
Porphyromonas gingivalis and Treponema denticola exhibit metabolic symbioses
Upload
independentCategory
view
0download
0
JOURNAL OF BACTERIOLOGY, July 2010, p. 3337–3344 Vol. 192, No. 130021-9193/10/$12.00 doi:10.1128/JB.00274-10Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Treponema denticola PrcB Is Required for Expression andActivity of the PrcA-PrtP (Dentilisin) Complex�
Valentina Godovikova, Hong-Tao Wang,‡ M. Paula Goetting-Minesky, Yu Ning, Ricardo F. Capone,†Claudia K. Slater, and J. Christopher Fenno*
Department of Biologic and Materials Sciences, School of Dentistry, University of Michigan, Ann Arbor, Michigan
Received 11 March 2010/Accepted 19 April 2010
The Treponema denticola surface protease complex, consisting of PrtP protease (dentilisin) and two auxiliarypolypeptides (PrcA1 and PrcA2), is believed to contribute to periodontal disease by degrading extracellularmatrix components and disrupting host intercellular signaling. Previously, we showed that transcription of theprotease operon initiates upstream of TDE0760 (herein designated prcB), the open reading frame immediately5� of prcA-prtP. The prcB gene is conserved in T. denticola strains. PrcB localizes to the detergent phase of TritonX-114 cell surface extracts and migrates as a 22-kDa polypeptide, in contrast to the predicted 17-kDacytoplasmic protein encoded in the annotated T. denticola genome. Consistent with this observation, the PrcBN terminus is unavailable for Edman sequencing, suggesting that it is acylated. Nonpolar deletion of prcB inT. denticola showed that PrcB is required for production of PrtP protease activity, including native PrtPcleavage of PrcA to PrcA1 and PrcA2. A 6�His-tagged PrcB protein coimmunoprecipitates with native PrtP,using either anti-PrtP or anti-His-tag antibodies, and recombinant PrtP copurifies with PrcB-6�His in nickelaffinity chromatography. Taken together, these data are consistent with identification of PrcB as a PrtP-binding lipoprotein that likely stabilizes the PrtP polypeptide during localization to the outer membrane.
Treponema denticola is an oral spirochete strongly associatedwith periodontal diseases. The predominance of spirochetes insubgingival plaque in severe periodontal diseases and theirspatial location within periodontal lesions are highly suggestiveof an important role in periodontal pathogenesis (13). SeveralT. denticola secreted proteinases and peptidases have beenidentified that likely contribute to periodontal pathogenesis asa consequence of their roles in processing host tissue proteinsand peptides to fulfill the nutritional requirements of thesehighly motile and invasive organisms (reviewed in references16 and 32). Among these, the outer membrane serine proteasecomplex (variously designated CTLP [40], dentilisin [26], andthe PrtP complex [5]) has several activities consistent with itsidentification as a virulence determinant in periodontal disease.The PrtP protease complex degrades extracellular matrix proteins(3, 23) as well as serum components involved in tissue homeosta-sis (22, 40), regulation of the complement pathway (33, 41), andother innate immune responses (1, 12, 38). PrtP protease activityalso contributes to T. denticola coaggregation with Porphyromo-nas gingivalis (25) and penetration of host tissue (8, 23).
The prcA-prtP locus is conserved in oral Treponema species,and the encoded products can be divided into two paralogousfamilies based on phylogenetic analysis and substrate specific-ity of the protease (9). PrtP is one of only two known acylatedmembers of the subtilisin family, the other being SphB1, asurface-anchored subtilisin autotransporter that catalyzes mat-
uration of the virulence factor FhaB (filamentous hemaggluti-nin) at the surface of Bordetella pertussis (10). PrcA has nohomologues outside the oral Treponema spp. Other than itslikely contribution to anchoring the protease complex in theouter membrane, the exact function of PrcA is not known. Thenative protease consists of a complex comprised of PrtP andthe PrcA1 and PrcA2 polypeptides that, if unheated, does notdissociate into its constituents in SDS-PAGE analyses (26, 29,40), suggesting that protein-protein interactions stabilize theprotease complex in the T. denticola outer membrane. Ourprior analysis of the T. denticola protease operon demon-strated that transcription of prtP initiates �580 bp upstream ofprcA and includes all of the TDE0760 open reading frame(ORF), immediately 5� of prcA (5). While our group and otherscontinue to characterize interactions between PrtP and PrcA, wechose to further investigate the role of this conserved, geneticallylinked open reading frame encoding a hypothetical protein inexpression and activity of the protease complex. Here we presentinitial characterization of the TDE0760 (herein designated prcB)gene product and provide evidence supporting a role of PrcB inexpression of the PrtP protease complex.
MATERIALS AND METHODS
Bacterial strains and growth conditions. T. denticola ATCC 35405, ATCC33520, and OTK and isogenic mutants of 35405 (Table 1) were grown in NOSbroth or NOS/GN semisolid medium as previously described (7, 24), with eryth-romycin (Em) (40 �g ml�1) added as appropriate. Cultures were examined byphase-contrast microscopy for purity and typical strain morphology before use.
Escherichia coli NovaBlue (Novagen, Inc., Madison, WI) and JM109 (42) wereused as hosts for cloning. E. coli was grown in LB agar or broth medium withampicillin (50 �g ml�1), kanamycin (30 �g ml�1), and Em (200 �g ml�1), asappropriate. Plasmid vector pSTBlue-1 (Novagen) was used for direct cloning ofPCR products, and 6�His-tagged constructs were made in pET28b (Novagen).
Construction of plasmids for expression and mutagenesis studies. A 1,070-bpfragment of T. denticola genomic DNA containing the entire prcB ORF andapproximately 500 bp 5� of prcB was amplified by a PCR using oligonucleotide
* Corresponding author. Mailing address: Department of Biologicand Materials Sciences, School of Dentistry, University of Michigan,Ann Arbor, MI 48109-1078. Phone: (734) 763-3331. Fax: (734) 647-2110. E-mail: [email protected].
† Present address: Jacobs School of Engineering, University of Cal-ifornia—San Diego, San Diego, CA.
‡ Present address: Shenzhen Microbiolab Ltd., Shenzhen, China.� Published ahead of print on 30 April 2010.
3337

primers CX516 and CX529. (Details of all plasmids and oligonucleotide primersused in this study are contained in Tables 2 and 3.) The PCR product, digestedwith NheI and XhoI, was ligated to similarly digested pET28b, yielding pCF414,in which prcB contains the coding sequence of an in-frame C-terminal 6�Hismotif derived from the vector. To move this construct into a standard cloningvector, the PrcB-6�His region of pCF414 was PCR amplified using oligonucle-otide primers CX516 and CX518 and cloned into pSTBlue-1, yielding pCF415.
In a previous study (29), we constructed pSY119, containing the 3� end of prcAand the entire prtP gene. The 2.1-kb ermF-ermAM cassette (erm) (21) carried onpSY118 (28) was released by EcoRI digestion and ligated to EcoRI-digestedpSY119 (29). In the resulting plasmid (pCF411), erm is inserted at the 5� end ofa DNA consisting of the 3� end of prcA and the entire prtP gene. Using oligo-nucleotide primers CX535 and CX536, the entire prcB-6�His fragment wasamplified from pCF414, digested with SacI and NheI, and ligated to similarlydigested pCF411, yielding pCF416. This plasmid, used to construct T. denticolamutant strain CF417, contains the native promoter region, prcB-6�His, erm, the3� end of prcA, and full-length prtP.
Using a similar strategy, we constructed a plasmid in which erm was insertedinto the intergenic region between TDE0758 and TDE0759, upstream of 6�His-tagged prcB. In this plasmid, the prcA 5� end is intact. This plasmid was used toconstruct T. denticola mutant strain CF499. Briefly, TDE0758, amplified usingprimers CX558 and CX578, was cloned into pSTBlue-1, yielding pCF472. DNAencoding TDE0759 and 6�His-tagged prcB was amplified from T. denticolaCF417 by use of primers CX562 and CX577 and was cloned into pSTBlue-1,yielding pCF471. The 0.9-kb plasmid insert, released by digestion with SnaBI andNdeI, was ligated to HincII/NdeI-digested pCF472, yielding pCF476. The ermfragment of pSY118 was released with NdeI and cloned into the NdeI site ofpCF476, between TDE0758 and TDE0759, yielding pCF480. DNA encoding the5� region of prcA was amplified from T. denticola 35405 by use of primers CX581and CX582 and was cloned into pSTBlue-1, yielding pCF478. The 1.9-kb prcAfragment released by digestion with XbaI/BstZ17I was cloned into pCF480 whichhad been digested with BglII (blunt ended with Klenow polymerase) and XbaI,yielding pCF483. This plasmid was used as the source of linear DNA to constructT. denticola mutant strain CF499.
For nonpolar deletion of prcB, a DNA fragment consisting of the 5� region ofprcA amplified from T. denticola 35405 by use of primers CX630 and CX582 wascloned downstream of the predicted protease operon promoter carried on
pCF480, yielding pCF521. This plasmid was used as the source of linear DNA toconstruct T. denticola CF522 by allelic replacement mutagenesis.
To compare recombinant expression of PrcB initiating at each of the threepossible start codons, DNA fragments including the coding region beginning atMet1, Met7, and Val38 were amplified using forward primers CX710, CX711, andCX712, respectively, and the reverse primer CX713. Each amplicon, digestedwith NdeI and HindIII, was cloned into pET30b such that the resulting plasmids(pCF569, pCF570, and pCF571) expressed PrcB-6�His from a different startcodon under the inducible control of T7 RNA polymerase.
Allelic replacement mutagenesis. Defined isogenic mutants were constructedas described previously (19, 31), by electroporation of T. denticola with linearizedplasmid DNA fragments consisting of the selectable erm cassette cloned betweenDNA fragments flanking the target gene. Mutants were selected for resistance toEm (Emr) in NOS/GN agar (7). Mutations were verified by PCR analysis and byDNA sequencing of the target region in genomic DNA.
DNA sequence analysis. Templates for DNA sequencing, including plasmidDNAs and PCR products, were sequenced at the University of Michigan DNASequencing Core Facility. Sequences of both DNA strands of regions of interestwere obtained. DNA sequences of the prcB ORF in T. denticola strains ATCC33520 and OTK were amplified from genomic DNA with primers CX402 andCX360. Annotation of the prcB open reading frame was done using theFGENESB algorithm trained to the T. denticola genome sequence (Softberry,Inc., Mt. Kisco, NY). A predicted �70 class promoter upstream of prcB wasidentified using BPROM software (Softberry, Inc., Mt. Kisco, NY). Identificationof potential signal peptidase cleavage sites was done using PSORT (14, 34),LipoP (27), and SpLip (37).
Transcription analysis. Total RNA was isolated from T. denticola cells har-vested during logarithmic growth (3-day cultures) with Trizol reagent (Invitro-gen, Carlsbad, CA), using the manufacturer’s protocol. DNase-treated RNAsamples were reversed transcribed with random hexamer primers, using theSuperScript first-strand synthesis system for reverse transcriptase PCR (RT-PCR) (Invitrogen). One microliter of the resulting first-strand cDNA was am-plified for 30 cycles by using Taq DNA polymerase (Invitrogen) and oligonucle-otide primer sets specific for the target gene. RT-PCR products, includingpositive controls (genomic DNA) and negative controls (no RT enzyme), wereanalyzed by agarose gel electrophoresis.
Protein electrophoresis and immunoblotting. Sodium dodecyl sulfate-poly-acrylamide gel electrophoresis (SDS-PAGE) and Western immunoblotting weredone as described previously (17). SDS-PAGE gels were stained with Coomassiebrilliant blue or silver nitrate. Cells were harvested by centrifugation at 10,000 �g (10 min, 4°C). Whole-cell lysates of T. denticola and E. coli were prepared bysuspending cells (1-ml cultures at an optical density at 600 nm [OD600] of 0.2) instandard SDS-PAGE sample buffer containing �-mercaptoethanol and 2 mMphenylmethylsulfonyl fluoride (PMSF) or by sonication of phosphate-bufferedsaline (PBS)-washed cells prior to suspension in sample buffer. For some exper-iments, T. denticola cells were treated with Triton X-114 to extract outer mem-brane and periplasmic components as described for Treponema pallidum (11),with slight modifications. Treponema cells were harvested by centrifugation at10,000 � g (10 min at 4°C), washed in PBS, suspended in a 1/40 volume of 20 mMTris-HCl (pH 7.5)–2 mM EDTA–1 mM dithiothreitol containing 1% TritonX-114, and stirred gently overnight at 4°C. Detergent-extracted cells were cen-trifuged at 17,000 � g for 10 min at 4°C. The supernatant, enriched for outer
TABLE 1. T. denticola strains used in this study
T. denticolastrain Relevant features Source or
referencea
35405 Parent for mutagenesis ATCC33520 ATCCOTK 20P0760 prcB, polar mutation 5CF417 prcB-6�His �prcA This studyCF499 prcB-6�His This studyCF522 �prcB, nonpolar mutation This study
a ATCC, American Type Culture Collection, Rockville, MD.
TABLE 2. Plasmids used for expression and mutagenesisa
E. coli plasmid Characteristics; use Reference
pSY118 ermF-ermAM cassette 28pSY119 3� end of prcA-prtP 29pCF411 ermF-ermAM inserted 5� of truncated prcA in pSY119; PrtP expression in E. coli This studypCF415 prcB-6�His in pSTBlue-1; PrcB-6�His expression in E. coli This studypCF416 prcB-6�His in pCF411; PrcB-6�His and PrtP expression in E. coli and
construction of T. denticola CF417This study
pCF483 ermF-ermAM between TDE0758 and TDE0759; prcB-6 � His in “native”protease locus; construction of T. denticola CF499
This study
pCF521 ermF-ermAM inserted between TDE0758 and TDE0759; �prcB; construction ofT. denticola CF522
This study
pCF569 prcB-6�His beginning at Met1, expressed in pET30b This studypCF570 prcB-6�His beginning at Met7, expressed in pET30b This studypCF571 prcB-6�His beginning at Val38, expressed in pET30b This study
a Construction of the plasmids, including intermediate steps, is described in Materials and Methods.
3338 GODOVIKOVA ET AL. J. BACTERIOL.

membrane components, was partitioned into aqueous and detergent phases at37°C, and each phase was reextracted in Triton X-114 and partitioned a secondtime. The extracted material was then precipitated in acetone and resuspendedin equal volumes to equalize sample loading. Prior to electrophoresis, sampleswere either heated at 100°C for 5 min or held on ice.
Proteins blotted onto nitrocellulose membranes were detected with rabbitpolyclonal antibodies raised against recombinant T. denticola proteins, followedby horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG (Thermo Sci-entific, Rockford, IL). 6�His-tagged proteins were detected with HisProbe HRPreagent (Thermo Scientific) or with mouse monoclonal anti-6�His antibodies(EMD Biosciences, San Diego, CA). Protein bands of interest were visualizedusing SuperSignal West Pico chemiluminescence substrate (Thermo Scientific).
Protein purification and sequencing. T. denticola and E. coli cells expressingPrcB-6�His were lysed by treatment with BugBuster (EMD Biosciences), and ex-tracts were further disrupted by sonication. PrcB-6�His was purified from cellextracts by nickel affinity chromatography (2). Eluted fractions containing PrcB-6�His were concentrated by acetone precipitation, subjected to SDS-PAGE, trans-ferred to nylon membranes, and stained with amido black or with HisProbe tovisualize eluted proteins. Bands of interest were submitted to the Protein StructureFacility at the University of Michigan for N-terminal amino acid sequencing.
Protease activity assays. Gelatin zymography to detect the active PrtP pro-tease was performed as described previously (15). Whole-cell extracts for zymog-raphy were prepared similarly to those for SDS-PAGE, except without heating,PMSF, or reducing agent. Following electrophoresis, the gel was stained withCoomassie brilliant blue to detect active protease. PrtP-dependent hydrolysis ofthe chromogenic substrate succinyl-L-alanyl-L-alanyl-L-prolyl-L-phenylalanine-p-nitroanilide (SAAPFNA) was assayed by the change in absorbance at 405 nm, asdescribed previously (30, 40).
Immunoprecipitation assay. Three-day-old 10-ml cultures of T. denticola(OD600 of 0.5) were harvested by centrifugation at 5,000 � g for 5 min, washedwith cold PBS, and incubated for 30 min at 4°C on a roller in 1 ml of Tris-buffered saline (TBS) (pH 7.4) containing 0.5% Triton X-100 and 10 �l proteaseinhibitor cocktail (Roche, Indianapolis, IN). The suspension was centrifuged at13,000 � g for 10 min at 4°C to remove unlysed cells. One-hundred-microliteraliquots of the suspension were diluted 1:4 in TBS, and 5 �l of rabbit polyclonalantiserum raised against PrtP, PrcA, FlaA, or Msp (5, 17, 29) was added to eachtube. Following overnight incubation at 4°C on a roller to form the antigen-antibody complexes, 50 �l of protein A-agarose slurry (Thermo Scientific) wasadded to each tube and incubated with gentle mixing for 2 h at room tempera-ture. The mixture was then centrifuged for 2 min at 2,500 � g and washed threetimes in TBS. The immune complex was eluted with 100 �l of 0.2 M glycine-HCl,pH 2.5. Eluted samples were neutralized with 10 �l of 1 M Tris (pH 8) andprocessed for SDS-PAGE and immunoblotting.
Nucleotide sequence accession numbers. The prcB sequences of T. denticolastrains ATCC 33520 and OTK have been assigned GenBank accession numbersFJ555200 and FJ555201, respectively.
RESULTS
Conservation of prcB. Prolyl-phenylalanyl protease activity isfound in diverse T. denticola strains (35). T. denticola strains35405, 33520, and OTK are antigenically distinct, consistentwith their assignment to distinct serotypes (20), and all exhibitprolyl-phenylalanyl protease activity (not shown). To deter-mine the level of intraspecies prcB conservation, the entireTDE0760 ORF and flanking DNAs were amplified from thegenomic DNAs of T. denticola strains 33520 and OTK, and theDNA sequences were compared with that of 35405. The threeT. denticola prcB DNA sequences were �97% identical overthe entire ORF, including 210 bp upstream of the first AUGcodon in the TDE060 ORF, resulting in a total of 7 amino aciddifferences between the three deduced polypeptides (Fig. 1and data not shown). Counting from the first AUG codon inthe ORF, the deduced amino acid sequences are identical forresidues 1 to 65, which includes the predicted untranslatedregion upstream of the GUG translation initiation codon in theannotated T. denticola genome sequence (36). Interstrain ho-mology is also very high for residues 120 to 198, which includea predicted transmembrane helical domain.
Expression and localization of PrcB. To determine whetherthe hypothetical protein encoded by prcB is expressed in T. den-ticola, we constructed strain CF417, in which prcB is modified toencode a C-terminal 6�His tag and the erm cassette is inserted atthe 5� end of prcA (Fig. 2). Because the N terminus of PrcB couldnot be deduced unambiguously from the DNA sequence, the6�His motif was added to the C terminus of PrcB. The ermcassette contains transcription terminators in both orientations,and native transcription downstream of the insertion site isblocked (5, 6). To determine whether addition of the 6�His tag
TABLE 3. Oligonucleotide primers used in this study
Primer Sequence (restriction enzyme)a Target; primer orientationb
CX360 GGGAAGCTGATTAGTAACCT 5� end of prcA; RCX402 TAAGTCTTGCTAAATCC TDE0759; F with respect to TDE0760CX516 GGCTAGCAAAATCTTTTTCATACTTCCCCCAAAATAG (NheI) prcB region; FCX518 GGGAGCTCAGCTTCCTTTCGGGCTTTG (SacI) 6�His tag in pCF414; RCX529 CCCTCGAGAAAATCTAATTTATCATTTTTAGCATCCCC (XhoI) prcB region; RCX535 GGGAGCTCAAAATCTTTTTCATACTTCCCCCAAAATAG (SacI) prcB-6�His; FCX536 GGCTAGCAGCTTCCTTTCGGGCTTTG (NheI) prcB-6�His; RCX558 AACTCCGGTTACGGCTA TDE0758; F with respect to TDE0760CX578 CCATATGGAATACCTGCCTGTAAAGGAC (NdeI) Between TDE0758 and TDE0759; RCX562 GTTGATCAGATCTCAGTGGTGGTGGTG (BclI) Upstream of prcB-6�His; FCX577 CCATATGTCTGACTTCGAGCTTATC (NdeI) Between TDE0758 and TDE0759; FCX581 GTATACTTATAGGAGGACGAGTATGTC (BstZ17I) prcA RBS; FCX582 GATTCCGCTTGAGCTGTTGTTTC (EcoRI) prcA near 3� end; RCX639 TACCGATGGTACCGGTG prtP internal; FCX640 ATGGGGTTGCCATTGAAG prtP internal; RCX267 CAATACATTCTCAGCCGAAC flaA internal; FCX268 GTCAATACGGGAGACTTAAG flaA internal; RCX710 GCATATGATATACTTAAGTACTATG (NdeI) prcB Met1; FCX711 GCATATGAGATTTTGTGTAATTTTAT (NdeI) prcB Met7; FCX712 GCATATGCCTGATGACAAAGTT (NdeI) prcB Val38; FCX713 AAGCTTCCTTTCGGGCTTTG (HindIII) prcB 3� end; R
a Engineered restriction enzyme sites are underlined.b F, forward; R, reverse.
VOL. 192, 2010 PrcB IS REQUIRED FOR T. DENTICOLA PrtP PROTEASE ACTIVITY 3339

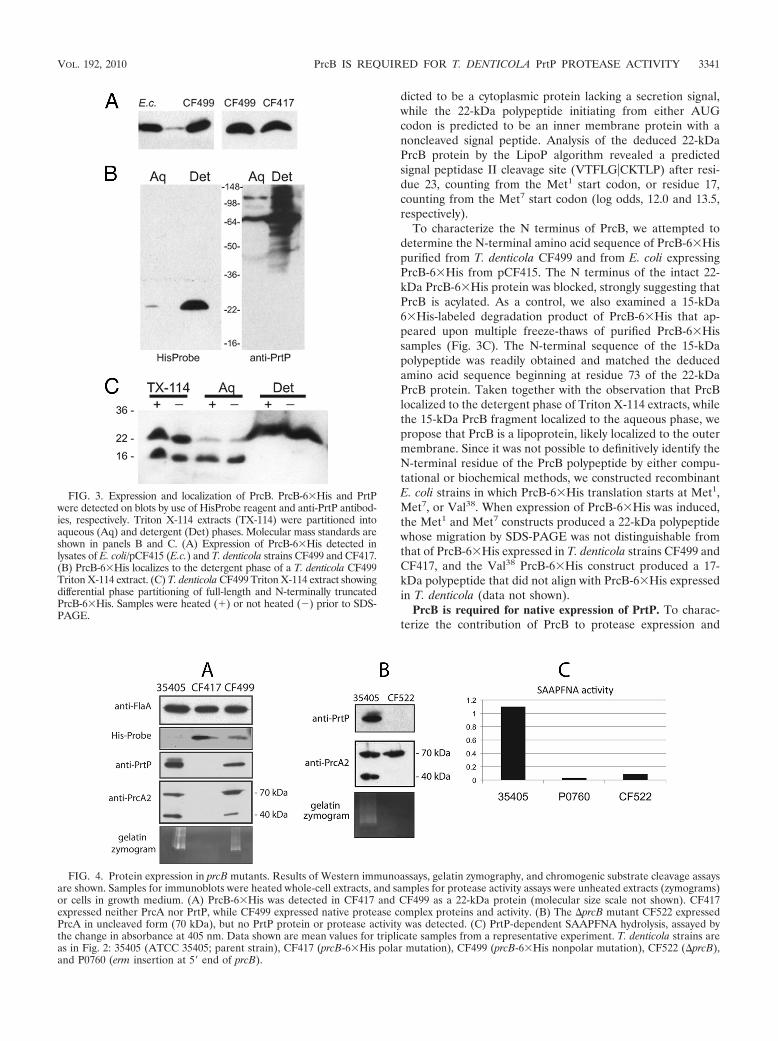
to PrcB affected protease expression and activity, we also con-structed CF499, in which the protease operon is wild type exceptfor the 6�His modification of prcB. In CF499, the erm cassette islocated between TDE0758 and TDE0759, upstream of the pro-tease locus (Fig. 2). The predicted protease operon promoterregion upstream of prcB is intact in both CF499 and CF417. Asshown in Fig. 3A, PrcB-6�His is a 22-kDa protein in T. denticolastrains CF417 and CF499 and in the recombinant E. coli straincarrying pCF415, in which expression of PrcB-6�His is initiatedfrom the native T. denticola promoter in the plasmid.
PrcB-6�His localized to the detergent phase of TritonX-114 extracts of T. denticola CF499 (Fig. 3B) and was foundonly in small amounts in the cell pellet fraction of the extract.This suggests that PrcB is very hydrophobic and may localize to
the outer membrane of T. denticola. Interestingly, a 15-kDa6�His-labeled degradation product of PrcB-6�His that ap-peared upon multiple freeze-thaws of PrcB-6�His sampleslocalized to the aqueous phase of Triton X-114 extracts (Fig.3C), demonstrating that the PrcB N terminus (not the C-terminal predicted transmembrane domain) is responsible forpartitioning of PrcB to the hydrophobic (detergent) phase ofthe Triton X-114 extract.
As expected, CF417 lacked PrcA, PrtP, and protease activitydue to polar effects of erm insertion at the 5� end of prcA (Fig.4A). In contrast, CF499 expressed protease activity and PrtPprotein at essentially wild-type levels, suggesting that the C-terminal modification of PrcB has only a minimal effect onprotease expression. In both CF499 and the parent strain,PrcA was detected as both the full-length form (70 kDa) andthe 40-kDa PrcA2 cleavage product resulting from PrtP activitywhen blots were probed with anti-PrcA2 antibodies (Fig. 4A).
N-terminal analysis of PrcB. Current annotations ofTDE0760 (prcB) by the J. Craig Venter Institute (JCVI) (http://cmr.jcvi.org/) and the Los Alamos National Laboratory(LANL) (http://www.oralgen.lanl.gov/) are somewhat ambigu-ous. Both identify the translation initiation codon as a GUGcodon 112 bp downstream of the initial AUG codon in theORF. However, predicted protein molecular masses are 17kDa (JCVI; consistent with the GUG start codon, encodingVal38) and 22 kDa (LANL; consistent with either the first orsecond AUG as the start codon, encoding Met1 or Met7 [re-sulting in a 21.8-kDa or 21.1-kDa protein, respectively]). Anal-ysis of the TDE0760 ORF by the FGENESB algorithm pre-dicted that translation of the TDE0760 ORF initiates at thesecond AUG codon (Met7) (data not shown). There is noclearly identifiable ribosome binding site (RBS) immediatelyupstream of either potential AUG start codon, although theJCVI annotation identifies the sequence GGGGA 10 bp up-stream of the proposed GUG start codon. It should be notedthat the RBS identification algorithm used by JCVI assumesthat the start codon has been predicted correctly (39). A po-tential �70 class promoter is located 62 bp upstream of theMet1 AUG codon (not shown). Using software designed toidentify type I signal peptides, the 17-kDa polypeptide is pre-
FIG. 1. Conservation of prcB in T. denticola. The deduced amino acid sequence of the complete ORF, including the PrcB coding region, isshown for T. denticola strains ATCC 35405, ATCC 33520, and OTK, aligned using Clustal 2.0. The N-terminal valine residue predicted by JCVI(Val38) and the two possible N-terminal methionine residues (Met1 and Met7) predicted by the present study are shaded gray. The predicted signalpeptidase II cleavage site is indicated by a black arrowhead. Amino acid identity throughout all three sequences is indicated by asterisks. Conservedand semiconserved amino acids are shown by colons and dots, respectively.
FIG. 2. Protease locus in T. denticola parent and prcB mutantstrains. The genes of the protease operon, prcB, prcA, and prtP, areshown as gray arrows. The location of the protease operon promoterregion is indicated by “P.” The protease mRNA transcript is shown asa thin arrow above the genes transcribed in each strain. The locationand orientation of the erm cassette are shown by a black arrow. The T.denticola strains are 35405 (wild-type parent), P0760 (erm insertion atthe 5� end of prcB), CF417 (prcB modified to encode a C-terminal Histag; erm insertion replaces the 5� end of prcA), CF499 (prcB modifiedto encode a C-terminal His tag; erm insertion is upstream of theprotease locus), and CF522 (�prcB; promoter region is intact; erminsertion as in CF499).
3340 GODOVIKOVA ET AL. J. BACTERIOL.
dicted to be a cytoplasmic protein lacking a secretion signal,while the 22-kDa polypeptide initiating from either AUGcodon is predicted to be an inner membrane protein with anoncleaved signal peptide. Analysis of the deduced 22-kDaPrcB protein by the LipoP algorithm revealed a predictedsignal peptidase II cleavage site (VTFLG�CKTLP) after resi-due 23, counting from the Met1 start codon, or residue 17,counting from the Met7 start codon (log odds, 12.0 and 13.5,respectively).
To characterize the N terminus of PrcB, we attempted todetermine the N-terminal amino acid sequence of PrcB-6�Hispurified from T. denticola CF499 and from E. coli expressingPrcB-6�His from pCF415. The N terminus of the intact 22-kDa PrcB-6�His protein was blocked, strongly suggesting thatPrcB is acylated. As a control, we also examined a 15-kDa6�His-labeled degradation product of PrcB-6�His that ap-peared upon multiple freeze-thaws of purified PrcB-6�Hissamples (Fig. 3C). The N-terminal sequence of the 15-kDapolypeptide was readily obtained and matched the deducedamino acid sequence beginning at residue 73 of the 22-kDaPrcB protein. Taken together with the observation that PrcBlocalized to the detergent phase of Triton X-114 extracts, whilethe 15-kDa PrcB fragment localized to the aqueous phase, wepropose that PrcB is a lipoprotein, likely localized to the outermembrane. Since it was not possible to definitively identify theN-terminal residue of the PrcB polypeptide by either compu-tational or biochemical methods, we constructed recombinantE. coli strains in which PrcB-6�His translation starts at Met1,Met7, or Val38. When expression of PrcB-6�His was induced,the Met1 and Met7 constructs produced a 22-kDa polypeptidewhose migration by SDS-PAGE was not distinguishable fromthat of PrcB-6�His expressed in T. denticola strains CF499 andCF417, and the Val38 PrcB-6�His construct produced a 17-kDa polypeptide that did not align with PrcB-6�His expressedin T. denticola (data not shown).
PrcB is required for native expression of PrtP. To charac-terize the contribution of PrcB to protease expression and
FIG. 3. Expression and localization of PrcB. PrcB-6�His and PrtPwere detected on blots by use of HisProbe reagent and anti-PrtP antibod-ies, respectively. Triton X-114 extracts (TX-114) were partitioned intoaqueous (Aq) and detergent (Det) phases. Molecular mass standards areshown in panels B and C. (A) Expression of PrcB-6�His detected inlysates of E. coli/pCF415 (E.c.) and T. denticola strains CF499 and CF417.(B) PrcB-6�His localizes to the detergent phase of a T. denticola CF499Triton X-114 extract. (C) T. denticola CF499 Triton X-114 extract showingdifferential phase partitioning of full-length and N-terminally truncatedPrcB-6�His. Samples were heated () or not heated (�) prior to SDS-PAGE.
FIG. 4. Protein expression in prcB mutants. Results of Western immunoassays, gelatin zymography, and chromogenic substrate cleavage assaysare shown. Samples for immunoblots were heated whole-cell extracts, and samples for protease activity assays were unheated extracts (zymograms)or cells in growth medium. (A) PrcB-6�His was detected in CF417 and CF499 as a 22-kDa protein (molecular size scale not shown). CF417expressed neither PrcA nor PrtP, while CF499 expressed native protease complex proteins and activity. (B) The �prcB mutant CF522 expressedPrcA in uncleaved form (70 kDa), but no PrtP protein or protease activity was detected. (C) PrtP-dependent SAAPFNA hydrolysis, assayed bythe change in absorbance at 405 nm. Data shown are mean values for triplicate samples from a representative experiment. T. denticola strains areas in Fig. 2: 35405 (ATCC 35405; parent strain), CF417 (prcB-6�His polar mutation), CF499 (prcB-6�His nonpolar mutation), CF522 (�prcB),and P0760 (erm insertion at 5� end of prcB).
VOL. 192, 2010 PrcB IS REQUIRED FOR T. DENTICOLA PrtP PROTEASE ACTIVITY 3341
activity, we constructed CF522, a �prcB mutant in which theputative protease locus promoter region is intact (Fig. 2). InCF522, as in CF499, erm is located upstream of the proteaselocus, between TDE0758 and TDE0759. As shown in Fig. 4B,CF522 produced an uncleaved full-length PrcA protein butproduced no detectable PrtP protein or gelatinase activity. Inboth the parent strain 35405 and in CF499, cleavage of PrcA
from the full-length form (70 kDa) to a cleaved form (PrcA2;40 kDa) was evident (Fig. 4A), while CF522 produced onlyuncleaved full-length PrcA (Fig. 4B), consistent with thelack of PrtP gelatinase activity. In a quantitative assay ofSAAPFNA degradation, CF522 had a level of activity sim-ilar to that of the negative-control strain P0760 (Fig. 4C).
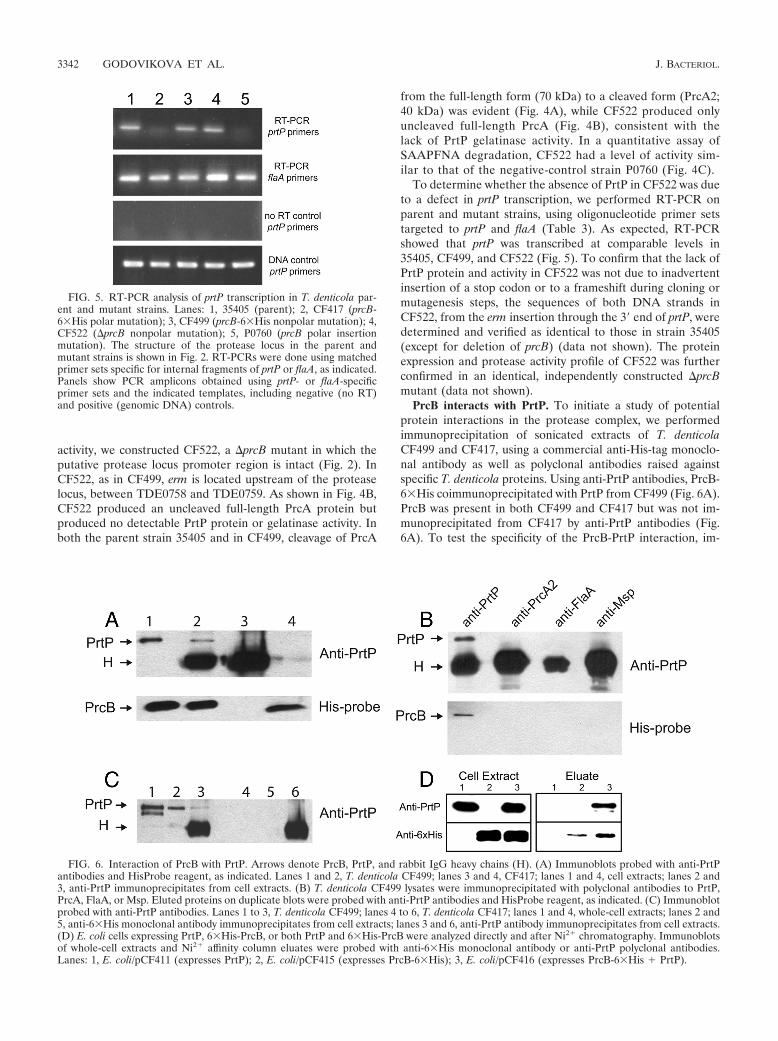
To determine whether the absence of PrtP in CF522 was dueto a defect in prtP transcription, we performed RT-PCR onparent and mutant strains, using oligonucleotide primer setstargeted to prtP and flaA (Table 3). As expected, RT-PCRshowed that prtP was transcribed at comparable levels in35405, CF499, and CF522 (Fig. 5). To confirm that the lack ofPrtP protein and activity in CF522 was not due to inadvertentinsertion of a stop codon or to a frameshift during cloning ormutagenesis steps, the sequences of both DNA strands inCF522, from the erm insertion through the 3� end of prtP, weredetermined and verified as identical to those in strain 35405(except for deletion of prcB) (data not shown). The proteinexpression and protease activity profile of CF522 was furtherconfirmed in an identical, independently constructed �prcBmutant (data not shown).
PrcB interacts with PrtP. To initiate a study of potentialprotein interactions in the protease complex, we performedimmunoprecipitation of sonicated extracts of T. denticolaCF499 and CF417, using a commercial anti-His-tag monoclo-nal antibody as well as polyclonal antibodies raised againstspecific T. denticola proteins. Using anti-PrtP antibodies, PrcB-6�His coimmunoprecipitated with PrtP from CF499 (Fig. 6A).PrcB was present in both CF499 and CF417 but was not im-munoprecipitated from CF417 by anti-PrtP antibodies (Fig.6A). To test the specificity of the PrcB-PrtP interaction, im-
FIG. 5. RT-PCR analysis of prtP transcription in T. denticola par-ent and mutant strains. Lanes: 1, 35405 (parent); 2, CF417 (prcB-6�His polar mutation); 3, CF499 (prcB-6�His nonpolar mutation); 4,CF522 (�prcB nonpolar mutation); 5, P0760 (prcB polar insertionmutation). The structure of the protease locus in the parent andmutant strains is shown in Fig. 2. RT-PCRs were done using matchedprimer sets specific for internal fragments of prtP or flaA, as indicated.Panels show PCR amplicons obtained using prtP- or flaA-specificprimer sets and the indicated templates, including negative (no RT)and positive (genomic DNA) controls.
FIG. 6. Interaction of PrcB with PrtP. Arrows denote PrcB, PrtP, and rabbit IgG heavy chains (H). (A) Immunoblots probed with anti-PrtPantibodies and HisProbe reagent, as indicated. Lanes 1 and 2, T. denticola CF499; lanes 3 and 4, CF417; lanes 1 and 4, cell extracts; lanes 2 and3, anti-PrtP immunoprecipitates from cell extracts. (B) T. denticola CF499 lysates were immunoprecipitated with polyclonal antibodies to PrtP,PrcA, FlaA, or Msp. Eluted proteins on duplicate blots were probed with anti-PrtP antibodies and HisProbe reagent, as indicated. (C) Immunoblotprobed with anti-PrtP antibodies. Lanes 1 to 3, T. denticola CF499; lanes 4 to 6, T. denticola CF417; lanes 1 and 4, whole-cell extracts; lanes 2 and5, anti-6�His monoclonal antibody immunoprecipitates from cell extracts; lanes 3 and 6, anti-PrtP antibody immunoprecipitates from cell extracts.(D) E. coli cells expressing PrtP, 6�His-PrcB, or both PrtP and 6�His-PrcB were analyzed directly and after Ni2 chromatography. Immunoblotsof whole-cell extracts and Ni2 affinity column eluates were probed with anti-6�His monoclonal antibody or anti-PrtP polyclonal antibodies.Lanes: 1, E. coli/pCF411 (expresses PrtP); 2, E. coli/pCF415 (expresses PrcB-6�His); 3, E. coli/pCF416 (expresses PrcB-6�His PrtP).
3342 GODOVIKOVA ET AL. J. BACTERIOL.

munoprecipitations were also done with antibodies raisedagainst PrtP, PrcA2, FlaA, and Msp (Fig. 6B). PrcB and PrtPwere coimmunoprecipitated from strain CF499 with anti-PrtPantibodies, but PrcB was not detected in the eluate fraction ofimmunoprecipitations done with polyclonal antibodies raisedagainst PrcA2, FlaA, or Msp. These data suggest that PrcBdoes not interact with PrcA, the other major component of theprotease complex. As shown in Fig. 6C, PrtP was coimmuno-precipitated with PrcB-6�His by use of anti-His-tag antibody,further demonstrating the PrtP-PrcB interaction. To generatea recombinant system for further study of protein interactionsin the protease complex, PrtP and PrcB-6�His were coex-pressed in E. coli. Extracts from E. coli strains expressing PrtP,PrcB-6�His, or both proteins were subjected to Ni2 affinitychromatography. As shown in Fig. 6D, PrtP eluted with PrcB-6�His from E. coli/pCF416 (lanes 3), but PrtP alone expressedin E. coli/pCF411 was not retained on the affinity column(lanes 1). These results provide further evidence of a directinteraction between PrtP and PrcB and provide a workablerecombinant system for molecular analysis of this interaction.
DISCUSSION
Considerable effort has gone into characterization of therole of T. denticola PrtP protease (dentilisin) in periodontaldisease, but little is known about how components of the pro-tease complex are secreted, translocated through the periplas-mic space, assembled, and anchored in the outer membrane.The present study addressed the role of PrcB in expression andactivity of the protease complex. The conserved structure ofthe prcB-prcA-prtP operon is consistent with functional linkageof PrcB with PrtP protease activity. While PrcB has not beenreported as a component of the protease complex, our resultsindicate that PrcB interacts with PrtP and contributes to stabilityof the native protease complex. The most likely reason that thisassociation has not been noted previously is that the PrtP-PrcBinteraction does not appear to be stable under SDS-PAGE con-ditions.
The addition of a C-terminal 6�His tag facilitated detectionof PrcB in T. denticola and did not appear to interfere with itsfunction, as reflected in protease expression and activity instrain CF499 that were comparable to those in the parentstrain. As noted, the choice of C- versus N-terminal 6�Hislabeling of PrcB was due to the ambiguous nature of the Nterminus of native PrcB. This is the first use of this technologyin T. denticola, and it provides a relatively rapid and econom-ical means of detecting expression of “hypothetical proteins”of interest identified in the genome annotation without gener-ating protein-specific detection reagents.
Native prtP transcription downstream of the insertion site inT. denticola P0760 and CF417 is blocked because the ermcassette contains transcription terminators that are functionalin both orientations (5, 6). However, prtP was transcribed atsimilar levels in the �prcB mutant CF522 and the parent strain35405. The absence of PrtP protein in CF522 was not due toeither a defect in transcription or the introduction of a frame-shift during mutagenesis. Taken together with the demon-strated PrcB-PrtP interaction, this suggests that PrtP is trans-lated in CF522 but is subsequently degraded due to misfoldingor incorrect localization in the absence of PrcB. Based on the
phenotype of CF522 and the current genome annotation ofPrcB as a cytoplasmic protein, we initially hypothesized thatPrcB serves as a PrtP-specific molecular chaperone to stabilizePrtP prior to or during its secretion across the inner mem-brane. However, our data demonstrating the molecular weight,cellular localization, and likely acylation of PrcB are not con-sistent either with this model or with the current genomeannotation, which predicts that PrcB translation initiates atVal38, counting from the first AUG codon. While we wereunable to experimentally identify the PrcB translation initia-tion site, our data are consistent with its being the second AUGcodon (Met7), as predicted by the FGENESB algorithm. Theperfect homology between deduced amino acid sequences ofthis N-terminal region of PrcB in diverse T. denticola strains isalso supportive of this interpretation. Annotation of prcB iscomplicated by the absence of an efficient consensus ribosomebinding site upstream of any of the three potential start codonsin the TDE0760 ORF. This is in stark contrast to the case forprcA and prtP, each of which has a consensus Shine-Dalgarnosequence (AGGAGG) within 8 bp of the respective AUG startcodon. Preliminary data (not shown) suggesting that the PrcBprotein is expressed at lower levels than PrcA and PrtP areconsistent with the DNA sequence information. This issue isbeing addressed in ongoing studies that are beyond the scopeof the present work.
Prediction of signal peptidase II cleavage and acylation sitesremains somewhat problematic for T. denticola and other spi-rochetes. As noted by Setubal et al. (37), spirochete lipoboxsequences are much less clearly defined than those of otherbacteria. PrcB, PrcA, and PrtP may or may not be identified aslipoproteins, depending on the algorithm used for analysis.The LipoP algorithm (27) identifies high-probability type IIsignal peptidase cleavage sites for all three proteins (forPrcB, VTFLG�CKTLP, with log odds of 12.0/13.5; for PrcA,FLFGS�CPQQK, with log odds of 15.4; and for PrtP,LIVSS�CNFGM, with log odds of 20.8). In contrast, the SpLipalgorithm, designed specifically for lipoprotein identification inspirochetes (37), predicts neither PrcB nor PrtP as a lipoprotein.Interestingly, although we have experimentally confirmed its ac-ylation (4, 18), the SpLip algorithm also fails to identify T. denti-cola OppA as a lipoprotein (data not shown).
Interactions between PrcB and PrtP were assayed both in T.denticola and in an E. coli background, for several reasons.First, as noted above, PrcB appears to be expressed at rela-tively low levels in T. denticola. Second, the PrtP polypeptideexpressed in E. coli does not have proteolytic activity (26),while high levels of PrtP protease activity in T. denticola ex-tracts complicate the possibility of obtaining unambiguous re-sults that are dependent on intact, stable protein. Third, theuse of two different ligand-receptor assay systems increases thereliability of results. The ability of PrcB-6�His to bind PrtP ineither environment is indicative of a direct interaction betweenPrtP and PrcB and supports the hypothesis that PrcB-PrtPinteraction is a key feature in expression of the PrtP proteasecomplex. The native protease consists of a complex of PrtP,PrcA1, and PrcA2 that, if unheated, does not dissociate into itsconstituent polypeptides in SDS-PAGE analysis (40). The acylmoieties of the PrcA and PrtP lipoproteins presumably serve inlocalization to the outer membrane. However, in their activeforms, both proteins are reported to undergo further process-
VOL. 192, 2010 PrcB IS REQUIRED FOR T. DENTICOLA PrtP PROTEASE ACTIVITY 3343

ing: PrtP is activated by cleavage at residue 158 (26), and the70-kDa PrcA protein is cleaved to PrcA1 (30 kDa) and PrcA2(40 kDa) by PrtP activity (29). While PrcB has not been de-tected as part of the SDS-stable active protease complex, ourdata suggest that the acyl moiety of PrcB serves to anchor it inthe outer membrane. It is not yet clear whether PrcB is an-chored at the periplasmic face of the outer membrane or isexposed on the cell surface. In either case, PrcB most likelyfunctions in either presentation or stabilization of the proteaseat the cell surface. The specific protein-protein interactionsrequired for stability and localization of the outer membraneprotease complex remain to be studied more fully. We arecontinuing to investigate the specific contribution of PrcB toexpression of the protease complex, including defining theinteracting domains of PrtP and PrcB.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grants DE013565and DE018221 (National Institute of Dental and Craniofacial Re-search), by the Office of the Vice President for Research (University ofMichigan), and by the Undergraduate Research Opportunity Program(University of Michigan).
REFERENCES
1. Asai, Y., T. Jinno, and T. Ogawa. 2003. Oral treponemes and their outermembrane extracts activate human gingival epithelial cells through Toll-likereceptor 2. Infect. Immun. 71:717–725.
2. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A.Smith, and K. Struhl (ed.). 1995. Current protocols in molecular biology, vol.1. Wiley-Interscience, New York, NY.
3. Bamford, C. V., J. C. Fenno, H. F. Jenkinson, and D. Dymock. 2007. Thechymotrypsin-like protease (CTLP) complex of Treponema denticola ATCC35405 mediates fibrinogen adherence and degradation. Infect. Immun. 75:4364–4372.
4. Bayer, C. H., Y. Ning, and J. C. Fenno. 2003. Recombinant expressionsystems for Treponema denticola OppA. J. Dent. Res. 82:S1416.
5. Bian, X.-L., H.-T. Wang, Y. Ning, S. Y. Lee, and J. C. Fenno. 2005. Mu-tagenesis of a novel gene in the prcA-prtP protease locus affects expression ofTreponema denticola membrane complexes. Infect. Immun. 73:1252–1255.
6. Bian, X. L., and J. C. Fenno. 2003. Transcription of msp and prcA-prtP inTreponema denticola parent and isogenic mutant strains. J. Dent. Res. 82:S1587.
7. Chan, E. C. S., A. DeCiccio, R. McLaughlin, A. Klitorinos, and R. Siboo.1997. An inexpensive solid medium for obtaining colony-forming units oforal spirochetes. Oral Microbiol. Immunol. 12:372–376.
8. Chi, B., M. Qi, and H. K. Kuramitsu. 2003. Role of dentilisin in Treponemadenticola epithelial cell layer penetration. Res. Microbiol. 154:637–643.
9. Correia, F. F., A. R. Plummer, R. P. Ellen, C. Wyss, S. K. Boches, J. L.Galvin, B. J. Paster, and F. E. Dewhirst. 2003. Two paralogous families of atwo-gene subtilisin operon are widely distributed in oral treponemes. J.Bacteriol. 185:6860–6869.
10. Coutte, L., E. Willery, R. Antoine, H. Drobecq, C. Locht, and F. Jacob-Dubuisson. 2003. Surface anchoring of bacterial subtilisin important formaturation function. Mol. Microbiol. 49:529–539.
11. Cunningham, T. M., E. M. Walker, J. N. Miller, and M. A. Lovett. 1988.Selective release of the Treponema pallidum outer membrane and associatedpolypeptides with Triton X-114. J. Bacteriol. 170:5789–5796.
12. Deng, Q. D., Y. Han, X. Xia, and H. K. Kuramitsu. 2001. Effects of the oralspirochete Treponema denticola on interleukin-8 expression from epithelialcells. Oral Microbiol. Immunol. 16:185–187.
13. Ellen, R. P., and V. B. Galimanas. 2005. Spirochetes at the forefront ofperiodontal infections. Periodontol. 2000 38:13–32.
14. Emanuelsson, O., S. Brunak, G. von Heijne, and H. Nielsen. 2007. Locatingproteins in the cell using TargetP, SignalP and related tools. Nat. Protoc.2:953–971.
15. Fenno, J. C., S. Y. Lee, C. H. Bayer, and Y. Ning. 2001. The opdB locusencodes the trypsin-like peptidase activity of Treponema denticola. Infect.Immun. 69:6193–6200.
16. Fenno, J. C., and B. C. McBride. 1998. Virulence factors of oral treponemes.Anaerobe 4:1–17.
17. Fenno, J. C., K.-H. Muller, and B. C. McBride. 1996. Sequence analysis,expression and binding activity of recombinant major outer sheath protein(Msp) of Treponema denticola. J. Bacteriol. 178:2489–2497.
18. Fenno, J. C., M. Tamura, P. M. Hannam, G. W. K. Wong, R. A. Chan, andB. C. McBride. 2000. Identification of a Treponema denticola OppA homo-logue that binds host proteins present in the subgingival environment. Infect.Immun. 68:1884–1892.
19. Fenno, J. C., G. W. K. Wong, P. M. Hannam, and B. C. McBride. 1998.Mutagenesis of outer membrane virulence determinants of the oral spiro-chete Treponema denticola. FEMS Microbiol. Lett. 163:209–215.
20. Fenno, J. C., G. W. K. Wong, P. M. Hannam, K.-H. Muller, W. K. Leung, andB. C. McBride. 1997. Conservation of msp, the gene encoding the majorouter membrane protein of oral Treponema spp. J. Bacteriol. 179:1082–1089.
21. Fletcher, H. M., H. A. Schenkein, R. M. Morgan, K. A. Bailey, C. R. Berry,and F. L. Macrina. 1995. Virulence of a Porphyromonas gingivalis W83mutant defective in the prtH gene. Infect. Immun. 63:1521–1528.
22. Grenier, D. 1996. Degradation of host protease inhibitors and activation ofplasminogen by proteolytic enzymes from Porphyromonas gingivalis andTreponema denticola. Microbiology 142:955–961.
23. Grenier, D., V.-J. Uitto, and B. C. McBride. 1990. Cellular location of a Trepo-nema denticola chymotrypsin-like protease and importance of the protease inmigration through the basement membrane. Infect. Immun. 58:347–351.
24. Haapasalo, M., U. Singh, B. C. McBride, and V.-J. Uitto. 1991. Sulfhydryl-dependent attachment of Treponema denticola to laminin and other proteins.Infect. Immun. 59:4230–4237.
25. Hashimoto, M., S. Ogawa, Y. Asai, Y. Takai, and T. Ogawa. 2003. Binding ofPorphyromonas gingivalis fimbriae to Treponema denticola dentilisin. FEMSMicrobiol. Lett. 226:267–271.
26. Ishihara, K., T. Miura, H. K. Kuramitsu, and K. Okuda. 1996. Character-ization of the Treponema denticola prtP gene encoding a prolyl-phenylala-nine-specific protease (dentilisin). Infect. Immun. 64:5178–5186.
27. Juncker, A. S., H. Willenbrock, G. Von Heijne, S. Brunak, H. Nielsen, and A.Krogh. 2003. Prediction of lipoprotein signal peptides in Gram-negativebacteria. Protein Sci. 12:1652–1662.
28. Kent, C., P. Gee, S. Y. Lee, X. Bian, and J. C. Fenno. 2004. A CDP-cholinepathway for phosphatidylcholine biosynthesis in Treponema denticola. Mol.Microbiol. 51:471–481.
29. Lee, S. Y., X. L. Bian, G. W. Wong, P. M. Hannam, B. C. McBride, and J. C.Fenno. 2002. Cleavage of Treponema denticola PrcA polypeptide to yieldprotease complex-associated proteins PrcA1 and PrcA2 is dependent onPrtP. J. Bacteriol. 184:3864–3870.
30. Lee, S. Y., and J. C. Fenno. 2004. Expression of Treponema denticolaoligopeptidase B in Escherichia coli. Curr. Microbiol. 48:379–382.
31. Li, H., J. Ruby, N. Charon, and H. Kuramitsu. 1996. Gene inactivation in theoral spirochete Treponema denticola: construction of an flgE mutant. J. Bac-teriol. 178:3664–3667.
32. Makinen, K. K., and P. L. Makinen. 1996. The peptidolytic capacity of thespirochete system. Med. Microbiol. Immunol. 185:1–10.
33. McDowell, J. V., B. Huang, J. C. Fenno, and R. T. Marconi. 2009. Analysisof a unique interaction between the complement regulatory protein factor Hand the periodontal pathogen Treponema denticola. Infect. Immun. 77:1417–1425.
34. Nakai, K., and M. Kanehisa. 1991. Expert system for predicting proteinlocalization sites in gram-negative bacteria. Proteins 11:95–110.
35. Rosen, G., R. Naor, S. Kutner, and M. N. Sela. 1994. Characterization offibrinolytic activities of Treponema denticola. Infect. Immun. 62:1749–1754.
36. Seshadri, R., G. S. Myers, H. Tettelin, J. A. Eisen, J. F. Heidelberg, R. J.Dodson, T. M. Davidsen, R. T. DeBoy, D. E. Fouts, D. H. Haft, J. Selengut,Q. Ren, L. M. Brinkac, R. Madupu, J. Kolonay, S. A. Durkin, S. C. Daugh-erty, J. Shetty, A. Shvartsbeyn, E. Gebregeorgis, K. Geer, G. Tsegaye, J.Malek, B. Ayodeji, S. Shatsman, M. P. McLeod, D. Smajs, J. K. Howell, S.Pal, A. Amin, P. Vashisth, T. Z. McNeill, Q. Xiang, E. Sodergren, E. Baca,G. M. Weinstock, S. J. Norris, C. M. Fraser, and I. T. Paulsen. 2004.Comparison of the genome of the oral pathogen Treponema denticola withother spirochete genomes. Proc. Natl. Acad. Sci. U. S. A. 101:5646–5651.
37. Setubal, J. C., M. Reis, J. Matsunaga, and D. A. Haake. 2006. Lipoproteincomputational prediction in spirochaetal genomes. Microbiology 152:113–121.
38. Sorsa, T., T. Ingman, K. Suomalainen, M. Haapasalo, Y. T. Konttinen, O.Lindy, H. Saari, and V.-J. Uitto. 1992. Identification of proteases fromperiodontopathogenic bacteria as activators of latent human neutrophil andfibroblast-type interstitial collagenases. Infect. Immun. 60:4491–4495.
39. Suzek, B. E., M. D. Ermolaeva, M. Schreiber, and S. L. Salzberg. 2001. Aprobabilistic method for identifying start codons in bacterial genomes. Bioin-formatics 17:1123–1130.
40. Uitto, V.-J., D. Grenier, E. C. Chan, and B. C. McBride. 1988. Isolation of achymotrypsin-like enzyme from Treponema denticola. Infect. Immun. 56:2717–2722.
41. Yamazaki, T., M. Miyamoto, S. Yamada, K. Okuda, and K. Ishihara. 2006.Surface protease of Treponema denticola hydrolyzes C3 and influences func-tion of polymorphonuclear leukocytes. Microbes Infect. 8:1758–1763.
42. Yanisch-Perron, C., J. Vieira, and J. Messing. 1985. Improved M13 phagecloning vectors and host strains: nucleotide sequences of the M13mp18 andpUC19 vectors. Gene 33:103–119.
3344 GODOVIKOVA ET AL. J. BACTERIOL.
Copyright © 2022 FDOKUMEN




















