
Phagocytosis of Borrelia burgdorferi and Treponema pallidum Potentiates Innate Immune Activation and...
18
Published Ahead of Print 12 January 2007. 2007, 75(4):2046. DOI: 10.1128/IAI.01666-06. Infect. Immun. Justin D. Radolf Amanda L. Marzo, Christian H. Eggers, Juan C. Salazar and Meagan W. Moore, Adriana R. Cruz, Carson J. LaVake, Interferon Production Immune Activation and Induces Gamma Potentiates Innate Treponema pallidum and Borrelia burgdorferi Phagocytosis of http://iai.asm.org/content/75/4/2046 Updated information and services can be found at: These include: REFERENCES http://iai.asm.org/content/75/4/2046#ref-list-1 at: This article cites 80 articles, 41 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from on October 8, 2013 by guest http://iai.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Phagocytosis of Borrelia burgdorferi and Treponema pallidum Potentiates Innate Immune Activation and...

Published Ahead of Print 12 January 2007. 2007, 75(4):2046. DOI: 10.1128/IAI.01666-06. Infect. Immun.
Justin D. RadolfAmanda L. Marzo, Christian H. Eggers, Juan C. Salazar and Meagan W. Moore, Adriana R. Cruz, Carson J. LaVake, Interferon Production Immune Activation and Induces Gamma
Potentiates InnateTreponema pallidum and Borrelia burgdorferiPhagocytosis of
http://iai.asm.org/content/75/4/2046Updated information and services can be found at:
These include:
REFERENCEShttp://iai.asm.org/content/75/4/2046#ref-list-1at:
This article cites 80 articles, 41 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from
on October 8, 2013 by guest
http://iai.asm.org/
Dow
nloaded from

INFECTION AND IMMUNITY, Apr. 2007, p. 2046–2062 Vol. 75, No. 40019-9567/07/$08.00�0 doi:10.1128/IAI.01666-06Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Phagocytosis of Borrelia burgdorferi and Treponema pallidum PotentiatesInnate Immune Activation and Induces Gamma
Interferon Production�
Meagan W. Moore,1 Adriana R. Cruz,1 Carson J. LaVake,1 Amanda L. Marzo,1† Christian H. Eggers,1Juan C. Salazar,3 and Justin D. Radolf1,2*
Departments of Medicine1 and Genetics and Developmental Biology,2 University of Connecticut Health Center,Farmington, Connecticut 06030-3715, and Division of Pediatric Infectious Diseases, Department of Pediatrics,
Connecticut Children’s Medical Center, Hartford, Connecticut 061063
Received 18 October 2006/Returned for modification 17 November 2006/Accepted 2 January 2007
We examined the interactions of live and lysed spirochetes with innate immune cells. THP-1 monocytoid cellswere activated to comparable extents by live Borrelia burgdorferi and by B. burgdorferi and Treponema pallidumlysates but were poorly activated by live T. pallidum. Because THP-1 cells poorly internalized live spirochetes,we turned to an ex vivo peripheral blood mononuclear cell system that would more closely reflect spirochete-mononuclear phagocyte interactions that occur during actual infection. In this system, B. burgdorferi inducedsignificantly greater monocyte activation and inflammatory cytokine production than did borrelial lysates orT. pallidum, and only B. burgdorferi elicited gamma interferon (IFN-�) from NK cells. B. burgdorferi wasphagocytosed avidly by monocytes, while T. pallidum was not, suggesting that the enhanced response to live B.burgdorferi was due to phagocytosis of the organism. When cytochalasin D was used to block phagocytosis oflive B. burgdorferi, cytokine production decreased to levels comparable to those induced by B. burgdorferilysates, while the IFN-� response was abrogated altogether. In the presence of human syphilitic serum, T.pallidum was efficiently internalized and initiated responses resembling those observed with live B. burgdorferi,including the production of IFN-� by NK cells. Depletion of monocytes revealed that they were the primarysource of inflammatory cytokines, while dendritic cells (DCs) directed IFN-� production from innate lympho-cytes. Thus, phagocytosis of live spirochetes initiates cell activation programs in monocytes and DCs that differqualitatively and quantitatively from those induced at the cell surface by lipoprotein-enriched lysates. Thegreater stimulatory capacity of B. burgdorferi versus T. pallidum appears to be explained by the successfulrecognition and phagocytosis of B. burgdorferi by host cells and the ability of T. pallidum to avoid detection anduptake by virtue of its denuded outer membrane rather than by differences in surface lipoprotein expression.
Borrelia burgdorferi and Treponema pallidum are the etio-logic agents of Lyme disease and venereal syphilis, respec-tively. These multisystem, spirochetal diseases share many clin-ical and immunopathogenic features but also have someimportant differences. Although transmitted by different means(tick bite versus sexual contact), B. burgdorferi and T. pallidumeach cause the development of a characteristic lesion at the siteof inoculation, erythema migrans in Lyme disease and thechancre in syphilis, and both pathogens disseminate from thesesites to invade diverse organ systems (33, 63). In the case ofLyme disease, constitutional symptoms often accompany ery-thema migrans despite what appear to be extremely low spi-rochetal burdens (33, 68). In contrast, systemic symptoms areuncommon in syphilis prior to the secondary stage of the dis-ease, during which spirochetal burdens in blood and tissues arebelieved to be at their highest levels (44, 63). Both B. burgdor-feri and T. pallidum can cause insidious ongoing inflammationand long-term sequelae in affected individuals, although the
molecular mechanisms which enable these pathogens to persistare poorly understood and are the subjects of considerabledebate (33, 63).
Genomic analysis of B. burgdorferi and T. pallidum has re-vealed that these pathogens lack orthologs of known exotoxinsas well as the specialized secretory machinery required for thedelivery of noxious molecules into host cells (14, 25, 26).Therefore, it is now generally accepted that the morbidity andmortality associated with syphilis and Lyme disease are a directconsequence of the inflammatory responses elicited by theiretiologic agents (63, 81). B. burgdorferi and T. pallidum lack theclassical gram-negative constituent lipopolysaccharide (LPS),which typically stimulates immune cells by Toll-like receptor 4(TLR4) (2). Instead, they express numerous bacterial lipopro-teins (BLPs) (14, 25, 26), which activate innate immune cellsvia TLR2/1 heterodimers in a CD14-dependent manner (2–4,11, 35, 70, 80). Intradermal injection of synthetic lipoproteinanalogs (lipopeptides) in both animals and humans has con-firmed that these molecules can exert proinflammatory activi-ties in vivo and that they have the capacity to recruit diverseleukocyte subsets into tissues (67, 71). In addition to initiatingthe innate immune response, TLR-derived signals also areessential in directing adaptive immune responses. Engagementof TLR2 by lipoproteins induces dendritic cell (DC) matura-tion whereby antigen processing is enhanced and major histo-
* Corresponding author. Mailing address: Department of Medicine,University of Connecticut Health Center, 263 Farmington Avenue,Farmington, CT 06030-3715. Phone: (860) 679-8480. Fax: (860) 679-1358. E-mail: [email protected].
† Present address: Department of Immunology, Rush Medical Cen-ter, Chicago, IL 60612.
� Published ahead of print on 12 January 2007.
2046
compatibility complex and costimulatory molecules are up-regulated (10). Activation of DCs via TLR2 also has beenshown to be critical for Th1 polarization during the priming ofnaı̈ve T cells (72).
TLR-ligand interactions originally were studied as cell sur-face phenomena (18); it is now well documented, however, thatsome pathogen-associated molecular patterns (PAMPs), suchas prokaryotic DNA, engage their cognate TLRs only fromwithin intracellular compartments (1, 41). In addition, recep-tors such as TLR1, TLR2, and TLR6 that act at the cell surfacealso can be recruited to phagolysosomes, where they sample
vacuolar contents (61, 77). Several years ago, we reported thatlive T. pallidum is markedly less stimulatory for THP-1 mono-cytic cells than live B. burgdorferi (69). We attributed this dif-ference to the fact that B. burgdorferi surface lipoproteins aredirectly accessible to TLRs on monocytes, while cellular acti-vation by T. pallidum, which has no demonstrated surface-exposed lipoproteins (13, 64), likely requires the liberation ofsequestered PAMPs following degradation of the organismswithin phagolysosomes (69). In this study, we sought to clarifythe relative importance of cell surface versus intracellular ac-tivation by spirochetal pathogens. Contrary to results obtained
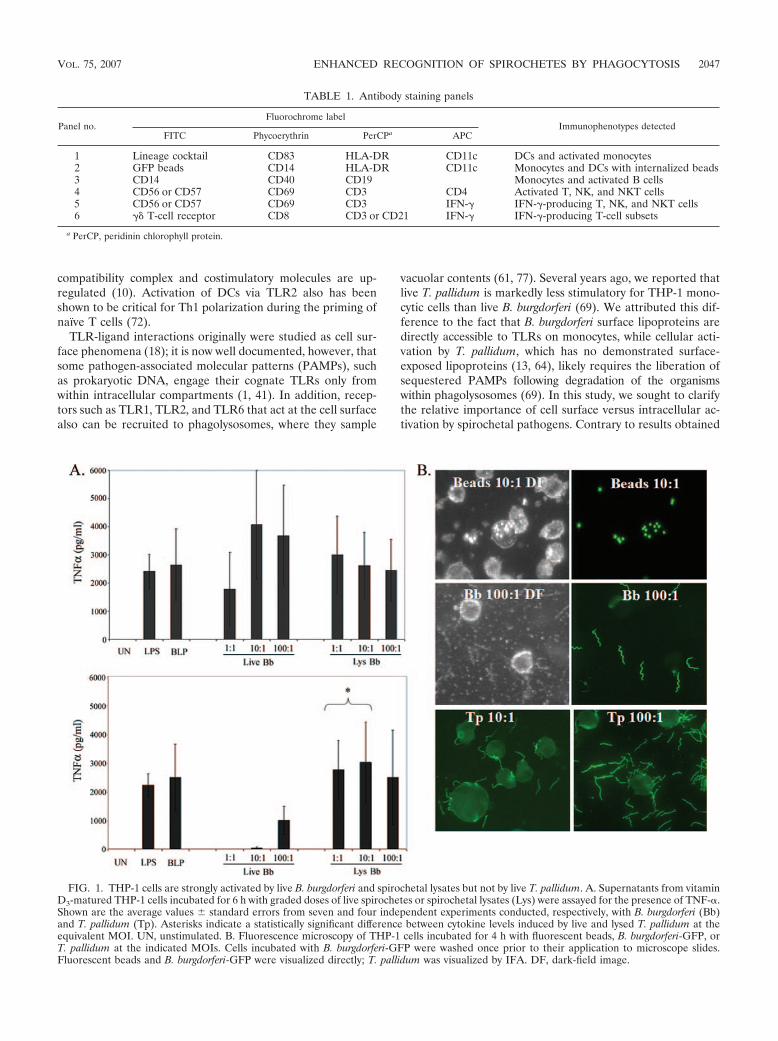
FIG. 1. THP-1 cells are strongly activated by live B. burgdorferi and spirochetal lysates but not by live T. pallidum. A. Supernatants from vitaminD3-matured THP-1 cells incubated for 6 h with graded doses of live spirochetes or spirochetal lysates (Lys) were assayed for the presence of TNF-�.Shown are the average values � standard errors from seven and four independent experiments conducted, respectively, with B. burgdorferi (Bb)and T. pallidum (Tp). Asterisks indicate a statistically significant difference between cytokine levels induced by live and lysed T. pallidum at theequivalent MOI. UN, unstimulated. B. Fluorescence microscopy of THP-1 cells incubated for 4 h with fluorescent beads, B. burgdorferi-GFP, orT. pallidum at the indicated MOIs. Cells incubated with B. burgdorferi-GFP were washed once prior to their application to microscope slides.Fluorescent beads and B. burgdorferi-GFP were visualized directly; T. pallidum was visualized by IFA. DF, dark-field image.
TABLE 1. Antibody staining panels
Panel no.Fluorochrome label
Immunophenotypes detectedFITC Phycoerythrin PerCPa APC
1 Lineage cocktail CD83 HLA-DR CD11c DCs and activated monocytes2 GFP beads CD14 HLA-DR CD11c Monocytes and DCs with internalized beads3 CD14 CD40 CD19 Monocytes and activated B cells4 CD56 or CD57 CD69 CD3 CD4 Activated T, NK, and NKT cells5 CD56 or CD57 CD69 CD3 IFN-� IFN-�-producing T, NK, and NKT cells6 �� T-cell receptor CD8 CD3 or CD21 IFN-� IFN-�-producing T-cell subsets
a PerCP, peridinin chlorophyll protein.
VOL. 75, 2007 ENHANCED RECOGNITION OF SPIROCHETES BY PHAGOCYTOSIS 2047

using THP-1 cells, we found by using an ex vivo peripheralblood mononuclear cell (PBMC) model that live B. burgdor-feri induced a response that differed quantitatively and qual-itatively from that of B. burgdorferi lysates and that thisaugmented response was dependent on phagocytosis of thespirochetes. While B. burgdorferi demonstrated greater stim-ulatory capacity than unopsonized T. pallidum, this differ-ence was essentially negated when opsonic antibody in hu-man syphilitic serum was employed to drive uptake oftreponemes into phagosomes. We also found that phagocy-tosis of spirochetes not only generated distinctive monocyteresponses but also induced DCs to stimulate gamma inter-feron (IFN-�) production by innate lymphocytes, potentiallysetting the stage for the intense Th1 polarization seen inboth syphilis and Lyme disease (68, 78). Thus, the greaterstimulatory capacity of B. burgdorferi versus T. pallidumappears to be explained by the successful recognition andphagocytosis of B. burgdorferi by host cells and the ability ofT. pallidum to avoid detection and uptake by virtue of itsdenuded outer membrane rather than by differences in sur-face lipoprotein expression.
MATERIALS AND METHODS
Human subjects. The University of Connecticut Health Center (UCHC) In-stitutional Review Board approved all protocols used here. Healthy volunteerswith no history of syphilis or Lyme disease were recruited by the General ClinicalResearch Center at UCHC. Written informed consent was obtained, and bloodwas collected by venipuncture. Volunteers were confirmed to be seronegative forsyphilis and/or Lyme disease by serological tests conducted in the UCHC clinicallaboratory.
Bacterial strains. Low-passage B. burgdorferi 297 was propagated in Barbour-Stoenner-Kelly medium containing 6% rabbit serum (Sigma). Spirochetes grownat 23°C were temperature shifted to 37°C; organisms were harvested at mid-logto late log phase (4 to 8 � 107/ml) by centrifugation at 8,000 � g, washed twicein CMRL (Gibco), and resuspended in RPMI medium (Gibco). For the micros-copy studies, virulent B. burgdorferi 297 engineered to express green fluorescentprotein (GFP) under the direction of the constitutive flaB promoter (20, 21) wasprepared as described above. T. pallidum subsp. pallidum (Nichols strain) waspropagated by intratesticular inoculation of adult New Zealand White rabbits aspreviously described (32) in strict accordance with a protocol approved by theUCHC Animal Care Committee. Treponemes were extracted from rabbit testesin CMRL, pelleted once by centrifugation at 14,000 � g, resuspended in RPMI,and enumerated by dark-field microscopy on a Petroff-Hausser counting cham-ber (Hausser Scientific).
Reagents. A synthetic bacterial lipohexapeptide (BLP) corresponding to the Nterminus of Escherichia coli murine lipoprotein (Bachem Bioscience) was used ata concentration of 10 �g/ml. LPS Re595 (Sigma) was suspended at 5 mg/ml in
FIG. 2. Enhanced activation of peripheral blood monocytes by live versus lysed B. burgdorferi. PBMCs were incubated for 8 h with live or lysed(Lys) B. burgdorferi (Bb), and monocytes were selectively gated on as described in Materials and Methods. A. Surface expression of CD40 andCD83. B. TNF-� and IL-1� levels in culture supernatants. Bars depict the means � standard errors from a minimum of four independentexperiments. Asterisks indicate a statistically significant difference between values induced by live and lysed B. burgdorferi at the equivalent MOI.UN, unstimulated.
2048 MOORE ET AL. INFECT. IMMUN.

FIG. 3. Live, but not lysed (Lys), B. burgdorferi (Bb) induces the production of IFN-� by NK cells. A. IFN-� levels in supernatants followingincubation of PBMCs with spirochetes for 8 h. Shown are the averages � standard errors from six experiments. B. Intracellular staining for IFN-�.Lymphocyte subpopulations within whole PBMCs were delineated via marker expression as described in Materials and Methods. The cytogramsshown are representative of three independent experiments. Numbers in the dot plots represent the percentage of cells positive for IFN-�. UN,unstimulated; TCR, T-cell receptor.
VOL. 75, 2007 ENHANCED RECOGNITION OF SPIROCHETES BY PHAGOCYTOSIS 2049

0.2% triethylamine, repurified as previously described (36), and used at a finalconcentration of 100 ng/ml. B. burgdorferi and T. pallidum lysates were preparedfrom live organisms by sonication. The equivalence of live and lysed spirochetepreparations was confirmed by sodium dodecyl sulfate-polyacrylamide gel elec-trophoresis and silver staining.
Cell preparation and stimulation. THP-1 cells were propagated in RPMIsupplemented with 10% fetal bovine serum (FBS) to a density of 1 � 106/ml andthen matured with 160 ng/ml vitamin D3 (Biomol) for 72 to 96 h. The maturedcells were washed in Hanks balanced salt solution (HBSS), plated at 1 � 106/mlin 24-well tissue culture plates, and stimulated with live or lysed spirochetes atmultiplicities of infection (MOIs) of 1, 10, and 100 or with LPS (100 ng/ml) orBLP (10 �g/ml). Cells were incubated for 6 h prior to collection of supernatantsfor cytokine analysis. For phagocytosis studies, 1-�m Fluoresbrite carboxylatedpolystyrene microspheres (Polysciences, Inc.) were used as controls. Bead dilu-tions were prepared to mimic the spirochete MOIs.
Human PBMCs were isolated by Ficoll density gradient centrifugation accord-ing to the instructions of the manufacturer (Amersham Biosciences). Cells werewashed three times with HBSS, counted on a hemocytometer, and resuspendedat 1 � 106/ml in RPMI–10% FBS. Cells were plated and stimulated with live orlysed spirochetes or control agonists as described above and incubated for 8 h at37°C with 5% CO2 prior to analysis. Selected samples were incubated with 10�M cytochalasin D (CytoD) (Sigma) or LysoTracker Red endosomal dye (Mo-lecular Probes). Some experiments with live T. pallidum included samples con-taining 10% heat-inactivated (56°C for 30 min) pooled human syphilitic serum(HSS) or normal human serum (NHS). When intracellular staining was per-formed, brefeldin A (Golgiplug; BD) was added at 4 h into the incubation at adilution of 1 �l/ml. As a control for the effects of opsonic antibody, beads were
incubated with 400 �g/ml of purified human immunoglobulin G (IgG) (Sigma)for 1 h at 37°C and then washed twice to remove unbound antibody. Antibodycoating was confirmed by counterstaining with anti-human IgG–allophycocyanin(APC) antibody (BD). Microscopy experiments were performed on PBMCsincubated with spirochetes or beads for 4 h. LPS levels in culture media andreagents were confirmed by Limulus assay (Cambrex) to be 10 pg/ml.
Cell Staining and flow cytometry. All antibody conjugates were purchasedfrom either BD Pharmingen or eBioscience. The antibody panels used in thefour-parameter staining of PBMCs are listed in Table 1. Unstimulated or stim-ulated PBMCs were harvested from tissue culture plates and washed once influorescence-activated cell sorting (FACS) buffer (phosphate-buffered saline,0.1% bovine serum albumin, 0.055% Na azide) in preparation for flow cytometry.Cells were incubated for 10 min at 4°C with 10 �g of purified human IgG (Sigma)for Fc receptor (FcR) blocking, followed by a 20-min incubation with fluoro-chrome-conjugated antibodies. After a final wash in FACS buffer, cells were fixed inFACS buffer plus 6.25% paraformaldehyde. Individual cell populations were selec-tively gated for analysis based on the following expression phenotypes: monocytes,CD14�; monocytoid DCs (mDCs), lineage HLA-DR� CD11c�; plasmacytoidDCs (pDCs), lineage HLA-DR� CD11c; NK cells, � CD56 or CD57� CD3;NKT cells, CD56 or CD57� CD3�; �� T cells, �� T-cell receptor� CD3�.
For intracellular cytokine staining (ICS), the cells were surface stained asdescribed above and then permeabilized in 250 �l of Cytofix/Cytoperm solution(BD) for 20 min at 4°C, followed by one wash in PermWash solution (FACSbuffer plus 0.5% saponin; Sigma). Cells were then resuspended in 50 �l ofPermWash plus 3 �l of anti-IFN-�–APC antibody and incubated for 30 min at4°C. After two washes in PermWash, the cells were resuspended in FACS bufferand analyzed immediately. Flow cytometry was performed using a FACSCalibur
FIG. 4. Live B. burgdorferi is a much more potent activator of human monocytes than live T. pallidum. A. Monocyte expression of CD40 andCD83 following an 8-h incubation of PBMCs with live B. burgdorferi (Bb) or live T. pallidum (Tp). Monocytes were gated as the CD14� population.B. TNF-� and IL-1� levels in culture supernatants. Data for B. burgdorferi represent a minimum of five experiments, including the four shown inFig. 2; data for T. pallidum represent at least three experiments. Statistical comparisons between B. burgdorferi and T. pallidum data sets wereperformed using an unpaired t test. Asterisks indicate a statistically significant difference between values induced by B. burgdorferi and T. pallidumat equivalent MOIs. UN, unstimulated.
2050 MOORE ET AL. INFECT. IMMUN.

dual-laser flow cytometer (BD); a minimum of 100,000 events and a maximum of500,000 events were collected, depending on the cell type of interest. Multipa-rameter files were analyzed using WinMDI v2.8 software (Joseph Trotter,Scripps Clinic).
Cytokine analysis. Simultaneous measurements of IFN-�, tumor necrosisfactor alpha (TNF-�), and interleukin-6 (IL-6) were performed using the humanTh1/Th2 Cytokine Bead Array (BD) per the manufacturer’s protocol. Alterna-tively, the human inflammatory Cytokine Bead Array kit was used to measureTNF-�, IL-6, IL-1�, and IL-12p70. IL-12p70 also was measured by ultrasensitiveenzyme-linked immunosorbent assay (Biosource).
Microscopy. Following a 4-h incubation with GFP-expressing B. burgdorferi (B.burgdorferi-GFP), cells were harvested, mounted onto a microscope slide, andviewed immediately on an Olympus Bx60 mercury arc lamp fluorescence micro-scope at �400 or �1,000 under oil immersion. Incorporation of GFP fluores-
FIG. 5. Internalization is a critical event for stimulation of cytokineproduction by live spirochetes. A. B. burgdorferi-GFP was incubated withPBMCs in the presence of LysoTracker Red for 4 h at an MOI of 10:1prior to fluorescence microscopy. The merged image (yellow) showscolocalization of GFP (green) with endosomal compartments (red). B.PBMCs were incubated with T. pallidum at an MOI of 10:1 in the pres-ence of LysoTracker Red for 4 h prior to IFA and fluorescence micros-copy. C. Effect of CytoD on the TNF-� response of PBMCs to live andlysed B. burgdorferi (Bb). Shown are results from one of two experiments;error bars represent the standard deviations for duplicate samples. D.CytoD inhibits uptake of live B. burgdorferi by monocytes. PBMCs werepreincubated with cytochalasin D (10 �g/ml) for 1 h prior to the additionof B. burgdorferi-GFP at various MOIs for 8 hours. The cells then werestained with CD14 and analyzed by flow cytometry to determine thepercentage of monocytes that had internalized spirochetes (GFP-positivecells) and the number of spirochetes taken up per monocyte (representedby the mean fluorescence intensity [MFI] of GFP-positive cells). Shownare the means � standard deviations from four independent experiments.Asterisks indicate P values of �0.05 for CytoD-treated and untreatedsamples. UN, unstimulated.
VOL. 75, 2007 ENHANCED RECOGNITION OF SPIROCHETES BY PHAGOCYTOSIS 2051

cence was visualized using a 470- to 490-nm excitation filter cube (Olympus). T.pallidum-cell associations were visualized by immunofluorescence assay (IFA) asdescribed by Lukehart and Miller (45). HSS pooled from five secondary syphilispatients was diluted 1:100 in CMRL–10% goat serum and used as the primaryantibody. NHS was used as an assay control. Secondary goat anti-human IgGantibody conjugated to fluorescein isothiocyanate (FITC) (Zymed) was used ata dilution of 1:250. Cells were visualized as described above for B. burgdorferi-GFP.
Depletion of monocytes from PBMCs by magnetic cell sorting. PBMCs weresuspended in ice-cold sorting buffer (phosphate-buffered saline, 2 mM EDTA,0.5% bovine serum albumin, pH 7.2) and incubated with metallic bead-conju-gated antibodies specific for CD14 or CD56 for the collection of monocytes orNK cells, respectively. Cells were then passed twice through separate ferromag-netic columns held in a magnetic field. The flowthrough was collected as the“depleted” cell fraction; the retained cells were harvested by flushing the columnoutside of the magnetic field. Sorted cells were washed twice with HBSS andresuspended in RPMI–10% FBS. The sorting magnet, separation columns, andall antibody conjugates were purchased from Miltenyi Biotech.
Statistical analysis. Statistical analysis was performed using Microsoft Excelsoftware. For comparison of each parameter, paired or unpaired Student t testswere performed, depending on whether the values were generated from withinthe same experiments or from parallel studies. All tests were two tailed andspecified to a 95% confidence interval. For each value, both the standard devi-ation and the standard error about the mean were calculated. P values of �0.05were considered significant.
RESULTS
Differential surface activation of human monocytic cellsby B. burgdorferi and T. pallidum. Initially, we used vitaminD3-matured THP-1 cells as a surrogate for human mono-cytes and compared their responses to individual PAMPs(LPS and BLP), live B. burgdorferi or T. pallidum, and son-icated preparations of spirochetes. As shown in Fig. 1A, liveB. burgdorferi induced a strong TNF-� response, which wasevident at the lowest MOI examined (1:1) and which wassimilar to the responses elicited by corresponding lysates. Incontrast, live T. pallidum proved to be a much poorer acti-vator of these cells in comparison to the equivalent doses oftreponemal lysates; an appreciable response to live T. pal-lidum was evident only at an MOI of 100:1. To determinewhether live spirochetes were taken up by the THP-1 cells,we used B. burgdorferi-GFP and IFA for T. pallidum. Al-though occasional binding of spirochetes to the cells wasapparent, neither B. burgdorferi nor T. pallidum appeared tobe internalized by this cell line (Fig. 1B). Thus, activation ofthese cells by spirochetes appears to occur predominantly orexclusively at the cell surface, while the difference in po-tency between the two spirochetal pathogens presumably isdue to the presence of abundant lipoproteins on the surfaceof B. burgdorferi and the paucity of these and other PAMPson the surface of T. pallidum (13, 64). As a test of generalphagocytic capacity, we incubated cells with fluorescentpolystyrene beads and found that our cell line was indeedcapable of phagocytosis (Fig. 1B).
Live B. burgdorferi is a far more potent immune cell activa-tor than lysates and live T. pallidum. The observation thatTHP-1 cells did not internalize spirochetes prompted us toutilize a system that would more accurately reflect spirochete-monocyte interactions that occur during infection. We rea-soned that an ex vivo model employing freshly isolated PBMCswould enable us to examine not only the interactions betweenspirochetes and native monocytes but also the responses ofother mononuclear cell types that spirochetes are likely to
2052 MOORE ET AL. INFECT. IMMUN.

encounter as they disseminate through blood and into tissues.We first used this system to compare the responses elicited bylive B. burgdorferi and borrelial lysates. Surprisingly in light ofour findings with THP-1 cells, fresh monocytes were morestrongly activated by live organisms as determined by surface
expression of CD40 and CD83 (Fig. 2A). Moreover, comparedto lysates, live B. burgdorferi induced much greater productionof TNF-� and IL-1� (Fig. 2B), as well as of IL-6 (data notshown). The results for IFN-� were particularly striking; al-though the relative amounts of this cytokine in culture super-
FIG. 6. Enhanced inflammatory cytokine production following opsonophagocytosis of T. pallidum. A. Effect of HSS on the uptake of T.pallidum. PBMCs were incubated with live T. pallidum at a density of 10:1 for 4 h in the presence of 10% HSS and LysoTracker Red before IFA.The merged image (yellow) demonstrates colocalization of T. pallidum material (green) with lysosomal vacuoles (red). B and C. Internalizationof T. pallidum (Tp) leads to enhanced macrophage activation (i.e., surface expression of CD40) (B) and greater production of TNF-� and IFN-�(C). Monocytes were gated as the CD14� population. Bar graphs represent the averages � standard deviations from at least three experiments.Asterisks indicate a statistically significant difference between unopsonized and opsonized T. pallidum at equivalent MOIs. D. Intracellular cytokinestaining for IFN-�. Lymphocyte subpopulations within whole PBMCs were delineated via marker expression as described in Materials andMethods. Numbers in the dot plots represent the percentage of cells positive for IFN-�; cytograms shown are representative of three experiments.TCR, T-cell receptor; PerCP, peridinin chlorophyll protein; UN, unstimulated.
VOL. 75, 2007 ENHANCED RECOGNITION OF SPIROCHETES BY PHAGOCYTOSIS 2053

natants varied widely between different cell donors, it wasconsistently and reproducibly detected only in response to liveB. burgdorferi (Fig. 3A). ICS revealed that NK cells were theprimary producers of IFN-� in response to B. burgdorferi (Fig.3B); also evident is the high degree of sensitivity of ICS fordetecting production of IFN-� in minor lymphocyte popula-tions. In contrast to live B. burgdorferi, which elicited a stronginflammatory response even at a spirochete-to-cell ratio of 1:1,monocyte activation and cytokine production in response tolive T. pallidum were significantly above background only atMOIs of 10:1 or greater. Furthermore, compared to B. burg-dorferi, live T. pallidum induced significantly less surface ex-pression of CD40 and CD83 by monocytes (Fig. 4A), de-creased production of TNF-� and IL-1� (Fig. 4B) and IL-6(data not shown), and no IFN-� (see Fig. 6D, left panels).Duplicate head-to-head comparisons of B. burgdorferi and T.pallidum confirmed that the difference in potency between thelive organisms was not due to experiment-to-experiment vari-ation (not shown).
Phagocytosis enhances monocyte and cytokine responses toviable spirochetes. It has been reported that monocytes avidlyphagocytose B. burgdorferi (8, 54), while the uptake of T. pal-lidum is inefficient in the absence of opsonins (45). We there-fore hypothesized that the augmented response to live B. burg-dorferi, compared to lysates and live T. pallidum, resulted fromuptake of B. burgdorferi by monocytes and liberation of borre-lial PAMPs following degradation within phagolysosomes.When PBMCs were incubated with B. burgdorferi-GFP, weobserved numerous vacuoles containing degraded organisms,but no intact spirochetes, inside a substantial fraction of thecells (Fig. 5A); colocalization of GFP fluorescence with Lyso-Tracker dye revealed that the digested borreliae were insidephagolysosomal vacuoles (Fig. 5A). In contrast, we were un-able to detect uptake of T. pallidum by IFA (Fig. 5B). Onaverage, 10% of the PBMCs contained ingested spirochetes, apercentage highly similar to the average monocyte compositionof the mixture determined by FACS analysis (12%). Flowcytometry subsequently confirmed that monocytes avidly inter-nalize B. burgdorferi-GFP (see Fig. 9, top row). The cytocha-lasins have been used extensively to inhibit phagocytosis ofmany different types of bacteria, and they also have beenshown to have no effect on the production of cytokines inresponse to agonists acting at the cell surface (15, 28, 56, 66).When CytoD was used to block phagocytosis of live B. burg-dorferi, production of proinflammatory cytokines decreased tolevels comparable to those induced by B. burgdorferi lysates,while the IFN-� response was abrogated altogether (Fig. 5C).Flow cytometry confirmed that CytoD at the dosage used (10�g/ml) virtually eliminated the uptake of B. burgdorferi-GFP bymonocytes at an MOI of 10:1 and markedly inhibited uptake ofspirochetes at the highest MOI (100:1) employed. (Fig. 5D).
In the rabbit model of experimental syphilis, ingestion andclearance of T. pallidum by macrophages are markedly facili-tated by the addition of rabbit immune serum but can beblocked by cytochalasin B (6, 43, 45); the ability of antibodiesin HSS to opsonize T. pallidum, however, has not been re-ported. Based on our results with B. burgdorferi, we hypothe-sized that incubation of treponemes with HSS would promotetheir internalization by PBMCs and that increased uptakewould result in enhanced immune cell activation. Indeed, as
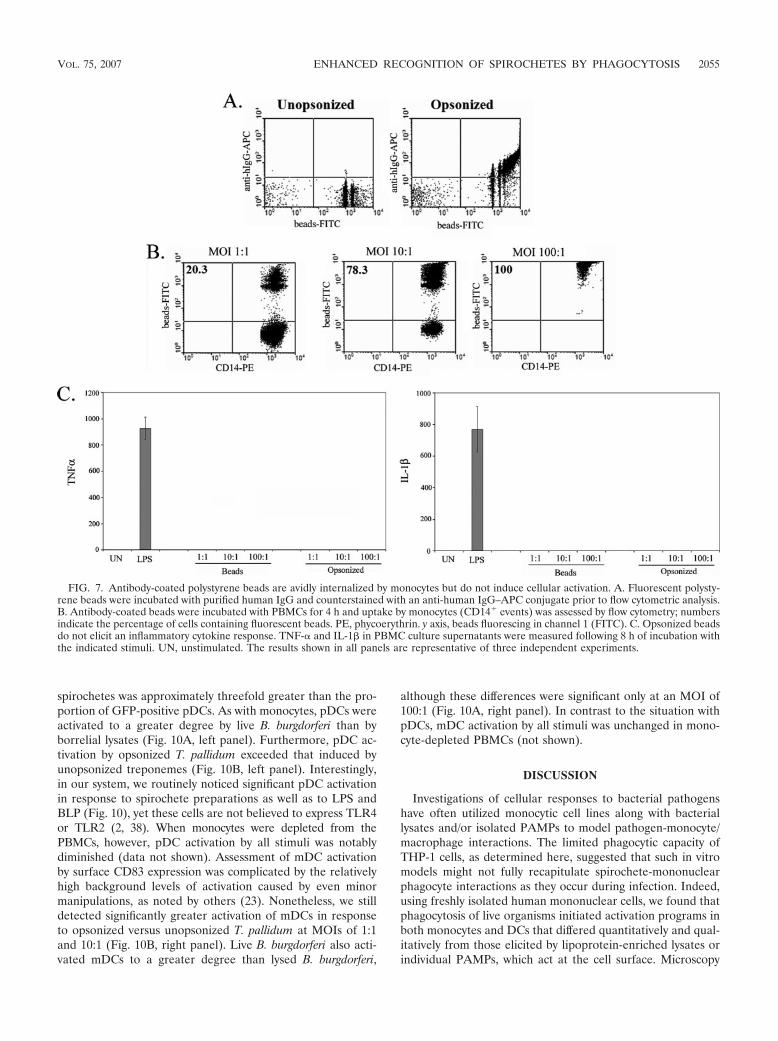
shown in Fig. 6A, degraded treponemes within phagolyso-somes were readily visualized when T. pallidum was incubatedwith PBMCs in the presence of 10% heat-inactivated HSS butnot heat-inactivated NHS (not shown). Enumeration of resid-ual spirochetes was consistent with the microscopy data; lessthan one-half of the input treponemes were recovered fromcell suspensions incubated at an MOI of 10 or 100 with HSS,while spirochete counts in cells incubated with NHS were es-sentially unchanged from input values (data not shown). Op-sonophagocytosis of T. pallidum markedly increased macro-phage activation as assessed by the surface expression of CD40(Fig. 6B) and CD83 (not shown) and the secretion of TNF-�(Fig. 6C), IL-6 (not shown), and IL-1� (not shown). The denovo induction of IFN-� by opsonized T. pallidum (Fig. 6C andD) further differentiated this response from that induced byunopsonized treponemes. The IFN-� response to low MOIs ofT. pallidum resembled that induced by B. burgdorferi in that itwas mediated primarily by NK cells (compare Fig. 6D with3D). High concentrations of opsonized T. pallidum, however,induced the production of this cytokine from a broader spec-trum of innate lymphocytes, namely, NK cells, NKT cells, and�� T cells (Fig. 6D). A small number of CD3� CD8� cells,likely representing CD8� �� and/or NKT cells (or iNKT cells)(37, 40), also produced IFN-� in response to high levels ofopsonized T. pallidum (Fig. 6D, bottom row). To confirm thatthe responses observed were due to uptake and degradation ofT. pallidum and not FcR signaling, we incubated PBMCs withcarboxylate beads coated with human IgG; opsonized beadswere highly internalized but did not induce cytokines (Fig. 7).
The enhanced response to internalized spirochetes is medi-ated by both monocytes and DCs. Our PBMC system containsseveral types of mononuclear phagocytes (monocytes, mDCs[CD11c�], and pDCs [CD11c]), with the capacity to activateinnate lymphocytes by contact-dependent and/or -independentmechanisms (29, 48, 73). To determine if monocytes stimulateIFN-� production in response to live spirochetes, we depletedthe CD14� cells from the mixture via magnetic cell sorting,typically achieving an absorption efficiency of �99%. Mono-cyte-depleted PBMCs stimulated with live B. burgdorferi (Fig.8A) or opsonized T. pallidum (Fig. 8B) produced dramaticallyless TNF-�, IL-1�, and IL-6 (not shown) than whole PBMCs,demonstrating conclusively that monocytes were the primarysource of these proinflammatory cytokines. In contrast, theIFN-� response elicited by B. burgdorferi and T. pallidum (Fig.8A and B) was unchanged by the depletion of monocytes fromPBMCs, indicating that DCs stimulated production of thiscytokine from innate lymphocytes. This conclusion was but-tressed by the observation that purified NK cells, incubatedalone or in coculture with purified monocytes, did not produceIFN-� in response to live B. burgdorferi (data not shown).Interestingly, using ultrasensitive cytokine assays, we were un-able to detect IL-12p70, a cytokine known to stimulate NK and�� T cells (9, 51), even in response to high MOIs of B. burg-dorferi or opsonized T. pallidum (data not shown).
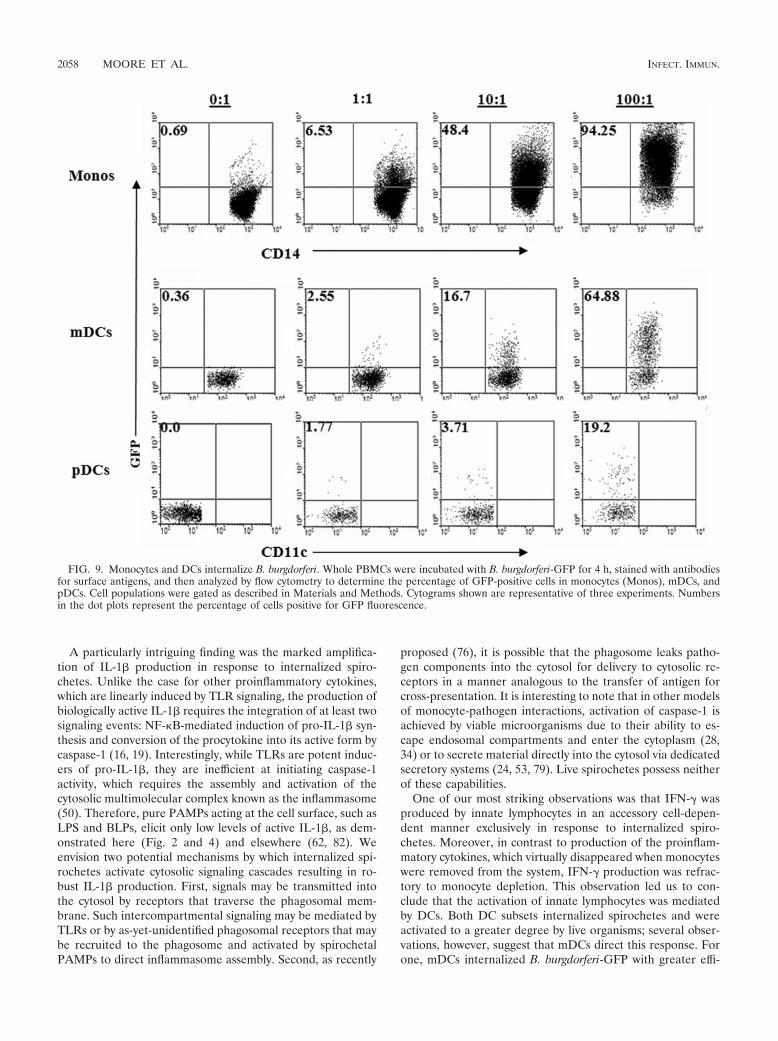
To shed further light on how DCs interact with B. burgdorferiand T. pallidum, we characterized their capacity to internalizespirochetes and their subsequent activation patterns. When weused B. burgdorferi-GFP to track the uptake of spirochetes byDCs, we were able to detect fluorescence in both mDCs andpDCs (Fig. 9). Notably, the percentage of mDCs containing
2054 MOORE ET AL. INFECT. IMMUN.
spirochetes was approximately threefold greater than the pro-portion of GFP-positive pDCs. As with monocytes, pDCs wereactivated to a greater degree by live B. burgdorferi than byborrelial lysates (Fig. 10A, left panel). Furthermore, pDC ac-tivation by opsonized T. pallidum exceeded that induced byunopsonized treponemes (Fig. 10B, left panel). Interestingly,in our system, we routinely noticed significant pDC activationin response to spirochete preparations as well as to LPS andBLP (Fig. 10), yet these cells are not believed to express TLR4or TLR2 (2, 38). When monocytes were depleted from thePBMCs, however, pDC activation by all stimuli was notablydiminished (data not shown). Assessment of mDC activationby surface CD83 expression was complicated by the relativelyhigh background levels of activation caused by even minormanipulations, as noted by others (23). Nonetheless, we stilldetected significantly greater activation of mDCs in responseto opsonized versus unopsonized T. pallidum at MOIs of 1:1and 10:1 (Fig. 10B, right panel). Live B. burgdorferi also acti-vated mDCs to a greater degree than lysed B. burgdorferi,
although these differences were significant only at an MOI of100:1 (Fig. 10A, right panel). In contrast to the situation withpDCs, mDC activation by all stimuli was unchanged in mono-cyte-depleted PBMCs (not shown).
DISCUSSION
Investigations of cellular responses to bacterial pathogenshave often utilized monocytic cell lines along with bacteriallysates and/or isolated PAMPs to model pathogen-monocyte/macrophage interactions. The limited phagocytic capacity ofTHP-1 cells, as determined here, suggested that such in vitromodels might not fully recapitulate spirochete-mononuclearphagocyte interactions as they occur during infection. Indeed,using freshly isolated human mononuclear cells, we found thatphagocytosis of live organisms initiated activation programs inboth monocytes and DCs that differed quantitatively and qual-itatively from those elicited by lipoprotein-enriched lysates orindividual PAMPs, which act at the cell surface. Microscopy
FIG. 7. Antibody-coated polystyrene beads are avidly internalized by monocytes but do not induce cellular activation. A. Fluorescent polysty-rene beads were incubated with purified human IgG and counterstained with an anti-human IgG–APC conjugate prior to flow cytometric analysis.B. Antibody-coated beads were incubated with PBMCs for 4 h and uptake by monocytes (CD14� events) was assessed by flow cytometry; numbersindicate the percentage of cells containing fluorescent beads. PE, phycoerythrin. y axis, beads fluorescing in channel 1 (FITC). C. Opsonized beadsdo not elicit an inflammatory cytokine response. TNF-� and IL-1� in PBMC culture supernatants were measured following 8 h of incubation withthe indicated stimuli. UN, unstimulated. The results shown in all panels are representative of three independent experiments.
VOL. 75, 2007 ENHANCED RECOGNITION OF SPIROCHETES BY PHAGOCYTOSIS 2055

studies demonstrated that internalized B. burgdorferi and T.pallidum were completely degraded within phagolysosomes,suggesting that the augmented responses of phagocytic cellsare triggered by the release of spirochetal products withinmaturing phagosomes.
Amplification of the proinflammatory cytokine response tospirochetes from within the phagosome may occur by a numberof nonexclusive mechanisms. TLR responses initiated fromwithin the phagosome may be augmented by a high concen-tration of PAMPs within a relatively small space. In addition,TLR2 and its cooperative pairing partners TLR1 and TLR6have been shown to be actively recruited to the phagosome (61,77), and this may lead to a greater receptor density in thiscompartment than at the cell surface. Phagolysosomes aredynamic organelles that mature, transmit signals to the cytosol,and direct their own interactions with other cellular compart-
ments via a complex set of membranous proteins on the or-ganelle surface (76). Thus, the presence of specific “docking”molecules may provide a particularly effective scaffold for TLRmachinery. Yet another possibility is that TLR signaling path-ways distinct from those at the cell surface are activated withinthe phagosome upon the liberation of sequestered PAMPs bybacterial disruption. The fact that some TLRs, such as TLR9,signal only from within phagolysosomes supports this conjec-ture (1, 41). Several spirochetal constituents, such as flagellin,DNA, and lipoproteins in the case of T. pallidum, would beavailable to cognate pattern recognition receptors only follow-ing spirochetal degradation. The activation of multiple TLRsmay then lead to synergistic amplification of the signaling cas-cade, resulting in greater cytokine secretion. The phenomenonof TLR cooperation to generate immune responses is becom-ing increasingly recognized (5, 59).
FIG. 8. Depletion of monocytes from PBMCs markedly diminishes production of TNF-� and IL-6, but not IFN-�, following incubation for 8 hwith live B. burgdorferi (Bb) (A) or opsonized T. pallidum (Tp) (B). Data for TNF-� and IL-1� depict the averages � standard deviations fromthree (B. burgdorferi) or two (T. pallidum) individual experiments. Dot plots for IFN-� are representative of at least two experiments; numbers inthe dot plots denote the percentage of cells positive for IFN-�. Lymphocyte subpopulations within whole PBMCs were delineated via markerexpression as described in Materials and Methods. Only NK cells are shown for B. burgdorferi experiments in panel A. CD4 T cells did not produceIFN-� and are not shown (B). UN, unstimulated; TCR, T-cell receptor; PerCP, peridinin chlorophyll protein.
2056 MOORE ET AL. INFECT. IMMUN.
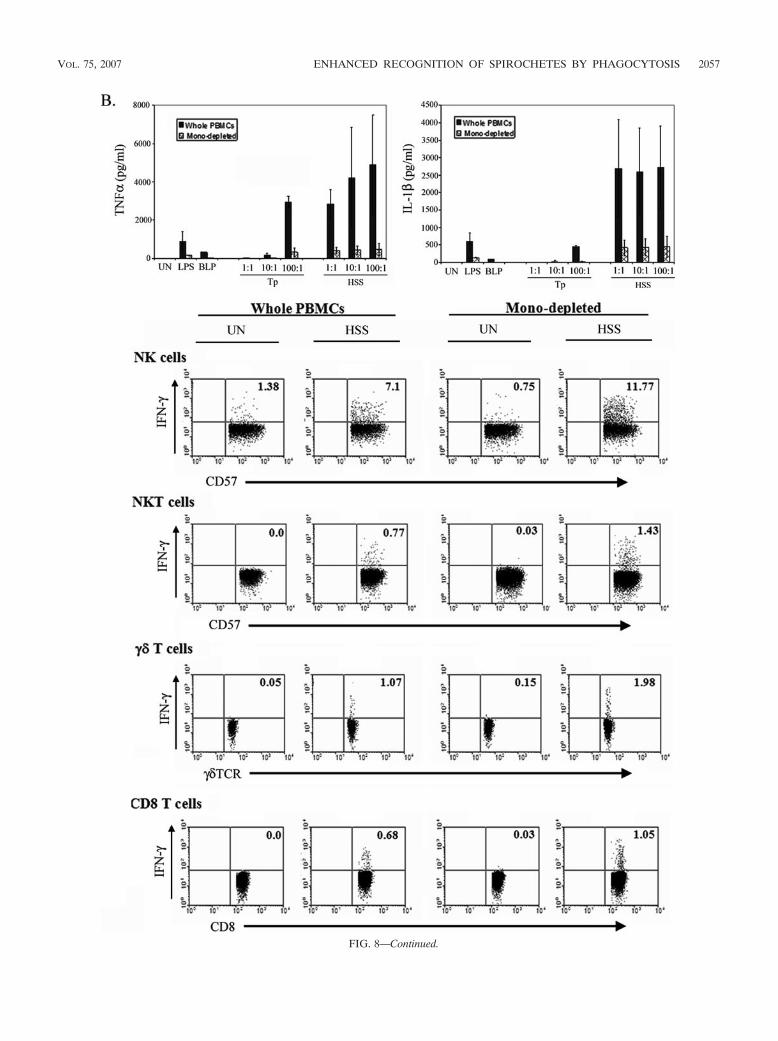
FIG. 8—Continued.
VOL. 75, 2007 ENHANCED RECOGNITION OF SPIROCHETES BY PHAGOCYTOSIS 2057
A particularly intriguing finding was the marked amplifica-tion of IL-1� production in response to internalized spiro-chetes. Unlike the case for other proinflammatory cytokines,which are linearly induced by TLR signaling, the production ofbiologically active IL-1� requires the integration of at least twosignaling events: NF-�B-mediated induction of pro-IL-1� syn-thesis and conversion of the procytokine into its active form bycaspase-1 (16, 19). Interestingly, while TLRs are potent induc-ers of pro-IL-1�, they are inefficient at initiating caspase-1activity, which requires the assembly and activation of thecytosolic multimolecular complex known as the inflammasome(50). Therefore, pure PAMPs acting at the cell surface, such asLPS and BLPs, elicit only low levels of active IL-1�, as dem-onstrated here (Fig. 2 and 4) and elsewhere (62, 82). Weenvision two potential mechanisms by which internalized spi-rochetes activate cytosolic signaling cascades resulting in ro-bust IL-1� production. First, signals may be transmitted intothe cytosol by receptors that traverse the phagosomal mem-brane. Such intercompartmental signaling may be mediated byTLRs or by as-yet-unidentified phagosomal receptors that maybe recruited to the phagosome and activated by spirochetalPAMPs to direct inflammasome assembly. Second, as recently
proposed (76), it is possible that the phagosome leaks patho-gen components into the cytosol for delivery to cytosolic re-ceptors in a manner analogous to the transfer of antigen forcross-presentation. It is interesting to note that in other modelsof monocyte-pathogen interactions, activation of caspase-1 isachieved by viable microorganisms due to their ability to es-cape endosomal compartments and enter the cytoplasm (28,34) or to secrete material directly into the cytosol via dedicatedsecretory systems (24, 53, 79). Live spirochetes possess neitherof these capabilities.
One of our most striking observations was that IFN-� wasproduced by innate lymphocytes in an accessory cell-depen-dent manner exclusively in response to internalized spiro-chetes. Moreover, in contrast to production of the proinflam-matory cytokines, which virtually disappeared when monocyteswere removed from the system, IFN-� production was refrac-tory to monocyte depletion. This observation led us to con-clude that the activation of innate lymphocytes was mediatedby DCs. Both DC subsets internalized spirochetes and wereactivated to a greater degree by live organisms; several obser-vations, however, suggest that mDCs direct this response. Forone, mDCs internalized B. burgdorferi-GFP with greater effi-
FIG. 9. Monocytes and DCs internalize B. burgdorferi. Whole PBMCs were incubated with B. burgdorferi-GFP for 4 h, stained with antibodiesfor surface antigens, and then analyzed by flow cytometry to determine the percentage of GFP-positive cells in monocytes (Monos), mDCs, andpDCs. Cell populations were gated as described in Materials and Methods. Cytograms shown are representative of three experiments. Numbersin the dot plots represent the percentage of cells positive for GFP fluorescence.
2058 MOORE ET AL. INFECT. IMMUN.

ciency than pDCs, consistent with the current paradigm thatthese cells are more phagocytic and function as the primaryantigen-processing and -presenting cells for the initiation ofadaptive responses (7, 17). Second, mDCs were highly acti-vated in a direct manner by live spirochetes. In contrast, thestrong pDC response to live spirochetes was diminished by thedepletion of monocytes, while the IFN-� response remainedunchanged, demonstrating a discordance between pDC activa-tion and IFN-� induction in our ex vivo system. Finally, mDCsare well known to activate NK, NKT, and �� T cells by a widevariety of mechanisms (27, 29, 40, 57, 58). Nonetheless, pDCshave been shown to activate NK cells (65) and therefore havenot been ruled out as a stimulator of IFN-� production in ourex vivo system. Furthermore, pDC activation in response to B.burgdorferi and T. pallidum is reported to occur in vivo (68,68a).
The clinical responses observed in both syphilis and Lymedisease are intensely Th1 polarized (68, 78). IFN-� producedby DC-activated NK (and other) cells has been shown to pro-vide a reciprocal signal to DCs for the induction of IL-12,ultimately generating a cytokine milieu that promotes Th1development in T cells as they are primed in the lymph nodes(46, 49, 74). Thus, the production of IFN-� by innate lympho-
cytes, as observed in our study, could provide the Th1 primingsignal during spirochetal infection. As IFN-� was elicited onlyby live organisms, and not by lysates or individual PAMPs, theinduction of this cytokine in innate lymphocytes could act as agauge at the innate-adaptive interface for the immune systemto differentiate between the presence of proinflammatory ago-nists and viable pathogens. There is considerable debate aboutthe benefit of a Th1 response in the case of Lyme disease. Th1responses are classically believed to target intracellular patho-gens by providing a signal for enhanced bactericidal killing bymacrophages in the form of IFN-�. As B. burgdorferi is anextracellular pathogen that is easily killed by monocytes, asshown here, and by macrophages in the absence of IFN-� (55),the advantage of a Th1 response in the case of B. burgdorferiinfection is unclear. Studies with the murine model have dem-onstrated no requirement for IFN-� in bacterial clearance ordisease susceptibility (12, 30) but have suggested that the Th1response is actually deleterious in the course of Lyme disease(52). On the other hand, a study that carefully examined thekinetics of Th cell polarization in Lyme arthritis-susceptibleand -resistant mice demonstrated that the Th1 response in factcorrelated with better control of infection early in disease andwas detrimental only when prolonged (39). IFN-� has been
FIG. 10. Activation of DCs by live B. burgdorferi and T. pallidum. Whole PBMCs were incubated with the indicated stimuli, and CD83expression on gated pDCs or mDCs in response to live versus lysed (Lys) B. burgdorferi (Bb) (A) or to unopsonized versus opsonized T. pallidum(Tp) (B) was determined. Shown are the averages � standard errors for four experiments. Asterisks indicate a statistically significant differencewhen comparing live and lysed B. burgdorferi or opsonized and unopsonized T. pallidum at equivalent MOIs. UN, unstimulated.
VOL. 75, 2007 ENHANCED RECOGNITION OF SPIROCHETES BY PHAGOCYTOSIS 2059

shown to enhance monocyte effector functions in response tospirochetal constituents (47) and so may function to expeditethe recruitment of key immune cells to the site of infectionduring the initial phases of the disease. In contrast to the casefor Lyme disease, the requirement for opsonic antibody helpsto explain the benefit of a Th1 response in syphilis. The IgGisotypes generated in a Th1 context, such as IgG2a (75), arebound with a high affinity by activating Fc receptors (Fc�RIand Fc�RIIIa) but poorly by the inhibitory Fc�RIIb (22, 60).Furthermore, IFN-� promotes the process of opsonophagocy-tosis via the upregulation of activating Fc�Rs and the down-regulation of the inhibitory Fc�Rs on monocytes (42).
The work presented here has helped to clarify the mecha-nisms by which B. burgdorferi and T. pallidum initiate innateand adaptive immune responses. Taken collectively, our datademonstrate that the difference in immunostimulatory capacitybetween B. burgdorferi and T. pallidum is a result of dramati-cally different levels of phagocytosis of these two organisms,not the relative abundance of surface-expressed lipoproteinsper se as we previously proposed (69). Presumably, live B.burgdorferi displays at least one moiety on its surface, which T.pallidum lacks, that promotes recognition and phagocytosis byhost cells. When the host counters this immune evasion strat-egy via the production of opsonic antibody, the efficiency of T.pallidum internalization is dramatically enhanced, leading tocellular and cytokine responses similar to those generated byB. burgdorferi. FcRs are known to initiate numerous signalingpathways and are reported to downregulate immune responses(31). Our data indicate, however, that the primary purpose ofFcRs is to deliver T. pallidum to phagolysosomes for PAMP-mediated recognition, rather than to modulate the pathogen-specific immune response. The delayed onset of constitutionalsymptoms in syphilis, compared to Lyme disease, may reflectthe fact that phagocytosis of T. pallidum and the subsequentgeneration of inflammatory responses do not occur efficientlyuntil opsonic antibody is generated as part of the adaptiveimmune response. The intrinsic ability of T. pallidum to evadephagocytosis highlights how exquisitely adapted it is as a hu-man pathogen.
ACKNOWLEDGMENTS
We thank Morgan LaVake, Cynthia Gonzales, and Ken Bourell forsuperb technical support; Melissa Caimano for assistance with passag-ing of B. burgdorferi; Timothy Sellati for insightful comments regardingthe manuscript; and Sheila Lukehart for thoughtful comments anddiscussions.
This work was supported by Public Health Service grants AI-26756,AI-29735, and AI-38894 to J.D.R. and by General Clinical ResearchCenter grant M01RR06192 from the National Institutes of Health.
REFERENCES
1. Ahmad-Nejad, P., H. Hacker, M. Rutz, S. Bauer, R. M. Vabulas, and H.Wagner. 2002. Bacterial CpG-DNA and lipopolysaccharides activate Toll-like receptors at distinct cellular compartments. Eur. J. Immunol. 32:1958–1968.
2. Akira, S., S. Uematsu, and O. Takeuchi. 2006. Pathogen recognition andinnate immunity. Cell 124:783–801.
3. Alexopoulou, L., V. Thomas, M. Schnare, Y. Lobet, J. Anguita, R. T. Schoen,R. Medzhitov, E. Fikrig, and R. A. Flavell. 2002. Hyporesponsiveness tovaccination with Borrelia burgdorferi OspA in humans and in TLR1- andTLR2-deficient mice. Nat. Med. 8:878–884.
4. Aliprantis, A. O., R. B. Yang, M. R. Mark, S. Suggett, B. Devaux, J. D.Radolf, G. R. Klimpel, P. Godowski, and A. Zychlinsky. 1999. Cell activationand apoptosis by bacterial lipoproteins through Toll-like receptor-2. Science285:736–739.
5. Bafica, A., C. A. Scanga, C. G. Feng, C. Leifer, A. Cheever, and A. Sher. 2005.TLR9 regulates Th1 responses and cooperates with TLR2 in mediatingoptimal resistance to Mycobacterium tuberculosis. J. Exp. Med. 202:1715–1724.
6. Baker-Zander, S. A., J. M. Shaffer, and S. A. Lukehart. 1993. Characteriza-tion of the serum requirement for macrophage-mediated killing of Trepo-nema pallidum ssp. pallidum: relationship to the development of opsonizingantibodies. FEMS Immunol. Med. Microbiol. 6:273–279.
7. Banchereau, J., F. Briere, C. Caux, J. Davoust, S. Lebecque, Y. J. Liu, B.Pulendran, and K. Palucka. 2000. Immunobiology of dendritic cells. Annu.Rev. Immunol. 18:767–811.
8. Benach, J. L., H. B. Fleit, G. S. Habicht, J. L. Coleman, E. M. Bosler, andB. P. Lane. 1984. Interactions of phagocytes with the Lyme disease spiro-chete: role of the Fc receptor. J. Infect. Dis. 150:497–507.
9. Biron, C. A., K. B. Nguyen, G. C. Pien, L. P. Cousens, and T. P. Salazar-Mather. 1999. Natural killer cells in antiviral defense: function and regulation byinnate cytokines. Annu. Rev. Immunol. 17:189–220.
10. Blander, J. M., and R. Medzhitov. 2006. Toll-dependent selection of micro-bial antigens for presentation by dendritic cells. Nature 440:808–812.
11. Brightbill, H. D., D. H. Libraty, S. R. Krutzik, R. B. Yang, J. T. Belisle, J. R.Bleharski, M. Maitland, M. V. Norgard, S. E. Plevy, S. T. Smale, P. J.Brennan, B. R. Bloom, P. J. Godowski, and R. L. Modlin. 1999. Host defensemechanisms triggered by microbial lipoproteins through Toll-like receptors.Science 285:732–736.
12. Brown, C. R., and S. L. Reiner. 1999. Experimental Lyme arthritis in theabsence of interleukin-4 or gamma interferon. Infect. Immun. 67:3329–3333.
13. Cameron, C. E. 2006. The T. pallidum outer membrane and outer membraneproteins, p. 237–266. In J. D. Radolf and S. A. Lukehart (ed.), Pathogenictreponemes: molecular and cellular biology. Caister Academic Press, Nor-wich, United Kingdom.
14. Casjens, S., N. Palmer, R. van Vugt, W. M. Huang, B. Stevenson, P. Rosa, R.Lathigra, G. Sutton, J. Peterson, R. J. Dodson, D. Haft, E. Hickey, M.Gwinn, O. White, and C. M. Fraser. 2000. A bacterial genome in flux: thetwelve linear and nine circular extrachromosomal DNAs in an infectiousisolate of the Lyme disease spirochete Borrelia burgdorferi. Mol. Microbiol.35:490–516.
15. Chiani, P., C. Bromuro, and A. Torosantucci. 2000. Defective induction ofinterleukin-12 in human monocytes by germ-tube forms of Candida albicans.Infect. Immun. 68:5628–5634.
16. Cogswell, J. P., M. M. Godlevski, G. B. Wisely, W. C. Clay, L. M. Leesnitzer,J. P. Ways, and J. G. Gray. 1994. NF-�B regulates IL-1 transcriptionthrough a consensus NF-�B binding site and a nonconsensus CRE-like site.J. Immunol. 153:712–723.
17. Colonna, M., G. Trinchieri, and Y. J. Liu. 2004. Plasmacytoid dendritic cellsin immunity. Nat. Immunol. 5:1219–1226.
18. Creagh, E. M., and L. A. O’Neill. 2006. TLRs, NLRs and RLRs: a trinity ofpathogen sensors that co-operate in innate immunity. Trends Immunol.27:352–357.
19. Dinarello, C. A. 1998. Interleukin-1 , interleukin-18, and the interleukin-1 converting enzyme. Ann. N. Y. Acad. Sci. 856:1–11.
20. Eggers, C. H., M. J. Caimano, M. L. Clawson, W. G. Miller, D. S. Samuels,and J. D. Radolf. 2002. Identification of loci critical for replication andcompatibility of a Borrelia burgdorferi cp32 plasmid and use of a cp32-basedshuttle vector for the expression of fluorescent reporters in the Lyme diseasespirochaete. Mol. Microbiol. 43:281–295.
21. Eggers, C. H., M. J. Caimano, and J. D. Radolf. 2006. Sigma factor selectivityin Borrelia burgdorferi: RpoS recognition of the ospE/ospF/elp promoters isdependent on the sequence of the 10 region. Mol. Microbiol. 59:1859–1875.
22. Fanger, N. A., D. Voigtlaender, C. Liu, S. Swink, K. Wardwell, J. Fisher,R. F. Graziano, L. C. Pfefferkorn, and P. M. Guyre. 1997. Characterizationof expression, cytokine regulation, and effector function of the high affinityIgG receptor Fc gamma RI (CD64) expressed on human blood dendriticcells. J. Immunol. 158:3090–3098.
23. Fonteneau, J. F., M. Larsson, A. S. Beignon, K. McKenna, I. Dasilva, A.Amara, Y. J. Liu, J. D. Lifson, D. R. Littman, and N. Bhardwaj. 2004.Human immunodeficiency virus type 1 activates plasmacytoid dendritic cellsand concomitantly induces the bystander maturation of myeloid dendriticcells. J. Virol. 78:5223–5232.
24. Franchi, L., A. Amer, M. Body-Malapel, T. D. Kanneganti, N. Ozoren, R.Jagirdar, N. Inohara, P. Vandenabeele, J. Bertin, A. Coyle, E. P. Grant, andG. Nunez. 2006. Cytosolic flagellin requires Ipaf for activation of caspase-1and interleukin 1 in Salmonella-infected macrophages. Nat. Immunol.7:576–582.
25. Fraser, C. M., S. Casjens, W. M. Huang, G. G. Sutton, R. Clayton, R.Lathigra, O. White, K. A. Ketchum, R. Dodson, E. K. Hickey, M. Gwinn, B.Dougherty, J. F. Tomb, R. D. Fleischmann, D. Richardson, J. Peterson, A.R. Kerlavage, J. Quackenbush, S. Salzberg, M. Hanson, R. van Vugt,N. Palmer, M. D. Adams, J. Gocayne, J. Weidman, T. Utterback, L. Watthey,L. McDonald, P. Artiach, C. Bowman, S. Garland, C. Fujii, M. D. Cotton, K.Horst, K. Roberts, B. Hatch, H. O. Smith, and J. C. Venter. 1997. Genomic
2060 MOORE ET AL. INFECT. IMMUN.

sequence of a Lyme disease spirochaete, Borrelia burgdorferi. Nature 390:580–586.
26. Fraser, C. M., S. J. Norris, G. M. Weinstock, O. White, G. G. Sutton, R.Dodson, M. Gwinn, E. K. Hickey, R. Clayton, K. A. Ketchum, E. Sodergren,J. M. Hardham, M. P. McLeod, S. Salzberg, J. Peterson, H. Khalak, D.Richardson, J. K. Howell, M. Chidambaram, T. Utterback, L. McDonald, P.Artiach, C. Bowman, M. D. Cotton, and J. C. Venter. 1998. Complete ge-nome sequence of Treponema pallidum, the syphilis spirochete. Science 281:375–388.
27. Fujii, S., K. Shimizu, M. Kronenberg, and R. M. Steinman. 2002. ProlongedIFN-�-producing NKT response induced with alpha-galactosylceramide-loaded DCs. Nat. Immunol. 3:867–874.
28. Gavrilin, M. A., I. J. Bouakl, N. L. Knatz, M. D. Duncan, M. W. Hall, J. S.Gunn, and M. D. Wewers. 2006. Internalization and phagosome escaperequired for Francisella to induce human monocyte IL-1� processing andrelease. Proc. Natl. Acad. Sci. USA 103:141–146.
29. Gerosa, F., B. Baldani-Guerra, C. Nisii, V. Marchesini, G. Carra, and G.Trinchieri. 2002. Reciprocal activating interaction between natural killercells and dendritic cells. J. Exp. Med. 195:327–333.
30. Glickstein, L., M. Edelstein, and J. Z. Dong. 2001. Gamma interferon is notrequired for arthritis resistance in the murine Lyme disease model. Infect.Immun. 69:3737–3743.
31. Grazia, C. M., F. S. Sutterwala, G. Trinchieri, D. M. Mosser, and X. Ma.2001. Suppression of IL-12 transcription in macrophages following Fc� re-ceptor ligation. J. Immunol. 166:4498–4506.
32. Hazlett, K. R. O., T. J. Sellati, T. T. Nguyen, D. L. Cox, M. L. Clawson, M. J.Caimano, and J. D. Radolf. 2001. The T. pallidumrK protein of Treponemapallidum is periplasmic and is not a target of opsonic antibody or protectiveimmunity. J. Exp. Med. 193:1015–1026.
33. Hengge, U. R., A. Tannapfel, S. K. Tyring, R. Erbel, G. Arendt, and T.Ruzicka. 2003. Lyme borreliosis. Lancet Infect. Dis. 3:489–500.
34. Hilbi, H., J. E. Moss, D. Hersh, Y. Chen, J. Arondel, S. Banerjee, R. A.Flavell, J. Yuan, P. J. Sansonetti, and A. Zychlinsky. 1998. Shigella-inducedapoptosis is dependent on caspase-1 which binds to IpaB. J. Biol. Chem.273:32895–32900.
35. Hirschfeld, M., C. J. Kirschning, R. Schwandner, H. Wesche, J. H. Weis,R. M. Wooten, and J. J. Weis. 1999. Inflammatory signaling by Borreliaburgdorferi lipoproteins is mediated by Toll-like receptor 2. J. Immunol.163:2382–2386.
36. Hirschfeld, M., Y. Ma, J. H. Weis, S. N. Vogel, and J. J. Weis. 2000. Cuttingedge: repurification of lipopolysaccharide eliminates signaling through bothhuman and murine toll-like receptor 2. J. Immunol. 165:618–622.
37. Imlach, S., C. Leen, J. E. Bell, and P. Simmonds. 2003. Phenotypic analysisof peripheral blood �� T lymphocytes and their targeting by human immu-nodeficiency virus type 1 in vivo. Virology 305:415–427.
38. Kadowaki, N., S. Ho, S. Antonenko, M. R. de Waal, R. A. Kastelein, F.Bazan, and Y. J. Liu. 2001. Subsets of human dendritic cell precursorsexpress different Toll-like receptors and respond to different microbial an-tigens. J. Exp. Med. 194:863–870.
39. Kang, I., S. W. Barthold, D. H. Persing, and L. K. Bockenstedt. 1997.T-helper-cell cytokines in the early evolution of murine Lyme arthritis.Infect. Immun. 65:3107–3111.
40. Kronenberg, M., and L. Gapin. 2002. The unconventional lifestyle of NKTcells. Nat. Rev. Immunol. 2:557–568.
41. Latz, E., A. Schoenemeyer, A. Visintin, K. A. Fitzgerald, B. G. Monks, C. F.Knetter, E. Lien, N. J. Nilsen, T. Espevik, and D. T. Golenbock. 2004. TLR9signals after translocating from the ER to CpG DNA in the lysosome. Nat.Immunol. 5:190–198.
42. Liu, Y., E. Masuda, M. C. Blank, K. A. Kirou, X. Gao, M. S. Park, and L.Pricop. 2005. Cytokine-mediated regulation of activating and inhibitory Fc�receptors in human monocytes. J. Leukoc. Biol. 77:767–776.
43. Lukehart, S. A. 1992. Immunology and pathogenesis of syphilis, p. 141–163.In T. C. Quinn (ed.), Advances in host defense mechanisms, vol. 8. Sexuallytransmitted diseases. Raven Press, New York, NY.
44. Lukehart, S. A. 2004. Syphilis, p. 977–985. In E. Brauwald, A. S. Faucci, S. L.Hauser, D. L. Longo, and J. L. Jameson (ed.), Harrison’s principles ofinternal medicine. McGraw Hill, New York, NY.
45. Lukehart, S. A., and J. N. Miller. 1978. Demonstration of the in vitrophagocytosis of Treponema pallidum by rabbit peritoneal macrophages.J. Immunol. 121:2014–2024.
46. Ma, X., J. M. Chow, G. Gri, G. Carra, F. Gerosa, S. F. Wolf, R. Dzialo, andG. Trinchieri. 1996. The interleukin 12 p40 gene promoter is primed byinterferon gamma in monocytic cells. J. Exp. Med. 183:147–157.
47. Ma, Y., and J. J. Weis. 1993. Borrelia burgdorferi outer surface lipoproteinsOspA and OspB possess B-cell mitogenic and cytokine-stimulatory proper-ties. Infect. Immun. 61:3843–3853.
48. Marshall, J. D., D. S. Heeke, C. Abbate, P. Yee, and N. G. Van. 2006.Induction of interferon-� from natural killer cells by immunostimulatoryCpG DNA is mediated through plasmacytoid-dendritic-cell-produced inter-feron-� and tumour necrosis factor-�. Immunology 117:38–46.
49. Martin-Fontecha, A., L. L. Thomsen, S. Brett, C. Gerard, M. Lipp, A.
Lanzavecchia, and F. Sallusto. 2004. Induced recruitment of NK cells tolymph nodes provides IFN-� for T(H)1 priming. Nat. Immunol. 5:1260–1265.
50. Martinon, F., K. Burns, and J. Tschopp. 2002. The inflammasome: a mo-lecular platform triggering activation of inflammatory caspases and process-ing of proIL-1 . Mol. Cell 10:417–426.
51. Matsuzaki, G., H. Yamada, K. Kishihara, Y. Yoshikai, and K. Nomoto. 2002.Mechanism of murine V�1� �� T cell-mediated innate immune responseagainst Listeria monocytogenes infection. Eur. J. Immunol. 32:928–935.
52. Matyniak, J. E., and S. L. Reiner. 1995. T helper phenotype and geneticsusceptibility in experimental Lyme disease. J. Exp. Med. 181:1251–1254.
53. Molofsky, A. B., B. G. Byrne, N. N. Whitfield, C. A. Madigan, E. T. Fuse, K.Tateda, and M. S. Swanson. 2006. Cytosolic recognition of flagellin by mousemacrophages restricts Legionella pneumophila infection. J. Exp. Med. 203:1093–1104.
54. Montgomery, R. R., D. Lusitani, C. A. de Boisfleury, and S. E. Malawista.2002. Human phagocytic cells in the early innate immune response to Bor-relia burgdorferi. J. Infect. Dis. 185:1773–1779.
55. Montgomery, R. R., and S. E. Malawista. 1996. Entry of Borrelia burgdorferiinto macrophages is end-on and leads to degradation in lysosomes. Infect.Immun. 64:2867–2872.
56. Moore, K. J., L. P. Andersson, R. R. Ingalls, B. G. Monks, R. Li, M. A.Arnaout, D. T. Golenbock, and M. W. Freeman. 2000. Divergent response toLPS and bacteria in CD14-deficient murine macrophages. J. Immunol. 165:4272–4280.
57. Morita, C. T., R. A. Mariuzza, and M. B. Brenner. 2000. Antigen recognitionby human gamma delta T cells: pattern recognition by the adaptive immunesystem. Springer Semin. Immunopathol. 22:191–217.
58. Morita, C. T., Y. Tanaka, B. R. Bloom, and M. B. Brenner. 1996. Directpresentation of non-peptide prenyl pyrophosphate antigens to humangamma delta T cells. Res. Immunol. 147:347–353.
59. Napolitani, G., A. Rinaldi, F. Bertoni, F. Sallusto, and A. Lanzavecchia.2005. Selected Toll-like receptor agonist combinations synergistically triggera T helper type 1-polarizing program in dendritic cells. Nat. Immunol.6:769–776.
60. Nimmerjahn, F., and J. V. Ravetch. 2006. Fc� receptors: old friends and newfamily members. Immunity 24:19–28.
61. Ozinsky, A., D. M. Underhill, J. D. Fontenot, A. M. Hajjar, K. D. Smith,C. B. Wilson, L. Schroeder, and A. Aderem. 2000. The repertoire for patternrecognition of pathogens by the innate immune system is defined by coop-eration between toll-like receptors. Proc. Natl. Acad. Sci. USA 97:13766–13771.
62. Ozoren, N., J. Masumoto, L. Franchi, T. D. Kanneganti, M. Body-Malapel,I. Erturk, R. Jagirdar, L. Zhu, N. Inohara, J. Bertin, A. Coyle, E. P. Grant,and G. Nunez. 2006. Distinct roles of TLR2 and the adaptor ASC in IL-1beta/IL-18 secretion in response to Listeria monocytogenes. J. Immunol.176:4337–4342.
63. Radolf, J. D., K. R. O. Hazlett, and S. A. Lukehart. 2006. Pathogenesis ofsyphilis, p. 197–236. In J. D. Radolf and S. A. Lukehart (ed.), Pathogenictreponemes: cellular and molecular biology. Caister Academic Press, Nor-folk, United Kingdom.
64. Radolf, J. D. 1995. Treponema pallidum and the quest for outer membraneproteins. Mol. Microbiol. 16:1067–1073.
65. Romagnani, C., C. M. Della, S. Kohler, B. Moewes, A. Radbruch, L. Moretta,A. Moretta, and A. Thiel. 2005. Activation of human NK cells by plasmacy-toid dendritic cells and its modulation by CD4(�) T helper cells andCD4(�) CD25(hi) T regulatory cells. Eur. J. Immunol. 35:2452–2458.
66. Rosenshine, I., S. Ruschkowski, and B. B. Finlay. 1994. Inhibitors of cy-toskeletal function and signal transduction to study bacterial invasion. Meth-ods Enzymol. 236:467–476.
67. Salazar, J. C., C. D. Pope, M. W. Moore, J. Pope, T. G. Kiely, and J. D.Radolf. 2005. Lipoprotein-dependent and -independent immune responsesto spirochetal infection. Clin. Diagn. Lab Immunol. 12:949–958.
68. Salazar, J. C., C. D. Pope, T. J. Sellati, H. M. Feder, Jr., T. G. Kiely, K. R.Dardick, R. L. Buckman, M. W. Moore, M. J. Caimano, J. G. Pope, P. J.Krause, and J. D. Radolf. 2003. Coevolution of markers of innate andadaptive immunity in skin and peripheral blood of patients with erythemamigrans. J. Immunol. 171:2660–2670.
68a.Salazar, J. C., A. R. Cruz, C. D. Pope, L. Valderrama, R. Trujillo, N. G.Saravia, and J. D. Radolf. 2007. Treponema pallidum elicits innate andadaptive cellular immune responses in skin and blood during secondarysyphilis: a flow-cytometric analysis. J. Infect. Dis. 195:879–887.
69. Sellati, T. J., D. A. Bouis, M. J. Caimano, J. A. Feulner, C. Ayers, E. Lien,and J. D. Radolf. 1999. Activation of human monocytic cells by Borreliaburgdorferi and Treponema pallidum is facilitated by CD14 and correlateswith surface exposure of spirochetal lipoproteins. J. Immunol. 163:2049–2056.
70. Sellati, T. J., D. A. Bouis, R. L. Kitchens, R. P. Darveau, J. Pugin, R. J.Ulevitch, S. C. Gangloff, S. M. Goyert, M. V. Norgard, and J. D. Radolf. 1998.Treponema pallidum and Borrelia burgdorferi lipoproteins and synthetic li-popeptides activate monocytic cells via a CD14-dependent pathway distinctfrom that used by lipopolysaccharide. J. Immunol. 160:5455–5464.
71. Sellati, T. J., S. L. Waldrop, J. C. Salazar, P. R. Bergstresser, L. J. Picker,
VOL. 75, 2007 ENHANCED RECOGNITION OF SPIROCHETES BY PHAGOCYTOSIS 2061

and J. D. Radolf. 2001. The cutaneous response in humans to Treponemapallidum lipoprotein analogues involves cellular elements of both innate andadaptive immunity. J. Immunol. 166:4131–4140.
72. Sieling, P. A., W. Chung, B. T. Duong, P. J. Godowski, and R. L. Modlin.2003. Toll-like receptor 2 ligands as adjuvants for human Th1 responses.J. Immunol. 170:194–200.
73. Siren, J., T. Sareneva, J. Pirhonen, M. Strengell, V. Veckman, I. Julkunen,and S. Matikainen. 2004. Cytokine and contact-dependent activation ofnatural killer cells by influenza A or Sendai virus-infected macrophages.J. Gen. Virol. 85:2357–2364.
74. Snijders, A., P. Kalinski, C. M. Hilkens, and M. L. Kapsenberg. 1998.High-level IL-12 production by human dendritic cells requires two signals.Int. Immunol. 10:1593–1598.
75. Stevens, T. L., A. Bossie, V. M. Sanders, R. Fernandez-Botran, R. L. Coffman,T. R. Mosmann, and E. S. Vitetta. 1988. Regulation of antibody isotype secre-tion by subsets of antigen-specific helper T cells. Nature 334:255–258.
76. Stuart, L. M., and R. A. Ezekowitz. 2005. Phagocytosis: elegant complexity.Immunity 22:539–550.
77. Underhill, D. M., A. Ozinsky, A. M. Hajjar, A. Stevens, C. B. Wilson, M.Bassetti, and A. Aderem. 1999. The Toll-like receptor 2 is recruited to
macrophage phagosomes and discriminates between pathogens. Nature 401:811–815.
78. Van Voorhis, W. C., L. K. Barrett, D. M. Koelle, J. M. Nasio, F. A. Plummer,and S. A. Lukehart. 1996. Primary and secondary syphilis lesions containmRNA for Th1 cytokines. J. Infect. Dis. 173:491–495.
79. Viala, J., C. Chaput, I. G. Boneca, A. Cardona, S. E. Girardin, A. P. Moran,R. Athman, S. Memet, M. R. Huerre, A. J. Coyle, P. S. DiStefano, P. J.Sansonetti, A. Labigne, J. Bertin, D. J. Philpott, and R. L. Ferrero. 2004.Nod1 responds to peptidoglycan delivered by the Helicobacter pylori cagpathogenicity island. Nat. Immunol. 5:1166–1174.
80. Wooten, R. M., T. B. Morrison, J. H. Weis, S. D. Wright, R. Thieringer, andJ. J. Weis. 1998. The role of CD14 in signaling mediated by outer membranelipoproteins of Borrelia burgdorferi. J. Immunol. 160:5485–5492.
81. Wooten, R. M., and J. J. Weis. 2001. Host-pathogen interactions promotinginflammatory Lyme arthritis: use of mouse models for dissection of diseaseprocesses. Curr. Opin. Microbiol. 4:274–279.
82. Yamamoto, M., K. Yaginuma, H. Tsutsui, J. Sagara, X. Guan, E. Seki, K.Yasuda, M. Yamamoto, S. Akira, K. Nakanishi, T. Noda, and S. Taniguchi.2004. ASC is essential for LPS-induced activation of procaspase-1 indepen-dently of TLR-associated signal adaptor molecules. Genes Cells 9:1055–1067.
Editor: D. L. Burns
2062 MOORE ET AL. INFECT. IMMUN.




















