
Gabaergic regulation of the neural organization of fear in the midbrain tectum

Uz
U
A
D2
nA
GiitwicFribse
f
2EAGpcihSr
Neuroscience 196 (2011) 131–146
ENHANCED GABAERGIC TONE IN THE VENTRAL PALLIDUM:
MEMORY OF UNPLEASANT EXPERIENCES?stp
M. SKIRZEWSKI,a,b* W. LÓPEZ,a E. MOSQUERA,a
L. BETANCOURT,b,c B. CATLOW,d M. CHIURILLO,e
N. LOUREIRO,a L. HERNÁNDEZb AND P. RADAb
aLaboratorio de Fisiología Celular, Decanato de Ciencias de la Salud,niversidad Centroccidental Lisandro Alvarado, Barquisimeto, Vene-uela, 3001-A
bLaboratorio de Fisiología de la Conducta, Facultad de Medicina,niversidad de Los Andes, Mérida, Venezuela, 5101-A
cLaboratorio de Histología, Facultad de Medicina, Universidad de Losndes, Mérida, Venezuela, 5101-A
dLaboratory of Molecular Biology, National Institute of Neurologicalisorders and Stroke, National Institutes of Health, Bethesda, MD0892, USA
eLaboratorio de Genética Molecular “Dr. Jorge Yunis-Turbay,” Deca-ato de Ciencias de la Salud, Universidad Centroccidental Lisandrolvarado, Barquisimeto, Venezuela, 3001-A
Abstract—The nucleus accumbens (NAc) has emerged as animportant part of the neural circuitry regulating depressive-like behaviors. Given that the NAc GABAergic medium spinyneurons project to the ventral pallidum (VP), it is reasonableto suggest that the VP may also be involved in these behav-iors. Consequently, we explored the role of the VP GABAer-gic terminals during depressive-like behaviors in rats usingthe forced swim test (FST) and the sucrose preference test(SPT). Microdialysis coupled with micellar electrokineticchromatography was used to monitor in vivo changes of
ABA in the VP during the FST. GABA levels significantlyncreased during day-1 and day-2 during swimming, return-ng to the pre-swimming levels after the test. Basal concen-rations of GABA on day-2 of the FST significantly increasedith respect to day-1. In another set of experiments, intra-VP
njections of vigabatrin (a GABA transaminase inhibitor) in-reased extracellular GABA and immobility behaviors in theST while the direct GABAA receptor antagonist bicucullineeduced immobility behaviors. In the SPT, intra-VP vigabatrinnjection significantly reduced preference for sucrose whileicuculline did not produce any change. At the postsynapticide, we used semiquantitative RT-PCR to measure mRNAxpression of 17 GABAA receptor subunits (�1–�6, �1–�3,
�2, �, �, �, �, and �1–�3) in rats subjected to the FST. Weound a significant reduction of �3 and �2 subunit expression
and an increase of � subunit expression after day-2 in rats
*Correspondence to: M. Skirzewski, Universidad CentroccidentalLisandro Alvarado, Laboratorio de Fisiología Celular, Decanato deCiencias de la Salud, Av. Libertador con Av. Andrés Bello, Barqui-simeto 3001, Venezuela. Tel: �58-251-2592049; fax: �58-251-591809.-mail address: [email protected] (M. Skirzewski).bbreviations: BSA, bovine serum albumin; FST, forced swim test;ABAA-R, GABA receptor type-A; GAPDH, glyceraldehyde-3-phos-hate dehydrogenase; IVIC, instituto venezolano de investigacionesientíficas; MEKC-LIFD, micellar electrokinetic chromatography, laser-nduced fluorescence detection; NAc, nucleus accumbens; PH, peakeight; p.s.i., pounds per square inch; SPT, sucrose preference test;
Q-RT-PCR, semiquantitative reverse transcription-polymerase chaineaction; TP, mild tail-pinch; VP, ventral pallidum.
0306-4522/11 $ - see front matter © 2011 IBRO. Published by Elsevier Ltd. All righdoi:10.1016/j.neuroscience.2011.08.058
131
ubject to the FST which might enhance tonic inhibition ofhe VP. Furthermore, immunoblot experiments revealed thatrotein expression of �2 and � subunits changed 6 days after
FST in a way similar to mRNA expression. These resultssuggest that the enhanced VP-GABAergic tone might triggera low motivational state, anhedonia and a possible memorymechanism for unpleasant experiences. © 2011 IBRO. Pub-lished by Elsevier Ltd. All rights reserved.
Key words: GABA, vigabatrin, GABA-A receptor, ventral pal-lidum, forced swim test, sucrose preference test.
Major depressive disorder in humans is characterized bydepressed mood, loss of interest or unpleasant feelings fordaily activities (anhedonia), altered sleep behavior, low en-ergy, psychomotor agitation or retardation activity, alteredappetite, and helplessness (DSMIV, 2000). Additionally, hu-man generalized anxiety disorders are characterized by feel-ings of wound-up, tension or restlessness, fatigue, concen-tration difficulty, irritability, significant muscular tension, andaltered sleep (DSMIV, 2000), and they are highly comorbidwith depression (Kessler et al., 2003). Several studies inhumans and in rodents have demonstrated that the prefrontaland cingulate cortex, hippocampus, striatum, amygdala, andthe thalamus are related to depression-like behaviors (Archeret al., 1984; MacLean, 1985; Berridge, 1996; Davidson et al.,1999; McFarland and Kalivas, 2001; Brambilla et al., 2003;Farrar et al., 2008). The nucleus accumbens (NAc) alsoseems to play a key role in depressive-like behaviors inrodents (Pliakas et al., 2001; Carlezon et al., 2005; Chau etal., 2011), probably through an abnormal function of theglutamatergic, cholinergic, and/or adenosinergic systems(Chau et al., 1999, 2001, 2011; Rada et al., 2003, 2006;Mingote et al., 2008). The NAc projects through the medialforebrain bundle to the ventral pallidum (VP) almost exclu-sively by GABAergic neurons (medium spiny neurons), sug-gesting that this nucleus and the GABA system could beinvolved in depressive-like behaviors and/or anxiety disor-ders (Walaas and Fonnum, 1979; Groenewegen and Russ-chen, 1984; MacLean, 1985; O’Donnell et al., 1997). Thepossible relationship between the GABAergic innervation ofthe VP with depressive-like behaviors and/or anxiety mayadd evidence to the GABAergic hypothesis of mood disor-ders as other authors have suggested occurs in the prefrontalcortex and the hippocampus (Lloyd et al., 1989; Brambilla etal., 2003).
The VP–GABAergic system has been linked to thecontrol of amphetamine-induced locomotor activity (Bour-delais and Kalivas, 1990; Hooks and Kalivas, 1995;Fletcher et al., 1998; Wisniecki et al., 2003), and more
recently with feeding, satiety, reward, and aversive behav-ts reserved.
eotUltaccuaG
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146132
iors (Stratford et al., 1999; Pérez et al., 2000; Smith andBerridge, 2005; Shimura et al., 2006; Inui et al., 2007;Farrar et al., 2008). In general, previous reports suggestthat enjoyable or hedonic activities reduce extracellularGABA release in the VP, whereas aversive or unwantedactivities increase it (Bourdelais and Kalivas, 1990, 1992a;Torregrossa et al., 2008; Inui et al., 2009). Moreover, theuse of drugs that modulate the activity of GABAergic re-ceptors, such as vigabatrin (an irreversible GABA-transaminase inhibitor) or bicuculline (competitive antago-nist of GABAA-R), have been a valuable pharmacologicalstrategy to establish the relationship of the GABAergicsystem with behavior (Jung et al., 1977a,b; Schechter etal., 1977; Lloyd et al., 1989). In spite of this tantalizinghypothesis, we found no previous reports associating di-rectly the GABAergic activity in the VP with depressive-likebehaviors and helplessness in rodents.
Major depressive disorder is difficult to study becauseno concurrent abnormality has proven to be sufficientlyrobust or consistent enough in humans to validate ananimal model in rodents (Krishnan and Nestler, 2008; Nes-tler and Hyman, 2010). However, the forced swim test(FST) has been suitable to evaluate antidepressant drugefficacy (Lucki, 1997; Chau et al., 1999, 2011; Skirzewskiet al., 2010) and to understand the neurochemical andneurobiological phenomena underlying this neuropsychiat-ric disorder (Chau et al., 2001; Yoshikawa et al., 2002;Gomez et al., 2003; Rada et al., 2003, 2006; Mombereauet al., 2004). Basically, the FST evaluates escape motiva-tional behaviors in rodents exposed to an inescapablewater tank experience (Porsolt, 1979; Nestler and Hyman,2010). Although this behavioral test models depressive-like behavior, several features differ from human depres-sion and extrapolation to the human disorder should betaken cautiously (Nestler and Hyman, 2010). Several au-thors have suggested that more than one animal modelshould be employed over the same hypothesis tostrengthen the conclusions reached by any single model ofstudy (Krishnan and Nestler, 2008; Nestler and Hyman,2010). The sucrose preference test (SPT) is another par-adigm that evaluates the animal’s interest in pleasurableactivities, by testing preference for a sucrose solution overwater. A reduced preference for the sucrose solution isinterpreted as anhedonia, a core symptom of the depres-sion in humans (Nestler and Hyman, 2010).
The mild tail-pinch test is an animal model that hasserved to study anxiety disorders and to assess for theimpact of anxiety-related behaviors on the FST. During thistest no signs of pain in the rats are generated. This animalmodel was originally employed to elucidate the neurobio-logical basis of stress-inducing eating behaviors in rodents(Rowland and Antelman, 1976; Levine and Morley, 1981)and more recently to study the relationship between anxi-ety–depression behaviors (Di Chiara et al., 1999; Rada etal., 2003, 2006).
Considering the aforementioned data, it is plausible tosuggest that depressive-like behavior, an emotional disordercharacterized by a generalized aversive and anhedonic state
(Carlezon and Chartoff, 2007; Bechtholt-Gompf et al., 2010),could produce an increase in the release of extracellularGABA in the VP, as has been demonstrated in other aversivebehavioral processes (Inui et al., 2007, 2009). Therefore,direct intra-VP administration of drugs, which enhanceextracellular GABA levels, such as vigabatrin, should po-tentiate depressive-like behavior, whereas antagonizingthe effect of GABA through bicuculline should block theVP-circuit that triggers this behavior. To verify this hypoth-esis, we monitored extracellular GABA levels in the VPbefore, during, and after the FST in microdialysis samplescollected every minute. In a parallel experiment, we mea-sured GABA in an independent group of rats subject to themild tail-pinch test (TP), a behavioral rat model of anxiety.We also evaluated the effect of a 20-min vigabatrin pulseover the release of extracellular intra VP GABA releasebefore, during, and after the FST. Then, we evaluated theeffects of intra-VP injections of vigabatrin or bicuculline onimmobility behaviors during the FST, anhedonia with theSPT, and locomotor activity in the open field test. In asecond set of experiments, we evaluated whether possiblechanges in GABA transmission in VP after FST or TP canaffect the relative mRNA expression of 17 GABAA-R sub-units (�1, �2, �3, �4, �5, �6, �1, �2, �3, �2, �, �, �, �, �1,�2, and �3), and protein expression of �3, �2, and �subunits in the VP by semiquantitative RT-PCR and im-munoblot, respectively, in rats.
EXPERIMENTAL PROCEDURES
Subjects
Male Sprague–Dawley rats obtained from the University Centroc-cidental Lisandro Alvarado and Universidad de los Andes bioteriaand weighing between 300–450 g were housed in pairs with foodand water ad libitum in a 12:12 h light–dark cycle at 22 °C. Allxperiments were conducted in accordance with the Declarationf Helsinki, followed the National Institutes of Health guidelines forhe care and use of laboratory animals and were approved by theniversity Centroccidental Lisandro Alvarado and Universidad de
os Andes bioethics committees. All efforts were made to minimizehe number of animals and to spare them suffering. For this report,
total of 35 rats were intra-VP microdialyzed to measure extra-ellular GABA levels, 122 rats were intra-VP intracerebrally mi-roinjected with saline, vigabatrin, or bicuculline, 90 rats weresed to measure transcript levels of 17 GABAA-R subunits in VP,nd 90 rats were employed to measure protein expression of threeABAA-R subunits in VP.
Surgery
A Stellar stereotaxic instrument (Stoelting, IL, USA) was used tochronically implant guide shafts aimed at the VP in rats anesthe-tized with a combination of (Calox International, C.A; Argentina)(50 mg/kg i.p.) and (Laboratories Calier, S.A; Barcelona, Spain)(10 mg/kg i.p.). For intracerebral microdialysis, unilateral 10 mm-long, 21-ga guide shafts aimed at the VP (AP: 0.0 mm, L: 2.4 mm,and V: 4.0 mm with respect to bregma, midsaggital sinus, andbrain surface) were implanted. Guide shafts were implanted aim-ing above the VP of the left or right hemispheres in a counterbal-anced manner. For intracerebral microinjections, bilateral 15 mm-long, 26-ga guide shafts aimed at the VP (AP: 0.0 mm, L: 2.4 mm,and V: 5.8 mm with respect to bregma, midsaggital sinus, andbrain surface) (Paxinos and Watson, 2005) were implanted. Also,an anatomical negative control group of animals was prepared for
microinjections at 2.5 mm above VP using the following stereo-
osrsa
st5sbsItdurb
fs
tir
oapm(
coa(
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146 133
taxic coordinates for the guide shafts: AP: 0.0 mm, L: 2.4 mm, andV: 3.3 mm with respect to bregma, midsaggital sinus, and brainsurface. Rats were allowed to recover for a week before startingany experimental procedure.
Intracerebral microdialysis. Microdialysis probes were con-structed in the laboratory as previously described (Hernandez et al.,1983, 1986). Briefly, the probe consisted of a 13 mm-long, 26-ga,stainless steel tube plus a tip of a 2 mm-long cellulose hollow fiber(16000 molecular weight cutoff), protruding 5 mm from the tip of theguide shaft. Inside of the 26-ga tube and the cellulose hollow fiberthere was a concentric fused silica capillary tubing that allowed tooutlet the dialysate. The probes were lowered through the guide shaftand cemented in place 18 h before the experiment. A modifiedRinger solution (136 mM NaCl, 3.7 mM KCl, 1.2 mM CaCl2, 1 mMMgCl2, and 10 mM NaHCO3 at pH 7.4) was perfused through theprobes at a flow rate of 1 l/min. For FST and TP, microdialysateswere collected over 1-min periods into hematocrit glass tubes flameshaped to a conical end to avoid evaporation (Hernandez et al.,2007). A group of rats (n�14) was directly subjected to the FST formonitoring extracellular GABA levels on VP as described later in thetext. To test for anxiety correlated changes in extracellular VP-GABA,a group of rats (n�7) was subjected to TP on two consecutive days.The dialysis procedure was similar in both FST and TP. Briefly, forday-1 and day-2 of FST and TP respectively, five samples werecollected in the home cage (basal) before placing the animal in thewater tank (FST) or tail-pinching with the forceps, followed by 10samples during FST or TP (10-min of test), six samples immediatelyafter the tests (home cage), and three samples at 30, 60, 120, and180 min after the tests. Finally, a control group was intra-VP dialyzedto monitor the effect of a 20-min vigabatrin (20 g/l) infusion byreverse microdialysis pulse infused at 1 l/min flow rate in the mod-ified ringer solution, and samples were collected before, during, andafter the swimming time at the second day of the FST (n�7 for saline,n�7 for vigabatrin). In this experiment, each sample was collectedver 5-min period in the following order: two basal samples, twoamples during 10-min saline or vigabatrin pulse before placing theat in the water tank, two samples during FST (10-min period) plusaline or vigabatrin pulse, two samples after the FST, one sample 1 hfter the FST, and one sample 2 h after the FST.
Intracerebral microinjections. Injectors were made of 33-gatainless steel tubing, 17-mm long, and protruded 2 mm from theip of the guide shaft. Rats were bilaterally intra-VP microinjectedmin before day-2 of the FST, the SPT, or the open field test with
aline (0.9% NaCl), vigabatrin (Marion Merrell Dow, MI, USA), oricuculline methbromide (Sigma, MO, USA). Microinjections con-isted of a total volume of 0.5 l/side infused over a 1-min period.njectors were left in place for another 1 min before withdrawinghe injectors to avoid retrograde mass flow. Before the injectionay, every animal was handled daily for 5 min during five consec-tive days to avoid handling anxiety. On day-2 of the FST, ratseceived bilateral intra-VP microinjections of saline (n�12); viga-atrin, 5 g/side (n�11), 10 g/side (n�11), 15 g/side (n�11);
and bicuculline, 0.1 g/side (n�10); 5 min before the test. To testor anatomical specificity, another group of rats (n�7) received aingle bilateral vigabatrin microinjection (10 g/side) at 2.5 mm
above the VP. Five minutes before the two-bottle sucrose prefer-ence test, a different group of rats received bilateral intra-VPmicroinjections of saline (n�10); vigabatrin, 10 g/side (n�10); orbicuculline, 0.1 g/side (n�10). Finally, 5 min before the openfield test, new rats were bilaterally intra-VP microinjected withsaline (n�10); vigabatrin, 10 g/side (n�10); and bicuculline, 0.1g/side (n�10). For all behavioral test, an investigator blind to thereatments, scored the total time of swimming-immobility behav-ors for FST, and locomotor activity for the open field test on video
ecorded.Behavioral methods
Forced swim test (FST). Scores of immobility behaviors aspreviously described for the FST served as a measure of a be-havioral depression-like state in rats (Porsolt, 1979; Nestler andHyman, 2010; Skirzewski et al., 2010). The test was conductedbetween 0800 and 1200 h on two consecutive days. On day-1,rats were placed individually in an opaque cylindrical water tank(30 cm diameter; 55 cm height; water level at 45 cm from thebottom, and temperature at 20–25 °C) for 10 min. The swimming-immobility time of each rat and for all experimental groups wasvideo recorded. Twenty-four hours later (day-2) subjects wereplaced again in the water tank for 10 min, and the length of theswimming time was video recorded. “Swimming” was defined byescape behaviors (i.e. diving, rigorous padding with all four legs,circling the tank, and clambering at the tank walls). “Immobility”was considered as floating and treading water just enough to keepthe nose above water. The water of the tank was changed for eachtest.
Mild tail-pinch test (TP). A separate group of rats werestressed by mild tail-pinch on two successive days for comparisonwith the FST and to differentiate anxiety from the depressive-likebehavior phenomena (Rada et al., 2006; Nestler and Hyman,2010). The test consisted of placing a forceps with rubber-coveredbranches 3 cm from the proximal end of the tail for 10 min andallowing the animal to freely move around the behavioral cage,and taking precautions to avoid any sign of pain.
Sucrose preference test (SPT). The SPT was used to de-termine anhedonia, which is considered as a core symptom ofdepression-like behaviors. The test was performed according tothe method described by Wang et al. (Wang et al., 2009). Briefly,before testing, the animals were individually placed in a plexiglassbehavioral cage and habituated to drink water from two bottles.Then, 24 h before the test, access to food and water was restrictedfor a period of 20 h. Finally, 5 min before the test, the rats receivedsingle bilateral microinjection of saline, vigabatrin (10 g/side), orbicuculline (0.1 g/side), and two bottles, one with water and thether with 1% sucrose solution were placed in the behavioral cagend consumption was measured over a 2-h period. The sucrosereference index was calculated according to the following for-ula: sucrose intake (ml)/[sucrose intake (ml) � water intake
ml)]. The test was conducted between 1700 and 2000 h.
Open field test. To test for nonspecific drug effects on lo-omotor activity, an open field test was performed in new rats. Thepen field test consisted of a wooden chamber (45 cm high) withsquare base (60�60 cm2) divided in nine squared sections
20�20 cm2). The apparatus was placed in a noise-isolated roomat 22 °C, illuminated with a white 70 W lamp placed 180 cm overthe center of the arena. Twenty minutes before the test, theanimals were individually placed in the open field behavioral cageto avoid altered locomotor activity due to novelty-exploratory be-havior (Berlyne et al., 1966). After this time, the rats (saline, 10g-vigabatrin/side, and 0.1 g-bicuculline/side) received singlebilateral intra-VP microinjections and then were placed again inthe behavioral cage. The open field test consisted in placing theanimals on the center section of the apparatus and counting thetotal number of entries to each section for 20 min. The woodenchamber was rinsed with 70% (v/v) of alcohol solution betweenevery animal. The entire test was video monitored, and an inves-tigator blind to the conditions counted the total number of entriesto each floor section.
Analytical methods
Micellar electrokinetic chromatography, laser-induced fluores-cence detection (MEKC-LIFD). In the present experiment, ex-
tracellular GABA was monitored in microdialysates of VP of rats
ica
satom5
opn
aq
w(stafaDs
Pdscg(ccappu
1(wUd
ppg
Stc
t
trdcggem
etemp�
pb
sww
MrrSI
WUcr(o
di
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146134
during FST and TP. The protocol for monitoring GABA by MEKC-LIFD has been reported elsewhere (Rada et al., 1999). Eachsample of 1 l of total volume was mixed in a 1:1 (v/v) ratio witha derivatizing solution. This one was prepared by mixing a 66 Msolution of fluorescein (Sigma) in 20 mM carbonate buffer at pH9.4 with a 2.5 mM solution of fluorescein isothiocyanate isomer-I(FITC) (Sigma) in acetone in a 1:1 (v/v) ratio. The samples reactedovernight (16 h) in dark at room temperature and in a watersaturated chamber to avoid evaporation (Hernandez et al., 1993).Ten microliters of derivatizing solution were mixed with 1 ml of a9.7 mM GABA solution and left to react overnight. We employeda Meridialysis® capillary zone electrophoresis system (ModelR2D2, Mérida, Venezuela) equipped with an argon laser (20 mW)(Omnichrom, CA, USA) tuned to 488 nm. A fiber optic conductedthe laser beam onto a dichroic mirror centered at 510 nm andfocused on the window of a capillary (Polymicro Technologies,Phoenix, AZ, USA) by a 0.85-numeric aperture objective. Thewindow was made by burning out the polyimide coating at 35 cmfrom the anodic end of a 45 cm long capillary, with a 26 m ofnternal diameter and 358 m of outside diameter, fused-silicaapillary. The fluorescence was captured through the objectivend focused on a multialkali photomultiplier tube.
The running buffer was a 40 mM borate buffer with 20 mModium dodecyl sulfate (SDS). The sample was loaded at thenodic end by a negative pressure (�12 p.s.i. for 0.5 s) applied athe cathodic end of the capillary. Electrophoretic separation wasbtained by applying 28 kV between the anode and cathode for 25in. Before each run, the capillary was rinsed with 1 M NaOH formin followed by water for 5 min and running buffer for 5 min.
After electrophoretic separation, an electropherogram wasbtained for every dialysate sample, and GABA and fluoresceineak heights (PH) were identified and measured. GABA PH wasormalized according to the following formula: (GABAsample PH �
fluoresceino PH)/fluoresceinsample PH. Then, a ratio of normalizedGABA PH vs. basal normalized GABA PH was calculated asfollows: [GABAnormalized PH/mean (basal GABAnormalized PH)]�1.
Semiquantitative reverse transcription-polymerase chain re-ction (SQ-RT-PCR). SQ-RT-PCR was employed to relativelyuantify mRNA expression of 17 subunits for the GABAA-R (�1–
�6, �1–�3, �2, �, �, �, �, and �1–�3) in the VP of rats. Total RNAas isolated immediately after day-2 with ice-cold Trizol reagent
Invitrogen, CA, USA) from fresh bilaterally VP dissected under atereoscopic microscope and using stereotaxic coordinates (AP: 0o �1 mm; L: 1.2 to 2.4 mm; V: 7.4 to 9 mm), from controls, TP,nd FST rats. Samples were independently collected and pooled
rom a total of five rats for each experimental group to obtain anppropriate amount of total RNA. All the RNA samples wereNase I-treated and reverse transcribed following a protocol de-cribed elsewhere (Rada et al., 2006).
Primers for detection of GABAA-R subunits transcripts byCR were designed into intronic DNA sequences from GenBankatabase for each GABAA-R subunit gene using Oligo Exploreroftware 1.1.2 (Teemu Kuulasmaa Kuopio, Finland) and pur-hased from Invitrogen. We also used a pair of primers to amplifyliceralgehyde-3-phosphate dehydrogenase (GAPDH) transcriptFort et al., 1985) as internal standard to normalize changes inDNA levels. Before examining each gene, a standard curve ofycle number within the linear and non-saturated range for PCRmplification was generated. The nucleic acid compositions for allrimers, GenBank access number, amplicon size, annealing tem-erature (°C) to each primer pairs, and number of PCR cyclesnder a linear range of amplification applied are listed in Table 1.
The amplified products were resolved in 2% agarose gel at00 V with 0.5 g/ml of ethidium bromide and 100 bp DNA LadderPromega, Madison, WI, USA) as a size marker, and visualizedith CCD system, Gel Logic Imaging System (Kodak, New Haven,SA), by 0.5 s to avoid over exposition of the bands. Then, bands
ensities (pixels2) were quantified with Molecular Imaging Soft- aware, V 4.0.1 (Kodak), for control, TP, and FST groups, andnormalized with GAPDH band density according to the followingformula: (GAPDH band densitycontrol/GAPDH band densitysample)�band densitysample. Finally, the expression index variation wascalculated for TP and FST groups, considering band density ofcontrol group as a baseline expression. We employed the follow-ing formula: (band densitysample/band densitycontrol)�1. Every ex-eriment that corresponds to a total of five pooled samples waserformed independently at least three times and the datarouped for analysis.
Nucleotide sequence of PCR fragments generated during aQ-RT-PCR for GABAA-R subunits were obtained at MGW Bio-
ech AG (Ebersberg, USA) and CESAAN (IVIC, Venezuela) toonfirm the amplified.
Immunoblot. Assay was performed to measure relative pro-ein expression of three GABAA-R subunits (�3, �, and �2) in VPof control, TP, and FST treated rats, using �-tubulin protein de-ection as internal standard. Bilateral VP of control, TP, and FSTats were dissected at day-2 or day-8 after tests as has beenescribed in the previous section. Samples were independentlyollected and pooled from a total of five rats for each experimentalroup for both day-2 and day-8 groups. VP samples of everyroup were pooled and homogenized independently in a homog-nization buffer (20 mM Tris–HCl, 137 mM NaCl, 2 mM EDTA, 1M DTT, 1 mM PMSF, 0.1 mM Na3VO4, 1 M NaF, 10 mM
�-glicerolphosphate, 1% nonidet P-40, 10 g/ml leupeptin, 5g/ml aprotinin at pH 7.4) at 4 °C. Protein concentrations werestimated by Bradford method (Bradford, 1976). Samples werehen diluted in SDS buffer (10% glycerol, 143 mM �-mercapto-thanol, 187.5 mM Tris–HCl, 0.1% Bromophenol Blue, and 22.9M SDS in homogenization buffer, pH 7.4) at 5 g/l of finalrotein concentrations, boiled at 100 °C for 10 min and stored at20 °C until used.
Protein samples were resolved in 12% SDS-PAGE (150 grotein/well) at 100 V and transferred to a nitrocellulose mem-rane (0.45 m) (Hybond ECL, Amersham GE Healthcare, Nor-
way). The very similar amounts of proteins loaded in gels weredemonstrated with a Ponceau staining. For immunodetection,blots were blocked as follows: for �3 subunit, 2 h in TBST and 5%casein; for � subunit, 1 h in TBST and 5% nonfat milk; for �2ubunit, 2 h in TBST and 5% casein-1% BSA. Then, incubationith commercial primary antibodies against rats overnight at 4 °Cas performed as follows: for �3 subunit, 1:100 of GABAA R�3
N-19 (sc-7353, Santa Cruz Biotechnology, CA, USA); for � sub-unit, 1:500 of GABAA R� H-100 (sc-25705, Santa Cruz); for �2subunit, 1:550 of rabbit-polyclonal IgG anti-�2 subunit (AB5954,
illipore, MA, USA). Incubation with secondary antibodies horseadish peroxidase-conjugated (IgG-HRP) were performed 1 h atoom temperature with donkey anti-goat IgG-HRP (sc-2020,anta Cruz) diluted at 1:4500 for �3 subunit and goat anti-rabbit
gG-HRP (AP187P, Millipore) diluted at 1:5000 for � and 1:6000for �2 subunit. Immunoreactivity was detected by SuperSignal
est Pico Chemiluminescent Substrate (Thermo Scientific, IL,SA), and x-ray films were exposed to blots under non-saturatingonditions. Blots were then stripped for 10 min of incubation atoom temperature with Restore Western Blot Stripping BufferThermo Scientific), blocked 1 h with 4% of nonfat milk, probedvernight with 1:100 of anti-�-tubulin P-16 antibody (sc-31779,
Santa Cruz) and 1 h of incubation with secondary 1:3000 donkeyanti-goat IgG-HRP (sc-2020, Santa Cruz) for chemiluminescentdetection. Finally, GABAA-R subunit blots with internal standardblots were digitalized under non-saturating conditions with a CCDsystem, Gel Logic Imaging System (Kodak) and densities quan-tified with Molecular Imaging Software, V 4.0.1 (Kodak), for con-trol, TP, and FST groups, and normalized with �-tubulin bandensity in the same way that was calculated for mRNA expression
n the previous section. Immunoblot conditions were verified using
total cerebellar protein extract as a positive control. Every ex-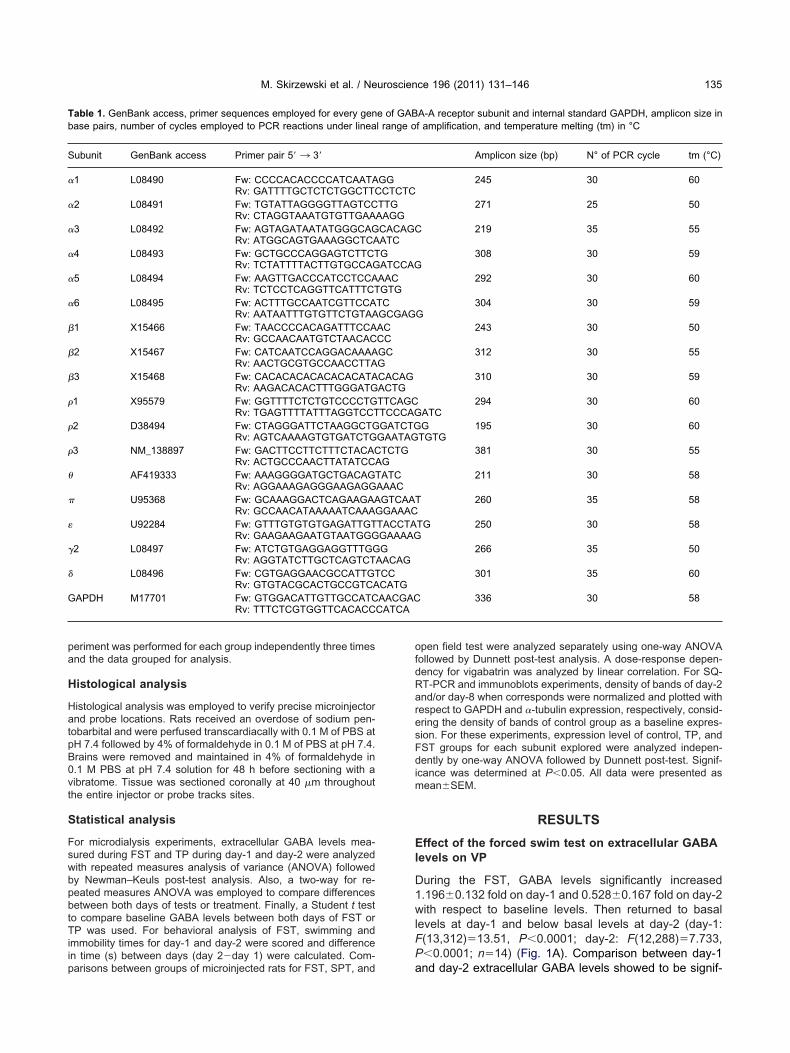
�
GCATCA
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146 135
periment was performed for each group independently three timesand the data grouped for analysis.
Histological analysis
Histological analysis was employed to verify precise microinjectorand probe locations. Rats received an overdose of sodium pen-tobarbital and were perfused transcardiacally with 0.1 M of PBS atpH 7.4 followed by 4% of formaldehyde in 0.1 M of PBS at pH 7.4.Brains were removed and maintained in 4% of formaldehyde in0.1 M PBS at pH 7.4 solution for 48 h before sectioning with avibratome. Tissue was sectioned coronally at 40 m throughoutthe entire injector or probe tracks sites.
Statistical analysis
For microdialysis experiments, extracellular GABA levels mea-sured during FST and TP during day-1 and day-2 were analyzedwith repeated measures analysis of variance (ANOVA) followedby Newman–Keuls post-test analysis. Also, a two-way for re-peated measures ANOVA was employed to compare differencesbetween both days of tests or treatment. Finally, a Student t testto compare baseline GABA levels between both days of FST orTP was used. For behavioral analysis of FST, swimming andimmobility times for day-1 and day-2 were scored and differencein time (s) between days (day 2�day 1) were calculated. Com-
Table 1. GenBank access, primer sequences employed for every genbase pairs, number of cycles employed to PCR reactions under linea
Subunit GenBank access Primer pair 5= ¡ 3=
�1 L08490 Fw: CCCCACACCCCATCAATARv: GATTTTGCTCTCTGGCTTC
�2 L08491 Fw: TGTATTAGGGGTTAGTCCRv: CTAGGTAAATGTGTTGAA
�3 L08492 Fw: AGTAGATAATATGGGCAGRv: ATGGCAGTGAAAGGCTCA
�4 L08493 Fw: GCTGCCCAGGAGTCTTCTRv: TCTATTTTACTTGTGCCAG
�5 L08494 Fw: AAGTTGACCCATCCTCCARv: TCTCCTCAGGTTCATTTCT
�6 L08495 Fw: ACTTTGCCAATCGTTCCARv: AATAATTTGTGTTCTGTAA
�1 X15466 Fw: TAACCCCACAGATTTCCARv: GCCAACAATGTCTAACAC
�2 X15467 Fw: CATCAATCCAGGACAAAARv: AACTGCGTGCCAACCTTA
�3 X15468 Fw: CACACACACACACACATARv: AAGACACACTTTGGGATG
�1 X95579 Fw: GGTTTTCTCTGTCCCCTGRv: TGAGTTTTATTTAGGTCCT
�2 D38494 Fw: CTAGGGATTCTAAGGCTGRv: AGTCAAAAGTGTGATCTG
�3 NM_138897 Fw: GACTTCCTTCTTTCTACACRv: ACTGCCCAACTTATATCC
� AF419333 Fw: AAAGGGGATGCTGACAGRv: AGGAAAGAGGGAAGAGG
� U95368 Fw: GCAAAGGACTCAGAAGAARv: GCCAACATAAAAATCAAA
U92284 Fw: GTTTGTGTGTGAGATTGTRv: GAAGAAGAATGTAATGGG
�2 L08497 Fw: ATCTGTGAGGAGGTTTGGRv: AGGTATCTTGCTCAGTCT
� L08496 Fw: CGTGAGGAACGCCATTGTRv: GTGTACGCACTGCCGTCA
APDH M17701 Fw: GTGGACATTGTTGCCATCRv: TTTCTCGTGGTTCACACC
parisons between groups of microinjected rats for FST, SPT, and
open field test were analyzed separately using one-way ANOVAfollowed by Dunnett post-test analysis. A dose-response depen-dency for vigabatrin was analyzed by linear correlation. For SQ-RT-PCR and immunoblots experiments, density of bands of day-2and/or day-8 when corresponds were normalized and plotted withrespect to GAPDH and �-tubulin expression, respectively, consid-ering the density of bands of control group as a baseline expres-sion. For these experiments, expression level of control, TP, andFST groups for each subunit explored were analyzed indepen-dently by one-way ANOVA followed by Dunnett post-test. Signif-icance was determined at P�0.05. All data were presented asmean�SEM.
RESULTS
Effect of the forced swim test on extracellular GABAlevels on VP
During the FST, GABA levels significantly increased1.196�0.132 fold on day-1 and 0.528�0.167 fold on day-2with respect to baseline levels. Then returned to basallevels at day-1 and below basal levels at day-2 (day-1:F(13,312)�13.51, P�0.0001; day-2: F(12,288)�7.733,P�0.0001; n�14) (Fig. 1A). Comparison between day-1
A-A receptor subunit and internal standard GAPDH, amplicon size inf amplification, and temperature melting (tm) in °C
Amplicon size (bp) N° of PCR cycle tm (°C)
245 30 60
271 25 50
219 35 55
308 30 59
292 30 60
G304 30 59
243 30 50
312 30 55
310 30 59
ATC294 30 60
GTGTG
195 30 60
381 30 55
211 30 58
T 260 35 58
TG 250 30 58
266 35 50
301 35 60
336 30 58
e of GABl range o
GGCTCTC
TTGAAGGCACAGCATCGATCCAGAACGTG
TCGCGAGACCCGCGCACAGACTGTTCAGCTCCCAGGATCTGGAATAGTCTG
AGTATCAAACGTCAA
GGAAACTACCTAGAAAAGG
AACAGCCCATGAACGAC
and day-2 extracellular GABA levels showed to be signif-
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146136
icantly different when compared by two-way for repeatedmeasures ANOVA (F�34.81, df�1/576, P�0.0001). Ab-solute baseline GABA levels measured in VP increasedsignificantly from day-1 (1.50�0.14 nM) to day-2(2.84�0.16 nM) in rats subject to FST (t(12)�8.253, P�0.0001)(Fig. 1B).
Effect of mild tail-pinch test on extracellular GABAlevels on VP
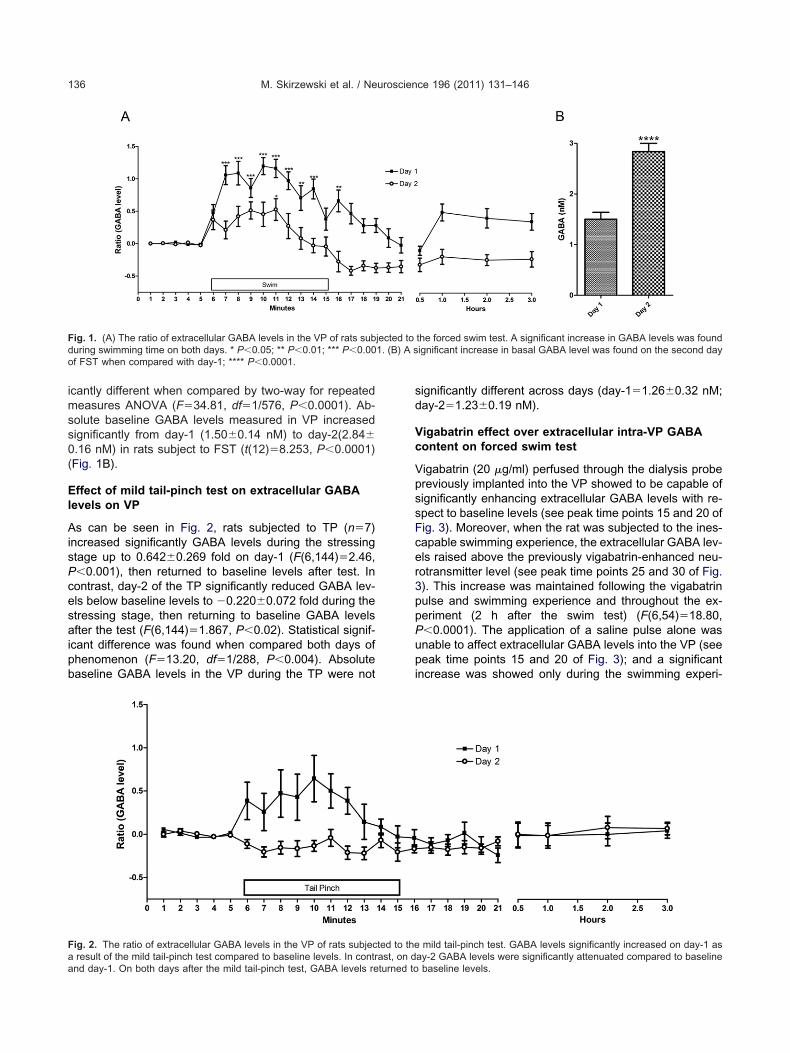
As can be seen in Fig. 2, rats subjected to TP (n�7)increased significantly GABA levels during the stressingstage up to 0.642�0.269 fold on day-1 (F(6,144)�2.46,P�0.001), then returned to baseline levels after test. Incontrast, day-2 of the TP significantly reduced GABA lev-els below baseline levels to �0.220�0.072 fold during thestressing stage, then returning to baseline GABA levelsafter the test (F(6,144)�1.867, P�0.02). Statistical signif-icant difference was found when compared both days ofphenomenon (F�13.20, df�1/288, P�0.004). Absolutebaseline GABA levels in the VP during the TP were not
Fig. 1. (A) The ratio of extracellular GABA levels in the VP of rats subduring swimming time on both days. * P�0.05; ** P�0.01; *** P�0.00of FST when compared with day-1; **** P�0.0001.
Fig. 2. The ratio of extracellular GABA levels in the VP of rats subjeca result of the mild tail-pinch test compared to baseline levels. In contr
and day-1. On both days after the mild tail-pinch test, GABA levels returned tosignificantly different across days (day-1�1.26�0.32 nM;day-2�1.23�0.19 nM).
Vigabatrin effect over extracellular intra-VP GABAcontent on forced swim test
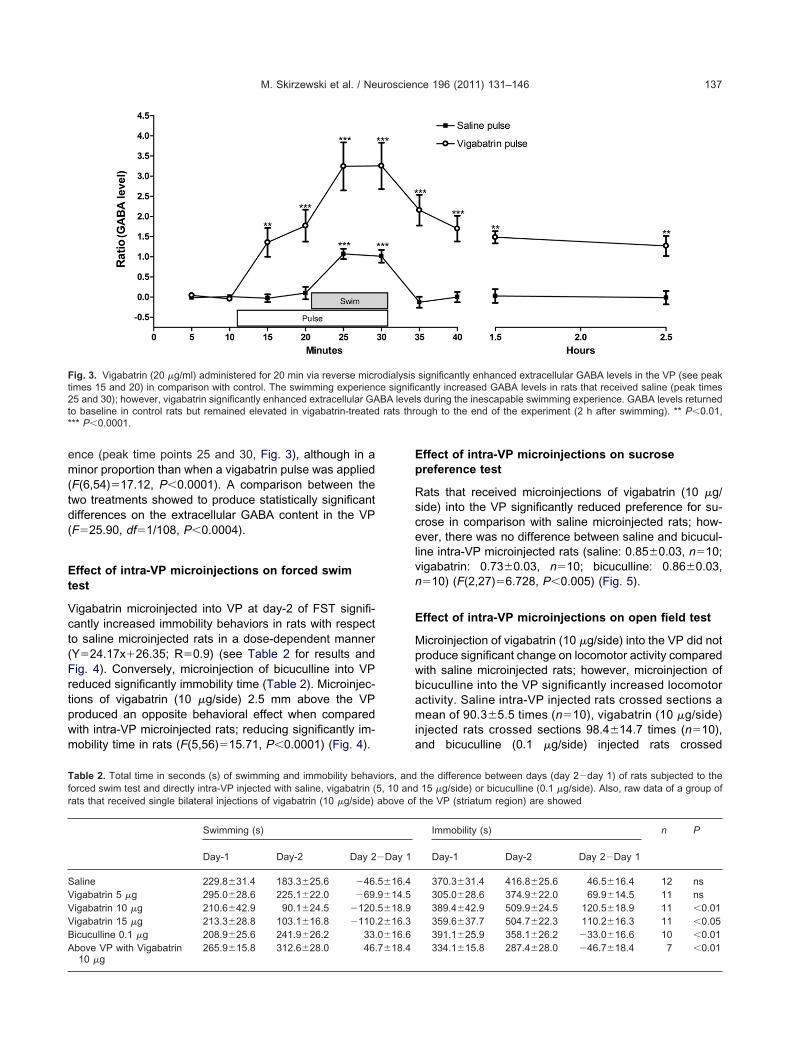
Vigabatrin (20 g/ml) perfused through the dialysis probepreviously implanted into the VP showed to be capable ofsignificantly enhancing extracellular GABA levels with re-spect to baseline levels (see peak time points 15 and 20 ofFig. 3). Moreover, when the rat was subjected to the ines-capable swimming experience, the extracellular GABA lev-els raised above the previously vigabatrin-enhanced neu-rotransmitter level (see peak time points 25 and 30 of Fig.3). This increase was maintained following the vigabatrinpulse and swimming experience and throughout the ex-periment (2 h after the swim test) (F(6,54)�18.80,P�0.0001). The application of a saline pulse alone wasunable to affect extracellular GABA levels into the VP (seepeak time points 15 and 20 of Fig. 3); and a significantincrease was showed only during the swimming experi-
the forced swim test. A significant increase in GABA levels was foundignificant increase in basal GABA level was found on the second day
mild tail-pinch test. GABA levels significantly increased on day-1 asay-2 GABA levels were significantly attenuated compared to baseline
jected to1. (B) A s
ted to theast, on d
baseline levels.
td(
pwm
Ep
Rscel
pwbamia
t2t*
Tf
VVBA
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146 137
ence (peak time points 25 and 30, Fig. 3), although in aminor proportion than when a vigabatrin pulse was applied(F(6,54)�17.12, P�0.0001). A comparison between thewo treatments showed to produce statistically significantifferences on the extracellular GABA content in the VPF�25.90, df�1/108, P�0.0004).
Effect of intra-VP microinjections on forced swimtest
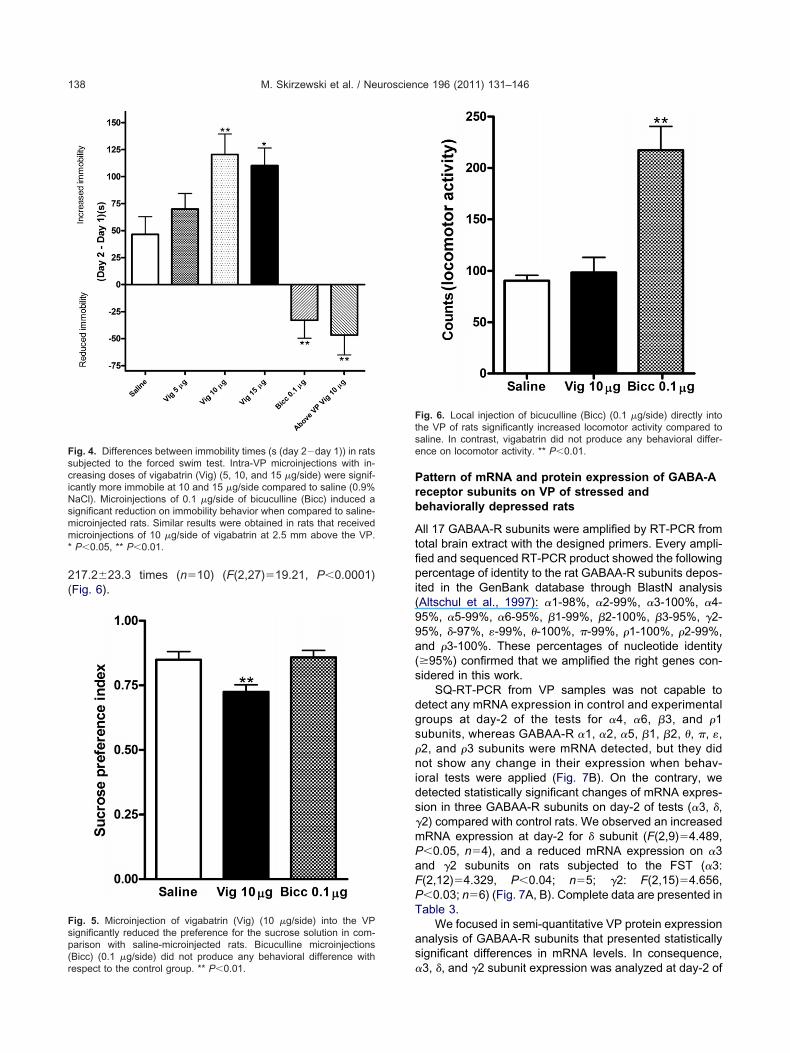
Vigabatrin microinjected into VP at day-2 of FST signifi-cantly increased immobility behaviors in rats with respectto saline microinjected rats in a dose-dependent manner(Y�24.17x�26.35; R�0.9) (see Table 2 for results andFig. 4). Conversely, microinjection of bicuculline into VPreduced significantly immobility time (Table 2). Microinjec-tions of vigabatrin (10 g/side) 2.5 mm above the VProduced an opposite behavioral effect when comparedith intra-VP microinjected rats; reducing significantly im-obility time in rats (F(5,56)�15.71, P�0.0001) (Fig. 4).
Fig. 3. Vigabatrin (20 g/ml) administered for 20 min via reverse micrimes 15 and 20) in comparison with control. The swimming experien5 and 30); however, vigabatrin significantly enhanced extracellular GAo baseline in control rats but remained elevated in vigabatrin-treated** P�0.0001.
able 2. Total time in seconds (s) of swimming and immobility behavorced swim test and directly intra-VP injected with saline, vigabatrin (rats that received single bilateral injections of vigabatrin (10 g/side)
Swimming (s)
Day-1 Day-2 Day 2
Saline 229.8�31.4 183.3�25.6 �46.Vigabatrin 5 g 295.0�28.6 225.1�22.0 �69.
igabatrin 10 g 210.6�42.9 90.1�24.5 �120.igabatrin 15 g 213.3�28.8 103.1�16.8 �110.icuculline 0.1 g 208.9�25.6 241.9�26.2 33.bove VP with Vigabatrin 265.9�15.8 312.6�28.0 46.
10 gffect of intra-VP microinjections on sucrosereference test
ats that received microinjections of vigabatrin (10 g/ide) into the VP significantly reduced preference for su-rose in comparison with saline microinjected rats; how-ver, there was no difference between saline and bicucul-
ine intra-VP microinjected rats (saline: 0.85�0.03, n�10;vigabatrin: 0.73�0.03, n�10; bicuculline: 0.86�0.03,n�10) (F(2,27)�6.728, P�0.005) (Fig. 5).
Effect of intra-VP microinjections on open field test
Microinjection of vigabatrin (10 g/side) into the VP did notroduce significant change on locomotor activity comparedith saline microinjected rats; however, microinjection oficuculline into the VP significantly increased locomotorctivity. Saline intra-VP injected rats crossed sections aean of 90.3�5.5 times (n�10), vigabatrin (10 g/side)
njected rats crossed sections 98.4�14.7 times (n�10),nd bicuculline (0.1 g/side) injected rats crossed
significantly enhanced extracellular GABA levels in the VP (see peakantly increased GABA levels in rats that received saline (peak times
s during the inescapable swimming experience. GABA levels returnedough to the end of the experiment (2 h after swimming). ** P�0.01,
the difference between days (day 2�day 1) of rats subjected to the15 g/side) or bicuculline (0.1 g/side). Also, raw data of a group ofthe VP (striatum region) are showed
Immobility (s) n P
Day-1 Day-2 Day 2�Day 1
370.3�31.4 416.8�25.6 46.5�16.4 12 ns305.0�28.6 374.9�22.0 69.9�14.5 11 ns389.4�42.9 509.9�24.5 120.5�18.9 11 �0.01359.6�37.7 504.7�22.3 110.2�16.3 11 �0.05391.1�25.9 358.1�26.2 �33.0�16.6 10 �0.01334.1�15.8 287.4�28.0 �46.7�18.4 7 �0.01
odialysisce significBA levelrats thr
iors, and5, 10 andabove of
�Day 1
5�16.49�14.55�18.92�16.30�16.67�18.4
9
a(s
dgs
nids
mPa
PT
as
iNsmm
sp(r
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146138
217.2�23.3 times (n�10) (F(2,27)�19.21, P�0.0001)(Fig. 6).
Fig. 4. Differences between immobility times (s (day 2�day 1)) in ratssubjected to the forced swim test. Intra-VP microinjections with in-creasing doses of vigabatrin (Vig) (5, 10, and 15 g/side) were signif-cantly more immobile at 10 and 15 g/side compared to saline (0.9%aCl). Microinjections of 0.1 g/side of bicuculline (Bicc) induced aignificant reduction on immobility behavior when compared to saline-icroinjected rats. Similar results were obtained in rats that receivedicroinjections of 10 g/side of vigabatrin at 2.5 mm above the VP.
* P�0.05, ** P�0.01.
Fig. 5. Microinjection of vigabatrin (Vig) (10 g/side) into the VPignificantly reduced the preference for the sucrose solution in com-arison with saline-microinjected rats. Bicuculline microinjectionsBicc) (0.1 g/side) did not produce any behavioral difference with
espect to the control group. ** P�0.01.Pattern of mRNA and protein expression of GABA-Areceptor subunits on VP of stressed andbehaviorally depressed rats
All 17 GABAA-R subunits were amplified by RT-PCR fromtotal brain extract with the designed primers. Every ampli-fied and sequenced RT-PCR product showed the followingpercentage of identity to the rat GABAA-R subunits depos-ited in the GenBank database through BlastN analysis(Altschul et al., 1997): �1-98%, �2-99%, �3-100%, �4-5%, �5-99%, �6-95%, �1-99%, �2-100%, �3-95%, �2-
95%, �-97%, �-99%, �-100%, �-99%, �1-100%, �2-99%,nd �3-100%. These percentages of nucleotide identity95%) confirmed that we amplified the right genes con-idered in this work.
SQ-RT-PCR from VP samples was not capable toetect any mRNA expression in control and experimentalroups at day-2 of the tests for �4, �6, �3, and �1ubunits, whereas GABAA-R �1, �2, �5, �1, �2, �, �, �,
�2, and �3 subunits were mRNA detected, but they didot show any change in their expression when behav-
oral tests were applied (Fig. 7B). On the contrary, weetected statistically significant changes of mRNA expres-ion in three GABAA-R subunits on day-2 of tests (�3, �,
�2) compared with control rats. We observed an increasedRNA expression at day-2 for � subunit (F(2,9)�4.489,�0.05, n�4), and a reduced mRNA expression on �3nd �2 subunits on rats subjected to the FST (�3:
F(2,12)�4.329, P�0.04; n�5; �2: F(2,15)�4.656,�0.03; n�6) (Fig. 7A, B). Complete data are presented inable 3.
We focused in semi-quantitative VP protein expressionnalysis of GABAA-R subunits that presented statisticallyignificant differences in mRNA levels. In consequence,
Fig. 6. Local injection of bicuculline (Bicc) (0.1 g/side) directly intothe VP of rats significantly increased locomotor activity compared tosaline. In contrast, vigabatrin did not produce any behavioral differ-ence on locomotor activity. ** P�0.01.
�3, �, and �2 subunit expression was analyzed at day-2 of
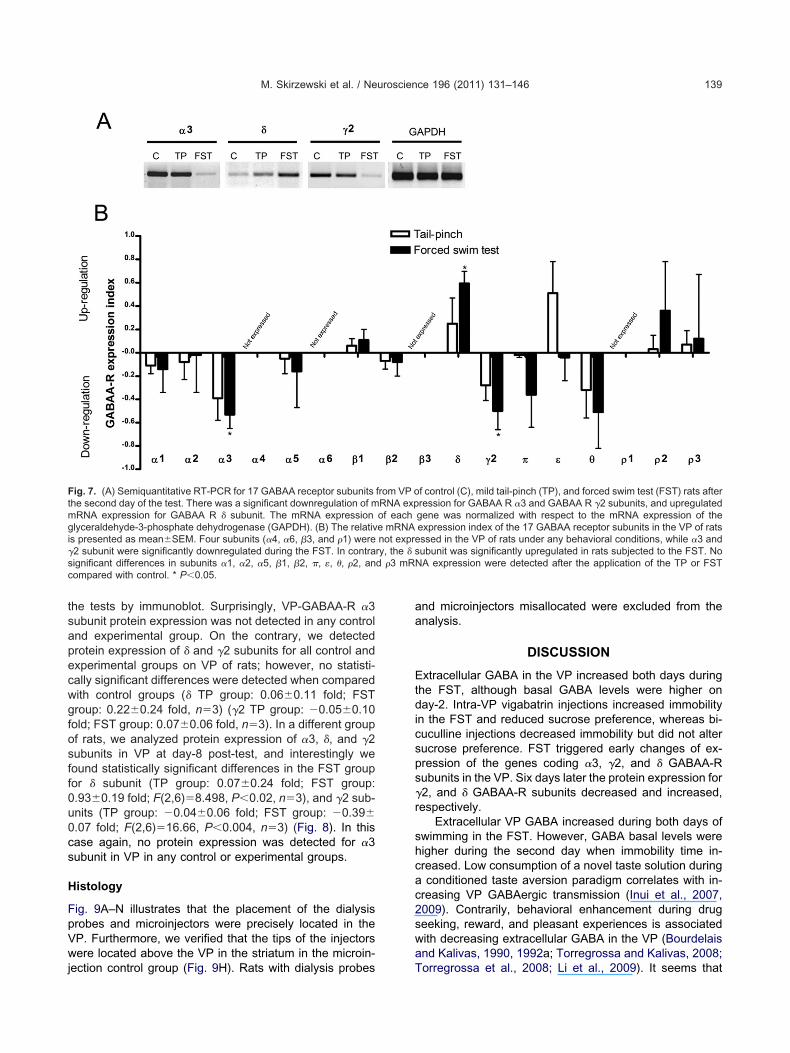
ecwgfosff
u
s
H
FpVwj
aa
s
mgi
d �3 mR
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146 139
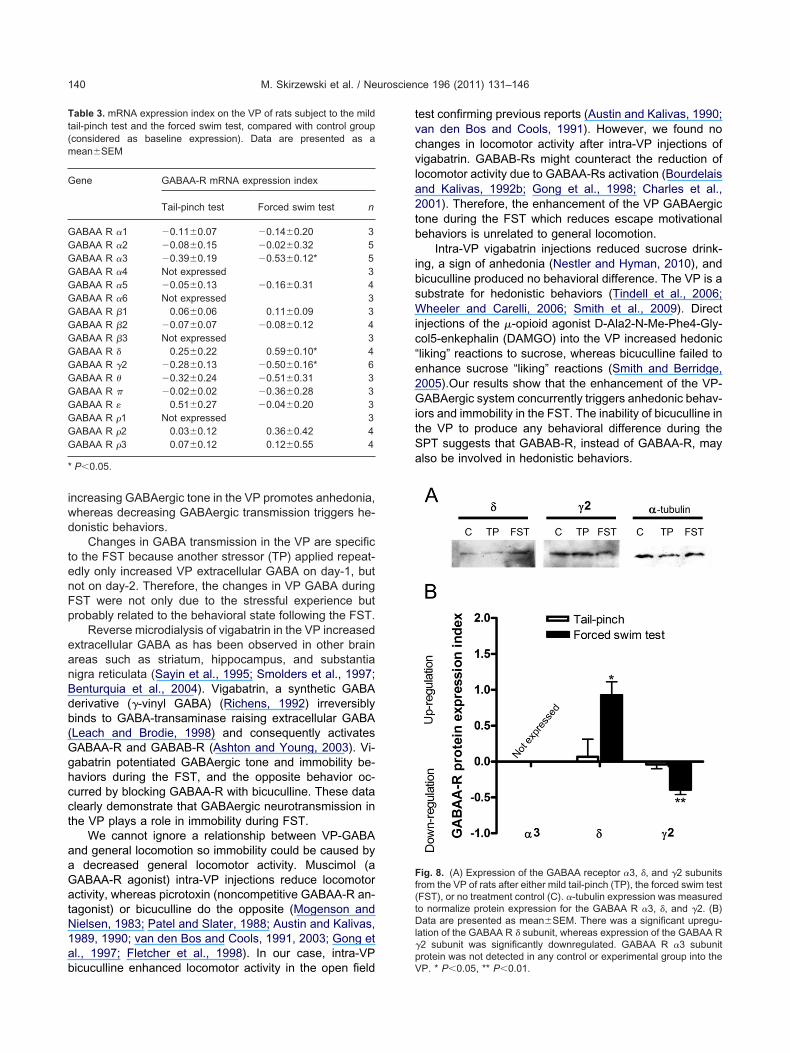
the tests by immunoblot. Surprisingly, VP-GABAA-R �3subunit protein expression was not detected in any controland experimental group. On the contrary, we detectedprotein expression of � and �2 subunits for all control andxperimental groups on VP of rats; however, no statisti-ally significant differences were detected when comparedith control groups (� TP group: 0.06�0.11 fold; FSTroup: 0.22�0.24 fold, n�3) (�2 TP group: �0.05�0.10old; FST group: 0.07�0.06 fold, n�3). In a different groupf rats, we analyzed protein expression of �3, �, and �2ubunits in VP at day-8 post-test, and interestingly weound statistically significant differences in the FST groupor � subunit (TP group: 0.07�0.24 fold; FST group:0.93�0.19 fold; F(2,6)�8.498, P�0.02, n�3), and �2 sub-nits (TP group: �0.04�0.06 fold; FST group: �0.39�
0.07 fold; F(2,6)�16.66, P�0.004, n�3) (Fig. 8). In thiscase again, no protein expression was detected for �3ubunit in VP in any control or experimental groups.
istology
ig. 9A–N illustrates that the placement of the dialysisrobes and microinjectors were precisely located in theP. Furthermore, we verified that the tips of the injectorsere located above the VP in the striatum in the microin-
Fig. 7. (A) Semiquantitative RT-PCR for 17 GABAA receptor subunitsthe second day of the test. There was a significant downregulation of m
RNA expression for GABAA R � subunit. The mRNA expressionlyceraldehyde-3-phosphate dehydrogenase (GAPDH). (B) The relativ
s presented as mean�SEM. Four subunits (�4, �6, �3, and �1) were�2 subunit were significantly downregulated during the FST. In contrasignificant differences in subunits �1, �2, �5, �1, �2, �, �, �, �2, ancompared with control. * P�0.05.
ection control group (Fig. 9H). Rats with dialysis probes
nd microinjectors misallocated were excluded from thenalysis.
DISCUSSION
Extracellular GABA in the VP increased both days duringthe FST, although basal GABA levels were higher onday-2. Intra-VP vigabatrin injections increased immobilityin the FST and reduced sucrose preference, whereas bi-cuculline injections decreased immobility but did not altersucrose preference. FST triggered early changes of ex-pression of the genes coding �3, �2, and � GABAA-Rubunits in the VP. Six days later the protein expression for
�2, and � GABAA-R subunits decreased and increased,respectively.
Extracellular VP GABA increased during both days ofswimming in the FST. However, GABA basal levels werehigher during the second day when immobility time in-creased. Low consumption of a novel taste solution duringa conditioned taste aversion paradigm correlates with in-creasing VP GABAergic transmission (Inui et al., 2007,2009). Contrarily, behavioral enhancement during drugseeking, reward, and pleasant experiences is associatedwith decreasing extracellular GABA in the VP (Bourdelaisand Kalivas, 1990, 1992a; Torregrossa and Kalivas, 2008;
f control (C), mild tail-pinch (TP), and forced swim test (FST) rats afterression for GABAA R �3 and GABAA R �2 subunits, and upregulatedgene was normalized with respect to the mRNA expression of theexpression index of the 17 GABAA receptor subunits in the VP of ratsssed in the VP of rats under any behavioral conditions, while �3 andubunit was significantly upregulated in rats subjected to the FST. No
NA expression were detected after the application of the TP or FST
from VP oRNA expof eache mRNAnot expre
ry, the � s
Torregrossa et al., 2008; Li et al., 2009). It seems that
c“e2GitSa
f(tDl
p
Tt(m
G
GG
G
GG
*
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146140
increasing GABAergic tone in the VP promotes anhedonia,whereas decreasing GABAergic transmission triggers he-donistic behaviors.
Changes in GABA transmission in the VP are specificto the FST because another stressor (TP) applied repeat-edly only increased VP extracellular GABA on day-1, butnot on day-2. Therefore, the changes in VP GABA duringFST were not only due to the stressful experience butprobably related to the behavioral state following the FST.
Reverse microdialysis of vigabatrin in the VP increasedextracellular GABA as has been observed in other brainareas such as striatum, hippocampus, and substantianigra reticulata (Sayin et al., 1995; Smolders et al., 1997;Benturquia et al., 2004). Vigabatrin, a synthetic GABAderivative (�-vinyl GABA) (Richens, 1992) irreversiblybinds to GABA-transaminase raising extracellular GABA(Leach and Brodie, 1998) and consequently activatesGABAA-R and GABAB-R (Ashton and Young, 2003). Vi-gabatrin potentiated GABAergic tone and immobility be-haviors during the FST, and the opposite behavior oc-curred by blocking GABAA-R with bicuculline. These dataclearly demonstrate that GABAergic neurotransmission inthe VP plays a role in immobility during FST.
We cannot ignore a relationship between VP-GABAand general locomotion so immobility could be caused bya decreased general locomotor activity. Muscimol (aGABAA-R agonist) intra-VP injections reduce locomotoractivity, whereas picrotoxin (noncompetitive GABAA-R an-tagonist) or bicuculline do the opposite (Mogenson andNielsen, 1983; Patel and Slater, 1988; Austin and Kalivas,1989, 1990; van den Bos and Cools, 1991, 2003; Gong etal., 1997; Fletcher et al., 1998). In our case, intra-VP
able 3. mRNA expression index on the VP of rats subject to the mildail-pinch test and the forced swim test, compared with control groupconsidered as baseline expression). Data are presented as aean�SEM
Gene GABAA-R mRNA expression index
Tail-pinch test Forced swim test n
GABAA R �1 �0.11�0.07 �0.14�0.20 3GABAA R �2 �0.08�0.15 �0.02�0.32 5GABAA R �3 �0.39�0.19 �0.53�0.12* 5GABAA R �4 Not expressed 3
ABAA R �5 �0.05�0.13 �0.16�0.31 4GABAA R �6 Not expressed 3
ABAA R �1 0.06�0.06 0.11�0.09 3ABAA R �2 �0.07�0.07 �0.08�0.12 4
GABAA R �3 Not expressed 3ABAA R � 0.25�0.22 0.59�0.10* 4
GABAA R �2 �0.28�0.13 �0.50�0.16* 6GABAA R � �0.32�0.24 �0.51�0.31 3GABAA R � �0.02�0.02 �0.36�0.28 3GABAA R � 0.51�0.27 �0.04�0.20 3GABAA R �1 Not expressed 3
ABAA R �2 0.03�0.12 0.36�0.42 4ABAA R �3 0.07�0.12 0.12�0.55 4
P�0.05.
bicuculline enhanced locomotor activity in the open field V
test confirming previous reports (Austin and Kalivas, 1990;van den Bos and Cools, 1991). However, we found nochanges in locomotor activity after intra-VP injections ofvigabatrin. GABAB-Rs might counteract the reduction oflocomotor activity due to GABAA-Rs activation (Bourdelaisand Kalivas, 1992b; Gong et al., 1998; Charles et al.,2001). Therefore, the enhancement of the VP GABAergictone during the FST which reduces escape motivationalbehaviors is unrelated to general locomotion.
Intra-VP vigabatrin injections reduced sucrose drink-ing, a sign of anhedonia (Nestler and Hyman, 2010), andbicuculline produced no behavioral difference. The VP is asubstrate for hedonistic behaviors (Tindell et al., 2006;Wheeler and Carelli, 2006; Smith et al., 2009). Directinjections of the -opioid agonist D-Ala2-N-Me-Phe4-Gly-ol5-enkephalin (DAMGO) into the VP increased hedonicliking” reactions to sucrose, whereas bicuculline failed tonhance sucrose “liking” reactions (Smith and Berridge,005).Our results show that the enhancement of the VP-ABAergic system concurrently triggers anhedonic behav-
ors and immobility in the FST. The inability of bicuculline inhe VP to produce any behavioral difference during thePT suggests that GABAB-R, instead of GABAA-R, maylso be involved in hedonistic behaviors.
Fig. 8. (A) Expression of the GABAA receptor �3, �, and �2 subunitsrom the VP of rats after either mild tail-pinch (TP), the forced swim testFST), or no treatment control (C). �-tubulin expression was measuredo normalize protein expression for the GABAA R �3, �, and �2. (B)ata are presented as mean�SEM. There was a significant upregu-
ation of the GABAA R � subunit, whereas expression of the GABAA R�2 subunit was significantly downregulated. GABAA R �3 subunitrotein was not detected in any control or experimental group into the
P. * P�0.05, ** P�0.01.
s
(
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146 141
Fig. 9. (A) Probe track locations from rats subjected to forced swim test or (B) mild tail-pinch test. (C) Probe track locations from rats subjected toforced swim test who also received a 20-min pulse of vehicle or (D) vigabatrin (20 g/ml). (E) Location of microinjectors aimed at the VP of ratsubjected to the forced swim test and treated with vehicle, (F) vigabatrin (5, 10, or 15 g/side of dose), or (G) bicuculline (0.1 g/side). (H) Tip location
of microinjectors aimed 2.5 mm above the VP (negative anatomical control) from rats subjected to the forced swim test. (I) Location of microinjectorsaimed at the VP of rats subjected to the sucrose preference test and treated with vehicle, (J) vigabatrin (10 g/side), or (K) bicuculline (0.1 g/side).L) Location of microinjectors aimed at the VP from rats subjected to the open field test and treated with vehicle, (M) vigabatrin (10 g/side), or (N)
bicuculline (0.1 g/side).

iida
sclm
t
od(bts(2
psaR
tiaafaeSO
uGl2
FctisaoncbacamutarirlYS
i
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146142
Intrastriatal-vigabatrin injections, contrary to the effectn the VP, enhanced swimming and suggest that immobilitynduced by intra-VP vigabatrin is location specific and notue to malaise. These results implicate both the striatumnd VP in the regulation of depressive-like behaviors.
The SQ-RT-PCR evaluates the expression of tran-cripts relative to the expression of a transcript from aontrol, house-keeping gene (such as the GAPDH) be-ieved to be unaffected by the FST. Genes coding for
RNAs of �1, �2, �3, �5, �1, �2, �, �2, �, �, �, �2, and �3GABAA-R subunits expressed in the VP, whereas �4, �6,�3, and �1 subunits were not. Previous reports haveshown the expression of �1, �1, �2, �1, and �2 subunits inhe VP, whereas �3, �5, �3, and �3 subunits are slightly orweakly expressed (Wisden et al., 1992; Pirker et al., 2000;Schwarzer et al., 2001). The present report corroboratesand extends these findings by demonstrating that �2, �, �2,�3, �, �, and � GABAA-R subunits are expressed in the VPf rats. �3 subunit expression by SQ-RT-PCR was notetected, corroborating the findings by Schwarzer et al.Schwarzer et al., 2001). However, weak expression haseen reported by others (Pirker et al., 2000). Cross-reac-ivity in the immunohistochemical techniques or the dis-ecting procedure could lead to false-positive detectionDolman et al., 2004; Holmseth et al., 2005; Pláteník et al.,005; Middeldorp et al., 2009).
FST diminished �3, �2, and enhanced � GABAA-R sub-unit mRNA expression and changed �2 and � GABAA-Rprotein expression 6 days later. The fast (24 h) mRNAchanges and the delayed protein changes are probablydue to the increased basal GABA extracellular level.GABAA-R subunit composition changed in neuronal cellcultures treated with GABA or other substances with anaffinity to GABAA-Rs for several consecutive days (5–7days), and the corresponding protein expression changedin a similarly delayed manner (Calkin and Barnes, 1994;Mhatre and Ticku, 1994; Platt et al., 1996; Yu et al., 1996;Zheng et al., 1996; Gravielle et al., 2005). Changes inGABAA-R composition observed in the VP likely leads to adepression-like phenotype.
The FST reduced the expression of the gene coding forthe �3 subunit in the VP, but we did not detect �3 subunitprotein in the VP in control or in experimental groups.Apparently, �3 does not follow the classical transcriptional/ost-transcriptional pattern of regulation for GABAA-Rubunits as others subunits do (Platt et al., 1996; Wan etl., 1997; Russek et al., 2000; Ma et al., 2004; Steiger andussek, 2004; Hu et al., 2008; Smith et al., 2008).
The most abundant variant of synaptic GABAA-Rs inhe central nervous system contain �2 subunit, for phasicnhibition/desensitizing kinetics (Essrich et al., 1998; Haasnd Macdonald, 1999; Farrant and Nusser, 2005; Olsennd Sieghart, 2008). �-containing GABAA-Rs with a 50-
old higher affinity for GABA are extra-synaptic, and medi-te tonic-inhibition/slow desensitization properties (Sax-na and Macdonald, 1994, 1996; Nusser et al., 1998;emyanov et al., 2004; Mtchedlishvili and Kapur, 2006;lsen and Sieghart, 2008). �2 and � subunit are mutually
exclusive in the same GABAA-R (Tretter et al., 2001; Peng
et al., 2002, 2004). Continuous GABAA-R stimulation, as itoccurs in the FST, decreases �2 and increases � subunitexpression, switching inhibition from phasic to tonic (Calkinand Barnes, 1994; Grimm and See, 2000; Farrant andNusser, 2005; Gravielle et al., 2005). Interestingly, theincreased VP �-subunit expression in rats that exhibit de-pressive-like behaviors could be associated to mood dis-orders (Stell et al., 2003).
�4 and �6 subunits are the natural partners of � sub-nits, and this co-assembly (�4� or �6�) gives to thisABAA-R their typical electrophysiological and pharmaco-
ogical properties (Pirker et al., 2000; Olsen and Sieghart,008). However, we found no gene expression for �4 and
�6 subunits in VP, whereas � subunit was expressed.Therefore, this canonical GABAA-R co-assembly does notoccur in the VP. Several reports have suggested that aminor proportion of native GABAA-Rs containing �4 or �6subunits also contain the � subunit (Nusser et al., 1998;Bencsits et al., 1999; Borghese and Harris, 2007), and it ispossible to find �-containing GABAA-Rs that do not contain�4 or �6 subunits (Bianchi et al., 2002; Feng et al., 2004;
eng and Macdonald, 2004a,b). Moreover, the � subunitan pair with �1� in hippocampal interneurons to mediateonic inhibitory currents (Glykys et al., 2007). This increasen GABAergic tone in the VP of rats triggered by a helplessituation as it occurs in the FST, causes and maintains annhedonic and low motivational state. The enhancementf the VP GABAergic transmission may be partially mag-ified by an enhancement of the expression of the geneoding the � subunit, which in turn increases tonic inhi-ition. The delayed enhancement of expression of thectual � subunit might assure persistence of the in-reased GABAergic inhibition in the VP. Both, presyn-ptic and postsynaptic GABAergic plasticity in the VPight be crucial components of the engram reminding thenpleasant experience. Several authors have proposedhat the VP is an interface of limbic motivational signalsnd motor output coding and causing enhancement ofeward learning, pleasure, and motivation with major rolesn feeding, sex, social affiliation, electrical brain stimulationeward, abuse of drugs, winning money, and depressive-ike behaviors (Mogenson et al., 1980; Mogenson andang, 1991; Cromwell and Berridge, 1993; Berridge, 1996;himura et al., 2006; Smith et al., 2009).
CONCLUSIONS
We demonstrated that the inescapable swimming experi-ence in the FST elevates extracellular GABA levels in theVP, and switches the composition of GABAA-Rs from pre-dominantly phasic (high content of �2 subunits) to predom-nantly tonic composition (high content of � subunits). In-terestingly, this change occurs rapidly for gene expression(24 h) and slower for protein (1 week). The excess ofGABAergic inhibition in the VP could be emulated by directbilateral intra-VP injections of vigabatrin that inhibits catab-olism of synaptic extracellular GABA and consequentlyit accumulates. These neurochemical and molecular
changes caused immobility and anhedonia as revealed by
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146 143
the FST and the SPT, respectively. In general, the en-hanced VP GABAergic tone seems to be related to de-pression-like behaviors and might be an important memorycomponent for unpleasant experiences. Increased GABAtransmission and variation of GABAA-R isoforms might con-tribute to the patho-physiological processes underlying alco-holism, drug addiction, eating disorders, schizophrenia, neu-rodegeneration, Alzheimer’s and Parkinson disease, panicdisorders, postpartum depression, post-traumatic stress dis-orders, and insensitivity to anxiolytic and antidepressanttreatments (Hattori, 2008; Darke et al., 2009; Elizabeth Sub-lette et al., 2009; Goossens et al., 2009; Khaled et al., 2009;Martens, 2009; Stevenson et al., 2009; Strober and Arnett,2009; Weinberger et al., 2009).
Acknowledgments—This work was supported by Consejo de De-sarrollo Científico, Humanístico y Tecnológico (CDCHT-UCLA)grant numbers 014-ME-2007 and 022-ME-2007.
REFERENCES
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W,Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new gener-ation of protein database search programs. Nucleic acids research25:3389–3402.
Archer T, Söderberg U, Ross SB, Jonsson G (1984) Role of olfactorybulbectomy and DSP4 treatment in avoidance learning in the rat.Behav Neurosci 98:496–505.
Ashton H, Young AH (2003) GABA-ergic drugs: exit stage left, enterstage right. J Psychopharmacol (Oxford, England) 17:174–178.
Austin MC, Kalivas PW (1989) Blockade of enkephalinergic andGABAergic mediated locomotion in the nucleus accumbens bymuscimol in the ventral pallidum. Jpn J Pharmacol 50:487–490.
Austin MC, Kalivas PW (1990) Enkephalinergic and GABAergic mod-ulation of motor activity in the ventral pallidum. J Pharmacol ExpTher 252:1370–1377.
Bechtholt-Gompf AJ, Walther HV, Adams MA, Carlezon WA Jr., OngürD, Cohen BM (2010) Blockade of astrocytic glutamate uptake inrats induces signs of anhedonia and impaired spatial memory.Neuropsychopharmacology 35:2049–2059.
Bencsits E, Ebert V, Tretter V, Sieghart W (1999) A significant part ofnative gamma-aminobutyric acid A receptors containing alpha4subunits do not contain gamma or delta subunits. J Biol Chem274:19613–19616.
Benturquia N, Parrot S, Sauvinet V, Renaud B, Denoroy L (2004)Simultaneous determination of vigabatrin and amino acid neu-rotransmitters in brain microdialysates by capillary electrophoresiswith laser-induced fluorescence detection. J Chromatogr B AnalTechnol Biomed Life Sci 806:237–244.
Berlyne DE, Koenig ID, Hirota T (1966) Novelty, arousal, and thereinforcement of diversive exploration in the rat. J Comp PhysiolPsychol 62:222–226.
Berridge KC (1996) Food reward: brain substrates of wanting andliking. Neurosci Biobehav Rev 20:1–25.
Bianchi MT, Haas KF, Macdonald RL (2002) Alpha1 and alpha6subunits specify distinct desensitization, deactivation and neuro-steroid modulation of GABA(A) receptors containing the delta sub-unit. Neuropharmacology 43:492–502.
Borghese CM, Harris RA (2007) Studies of ethanol actions on recom-binant delta-containing gamma-aminobutyric acid type A receptorsyield contradictory results. Alcohol 41:155–162.
Bourdelais A, Kalivas PW (1990) Amphetamine lowers extracellularGABA concentration in the ventral pallidum. Brain Res 516:
132–136.Bourdelais A, Kalivas PW (1992a) Apomorphine decreases extracel-lular GABA in the ventral pallidum of rats with 6-OHDA lesions inthe nucleus accumbens. Brain Res 577:306–311.
Bourdelais AJ, Kalivas PW (1992b) Modulation of extracellular gam-ma-aminobutyric acid in the ventral pallidum using in vivo micro-dialysis. J Neurochem 58:2311–2320.
Bradford MM (1976) A rapid and sensitive method for the quantitationof microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254.
Brambilla P, Perez J, Barale F, Schettini G, Soares JC (2003)GABAergic dysfunction in mood disorders. Mol Psychiatry 8:721–737, 715.
Calkin PA, Barnes EM Jr. (1994) gamma-Aminobutyric acid-A(GABAA) agonists down-regulate GABAA/benzodiazepine recep-tor polypeptides from the surface of chick cortical neurons. J BiolChem 269:1548–1553.
Carlezon WA Jr., Chartoff EH (2007) Intracranial self-stimulation(ICSS) in rodents to study the neurobiology of motivation. NatProtoc 2:2987–2995.
Carlezon WA Jr., Duman RS, Nestler EJ (2005) The many faces ofCREB. Trends Neurosci 28:436–445.
Charles KJ, Evans ML, Robbins MJ, Calver AR, Leslie RA, PangalosMN (2001) Comparative immunohistochemical localisation ofGABA(B1a), GABA(B1b) and GABA(B2) subunits in rat brain, spi-nal cord and dorsal root ganglion. Neuroscience 106:447–467.
Chau D, Rada PV, Kosloff RA, Hoebel BG (1999) Cholinergic, M1receptors in the nucleus accumbens mediate behavioral depres-sion. A possible downstream target for fluoxetine. Ann N Y AcadSci 877:769–774.
Chau DT, Rada P, Kosloff RA, Taylor JL, Hoebel BG (2001) Nucleusaccumbens muscarinic receptors in the control of behavioral de-pression: antidepressant-like effects of local M1 antagonist in thePorsolt swim test. Neuroscience 104:791–798.
Chau DT, Rada PV, Kim K, Kosloff RA, Hoebel BG (2011) Fluoxetinealleviates behavioral depression while decreasing acetylcholinerelease in the nucleus accumbens shell. Neuropsychopharmacol-ogy 36:1729–1737.
Cromwell HC, Berridge KC (1993) Where does damage lead to en-hanced food aversion: the ventral pallidum/substantia innominataor lateral hypothalamus? Brain Res 624:1–10.
Darke S, Mills K, Teesson M, Ross J, Williamson A, Havard A (2009)Patterns of major depression and drug-related problems amongstheroin users across 36 months. Psychiatry Res 166:7–14.
Davidson RJ, Abercrombie H, Nitschke JB, Putnam K (1999) Regionalbrain function, emotion and disorders of emotion. Curr Opin Neu-robiol 9:228–234.
Di Chiara G, Loddo P, Tanda G (1999) Reciprocal changes in prefron-tal and limbic dopamine responsiveness to aversive and rewardingstimuli after chronic mild stress: implications for the psychobiologyof depression. Biol Psychiatry 46:1624–1633.
Dolman DE, Lawrenson JG, Abbott NJ (2004) Cross reactivity ofpolyclonal GFAP antiserum: implications for the in-vitro character-isation of brain endothelium. Brain Res 1012:185–186.
DSMIV (2000) Diagnostic and statistical manual IV. Washington, D.C.:American Psychiatric Press.
Elizabeth Sublette M, Carballo JJ, Moreno C, Galfalvy HC, Brent DA,Birmaher B, John Mann J, Oquendo MA (2009) Substance usedisorders and suicide attempts in bipolar subtypes. J Psychiatr Res43:230–238.
Essrich C, Lorez M, Benson JA, Fritschy JM, Lüscher B (1998) Post-synaptic clustering of major GABAA receptor subtypes requires thegamma 2 subunit and gephyrin. Nat Neurosci 1:563–571.
Farrant M, Nusser Z (2005) Variations on an inhibitory theme: phasicand tonic activation of GABA(A) receptors. Nat Rev 6:215–229.
Farrar AM, Font L, Pereira M, Mingote S, Bunce JG, Chrobak JJ,Salamone JD (2008) Forebrain circuitry involved in effort-related
choice: injections of the GABA(A) agonist muscimol into ventral
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146144
pallidum alter response allocation in food-seeking behavior.Neuroscience 152:321–330.
Feng HJ, Bianchi MT, Macdonald RL (2004) Pentobarbital differen-tially modulates alpha1beta3delta and alpha1beta3gamma2LGABAA receptor currents. Mol Pharmacol 66:988–1003.
Feng HJ, Macdonald RL (2004a) Multiple actions of propofol on al-phabetagamma and alphabetadelta GABAA receptors. Mol Phar-macol 66:1517–1524.
Feng HJ, Macdonald RL (2004b) Proton modulation of alpha 1 beta 3delta GABAA receptor channel gating and desensitization. J Neu-rophysiol 92:1577–1585.
Fletcher PJ, Korth KM, Sabijan MS, DeSousa NJ (1998) Injections ofD-amphetamine into the ventral pallidum increase locomotor activ-ity and responding for conditioned reward: a comparison withinjections into the nucleus accumbens. Brain Res 805:29–40.
Fort P, Marty L, Piechaczyk M, el Sabrouty S, Dani C, Jeanteur P,Blanchard JM (1985) Various rat adult tissues express only onemajor mRNA species from the glyceraldehyde-3-phosphate-dehy-drogenase multigenic family. Nucleic Acids Res 13:1431–1442.
Glykys J, Peng Z, Chandra D, Homanics GE, Houser CR, Mody I(2007) A new naturally occurring GABA(A) receptor subunit part-nership with high sensitivity to ethanol. Nat Neurosci 10:40–48.
Gomez R, Vargas CR, Wajner M, Barros HM (2003) Lower in vivobrain extracellular GABA concentration in diabetic rats duringforced swimming. Brain Res 968:281–284.
Gong W, Neill D, Justice JB Jr. (1997) 6-hydroxydopamine lesion ofventral pallidum blocks acquisition of place preference conditioningto cocaine. Brain Res 754:103–112.
Gong W, Neill DB, Justice JB Jr. (1998) GABAergic modulation ofventral pallidal dopamine release studied by in vivo microdialysis inthe freely moving rat. Synapse 29:406–412.
Goossens L, Braet C, Van Vlierberghe L, Mels S (2009) Loss of controlover eating in overweight youngsters: the role of anxiety, depres-sion and emotional eating. Eur Eat Disord Rev 17:68–78.
Gravielle MC, Faris R, Russek SJ, Farb DH (2005) GABA inducesactivity dependent delayed-onset uncoupling of GABA/benzodiaz-epine site interactions in neocortical neurons. J Biol Chem280:20954–20960.
Grimm JW, See RE (2000) Chronic haloperidol-induced alterations inpallidal GABA and striatal D(1)-mediated dopamine turnover asmeasured by dual probe microdialysis in rats. Neuroscience100:507–514.
Groenewegen HJ, Russchen FT (1984) Organization of the efferentprojections of the nucleus accumbens to pallidal, hypothalamic,and mesencephalic structures: a tracing and immunohistochemicalstudy in the cat. J Comp Neurol 223:347–367.
Haas KF, Macdonald RL (1999) GABAA receptor subunit gamma2and delta subtypes confer unique kinetic properties on recombi-nant GABAA receptor currents in mouse fibroblasts. J Physiol514(Pt 1):27–45.
Hattori H (2008) [Depression in the elderly]. Nippon Ronen IgakkaiZasshi 45:451–461.
Hernandez L, Paez X, Hamlin C (1983) Neurotransmitters extractionby local intracerebral dialysis in anesthetized rats. PharmacolBiochem Behav 18:159–162.
Hernandez L, Rossell S, Tucci S, Paredes D, Rada P (2007) Improve-ment of the temporal resolution of brain microdialysis: sampling inseconds. San Diego, CA: Academic Press.
Hernandez L, Stanley BG, Hoebel BG (1986) A small, removablemicrodialysis probe. Life Sci 39:2629–2637.
Hernandez L, Tucci S, Guzman N, Paez X (1993) In vivo monitoring ofglutamate in the brain by microdialysis and capillary electrophore-sis with laser-induced fluorescence detection. J Chromatogr A652:393–398.
Holmseth S, Dehnes Y, Bjørnsen LP, Boulland JL, Furness DN,Bergles D, Danbolt NC (2005) Specificity of antibodies: unex-pected cross-reactivity of antibodies directed against the excitatory
amino acid transporter 3 (EAAT3). Neuroscience 136:649–660.Hooks MS, Kalivas PW (1995) The role of mesoaccumbens—pallidalcircuitry in novelty-induced behavioral activation. Neuroscience64:587–597.
Hu Y, Lund IV, Gravielle MC, Farb DH, Brooks-Kayal AR, Russek SJ(2008) Surface expression of GABAA receptors is transcriptionallycontrolled by the interplay of cAMP-response element-binding pro-tein and its binding partner inducible cAMP early repressor. J BiolChem 283:9328–9340.
Inui T, Shimura T, Yamamoto T (2007) The role of the ventral pallidumGABAergic system in conditioned taste aversion: effects of micro-injections of a GABAA receptor antagonist on taste palatability of aconditioned stimulus. Brain Res 1164:117–124.
Inui T, Yamamoto T, Shimura T (2009) GABAergic transmission in therat ventral pallidum mediates a saccharin palatability shift in con-ditioned taste aversion. Eur J Neurosci 30:110–115.
Jung MJ, Lippert B, Metcalf BW, Bohlen P, Schechter PJ (1977a)gamma-Vinyl GABA (4-amino-hex-5-enoic acid), a new selectiveirreversible inhibitor of GABA-T: effects on brain GABA metabolismin mice. J Neurochem 29:797–802.
Jung MJ, Lippert B, Metcalf BW, Schechter PJ, Bohlen P, SjoerdsmaA (1977b) The effect of 4-amino hex-5-ynoic acid (gamma-acety-lenic GABA, gammma-ethynyl GABA), a catalytic inhibitor of GABAtransaminase, on brain GABA metabolism in vivo. J Neurochem28:717–723.
Kessler RC, Berglund P, Demler O, Jin R, Koretz D, Merikangas KR,Rush AJ, Walters EE, Wang PS (2003) The epidemiology of majordepressive disorder: results from the National Comorbidity SurveyReplication (NCS-R). JAMA 289:3095–3105.
Khaled SM, Bulloch A, Exner DV, Patten SB (2009) Cigarette smoking,stages of change, and major depression in the Canadian popula-tion. Can J Psychiatry 54:204–208.
Krishnan V, Nestler EJ (2008) The molecular neurobiology of depres-sion. Nature 455:894–902.
Leach JP, Brodie MJ (1998) Tiagabine. Lancet 351:203–207.Levine AS, Morley JE (1981) Stress-induced eating in rats. Am J
Physiol 241:R72–R76.Li X, Li J, Peng XQ, Spiller K, Gardner EL, Xi ZX (2009) Metabotropic
glutamate receptor 7 modulates the rewarding effects of cocaine inrats: involvement of a ventral pallidal GABAergic mechanism. Neu-ropsychopharmacology 34:1783–1796.
Lloyd KG, Zivkovic B, Scatton B, Morselli PL, Bartholini G (1989) Thegabaergic hypothesis of depression. Prog Neuro-Psychopharma-col Biol Psychiatry 13:341–351.
Lucki I (1997) The forced swimming test as a model for core andcomponent behavioral effects of antidepressant drugs. BehavPharmacol 8:523–532.
Ma L, Song L, Radoi GE, Harrison NL (2004) Transcriptional regula-tion of the mouse gene encoding the alpha-4 subunit of the GABAAreceptor. J Biol Chem 279:40451–40461.
MacLean P (1985) Fiber systems of the forebrain. In: The rat nervoussystem, Vol. 1 (Paxinos G, ed), pp 417–436. Florida, USA: Aca-demic press.
Martens WH (2009) The role of self-complexity in reducing harmfulinsight among persons with schizophrenia. Theoretical and thera-peutic implications. Am J Psychother 63:53–68.
McFarland K, Kalivas PW (2001) The circuitry mediating cocaine-induced reinstatement of drug-seeking behavior. J Neurosci21:8655–8663.
Mhatre MC, Ticku MK (1994) Chronic GABA treatment downregulatesthe GABAA receptor alpha 2 and alpha 3 subunit mRNAS as wellas polypeptide expression in primary cultured cerebral corticalneurons. Brain Res 24:159–165.
Middeldorp J, van den Berge SA, Aronica E, Speijer D, Hol EM (2009)Specific human astrocyte subtype revealed by affinity purifiedGFAP antibody; unpurified serum cross-reacts with neurofilament-L in Alzheimer. PLoS One 4:e7663.
Mingote S, Font L, Farrar AM, Vontell R, Worden LT, Stopper CM, Port
RG, Sink KS, Bunce JG, Chrobak JJ, Salamone JD (2008) Nucleus
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146 145
accumbens adenosine A2A receptors regulate exertion of effort byacting on the ventral striatopallidal pathway. J Neurosci 28:9037–9046.
Mogenson GJ, Jones DL, Yim CY (1980) From motivation to action:functional interface between the limbic system and the motor sys-tem. Prog Neurobiol 14:69–97.
Mogenson GJ, Nielsen MA (1983) Evidence that an accumbens tosubpallidal GABAergic projection contributes to locomotor activity.Brain Res Bull 11:309–314.
Mogenson GJ, Yang CR (1991) The contribution of basal forebrain tolimbic-motor integration and the mediation of motivation to action.Adv Exp Med Biol 295:267–290.
Mombereau C, Kaupmann K, Froestl W, Sansig G, van der Putten H,Cryan JF (2004) Genetic and pharmacological evidence of a rolefor GABA(B) receptors in the modulation of anxiety- and antide-pressant-like behavior. Neuropsychopharmacology 29:1050–1062.
Mtchedlishvili Z, Kapur J (2006) High-affinity, slowly desensitizingGABAA receptors mediate tonic inhibition in hippocampal dentategranule cells. Mol Pharmacol 69:564–575.
Nestler EJ, Hyman SE (2010) Animal models of neuropsychiatricdisorders. Nat Neurosci 13:1161–1169.
Nusser Z, Sieghart W, Somogyi P (1998) Segregation of differentGABAA receptors to synaptic and extrasynaptic membranes ofcerebellar granule cells. J Neurosci 18:1693–1703.
O’Donnell P, Lavín A, Enquist LW, Grace AA, Card JP (1997) Inter-connected parallel circuits between rat nucleus accumbens andthalamus revealed by retrograde transynaptic transport of pseudo-rabies virus. J Neurosci 17:2143–2167.
Olsen RW, Sieghart W (2008) International Union of Pharmacology.LXX. Subtypes of gamma-aminobutyric acid(A) receptors: classifi-cation on the basis of subunit composition, pharmacology, andfunction. Update. Pharmacol Rev 60:243–260.
Patel S, Slater P (1988) Effects of GABA compounds injected into thesubpallidal regions of rat brain on nucleus accumbens evokedhyperactivity. Behav Neurosci 102:596–600.
Paxinos G, Watson C (2005) The rat brain in stereotaxic coordinates.San Diego, CA: Elsevier Academic Press.
Peng Z, Hauer B, Mihalek RM, Homanics GE, Sieghart W, Olsen RW,Houser CR (2002) GABA(A) receptor changes in delta subunit-deficient mice: altered expression of alpha4 and gamma2 subunitsin the forebrain. J Comp Neurol 446:179–197.
Peng Z, Huang CS, Stell BM, Mody I, Houser CR (2004) Alteredexpression of the delta subunit of the GABAA receptor in a mousemodel of temporal lobe epilepsy. J Neurosci 24:8629–8639.
Pérez J, Colasante C, Tucci S, Hernández L, Rada P (2000) Effects offeeding on extracellular levels of glutamate in the medial andlateral portion of the globus pallidus of freely moving rats. BrainRes 877:91–94.
Pirker S, Schwarzer C, Wieselthaler A, Sieghart W, Sperk G (2000)GABA(A) receptors: immunocytochemical distribution of 13 sub-units in the adult rat brain. Neuroscience 101:815–850.
Pláteník J, Balcar VJ, Yoneda Y, Mioduszewska B, Buchal R, HynekR, Kilianek L, Kuramoto N, Wilczynski G, Ogita K, Nakamura Y,Kaczmarek L (2005) Apparent presence of Ser133-phosphorylatedcyclic AMP response element binding protein (pCREB) in brainmitochondria is due to cross-reactivity of pCREB antibodies withpyruvate dehydrogenase. J Neurochem 95:1446–1460.
Platt KP, Zwartjes RE, Bristow DR (1996) The effect of GABA stimu-lation on GABAA receptor subunit protein and mRNA expression inrat cultured cerebellar granule cells. Br J Pharmacol 119:1393–1400.
Pliakas AM, Carlson RR, Neve RL, Konradi C, Nestler EJ, CarlezonWA Jr. (2001) Altered responsiveness to cocaine and increasedimmobility in the forced swim test associated with elevated cAMPresponse element-binding protein expression in nucleus accum-
bens. J Neurosci 21:7397–7403.Porsolt RD (1979) Animal model of depression. Biomedicine 30:139–140.
Rada P, Colasante C, Skirzewski M, Hernandez L, Hoebel B (2006)Behavioral depression in the swim test causes a biphasic, long-lasting change in accumbens acetylcholine release, with partialcompensation by acetylcholinesterase and muscarinic-1 rece-ptors. Neuroscience 141:67–76.
Rada P, Moreno SA, Tucci S, Gonzalez LE, Harrison T, Chau DT,Hoebel BG, Hernandez L (2003) Glutamate release in the nucleusaccumbens is involved in behavioral depression during thePORSOLT swim test. Neuroscience 119:557–565.
Rada P, Tucci S, Teneud L, Paez X, Perez J, Alba G, Garcia Y,Sacchettoni S, del Corral J, Hernandez L (1999) Monitoring gam-ma-aminobutyric acid in human brain and plasma microdialysatesusing micellar electrokinetic chromatography and laser-inducedfluorescence detection. J Chromatogr 735:1–10.
Richens A (1992) New drugs for epilepsy: a rapidly changing scene.Acta Neurol Scand Suppl 140:65–70.
Rowland NE, Antelman SM (1976) Stress-induced hyperphagia andobesity in rats: a possible model for understanding human obesity.Science 191:310–312.
Russek SJ, Bandyopadhyay S, Farb DH (2000) An initiator elementmediates autologous downregulation of the human type A gamma-aminobutyric acid receptor beta 1 subunit gene. Proc Natl AcadSci U S A 97:8600–8605.
Saxena NC, Macdonald RL (1994) Assembly of GABAA receptorsubunits: role of the delta subunit. J Neurosci 14:7077–7086.
Saxena NC, Macdonald RL (1996) Properties of putative cerebellargamma-aminobutyric acid A receptor isoforms. Mol Pharmacol49:567–579.
Sayin U, Timmerman W, Westerink BH (1995) The significance ofextracellular GABA in the substantia nigra of the rat during seizuresand anticonvulsant treatments. Brain Res 669:67–72.
Schechter PJ, Tranier Y, Jung MJ, Böhlen P (1977) Audiogenicseizure protection by elevated brain GABA concentration inmice: effects of gamma-acetylenic GABA and gamma-vinylGABA, two irreversible GABA-T inhibitors. Eur J Pharmacol45:319 –328.
Schwarzer C, Berresheim U, Pirker S, Wieselthaler A, Fuchs K,Sieghart W, Sperk G (2001) Distribution of the major gamma-aminobutyric acid(A) receptor subunits in the basal ganglia andassociated limbic brain areas of the adult rat. J Comp Neurol433:526 –549.
Semyanov A, Walker MC, Kullmann DM, Silver RA (2004) Tonicallyactive GABA A receptors: modulating gain and maintaining thetone. Trends Neurosci 27:262–269.
Shimura T, Imaoka H, Yamamoto T (2006) Neurochemical modulationof ingestive behavior in the ventral pallidum. Eur J Neurosci23:1596–1604.
Skirzewski M, Hernandez L, Schechter LE, Rada P (2010) Acutelecozotan administration increases learning and memory in ratswithout affecting anxiety or behavioral depression. PharmacolBiochem Behav 95:325–330.
Smith KR, McAinsh K, Chen G, Arancibia-Carcamo IL, Haucke V, YanZ, Moss SJ, Kittler JT (2008) Regulation of inhibitory synaptictransmission by a conserved atypical interaction of GABA(A) re-ceptor beta- and gamma-subunits with the clathrin AP2 adaptor.Neuropharmacology 55:844–850.
Smith KS, Berridge KC (2005) The ventral pallidum and hedonicreward: neurochemical maps of sucrose “liking” and food intake.J Neurosci 25:8637–8649.
Smith KS, Tindell AJ, Aldridge JW, Berridge KC (2009) Ventral palli-dum roles in reward and motivation. Behav Brain Res 196:155–167.
Smolders I, Khan GM, Lindekens H, Prikken S, Marvin CA, Manil J,Ebinger G, Michotte Y (1997) Effectiveness of vigabatrin againstfocally evoked pilocarpine-induced seizures and concomitant
changes in extracellular hippocampal and cerebellar glutamate, gam-
M. Skirzewski et al. / Neuroscience 196 (2011) 131–146146
ma-aminobutyric acid and dopamine levels, a microdialysis-electrocorticography study in freely moving rats. J Pharmacol ExpTher 283:1239–1248.
Steiger JL, Russek SJ (2004) GABAA receptors: building the bridgebetween subunit mRNAs, their promoters, and cognate transcrip-tion factors. Pharmacol Ther 101:259–281.
Stell BM, Brickley SG, Tang CY, Farrant M, Mody I (2003) Neuroactivesteroids reduce neuronal excitability by selectively enhancing tonicinhibition mediated by delta subunit-containing GABAA receptors.Proc Natl Acad Sci U S A 100:14439–14444.
Stevenson JR, Schroeder JP, Nixon K, Besheer J, Crews FT, HodgeCW (2009) Abstinence following alcohol drinking produces depres-sion-like behavior and reduced hippocampal neurogenesis in mice.Neuropsychopharmacology 34:1209–1222.
Stratford TR, Kelley AE, Simansky KJ (1999) Blockade of GABAAreceptors in the medial ventral pallidum elicits feeding in satiatedrats. Brain Res 825:199–203.
Strober LB, Arnett PA (2009) Assessment of depression in threemedically ill, elderly populations: Alzheimer’s disease, Parkinson’sdisease, and stroke. Clin Neuropsychol 23:205–230.
Tindell AJ, Smith KS, Peciña S, Berridge KC, Aldridge JW (2006)Ventral pallidum firing codes hedonic reward: when a bad tasteturns good. J Neurophysiol 96:2399–2409.
Torregrossa MM, Kalivas PW (2008) Neurotensin in the ventral palli-dum increases extracellular gamma-aminobutyric acid and differ-entially affects cue- and cocaine-primed reinstatement. J Pharma-col Exp Ther 325:556–566.
Torregrossa MM, Tang XC, Kalivas PW (2008) The glutamatergicprojection from the prefrontal cortex to the nucleus accumbenscore is required for cocaine-induced decreases in ventral pallidalGABA. Neurosci Lett 438:142–145.
Tretter V, Hauer B, Nusser Z, Mihalek RM, Höger H, Homanics GE,Somogyi P, Sieghart W (2001) Targeted disruption of the GABA(A)receptor delta subunit gene leads to an up-regulation of gamma 2subunit-containing receptors in cerebellar granule cells. J BiolChem 276:10532–10538.
van den Bos R, Cools AR (1991) Motor activity and the GABAA-receptor in the ventral pallidum/substantia innominata complex.Neurosci Lett 124:246–250.
van den Bos R, Cools AR (2003) Switching to cue-directed behavior:specific for ventral striatal dopamine but not ventral pallidum/sub-
stantia innominata GABA as revealed by a swimming-testprocedure in rats. Neuroscience 118:1141–1149.
Walaas I, Fonnum F (1979) The distribution and origin of glutamatedecarboxylase and choline acetyltransferase in ventral pallidumand other basal forebrain regions. Brain Res 177:325–336.
Wan Q, Man HY, Braunton J, Wang W, Salter MW, Becker L, Wang YT(1997) Modulation of GABAA receptor function by tyrosine phos-phorylation of beta subunits. J Neurosci 17:5062–5069.
Wang SH, Sun ZL, Guo YJ, Yuan Y, Yang BQ (2009) Diabetes impairshippocampal function via advanced glycation end product medi-ated new neuron generation in animals with diabetes-related de-pression. Toxicol Sci 111:72–79.
Weinberger AH, Maciejewski PK, McKee SA, Reutenauer EL, MazureCM (2009) Gender differences in associations between lifetimealcohol, depression, panic disorder, and posttraumatic stress dis-order and tobacco withdrawal. Am J Add 18:140–147.
Wheeler RA, Carelli RM (2006) The neuroscience of pleasure. Focuson “Ventral pallidum firing codes hedonic reward: when a bad tasteturns good”. J Neurophysiol 96:2175–2176.
Wisden W, Laurie DJ, Monyer H, Seeburg PH (1992) The distributionof 13 GABAA receptor subunit mRNAs in the rat brain. I. Telen-cephalon, diencephalon, mesencephalon. J Neurosci 12:1040–1062.
Wisniecki A, Correa M, Arizzi MN, Ishiwari K, Salamone JD (2003)Motor effects of GABA(A) antagonism in globus pallidus: studies oflocomotion and tremulous jaw movements in rats. Psychopharma-cology (Berl) 170:140–149.
Yoshikawa T, Watanabe A, Ishitsuka Y, Nakaya A, Nakatani N (2002)Identification of multiple genetic loci linked to the propensity for“behavioral despair” in mice. Genome Res 12:357–366.
Yu R, Follesa P, Ticku MK (1996) Down-regulation of the GABAreceptor subunits mRNA levels in mammalian cultured corticalneurons following chronic neurosteroid treatment. Brain Res41:163–168.
Zheng TM, Caruncho HJ, Zhu WJ, Vicini S, Ikonomovic S, GraysonDR, Costa E (1996) Chronic flumazenil alters GABA(A) receptorsubunit mRNA expression, translation product assembly and chan-nel function in neuronal cultures. J Pharmacol Exp Ther 277:
525–533.(Accepted 25 August 2011)(Available online 30 August 2011)
Copyright © 2022 FDOKUMEN




















