CDUE: Chouaib DoukkaliUniversity El Jadida, Morocco. “C” added to specimen num-bers of this collection indicates collected specimen (originalmaterial), “M” means mold (artificial cast), added numbers 1-nrefer to successive sets in a trackway.
Ichnological abbreviations. pl, length of pes; pw, widthof pes; pl/pw, length to width ratio of pes; ml, length ofmanus; mw, width of manus; ml/mw, length to width ratio ofmanus; SL, length of stride; TW, trackway width; PA, paceangulation.
GEOLOGIC SETTINGThe Argana Basin is located in the Western High Atlas
of Central Morocco, NE of Agadir. The study area with thefootprint localities lies close to the village of Irerhi at about halfthe distance between Argana and Timezgadiouine (Fig. 1A–C).Permian and Triassic strata crop out extensively in a successionof typical continental red-beds. The sediment accumulation ofthe Argana red-beds took place at the eastern margin of anintracontinental rift-basin of the North American and Africanplates (Olsen, 1997; Hofmann et al., 2000; Tourani et al., 2000).A large number of such basins are located in Morocco and ineastern North America. They demarcate the breakup of Pangeabetween NW Gondwana and Laurasia. Palaeogeographically,the northwestern part of Africa is adjacent to eastern NorthAmerica in Triassic time. The Fundy basin of Nova Scotiaand the Argana basin of Morocco were paralleling the suturethat marks the rifting and early opening of the Central Atlantic(Olsen et al., 2000; Olsen and Et Touhami, 2008). In NorthAmerica, deposits of the Newark Supergroup (?Middle/UpperTriassic—Lower Jurassic) represent the filling between NovaScotia in the north and North Carolina in the south. Depositiontook place basically under fluvial and lacustrine conditions(Olsen et al., 2000; Schlische, 2003). In Morocco, the sed-imentary input is essentially controlled by alternating andinterfingering alluvial fan, braided to meandering river andplaya environments. Successions with conglomerates, sand-,silt-, and mudstones in channel and overbank settings reflectrepeating cycles of high- and low-energy regime deposition.They were obviously related to both long-term climatic changesand seasonal fluctuation of precipitation under semi-arid toarid conditions (Hofmann et al., 2000; Tourani et al., 2000).
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010
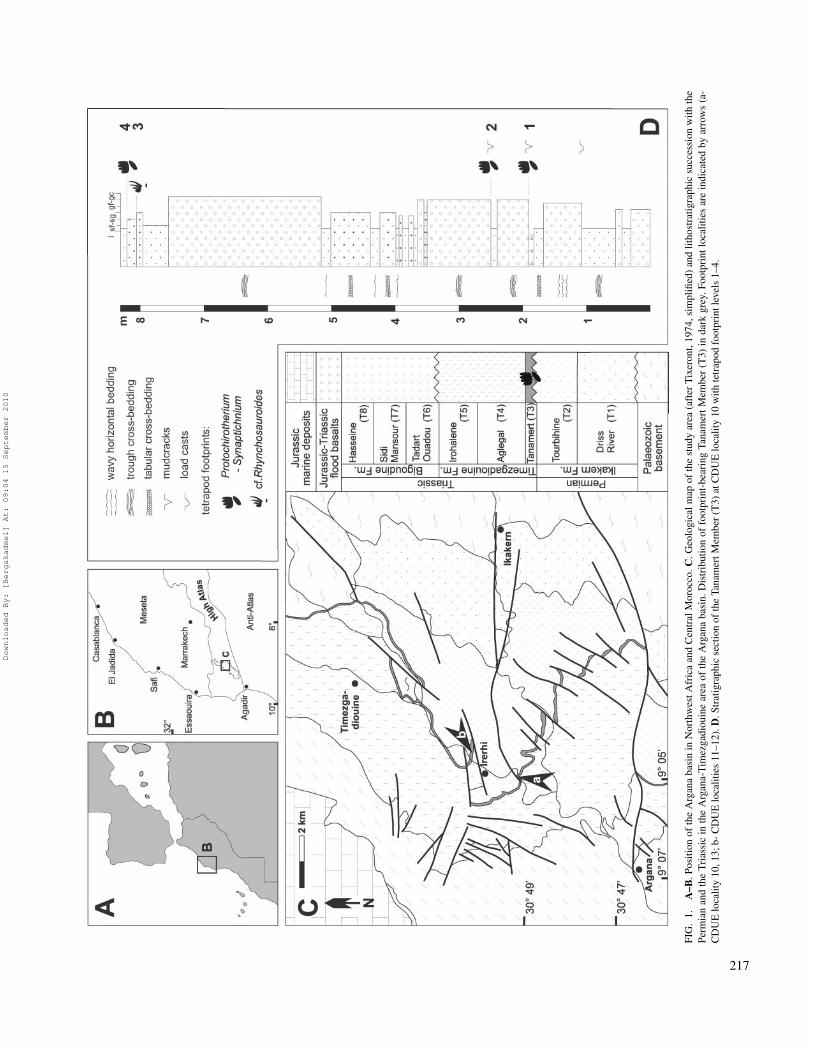
FIG
.1.
A–B
.Pos
ition
ofth
eA
rgan
aba
sin
inN
orth
wes
tAfr
ica
and
Cen
tral
Mor
occo
.C.G
eolo
gica
lmap
ofth
est
udy
area
(aft
erT
ixer
ont,
1974
,sim
plifi
ed)
and
litho
stra
tigra
phic
succ
essi
onw
ithth
ePe
rmia
nan
dth
eT
rias
sic
inth
eA
rgan
a-T
imez
gadi
ouin
ear
eaof
the
Arg
ana
basi
n.D
istr
ibut
ion
offo
otpr
int-
bear
ing
Tana
mer
tM
embe
r(T
3)in
dark
grey
.Foo
tpri
ntlo
calit
ies
are
indi
cate
dby
arro
ws
(a-
CD
UE
loca
lity
10,1
3;b-
CD
UE
loca
litie
s11
–12)
.D.S
trat
igra
phic
sect
ion
ofth
eTa
nam
ertM
embe
r(T
3)at
CD
UE
loca
lity
10w
ithte
trap
odfo
otpr
intl
evel
s1–
4.
217
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010

218 H. KLEIN ET AL.
After Tourani et al. (2000), the sedimentary environment ofthe Tanamert Member (T3) is interpreted as that of braidedrivers.
The total thickness of the Permian and Triassic red-bedshas been differently evaluated between 2,500 and 5,000 m(Tixeront, 1973; Jones, 1975; Brown, 1980; Olsen et al., 2000;Hofmann et al., 2000; Tourani et al., 2000). These predomi-nantly siliciclastic deposits unconformably rest on Palaeozoicbasement rocks and are themselves overlain by tholeiitic basaltsof the Central Atlantic Magmatic Province (Argana basalt;see Ait Chayeb et al., 1998; Olsen et al., 2000; Olsen andEt-Touhami, 2008). The sequence was subdivided by Tixeront(1973) into eight lithostratigraphic units: T1–T8. These wereformally attributed by Brown (1980) as members to the Ikakern(T1–T2: 900–1800m), Timezgadiouine (T3–T5: 1000–2000m),and Bigoudine formations (T6–T8: 300–1500m; Fig. 1C). Thebiostratigraphic record indicates a Permian age for T1 andT2 (Driss and Tourbihine Member of the Ikakern Fm.) and aTriassic age for T4–T8 (Aglegal, Irohalene, Tadart Ouadou,Sidi Mansour, and Hasseine Member of the Timezgadiouineand Bigoudine formations; see below). Based on palynologicaldata, the boundary with the Jurassic was formerly positioned afew meters below the Argana basalt within a succession of greyand black shales (Olsen et al, 2000, 2002 Et-Touhami et al.,2008). However, recent interdisciplinary studies by Deenan et al.(2010) comprising data from astrochronology, paleomagnetism,basalt geochemistry, and geobiology suggest that the Triassic-Jurassic boundary is above the basalt. Thus far, the TanamertMember (T3) had provided no age indicative fossils, but itwas linked to the Triassic Timezgadiouine Formation based onthe conformable contact with the overlying T4 and the angularunconformity separating it from underlying Permian T2. It wassupposed to be of Middle or Late Triassic age (Hofmann et al.,2000; Tourani et al., 2000). Estimated thickness of T3 is up to10 m.
The footprints described herein come from several horizonsnear to the base of T3. At CDUE locality 10, four differenttrackway levels (1–4) occur in a succession of fine to coarseclastics (Fig. 1D). Level 1 and 2 characterize the boundarybetween siltstones or fine-grained sandstones overlain bymedium- to coarse-grained trough cross-bedded conglomerates.Footprint surfaces were mudcracked prior to embedding.
The sedimentation pattern is typical for fluvial sequenceswith preserved footprint surfaces: (1) high energy flow withtransport and deposition of coarse-grained sand and gravel,in particular in channel bodies; (2) decreasing flow energyand deposition of fine-grained sand and silt; (3) drop ofwater level and drying out of small pool-shaped water bod-ies, preservation of footprints, and formation of mudcracks;(4) moderate flooding and covering of surfaces with thinlayer of fine-grained sand conserving footprints; (5) highenergy flow and cutting by coarse-grained gravel as in(1).
SYSTEMATIC PALAEONTOLOGYPlexus Protochirotherium—Synaptichnium (Fig. 2–Fig. 3,
5G–L; Appendix 1–2)Term introduced here for chirotherians with plantigrade to
semiplantigrade pes and manus imprints with relatively longdigits IV and V. Digit IV in the pes longer than III to slightlyshorter. Imprints with wide variation between broad and slendershape (see discussion below).
Ichnogenus Protochirotherium Fichter and Kunz, 2004Type ichnospecies: Protochirotherium wolfhagense Fichter
and Kunz, 2004 (monotypic).Diagnosis (after Fichter and Kunz, 2004): Pes imprints
pentadactyl, plantigrade to semiplantigrade, average length 109mm, average width 81 mm. Digit proportions V > III > IV >
II > I. Digit angle I-IV 28◦, digit angle I-V 67◦ on average. Aver-age cross-axis angle 76◦. Manus imprints: Like the pes imprintspentadactyl and plantigrade to semiplantigrade, average length:59 mm, average width 62 mm. Digit proportions V > III >
IV = II > I. Digit angle I-IV 46◦, digit angle I-V 73◦ on average.Ichnogenus Synaptichnium Nopcsa, 1923Type ichnospecies: Synaptichnium pseudosuchoides Nopcsa,
1923.Diagnosis (after Haubold, 1971b): Lacertoid chirotherians
of relative small size, average pes length up to 100 mm; Pedaldigits I-IV increasing in length, digit V positioned more laterallythan proximally behind anterior digit group. Palmar : plantarsurface ratio = 1:1,8 to 3,7; 1:2,5 on average. No distinct narrowgait, pace angulation rarely exceeding 160◦, 140◦ on average.
Material referred to Protochirotherium—SynaptichniumCDUE 60, trackway consisting of 4 successive pes-manus
sets; molds taken from 3 steps, CDUE 60–1M, 60–2M, 60–4M;CDUE 61, trackway, with 3 successive pes-manus sets, CDUE61–1C, CDUE 61–1M; several isolated imprints CDUE 62M-65M; all from same surface (level 2) at CDUE locality 10 (Fig.2A–F). CDUE 66, partial trackway consisting of two successivepes-manus sets, CDUE 66–2M from lower surface of a loosesandstone block at CDUE locality 12 (Fig. 3A–D; Hmich et al.,2006, Figs. 12F, 13A). CDUE 67C, isolated fragmentary pesfrom reddish gray sandstone layer at CDUE locality 11. Severalisolated imprints of different size and partial trackways recordedby outline drawings and photographs only from CDUE localities10 (levels 1–2) and 12 (Figs. 2A–B, E, 3A, E) and from CDUElocality 13, level 4.
Description: Trackway CDUE 60 (Figs. 2A, C, D) with foursuccessive pes and manus imprints. The trackway pattern showsa narrow trackway width and relatively low pace angulation.Pes broad, rotated strongly outward with respect to the midline,pentadactyl, plantigrade to semi-plantigrade. Digits increase inlength from I–III. Digit IV subequal with III or slightly shorter,V elongated proximally in a massive “heel,” representing abouthalf of total pes length. A phalangeal part is indicated in digit Vby the constricted and partly laterally curved distal portion.Shape of digits broad and rounded, terminating in robust,
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010
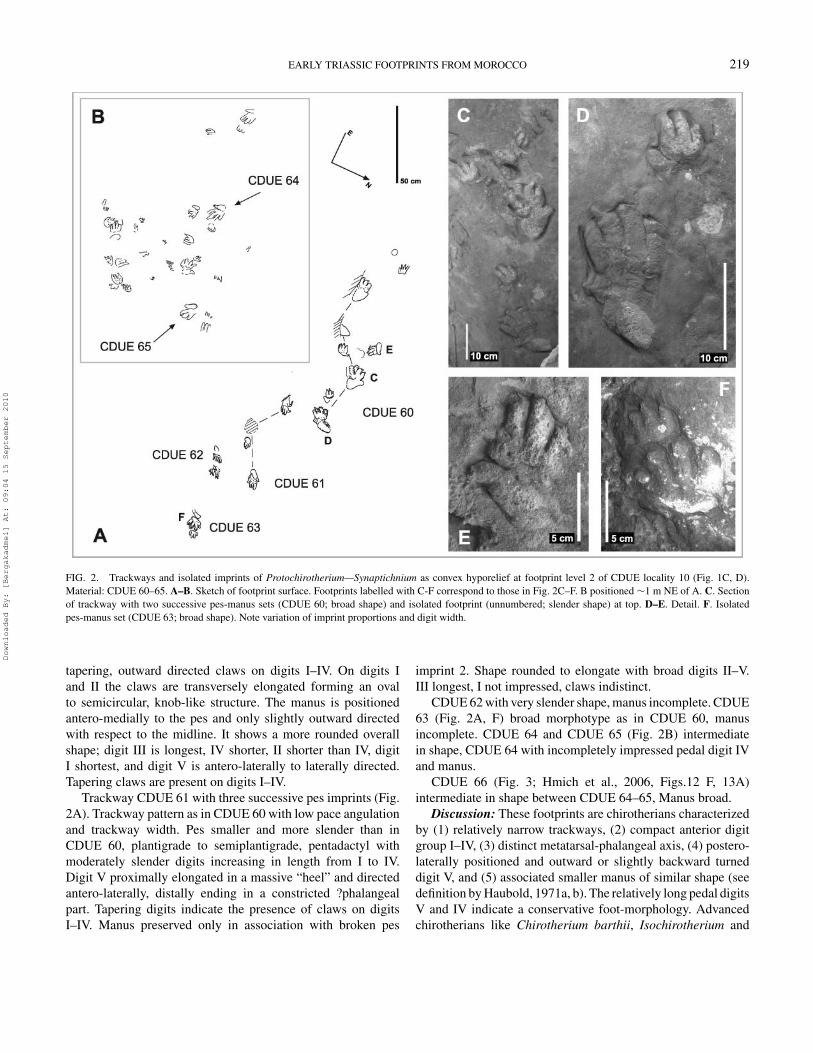
EARLY TRIASSIC FOOTPRINTS FROM MOROCCO 219
FIG. 2. Trackways and isolated imprints of Protochirotherium—Synaptichnium as convex hyporelief at footprint level 2 of CDUE locality 10 (Fig. 1C, D).Material: CDUE 60–65. A–B. Sketch of footprint surface. Footprints labelled with C-F correspond to those in Fig. 2C–F. B positioned ∼1 m NE of A. C. Sectionof trackway with two successive pes-manus sets (CDUE 60; broad shape) and isolated footprint (unnumbered; slender shape) at top. D–E. Detail. F. Isolatedpes-manus set (CDUE 63; broad shape). Note variation of imprint proportions and digit width.
tapering, outward directed claws on digits I–IV. On digits Iand II the claws are transversely elongated forming an ovalto semicircular, knob-like structure. The manus is positionedantero-medially to the pes and only slightly outward directedwith respect to the midline. It shows a more rounded overallshape; digit III is longest, IV shorter, II shorter than IV, digitI shortest, and digit V is antero-laterally to laterally directed.Tapering claws are present on digits I–IV.
Trackway CDUE 61 with three successive pes imprints (Fig.2A). Trackway pattern as in CDUE 60 with low pace angulationand trackway width. Pes smaller and more slender than inCDUE 60, plantigrade to semiplantigrade, pentadactyl withmoderately slender digits increasing in length from I to IV.Digit V proximally elongated in a massive “heel” and directedantero-laterally, distally ending in a constricted ?phalangealpart. Tapering digits indicate the presence of claws on digitsI–IV. Manus preserved only in association with broken pes
imprint 2. Shape rounded to elongate with broad digits II–V.III longest, I not impressed, claws indistinct.
CDUE 62 with very slender shape, manus incomplete. CDUE63 (Fig. 2A, F) broad morphotype as in CDUE 60, manusincomplete. CDUE 64 and CDUE 65 (Fig. 2B) intermediatein shape, CDUE 64 with incompletely impressed pedal digit IVand manus.
CDUE 66 (Fig. 3; Hmich et al., 2006, Figs.12 F, 13A)intermediate in shape between CDUE 64–65, Manus broad.
Discussion: These footprints are chirotherians characterizedby (1) relatively narrow trackways, (2) compact anterior digitgroup I–IV, (3) distinct metatarsal-phalangeal axis, (4) postero-laterally positioned and outward or slightly backward turneddigit V, and (5) associated smaller manus of similar shape (seedefinition by Haubold, 1971a, b). The relatively long pedal digitsV and IV indicate a conservative foot-morphology. Advancedchirotherians like Chirotherium barthii, Isochirotherium and
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010
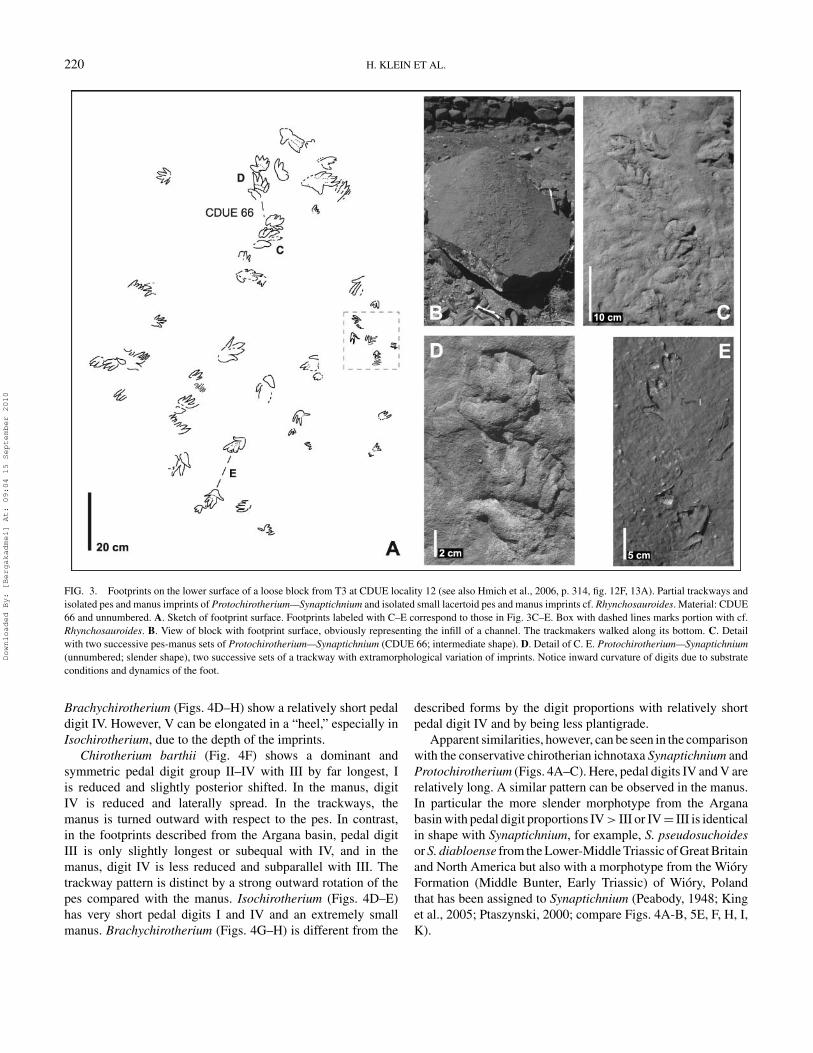
220 H. KLEIN ET AL.
FIG. 3. Footprints on the lower surface of a loose block from T3 at CDUE locality 12 (see also Hmich et al., 2006, p. 314, fig. 12F, 13A). Partial trackways andisolated pes and manus imprints of Protochirotherium—Synaptichnium and isolated small lacertoid pes and manus imprints cf. Rhynchosauroides. Material: CDUE66 and unnumbered. A. Sketch of footprint surface. Footprints labeled with C–E correspond to those in Fig. 3C–E. Box with dashed lines marks portion with cf.Rhynchosauroides. B. View of block with footprint surface, obviously representing the infill of a channel. The trackmakers walked along its bottom. C. Detailwith two successive pes-manus sets of Protochirotherium—Synaptichnium (CDUE 66; intermediate shape). D. Detail of C. E. Protochirotherium—Synaptichnium(unnumbered; slender shape), two successive sets of a trackway with extramorphological variation of imprints. Notice inward curvature of digits due to substrateconditions and dynamics of the foot.
Brachychirotherium (Figs. 4D–H) show a relatively short pedaldigit IV. However, V can be elongated in a “heel,” especially inIsochirotherium, due to the depth of the imprints.
Chirotherium barthii (Fig. 4F) shows a dominant andsymmetric pedal digit group II–IV with III by far longest, Iis reduced and slightly posterior shifted. In the manus, digitIV is reduced and laterally spread. In the trackways, themanus is turned outward with respect to the pes. In contrast,in the footprints described from the Argana basin, pedal digitIII is only slightly longest or subequal with IV, and in themanus, digit IV is less reduced and subparallel with III. Thetrackway pattern is distinct by a strong outward rotation of thepes compared with the manus. Isochirotherium (Figs. 4D–E)has very short pedal digits I and IV and an extremely smallmanus. Brachychirotherium (Figs. 4G–H) is different from the
described forms by the digit proportions with relatively shortpedal digit IV and by being less plantigrade.
Apparent similarities, however, can be seen in the comparisonwith the conservative chirotherian ichnotaxa Synaptichnium andProtochirotherium (Figs. 4A–C). Here, pedal digits IV and V arerelatively long. A similar pattern can be observed in the manus.In particular the more slender morphotype from the Arganabasin with pedal digit proportions IV > III or IV = III is identicalin shape with Synaptichnium, for example, S. pseudosuchoidesor S. diabloense from the Lower-Middle Triassic of Great Britainand North America but also with a morphotype from the WioryFormation (Middle Bunter, Early Triassic) of Wiory, Polandthat has been assigned to Synaptichnium (Peabody, 1948; Kinget al., 2005; Ptaszynski, 2000; compare Figs. 4A-B, 5E, F, H, I,K).
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010

EARLY TRIASSIC FOOTPRINTS FROM MOROCCO 221
FIG. 4. Comparison of chirotherian ichnogenera showing different overallshape and proportions of digits. A–B. Synaptichnium. C. Protochirotherium.D–E. Isochirotherium. F. Chirotherium. G–H. Brachychirotherium. AfterPeabody, 1948; Haubold, 1971b; Karl and Haubold, 1998; Klein and Haubold,2007.
The broad imprints in trackway CDUE 60 show strongresemblance with Protochirotherium wolfhagense, a taxonnamed by Fichter and Kunz, 2004 for material from theDetfurth Formation (Middle Bunter, Early Triassic) of Ger-many (compare Figs. 5A–B, G, J). Furthermore, they seemto be identical in shape with a more robust morphotypefrom the Wiory Formation (Middle Bunter, Early Triassic) ofWiory, Poland, that has been described under Chirotherium,Brachychirotherium, and Isochirotherium (Fuglewicz et al.,1990; Ptaszynski, 2000; Niedzwiedzki and Ptaszynski, 2007)and referred to Protochirotherium (Klein and Haubold, 2007;compare Figs. 5C–D, G, J). The relatively low pace angulationcompared with the data from other chirotherians (see Haubold,1971a) is also in the range of values given for trackwaysfrom Wiory (Ptaszynski, 2000). Fichter and Lepper (1997)described pes and manus imprints of similar shape fromthe Hardegsen Formation (Middle Bunter, Early Triassic) ofGermany and assigned them to Isochirotherium sanctacrucense,an ichnotaxon named by Fuglewicz et al. (1990) for materialfrom the Wiory Formation of Poland. All show long pedal
FIG. 5. Protochirotherium—Synaptichnium from different localities of Eu-rope and from Argana basin, Morocco. A–B. Protochirotherium wolfha-gense, paratype and holotype, Detfurth Fm., Early Triassic, northern Hes-sen, Germany. C–F. Protochirotherium—Synaptichnium (Brachychirotherium,Isochirotherium, Synaptichnium after Ptaszynski, 2000) Wiory Fm., EarlyTriassic, Wiory, Poland. G–L. Protochirotherium—Synaptichnium, T3 ofTimezgadiouine Fm., Early Triassic, Argana basin, Morocco. A-F after Kleinand Haubold, 2007; Ptaszynski, 2000.
digits IV and V, the latter with a massive basal pad. Also, themanus is similar in overall shape and the relatively long digit IV.Similarities can be observed also with footprints from probableEarly Triassic strata of the Teloua Formation of Niger (Ginsburget al., 1968; Taquet, 1976). This material should be restudiedand re-evaluated in a detailed comparison with footprints fromMorocco and Europe.
The described chirotherian footprints from the TanamertMember show strong congruence with Protochirotherium andSynaptichnium and therefore could be assigned to these ichno-genera. However, the determination is complicated by imprintsthat are intermediate in shape between the slender and the broadmorphotype. This might be interpreted as a size-related feature.CDUE 60 (Protochirotherium) is from a large individual,CDUE 61–62 and CDUE 64–66 (Synaptichnium) reflect smalleranimals. CDUE 63 (Figs. 2A, F) is also from a larger individual.The low pes length value is due to the lack of the “heel”in digit V. Another, probably better, explanation for variation
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010

222 H. KLEIN ET AL.
in overall-shape, thickness of digits, shape of claws, etc., isthe influence of the substrate. Extramorphological variationin chirotherian footprints has been demonstrated by Kleinand Haubold (2004). It cannot be decided if the chirotherianfootprints from surfaces of the T3 represent different ichnotaxa.As long as clear differentiation of the footprints from the T3as either Protochirotherium or Synaptichnium is doubtful, weassign them to the plexus Protochirotherium—Synaptichnium,which comprises both robust and slender forms from thesesurfaces.
Trackmaker affinities: Chirotherians can be assigned toarchosaurs. This is suggested by (1) narrow trackways due to theposition of extremities under the body, (2) small manus relativeto the pes indicating an early tendency toward bipedality, and(3) pes and manus imprints with compact anterior digit groupI–IV and postero-laterally positioned, strongly reduced digitV (Haubold, 1971a, b, 1984, 2006). Supposed trackmakersof Protochirotherium—Synaptichnium are either members ofthe crurotarsan clade or Archosauriformes (sensu Sereno,1991; Sereno and Arcucci, 1990) or archosaurs outside of thecrown-group Archosauria (Avesuchia sensu Benton 1999). Acomparison with foot skeletons of these groups is difficultbecause the bone material is mostly fragmentary. If preserved,the pes often lacks the phalangeal part and the exact digitproportions remain obscure. They can be coarsely estimatedonly by the relative length of the metatarsals. In footprintsof the Protochirotherium—Synaptichnium plexus pedal digitsIV and V are relatively long. This indicates trackmakerswith a conservative foot structure as can be seen in somebasal archosaurs, for example proterosuchids, erythrosuchidsor euparkeriids (Parrish, 1989; Gower, 1996). Obviously, inErythrosuchus and Euparkeria pedal digit IV is shorter, and inProterosuchus longer than III (Gower, 1996). This correspondswith digit proportions and variation seen in the footprints fromthe Tanamert member.
In most crurotarsans, pedal digit IV is relatively shortcompared with III, however in some rauisuchians such asTicinosuchus and Batrachotomus from the Middle Triassic,proportions might vary between slightly shorter to subequalwith or longer than III (Krebs, 1965; Gower and Schoch, 2009).The condition in Arganasuchus from the Carnian IrohaleneMember (T5), Timezgadiouine Formation of the Argana basinis unknown (Dutuit, 1979; Jalil and Peyer, 2007). The foot ofknown proterosuchids and erythrosuchids was relatively wideand might have left broad imprints as can be seen, for example,in Protochirotherium. The slender imprints of Synaptichniumrather match the pes of some rauisuchians. Gait and postureof basal archosaur taxa was different, that of Erythrosuchusseems to have been more sprawling (Sereno, 1991; Kuboand Benton, 2007). This contradicts the narrow trackways ofProtochirotherium—Synaptichnium. However, Euparkeria andrauisuchians such as Batrachotomus obviously had a semi-erector erect posture that might better match this pattern (Sereno,
1991; Haubold, 2006; Kubo and Benton, 2007; Gower andSchoch, 2009).
The documented stratigraphic range of proterosuchids isLate Permian to Middle Triassic, that of erythrosuchids Earlyto Middle Triassic (Gower and Sennikov, 2000). Some basalarchosauriforms such as Doswellia are also known from the LateTriassic (Dilkes and Sues, 2009). Occurrences of rauisuchians(which are probably para- or polyphyletic) range from Early toLate Triassic.
The record of pes skeletons of all these groups is toofragmentary and the preserved archosaur footprints from T3might also belong to still unknown osteological taxa. At thepresent state of knowledge, basal archosaurs closely related tothe crowngroup represent the best candidate trackmakers.
Tetrapod footprints cf. Rhynchosauroides Maidwell, 1911(Figs. 3A, 6A–D; Appendix 1)
Type ichnospecies of Rhynchosauroides: Rhynchosau-roides rectipes Maidwell, 1911.
Diagnosis (after Haubold, 1966, 1971a, b): Broadquadrupedal trackways. Manus positioned medially to the pesor being overstepped by the latter at normal gait. Dependingon the velocity of movement, the pace angulation of the pesvaries between 70◦ and 125◦. The pes is slender and digitigrade,the smaller manus more stout and plantigrade. Digits increasein length from I to IV, V is much shorter, I–IV can be curvedinward. Drag marks of the tail occasionally present.
Referred material: Several isolated pes and manus imprintson surface of loose block at CDUE locality 12 (Fig. 3A). Twocollected specimens (CDUE 68C–69C) with isolated pes/manusimprints from CDUE locality 11. CDUE 70C with severaltrackways and isolated pes and manus imprints from CDUElocality 13 (Figs. 6A–D).
Description: Moderately broad trackways with digitigradeto semiplantigrade small pes and manus imprints that are turnedoutward with respect to the midline (Figs. 6A–D). Pes length2–3.5 cm. Manus smaller, at 1.2–2.4 cm. Manus is in frontof the pes or overstepped laterally by the latter. Digits longand slender, increasing in length from I to IV, digit V can berelatively long. Small, tapering claws are present. Digits can becurved inward, nearly straight or slightly curved outward, oftenwith hook-shaped distal ends. Digits I and V are often missing inthe imprint with tendency toward functional tridactyly of digitsII, III, IV. Some manus imprints show a relatively short digit IV(Figs. 6C–D).
Discussion: The small lacertoid imprints with slenderdigits and IV longest resemble Rhynchosauroides especiallyin specimens where the digits are curved inward. The positionof the manus anterior to the pes in some trackways is unusual(CDUE 70C; Figs. 6A–D). However, this pattern may be highlyvariable, dependent on the velocity of movement (see Haubold,1984, fig. 98; Ptaszynski, 2000, Figs.21–26). The relatively longpedal digit V, the outward curvature of digits and the shortmanual digit IV in some imprints is also atypical (Figs. 6C–D),
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010

EARLY TRIASSIC FOOTPRINTS FROM MOROCCO 223
FIG. 6. Cf. Rhynchosauroides (CDUE 70C) from Tanamert Member (T3) of the Timezgadiouine Fm., CDUE locality 13 (level 3). A–B. Several trackwayspartly with tail traces. C. Detail. D. Sketch of C. Notice manus in front of the pes, outward curvature of digit IV, relatively long digit V in the pes, short digit IV inthe manus in C–D.
but it can be due to the dynamics of the foot and differentsubstrate conditions.
The features observed in the small lacertoid footprintsfrom T3 coarsely match those listed in the diagnosis ofRhynchosauroides Maidwell 1911. The described variationof digit proportions and orientation, however, can not beevaluated as either anatomically or extramorphologically based.Furthermore, the material and the knowledge of these earlylacertoid forms from Permo-Triassic strata is still restricted.Therefore, we herein determine these footprints from surfacesof the T3 tentatively as cf. Rhynchosauroides.
Trackmaker affinities: The stratigraphic range of the ich-notaxon Rhynchosauroides from the Late Permian to the EarlyJurassic (Haubold, 1966, 1971a, b; Conti et al., 1977; Silvestriand Szajna, 1993; Lucas, 2007) implies different trackmakerswith similar foot-morphology. Lepidosauromorphs as well asarchosauromorphs were considered (Haubold, 1966, 1971a,b; Avanzini and Renesto, 2002). With respect to their footskeletons showing long and slender, inward curved digits withIV being longest, these taxa are also candidate trackmakers ofcf. Rhynchosauroides from T3.
BIOSTRATIGRAPHIC IMPLICATIONSAgainst the background of the described footprint assem-
blage, the stratigraphic age of the Tanamert Member (T3) ofthe Timezgadiouine Formation has to be discussed. Therefore,known biostratigraphic data from the Timezgadiouine Forma-tion (T3–T5) and from underlying Ikakern Formation (T1–T2)are briefly reviewed.
In early studies, the entire sequence of red-beds in the Arganabasin (T1–T8) was considered to be of Triassic age (Tixeront,
1973; Jones, 1975; Brown, 1980). This was based on the allegedpresence of fossils of the Triassic conifer Voltzia heterophyllaat the base of the succession (De Koning, 1957). The vertebrateskeletal record, however, suggests a Late Permian age for theuppermost part of T2 (Dutuit, 1976b, 1988; Jalil and Dutuit,1996; Jalil, 1999; Jalil and Janvier, 2005; Khaldoune et al.,2009). Tetrapod ichnofaunas occuring at different levels in T2are also typical Permian assemblages (Hminna et al., 2009b;Voigt et al., 2009; Voigt et al., 2010).
The Timezgadiouine Formation (T3–T5) has been differentlyevaluated as Late Triassic or ? Middle to Late Triassic in age(Hofmann et al., 2000; Tourani et al., 2000). A characteristicLate Triassic (Carnian) vertebrate assemblage was describedfrom the base of T5 (Dutuit, 1972, 1976a, 1977a, 1977b;Gauffre, 1993; Jalil, 1999; Lucas, 1998, 1999; Jalil and Peyer,2007; Khaldoune et al., 2009; Steyer et al., 2009). A MiddleTriassic flora and fauna is known from T4 (Aglegal Member)by charophytes, ostracodes, and vertebrates (Medina et al.,2001; Jalil et al., 2009). Vertebrate remains from the top ofT4 include cyclotosaurid temnospondyls typical of the earlyAnisian (Lucas, 1998, 1999; Jalil et al., 2009). Recent discoveryof a characteristic ichnofauna with Chirotherium barthii by theauthors support a Middle Triassic age of T4. The material iscurrently under study.
Thus far, the Tanamert Member (T3) lacked distinct fossilremains, and its geologic age was uncertain. However, it waslinked to the (Triassic) Timezgadiouine Formation (Hofmann etal., 2000; Tourani et al., 2000; see above). The disvovery of aProtochirotherium—Synaptichnium footprint assemblage in T3is of biostratigraphic importance (see Klein and Haubold, 2007;Klein and Lucas, 2010). Protochirotherium—Synaptichniumfrom T3 are identical with ichnofaunas from the Early
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010

224 H. KLEIN ET AL.
Triassic Detfurth and Wiory formations of Germany and Poland(see above). Beyond lithostratigraphic linkages, the EarlyTriassic age of the latter units is based on biostratigraphicdata from conchostracans and palynomorphs (Bachmann andKozur, 2004; Kozur and Bachmann, 2005; Niedzwiedzkiand Ptaszynski, 2007). Chirotherian imprint morphology andtrackway pattern reflect distinct evolutionary stages and de-velopments of the archosaurian locomotor apparatus. Thisimplies that certain ichnogenera and ichno-assemblages arerestricted by their stratigraphic range and accordingly areuseful for biostratigraphy and biochronology (Lucas, 2003,2007). Protochirotherium—Synaptichnium are widespread andcharacteristic of the Early Triassic (Klein and Haubold, 2007;Klein and Lucas, 2010). Derived from the European record,these taxa precede typical Middle Triassic associations withChirotherium barthii that first appear near the Olenekian-Anisian boundary (Peabody, 1948; Haubold, 1967, 1969, 1971a,b, 1984, 2006; Avanzini et al., 2001; Avanzini and Mietto, 2008;Demathieu 1970; Demathieu and Haubold, 1972; Demathieuand Oosterink, 1983; Demathieu and Demathieu, 2004; Gand,1979; Gand and Demathieu, 2005; Haubold and Klein,2000,2002; King et al., 2005; Klein and Haubold, 2004, 2007; Kleinand Lucas, 2010). Their occurence in T3 indicates an EarlyTriassic age of this unit. Furthermore, the stratigraphic rangeof Protochirotherium—Synaptichnium in the Permo-Triassicred-bed sequence of the Argana Basin is restricted to T3. Typicalelements of Middle Triassic ichnofaunas first appear in theoverlying T4 where Protochirotherium—Synaptichnium has notbeen recorded so far.
In summary, T3 is considered to be of Early Tri-assic age based on (1) the presence of a Protochi-rotherium—Synaptichnium dominated ichnofauna, identicalwith characteristic Early Triassic assemblages from Europe;(2) lack of typical Middle Triassic footprints; and (3) under-/overlying Late Permian/ Middle Triassic strata with character-istic ichno- and body fossil record.
CONCLUSIONS AND PERSPECTIVESThe footprints described from the Tanamert Member
(T3) of the Timezgadiouine Formation in the ArganaBasin represent the first occurrence of an assemblage withProtochirotherium—Synaptichnium outside of central Europe.By indirect evidence our results suggest that trackmakers, basalarchosaurs with conservative foot structure, were widespread incentral Pangaea.
Biostratigraphic studies based on the European recordshow that this assemblage is characteristic of the pre-AnisianTriassic and precedes typical Middle Triassic associations withChirotherium barthii (Klein and Haubold, 2007; Klein andLucas, 2010). Its presence in T3 of the Argana basin suggestsan Early Triassic age of this unit. It is the first evidence of thecontinental Early Triassic in Morocco.
The study area offers a high potential for ichnotaxonomic andbiostratigraphic studies based on a well-established database.This is indicated by the abundance of well-preserved footprintsand excellent outcrops that show a more or less completesequence of Permian to Lower Jurassic continental strata. More-over, this knowledge may significantly contribute to locomotorevolution, behaviour, ecology, and paleobiogeography of earlyarchosaurs.
ACKNOWLEDGMENTSThe authors appreciate helpful reviews of Spencer G.
Lucas, New Mexico Museum of Natural History and Science,Albuquerque, and Andrew B. Heckert, Appalachian StateUniversity, Boone, North Carolina, that significantly improvedthe manuscript. Philippe Taquet, Museum National d’HistoireNaturelle Paris, is acknowledged for photos and informationsconcerning Triassic footprints from Niger. A. Hminna thanks theGerman Academic Exchange Service for financial support in theframe of the project DAAD A/07/80641. J.W. Schneider and S.Voigt acknowledge the financial suport of the German ResearchFoundation, project SCHN 408/17–1, “Tetrapod ichnofaunaMorroco.”
REFERENCESAit Chayeb, E., Youbi, N., El-Boukhari, A., Bouabdelli, M., and Amrhar, M.
1998. Le volcanisme permien et mesozoique inferieur du bassin d’Argana(Haut-Atlas occidental, Maroc): un magmatisme intraplaque associe al’ouverture de l’Atlantique central. Journal of African Earth Sciences, 26:499–519.
Avanzini, M. and Mietto, P. 2008. Lower and Middle Triassic footprint-basedBiochronology in the Italian Southern Alps. Oryctos, 8: 3–13.
Avanzini, M. and Renesto, S. 2002. A review of Rhynchosauroides tirolicusAbel, 1926 ichnospecies (Middle Triassic: Anisian-Ladinian) and some in-ferences on Rhynchosauroides trackmaker. Rivista Italiana di Paleontologiae Stratigrafia, 108(1): 51–66.
Avanzini, M., Ceoloni, P., Conti, M.A., Leonardi, G., Manni, R., Mariotti, N.,Mietto, P., Muraro, C., Nicosia, U., Sacchi, E., Santi, G., and Spezzamonte,M. 2001. Permian and Triassic tetrapod ichnofaunal units of northern Italy:Their potential contribution to continental biochronology. Natura Bresciana(Annali del Museo Civico di Scienze Naturali di Brescia), Monografia N,25: 89–107.
Bachmann, G.H. and Kozur, H.W. 2004. The Germanic Triassic: Correlationswith the international chronostratigraphic scale, numerical ages andMilankovitch cyclicity. Hallesches Jahrbuch fur Geowissenschaften B, 26:17–62.
Benton, M. J. 1999. Scleromochlus taylori and the origin of dinosaurs andpterosaurs. Philosophical Transactions of the Royal Society of London B,354: 1423–1446.
Benton, M. J. 2008. Presidential address 2007: The end-Permian massextinction—events on land in Russia. Proceedings of the Geologists’Association, 119: 119–136.
Benton, M. J., Tverdokhlebov, V. P., and Surkov, M. V. 2004. Ecosystemremodeling among vertebrates at the Permian-Triassic boundary in Russia.Nature, 432: 97–100.
Biron, P.E. and Dutuit, J.-M. 1981. Figurations sedimentaires et traces d’activiteau sol dans le Trias de la formation d’Argana et de l’Ourika (Maroc). Bulletindu Museum National d’ Histoire Naturelle Paris, 4e serie, 3, section C, 4:399–427.
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010

EARLY TRIASSIC FOOTPRINTS FROM MOROCCO 225
Botha, J. and Smith, R.M.H. 2006. Rapid vertebrate recuperation in the KarooBasin of South Africa following the end-Permian extinction. Journal ofAfrican Earth Sciences, 45: 502–514.
Brown, R.H. 1980. Triassic rocks of Argana Valley, Southern Morocco andtheir regional structural implications. Bulletin of the American Associationof Petroleum Geologists, 64: 988–1003.
Brusatte, S.L., Benton, M.J., Ruta, M., and Lloyd, G.T. 2008. Superiority,competition, and opportunism in the evolutionary radiation of dinosaurs.Science, 321: 1485–1488.
Conti, M.A., Leonardi, G., Mariotti, N., and Nicosia, U. 1977. Tetrapodfootprints of the “Val Gardena Sandstone” (North Italy). PaleontographicaItaliana N.S., 40, 91 p.
Deenen, M.H.L., Ruhl, M., Bonis, N.R., Krijgsman, W., Kuerschner, W.M.,Reitsma, M., and van Bergen, M.J. 2010. A new chronology for theend-Triassic mass extinction. Earth and Planetary Science Letters, 291:113–125.
Demathieu, G. 1970. Les empreintes de pas de vertebres du Trias de la bordureNord-Est du Massif Central. Cahiers de Paleontologie, 211 p.
De Koning, G. 1957. Geologie des Ida ou Zal (Maroc). Stratigraphie,petrographie et tectonique de la partie SW du bloc occidental du massifancien du Haut-Atlas. Leidse Geologische Mededelingen, 22, 209 p.
Demathieu, G. and Demathieu, P. 2004. Chirotheria and other ichnotaxa of theEuropean Triassic. Ichnos, 11: 79–88.
Demathieu, G. and Haubold, H. 1972. Stratigraphische Aussagen der Tetrapo-denfahrten aus der terrestrischen Trias Europas. Geologie, 21: 802–836.
Demathieu, G. and Haubold, H. 1982. Reptilfahrten aus dem MittlerenBuntsandstein von Hessen. Hallesches Jahrbuch fur Geowissenschaften,7: 97–110.
Demathieu, G. and Oosterink, H.W. 1983. Die Wirbeltier-Ichnofauna aus demUnteren Muschelkalk von Winterswijk (Die Reptilfahrten aus der Mitteltriasder Niederlande). Staringia, 7: 1–51.
Dilkes, D. and Sues, H.-D. 2009. Redescription and phylogenetic relation-ships of Doswellia kaltenbachi (Diapsida: Archosauriformes) from theUpper Triassic of Virginia. Journal of Vertebrate Paleontology, 29(1):58–79.
Dutuit, J.-M. 1972. Decouverte d’un Dinosaure ornithischien dans le Triassuperieur de I’Atlas occidental marocain. Comptes rendus de l
′Academie
des Sciences, 275: 2841–2844.Dutuit, J.-M. 1976a. Introduction a l’etude paleontologique du Trias continental
marocain. Description des premiers Stegocephales recueillis dans le couloird’Argana (Atlas occidental). Memoires du Museum National d’ HistoireNaturelle, Paris, C, 36: 1–253.
Dutuit, J.-M. 1976b. II est probable que les Rhynchocephales sont representesdans la faune du Trias marocain. Comptes Rendus de l’Academie desSciences, Paris, 238: 483–486.
Dutuit, J.-M. 1977a. Paleorhinus magnoculus, Phytosaure du Trias superieurde I’Atlas marocain. Annales de l’Universite de Provence, GeologieMediterraneenne, 4(3): 255–268.
Dutuit, J.-M. 1977b. Description du crane de Angistorhinus talainti n. sp.un nouveau phytosaure du Trias atlasique marocain. Bulletin du MuseumNational d’Histoire Naturelle, Paris, Serie 3, 489: 297–336.
Dutuit, J.-M. 1979. Un pseudosuchien du Trias continental marocain. Annalesde Paleontologie (Vertebres), 65: 55–68.
Dutuit, J.-M. 1988. Diplocaulus minimus n. sp. (Amphibia, Nectridea)lepospondyle de la formation d’Argana, dans l’Atlas occidental marocain.Comptes rendus de l’Academie des Sciences, Paris, 307: 851–854.
Fichter, J. and Lepper, J. 1997. Die Fahrtenplatte vom Heuberg bei Gieselwerder.Philippia, 8/1: 35–60.
Fichter, J. and Kunz, R. 2004. New genus and species of chirotheroid tracks in theDetfurth-Formation (Middle Bunter, Lower Triassic) of Central Germany.Ichnos, 11: 183–193.
Fuglewicz, R., Ptaszynski, T., and Rdzanek, K. 1990. Lower Triassic footprintsfrom the Swietokrzyskie (Holy Cross) Mountains, Poland. Acta Palaeonto-logica Polonica, 35: 109–164.
Gand, G. 1979. Note sur quelques nouvelles pistes de reptiles observees dans leTrias moyen du Plateau d’Antully (Saone-et-Loire—France). Bulletin de laSociete d’Histoire Naturelle d’Autun, 92: 7–20.
Gand, G. and Demathieu, G. 2005. Les pistes dinosauroides du Trias moyenfrancais: Interpretation et reevaluation de la nomenclature. Geobios, 38:725–749.
Gauffre, F.-X. 1993. The prosauropod dinosaur Azendohsaurus laaroussii fromthe Upper Triassic of Morocco. Palaeontology, 36(4): 897–908.
Ginsburg, L., Lapparent, A.F. de, and Taquet, P. 1968. Piste de Chirotheriumdans le Trias du Niger. Comptes Rendus de l’ Academie des Sciences ParisSerie D, 266: 2056–2058.
Gower, D.J. 1996. The tarsus of erythrosuchid archosaurs, and implications forearly diapsid phylogeny. Zoological Journal of the Linnean Society, 116:347–375.
Gower, D.J. and Schoch, R.R. 2009. Postcranial anatomy of the rauisuchian ar-chosaur Batrachotomus kupferzellensis. Journal of Vertebrate Paleontology,29(1): 103–122.
Gower, D.J. and Sennikov, A.G. 2000. Early archosaurs from Russia. In Benton,M.J., Shishkin, M.A., Unwin, D.M., and Kurochkin, E.N. (eds.), The Age ofDinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge,pp. 140–159.
Haubold, H. 1966. Therapsiden- und Rhynchocephalen-Fahrten aus demBuntsandstein Sudthuringens. Hercynia, NF, 3(2): 147–183.
Haubold, H. 1967. Eine Pseudosuchier-Fahrtenfauna aus dem BuntsandsteinSudthuringens. Hallesches Jahrbuch fur Mitteldeutsche Erdgeschichte, 8:12–48.
Haubold, H. 1969. Parallelisierung terrestrischer Ablagerungen der tieferenTrias mit Pseudosuchier-Fahrten. Geologie, 18: 836–843.
Haubold, H. 1971a. Die Tetrapodenfahrten des Buntsandsteins. Palaontologis-che Abhandlungen A, IV: 395–548.
Haubold, H. 1971b. Ichnia Amphibiorum et Reptiliorum fossilium. Encyclope-dia of Paleoherpetology, 18: 1–124.
Haubold, H. 1984. Saurierfahrten. Wittenberg, Ziemsen, 231 p.Haubold, H. 2006. Die Saurierfahrten Chirotherium barthii Kaup, 1835—das
Typusmaterial aus dem Buntsandstein bei Hildburghausen/Thuringen unddas Chirotherium-Monument. Veroffentlichungen Naturhistorisches Mu-seum Schleusingen, 21: 3–31.
Haubold, H. and Klein, H. 2000. Die dinosauroiden FahrtenParachirotherium–Atreipus–Grallator aus dem unteren Mittelkeuper(Obere Trias: Ladin, Karn, ?Nor) in Franken. Hallesches Jahrbuch furGeowissenschaften B, 22: 59–85.
Haubold, H. and Klein, H. 2002. Chirotherien und Grallatoriden aus derUnteren bis Oberen Trias Mitteleuropas und die Entstehung der Dinosauria.Hallesches Jahrbuch fur Geowissenschaften B, 24: 1–22.
Hmich, D., Schneider, W., Saber, H., Voigt, S., and El Wartiti, M. 2006. Newcontinental Carboniferous and Permian faunas of Morocco: Implications forbiostratigraphy, palaeobiogeography and palaeoclimate. Geological SocietyLondon, Special Publications, 265: 297–324.
Hminna, A., Saber, H., Schneider, J.W., Klein, H., Voigt, S., and Hmich,D. 2009a. Brachychirotherium ichnofauna from the Upper Triassic ofOued Oum Er Rbiaa Formation of Sidi Saıd M’aachou basin, CoastalMeseta, Morocco. 1st International Congress on North African VertebratePalaeontology, Abstracts: 32.
Hminna, A., Voigt, S., Saber, H., Schneider, J.W., and Hmich, D. 2009b.Tetrapod footprints from Ikakern Formation (Argana Basin, Western HighAtlas, Morocco). 1st International Congress on North African VertebratePalaeontology, Abstracts: 61–62.
Hofmann, A., Tourani, A., and Gaupp, R. 2000. Cyclicity of Triassic to LowerJurassic continental red beds of the Argana Valley, Morocco: Implicationsfor palaeoclimate and basin evolution. Palaeogeography, Palaeoclimatology,Palaeoecology, 161: 229–266.
Jalil, N.-E. 1999. Continental Permian and Triassic vertebrate localities fromAlgeria and Morocco and their stratigraphical correlations. Journal ofAfrican Earth Sciences, 29(1): 219–226.
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010

226 H. KLEIN ET AL.
Jalil, N.-E. and Dutuit, J.-M. 1996. Permian captorhinid reptiles from the ArganaFormation, Morocco. Palaeontology, 39(4): 907–918.
Jalil, N.-E. and Janvier, P. 2005. Les pareiasaures (Amniota, Parareptilia) duPermien superieur du Bassin d’Argana, Maroc. Geodiversitas, 27: 35–132.
Jalil, N.-E. and Peyer, K. 2007. A new rauisuchian (Archosauria, Suchia) fromthe Upper Triassic of the Argana Basin, Morocco. Palaeontology, 50(2):417–430.
Jalil, N.-E., Janvier, P., and Steyer, J.-S. 2009. A new cyclotosaurid (Amphibia,Temnospondyli) from the Triassic of Argana Basin (High Atlas Mountains,Morocco); biostratigraphic implications. 1st International Congress onNorth African Vertebrate Palaeontology, Abstracts: 36–37.
Jones, D.F. 1975. Stratigraphy, environments of deposition, petrology, age andprovenance of the basal red beds of the Argana valley, western High AtlasMountains, Morocco. New Mexico Institute of Mining and Technology,Master Thesis, 148 p.
Karl, C. and Haubold, H. 1998. Brachychirotherium aus dem Coburger Sand-stein (Mittlerer Keuper, Karn/Nor) in Nordbayern. Hallesches Jahrbuch furGeowissenschaften B, 20: 33–58.
Khaldoune, F., Jalil, N.-E., and Peyer, K. 2009. Les vertebres fossiles du bassind’Argana (Permien-Trias, Haut Atlas occidental): Liste faunique et niveauxstratigraphiques. 1st International Congress on North African VertebratePalaeontology, Abstracts: 68.
King, M.J., Sarjeant, W.A.S., Thompson, D.B., and Tresise, G. 2005. Arevised systematic ichnotaxonomy and review of the vertebrate footprintichnofamily Chirotheriidae from the British Triassic. Ichnos, 12:241–299.
Klein, H. and Haubold, H. 2004. Uberlieferungsbedingte Variation beiChirotherien und Hinweise zur Ichnotaxonomie nach Beispielen aus derMittel- bis Ober-Trias (Anisium–Karnium) von Nordbayern. HalleschesJahrbuch fur Geowissenschaften B, 26: 1–15.
Klein, H. and Haubold, H. 2007. Archosaur footprints—potential forbiochronology of Triassic continental sequences. New Mexico Museum ofNatural History and Science, Bulletin, 41: 120–130.
Klein, H. and Lucas, S.G. 2010. Tetrapod footprints—their use in biostratigraphyand biochronology of the Triassic. Geological Society, London, SpecialPublications, 334: 419–446.
Klein, H., Saber, H., Voigt, S., Schneider, J.W., Hmich, D., and Hminna, A. 2009.An archosaur-dominated footprint assemblage in Permo-Triassic red-bedsof Morocco and the global record of early chirotherians. 1st InternationalCongress on North African Vertebrate Palaeontology, Abstracts: 18.
Kotanski, Z., Gierlinski, G., and Ptaszynski, T. 2004. Reptile tracks (Roto-dactylus) from the Middle Triassic of the Djurdjura Mountains in Algeria.Geological Quarterly, 48(1): 89–96.
Kozur, H.W. and Bachmann, G.H. 2005. Correlation of the Germanic Triassicwith the international scale. Albertiana, 32: 21–35.
Krebs, B. 1965. Ticinosuchus ferox n. g. n. sp., ein neuer Pseudosuchier aus derTrias des Monte San Giorgio. Schweizer Palaontologische Abhandlungen,81: 1–141.
Kubo, T. and Benton, M.J. 2007. Evolution of hindlimb posture in ar-chosaurs: Limb stresses in extinct vertebrates. Palaeontology, 50(6): 1519–1529.
Leonardi, G. (ed.). 1987. Glossary and Manual of Tetrapod Footprint Palaeoich-nology. Ministerio Minas Energie, Departemento Nacional da ProducaoMineral, Brasilia, 117 p.
Lucas, S.G. 1998. Global Triassic tetrapod biostratigraphy and biochronology.Palaeogeography, Palaeoclimatology, Palaeoecology, 143: 347–384.
Lucas, S.G. 1999. Tetrapod-based correlation of the nonmarine Triassic.Zentralblatt fur Geologie und Palaontologie I, 7–8: 497–521.
Lucas, S.G. 2003. Triassic tetrapod footprint biostratigraphy and biochronology.Albertiana, 28: 75–84.
Lucas, S.G. 2007. Tetrapod footprint biostratigraphy and biochronology. Ichnos,14: 5–38.
MacDonald, D.I.M., Isbell, J.L., and Hammer, W.R. 1991. Vertebrate trackwaysfrom the Triassic Fremouw Formation, Queen Alexandra Range, Antarctica.Antarctic Journal of the United States, 26: 20–21.
Maidwell, F.T. 1911. Notes on footprints from the Keuper of Runcorn Hill.Proceedings of the Liverpool Geological Society, 11: 140–152.
Medina, F., Vachard, D., Colin, J.-P., Ouarhache, D.D., and Ahmamou, M.2001. Charophytes et ostracodes du niveau carbonate de Taourirt Imzilen(Membre d’Aglegal, Trias d’Argana); implications stratigraphiques. Bulletinde l’ Institut scientifique, Rabat, section Sciences de la Terre, 23: 21–26.
Niedzwiedzki, G. and Ptaszynski, T. 2007. Large Chirotheriidae tracks in theEarly Triassic of Wiory, Holy Cross Mountains, Poland. Acta GeologicaPolonica, 57(3): 325–342.
Nopcsa, F. von. 1923. Die Familien der reptilien. Fortschritte der Geologie undPalaontologie, 2: 1–210.
Olsen, P.E. 1997. Stratigraphic record of the early Mesozoic breakup ofPangea in the Laurasia-Gondwana rift system. Annual Review of Earthand Planetary Science, 25: 337–401.
Olsen, P.E. and Et-Touhami, M. (eds.). 2008. Fieldtrip Guidebook. CentralAtlantic Conjugate Margins Conference, Halifax.
Olsen, P.E., Kent, D.V., Fowell, S.J., Schlische, R.W., Withjack, M.O., andLeTourneau, P.M. 2000. Implications of a comparison of the stratigraphyand depositional environments of the Argana (Morocco) and Fundy (NovaScotia, Canada) Permian-Jurassic basins. In Oujidi, M. and Et-Touhami, M.(eds.), Le Permien et le Trias du Maroc. Actes de la Premiere Reunion duGroupe Marocain du Permien et du Trias, Hilal Impression Oujda, 165–183.
Olsen, P.E., Kent, D.V., Et-Touhami, M., Fowell, S.J., and Witte, B. 2002.Cyclicity, time scale, and correlation of the Triassic and Early Jurassic riftstrata of the Argana basin, Morocco. In Essamoud, R. and Ekamel, F. (eds.),Troiseme Reunion du Groupe Marocain du Permien et du Trias, Casablanca,18–20 Avril, Abstracts, pp. 29–30.
Padian, K., Li, C., and Pchelnikova, J. 2010. The trackmaker of Apatopus (LateTriassic, North America): Implications for the evolution of archosaur stanceand gait. Palaeontology, 53(1): 175–189.
Parrish, M.J. 1989. Phylogenetic patterns in the manus and pes of earlyMesozoic Archosauromorpha. In Gillette, D.D. and Lockley, M.G. (eds.),Dinosaur Tracks and Traces. Cambridge, UK, Cambridge University Press,pp. 249–258.
Peabody, F.E. 1948. Reptile and amphibian trackways from the MoenkopiFormation of Arizona and Utah. University of California Publications,Bulletin of the Department of Geological Sciences, 27: 295–468.
Ptaszynski, T. 2000. Lower Triassic vertebrate footprints from Wiory, HolyCross Mountains, Poland. Acta Palaeontologica Polonica, 45: 151–194.
Retallack, G.J. 1996. Early Triassic therapsid footprints from the Sydney basin,Australia. Alcheringa, 20: 301–314.
Sahney, S. and Benton, M.J. 2008. Recovery from the most profound massextinction of all time. Proceedings of the Royal Society B, 275: 759–765.
Schlische, R.W. 2003. Progress in understanding the structural geology, basinevolution, and tectonic history of the eastern North American rift system. InLe Tourneau, P.M. and Olsen, P.E. (eds.), The Great Rift Valleys of Pangaeain Eastern North America, Vol. 1. Columbia University Press, New York,pp. 21–64.
Sereno, P.C. 1991. Basal archosaurs: phylogenetic relationships and functionalimplications. Journal of Vertebrate Paleontology, 11 (Suppl.): 1–65.
Sereno, P.C. and Arcucci, A.B. 1990. The monophyly of crurotarsal archosaursand the origin of bird and crocodile ankle joints. Neues Jahrbuch furGeologie und Palaontologie, Abhandlungen, 180: 21–52.
Silvestri, S.M. and Szajna, M.J. 1993. Biostratigraphy of vertebrate footprints inthe Late Triassic section of the Newark Basin, Pennsylvania: Reassessmentof stratigraphic ranges. In Lucas, S.G. and Morales, M. (eds.), TheNonmarine Triassic. New Mexico Museum of Natural History and ScienceBulletin, 3: 439–444.
Steyer, J.-S., Jalil, N.-E., Taquet, P., Bitam, I., Aitouali, R., Nedjari, A., Bouzidi,O., and Kedadra, B. 2009. Temnospondyls from North Africa: Reviewand perspectives. 1st International Congress on North African VertebratePalaeontology, Abstracts: 62.
Taquet, P. 1976. Geologie et Paleontologie du gisement de Gadoufaoua (Aptiendu Niger). Cahiers de Paleontologie, CNRS, Paris: 1–191.
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010

EARLY TRIASSIC FOOTPRINTS FROM MOROCCO 227
Tixeront, M. 1973. Lithostratigraphie et mineralisation cupriferes et uraniferesstratiformes syngenetiques et familieres des formations detriques permo-triasiques du Couloir d’Argana (Haut-Atlas occidental, Maroc). Notes etMemoires du Service Geologique du Maroc, 33(249): 147–177.
Tixeront, M. 1974. Carte geologique et mineralogique du Couloir d’Argana,1/100 000. Edition du Service Geologique du Maroc, Notes et Memoires205.
Et-Touhami, M., Olsen, P.E., Kent, D.V., Fowell, S.J., and Whiteside, J.H.2008. Tectonostratigraphy, biostratigraphy, and magnetostratigraphy of LateTriassic-Early Jurassic red beds in Morocco. First International Conference& Exhibition, Marrakech, Morocco, Abstracts.
Tourani, A., Lund, J.J., Benaouiss, N., and Gaupp, R. 2000. Stratigraphy ofTriassic syn-rift deposition in Western Morocco. Zentralblatt fur Geologieund Palaontologie Teil I, 9–10: 1193–1215.
Voigt, S., Saber, H., Schneider, J.W., Hminna, A., Hmich, D., and Klein, H. 2009.Large imprints of Hyloidichnus Gilmore, 1927 from the Permian of Moroccoin the light of captorrhinid phylogeny and biogeography. 1st InternationalCongress on North African Vertebrate Palaeontology, Abstracts: 22.
Voigt, S., Hminna, A., Saber, H., Schneider, J.W., and Klein, H. 2010. Tetrapodfootprints from the uppermost level of the Permian Ikakern Formation(Argana basin, western High Atlas, Morocco). Journal of African EarthSciences, 57: 470–478.
APPENDIX 1Overview of locations and numbered specimens of Protochirotherium—Synaptichnium and cf. Rhynchosauroides.
CDUE location Coordinates CDUE specimen
10-Tikida N 30◦ 49’ 22.9” W 009◦04’ 58.9”
60–65 (Protochirotherium-Synaptichnium)
11 -Aguenza N 30◦ 50’ 19.05” W 009◦04’ 12.06”
67 (Protochirotherium-Synaptichnium)68–69 (cf. Rhynchosauroides)
12 -Aguenza N 30◦ 50’ 17. 10” W 009◦04’ 15.27”
66 (Protochirotherium-Synaptichnium)
13-Tikida N 30◦ 49’ 22.87” W 009◦05’ 00.78”
70 (cf. Rhynchosauroides)
APPENDIX 2Footprint and trackway parameters of Protochirotherium—Synaptichnium from CDUE localities 10, 12, and 13.
Measurements in centimeter and degrees. Asterisk indicates estimated data from incomplete imprints and trackways.
CDUE 60–1 60–2 60–4 61–1 61–2 61–3 62 63 64 65 66–1 66–2
Pl 19.0 16.0 16.0 13.0∗ — 13.0 12.0 11.0 13.5 12.0 10.0 9.5pw 11.5 14.5 11.0∗ 6.5 — 7.0 5.2 9.0 8.0 8.0∗ 6.5 7.0pl/pw 1.7 1.1 1.5∗ 2.0∗ — 1.9 2.3 1.2 1.7 1.5∗ 1.5 1.4mL 9.0 8.0 — — 7.5 — 6.0 5.5∗ 5.5∗ — 6.0 6.0mw 7.0∗ 8.0 — — 5.0∗ — 5.5∗ 5.5∗ 5.5∗ — 6.0 6.0mL/mw 1.3∗ 1.0 — — 1.5∗ — 1.1∗ 1.0∗ 1.0∗ — 1.0 1.0SL 57.0 51.0 — — — — — —
TW 18.0∗ 15.0∗ — — — — — —
PA 120–130∗ 120∗ — — — — — —
Downloaded By: [Bergakadmei] At: 09:04 15 September 2010




















