
Effects of ventral and dorsal CA1 subregional lesions on trace fear conditioning
Upload
independentCategory
view
2download
0
Official Journal of the Japanese Society of Child Neuroloov
ELSEVIER Brain & Development 2.5 Suppl. 1 (2003) S3-S 14
www.elsevier.com/locate/braindev
Does an imbalance between the dorsal and ventral striatopallidal systems play a role in Tourette’s syndrome? A neuronal circuit approach
Henk J. Groenewegen a, Odile A. van den Heuvelb, Danielle C. Cathb, P. Voorn a, Dick J. Veltmanb
aDepartment of Anatomy, bDepartment of Psychiatry, Research Institute Neurosciences Vrije Universiteit, VU University Medical Center (VlJmc), Amsterdam, The Netherlands
Abstract
Tourette’s syndrome is characterized by simple, involuntary muscle contractions and/or more complex movements or stereotyped behaviors, including vocalizations. There are strong indications that the basal ganglia play an important role in the pathophysiology of Tourette’s syndrome. The present account reviews the functional anatomy of the basal ganglia, with an emphasis on the prefrontal cortex-ventral striatopallidal system. Different parts of the basal ganglia and thalamocortical system, with a focus on the premotor and prefrontal cortices, are connected with each other via parallel, functionally segregated basal ganglia-thalamocortical systems. These parallel circuits, representing sensorimotor, cognitive and emotional-motivational behavioral processes, are connected with each other through specific pathways that serve to integrate these various functions. In the context of the discussion on the pathophysiological mechanisms that lead to the expression of tics, emphasis is placed on the pathways that lead from the ventral striatum via the dopaminergic substantia nigra to the dorsal striatum. The dorsal striatum is crucial for habit formation. A conclusion of this overview of the anatomical organization of the basal ganglia is that via dopaminergic pathways limbic-relation information can influence the expression of (fragments of) motor and behavioral repertoires. Whether such mechanisms indeed play a role in the expression of tics in Tourette’s syndrome remains to be established. 0 2003 Elsevier Science B .V. All rights reserved.
1. Introduction
Tourette’s syndrome (TS) is a neuropsychiatric disorder that has its onset in childhood and is characterized by multiple chronic tics. Tics in TS can be simple, involuntary focused muscle contractions, or they may encompass more complex movements or stereotyped behaviors, including vocalizations [l-5]. For the diagnosis of TS, it is important that the tics occur frequently, i.e., several times daily, and that they have been present throughout a period of more than a year. However, over time symptoms may wax and
*Correspondence to: H.J. Groenewegen, MD, PhD, Department of Anatomy, VU University Medical Center (VUmc), MF-G-102, P.O. Box 7057, 1007 MB Amsterdam, The Netherlands. Tel.: +31-20.4448040 or 4448033; Fax: +31-20-44480.54.
E-mail address: [email protected]
wane, and often the character of the tics changes over time, subsequently involving muscles or muscle groups of different parts of the body. Simple tics concern contractions of single muscles or small groups of muscles, more complex tics involve seemingly coordinated movements or fragments of behaviors that could be considered as elements of normal movements or of purposeful behavioral acts. However, tics in TS are largely beyond voluntary control, in essence purposeless, and with a strong repetitive character. In many TS patients, tics are preceded by sensory phenomena, i.e., itching sensations in particular body regions. Subsequently, tics often are expressed to relieve these sensations. A characteristic feature of tics - in contrast to other movement disorders - is that patients can suppress tics for a period of time. Thus, tics are partly under voluntary control [6,7]. However, tic suppression often leads to an urge to ‘release’
0387.7604/03/$ - see front matter 0 2003 Elsevier Science B.V. All rights reserved.

S4 H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14
the tics, rebound and an increase of emotional tension. The subsequent release of the suppressed tics may lead to emotional relief. Stress and hormones appear to have an influence on the severity and the frequency of tics, and this influence may be in either direction, thus either worsening or alleviating the symptoms. Finally, during concentration on specific behavioral tasks, tics may also be suppressed.
Various neuropsychiatric disorders have been associated with Tourette’s syndrome, most prominently (obsessive) compulsive behaviors, but also Attention Deficit Hyperac- tivity Disorder (ADHD) [S-14]. Compulsive behaviors are in most instances accompanied by repetitive thoughts that are involuntary, anxiety provoking (intrusions) and sense- less. By definition, compulsions are designed to reduce or neutralize anxiety or tension, provoked by the obsessions [15]. Such compulsive behaviors may encompass complex behavioral sequences or they can be rather simple and in that case difficult to distinguish from complex tics. However, complex tics are most often not accompanied by intrusive thoughts. Repetitive compulsive behaviors that are accompanied by anxiety and have become ritualistic in an attempt to relieve anxiety, are diagnosed as obsessive- compulsive behaviors. Symptoms of obsessive-compulsive disorder (OCD) and tic disorders like TS may co-occur. In other words, patients with obsessive-compulsive disor- der may present with or without tics, while TS patients may or may not have obsessive-compulsive symptoms. Furthermore, these disorders have been shown to exhibit a familial character. Therefore, OCD and TS have been con- sidered to be linked and to form exponents of a spectrum of disorders with overlapping phenotypes and underlying neuropathology [4]. At the phenotypical level, a common feature of all three disorders, i.e., TS, OCD and ADHD, is the poor control of impulses that lead to movements or complex behaviors that are in most cases inappropriate in the given context [16]. This conclusion is important in the light of inferences that have been made about the involvement of the basal ganglia in the pathophysiological mechanisms of these neuropsychiatric disorders, including TS. For example, PaIumbo et al. [17] have formulated a hypothesis of ‘developmental basal ganglia syndromes’, in which tics, OCD, ADHD and autism are considered to be the consequence of a deficit in basal ganglia development, either genetic or acquired.
1.1. A role for the basal ganglia in TouretteS syndrome?
Several lines of evidence indeed indicate that dysfunc- tions of the basal ganglia in their relationships with the cerebral cortex play an important role in the pathophysiol- ogy of Tourette’s syndrome. Thus, neuroimaging data show abnormalities, both in structural and functional aspects of the basal ganglia (reviews: Refs. 18, 19). Structurally, the results of morphometric MRI studies indicate that the normal (a)symmetrical relationships of the basal ganglia and associated structures may be changed in TS (review:
Ref. 18) [20,21]. Functional neuroimaging studies in TS have been performed mostly in resting state conditions, comparing baseline blood flow or glucose metabolism em- ploying Positron Emission Tomography (PET) or Single Photon Emission Tomography (SPECT) in TS patients and normal controls. The results of such studies likewise show abnormalities in TS patients in both basal ganglia struc- tures and, although more variably, in connectionally related orbitofrontal cortical and thalamic structures [l&19]. Un- fortunately, to date only few symptom-related functional neuroimaging studies have been executed in TS patients. However, the results of a functional MRI study in TS patients comparing conditions of active tic suppression with a baseline condition in which tics were allowed to be freely expressed, demonstrated differences in activity in basal ganglia structures as well as in cerebral cortical regions between these conditions. In particular, ventral parts of the basal ganglia and limbic-related cortical areas appeared to be involved in tic suppression [22]. Finally, in line with the clinical experience that dopaminergic antagonists are effective in the symptomatic treatment of TS patients, the results of a number of functional neuroimaging studies have shown abnormalities in dopamine transporter and dopamine receptor binding in the striatum of TS patients (review: Ref. 18) [23,24].
From the previous paragraphs, it can be concluded that (dysfunctions of) basal ganglia structures may play a crucial role in the pathophysiological mechanisms that underly the symptomatology of Tourette’s syndrome. All the above- mentioned findings nicely link with recent insights in the functional anatomy of the basal ganglia and their role in motor, cognitive and behavioral control (e.g. Refs. 16, 25, 26). In this context, the present paper contains a brief review of the functional-anatomical aspects of the basal ganglia and their relationships with the cerebral cortex and the thalamus. Interestingly, both the sensorimotor- and the limbic-innervated parts of the basal ganglia including the dorsal and ventral striatum, respectively, have been implicated in the pathophysiology of TS [20,22,27,28]. Although the focus of the present account will be on the ventral, limbic-innervated parts of the basal ganglia, a brief description of the main organizational aspects of the basal ganglia as a whole will be given.
In the last decades, the circuitries in which the dorsal and ventral components of the basal ganglia are implicated, have been viewed as anatomically and functionally largely segregated and arranged in parallel. However, recently more emphasis has been placed on possible ways of reciprocal communication between the dorsal, sensorimotor cortical- basal ganglia circuits and the ventral, ‘limbic-related’ basal ganglia circuits. These ways of communication between functionally different cortical-basal ganglia circuits may prove to be particularly important for a better understand- ing of the pathophysiological mechanisms that underlie neuropsychiatric disorders like TS.
H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14 S.5
2. Some basic principles of cortical-basal ganglia relationships
2.1. Basal ganglia outputs ‘focus’ on the frontal lobe
The basal ganglia, consisting of the striatum, the pal- lidum, the subthalamic nucleus and the substantia nigra have a special functional-anatomica relationship with the cerebral cortex. All parts of the cerebral cortex, ranging from the primary sensory and motor cortices to the highest association cortices, i.e., the hippocampal, parahippocampal and prefrontal areas, send projections to the input structure of the basal ganglia, i.e. the striatum. In turn, the out- put structures of the basal ganglia, i.e., specific parts of the pallidurn and the substantia nigra (see below), project via the thalamus back to the cerebral cortex. Importantly, these basal ganglia-thalamocortical projections are directed primarily, although not exclusively, to the frontal lobe, and they involve in particular the premotor and prefrontal cortices (e.g. Refs. 29, 30, 31). The abundance of the basal ganglia projections to the frontal lobe emphasizes the role of the basal ganglia complex in premotor and prefrontal functions, i.e. in motor, executive behavioral and cognitive functions. However, it must be noted that the basal ganglia also have a considerable output to various brainstem struc- tures, prominently including the pedunculopontine nucleus. These descending projections signify a role for the basal ganglia in a number of basic aspects of motor control, like muscle tone, posture and balance, as well as in innate movement patterns like locomotion, mastication, etc.
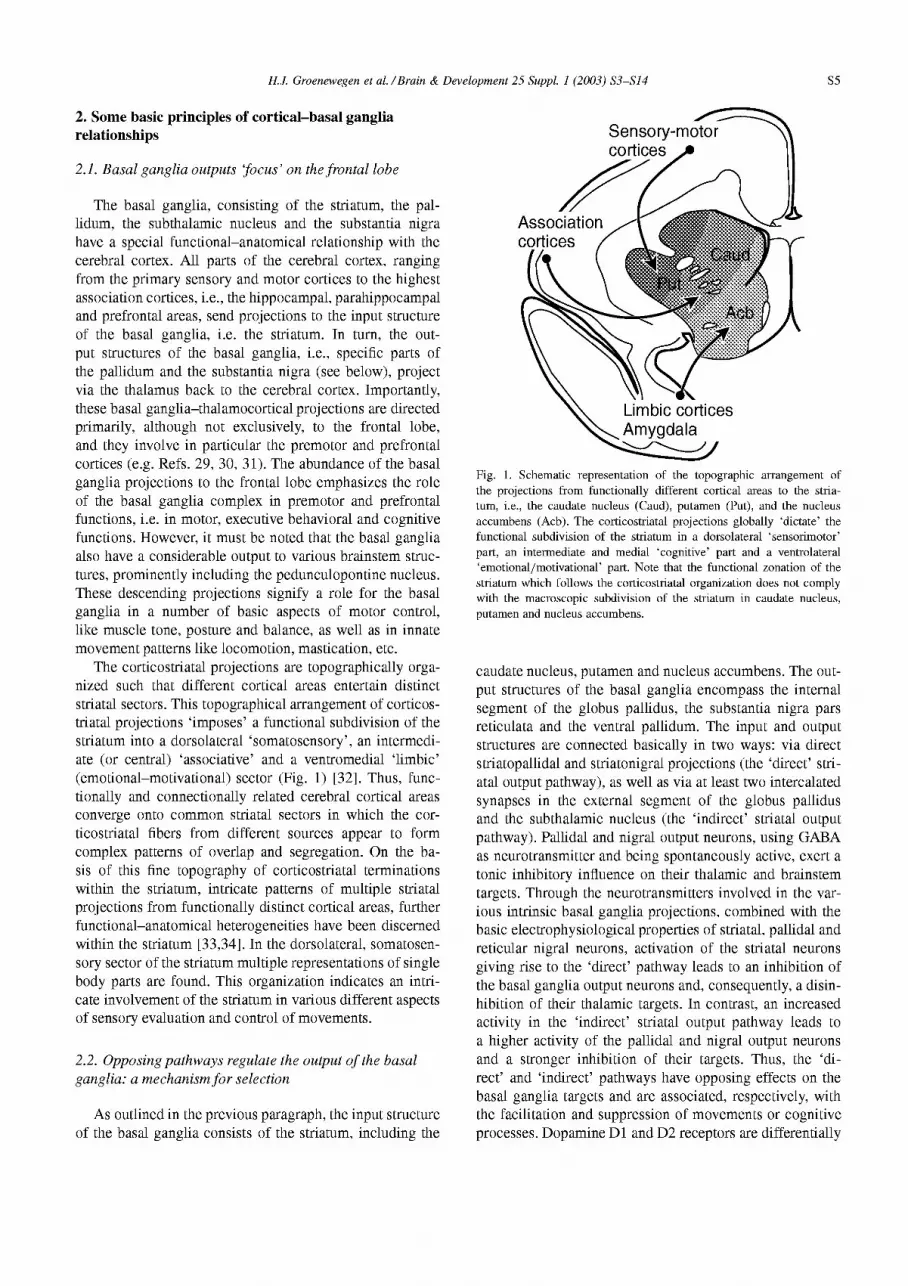
The corticostriatal projections are topographically orga- nized such that different cortical areas entertain distinct striatal sectors. This topographical arrangement of corticos- triatal projections ‘imposes’ a functional subdivision of the striatum into a dorsolateral ‘somatosensory’, an intermedi- ate (or central) ‘associative’ and a ventromedial ‘limbic’ (emotional-motivational) sector (Fig. 1) [32]. Thus, func- tionally and connectionally related cerebral cortical areas converge onto common striatal sectors in which the cor- ticostriatal fibers from different sources appear to form complex patterns of overlap and segregation. On the ba- sis of this fine topography of corticostriatal terminations within the striatum, intricate patterns of multiple striatal projections from functionally distinct cortical areas, further functional-anatomical heterogeneities have been discerned within the striatum [33,34]. In the dorsolateral, somatosen- sory sector of the striatum multiple representations of single body parts are found. This organization indicates an intri- cate involvement of the striatum in various different aspects of sensory evaluation and control of movements.
2.2. Opposing pathways regulate the output of the basal ganglia: a mechanism for selection
As outlined in the previous paragraph, the input structure of the basal ganglia consists of the striatum, including the
T Sensory-motor
Fig. 1. Schematic representation of the topographic arrangement of the projections from functionally different cortical areas to the stria- turn, i.e., the caudate nucleus (Caud), putamen (Put), and the nucleus accumbens (Acb). The corticostriatal projections globally ‘dictate’ the functional subdivision of the striatum in a dorsolateral ‘sensorimotor’ part, an intermediate and medial ‘cognitive’ part and a ventrolateral ‘emotional/motivational’ part. Note that the functional zonation of the striatum which follows the corticostriatal organization does not comply with the macroscopic subdivision of the striatum in caudate nucleus, putamen and nucleus accumbens.
caudate nucleus, putamen and nucleus accumbens. The out- put structures of the basal ganglia encompass the internal segment of the globus pallidus, the substantia nigra pars reticulata and the ventral pallidurn. The input and output structures are connected basically in two ways: via direct striatopallidal and striatonigral projections (the ‘direct’ stri- atal output pathway), as well as via at least two intercalated synapses in the external segment of the globus pallidus and the subthalamic nucleus (the ‘indirect’ striatal output pathway). Pallidal and nigral output neurons, using GABA as neurotransmitter and being spontaneously active, exert a tonic inhibitory influence on their thahtmic and brainstem targets. Through the neurotransmitters involved in the var- ious intrinsic basal ganglia projections, combined with the basic electrophysiological properties of striatal, pallidal and reticular nigral neurons, activation of the striatal neurons giving rise to the ‘direct’ pathway leads to an inhibition of the basal ganglia output neurons and, consequently, a disin- hibition of their thahtmic targets. In contrast, an increased activity in the ‘indirect’ striatal output pathway leads to a higher activity of the pallidal and nigral output neurons and a stronger inhibition of their targets. Thus, the ‘di- rect’ and ‘indirect’ pathways have opposing effects on the basal ganglia targets and are associated, respectively, with the facilitation and suppression of movements or cognitive processes. Dopamine Dl and D2 receptors are differentially

S6 H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14
localized in striatal neurons that give rise to the direct and indirect pathways, respectively [35]. Striatal dopamine levels are thought to largely determine the balance between the direct and indirect pathways and, consequently, the degree of basal ganglia output. Higher levels of striatal dopamine, via opposing effects on striatal neurons giving rise to the direct and indirect pathways, lead to a lower (inhibitory) activity of the basal ganglia output neurons and, consequently, a disinhibition of the thahtmic and brain stem targets. This is accompanied with the facilitation of movements and cognitive processes (review: see Ref. 36). The reverse is true for situations with low levels of striatal dopamine, leading in pathological conditions like in Parkin- son’s disease to hypokinesia and bradyphrenia. Moreover, considering the various interconnections between the dif- ferent basal ganglia components and the fact that extrinsic cortical projections can also reach the subthahtmic nucleus directly, thus bypassing the striatum, Bolam et al. [37] have proposed to view these indirect connections as part of an indirect modulatory ‘network’ rather than an indirect ‘pathway’. The ‘network’ view of the indirect connections between input and output structures of the basal ganglia suggests a richer functional repertoire for the basal ganglia than just facilitation or inhibition.
Indeed, the global description of the basal ganglia circuit model, as outlined above, could give rise to the idea that the basal ganglia in a rather general way act as a brake or a facilitator for the expression of movements, and cognitive and behavioral processes. However, the highly ordered way in which the extrinsic and intrinsic connections of the basal ganglia are organized, points to a more sophisticated role of the basal ganglia in motor and behavioral processes. It has been suggested by several authors [38,39], that one of the primary functions of the basal ganglia entails the selection of motor or behavioral programs, or elements thereof, that are appropriate for a particular context. More specifically, Mink [38] has proposed that basal ganglia are crucial for the facilitation of desired movements and the suppression of unwanted, competing movements. Thus, in resting conditions the basal ganglia, with their strong and tonic inhibitory input on the thahtmic and brain stem target areas, act as a brake on the expression of motor and behavioral output. At the moment it is being ‘decided’ in the prefrontal and premotor cortices to execute a motor program, this information is sent to the striatum as well as, in a corollary manner, to the subthalamic nucleus. Activation of striatal neurons that give rise to the direct output pathway to either the internal segment of the globus pallidus or the substantia nigra pars reticulata leads to a disinhibition of the thalamocortical system that, in mm, provides the final output for the desired movement. The ‘parallel’ cortical excitation of the subthalamic nucleus leads to a higher activity of the basal ganglia output neurons not concerned with the intended movement or complex of movements, resulting in the suppression of potentially competing motor output [38].
In the present context, it is of importance to note that the striatum has been hypothesized to function as a context detector [40]. As will be discussed below with respect to the ‘ventral striatum’, activity of convergent corticostriatal inputs are probably necessary to activate striatal neurons, leading to an output of the basal ganglia. Therefore, the basal ganglia might play an important role in the mechanisms that in the cerebral cortex lead to the ‘decision’ to perform a particular movement or behavioral act in a particular context.
2.3. Parallel organization of basal ganglia-thalamocortical circuits: segregation and integration
The topographical organization of the corticostriatal projections, as well as of the subsequent striatopaIlidaI/ striatonigral, pallidothaIamic/nigrothalamic and thalamo- cortical projections forms the anatomical basis for the recognition of a number of parallel, functionally segregated cortical-basal ganglia-thalamocortical circuits that, at least in part, form re-entrant loops [29-3 11. The functional con- notation of the various circuits relates to the predominant cerebral cortical input to the sector of the striatum that forms the striatal node in this circuit. Thus, the three differ- ent striatal sectors briefly outlined above, i.e., sensorimotor, associative and limbic, may be viewed as part of three ‘macro-circuits’ that entertain different parts of the frontal lobe, i.e., the premotor, and the dorsolateral prefrontal and orbitomedial prefrontal cortices, respectively. Within these macrocircuits, smaller (micro)circuits can be recognized that subserve specific functions within the broader func- tional domain, i.e. sensorimotor (movements), associative (cognitive functions) or limbic (emotional-motivational be- haviors). This more detailed organization has most clearly be shown for the sensorimotor macrocircuit within which specific microcircuits are related to different parts of the body and subserve various aspects of movements (direc- tion, force, etc.) [31]. However, the fine topography that exists within the prefrontal cortical projections to the ven- tral striatum likewise indicates that within the ‘limbic’ macrocircuit various subcircuits subserving different as- pects of emotional-motivational behavior exist [30,41]. The functional-anatomica arrangement of these basal ganglia- thalamocortical macro- and microcircuits, that all contain re-entrant components leading back to the distinct areas of the (pre)frontaI cortex that originally formed the starting point of that particular circuit (closed component), sug- gests a large degree of segregation between these circuits. However, it is clear that the various functions that are subserved by the different basal ganglia-thalamocortical circuits cannot be executed independently and it has been hypothesized that there exist several ways of communica- tion between these circuits (e.g. Refs. 42-45). As will be discussed below, these interconnections between the vari- ous functionally identifiable basal ganglia-thalamocortical circuits might be important for the interpretation of the role

H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14 s7
of the ‘limbic’ ventral striatopallidal circuit in sensorimotor functions.
2.4. Compartmental organization of the striatum
Since more than 25 years it has been recognized that the striatum consists basically of two compartments, i.e., the striosome and the matrix (cats and primates) [46], or patch and matrix (rats) [47] compartments. These compartments have first been described on the basis of their histochemical differences, but it has subsequently been shown that they differ also with respect to their input-output relationships [35,48] and ontogenetic origin [49]. The striatal matrix compartment is by far the largest, occupying about 90% of the striatal volume. The striosome or patch compartment represents only about 10% of the striatal volume, although there may be species differences in the volumetric relation- ships between striosome/patch and matrix compartments [50,5 11. Although the functional distinction and significance of the two striatal compartments is not yet understood, the basis most probably lies in the differences with respect to their inputs and outputs. The general striatal input-output relationships outlined above, primarily hold for the matrix compartment. Thus, topographical relationships of corticos- triatal inputs and the projections to the basal ganglia output structures all concern the matrix compartment. Inputs to the striosome/patch compartment are derived from specific parts of the cortex, i.e. the medial prefrontal cortex [52,53] and specific cortical layers [53,54]. Other inputs to the strisome/patch compartment are derived from the paraven- tricular thalamic nucleus [55] as well as specific subnuclei of the basal amygdaloid complex [56,57]. With respect to the outputs of the striosome/patch compartment, it has been shown that this compartment projects preferentially to a particular part of the pars compacta of the substantia nigra, i.e., the ventral tier of dopaminergic neurons (see Fig. 3) [35,58]. Thus, the striosome/patch compartment does not appear to entertain the output neurons of the basal ganglia in the internal segment of the pallidum or the substantia nigra pars reticulata, but to reach pars compacta neurons that give rise to ascending dopaminergic pathways. Interestingly, it has been suggested that those dopaminergic neurons that re- ceive inputs from the striosome/patch compartment project back to that same striatal compartment. In contrast, the matrix compartment is receiving inputs from a different set of dopaminergic neurons that are beyond the direct control of strisome/patch projections [58,59]. Of particular interest in the context of the pathophysiology of TS, it has recently been suggested that the striosome/patch compartment and its relationships with a particular set of dopaminergic neu- rons plays an important role in the regulation of the motor output of the surrounding matrix [60].
3. Functional-anatomical organization of the ventral striatum
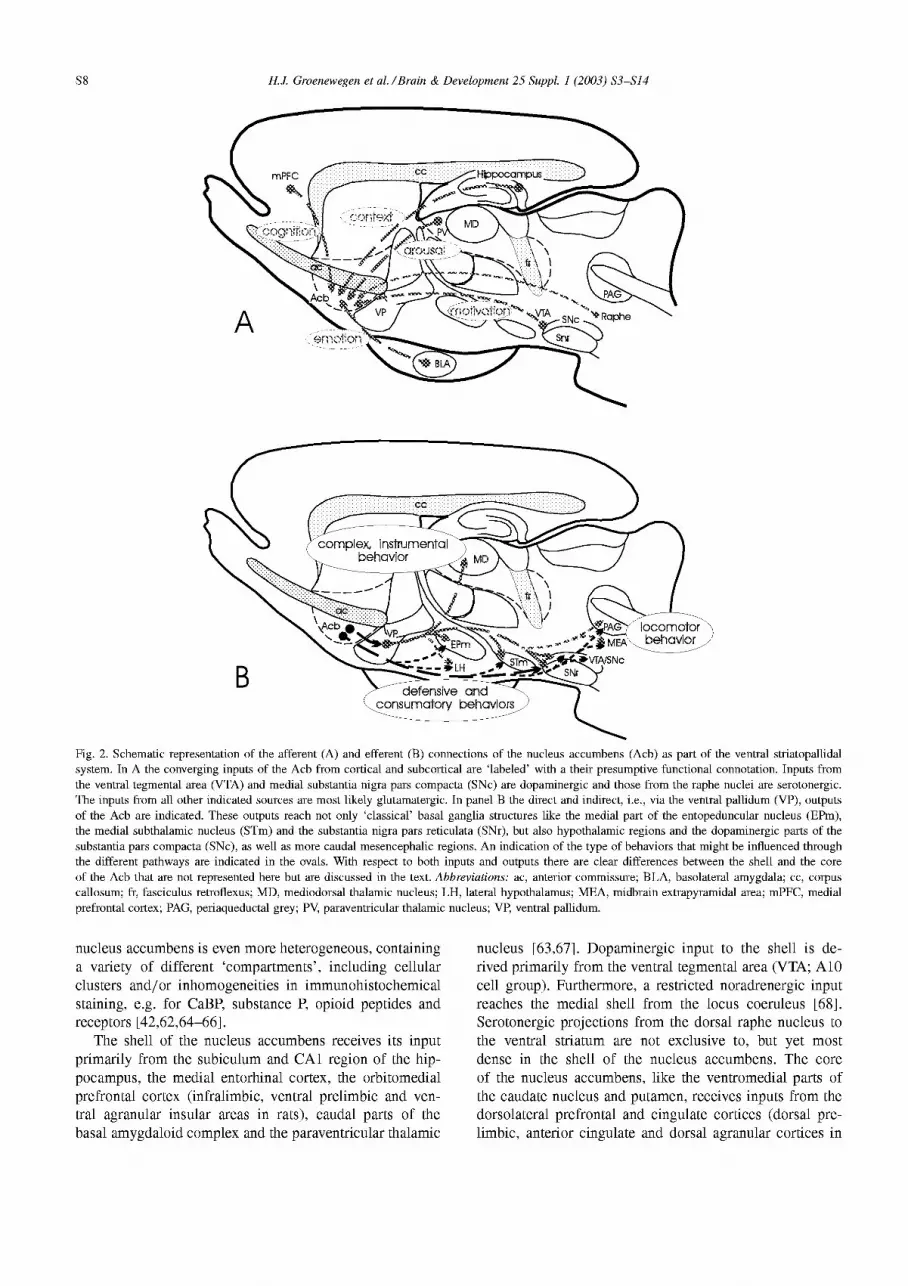
As indicated above, the ventral striatum is character- ized by inputs from ‘limbic’ cortices, in particular from hippocampal, parahippocampal and orbitomedial prefrontal areas. According to these connectional ‘criteria’, the ventral striatum not only includes the nucleus accumbens but also ventral and medial parts of the caudate nucleus and puta- men. Further inputs to the ventral striatum are derived from the basal amygdaloid complex, midline thalamic nuclei, and the dopaminergic ventral tegmental area (VTA) (Fig. 2A). This combination of inputs implies that the ventral striatum is the site of convergence of limbic cortical and amygdaloid inputs with the mesolimbic dopamine system. The efferent projections of the ventral striatum, like those of the rest of the striatum, reach the pallidal complex and substantia nigra, in this way establishing an output to motor-related structures. However, the organization and distribution of ventral striatal efferent projections differs in a number of aspects from those of the dorsal striatum, notably in its projections to hypothalamic regions and the dopaminergic cell groups in the ventral mesencephalon (Fig. 2B). Further details of the organization of inputs and outputs of the ven- tral striatum will be briefly discussed below, as they might be important for an understanding of a possible role of this part of the basal ganglia in TS. First, a brief outline will be given of the (immuno)histochemical and connectional heterogeneity of the ventral striatum.
3.1. Shell-core distinction
The ventral striatum, like its dorsal counterpart, is a rather heterogeneous structure. The nucleus accumbens occupies an extensive part of the ventral striatum. Within the nucleus accumbens there exists a very prominent distinction between two larger subregions, i.e., a peripheral (medially, ventrally and laterally located) shell subregion and a centrally located core subregion [42,61,62]. Shell and core subregions of the nucleus accumbens differ with respect to cytoarchitectonics, immunohistochemical and receptor composition as well as their input- and output relationships. These differences are reflected in important functional differences between shell and core that have attracted much attention in the last decade [42,62-641.
The most widely used marker for a distinction between shell and core is the differential immunoreactivity for cal- bindin D2s~ (CaBP), a calcium-binding protein that is abundantly expressed in the core and is only weakly to moderately present in the shell. Immunoreactive staining in- tensity for CaBP in the core is very comparable to that in the dorsally adjacent caudate nucleus and putamen. Within the caudate nucleus and putamen, the CaBP staining differenti- ates between the striosome/patch and matrix compartments in that the former stand out as weakly immunoreactive ‘islands’ in the strongly staining matrix. The shell of the
S8 H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14
Fig. 2. Schematic representation of the afferent (A) and efferent (B) connections of the nucleus accumbens (Acb) as part of the ventral striatopallidal system. In A the converging inputs of the Acb from cortical and subcortical are ‘labeled’ with a their presumptive functional connotation. Inputs from the ventral tegmental area (VTA) and medial substantia nigra pars compacta (SNc) are dopaminergic and those from the raphe nuclei are serotonergic. The inputs from all other indicated som-ces are most likely glutamatergic. In panel B the direct and indirect, i.e., via the ventral pallidum (VP), outputs of the Acb are indicated. These outputs reach not only ‘classical’ basal ganglia structures like the medial part of the entopeduncular nucleus (Elk), the medial subthalamic nucleus (STm) and the substantia nigra pars reticulata (SNr), but also hypothalamic regions and the dopaminergic parts of the substantia pars compacta (SNc), as well as more caudal mesencephalic regions. An indication of the type of behaviors that might be influenced through the different pathways are indicated in the ovals. With respect to both inputs and outputs there are clear differences between the shell and the core of the Acb that are not represented here but are discussed in the text. Abbreviations: ac, anterior commissure; BLA, basolateral amygdala; cc, corpus callosum; fr, fasciculus retroflexus; MD, mediodorsal thalamic nucleus; LH, lateral hypothalamus; MEA, midbrain extrapyramidal area; mPFC, medial prefrontal cortex; PAG, periaqueductal grey; PV, paraventricular thalamic nucleus; VP, ventral pallidum.
nucleus accumbens is even more heterogeneous, containing a variety of different ‘compartments’, including cellular clusters and/or inhomogeneities in immunohistochemical staining, e.g. for CaBP, substance P, opioid peptides and receptors [42,62,64-661.
The shell of the nucleus accumbens receives its input primarily from the subiculum and CA1 region of the hip- pocampus, the medial entorhinal cortex, the orbitomedial prefrontal cortex (infralimbic, ventral prelimbic and ven- tral agranular insular areas in rats), caudal parts of the basal amygdaloid complex and the paraventricular thalamic
nucleus [63,67]. Dopaminergic input to the shell is de- rived primarily from the ventral tegmental area (VTA; A10 cell group). Furthermore, a restricted noradrenergic input reaches the medial shell from the locus coeruleus [68]. Serotonergic projections from the dorsal raphe nucleus to the ventral striatum are not exclusive to, but yet most dense in the shell of the nucleus accumbens. The core of the nucleus accumbens, like the ventromedial parts of the caudate nucleus and putamen, receives inputs from the dorsolateral prefrontal and cingulate cortices (dorsal pre- limbic, anterior cingulate and dorsal agranular cortices in

H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14 s9
rats), the parahippocampal cortex, including the lateral en- torhinal and perirhinal cortices, the rostra1 part of the basal amygdaloid complex and caudal midline and intralaminar thalamic nuclei. Dopaminergic input to the nucleus accum- bens core is derived from the lateral part of the VTA and the substantia nigra pars compacta [63]. On the basis of these patterns of afferent connections, the shell can be considered as the main site of convergence of visceral-limbic (orbito- medial prefrontal cortex and hippocampus/amygdala) and arousing stimuli (midline thalamus), while the core and adjacent parts of the caudate nucleus and putamen receive converging stimuli from higher limbic-cognitive and exec- utive behavioral origins (dorsolateral prefrontal cortex and rostra1 amygdala). Furthermore, the latter striatal regions receive arousing stimuli from the midline and intralaminar thalamus. Thus, whereas the basic patterns of cortical and subcortical afferents to shell and core are rather similar, the afferents of the core are more comparable to those of the adjacent caudate-putamen than to those of the shell. These differences between shell and core are further emphasized by the differences in the dopaminergic innervation of the shell and core subregions [69].
The differences in efferent connectivity of shell and core are even more pronounced. Thus, the core projects to the subcommisural ventral pallidum, the most medial part of the entopeduncular nucleus (internal pallidal segment) and the dorsomedial part of the pars reticulata of the substantia nigra, i.e., ‘classical’ striatal targets. Whereas the shell also projects to the ventral pallidum and the substantia nigra, this part of the accumbens also reaches lateral preoptic, and lateral and medial hypothalamic areas and caudal mesencephalic regions including the midbrain extrapyramidal area and ventromedial part of the periaque- ductal gray matter (Fig. 2B). More importantly, the ventral pallidal and nigral targets of the shell differ considerably from the areas in these structures that are reached by the core subregion. Thus, the medial and ventral parts of the ventral pallidum, the area primarily reached by shell affer- ents, immunohistochemically has characteristics of both the external and internal segments of the globus pallidus. With respect to its connections, it is remarkable that this part of the ventral pallidum projects heavily to the mediodorsal thalamic nucleus, but not to the subthalamic nucleus. By contrast, the dorsal, subcommissural part of the ventral pallidum, i.e., the prime recipient of core afferents, has strong reciprocal connections with the subthalamic nucleus and hardly any projections to the thalamus, a pattern that is very comparable with the dorsal globus pallidus (in rats; or the external pallidum in primates). Most interesting are the projections from the accumbens shell subregion to the nigral complex: the primary targets of the shell projections are the dopaminergic neurons of the VTA and the dorsal tier of the substantia nigra pars compacta [43,45,62,70]. In contrast, the projections from the accumbens core to the substantia nigra primarily target the dorsomedial part of the pars reticulata [71]. This brief summary indicates that
the shell and core of the nucleus accumbens are involved in quite different circuitries and, consequently, subserve different functions (see below).
3.2. Compartments and ensembles of neurons
As indicated above, the ventral striatum is a highly compartmentalized structure. Patch/striosome-matrix com- partments exist within the core of the nucleus accumbens that are very comparable to such compartments in the other parts of the striatum. However, the neurochemical ‘identity’ of these compartments, at least in rats, in rostraI and caudal parts of the core differs considerably [54,59,65]. Furthermore, the shell subregion contains an even greater diversity of larger and smaller compartments that present in part as cell clusters or cell-sparse regions, or as im- munohistochemically identifiable subregions that all seem to have different input-output relationships. Also in humans the compartmental structure of the ventral striatum is more complex than in its dorsal counterpart [66,72,73]. The ventral striatum has been associated with a great variety of functional and behavioral aspects, ranging from rather simple locomotor activity to highly complex behaviors. Among others based on the highly intricate anatomical organization, it has been suggested that within the ven- tral striatum a multitude of ensembles of neurons may be variably active in different behavioral situations [74]. Which ensembles of neurons become active and provide an output of the ventral striatopallidal system, depends upon the patterns of convergence of active excitatory inputs from cortical, hippocampal, thalamic and amygdaloid origin, in combination with the dopaminergic input (e.g. Refs. 74- 76). In that respect, basic electrophysiological mechanisms in the ventral striatum are very similar to those in its dorsal counterpart, albeit that in the ventral striatum convergence of highly integrated and evaluated information (e.g. from hippocampus, amygdala and prefrontal cortex) leads to a particular output while, at the other extreme, i.e., in the dorsolateral striatum, it is convergence of relatively more ‘simple’ sensory and motor signals that elicits a particular output.
4. Interactions between the dorsal and ventral striatum
In the past two decades, the principle of a parallel, functionally segregated organization of basal ganglia- thalamocortical circuits has given an enormous impetus to hypotheses about the basic functions of the basal gan- glia and the role of these structures in neurological and psychiatric diseases. However, several authors have empha- sized the importance of an integration of the information ‘flowing’ through the various circuits for the production of coherent behavior. Several ways of communication and integration between these circuits have been hypothesized. Zahm and Brog [42] have been among the first authors to

SlO H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14
emphasize the existence of ‘open’ components in the con- nections between the cerebral cortex, basal ganglia and tha- lamus, suggesting a ‘spiral’ of connections leading from the limbic-innervated parts of the basal ganglia via the thaIa- mus and subsequent cortical-basal ganglia-thalamocortical loops to the premotor cortex (cf. also Ref. 64). In a similar vein, Joel and Weiner [44] have proposed the existence of so-called ‘open-interconnected’ basal ganglia- thalamocortical circuits (so-called ‘split circuits’) that like- wise have a tendency to lead from limbic-innervated parts of the corticostriatal system to motor-related cortical-basal ganglia circuits. However, according to their hypothesis, for specific circuits the reverse might also hold true. Several neural substrates for a ‘communication’ between different basal ganglia-thalamocortical circuits have been suggested, such as via cortico-cortical pathways within the frontal lobe, via different thahtmic nuclei, or through intrastriatal connections [43,44,77,78]. These various pathways would allow activity in one cortical-subcortical circuit to in- fluence information processing in another circuit. In this way, emotional and motivational processes might influence sensori-motor functions. Yet another, most interesting route through which the information processing in different basal ganglia-thalamocortical circuits is possibly integrated, oc- curs via the dopaminergic system. Since the landmark paper by Nauta et al. [70] on the efferents of the nucleus accumbens, it is known that the ventral striatum reaches a broad array of dopaminergic neurons in the ventral mesencephalon. These neurons in turn project to extensive regions of the dorsal striatum. As indicated above, in rats the substantia nigra pars compacta contains at least two subpopulations of dopaminergic neurons. First, a dorsal tier of neurons with horizontally oriented dendrites that project to the matrix compartment of the striatum and, second, a ventral tier of neurons with more vertically ori- ented dendrites extending into the pars reticulata of the substantia nigra that primarily project to the striatal patch compartment (Fig. 3). The dorsal tier neurons, together with the lateral VTA receive projections from the shell of the nucleus accumbens. As a consequence, the striatal output neurons in the shell are in a position to influence the dopaminergic activity in extensive parts of the matrix compartments of the core of the nucleus accumbens and the caudate-putamen complex via the ascending nigrostriatal pathways [45,79,80]. The caudo-medial part of the shell region is an exception: this region is reciprocally connected with the medial part of the VTA [45,79]. Furthermore, as indicated above, the ventral tier of dopaminergic neurons is reciprocally connected with the patch compartment in both the core and caudate-putamen. It must further be realized that the ventral striatum, in particular the shell region, can influence the dopaminergic neurons also indirectly via the ventral pallidum and the subthahtmic nucleus. In addition, the medial prefrontal cortex directly projects to the VTA and substantia nigra pars compacta, as well as indirectly via the nucleus accumbens [Sl]. In that respect the pro-
habits
Fig. 3. Schematic representation of the interrelationships between the striatum and the substantia nigra, illustrating the connections between the ventral striatum, in particular the shell of the nucleus accumbens, and the dorsal striatum via the dopaminergic system, specifically the dorsal tier of the substantia nigra pars compacta (see also text). In ad- dition the reciprocal relationships between the striatal striosome/patch compartment and the dopaminergic ventral tier nem-ens of the sub- stantia nigra pars compacta are depicted. Possible interactions between the striosome/patch compartment and the surrounding matrix has been represented by arrows and a question mark. At the right-hand side of the figure different types of behavior related to particular sectors of the striatum are represented. The dopaminergic projections from the dorsal tier nem-ens to the dorsal striatum might represent a neural pathway through which limbic-related information can influence the motor-related (habits) outputs of the dorsal striatum. For further consid- erations, see text. Abbreviations: SNC, substantia nigra pars compacta; SNR, substantia nigra pars reticulata; VTA, ventral tegmental area.
jections from the shell of the nucleus accumbens to the dopaminergic neurons are not to be considered in isolation. Furthermore, it should be noted that the influence of the prefrontal cortico-striato-pallidal system on the dopaminer- gic system is not a ‘simple’ summation of excitatory and inhibitory inputs but that this influence might be ‘translated’ in changes in firing patterns of populations of dopaminergic neurons. Whereas the above-given description is primarily based on data obtained in rats, similar arrangements of projections from the ventral striatum, via the dopaminergic system, to the dorsal striatum have been shown to exist in primates. However, in the latter species such interactions might be more elaborate, including various intermediairy steps back and forth between striatum and substantia nigra [45,82].

H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14 Sll
5. Functional aspects of the dorsal and ventral striatum
To date, the functions of the basal ganglia are not completely understood, but it is clear that such functions should be considered, at least in part, in the context of their inclusion in the thahuno-cortical circuitry. In rather general terms a characterization of basal ganglia functions is that these structures ‘assist’ the (pre)frontal cortex in selecting appropriate motor and behavioral output in a particular context [36,38,39]. The addition ‘in a
particular context’ signifies the crucial role of the basal ganglia in the integration of sensory (and mnemonic = past experience) aspects with motor and behavioral expression. This integration of perceptive and executive functions has, within the basal ganglia, its neuronal substrate most clearly at the level of the striatum, i.e., in the convergence of corticostriatal projections originating from cerebral cortical areas with, respectively, sensory and motor functions.
Two main aspects of the ‘behavioral selection’ function of the basal ganglia should be considered. First, the basal ganglia play an important role in the adaptive processes that lead to new motor and behavioral outputs, i.e., in the phase of learning new behaviors. Second, the basal ganglia maintain a role in selecting a specific output in a particular context even when learned behaviors have become routine. As to the first aspect, it has been well-established that the striatum and the dopaminergic system are crucial elements in the associative processes that underlie adaptive behavioral learning and selection [83,84]. In this learning process, dopamine signals the occurrence of novel stimuli that may be of significance for the organism, as well as the presentation of rewarding stimuli or stimuli that predict upcoming rewarding events [85]. In other words, the basal ganglia are thought to play a role in predicting the (positive) outcome of the selection of a certain behavioral output in a particular situation and, in this way, the basal ganglia help to acquire and guide adaptive behavior. In general, these processes are characterized as a procedural learning process that leads to implicit memory for contextually appropriate behavior. Well-established, routine behavioral procedures or motor sequences are indicated as ‘habits’. Indeed, habit formation has been attributed to the basal ganglia since many years [84,86].
In association with the above-described functionally segregated basal ganglia-thalamocortical circuits, different sectors of the striatum are involved in different aspects of behavioral learning (Fig. 3). It has been clearly shown that the ventral striatum is involved in various aspects of associative, reward-based learning processes underlying the formation of complex behavioral sequences. Thus, through its connections with limbic structures like the amygdala and hippocampus, the ventral striatum ‘translates’ stimulus- reward associations into a behavioral output [87]. A crucial role in this process is being played by the mesolimbic dopamine system. Furthermore, there appears to be a differential involvement of the shell and core subregions
of the nucleus accumbens in these processes (review: Ref. 88). The shell mediates the motivational significance of unconditioned stimuli on behavior and is primarily involved in unlearned behaviors. In contrast, the core is important for the mediation of conditioned stimuli and forms a crucial link in the expression of complex, learned (instrumental) behaviors (cf. Ref. 88). The dorsal striatum appears to become involved when, through overtraining, behaviors become inflexible and a particular stimulus automatically leads to a specific response, i.e., when motor responses and behaviors have become habits.
In the context of neural mechanisms that underlie the development of drug addiction and drug-seeking behavior, it has been suggested that different sectors of the striatum are involved in different phases of the development of the addictive behavior. In early phases, interactions between the prefrontal cortex, the amygdala, the ventral striatum and the mesolimbic dopamine system are included, whereas in a later phase when drug-taking has become a habit, the caudate-putamen is involved as well [89]. In those later phases, even after longer periods of abstinence, confrontation of the addict with stimuli that are associated with drug-intake, may lead to a relapse. In the transition from the early phases of addictive behavior to the later, habitual phases, the above-described interactions between different parts of the striatum via the dopaminergic system (the spirals of Haber et al. [45]) are thought to play a role (e.g. Ref. 89). Although it might be argued that this shift in the involvement of functionally different prefrontal corticostriatal circuits is related to maladaptive behaviors such as drug abuse, it is tempting to speculate that similar mechanisms play a role in normal, adaptive behavior related to classical conditioning in various situations.
6. Does the ventral striatum play a role in Tourette’s syndrome?
As indicated in the introduction, the basal ganglia are thought to play an important role in the development and symptomatology of Tourette’s syndrome (TS). Not only differences in the structure of basal ganglia nuclei, but also functional differences between TS patients and controls form the basis of this hypothesis. In particular the dorsal striatum in association with the prefrontal cortex, a system that is involved in the above-discussed habit formation, has been the main focus of attention [26,90]. However, also cortical and subcortical structures involved in limbic-related basal ganglia-thalamocortical circuits have been shown or hypothesized to be involved in TS (e.g. Ref. 22 and 91).
What could be the role for the ventral striatum and associated cortical areas in tic disorders? This role might be hypothesized in two different phases, i.e., 1. the striatal learning phase that follows the transition from conditioned, instrumental learning to habit formation, and 2. the phase during which habits have been firmly laid down and must

s12 H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14
be executed in the appropriate context. First, as discussed in the previous paragraphs, the ventral striatum might in- fluence the formation of habitual motor and behavioral processes by way of its influence on the dopaminergic in- nervation of the dorsal striatum. If during development (i.e., largely during childhood) the interplay between the moti- vational, cognitive and sensorimotor circuits is disturbed, it might be hypothesized that this leads to an aberrant habit formation. This could be interpreted as an imbalance be- tween the dorsal and ventral cortical-basal ganglia circuits and might underlie the ultimate symptomatology of tics. Thus, as a result of this disturbed development, fragments of movements, and/or cognitive or emotional processes are expressed in an inappropriate context and in an aberrant frequency. In other words, the role of the basal ganglia in focussed attention and the selection of appropriate motor or behavioral acts is disturbed [38].
Second, the ventral striatal system, together with its related prefrontal cortical areas like the orbitofrontal cortex, might influence the expression of habits, once they have been firmly established. Again, it is possible that this oc- curs through the influence of the ventral striatum on the dopaminergic neurons that project to the dorsal striatum. For example, the prefrontal-ventral striatal circuitry might be important at the moment the TS patient feels the urge to tic but consciously suppresses the expression of the tics. This requires a highly motivated brain action of which it is very conceivable that it includes the mesolimbic dopamine system as well as the orbitofrontal cortex and ventral stria- mm. The emotional tension that is felt with the suppression of the tics could be associated with the activity in this circuitry. Thus, dopaminergic mechanisms at the level of both the mesolimbic and the nigrostriatal dopamine systems would be involved in these processes. This might explain why in a functional-MRI study active tic suppression is associated with activation of ventral parts of the basal ganglia and orbitofrontal, limbic-related cortical areas [22].
Finally, if indeed an imbalance between striosome/patch and matrix compartments is part of the pathophysiol- ogy of tic disorders [60], the specific innervation of the striosome/patch compartment might explain the sensitivity of tic expression to stressful and emotional situations. The striosome/patch compartment is specifically innervated by the paraventricular thahtmic nucleus, a key area in mediat- ing stress and arousal [55,92], as well as particular subnu- clei of the amygdaloid complex [56,57]. This compartment also receives specific inputs from medial prefrontal areas and/or particular layers of the prefrontal cortex [52-541. These inputs to the patch/striosome compartment are all excitatory and could influence the balance between the two striatal compartments. This could lead to a higher output of the surrounding matrix which may be accompanied by the release of motor or behavioral programs. In a disturbed striatum, as in TS patients, this may lead to a higher expression of tics. Whether such mechanisms indeed play a role, remains to be established.
References
[l] Tour&e Syndrome Classification Study Group. Definitions and classification of tic disorders. Arch Nemo1 1993;50:1013-16.
[2] Kurlan R. Tourette’s syndrome. In: Watts RL, Koller WC, editors. Movement Disorders: Neurologic Principles and Practice. New York, NY: McGraw-Hill, 1997. p. 569-7.5.
[3] Kushner HI. A Cursing Brain? The Histories of Tourette Syn- drome. London: Harvard University Press, 1999.
[4] Swerdlow NR, Zinner S, Farber RH, Seacrist C, Hartston H. Symp- toms in obsessive-compulsive disorder and Tourette syndrome: a spectrum? CNS Spectrums 1999;4:21-33.
[5] Robertson MM, Kurlan R, Cohen DJ, Leckman JF, McMahon W, Pauls DL, Sandor P, van de Wetering BJ. The Tourette syndrome diagnostic confidence index: development and clinical associations. Neurology 1999;53:2108-12.
[6] Leckman JF, Walker DE, Cohen DJ. Premonitory urges in Tourette’s syndrome. Am .I Psychiatry 1993;150:98-102.
[7] Scahill LD, Leckman JF, Marek KL. Sensory phenomena in Tourette’s syndrome. Adv Nemo1 1995;65:273-80.
[8] Comings DE, Comings BG. Tourette’s syndrome and attention deficit disorder with hyperactivity: are they genetically related? J Am Acad Child Psychiatry 1984;23: 138-46.
[9] Frankel M, Cummings JL, Robertson MM, Trimble MR, Hill MA, Benson DE Obsessions and compulsions in Gilles de la Tourette’s syndrome. Neurology 1986;36:378-82.
[lo] Pauls DL, Towbin KE, Leckman JF, Zahner GE, Cohen DJ. Gilles de la Tourette’s syndrome and obsessive-compulsive disorder. Ev- idence supporting a genetic relationship. Arch Gen Psychiatry 1986;43:1180-2.
[ll] Pauls DL, Raymond CL, Stevenson JM, Leckman JF. A fam- ily study of Gilles de la Tourette syndrome. Am J Hum Genet 1991;48:154-63.
[12] Pauls DL, Leckman JF, Cohen DJ. Familial relationship between Gilles de la Tourette’s syndrome, attention deficit disorder, learning disabilities, speech disorders, and stuttering. J Am Acad Child Adolesc Psychiatry 1993;32: 1044-50.
[ 131 Como PG. Obsessive-compulsive disorder in Tourette’s syndrome. Adv Nemo1 1995;65:281-91.
[14] Comings DE. Clinical and molecular genetics of ADHD and Tourette syndrome. Two related polygenic disorders. Ann N Y Acad Sci 2001;931:50-83.
[15] DSM-IV task force. Diagnostic and statisical manual of mental disorders, fourth edition. In: Frances A, Pincus HA, First MB, editors. Washington DC: American Psychiatric Association, 1994. p. 100-5.
[ 161 Mink JW. Basal ganglia dysfunction in Tourette’s syndrome: a new hypothesis. Pediatr Nemo1 2001;25:190-8.
[17] Palumbo D, Maughan A, Kurlan R. Hypothesis III. Tourette syn- drome is only one of several causes of a developmental basal ganglia syndrome. Arch Nemo1 1997;54:475-83.
[18] Wright CI, Bradley S, Peterson MD, Rauch SL. Neuroimaging studies in Tourette syndrome. CNS Spectrums 1999;4:54-60.
[19] Rauch SL, Whalen PJ, Curran T, Shin LM, Coffey BJ, Savage CR, McInerney SC, Baer L, Jenike MA. Probing striate-thalamic function in obsessive-compulsive disorder and Tourette syndrome using neuroimaging methods. Adv Nemo1 2001; 85:207-24.
[20] Peterson BS, Riddle MA, Cohen DJ. Reduced basal ganglia volumes in Tourette’s syndrome using three-dimensional recon- struction techniques from magnetic resonance images. Neurology 1993;43:941-9.
[21] Singer HS,. Volumetric MRI changes in basal ganglia of children with Tourette’s syndrome. Neurology 1993;43:950-6.
[22] Peterson BS, Skudlarski P, Anderson AW, Zhang H, Gatenby JC, Lacadie CM, Leckman JF, Gore JC. A functional magnetic resonance imaging study of tic suppression in Tourette syndrome. Arch Gen Psychiatry 1998;55:326-33.

H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14 s13
[23] Ernst M, Zametkin AJ; Jons PH; Matochik JA; Pascualvaca D; Cohen RM. High presynaptic dopaminergic activity in children with Tourette’s disorder. .I Am Acad Child Adolesc Psychiatry 1999;38:86-94.
[24] Singer HS, Szymanski S, Giuliano J, Yokoi F, Dogan AS, Brasic JR, Zhou Y, Grace AA, Wong DE Elevated intrasynaptic dopamine release in Tourette’s syndrome measured by PET. Am J Psychiatry 2002;159:1329-36.
[25] Rauch SL, Savage CR. Neuroimaging and neuropsychology of the striatum. Bridging basic science and clinical practice. Psychiatr Clin North Am 1997;20:741-68.
[26] Graybiel AM, Rauch SL. Toward a neurobiology of obsessive- compulsive disorder. Neuron 2000;28: 343-7.
[27] Stern E, Silbersweig DA, Chee KY, Holmes A, Robertson MM, Trimble M, Frith CD, Frackowiak RS, Dolan RJ. A functional neuroanatomy of tics in Tourette syndrome. Arch Gen Psychiatry 2000; 57:741-8.
[28] Peterson BS, Thomas P, Kane MJ, Scahill L, Zhang H, Bronen R, King RA, Leckman JF, Staib L.. Basal Ganglia volumes in patients with Gilles de la Tourette syndrome. Arch Gen Psychiatry 2003;60:415-24.
[29] Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci 1986;9:357-81.
[30] Groenewegen HJ, Berendse HW, Wolters JG, Lohman AH. The
[31
~32
anatomical relationship of the prefrontal cortex with the striatopall- idal system, the thalamus and the amygdala: evidence for a parallel organization. Prog Brain Res 1990;85:95-116.
] Alexander GE, Crutcher MD, DeLong MR. Basal ganglia- thalamocortical circuits: parallel substrates for motor, oculomotor, ‘prefrontal’ and ‘limbic’ functions. Prog Brain Res 1990;85:119- 46.
,] Parent A, Hazrati LN. Functional anatomy of the basal ganglia. I. The cortico-basal ganglia-thalamo-cortical loop. Brain Res Brain Res Rev 1995;20:91-127.
[33] Selemon LD, Goldman-Rakic PS. Longitudinal topography and interdigitation of corticostriatal projections in the rhesus monkey. J Neurosci 1985;5:77694.
[34] Flaherty AW, Graybiel AM. Two input systems for body represen- tations in the primate striatal matrix: experimental evidence in the squirrel monkey. J Neurosci 1993;13:1120-37.
[35] Gerfen CR. The neostriatal mosaic: multiple levels of compar- mental organization in the basal ganglia. Annu Rev Neurosci 1992;15:285-320.
[36] Wise SP, Murray EA, Gerfen CR. The frontal cortex-basal ganglia system in primates. Crit Rev Neurobiol 1996;10:317-56.
[37] Bolam JP, Hanley JJ, Booth PA, Bevan MD. Synaptic organisation of the basal ganglia. J Anat 2000;196:527-42.
[38] Mink JW. The basal ganglia: focused selection and inhibition of competing motor programs. Prog Neurobiol 1996;50:381-425.
[39] Redgrave P, Prescott TJ, Gurney K. The basal ganglia: a vertebrate solution to the selection problem? Neuroscience 1999;89: 1009-23.
[40] Houk JC, Adams JL, Barto AG. A model how the basal ganglia generate and use neural signals that predict reinforcement. In: Houk JC, Davis IL, Beiser DG, editors. Models of Information Processing in the Basal Ganglia. Cambridge, MA: MIT Press, 1995. p. 117-30.
] Haber SN, Kunishio K, Mizobuchi M, Lynd-Balta E. The orbital [41 and medial prefrontal circuit through the primate basal ganglia. J Neurosci 1995;15:4851-67.
~42 ,] Zahm DS, Brog JS. On the significance of subterritories in the ‘accumbens’ part of the rat ventral striatum. Neuroscience 1992; 50:751-67.
[43] Groenewegen HJ, Berendse HW. Anatomical relationships between the prefrontal cortex and the basal ganglia in the rat. In: Thierry AM, Glowinski J, Goldman-Rakic P, Christen Y, editors. Motor and Cognitive Functions of the Prefrontal Cortex. Berlin: Springer Verlag, 1994. p. 51-77.
[44] Joel D, Weiner I. The organization of the basal ganglia- thalamocortical circuits: open interconnected rather than closed segregated. Neuroscience 1994;63:363-79.
[45] Haber SN, Fudge JL, McFarland NR. Striatonigrostriatal path- ways in primates form an ascending spiral from the shell to the dorsolateral striatum. J Neurosci 2000;20:2369-82.
[46] Graybiel AM, Ragsdale CW, Jr. Histochemically distinct compar- merits in the striatum of human, monkeys, and cat demonstrated by acetylthiocholinesterase staining. Proc Nat1 Acad Sci U S A 1978;75:5723-6.
[47] Gerfen CR, Baimbridge KG, Miller JJ. The neostriatal mosaic: compartmental distribution of calcium-binding protein and parval- bumin in the basal ganglia of the rat and monkey. Proc Nat1 Acad Sci U S A 1985;82:8780-4.
[48] Graybiel AM. Neurotransmitters and neuromodulators in the basal ganglia. Trends Neurosci 1990;13:244-54.
[49] Van der Kooy D, Fishell G. Neuronal birthdate underlies the development of striatal compartments. Brain Res 1987;401:155- 61.
[50] Johnston JG, Gerfen CR, Haber SN, van der Kooy D. Mecha- nisms of striatal pattern formation: conservation of mammalian compartmentalization. Brain Res Dev Brain Res 1990;57:93-102.
[51] Holt DJ, Hersh LB, Saper CB. Cholinergic innervation in the human striatum: a three-compartment model. Neuroscience 1996; 74~67-87.
[52] Eblen F, Graybiel AM. Highly restricted origin of prefrontal cor- tical inputs to striosomes in the macaque monkey. J Neurosci 1995;15:5999-6013.
[53] Gerfen CR. The neostriatal mosaic: striatal patch-matrix organiza- tion is related to cortical lamination. Science 1989;246:385-8.
[54] Berendse HW, Galis-de Graaf Y, Groenewegen HJ. Topographical organization and relationship with ventral striatal compartments of prefrontal corticostriatal projections in the rat. J Comp Nemo1 1992;316:314-47.
[55] Berendse HW, Groenewegen HJ. Organization of the thalamostr- atal projections in the rat, with special emphasis on the ventral striatum. J Comp Nemo1 1990; 299:187-228.
[56] Ragsdale CW, Jr., Graybiel AM. Fibers from the basolateral nu- cleus of the amygdala selectively innervate striosomes in the cau- date nucleus of the cat. J Comp Nemo1 1988; 269:506-22.
[57] Wright CI, Beijer AV, Groenewegen HJ. Basal amygdaloid com- plex afferents to the rat nucleus accumbens are compartmentally organized. J Neurosci 1996;16:1877-93.
[58] Gerfen CR, Herkenham M, Thibault J. The neostriatal mosaic: II. Patch- and matrix-directed mesostriatal dopaminergic and non- dopaminergic systems. J Neurosci 1987;7:3915-34.
[59] Berendse HW, Groenewegen HJ, Lohman AH. Compartmental distribution of ventral striatal nem-ens projecting to the mesen- cephalon in the rat. J Neurosci 1992;12:2079-103.
[60] Canales JJ, Graybiel AM. A measure of striatal function predicts motor stereotypy. Nat Neurosci 2000;3:377-83.
[61] Meredith GE, Pattiselanno A, Groenewegen HJ, Haber SN. Shell and core in monkey and human nucleus accumbens identified with antibodies to calbindin-D28k. J Comp Nemo1 1996;365:628-39.
[62] Groenewegen HJ, Wright CI, Beijer AV. The nucleus accumbens: gateway for limbic structures to reach the motor system? Prog Brain Res 1996;107:485-511.
[63] Groenewegen HJ, Wright CI, Beijer AV, Voorn l? Convergence and segregation of ventral striatal inputs and outputs. Ann N Y Acad Sci 1999;877:49-63.
[64] Zahm DS. Functional-anatomical implications of the nucleus ac- cumbens core and shell subterritories. Ann N Y Acad Sci 1999;877:113-28.
[65] Voorn P, Gerfen CR, Groenewegen HJ. Compartmental organiza- tion of the ventral striatum of the rat: immunohistochemical distr- bution of enkephalin, substance P, dopamine, and calcium-binding protein. J Comp Nemo1 1989;289:189-201.

s14 H.J. Groenewegen et al. /Brain & Development 25 Suppl. I (2003) S3-S14
[66] Voom P, Brady LS, Berendse HW, Richfield EK. Densitometical analysis of opioid receptor ligand binding in the human striatum - I. Distribution of mu opioid receptor defines shell and core of the ventral striatum. Neuroscience 1996;75:777-92.
[67] Fudge JL, Kunishio K, Walsh P, Richard C, Haber SN. Amyg- daloid projections to ventromedial striatal subterritories in the primate. Neuroscience 2002;110:257-7.5.
[68] Berridge CW, Stratford TL, Foote SL, Kelley AE. Distribution of dopamine beta-hydroxylase-like immunoreactive fibers within the shell subregion of the nucleus accumbens. Synapse 1997;27:230- 41.
[69] Zahm DS. Compartments in rat dorsal and ventral striatum re- vealed following injection of 6-hydroxydopamine into the ventral mesencephalon. Brain Res 1991;552:164-9.
[70] Nauta WJ, Smith GP, Faull RL, Domesick VB. Efferent connec- tions and nigral afferents of the nucleus accumbens septi in the rat. Neuroscience 1978;3:385-401.
[71] Deniau JM, Menetrey A, Thierry AM. Indirect nucleus accumbens input to the prefrontal cortex via the substantia nigra pars reticu- lata: a combined anatomical and electrophysiological study in the rat. Neuroscience 1994;61:533-45.
[72] Berendse HW, Richfield EK. Heterogeneous distribution of dopamine Dl and D2 receptors in the human ventral striatum. Neurosci Lett 1993;150:75-9.
[73] Heimer L, de Olmos JS, Alheid GF, Pearson .I, Sakamoto N, Shinoda K, Marksteiner J, Switser III R.C.. The human basal forebrain. Part II. In: Bloom FE, Bj rklund A, H kfelt T, editors. Handbook of Chemical Neuroanatomy. 1999;15:57-225.
[74] Pennartz CM, Groenewegen HJ, Lopes da Silva FH. The nu- cleus accumbens as a complex of functionally distinct neuronal ensembles: an integration of behavioural, electrophysiological and anatomical data. Prog Neurobiol 1994;42:719-61.
[75] O’Donnell P, Grace AA. Synaptic interactions among excitatory afferents to nucleus accumbens neurons: hippocampal gating of prefrontal cortical input. J Neurosci 1995;15:3622-39.
[76] O’Donnell l? Dopamine gating of forebrain neural ensembles. Em- J Neurosci 2003; 17:429-35.
[77] McFarland NR, Haber SN. Thalamic relay nuclei of the basal gan- glia form both reciprocal and nonreciprocal cortical connections, linking multiple frontal cortical areas. J Neurosci 2002;22:8117- 32.
[78] Van Dongen YC, Groenewegen HJ. Core and shell of the nucleus accumbens are interconnected via intrastriatal projections. An an- terograde tracing study in rats. In: Nicholson L, Faull RL, editors.
The Basal Ganglia VII. New York, NY: Kluwer Academic/ Plenum, 2002.
[79] Groenewegen HJ, Berendse HW, Wouterlood FG. Organization of the projections form the ventral striatopallidal system to ventral mesencephalic dopaminergic neurons. In: Percheron G, McKenzie JS, editors. The Basal Ganglia IV. New York, NY: Plenum Press, 1994. p. 81-93.
[80] Kolomiets B, van Dongen YC, Thierry AM, Glowinski J, Groe- newegen HJ, Deniau JM. Mesencephalic dopaminergic nem-ens as an interface between the ventral and dorsal striatum: anatomical and electrophysiological evidence. Em- J Neurosci 2002. Abstract
[81] Sesack SR, Deutch AY, Roth RH, Bunney BS. Topographical organization of the efferent projections of the medial prefrontal cortex in the rat: an anterograde tract-tracing study with Phaseolus vulgaris leucoagglutinin. J Comp Nemo1 1989;290:213-42.
[82] Fran ois C, Yelnik J, Tand D, Agid Y, Hirsch EC. Dopaminer- gic cell group A8 in the monkey: Anatomical organization and projections to the striatum. J Comp Nemo1 1999;414:334-347.
[83] Hikosaka 0. Role of basal ganglia in control of innate movements, learned behavior and cognition - a hypothesis. In: Percheron G, McKenzie JS, editors. The Basal Ganglia IV. New York, NY: Plenum Press, 1994. p. 589-96.
[84] Packard MG, Knowlton BJ. Learning and memory functions of the Basal Ganglia. Annu Rev Neurosci 2002;25:563-93.
[85] Schultz W. Getting formal with dopamine and reward. Neuron 2002;36:241-63.
[86] Bachevalier J, Mishkin M. An early and a late developing system for learning and retention in infant monkeys. Behav Neurosci 1984;98:770-8.
[87] Robbins TW, Everitt BJ. Neurobehavioural mechanisms of reward and motivation. Cur Opin Neurobiol 1996;6:228-36.
[88] Cardinal RN, Parkinson JA, Hall J, Everitt BJ. Emotion and mo- tivation: the role of the amygdala, ventral striatum, and prefrontal cortex. Neurosci Biobehav Rev 2002;26:321-52.
[89] Everitt BJ, Wolf ME. Psychomotor stimulant addiction: a neural systems perspective. J Neurosci 2002;22:3312-20.
[90] Leckman JF, Riddle MA. Tourette’s syndrome: when habit-forming systems form habits of their own? Neuron 2000;28:349-54.
[91] Brito GN. A neurobiological model for Tourette syndrome centered on the nucleus accumbens. Med Hypotheses 1997;49:133-42.
[92] Van der Werf YD, Witter MP, Groenewegen HJ. The intralaminar and midline nuclei of the thalamus. Anatomical and functional evidence for participation in processes of arousal and awareness. Brain Res Brain Res Rev 2002;39: 107-40.
Copyright © 2022 FDOKUMEN




















