
Effects of temperature, body size, and starvation on feeding in a major echinoderm predator

229
J. Paleont., 80(2), 2006, pp. 229–244Copyright � 2006, The Paleontological Society0022-3360/06/0080-229$03.00
AN EDRIOASTEROID-DOMINATED ECHINODERM ASSEMBLAGE FROM ALOWER PENNSYLVANIAN MARINE CONGLOMERATE IN OKLAHOMA
COLIN D. SUMRALL,1 JAMES SPRINKLE,2 AND RENA M. BONEM3
1Department of Earth and Planetary Sciences, University of Tennessee, Knoxville 37996, �[email protected]�, 2Department of GeologicalSciences, Jackson School of Geosciences, University of Texas, Austin 78712-0254, �[email protected]�, and 3Department of Geology, Baylor
University, Waco, Texas 76798-7354, �Rena�[email protected]�
ABSTRACT—Four species of isorophinid edrioasteroids, Ulrichidiscus forbesi n. sp., Neoisorophusella whitesidei n. sp., Postibullawestergaardi n. sp., and Parapostibulla graysoni n. sp., are described from unnamed unit 1 of the Golf Course Formation, DornickHills Group, Lower Pennsylvanian (Morrowan), at Forbes Ranch, north of Marietta, southern Oklahoma. This locality is the first reportof edrioasteroids attached to extraformational clasts, and they exhibit extremely high specimen density and the highest taxonomicdiversity known from the post-Ordovician. The associated fauna consists of the stemless crinoid Paragassizocrinus tarri, serpulid wormtubes, and the acrothoracic barnacle boring, Rogerella, along with a few brachiopods, rugose corals, and other open marine organisms.Edrioasteroids show no preference for clast type, size, or shape. Intraspecific spatial competition is shown by polygonal specimenoutlines in highly crowded conditions, whereas interspecific competition results in specimen overgrowth in a predictable manner.
INTRODUCTION
ISOROPHIDS ARE a clade of dorsoventrally flattened edrioasteroidechinoderms that spanned the Paleozoic, and are presently
known from North America, Asia, Europe, and Australia. Thisgroup lived in a variety of open-marine habitats, but because oftheir attachment disk (peripheral rim), they required a hard sub-strate for attachment (Bell, 1976). Throughout their history, iso-rophids have utilized several types of hard substrate, includingexposed concretions (Wilson, 1985), hardgrounds (Koch andStrimple, 1968; Brett and Liddell, 1978; Waddington, 1980;Guensburg and Sprinkle, 1994; Sumrall, 2001; Sumrall et al.,2001; Cornell et al., 2003), shell pavements (Kammer et al., 1987;Meyer, 1990), isolated skeletal debris (Sumrall, 1992, 1996; Sum-rall et al., 2000), microbial buildups and encrusting bryozoan col-onies (Guensburg and Sprinkle, 1994; Sumrall et al., 2001), andliving organisms (Bell, 1976; Sumrall, 2000; Glass, in press).However, the locality described here is the first known exampleof edrioasteroids attached to cobbles and pebbles in extraforma-tional conglomerate. These isolated hard substrates are shared byother encrusting organisms, including acrothoracic barnacle bor-ings, corals, serpulid worms, and bryozoans. The stalkless crinoid,Paragassizocrinus tarri Strimple, 1938, is found in small numbersassociated with the edrioasteroids and in large numbers in theoverlying bed. This faunal association for ParagassizocrinusMoore and Plummer, 1940 is unusual and contrasts sharply withthe previous paleoenvironmental interpretation of dark calcareousshales and marls as prodeltaic facies prograding into shallow seas,or brief shallow-water transgressions onto the delta plain (Etten-sohn, 1980). The conglomerate and edrioasteroid associationtherefore shows an environmental expansion of the paleoenviron-mental tolerances of Paragassizocrinus.
This locality is also unusual because of its age and edrioasteroiddiversity. Until recently, Pennsylvanian edrioasteroids were poor-ly known (Fraunfelter and Utgaard, 1970; Bell, 1976), but morerecently a clearer picture of true Pennsylvanian edrioasteroid di-versity has emerged (Sumrall, 1992; Sumrall and Bowsher, 1996;Sumrall et al., 2000). These studies have shown that rather thanbeing a time of decline, three different clades of edrioasteroidspassed into the Pennsylvanian, and one (Yukutidiscus Arendt,1983) persisted into the Permian (Arendt, 1983). The ForbesRanch locality is significant in that 1) two additional clades (Ul-richidiscus Bassler, 1935 and Postibulla Bell, 1976) are extendedinto the Pennsylvanian, and 2) the presence of four species co-occurring is equal to the most diverse Ordovician edrioasteroidoccurrences (Sumrall et al., 2001).
Original discovery and collection of edrioasteroids.⎯TheForbes Ranch locality north of Marietta in southern Oklahomawas discovered by E. Westergaard on a Baylor University geo-logical field trip led by R. Grayson in the fall of 1981. Whiteside(1987) did the initial study on the locality, describing the occur-rence and paleoecology and assigning the edrioasteroids to threeunnamed genera and species. Since that time, numerous facultyand students at Baylor University and the University of Texas atAustin have collected additional specimens described in thisstudy.
STRATIGRAPHY AND DEPOSITIONAL SETTING
The extraformational clasts to which the edrioasteroids are at-tached occur within a 15 cm conglomerate layer with a calcareoussilty matrix and scattered, well-rounded sand grains. This layeroccurs within ‘‘unnamed unit one’’ of the Lower PennsylvanianGolf Course Formation, Dornick Hills Group (Fig. 1), which isexposed on the eastern side of the Overbrook Anticline in thesouthern Ardmore Basin. Unnamed unit 1 lies between the Jolliffand Otterville Members and is dominantly shale with thin lime-stones and conglomerates having a total thickness of 49.4 m atthe Forbes Ranch locality north of Marietta, Oklahoma (Dickey,1982). Whiteside (1987, see also Sutherland and Grayson, 1992)recorded a thickness of about 280 m for unnamed unit 1 at thislocality, but apparently included faulted and folded sequences.The conglomerate layer containing the edrioasteroids is nearlyvertical in the bank of a small stream, and the north end of thissmall exposure has an overturned fold that apparently resultedfrom soft-sediment deformation. The unit is highly weathered andsome clasts have slickenslides on their surfaces that distort theattached edrioasteroids. Although thicker conglomerate beds oc-cur up- and down-section, this thin conglomerate layer is the onlyone that has been found with bored and encrusted clasts. Thislayer contains conodonts from the Idiognathoides sinuatus Zone,indicating a Morrowan age for the unit (Sutherland and Grayson,1992).
Most pebbles and cobbles with attached edrioasteroids are wellrounded to subrounded, with only a small percent showing sharpedges, probably from recent breakage. Disks (47.3%) are some-what more common than blocky, subrounded clasts (33.4%), withfewer cylinders (11.3%), and relatively few oblate spheroids, bro-ken disks, and spheres (7.8% total). Carbonate, sandstone, andsiltstone lithologies compose most of the disks and cylinders,whereas chert forms most of the blocky clasts, probably becausethey began as broken-up beds and were less easily rounded. The

230 JOURNAL OF PALEONTOLOGY, V. 80, NO. 2, 2006
FIGURE 1—Stratigraphic location of the thin conglomerate layer with ed-rioasteroids and acrothoracic barnacle borings (arrow) in unnamed unit1 of the Golf Course Formation (Lower Pennsylvanian), and sourcerock for 220 clasts with edrioasteroids in the Baylor Collection thatcould be identified as to their unit of origin lower in the section (num-bers in the pebbles column). Abbreviations: Ls � limestone, Sh �
←
shale, Fm � Formation, Gr � Group. Stratigraphic column slightlymodified from Ham (1969, fig. 4).
few spheres are chert and appear to have originated as concentricconcretions.
Clasts within the conglomerate layer represent units rangingfrom the Ordovician Arbuckle and Simpson groups, and Silurianand Devonian Hunton Group (including Henryhouse, Haragan,Bois d’Arc, and Woodford formations) to the Mississippian Syc-amore Formation (Fig. 1). These clasts were probably eroded offthe Criner Uplift during an early pulse of the Wichita—ArbuckleOrogeny and were carried eastward by braided streams on analluvial fan to be deposited as a prograding fan delta sequence ina shallow-marine environment (Dickey, 1982). The number ofclasts from each unit found in this conglomerate layer probablyreflects the resistance to weathering of the source lithology ratherthan the thickness of each formation or the exposed area beingeroded.
PALEOECOLOGY
Faunal composition.⎯The fauna collected from the outcrop ofunnamed unit 1 of the Golf Course Formation at the Forbes Ranchlocality includes hard-substrate sessile organisms, soft-substratesessile organisms, and vagrant benthic/nektic forms. Organismsthat attached to the conglomerate cobbles and pebbles include (a� abundant, c � common, u � uncommon, r � rare): the ac-rothoracic barnacle boring Rogerella Saint-Seine, 1951 (a) (Fig.2.19), four genera of edrioasteroids described here (c, c, u, andu), small encrusting worm tubes (c) (Fig. 2.13, 2.17), the solitaryrugose coral Lophotichium amoenum Moore and Jeffords, 1945(u) (Fig. 2.16), the alloporid coral Cladochonus M’Coy, 1847 (r)(Fig. 2.9), small erect bryozoan colonies (u) and a large encrust-ing bryozoan (r) (Fig. 2.8), and small spiriferid and productidbrachiopods—mostly fragmental (u) (Fig. 2.14, 2.18). Soft-sub-strate and vagrant/nektic organisms include: Paragassizocrinustarri (c) (Fig. 2.1–2.7, 2.11, 2.12), several unidentified crinoidspreserved as isolated skeletal debris (u) (Fig. 2.10, 2.15), severalconodonts, including zone markers Idiognathodus delicates Gun-nell, 1931 and I. sinuatus Harris and Hollingsworth, 1933 (r),unidentified small ostracodes (u), an unidentified rostrochonch (r),echinoid spines and plates (u), and holothurian sclerites (wheelsand hooks) (r). This fauna is consistent with a shallow-marinedepositional setting, although the diversity is unusually low for afauna of Morrowan age.
Pebble preference.⎯The edrioasteroids from the Forbes Ranchlocality show no preference for any particular substrate, and arefound attached to clasts of sandstone, siltstone, limestone, andchert (Fig. 1). Large edrioasteroid concentrations of up to 4,000/m2 are also found on a number of different lithologies. There isapparently no preference for clast size with edrioasteroids beingfound on cobbles as large as 112 mm long and on pebbles assmall as 7 mm long. Some very large clasts are uncolonized whileothers are densely colonized (Fig. 3.11, 3.12). Likewise smallpebbles are often found either colonized or uncolonized. In somecases, edrioasteroids are found attached to pebbles much smallerthan their diameter (Fig. 3.2–3.4, 3.7). This strongly suggests thatedrioasteroids were incapable of moving from pebble to pebbleas they outgrew their attachment surface. Edrioasteroids are alsofound attached to both bored and unbored pebbles.
Unlike edrioasteroids, acrothoracic barnacles were much moreselective in pebble colonization. Limestone was preferred to allother clast types, and borings in chert are unknown in the sample.In some cases a single clast that had both limestone and chert

231SUMRALL ET AL.—EDRIOASTEROIDS ON PEBBLES
→
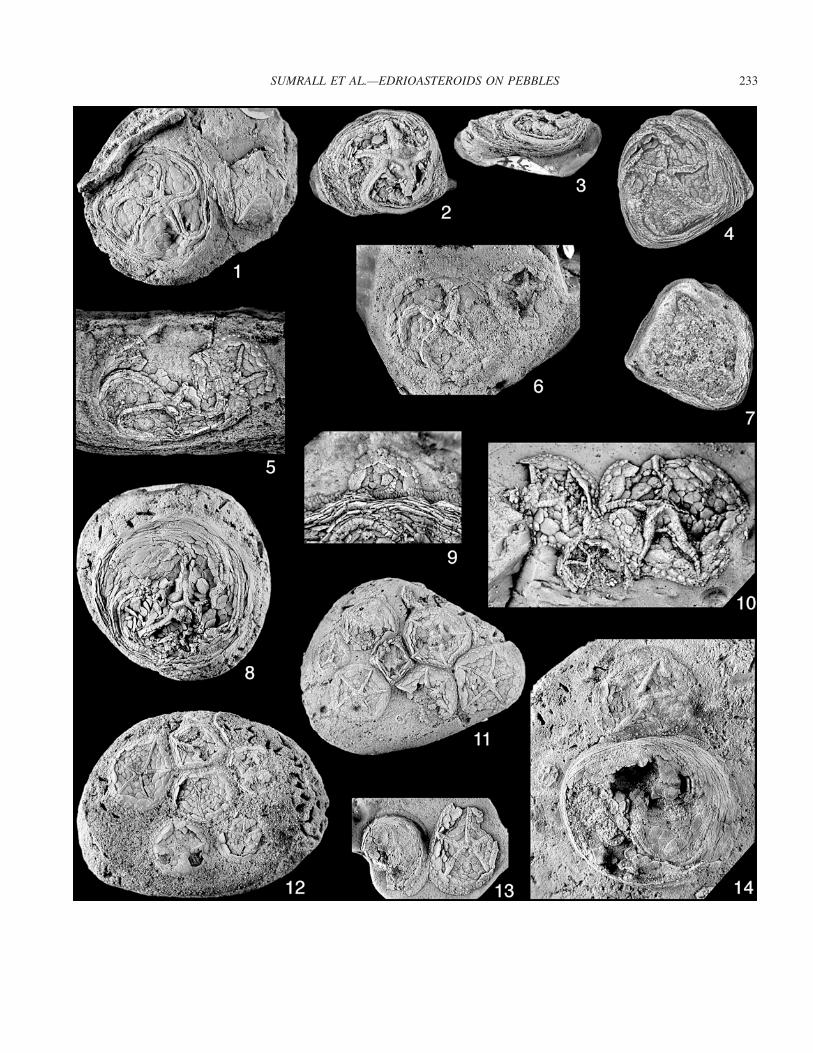
FIGURE 3—Photographs of edrioasteroids on pebbles. All specimens whitened. 1, Large specimen of Ulrichidiscus forbesi n. sp. overgrowing smallspecimen of Neoisorophusella whitesidei n. sp. 1978TX114. Note that the Ulrichidiscus Bassler, 1935 has an abnormal clockwise curvature on theD ambulacrum, �2; 2, 3, small specimen of Ulrichidiscus forbesi 1978TX37 attached to a small pebble, �2; 4, 7, large specimen of Ulrichidiscusforbesi 1978TX295 attached to a pebble smaller than its diameter. Note that the specimen engulfs the pebble, �2; 5, large specimen of Ulrichidiscusforbesi overgrowing small specimen of Neoisorophusella whitesidei 1978TX262. Note that the Neoisorophusella Kammer et al., 1987 is halfovergrown, including the peristome, hydro-gonopore, and periproct, �2; 6, small specimen of Ulrichidiscus forbesi overgrowing small specimenof Neoisorophusella whitesidei 1978TX25, �2. 8, large specimen of Ulrichidiscus forbesi 1978TX222 attached to a small bored pebble. Note thatthe edrioasteroid is overgrowing the Rogerella borings and that some of the peripheral rim plates have fallen into the borings, �2; 9, very largespecimen of Ulrichidiscus forbesi overgrowing very small specimen of Neoisorophusella whitesidei 1978TX64. Note that the Neoisorophusella isnearly half covered by the larger edrioasteroid, �4; 10, medium-sized specimens of Neoisorophusella whitesidei being overgrown by small specimenof Parapostibulla graysoni n. sp., 1978TX269, �4; 11, crowded pebble 1978TX21 encrusted by 10 specimens of Neoisorophusella whitesidei.Note that nowhere do any of the edrioasteroids overgrow adjacent individuals, �1; 12, crowded and bored pebble 1978TX29 encrusted with eightspecimens of Neoisorophusella whitesidei, �1; 13, small pebble with a small specimen of Ulrichidiscus forbesi overgrowing small specimen ofNeoisorophusella whitesidei 1978TX107, �2; 14, pebble with a large specimen of Ulrichidiscus forbesi slightly overgrowing small specimen ofNeoisorophusella whitesidei 1978TX66, �2.
→
FIGURE 2—Photographs of the associated fauna at the Forbes Ranch Locality. All specimens whitened. 1, 3, Infrabasal circlet of Paragassizocrinustarri low cone ecophenotype 1978TX298, top and lateral views, �2; 2, 4, infrabasal circlet of Paragassizocrinus tarri low cone ecophenotype,1978TX299, top and lateral views, �2; 5–7, unusual cup of Paragassizocrinus tarri low cone ecophenotype showing infrabasal sutures, 1978TX300,upper, lower, and lateral CD views, �2; 8, large encrusting bryozoan colony on small pebble, 1978TX119, �1; 9, alloporid coral CladochonusM’Coy, 1847, 1978TX303, �3; 10, star-shaped crinoid columnal, 1978TX304, �3; 11, 12, infrabasal circlet of Paragassizocrinus tarri low coneecophenotype, 1978TX301, top and lateral views. Note that this specimen was eroded and bored by barnacles, �2; 13, surface of a pebble,1978TX56, top and lateral views. Note that this specimen was eroded and bored by barnacles, �2; 13, surface of a pebble, 1978TX56. Note thatthere are numerous tube worms, including several individuals that are exposed where the central edrioasteroid is eroded away, �2; 14, brachialview of a small spiriferid brachiopod, 1978TX307 �3; 15, circular crinoid columnal, 1978TX305, �3; 16, small rugose coral Lophotichiumamoenum Moore and Jeffords, 1945, 1978TX306, �3; 17, small tube worms overgrowing the peripheral rim of Neoisorophusella whitesidei n.sp., 1978TX29A, �15; 18, broken pedicle valve of the brachiopod Composita Brown, 1849, 1978TX308, �3; 19, large pebble 1978TX34 with alimestone upper surface and chert lower surface. Note that the Rogerella barnacle borings are confined to the limestone where they are extremelydense, �1.
lithologies was heavily bored in the limestone and unbored in thechert (Fig. 2.19). Borings were also preferentially found in certainlayers within a given clast (probably softer) and along sharp clastedges. Many pebbles and cobbles are bored on multiple surfaces,suggesting several generations of borings when pebbles over-turned.
Intra- and interspecific competition.⎯Isorophinid edrioaster-oids require a firm to hard substrate for attachment. At the ForbesRanch locality, these substrates are provided by an accumulationof cobbles and pebbles that were exposed on the seafloor. Severalof these cobbles and pebbles have extremely high edrioasteroidpopulation densities ranging up to 4,000 individuals/m2, althoughthe majority of clasts were not colonized. About 64% of the en-crusted clasts have only a single edrioasteroid attached, another27% have 2–4 edrioasteroids, and only 9% have as many as 5–10 edrioasteroids (Fig. 3.11). In some cases, edrioasteroids werecrowded by conspecific adjacent individuals, and the typical cir-cular outline of the edrioasteroids grown unencumbered (Fig. 3.8)becomes polygonal where individuals abut (Fig. 3.11, 3.12). Inthese conspecific cases, no species of edrioasteroid is ever seenwith the margin of the peripheral rim overgrowing its neighbor.In other cases, crowding will occur interspecifically. In these sit-uations, there is a pecking order in which one species will extendits peripheral rim over its neighbor. Eight cases were documentedin which Ulrichidiscus forbesi n. sp. has overgrown Neoisoro-phusella whitesidei n. sp. and no cases are observed where thereverse is true (Fig. 3.1, 3.5, 3.6, 3.9, 3.13, 3.14). In the earliestphases of overgrowing, the Neoisorophusella Kammer et al., 1987appears to resist the overgrowth and both specimens become flat-sided (Fig. 3.13, 3.14), but when the overgrowth is advanced, theUlrichidiscus regains a circular outline as if the Neoisorophusellawere not present (Fig. 3.5, 3.9). Interestingly, overgrown Neoiso-rophusella specimens can have much of their theca covered, in-cluding the peristome, hydro-gonopore, and periproct, yet remain
articulated, suggesting that they could survive for some time withlimited body function. One example of Parapostibulla graysonin. sp. overgrowing two individuals of Neoisorophusella whitesideiwas also documented (Fig. 3.10). In addition, the ParapostibullaSumrall et al., 2000 is much smaller than the Neoisorophusellaspecimens it is overgrowing, suggesting that ability to overgrowanother species is a characteristic of the species rather than itssize or maturity.
There are also cases of edrioasteroids overgrowing other mem-bers of the hard-substrate fauna. Edrioasteroids are noted partiallyovergrowing rugose corals, ramose bryozoans, tubeworms, andborings left by acrothoracic barnacles (Figs. 2.13, 3.8). There isno clear evidence for competition or lack thereof and thereforeno way to determine with certainty whether these faunal elementsare older than or coeval with the edrioasteroids or both. Somevery small tubeworms are seen overgrowing plates of Neoisoro-phusella whitesidei (Fig. 2.17). The taphonomically fragile natureof the edrioasteroid theca strongly suggests that these worms wereliving as an epibiont upon the edrioasteroids while they werealive. This relationship has not been observed on any other spe-cies of edrioasteroid.
POPULATION DYNAMICS AND ONTOGENY
A total of 451 edrioasteroid specimens belonging to four spe-cies were collected. Of these 53.7% were Neoisorophusella,37.9% were Ulrichidiscus, 3.3% were Postibulla, 2.9% were Par-apostibulla, 1.1% were undifferentiated postibullinid, and 1.1%were unidentified (Fig. 4). Total postibullinids comprise 7.3% ofthe edrioasteroid fauna.
Neoisorophusella.⎯Neoisorophusella is known from 242 spec-imens that range in thecal diameter from 1 mm to 19 mm withan average of 11.5 mm (n � 200, with 42 unmeasurable speci-mens). The distribution is left-skewed with a standard deviationof 3.8, and a g1 of �0.83, indicating a dominance of mature

232 JOURNAL OF PALEONTOLOGY, V. 80, NO. 2, 2006
233SUMRALL ET AL.—EDRIOASTEROIDS ON PEBBLES
234 JOURNAL OF PALEONTOLOGY, V. 80, NO. 2, 2006
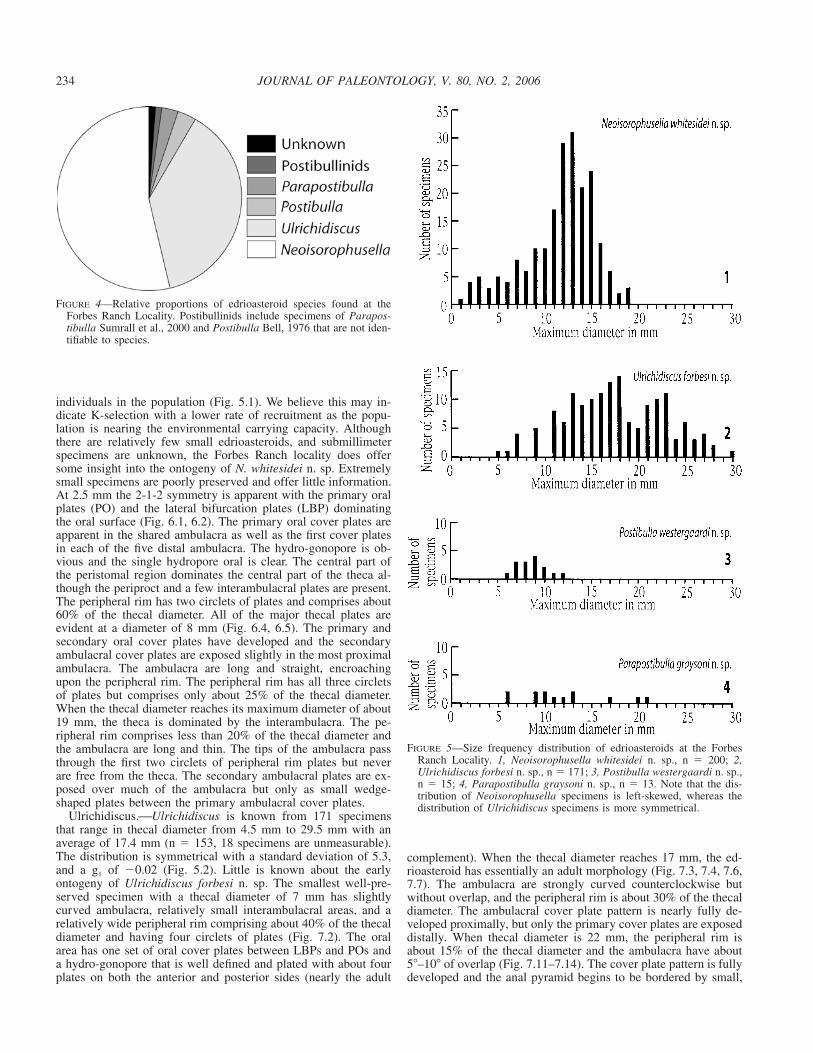
FIGURE 4—Relative proportions of edrioasteroid species found at theForbes Ranch Locality. Postibullinids include specimens of Parapos-tibulla Sumrall et al., 2000 and Postibulla Bell, 1976 that are not iden-tifiable to species.
FIGURE 5—Size frequency distribution of edrioasteroids at the ForbesRanch Locality. 1, Neoisorophusella whitesidei n. sp., n � 200; 2,Ulrichidiscus forbesi n. sp., n � 171; 3, Postibulla westergaardi n. sp.,n � 15; 4, Parapostibulla graysoni n. sp., n � 13. Note that the dis-tribution of Neoisorophusella specimens is left-skewed, whereas thedistribution of Ulrichidiscus specimens is more symmetrical.
individuals in the population (Fig. 5.1). We believe this may in-dicate K-selection with a lower rate of recruitment as the popu-lation is nearing the environmental carrying capacity. Althoughthere are relatively few small edrioasteroids, and submillimeterspecimens are unknown, the Forbes Ranch locality does offersome insight into the ontogeny of N. whitesidei n. sp. Extremelysmall specimens are poorly preserved and offer little information.At 2.5 mm the 2-1-2 symmetry is apparent with the primary oralplates (PO) and the lateral bifurcation plates (LBP) dominatingthe oral surface (Fig. 6.1, 6.2). The primary oral cover plates areapparent in the shared ambulacra as well as the first cover platesin each of the five distal ambulacra. The hydro-gonopore is ob-vious and the single hydropore oral is clear. The central part ofthe peristomal region dominates the central part of the theca al-though the periproct and a few interambulacral plates are present.The peripheral rim has two circlets of plates and comprises about60% of the thecal diameter. All of the major thecal plates areevident at a diameter of 8 mm (Fig. 6.4, 6.5). The primary andsecondary oral cover plates have developed and the secondaryambulacral cover plates are exposed slightly in the most proximalambulacra. The ambulacra are long and straight, encroachingupon the peripheral rim. The peripheral rim has all three circletsof plates but comprises only about 25% of the thecal diameter.When the thecal diameter reaches its maximum diameter of about19 mm, the theca is dominated by the interambulacra. The pe-ripheral rim comprises less than 20% of the thecal diameter andthe ambulacra are long and thin. The tips of the ambulacra passthrough the first two circlets of peripheral rim plates but neverare free from the theca. The secondary ambulacral plates are ex-posed over much of the ambulacra but only as small wedge-shaped plates between the primary ambulacral cover plates.
Ulrichidiscus.⎯Ulrichidiscus is known from 171 specimensthat range in thecal diameter from 4.5 mm to 29.5 mm with anaverage of 17.4 mm (n � 153, 18 specimens are unmeasurable).The distribution is symmetrical with a standard deviation of 5.3,and a g1 of �0.02 (Fig. 5.2). Little is known about the earlyontogeny of Ulrichidiscus forbesi n. sp. The smallest well-pre-served specimen with a thecal diameter of 7 mm has slightlycurved ambulacra, relatively small interambulacral areas, and arelatively wide peripheral rim comprising about 40% of the thecaldiameter and having four circlets of plates (Fig. 7.2). The oralarea has one set of oral cover plates between LBPs and POs anda hydro-gonopore that is well defined and plated with about fourplates on both the anterior and posterior sides (nearly the adult
complement). When the thecal diameter reaches 17 mm, the ed-rioasteroid has essentially an adult morphology (Fig. 7.3, 7.4, 7.6,7.7). The ambulacra are strongly curved counterclockwise butwithout overlap, and the peripheral rim is about 30% of the thecaldiameter. The ambulacral cover plate pattern is nearly fully de-veloped proximally, but only the primary cover plates are exposeddistally. When thecal diameter is 22 mm, the peripheral rim isabout 15% of the thecal diameter and the ambulacra have about5�–10� of overlap (Fig. 7.11–7.14). The cover plate pattern is fullydeveloped and the anal pyramid begins to be bordered by small,

235SUMRALL ET AL.—EDRIOASTEROIDS ON PEBBLES
differentiated interambulacral plates (Fig. 7.19). The largest spec-imens (Fig. 7.21) are essentially the same morphology as thosethat are 22 mm, but with about 30� of ambulacral overlap.
Postibulla.⎯Postibulla westergaardi n. sp. is known from 15specimens that range in thecal diameter from 5.5 mm to 12 mmwith an average of 8.5 mm (n � 15, five additional specimensare unmeasurable or uncertain whether they belong to Postibullaor Parapostibulla). The distribution is right-skewed with a stan-dard deviation of 1.7, and a g1 of 0.35 (Fig. 5.3). Given the smallsample size, little is inferred about this size-frequency distribu-tion. Little ontogenetic information is available except that small-er specimens have shorter ambulacra and more pronounced pe-ripheral rims than large specimens (compare Fig. 8.1 to Fig. 8.10).
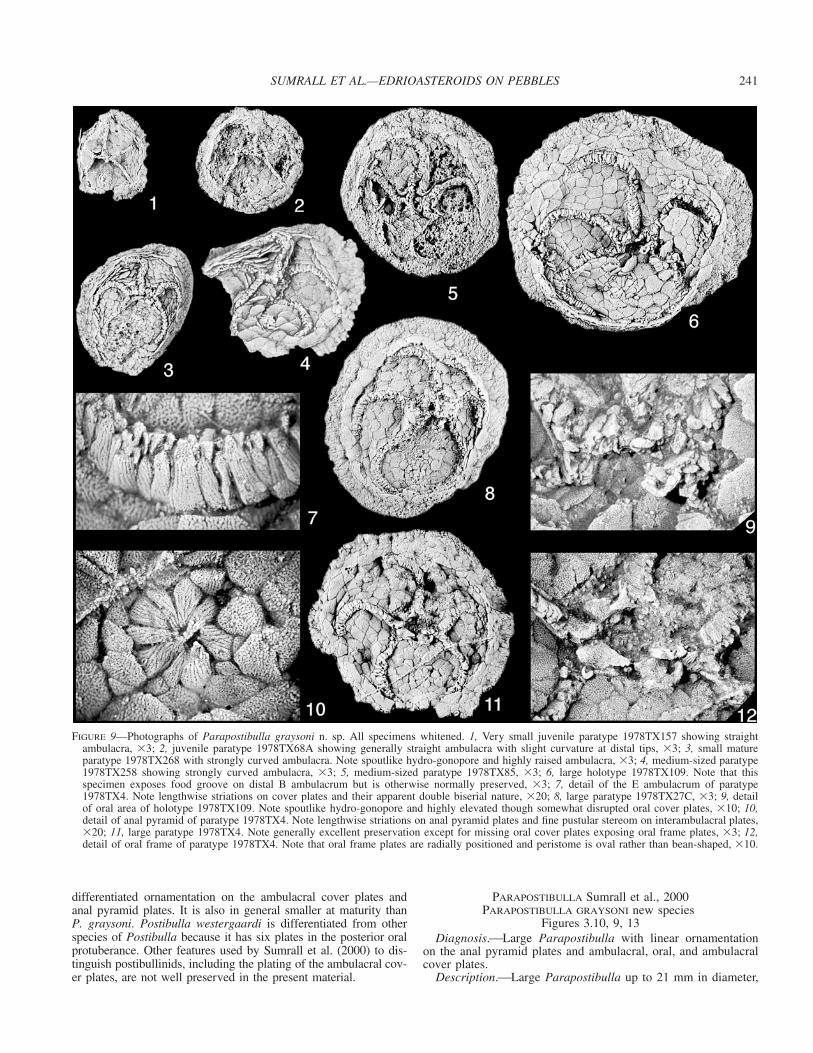
Parapostibulla.⎯Parapostibulla graysoni n. sp. is known from13 specimens that range in thecal diameter from 5.5 mm to 21mm with an average of 11.8 mm (n � 13, five additional speci-mens are unmeasurable or uncertain whether they belong to Pos-tibulla or Parapostibulla). The distribution is right-skewed witha standard deviation of 4.9, and a g1 of 0.66 (Fig. 5.4). Given thesmall sample size, little is inferred about this size-frequency dis-tribution. Little is known of the early ontogeny. Juvenile speci-mens less than 9 mm have straight ambulacra and can easily beconfused with Neoisorophusella whitesidei (Fig. 9.1, 9.2). Thesecan be differentiated by the protuberant nature of the hydroporeand the unusual ornamentation on the highly elevated coverplates. These small individuals have a wide peripheral rim com-prising about 30% of the thecal diameter. When the thecal di-ameter reaches 12 mm, the ambulacra are curved most noticeablyin the distal portions but without ambulacral overlap (Fig. 9.3).With a thecal diameter of 15 mm, the theca essentially has anadult morphology and an ambulacra curve of about 40� (Fig. 9.4).The peripheral rim remains wide, covering about 25% of the the-cal diameter. Continued growth makes the interambulacral areasslightly more prominent, the peripheral rim slightly less promi-nent, and the ambulacra slightly more curved (Fig. 9.5, 9.6).
Developmental anomalies.⎯Only two specimens in the collec-tion have a developmental anomaly. A single specimen of Ulri-chidiscus forbesi 1978TX114A is normal in every respect exceptthat the D ambulacrum curves clockwise instead of the normalcounterclockwise direction (Fig. 3.1). The tip of the D ambula-crum extends distally to the counterclockwise E ambulacrum,which appears to be foreshortened because its path is blocked.Interestingly, this specimen also overgrew a medium-sized indi-vidual of Neoisorophusella whitesidei, but this is interpreted aspurely coincidental. The second anomalous specimen is a verypoorly preserved unfigured specimen of Neoisorophusella whi-tesidei 1978TX56B that has six ambulacra resulting from ananomalous bifurcation of the A ambulacrum. The specimen isotherwise normal.
TAPHONOMY
Edrioasteroids have a very fragile and easily disarticulated skel-eton composed of several hundred to several thousand plates. Aswith most echinoderms, the thecal plates are articulated with softtissue that easily degrades within a few hours to days after death.Unless thecal burial is immediate or nearly so with death, theskeleton rapidly disaggregates into sand-sized particles. Althoughthese plates are easily recognizable as edrioasteroid, they are eas-ily overlooked and of very little use at lower taxonomic levels(Sumrall and Parsley, 2003). Isorophid edrioasteroids, unlikemany more familiar echinoderms, have a theca that is imbricatelyplated, giving the organism great flexibility when alive. Conse-quently, there are no skeletally fused thecal elements that holdtogether as modules after death except for the peripheral rim(Brett et al., 1997). During postmortem thecal collapse and burialcompaction, the dorsal plating of the oral surface is pressed
against the substrate as the viscera degrade and the specimen isflattened (Fig. 3.8, 3.11). The relatively high relief of the ambu-lacra, anal pyramid, and peripheral rim results from the greaterthickness of these plates over the low interambulacral plating (Fig.6.4, 6.12).
Several features are unusual about the thecal collapse amongthe edrioasteroids at the Forbes Ranch locality. Edrioasteroids at-tached to smooth, unbored pebbles undergo thecal collapse asdescribed above (Fig. 3.10). Where edrioasteroids are attached tobored pebbles, there is no underlying support for the theca overthe borings during compaction. In many cases, this results in anirregular thecal surface reflecting the uneven underlying surface(Figs. 6.9, 7.16). In some cases, borings appear to pass throughthe edrioasteroids (Figs. 6.9, 6.13, 7.13). Under careful exami-nation, these are places where thecal plates have collapsed intothe borings and are in some cases broken. They lack crosscuttingof the thecal plates that would be expected if truly bored (seeexamples of acrothoracic borings into other calcite fossils in Rod-da and Fisher, 1962).
The extremely tall ambulacral cover plates of the postibullinidsare often preserved slumped over to one side (Figs. 8.14, 9.7),adding to the difficulty of their interpretation. Ulrichidiscus andNeoisorophusella typically have well-preserved cover plates thatform a low dome over the food groove, but in a few cases com-paction has pressed the ambulacral cover plates into the foodgroove, giving the ambulacra a concave look (Figs. 6.11, 7.21).In no cases are the cover plates disarticulated from the floor platesbefore burial, suggesting rapid burial of the fauna.
Ulrichidiscus forbesi n. sp., unlike the other edrioasteroids inthe fauna, has a more fully developed pedunculate zone (the cir-clets of plates between the oral surface and the peripheral rim)than other edrioasteroids (Fig. 7.5, 7.8, 7.15). It is variably ex-pressed, probably because of ecophenotypic variation and whetheror not the specimen was fully retracted at the time of burial. Thisis most evident in cases where individuals are attached to pebblesthat are smaller than or equal to the diameter of the edrioasteroid(Fig. 3.1–3.4, 3.7). In some cases, the pebble is effectively en-gulfed by the edrioasteroid. In other cases lateral slumping of thepedunculate zone and the oral surface results in these plates lyingover the peripheral rim (Figs. 3.8, 7.13, 7.15) and in a few casesover abutting specimens, giving them the appearance of over-growing.
Preservation of the stereom microstructure in many specimensis exquisite. Although the inner cores of the plates are perminer-alized by calcite overgrowths of the microporous stereom, theouter surfaces of many plates are still porous (Figs. 2.17, 9.7,9.10).
SYSTEMATIC PALEONTOLOGY
CRINOIDEA Miller, 1821CLADIDA Moore and Laudon, 1943
AMPELOCRINIDAE Kirk, 1942PARAGASSIZOCRININAE Strimple and Watkins, 1969
PARAGASSIZOCRINUS Moore and Plummer, 1940PARAGASSIZOCRINUS TARRI Strimple, 1938
Figure 2.1–2.7, 2.11, 2.12
Material examined.⎯Figured specimens of P. tarri low coneecophenotype 1978TX298–1978TX301, and nearly 100 unsortedinfrabasal circlets in the collections at the Texas Memorial Mu-seum.
Discussion.⎯Numerous infrabasal circlets of the low-cone eco-phenotype sensu Ettensohn (1980) associated with disarticulatedbasals, radials, and arm plates were recovered from the ForbesRanch locality of unnamed unit 1 of the Golf Course Formation.Only one complete cup was recovered (Fig. 2.5–2.7) although it
236 JOURNAL OF PALEONTOLOGY, V. 80, NO. 2, 2006
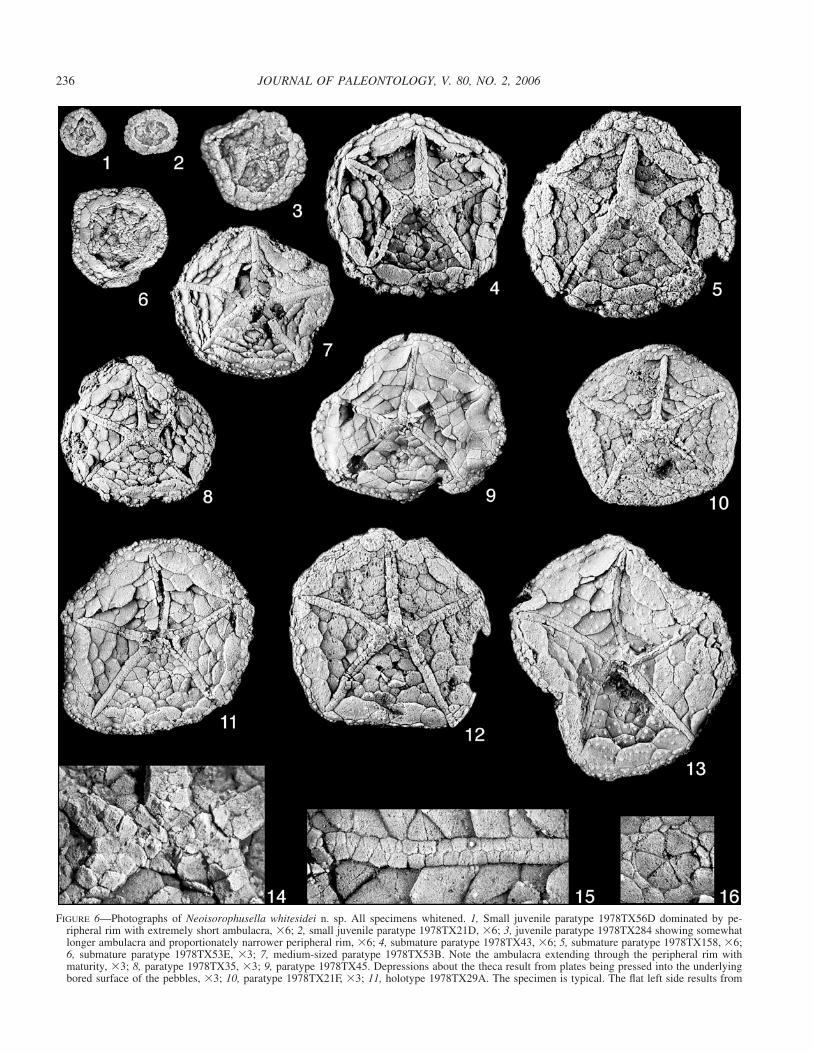
FIGURE 6—Photographs of Neoisorophusella whitesidei n. sp. All specimens whitened. 1, Small juvenile paratype 1978TX56D dominated by pe-ripheral rim with extremely short ambulacra, �6; 2, small juvenile paratype 1978TX21D, �6; 3, juvenile paratype 1978TX284 showing somewhatlonger ambulacra and proportionately narrower peripheral rim, �6; 4, submature paratype 1978TX43, �6; 5, submature paratype 1978TX158, �6;6, submature paratype 1978TX53E, �3; 7, medium-sized paratype 1978TX53B. Note the ambulacra extending through the peripheral rim withmaturity, �3; 8, paratype 1978TX35, �3; 9, paratype 1978TX45. Depressions about the theca result from plates being pressed into the underlyingbored surface of the pebbles, �3; 10, paratype 1978TX21F, �3; 11, holotype 1978TX29A. The specimen is typical. The flat left side results from

237SUMRALL ET AL.—EDRIOASTEROIDS ON PEBBLES
←
crowding by an adjacent individual, �3; 12, paratype 1978TX21A, �3; 13, paratype 1978TX89. Note the large nodes covering the theca, �3;14, detail of oral area of paratype 1978TX21A. Note the relatively large primary oral plates and the undifferentiated lateral bifurcation plates,�10; 15, detail of the D ambulacrum of holotype 1978TX29A. Note the double biserial pattern of ambulacral cover plates proximally that gradeinto a single biseries distally (right), �10; 16, detail of periproct of paratype 1978TX29B. Note the relatively few anal pyramid plates (this onehas five; four is also common) and its circular outline, �10.
is developmentally anomalous. This specimen shows weak butunambiguous lack of fusion of the infrabasals—a condition un-known in other mature specimens of Paragassizocrinus. Accord-ing to Ettensohn (1980), Paragassizocrinus ontogenetically shiftsfrom a juvenile with a small stem and separate infrabasals to amature individual lacking a stem and with fused infrabasals. Thisspecimen is similar to a condition seen in the similar, but distantlyrelated Agassizocrinus Owen and Shumard, 1851, documented byEttensohn (1975, personal commun., 2004).
EDRIOASTEROIDEA Billings, 1858ISOROPHIDA Bell, 1976ISOROPHINA Bell, 1976
AGELACRINITIDAE Chapman, 1860NEOISOROPHUSELLA Kammer et al., 1987
Type species.⎯Neoisorophusella lanei Kammer et al., 1987.Discussion.⎯Kammer et al. (1987) described Neoisorophusella
based on two species, N. lanei from the Upper Mississippian Kin-kaid Formation, and N. berryi Kammer et al., 1987 from the sim-ilar-aged Greenbrier Formation. They placed Neoisorophusellainto Isorophinae based on a supposed close relationship with Iso-rophusella Bassler, 1935, an Ordovician edrioasteroid withstraight ambulacra following a taxonomic revision of Bell (1976)by Smith (1985). Sumrall (1993) followed the Bell (1976) clas-sification, placing Neoisorophusella in Isorophidae rather than inIsorophinae. Based on new evidence including the lack of intra-thecal extensions of the ambulacral cover plates and complex oralcover plates, Neoisorophusella is here considered to be a paedo-morphic agelacrinitid, s. Bell (1976).
NEOISOROPHUSELLA WHITESIDEI new speciesFigures 2.13, 2.17, 3.10–3.14, 6, 10
Diagnosis.⎯Neoisorophusella with small, evenly spaced nodeson interambulacra and peripheral rim; ambulacra extend throughnotches in peripheral rim; one primary oral cover plate betweenPOs and LBPs; peripheral rim with three circlets of plates.
Description.⎯Large Neoisorophusella up to 19 mm in diam-eter, circular in outline if growth unencumbered. Oral area trans-versely elongate with ambulacra arranged into 2-1-2 symmetry;perradial suture over peristome slightly concave posteriorly; POsdifferentiated from other cover plates, LBPs same size as coverplates (Fig. 10.1); one primary oral cover plate and secondaryplate pair separates POs from LBPs on both sides (Figs. 6.14,10.1); posterior oral cover plates similar to anterior plates; hydro-gonopore semi-integrated into oral rise, with one posterior hydro-pore oral, aperture slitlike (Figs. 6.14, 10.1). Ambulacra linear,narrow, extremely long, pass through notch in peripheral rim inlarge specimens (Fig. 6.7–6.13), slight taper throughout with blunttermination, plated with cover plates and underlying floor plates;cover plates double biserial, primary cover plates extremely wideand boxlike, secondary cover plates exposed only in large spec-imens, wedge-shaped between primary cover plates along nearlylinear perradial suture (Figs. 6.15, 10.2); floor plates with U-shaped trough for food groove, poorly constrained; periproct cov-ered by large anal pyramid centered in CD interambulacral areaalong midline, plated with four or five lathe-shaped plates (Fig.6.16). Interambulacral areas plated with large, thin, squamose, andhighly imbricate interambulacral plates; plates have evenly but
widely spaced nodes along proximal edges; large specimens with10–12 interambulacral plates per interambulacrum; pedunculatezone absent; peripheral rim wide, with three irregular circlets ofhighly imbricate plates, most proximal circlet extremely wide, upto 70% of rim width, more equant than typical edrioasteroid; mid-dle and distal circlets progressively smaller distally, both withequant plates (Fig. 6.13); peripheral rim is typically notched inproximal circlet for distal ambulacra tips; all peripheral rim plateswith small evenly spaced nodes except occasionally in distal cir-clet.
Etymology.⎯Named for Joseph Whiteside, a former student atBaylor University, who originally described the Forbes Ranch ed-rioasteroid occurrence for his bachelor’s thesis (Whiteside, 1987).
Types.⎯The type series for Neoisorophusella whitesidei in-cludes holotype 1978TX29A and paratypes 1978TX21A, 21D,21F, 29B, 35, 43, 45, 53B, 53E, 56D, 89, 158, and 284. Additionalspecimens found on numerous clasts of the 1978TX number seriesat the Texas Memorial Museum, Austin.
Occurrence.⎯Neoisorophusella whitesidei is known only fromunnamed unit 1 of the Golf Course Formation, Dornick HillsGroup, Lower Pennsylvanian (Morrowan), at Forbes Ranch, northof Marietta, southern Oklahoma.
Discussion.⎯Neoisorophusella whitesidei is the largest speciesof Neoisorophusella known. It differs from other species assignedto the genus by the presence of three circlets of peripheral rimplates, notched peripheral rim plates accommodating the distalambulacra, a relatively narrow oral area, and the presence of smallnodes that cover the interambulacral and peripheral rim plates.Neoisorophusella lanei has smaller proximal peripheral rim platesand small nodes on the adradial tips of the ambulacral coverplates. Neoisorophusella berryi has relatively smaller and morenumerous interambulacral plates. Although the differences be-tween N. whitesidei, other members of Neiosorophusella, andprobably species assigned to Yukutidiscus are small, these specieshave not been synonymized, pending a thorough phylogeneticanalysis of this clade that is beyond the scope of this manuscript.
ULRICHIDISCUS Bassler, 1935ULRICHIDISCUS FORBESI new species
Figures 3.1–3.9, 3.13, 3.14, 7, 11Diagnosis.⎯Ulrichidiscus lacking spines, with six-plate pattern
of ambulacral cover plates that are poorly developed at maturity;ambulacra with much overlap in mature specimens.
Description.⎯Large Ulrichidiscus up to 29.5 mm in diameter,theca circular in outline; oral area transversely elongate with am-bulacra arranged with 2-1-2 symmetry (Figs. 7.20, 11.1); perradialsuture concave posteriorly. LBPs and POs long and extremelynarrow, separated by three sets of primary and secondary coverplates anteriorly (Figs. 7.20, 11.1); posterior oral cover plates sim-ilar to anterior plates except somewhat foreshortened, allowingroom for the hydro-gonopore; hydro-gonopore highly elevated,separated from oral rise, located in proximal right CD interam-bulacrum, with five hydropore oral plates proximally, five platesdistally, and poorly developed left hydropore oral plate, apertureslitlike (Fig. 7.20); periproct large, located in distal CD interam-bulacrum, with well-developed low conical anal pyramid of innerand outer anal pyramid plates, irregular with 8–11 plates per cir-clet, surrounded by many smaller imbricate plates (Fig. 7.19,
238 JOURNAL OF PALEONTOLOGY, V. 80, NO. 2, 2006

239SUMRALL ET AL.—EDRIOASTEROIDS ON PEBBLES
←
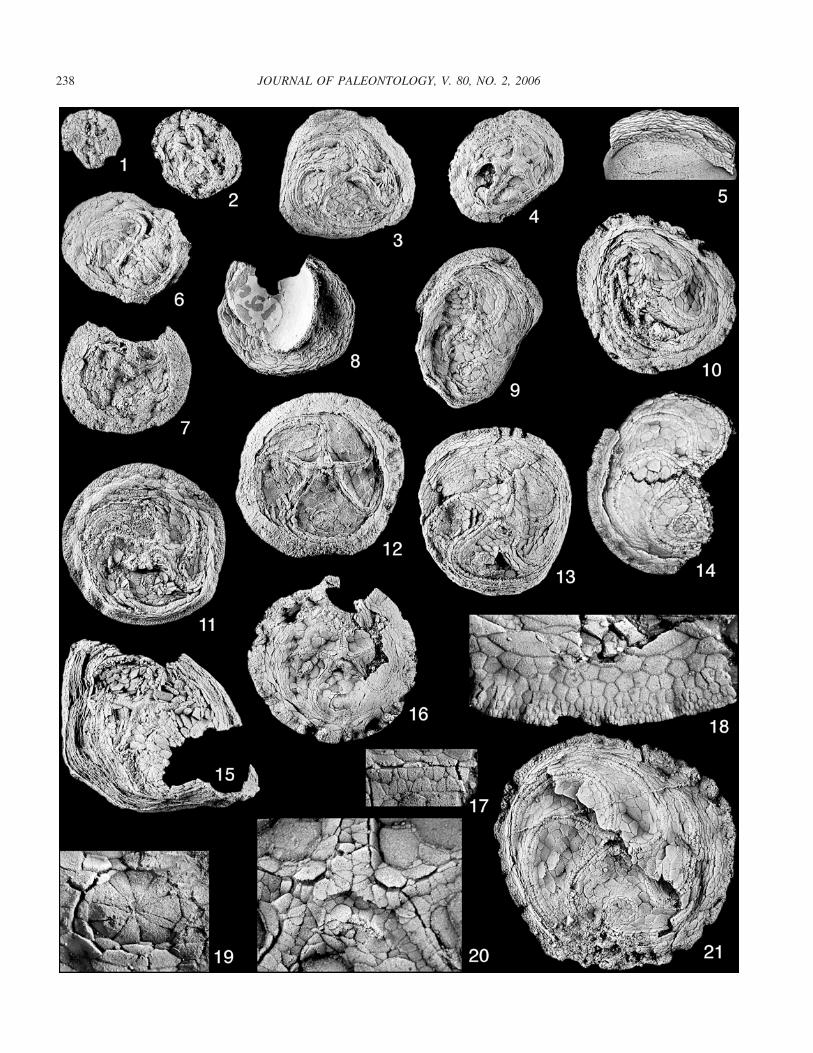
FIGURE 7—Photographs of Ulrichidiscus forbesi n. sp. All specimens whitened. 1, Small paratype 1978TX20 with slight curvature of the ambulacra,�2; 2, small paratype 1978TX107A showing slightly more ambulacral curvature, �2; 3, medium-sized paratype 1978TX73 showing long curvedambulacra, �2; 4, small paratype 1978TX206, �2; 5, fragmentary paratype 1978TX266 showing peripheral rim and slightly extended pedunculatezone, �2; 6, medium-sized paratype 1978TX100, �2; 7, medium-sized paratype 1978TX95 with relatively wide peripheral rim, �2; 8, medium-sized paratype 1978TX159 attached to brachiopod fragment. Note that small attachment surface resulted in small peripheral rim and multiplatedpedunculate zone, �2; 9, large paratype 1978TX164. The unusual shape results from specimen extending over the edge of a pebble, �2; 10, largeparatype 1978TX50. Note long ambulacra with about 10� of ambulacral overlap, �2; 11, large paratype 1978TX7 showing well-developed peripheralrim and anal pyramid, �2; 12, large paratype 1978TX14, �2; 13, large holotype 1978TX205. Note slight disruption to the right of periproct, butotherwise well preserved, �2; 14, large paratype 1978TX296 missing most of peripheral rim and BC interambulacral area, �2; 15, very largeparatype 1978TX62. Note extreme development of pedunculate zone bearing about 24 plate circlets, �2; 16, well-preserved but incomplete largeparatype 1978TX63, �2; 17, detail of proximal B ambulacrum of paratype 1978TX63. Note cycles of three primary cover plates and poorlyexpressed secondary cover plates, �10; 18, detail of peripheral rim along B radius of paratype 1978TX63. Note progressively smaller and moreradially elongate plates distally, �8; 19, detail of periproct of paratype 1978TX63; �10; 20, detail of oral area of paratype 1978TX63. Noterelatively undifferentiated primary oral plates, lateral bifurcation plates, and plating of the hydro-gonopore, �10; 21, gerontic paratype 1978TX113.Note extremely long ambulacra, relatively narrow peripheral rim, and well-defined periproct, �2.
7.21). Ambulacra with 5–0 counterclockwise arrangement, evenlycurved throughout length with little taper and blunt termination,mature specimens with up to 30� overlap in 29 mm specimen, 20mm specimen without overlap; ambulacral cover plates in six-plate cycles with three primary and three secondary cover platesper cycle (Fig. 11.2), secondary cover plates extremely small andcover plate pattern only fully developed proximally in largestspecimens; perradial zigzag poorly developed, cycles from op-posite sides of perradial suture mirror image offset by half cycle(Fig. 11.2); floor plates with U-shaped food groove, otherwisepoorly constrained. Interambulacral areas plated with thin semi-imbricate, squamose to tessellate interambulacral plates. Maturespecimens with 25–30 plates per interambulacral area; peduncu-late zone variably developed in material, with 5–24 circlets ofplates at maturity (Fig. 7.15); plates squamose, highly imbricate,forming irregular circlets; peripheral rim extremely narrow (Fig.7.21), with five irregular circlets of highly imbricate plates; mostproximal circlet not differentiated, transversely elongate, moredistal circlets gradually transition from transversely elongate toradially elongate (Fig. 7.18), bottom surface unknown.
Etymology.⎯Named for the late Joe Forbes, who owned theranch locality.
Types.⎯The type series for Ulrichidiscus forbesi includes ho-lotype 1978TX205 and paratypes 1978TX7, 14, 20, 50, 62, 63,73, 95, 100, 107A, 113, 114A, 159, 164, 206, 266, and 296.Additional specimens found on numerous clasts of the 1978TXnumber series at the Texas Memorial Museum, Austin.
Occurrence.⎯Ulrichidiscus forbesi is known only from un-named unit 1 of the Golf Course Formation, Dornick Hills Group,Lower Pennsylvanian (Morrowan), at Forbes Ranch, north ofMarietta, southern Oklahoma.
Discussion.⎯Ulrichidiscus forbesi is distinguished from otherspecies of Ulrichidiscus by its six-plate pattern of ambulacral cov-er plates that have poorly exposed secondary cover plates, and itslong, strongly overlapping ambulacra at maturity. Ulrichidiscuspulaskiensis Miller and Gurley, 1894 has much more ambulacraloverlap, much greater exposure of the secondary ambulacral cov-er plates, and a seven-plate pattern of ambulacral cover plates.Ulrichidiscus spinosus Sumrall, 2001 has a six-plate pattern ofambulacral cover plates, but has long spines on the primary coverplates and the outer anal pyramid plates.
POSTIBULLINAE Sumrall et al., 2000POSTIBULLA Bell, 1976
POSTIBULLA WESTERGAARDI new speciesFigures 8, 12
Diagnosis.⎯Postibulla with extremely large posterior oral pro-tuberance formed from six paired plates; hydropore formed fromthree hydropore orals.
Description.⎯Medium-sized Postibulla up to 12 mm in di-ameter, circular in outline; all plates with fine laminar stereom onouter surface; oral area relatively wide with ambulacra arrangedin 2–1–2 symmetry (Figs. 8.12, 8.13, 12); primary oral coverplates and lateral bifurcation plates undifferentiated; oral coverplates with three sets of primary and secondary cover plates be-tween POs and LBPs (Fig. 12); posterior oral cover plates similarto anterior cover plates, foreshortened on right to allow room forhydro-gonopore; perradial suture over peristome chevron-shaped,concave toward posterior; posterior oral protuberance very large,oriented along midline, formed proximally from two enlargedposterior oral cover plates and posteriorly from six enlarged oralprotuberance plates (Figs. 8.12, 8.13, 12); hydropore offset fromoral rise, in proximal right CD interradius, formed from threeplates, with slitlike aperture (Fig. 12). Ambulacra with 3-2 cur-vature with slight counter flexure prior to main curvature in ma-ture specimens, curved throughout length without ambulacraloverlap, taper slightly with blunt termination without overlap;plated with high ambulacral cover plates over floor plates; coverplates apparently in complex pattern, but details unclear in presentmaterial (Fig. 8.14); floor plates poorly constrained; periproct lo-cated in center of CD interambulacrum along midline, small, cir-cular in outline; covered by inner and outer circlet of lathe-shapedplates typically eight in each circlet (Fig. 8.15). Interambulacraplated with thin, highly imbricate, squamose plates; plates rathersmall with about 20 plates per interambulacrum in mature spec-imens, but number poorly constrained. Pedunculate zone poorlydeveloped with typically about two to three circlets of highlyimbricate plates between distal ambulacra and peripheral rim; pe-ripheral rim with four irregular circlets of plates (Fig. 8.10); mostproximal circlet with transversely elongate plates, much widerthan other circlets; middle two circlets with equant plates pro-gressively smaller distally; distalmost circlet with extremely smallplates, radially elongate; bottom surface unknown.
Etymology.⎯Named for Edwin Westergaard, a former studentat Baylor University, who found the first edrioasteroid specimensat the Forbes Ranch locality.
Types.⎯The type series for Postibulla westergaardi includesholotype 1978TX183 and paratypes 1978TX27A, 40, 105, 108,148, 169, 191, 255, 288, and 291. Additional specimens foundon a few other clasts of the 1978TX number series at the TexasMemorial Museum, Austin.
Occurrence.⎯Postibulla westergaardi is known only from un-named unit 1 of the Golf Course Formation, Dornick Hills Group,Lower Pennsylvanian (Morrowan), at Forbes Ranch, north ofMarietta, southern Oklahoma.
Discussion.⎯Although similar to the co-occurring Paraposti-bulla graysoni n. sp., Postibulla westergaardi can be easily distin-guished by the presence of a posterior oral protuberance, the gen-erally less highly elevated ambulacral cover plates, and the lack of

240 JOURNAL OF PALEONTOLOGY, V. 80, NO. 2, 2006
FIGURE 8—Photographs of Postibulla westergaardi n. sp. All specimens whitened. 1, Small paratype 1978TX27A. Note that unlike Parapostibullagraysoni n. sp., Postibulla westergaardi has curved ambulacra while still very immature, �5; 2, incomplete paratype 1978TX255, �5; 3, advancedjuvenile paratype 1978TX40 showing relatively wide peripheral rim and small interambulacra, �5; 4, medium-sized paratype 1978TX169 showingwell-developed posterior oral protuberance, �5; 5, incomplete medium-sized paratype 1978TX291, �5; 6, medium-sized paratype 1978TX108,�5; 7, medium-sized paratype 1978TX148, �5; 8, medium-sized paratype 1978TX105, �5; 9, poorly preserved large paratype 1978TX191. Notelong curved ambulacra, posterior oral protuberance, and relatively narrow peripheral rim, �5; 10, large holotype 1978TX183. Specimen is mostlycomplete except for portions of peripheral rim especially in CD interradius, �5; 11, incomplete large paratype 1978TX288, �5; 12, detail of oralarea of paratype 1978TX169. Note plating of oral cover plates, primary oral plates, and lateral bifurcation plates, �10; 13, detail of oral area ofparatype 1978TX288. Note plating of posterior oral protuberance and hydro-gonopore, �10; 14, detail of D ambulacral cover plates of paratype1978TX288, �20; 15, detail of anal pyramid of paratype 1978TX169, �20.
241SUMRALL ET AL.—EDRIOASTEROIDS ON PEBBLES
FIGURE 9—Photographs of Parapostibulla graysoni n. sp. All specimens whitened. 1, Very small juvenile paratype 1978TX157 showing straightambulacra, �3; 2, juvenile paratype 1978TX68A showing generally straight ambulacra with slight curvature at distal tips, �3; 3, small matureparatype 1978TX268 with strongly curved ambulacra. Note spoutlike hydro-gonopore and highly raised ambulacra, �3; 4, medium-sized paratype1978TX258 showing strongly curved ambulacra, �3; 5, medium-sized paratype 1978TX85, �3; 6, large holotype 1978TX109. Note that thisspecimen exposes food groove on distal B ambulacrum but is otherwise normally preserved, �3; 7, detail of the E ambulacrum of paratype1978TX4. Note lengthwise striations on cover plates and their apparent double biserial nature, �20; 8, large paratype 1978TX27C, �3; 9, detailof oral area of holotype 1978TX109. Note spoutlike hydro-gonopore and highly elevated though somewhat disrupted oral cover plates, �10; 10,detail of anal pyramid of paratype 1978TX4. Note lengthwise striations on anal pyramid plates and fine pustular stereom on interambulacral plates,�20; 11, large paratype 1978TX4. Note generally excellent preservation except for missing oral cover plates exposing oral frame plates, �3; 12,detail of oral frame of paratype 1978TX4. Note that oral frame plates are radially positioned and peristome is oval rather than bean-shaped, �10.
differentiated ornamentation on the ambulacral cover plates andanal pyramid plates. It is also in general smaller at maturity thanP. graysoni. Postibulla westergaardi is differentiated from otherspecies of Postibulla because it has six plates in the posterior oralprotuberance. Other features used by Sumrall et al. (2000) to dis-tinguish postibullinids, including the plating of the ambulacral cov-er plates, are not well preserved in the present material.
PARAPOSTIBULLA Sumrall et al., 2000PARAPOSTIBULLA GRAYSONI new species
Figures 3.10, 9, 13Diagnosis.⎯Large Parapostibulla with linear ornamentation
on the anal pyramid plates and ambulacral, oral, and ambulacralcover plates.
Description.⎯Large Parapostibulla up to 21 mm in diameter,

242 JOURNAL OF PALEONTOLOGY, V. 80, NO. 2, 2006
FIGURE 10—Camera lucida drawings of oral area and ambulacral coverplates of Neoisorophusella whitesidei n. sp. 1, Oral area of paratype1978TX21A. Stippled plates are lateral bifurcation plates (LBP) andprimary orals (PO). Note that hydro-gonopore is bordered by singlehydropore oral (HO); 2, proximal D ambulacral cover plates of holo-type 1978TX29A (compare with Fig. 6.15) with peristome to left. Notethat secondary cover plates rapidly diminish in size distally.
FIGURE 11—Camera lucida drawings of oral area and ambulacral coverplates of Ulrichidiscus forbesi n. sp. 1, Oral area of paratype 1978TX63(compare with Fig. 7.20). Stippled plates are lateral bifurcation plates(LBP) and primary orals (PO). Note that hydro-gonopore is borderedby four and five hydropore oral plates (H) proximally and distally andaperture is slitlike; 2, proximal A ambulacral cover plates of paratype1978TX63 (compare with Fig. 7.17) with peristome to left. Note slightperradial zigzag and irregular expression of extremely small secondarycover plates. Cover plate designations follow Sumrall (1996).
FIGURE 12—Camera lucida drawings of oral area of Postibulla wester-gaardi n. sp. paratype 1978TX288 (compare with Fig. 8.11). Stippledplates are lateral bifurcation plates (LBP) and primary orals (PO). Theposterior oral protuberance is the series of six plates to the left of thehydro-gonopore. H, Hydropore oral plates.
circular in outline; oral area relatively wide with ambulacra ar-ranged into 2-1-2 symmetry (Fig. 9); primary oral plates large,lateral bifurcation plates undifferentiated; oral cover plates withthree sets of primary and secondary cover plates between PO andLBP (Figs. 9.9, 13.1); posterior oral cover plates similar to an-terior cover plates; perradial suture over peristome linear; peristo-mal opening with radially positioned oral frame plates, openingroughly oval (Figs. 9.12, 13.2); A oral frame plate symmetricalabout midline, D and E oral frame plates foreshortened along theircommon suture (Figs. 9.12, 13.2); hydropore in proximal rightCD interambulacrum, separated from oral rise, formed from twoplates, one anterior, one posterior, elevated into high spoutlikeopening with broad lip (Figs. 9.9, 9.12, 13.1, 13.2); posterior oralprotuberance absent. Ambulacra in 3–2 arrangement in maturespecimens, linear in specimens up to thecal diameter of 9 mm(Fig. 9.1, 9.2), evenly curved throughout most of length withoutoverlap in mature specimens, little taper throughout length withblunt termination, plated with high cover plates and uniserial am-bulacral floor plates; cover plates highly domed over food groove,with primary and secondary plates, each with interambulacral ex-tension; ornamented with linear striations that extend along lengthof plate; plates arranged into double biseries with secondary coverplates exposed as slivers along sutures of primary cover plates,some hint of primary cover plates arranged into more complexcycles, but this detail unclear in present material (Figs. 9.7, 13.3);floor plates poorly known in present material; periproct coveredby highly conical anal pyramid of typically nine lathe-shapedplates, anal pyramid plates with linear striations similar to orna-mentation on ambulacral cover plates (Fig. 9.10); interambulacralplates medium-sized, approximately 20–24 per interambulacrumin large specimens, imbricate, squamose to somewhat tessellate,surface with coarse-mesh lamellar stereom; pedunculate zone withperhaps one circlet of plates in mature specimens; peripheral rimplated with five irregular circlets of highly imbricate plates (Fig.9.6), exterior surface with coarse lamellar stereom; most proximalcirclet with largest plates, much wider than long, middle threecirclets decrease in size distally, with equant plates; distalmostcirclet with very small radially elongate plates; underside of pe-ripheral rim unknown in present material.
Etymology.⎯Named for the Robert Grayson, former professorat Baylor University who led the field trip to Forbes Ranch wherethe edrioasteroid locality was first discovered.
Types.⎯The type series for Parapostibulla graysoni includesholotype 1978TX109 and paratypes 1978TX4, 27C, 66A, 85, 157,258, and 268. Additional specimens found on a few other clasts

243SUMRALL ET AL.—EDRIOASTEROIDS ON PEBBLES
FIGURE 13—Camera lucida drawings of oral area and ambulacral coverplates of Parapostibulla graysoni n. sp. 1, Oral area of holotype1978TX109 (compare with Fig. 9.9). Stippled plates are lateral bifur-cation plates (LBP), primary orals (PO), and hydropore oral (HO). Notespoutlike hydro-gonopore. 2, Partially exposed oral frame of paratype1978TX4 (compare with Fig. 9.12), cover plates stippled; 3, ambulacralcover plates of paratype 1978TX4 (compare with Fig. 9.7). Note smallwedgelike secondary cover plates and hint of more complex cover platepattern. Plates from opposite side of ambulacrum stippled.
of the 1978TX number series at the Texas Memorial Museum,Austin.
Occurrence.⎯Parapostibulla graysoni is known only from un-named unit 1 of the Golf Course Formation, Dornick Hills Group,Lower Pennsylvanian (Morrowan), at Forbes Ranch, north ofMarietta, southern Oklahoma.
Discussion.⎯Parapostibulla graysoni conforms well to theconcept of Parapostibulla in Sumrall et al. (2000). It differs fromother species of Parapostibulla in the slightly more complex cov-er plate pattern and the unusual ornament on the cover plates andanal pyramid plates. Interestingly, unusual modifications to bothcover plates and anal pyramid plates are also known in Ulrichi-discus spinosus, which has small spines on both of these plate
types (Sumrall, 2001). Modifications to ambulacral cover platesand anal pyramid plates likely have similar genetic control.
ACKNOWLEDGMENTS
We thank former Baylor University students E. H. Westergaard,who found the first edrioasteroids on a sedimentary petrologyfield trip in 1981, J. Whiteside, who helped collect many of theedrioasteroids in the mid-1980s and wrote the original descriptionof the locality and specimens for his bachelor’s thesis (1987), K.Pate, who studied the edrioasteroid faunal community in the late1980s, and W. York, who helped collect additional specimens inthe late 1990s and contributed them to this study. Former BaylorProfessor R. Grayson led the original and several later field tripsand helped date unnamed unit 1 using conodont biostratigraphy.The late Joe and Margaret Forbes of Marietta, Oklahoma, andtheir children graciously allowed the authors access to the fossillocality on their ranch property. T. Kammer, T. Guensburg, andB. Pratt reviewed the manuscript and made many helpful sugges-tions for improvements. Both the Department of Geology, BaylorUniversity, and the Geology Foundation, University of Texas atAustin, funded field trips by Grayson and Bonem and by Sprinkleand Sumrall to visit the locality, and also research and publicationexpenses for this paper.
REFERENCES
ARENDT, Y. A. 1983. About some echinoderms. Bulletin of the MoscowSociety of Naturalists, Geology, 58(4):135–136.
BASSLER, R. S. 1935. The classification of the Edrioasteroidea. Smith-sonian Miscellaneous Collections, 93:1–11.
BELL, B. M. 1976. A Study of North American Edrioasteroidea. NewYork State Museum Memoir, 21, 446 p.
BILLINGS, E. 1858. On the Asteriadae of the Lower Silurian rocks ofCanada. Geological Survey of Canada, figures and descriptions of Ca-nadian organic remains, Decade 3:75–85.
BRETT, C. E., AND W. D. LIDDELL. 1978. Preservation and paleoecologyof an Ordovician hardground community. Paleobiology, 4:329–348.
BRETT, C. E., H. A. MOFFAT, AND W. L. TAYLOR. 1997. Echinodermtaphonomy, taphofacies, and lagerstatten, p. 147–190. In J. A. Watersand C. G. Maples (eds.), Geobiology of Echinoderms, PaleontologicalSociety Papers, 3.
BROWN, T. 1849. Illustrations of the Fossil Conchology of Great Britainand Ireland, With Descriptions and Localities of All Species. London,273 p.
CHAPMAN, E. J. 1860. On a new species of Agelacrinites and on thestructural relations of that genus. Canadian Journal of Industry, Sci-ence, and Art, 5:358–365.
CORNELL, S. R., C. E. BRETT, AND C. D. SUMRALL. 2003. Paleoecologyand taphonomy of an edrioasteroid-dominated hardground associationfrom tentaculutid limestones in the Early Devonian of New York: APaleozoic rocky peritidal community. Palaios, 18:212–224.
DICKEY, R. T. 1982. Lithostratigraphy and conodont biostratigraphy ofthe Jolliff Member, Golf Course Formation, in the Ardmore Basin,Oklahoma. Bachelor’s thesis, Baylor University, Waco, Texas, 83 p.
ETTENSOHN, F. R. 1975. The autecology of Agassizocrinus. Journal ofPaleontology, 49:1044–1061.
ETTENSOHN, F. R. 1980. Paragassizocrinus: Systematics, phylogeny andecology. Journal of Paleontology, 54:978–1007.
FRAUNFELTER, G. H., AND J. UTGAARD. 1970. Middle Pennsylvanianedrioasteroid from southern Illinois. Journal of Paleontology, 44:279–303.
GLASS, A. In press. Two isorophid edrioasteroids (Echinodermata) en-crusting conulariids from the Hunsruck Slate (Lower Devonian, Em-sian; Rheinische Schiefergebirge) of Germany. Senckenbergiana Le-thaea.
GUENSBURG, T. E., AND J. SPRINKLE. 1994. Revised phylogeny and func-tional interpretation of the Edrioasteroidea based on new taxa from theEarly Ordovician of western Utah. Fieldiana (Geology), n. s., 29:1–43.
GUNNELL, F. H. 1931. Conodonts from the Fort Scott Limestone of Mis-souri. Journal of Paleontology, 5:244–252.
HAM, W. E. 1969. Regional Geology of the Arbuckle Mountains,Oklahoma. Oklahoma Geological Survey Guidebook no. 3, 52 p.

244 JOURNAL OF PALEONTOLOGY, V. 80, NO. 2, 2006
HARRIS, R. W., AND R. V. HOLLINGSWORTH. 1933. New Pennsylvanianconodonts from Oklahoma. American Journal of Science, series 5, 25:193–204.
KAMMER, T. W., E. C. TISSUE, AND M. A. WILSON. 1987. Neoisorophu-sella, a new edrioasteroid genus from the Upper Mississippian of theeastern United States. Journal of Paleontology, 61:1033–1042.
KIRK, E. 1942. Ampelocrinus, a new genus from the Upper Mississippian.American Journal of Science, 240:22–28.
KOCH, D. L., AND H. L. STRIMPLE. 1968. A new Upper Devonian cystoidattached to a discontinuity surface. Iowa Geological Survey, report ofinvestigations, 5:1–49.
M’COY, F. 1847. On the fossil botany and zoology of the rocks associatedwith the coal of Australia. Annals and Magazine of Natural History(series 1), 20:145–147, 226–236, 298–312.
MEYER, D. L. 1990. Population paleoecology and comparative taphono-my of two edrioasteroid (Echinodermata) pavements: Upper Ordovi-cian of Kentucky and Ohio. Historical Biology, 4:155–178.
MILLER, J. S. 1821. A Natural History of the Crinoidea of Lily-ShapedAnimals, with Observations on the Genera Asteria, Euryale, Comatula,and Marsupites. Bryan, Bristol, England, 150 p.
MILLER, S. A., AND W. F. E. GURLEY. 1894. Descriptions of new speciesof Palaeozoic Echinodermata. Illinois State Museum of Natural HistoryBulletin, 6:62.
MOORE, R. C., AND R. M. JEFFORDS. 1945. Description of Lower Penn-sylvanian corals from Texas and adjacent states. University of TexasPublication 4401:77–208.
MOORE, R. C., AND L. R. LAUDON. 1943. Evolution and classification ofPaleozoic crinoids. Geological Society of America Special Paper, 46:1–167.
MOORE, R. C., AND F. B. PLUMMER. 1940. Crinoids from the UpperCarboniferous and Permian strata in Texas. University of Texas SpecialPublication, 3945:9–468.
OWEN, D. D., AND B. F. SHUMARD. 1851. On the number and distributionof fossil species in the Paleozoic rocks of Iowa, Wisconsin, and Min-nesota. American Association for the Advancement of Science, pro-ceedings, 235–239.
RODDA, P. U., AND W. L. FISHER. 1962. Upper Paleozoic acrothoracicbarnacles from Texas. Texas Journal of Science, 14(4):460–479.
SAINT-SEINE, R. D. 1951. Un Cirripede acrothoracique du Cretace: Ro-gerella lecontrei n. g., n. sp. Academie des Sciences (Paris), comptesrendus, 233:1051–1053.
SMITH, A. B. 1985. Cambrian eleutherozoan echinoderms and the earlydiversification of edrioasteroids. Palaeontology, 28:715–756.
STRIMPLE, H. L. 1938. A Group of Crinoids from the Pennsylvanian ofNortheastern Oklahoma. Private Publication, 15 p.
STRIMPLE, H. L., AND W. T. WATKINS. 1969. Carboniferous crinoids of
Texas with stratigraphic implications. Paleontologica Americana, 6(40):139–275 (�27 pls.).
SUMRALL, C. D. 1992. Spiraclavus nacoensis, a new clavate agelacrinitidedrioasteroid from central Arizona. Journal of Paleontology, 66:90–98.
SUMRALL, C. D. 1993. Thecal designs in isorophinid edrioasteroids. Le-thaia, 26:289–302.
SUMRALL, C. D. 1996. Late Paleozoic edrioasteroids from the NorthAmerican mid-continent. Journal of Paleontology, 70:969–985.
SUMRALL, C. D. 2000. The biological implications of an edrioasteroidattached to a pleurocystitid rhombiferan. Journal of Paleontology, 74:67–71.
SUMRALL, C. D. 2001. Paleoecology and taphonomy of two new edrioas-teroids from a Mississippian hardground in Kentucky. Journal of Pa-leontology, 75:136–146.
SUMRALL, C. D., AND A. L. BOWSHER. 1996. Giganticlavus, a new genusof Pennsylvanian edrioasteroid from North America. Journal of Pale-ontology, 70:986–993.
SUMRALL, C. D., AND R. L. PARSLEY. 2003. Morphology and biome-chanical implications of isolated discocystinid plates (Edrioasteroidea,Echinodermata) from the Carboniferous of North America. Palaeon-tology, 46:113–138.
SUMRALL, C. D., J. GARBISCH, AND J. P. POPE. 2000. The systematics ofpostibullinid edrioasteroids. Journal of Paleontology, 74:72–83.
SUMRALL, C. D., C. E. BRETT, P. T. WORK, AND D. L. MEYER. 2001.Taphonomy and paleoecology of an edrioasteroid encrusted hardgroundin the lower Bellevue Formation at Maysville, Kentucky, p. 123–131.In T. J. Algeo and C. E. Brett (eds.), Sequence, Cycle & Event Stra-tigraphy of Upper Ordovician & Silurian Strata of the Cincinnati ArchRegion. Field Trip Guidebook in conjunction with the 1999 Field Con-ference of the Great Lakes Section SEPM-SSG and the Kentucky So-ciety of Professional Geologists, guidebook 1, series XII. KentuckyGeological Survey, Lexington.
SUTHERLAND, P. K., AND R. C. GRAYSON JR. 1992. Morrowan and Ato-kan (Pennsylvanian) biostratigraphy in the Ardmore Basin, Oklahoma,p. 81–99. In P. K. Sutherland and W. L. Manger (eds.), Recent Ad-vances in Middle Carboniferous Biostratigraphy—A Symposium.Oklahoma Geological Survey Circular, 94, 181 p.
WADDINGTON, J. B. 1980. A soft-substrate community with edrioaster-oids from the Verulam Formation (Middle Ordovician) at Cambridge,Ontario, Canada. Canadian Journal of Earth Sciences, 17:674–679.
WHITESIDE, J. R. 1987. A study of Pennsylvanian (Late Morrowan) Ed-rioasteroidea from the Ardmore Basin, southern Oklahoma. Bachelor’sthesis, Baylor University, Waco, Texas, 50 p.
WILSON, M. A. 1985. Disturbance and ecological successions in an UpperOrdovician cobble-dwelling hardground fauna. Science, 228:575–577.
ACCEPTED 3 FEBRUARY 2005
Copyright © 2022 FDOKUMEN




















