
Identification of novel genes regulated in the developing human ventral mesencephalon
Upload
independentCategory
view
3download
0
THE JOURNAL OF COMPARATIVE NEUROLOGY 235~322-335 (1985)
Efferent Connections of the Ventral Pallidum: Evidence of a Dual Striato
Pallidofugal Pathway
S.N. HABER, H.J. GROENEWEGEN, E.A. GIROVE, ANI) W.J.H. NAUTA Department of Anatomy, The University of Rochester, Rochester, New York 14642
(S.N.H.), Department of Anatom,y, Vrije Universiteit, 1081 BT Amsterdam, The Netherlands (H. J.G.), Department of Psychology and Brain Science, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139, and Mailman Research
Center, McLean Hospital, Belniont, Massachusetts 02178 (E.A.G., W.J.H.N.)
ABSTRACT Previous histological and liistochemical studies have provided evidence
that the globus pallidus (external pallidal segm’ent) as conventionally delin- eated in the rat extends ventra.lly and rostrally beneath the transverse limb of the anterior commissure, invading the olfactory tubercle with its most ventral ramifications. This infracommissural subdivision of the globus pal- lidus or ventral pallidum (VP) is most selectively identified by being per- vaded by a dense plexus of substance-P-positive striatofugal fibers; the extent of this plexus indicates that the VP behi.nd the anterior commissure continues dorsally over some distance into the anteroventromedial part of the generally recognized (supracommissural) globus pallidus; the adjoining anterodorsolateral pallidal region, here named dorsal pallidum (DP), re- ceives only few substance-P-positive fibers, but contains a dense plexus of enkephalin-positive striatal af‘ferents that also pervades VP. Available au- toradiographic data indicate that VP and DP receive their striatal innerva- tion from two different subdivisions of the striatum: whereas VP is innervated by a large, anteroventromedial striatal region receiving substan- tial inputs from a variety of limbic and limbic-system-associated structures (and therefore called “limbic striatum”), DP receives its striatal input from ;an anterodorsolateral striatal sector receiving only sparse limbic afferents Ynonlimbic” striatum) but instead heavily innervated by the sensorimotor cortex. The present autoradiographic study has produced evidence that this dichotomy in the striatopallidal projection is to a large extent continued beyond the globus pallidus: whereas the efferents of DP were traced to the subthalamic nucleus and substantia nigra, those of VP were found to involve not only the subthalamic nucleus and substantia nigra but also the fronto- cingulate (and ajoining medial sensorimotor) cortex, the amygdala, lateral habenular and mediodorsal tlnalamic nucleus, hypothalamus, ventral teg- mental area, and tegmental regions farther caudal and dorsal in the mid- brain. These findings indicate that the veintral pallidum can convey ,striatopallidal outflow of limbic antecedents not only into extrapyramidal ‘circuits but also back into the circuitry of the limbic system.
Key words: corpus striatum, efferent connections, basal forebrain, ventral pallidum
The ventral palliduin was first described by Heimer and Wilson (‘75) in the rat as an extension of the globus palli- dus, stretching ventrally and rostrally beneath the anterior commissure. Its identification as a pallidal subdivision of the infracommissural forebrain region was originally based
on cytological characteristics, as well as on the finding that it receives a massive striatal projection originating in par- ticular from the nucleus accumbens (Wilson, ’72; Williams ~-
Accepted January 11, 1985.
0 1985 ALAN R. LISS, INC.

EFFERENTS OF VENTRAL, PALLIDUM
et al., ’77; Nauta et al., ’78). Its pallidal nature was later documented further by evidence that it also shares with the main mass of the globus pallidus a substantial projection from the subthalamic nucleus (Ricardo, ’80), as well as the histochemical characteristics of a high iron content (Switzer and Hill ’79; Hill and Switzer, ’84) and a strong enkephalin- like (Haber and Nauta, ’81, ’83; Switzer et al., ’82) and glutamic-acid-decarboxylase-like immunoreactivity (Fallon and Ribak, ’80).
Despite these similarities, the ventral pallidum must be viewed as distinct from the main, conventionally recog- nized, retro- and supracommissural mass of the globus pal- lidus. A principal reason for this distinction is histochemical: whereas both the ventral pallidum and the main mass of the globus pallidus are infiltrated by a dense plexus of enkephalin-positive fibers, the ventral pallidum contains an additional and almost equally dense plexus of substance-P-positive fibers that extends over only a short distance into the main pallidal mass. For a proper under- standing of the criteria followed in defining the ventral pallidum for the purposes of the present study, a brief elaboration of this last point is necessary.
It is important to note that intrapallidal enkephalin and substance P are not intrinsic to the globus pallidus proper, but are associated with the striatopallidal projection. As first observed by Fox et al. (’74) in Golgi material, striato- pallidal fibers enmesh the dendrites of pallidal neurons individually in sleevelike plexuses. The characteristic his- tological picture reflecting this junctional arrangement in material tested immunocytochemically for enkephalin (“woolly fibers,” Haber and Nauta, ’83) provides a light- microscopic criterion for determining the extent of the glo- bus pallidus. In such material the dense plexus of enke- phalin-positive “woolly fibers” filling the main mass of the globus pallidus is seen to continue without interruption ventrally into the infracommissural region where it ex- tends rostrally over a wide area bordering the ventral side of the striatum and bed nucleus of the stria terminalis (Switzer et al., ’82; Haber and Nauta, ’83). Interestingly, such material also shows that this infracommissural plexus extends around the lateral border of the nucleus of the diagonal band into the deep, fibrocellular layer of the olfac- tory tubercle, in agreement with an earlier suggestion, made by Heimer (’78) on cytoarchitectural grounds, that the tubercle is a striatopallidal complex, as well as with the description by Ribak and Fallon (’83) of large cells associ- ated with the islands of Calleja and electron-microscopi- cally indistinguishable from pallidal neurons.
Since the density of the enkephalin-positive intrapallidal fiber plexus varies little through its extent, it cannot serve to delineate the ventral pallidum from the main, supra- and retrocommissural, mass of the globus pallidus. Such a de- lineation is, however, possible in sections processed for sub- stance-P-like immunoreactivity. Such material reveals a dense plexus of substancep-positive “woolly fibers” that, with minor exceptions, is coextensive with the infracom- missural enkephalin-positive plexus; but, unlike the latter, it extends over only a short distance into the main pallidal mass, then abruptly declines in density, and only very sparsely extends through the large remainder of the globus pallidus. The curved line along which this decline in den- sity of the substance-P-positive “woolly fibers” plexus oc- curs suggests itself as the border between the ventral pallidum (VP) and the remaining part of the globus palli- dus, here for purposes of contrast to be referred to as dorsal
323
pallidurn (DP) (Haber and Nauta, ’81, ’83; see also Fig. 3, in which the extent of the substance-P-positive plexus is indicated by a stipple pattern).
In summary, the ventral pallidum is here defined as the largely infracommissural region massively pervaded by en- kephalin-positive and substancep-positive (and probably additional) striatofugal fibers terminating in the periden- dritic manner characteristic of the striatopallidal projec- tion. As illustrated by Young et al. (’84), this region in Nissl- stained material displays throughout its extent the pallidal feature of predominantly medium-sized, triangular, or fusi- form cells, relatively widely spaced either singly or in strandlike clusters, mixed in with which are some large cells of more multangular or round shape that are most likely acetylcholinesterase (AChE)-positive elements of the magno-cellular nucleus of the basal forebrain (see below). Probably in part as a result of uneven grouping of these AChE cells, the cytoarchitectural appearance of the ventral pallidum is somewhat variable locally, and this may ex- plain the fact that before the appropriate histochemical methods came into use the region was not widely recog- nized as a continuous expanse of pallidal tissue. To be more specific: the ventral pallidum as outlined by its substance- P-positive fiber plexus (cf. Fig. 3 of Haber and Nauta, ’83) comprises: (1) an anterior, ventral, and medial part of the conventionally recognized globus pallidus, (2) the region sometimes labelled “substantia innominata” (Swanson, ’76; Mogenson et al., ’83), and (3) an anterior and dorsal part, at least, of the region traditionally outlined as lateral preoptic area.
THE STRIATAL INNERVATION OF THE VENTRAL PALLIDUM
Autoradiographic analysis of the striatopallidal projec- tion (V.B. Domesick, personal communication) indicates that the entire extent of the ventral pallidum as defined by its dense substance-P-positive fiber plexus receives its striatal innervation selectively from a large anterior, ventromedial striatal region that includes, but is not confined to, the nucleus accumbens and olfactory tubercle. The general dis- tribution area of this striatal projection was already indi- cated by earlier autoradiographic findings (Swanson and Cowan, ’75; Nauta et al, ’781, and some features of its topographic organization were demonstrated more recently (Mogenson et al., ’83). The large anteroventromedial stria- tal region from which it originates is, in turn, projected upon in a partially overlapping manner by various struc- tures implicated in the circuitry of the limbic system (hip- pocampus, basolateral nucleus of the amygdala, frontocingulate cortex, and ventral tegmental area) and has therefore been distinguished as “limbic striatum” (Kelley et al., ’83) from the adjacent anterodorsolateral striatal region that receives only sparse limbic afferents but is heavily projected upon by the sensorimotor cortex. The striatopallidal projection from the latter, “nonlimbic” sec- tor of the anterior striatum involves almost exclusively the anterior region of the dorsal pallidum (V.B. Domesick, per- sonal communication, and cf. Fig. 4 of Mogenson et al., ’831, and it thus appears that the limbic and the nonlimbic striatal sector project to topographically and histochemi- cally different subdivisions of the globus pallidus. As this difference raised the question of whether the segregation of the two striatofugal conduction lines might not persist be- yond the globus pallidus, an attempt was made to deter- mine whether the projections from the ventral pallidum

S.N. HABER ET AL. 324
Fig. 1. Fiber labeling following a microelectrophoretic injection of triti- ated leucine in the ventral pallidum. Case RR 37; survival time 4 days, exposure time 12 weeks. Shown in jet black in A is the region in which neurons exhibited evidence of label uptake. The injection site i:; further illustrated by Figure 2A.B. The microelectrode track in this case had re- mained entirely free from contamination with isotope. Abbreviations in this and all subsequent illustrations: abl, basolateral amygdaloid nucleus; AC, anterior commissure; ace, central amygdaloid nucleus; am, medial amyg~ daloid nucleus; A8, lcoation of dopamine cell group A8 (Dalstrom and Fuxe, '64); bns, bed nucleus of stria terminalis; CC, crus cerebri; cgs, central gray substance; CI, capsula interna; CP, caudatoputamen; dbc, decussation of brachium conjunctivum; dm, dorsomedial hypothalamic nucleus; dp, dorsal
pallidum; ep, entopeduncular nucleus; F, fornix; F-CC, frontocingulate cor- tex; FR, fasciculus retroflexus; GP, globus pallidus; H, hippocampus; hl, lateral habenular nucleus; hm, medial habenular nucleus; ht, hypothala- mus; ip, interpeduncular nucleus; Ih, lateral hypothalamic region; md, mediodors;al thalamic nucleus; ML, medial lemniscus; na, nucleus accum- bens; ndb, nucleus of the diagonal band; pf, parsfascicular nucleus; ph, posterior hypothalamic nucleus; pt, parataenial nucleus; rt, nucleus reticu- laris thalami; SM, stria medullaris; SMC: sensorimotor cortex; sn, substan- tia nigra; snc, substantia nigra, pars compacta; snr, substantia nigra, pars reticulata; st, subthalamic nucleus; tpn, nucleus tegmenti pedunculopon- tinus; va, nucleus ventralis anterior thalami; vm, ventromedial bypotha- lamic nucleus; vp, ventral pallidum; vta, ventral tegmental area.

EFFERENTS OF VENTRAL PALLIDUM 325
Figure 1 (continued)
differ from those of the main or dorsal pallidum. The re- sults, described below, have in part been reported earlier in the form of an abstract (Haber et al., ’82).
METHODS In order to label the efferent connections of the VP, mi-
croelectrophoretic injections of tritiated proline and leu- cine, or tritiated leucine alone, were placed in or adjacent to this structure in 28 female adult albino rats (Charles River CD strain) anesthetized with Chloropent, a veteri- nary anesthetic containing chloral hydrates and pentobar- bital (Ft. Dodge Labs). Four days to 2 weeks after the injection the animals were once again deeply anesthetized and then perfused through the heart with saline followed by 4% formaldehyde. The brains were cut on a freezing microtome at 30 pm; the sections were mounted on slides “subbed” with chrome-alum gelatin and coated with Kodak NTB 2 or NTB 3 photographic emulsion. Following autora- diographic exposure for 8-16 weeks, the sections were de- veloped in Kodak D-19 and counterstained with cresyl violet.
To verify apparent projections of the ventral pallidum further, electrophoretic injections of wheat-germ-agglu- tinin-conjugated HRP (WGA-HRP) were placed in areas in which anterograde fiber labeling had been found. Two to 3 days later the animals were anesthetized and perfused with a fixative containing 1.25% glutararaldehyde and 0.1% par- aformaldehyde. Sections of 50 pm were cut on a freezing microtome and processed according to the tetramethylben- zidine method of Mesulam (’78).
RESULTS Of the 28 injections of tritiated amino acids, six were
placed in the center of the infracommissural pallidum, while in each of the remaining cases the injection primarily or solely involved one of the following neighboring structures: the preoptic area, the bed nucleus of the stria terminalis, the striatum, the dorsal pallidum, and the nucleus of the diagonal band of Broca. In brief, comparison of the results of these various injections indicates that the ventral palli- dum shares with the main, or dorsal pallidum projections to the subthalamic nucleus, substantia nigra, and striatum,
but unlike the latter projects in addition to several struc- tures associated with the limbic system. To illustrate this, case RR 37 will be briefly described (Fig. 1).
The injection in this case (Figs. lA, 2A,B) was small and circumscript. It appeared to be confined entirely to the ventral palliduim: even in sections exposed for 16 weeks no evidence of label uptake by neurons could be detected in either the adjacent bed nucleus of the stria terminalis or the nucleus of the diagonal band, Moreover, and quite for- tuitously, the pipette track in this case was virtually uncon- taminated by isotope leakage and consequently was difficult to trace through the cortex and striatum.
From the injection site labeled fibers travel caudally in several directions. One large contingent gathers medially (Fig. 1B) and descends with the medial forebrain bundle, giving rise to a dense labeling of the dorsomedial part of the subthalamic nucleus (Figs. lE, 2C) and sparser labeling of the substantia nigra and ventral tegmental area (Fig. 1F). Throughout its descent the group of labeled fibers decreases in volume, apparently by issuing fibers to various parts of the hypothalamus (Fig. 1C-E). Sparse fiber label- ing extends farther caudally along paramedian and lateral- tegmental routes to the mesencephalic raphe nuclei, cen- tral gray substance, and parabrachial region (Fig. lG,H). Since no loci of increased grain density appear anywhere within either the hypothalamic or tegmental distribution of labeled fibers, the present autoradiographic findings al- low no detailed statement concerning the actual termina- tion sites of VP efferents in these brain regions.
Labeled fibers leaving the VP reach the amygdaloid com- plex by two different routes. A few fibers follow the stria terminalis, while a much larger number trace a more ven- tral path, beneath the lentiform nucleus (Fig. 1C). In the amygdala labeled fibers are widely dispersed but appear to terminate preferentially in the basolateral nucleus (Fig. 1C,D). The present evidence, consistent with earlier find- ings (Otterson, ’80), indicates that basal-forebain projec- tions to the basolateral nucleus originate largely from VP: injection of the basolateral nucleus with WGA-HRP labels numerous VP neurons, including those in the olfactory tubercle, and tritiated amino acids injected into regions immediately surrounding VP label only sporadic fibers to the basolateral nucleus.

326 S.N. HABER ET AL.
throughout VP, including the latter's eKtension into the olfactory tubercle. Regarding the mediodorsal nucleus such retrograde evidence was reported earlier by Velayos et al. ('80), Fallon and Ribak ('80), Goldschmidt and Heimer ('801, and Young et al. ('84).
Finally, a group of dispersed labeled fibers extends ros- trally to the nucleus accumbens and olfactory tubercle. Rostra1 to the septum, sparser but longer fibers of this group curve dorsally, passing along both the medial and lateral side of the striatum to the frontocingulate cortex and an a,djoining dorsal-convexity region including at least a medial part of the sensorimotor area.
Contrasting cases; the dorsal pallidum The more caudal, retrocommissural levels of the ventral
palliduni as defined in the rat by its dense substance-P- positive striatofugal-fiber plexus are represented in Figure 3B by stippling (adapted from Haber add Nauta, '83). As indicated in this drawing, the VP does not border the dorsal palliduni (DP) along a horizontal plane through the ante- rior commissure, but continues dorsally from here over some distance, in particular medially, along the lateral face of the internal capsule. As a result of this dorsal incursion, the main (retro- and supracommissural) mass of the globus pallidus appears divided at these rostral levels into a ven- tromedial zone representing VP, and a dorsolateral region that contains only sparse substance-P-positive fibers and must be considered the rostral part of DP proper. In line with this interpretation, the ventromedihl zone receives its striatal innervation from the limbic-derented striatal re- gion (cf. Nauta et al., '78), whereas the lateral pallidal region (DP proper) is innervated by the nonlimbic striatal sector CV.B. Domesick, personal communication; and see Fig. 4 of Mogenson et al., '83). Of further interest is that, as first reported by Jacobowitz and Palkovits ('741, the me- dial pallidal zone contains numerous strongly AChE-posi- tive cells (heavy black dots in Fig. 3B), and has actually been suggested as the rodent homologue of the primate nucleus basalis (Butcher, '83). Strongly AChE-positive neu- rons are common also in the infracommissural extent of VP (Fig. 3A), but such cells are rare in the dorsolateral pallidal region (Fig. 3Bl.I
The present control material includes nine cases of triti- ated amino-acid injection confined to the rostral part of the retro- and supracommissural (i.e., conventionally recog- nized) globus pallidus. In six of these the injection involved both its medial (AChE-cell- and substapce-P-rich) and lat- eral parts. In no case was the injection confined to the medial part, but an injection restricted to the lateral (DP) part was obtained in three cases, one of which (RR 192) is
Three darkfield photographs of case RR 37 A,B The injection site illustrated by Figure 4. As shown by the chartings, labeled as appearing in sections of 16 week autoradiographic exposure, (A) beneath fibers in these extended (1) caudally to the subthal- the transverse limb of the antenor commissure, (B) 200 pm farther (caudally C Fiber labeling in medial quarter and dorsal rim of the subthalamic amic nllcleus and substantia ni@a the pars reticu- nucleus (autoradiographic exposure 12 weeks) Abbreviations see legend to lata), (2) medially to the nucleus reticularis thalami, Figure 1 confirming an earlier finding in the cat (Nauta, '79, and (3)
Fig 2
_____ A third group Of labeled turns from the
injection site and sequentially follows the inferior thalamic peduncle (Fig. 1B-D) and stria medullaris to the lateral
'In the human brain, the neurons of the magnocellular nucleus ofthe basal forebrain are for the most part aggregated into a large and fairly circumscriot sublenticular mouu. the basal nucleus of - .
habenular nucleus and medial parts of the mediadorsal nucleus (Fig. 1C,D). Accordingly, WGA-HRP injected into either of these thalamic nuclei was found to label neurons
Meynert" Even in the human, however, ramifications of the nu cleus extend dorsally into and along the medullary laminae of the globus pallidus (Folx and Nicolesco, '25)

EFFERENTS OF VENTRAL PALLIDUM 327
C P
Fig. 3. Drawings of two frontal sections, respectively, a t the level of the acetylcholinesterase (AChE)-positive intrapallidal neurons. The numerous anterior commissure (A) and a short distance behind it (B). The substance- AChE-positive cells in the nucleus of the diagonal band and other sublenti- P-positive plexus, co-extensive with the ventral pallidum, is indicated by cular structures (cf. Butcher, '83; Mesulam et al., '83) have been omitted stippling (cf. Haber and Nauta, '83). Coarser black dots represent strongly from these drawings. Abbrevations: see legend to Figure 1.
rostrally and laterally into the striatum (H.J.W. Nauta, '79; Staines et al., '81; Jayaraman, '83; Beckstead, '83); none could be traced to the mediodorsal nucleus, habenula, amygdala, or midbrain tegmentum.
A similar labeling pattern was seen following isotope injections involving both the lateral and the medial (VP) part of the rostra1 globus pallidus, but in these cases nu- merous additional fibers were labeled. A case in point (RR 49) is illustrated by Figure 5 . The isotope injection in this case (Fig. 5B) was confined to supra- and retrocommissural levels, and thus was located entirely within the borders of the globus pallidus as conventionally outlined. The result- ing anterograde labeling combined evidence of both dorsal- pallidum (RR 192, Fig. 4) and ventral-pallidum (RR 37, Fig. 1) efferents from the injected region. In common with RR 192 it included fiber labeling in the reticular nucleus of the thalamus (Fig. 5B), while it shared with both cases a sub- stantial labeling in the subthalamic nucleus and substantia nigra (Fig. 5D,E), and only with case RR 37 further labeled fibers to the anteromedial and adjacent convexity cortex (Fig. 5A), amygdala (Fig. 5C), lateral habenular and medi- odorsal thalamic nucleus (Fig. 5C,D), ventral tegmental area (Fig. 5E), and more caudal and dorsal regions of the midbrain tegmentum (Fig. 5F). The only notable difference with case RR 37 was that fewer labeled fibers descended in the medial forebrain bundle and that, accordingly, fiber labeling in the hypothalamus was sparser.
Perhaps the most significant conclusion to be drawn from case RR 49 is that projections to the various limbic-system- associated fore- and midbrain structures enumerated above originate not only ventral to the anterior commissure (as shown by RR 37) but also farther caudally and dorsally, from within the generally accepted limits of the globus pallidus. All of the available evidence indicates that the region giving rise to such projections is a continuum that extends from the anteroventromedial region of the globus pallidus ventrally and forward underneath the anterior commissure.
DISCUSSION The present findings further document the pallidal na-
ture of the infracommissural region designated ventral pal- lidum (VP). Not only does the region share with the main, retrocommissural mass of the globus pallidus (dorsal palli- dum; DP) several features of its cytology, histochemistry, and afferent connectivity; the combination of its efferent connections with the subthalamic nucleus and substantia nigra likewise is characteristically pallidal. That the VP must nonetheless be viewed as a distinctive part of the globus pallidus is indicated by (1) its dense plexus of sub- stance-P-positive striatopallidal fibers, not shared by the DP; (2) its selective innervation by the anteroventromedial, limbic-afferented striatal quadrant; and (3) the evidence that its projections involve not only the subthalamic nu- cleus and substantia nigra but also various limbic-system- associated structures: amygdala, anteromedial cortex, lat- eral habenular nucleus, mediodorsal thalamic nucleus, hy- pothalamus, ventral tegmental area, and more caudal and dorsal regions of the midbrain tegmentum.
As to these additional projections of the ventral pallidum, the question arises whether they might not originate selec- tively from the numerous strongly AChE-positive neurons within the VP. These cells differ from the larger remaining population of intrapallidal neurons not only by their high AChE content but also by morphological criteria. The con- trast between the two cell categories is seen clearly in AChE-counterstained sections from brains in which a large number of AChE-negative pallidal neurons were retro- gradely labeled by HRP injection of the subthalamic nu- cleus. In such sections, the AChE-negative labeled cells, in accord with existing descriptions of Golgi impregnated pal- lidal nuerons (Fox et al., '74), appear as medium-sized, triangular or fusiform neurons with long and straight, sparsely ramified dendrites, whereas the strongly AChE- positive cells, which have remained unlabeled, are gener- ally larger with a more variably shaped, often multangular
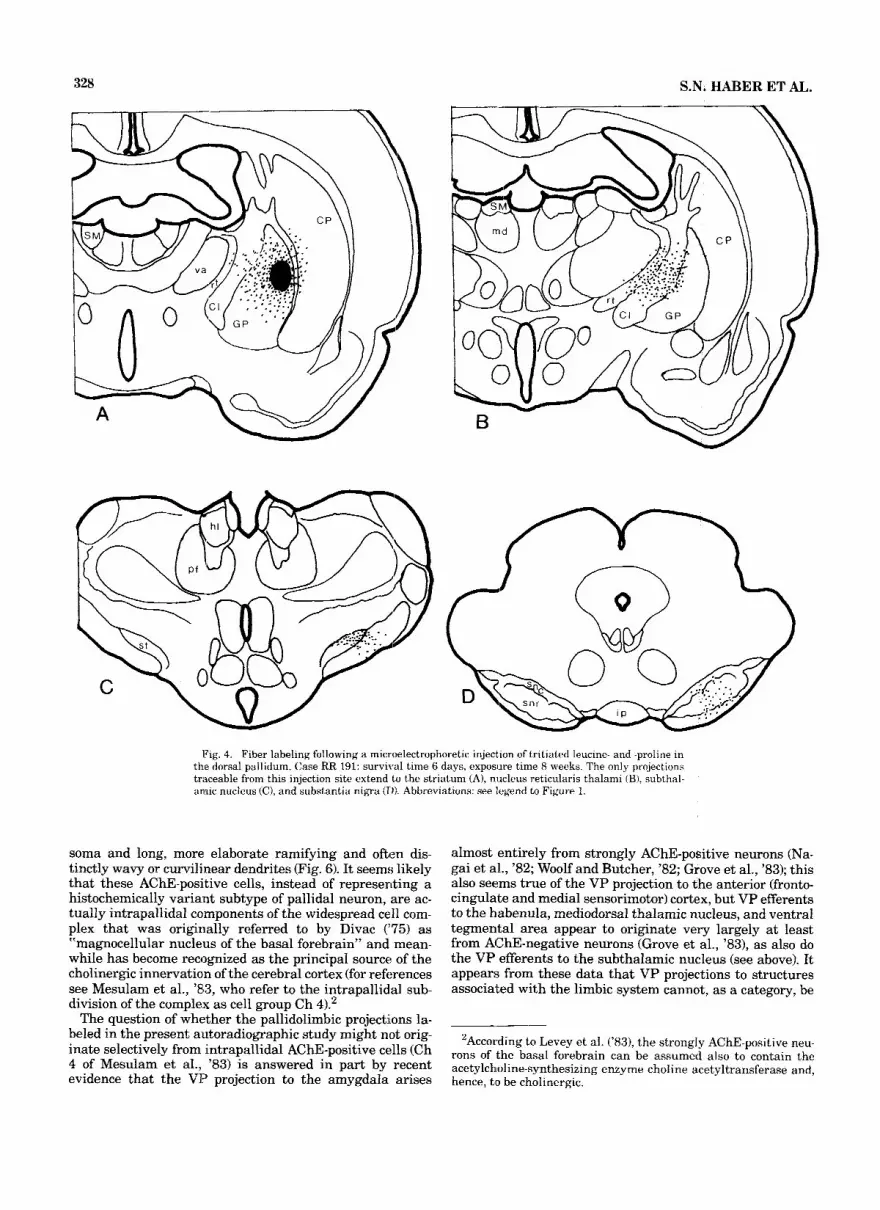
328 S.N. HABER ET AL.
Fig. 4. Fiber labeling following a microelectrophoretic injection of tritiated leucine- and -proline in the dorsal pallidum. Case RR 191: survival time 6 days, exposure time 8 weeks. The only projections traceable from this injection site extend to the striatum (A), nucleus reticularis thalami (B), subthal- annic nucleus (C), and substantia nigra (1)). Abbreviations: see legend to Figure 1.
soma and long, more elaborate ramifying and often dis- tinctly wavy or curvilinear dendrites (Fig. 6). It seems; likely that these AChE-positive cells, instead of representing a histochemically variant subtype of pallidal neuron, are ac- tually intrapallidal components of the widespread cell com- plex that was originally referred to by Divac (‘75) as “magnocellular nucleus of the basal forebrain” and mean- while has become recognized as the principal source of the cholinergic innervation of the cerebral cortex (for reft, brences see Mesulam et al., ’S3, who refer to the intrapallidal sub- division of the complex as cell group Ch 41.’
The question of whether the pallidolimbic projections la- beled in the present autoradiographic study might not orig- inate selectively from intrapallidal AChE-positive cells (Ch 4 of Mesulam et al., ’83) is answered in part by recent evidence that the VP projection to the amygdala arises
almost entirely from strongly AChE-positive neurons (Na- gai et al., ’82; Woolf and Butcher, ’82; Grove et al., ’83); this also seems true of the VP projection to the anterior (fronto- cingulate and medial sensorimotor) cortex, but VP efferents to the habenula, mediodorsal thalamic nucleus, and ventral tegmental area appear to originate very largely at least from AChE-negative neurons (Grove et al., ’831, as also do the VP efferents to the subthalamic nucleus (see above). It appears; from these data that VP projections to structures associated with the limbic system cannot, as a category, be
‘According to Levey et al. (’83), the strongly AChE-positive neu- rons of the basal forebrain can be assumed also to contain the acetylchloline-synthesizing enzyme choline acetyltransferase and, hence, t o be cholinergic.

Fig. 5. Fiber labeling in a case CRR 49; survival time 6 days, exposure time 8 weeks) of microelectrophoretic injection of tritiated leucine- and - proline into the globus pallidus. The injection involved both the medial (substance-P- and AChEcell-rich) pallidal region along the lateral border of
the internal capsule, and an adjacent region of the dorsal pallidum. Note that the resulting labeling pattern combines aspects of both RR 37 (Fig. 1) and RR 192 (Fig. 4). Abbreviations: see legend to Figure 1.

Figure 6

EFFERENTS OF VENTRAL PALLIDUM 331
attributed to the AChE-positive cell population of VP: while the projections to cortex and amygdala indeed originate very largely from strongly AChE-positive cells, others ap- pear to arise for the most part from AChE-negative VP neurons.
The differential origin of ventral-pallidal efferents gains in importance when the possibility is considered that only the AChE-negative category of pallidal neurons may be innervated by the striatum, while AChE-positive VP cells- presumably extraneous to the pallidum-may receive their input or inputs exclusively from nonextrapyramidal sources. This possibility is by no means negligible even though both the AChE-positive and AChE-negative cells lie embedded in the dense plexus of striatopallidal fibers infil- trating the VP, and even though the only other system of VP afferents known to be of substantial volume originates from the likewise extrapyramidal subthalamic nucleus (Ri- cardo, ’80). It is therefore of interest that two recent studies combining AChE staining with, respectively, enkephalin- like immunoreactivity (Haber, ’84) and Gerfen and Saw- chenko’s (’84) immunohistochemical method for anterogradely transported Phaseolus vulgaris leuco-agglu- tinin (PHA-L) (Grove and Nauta, ’841, have produced light- microscopic evidence of efferents from the limbic-inner- vated striatal region coursing alongside, or forming plex- uses of varicose fibers around, soma and dendrites of individual, strongly AChE-positive, VP cells. These find- ings, while not entirely conclusive in the absence of elec- tron-microscopic verification of synaptic junction, suggest that not only AChE-negative VP neurons, but also strongly AChE-positive cells in dwelling in VP can form links in the exit pathway from the limbic-af€erented striatum.
The outcome of the present experiments indicates that the limbic-versus-nonlimbic dichotomy in the innervation of the striatum reflects itself in the existence of two partly segre- gated striatofugal conduction lines. The present evidence of this continued dualism is schematically illustrated by F i g ure 7, and can be summarized as follows. A dorsolateral sec- tor of the anterior striatum receiving only sparse afferents of limbic origin but instead heavily innervated by the sensori- motor cortex (Kelley et al., ’82) projects to an anterodorsola- teral region of the main, supra- and retrocommissural (i.e., conventionally recognized), mass of the globus pallidus; this pallidal region (“dorsal pallidum”) in turn gives rise to a pro- jection that forms part of the classical pallidofugal pathway descending to the subthalamic nucleus and substantia nigra (Fig. 7B). By contrast, the heavily limbic-innervated anter- oventromedial striatal region (shaded in Fig. 7A) projects to the largely infracommissural, but partly also supra- and re-
Fig. 6. photographs representing two regions of the rostra1 globus palli- dus in a case of WGA-HRP injection of the suhthalamic nucleus, with counterstaining for AChE following Mesulam (’821. Frame A represents the medial, substance-P- and AChE-cell-rich ventromedial pallidal region along- side the internal capsule (CI) (see Fig. 3) and contains five strongly AChE- positive neurons (large arrowheads), only one of which appears in focus and none of which is retrogradely labeled. Frame A also contains nine smaller neurons (small arrowheads), AChE-negative and visible only because of retrograde labeling with HRP. Frame B represents the dorsolateral pallidal region only sparsely supplied with substance-P-positive striatofugal fibers, and labeled dp in Figure 3. This frame contains only a single, large, unla- beled but strongly AChE-positive neuron (arrowhead); all other dark pro- files represent AChE-negative neurons retrogradely labeled with HRP. Scale = 100 urn.
trocommissural ventral pallidum, from which in turn effer- ents pass not only to the subthalamic nucleus and substantia nigra but also to various limbic-system-associated struc- tures, in particular the amygdala, frontocingulate (and ad- joining sensorimotor) cortex, mediodorsal thalamic nucleus, lateral habenular nucleus, hypothalamus, ventral tegmen- tal area, and more caudal and dorsal regions of the midbrain tegmentum (Fig. 7C).
As thus defined, the two striatopallidofugal lines share the subthalamic nucleus and substantia nigra as common distribution targets. If it is assumed that both these target structures first and foremost are components of the somatic motor system, the present findings suggest both as sites where central skeletomuscular mechanisms are accessible to striatopallidal output of limbic antecedents, conveyed by the line leading from the limbic-afferented ventromedial striatum over the ventral pallidum. It is possible that fur- ther sites of such access lie farther caudally in the mid- brain. In the present experiments, isotope injections in either the infracommissural (RR 37) or retro- and supra- commissural VP region (RR 49) were found to label rather sparse fibers distributed over wide regions of the midbrain tegmentum including the dorsolateral area containing the nucleus tegmenti pedunculopontinus. It is of interest to note that fibers of very similar tegmental distribution have been traced from the nucleus accumbens in both the rat (Nauta et al., ’78) and cat (Groenewegen and Russchen, ’841, and it would thus seem that the system in question contains striatal as well as pallidal efferents. The available autoradiographic material provided no evidence that either of these fiber categories actually terminates in the region of the pedunculopontine nucleus (rather than merely pass- ing through it to the adjacent central gray substance), but recent studies with the aid of the anterograde marker PHA- L have shown that some efferents from the region here called ventral pallidum indeed terminate in and about the pedunculopontine nucleus (Swanson et al., ’84; Groenewe- gen and van Dijk, ’84). Swanson et al. (’84) interpret this as suggesting a limbic innervation of the “mesencephalic lo- comotor region” of Grillner and Shik (‘73).
A third route by which the limbic system could be thought to have access to central somatic-motor mechanisms could lead from the limbic-innervated striatum over the strongly AChE-positive cells embedded in the ventral pallidum. The present anterograde evidence suggesting that at least some of these cells project to the sensorimotor cortex only con- firms earlier findings in retrograde-labeling experiments (McKinney et al., ’83; Saper, ’84).
Even more difficult to interpret from a functional point of view are the limbicostriatopallidofugal lines leading to var- ious limbic and limbic-system-associated structures, sum- marized in Figure 7C. From one vantage point, these connections could be viewed collectively as a differentiated “return loop” enabling the limbic system to monitor the effect of its outputs to the striatum (and thus possibly act- ing as an aid in adjusting the organism’s motivational set to the output efficacy of the central somatic-motor system). But other possibilities must be considered. The question arises, for example, if significance of the much-interrupted limbicostriatopallidolimbic circuit here proposed may not lie in the fact that the striatum is the site of an unusually wide variety of intricately interspersed neurotransmitters and neuromodulators (cf. Graybiel and Ragsdale, ’83), par- ticular combinations of which might be required for optimal functioning of limbic and cortical mechanisms no less strin-

A
B
Fig. 7. Semischematic illustration of the two partly segregated striato- fugal conduction lines described in the text. A. “Limbic” (shadc?d) and “nonlimbic” striatal subdivi:sions and their respective main telencephalic afferents. B. The conventionally described striatofugal line leading from the motor-cortex-innervated “nonlimbic” striatal subdivision to the dorsal pallidum (heavy arrow) which in turn projects to the subthalamic nucleus
and substaiitia nigra. C. The line leading from the limbic-aflerented striatal subdivision, to the ventral pallidum (heavy arrow) and from there not only t o subthalamic nucleus and substantia nigra but also to the frontocingulate and medial sensorimotor cortex, lateral habenuiar and mediodorsal tha- lamic nucleus, amygdala, hypothalamus, ventral teynental area, and more caudal and dorsal tegmental regions. Abbreviations: see legend to Figure 1.

EFFERENTS OF VENTRAL PALLIDUM 333
Fig. 8. The extrapyramidal circuit of the motor cortex (A) compared with analogous trans~striatal circuits involving the amygdala and frontocingulate cortex (B). To avoid excesswe complexity of‘ the diagrams no attempt has been made to include possible cross~links between A and B, such as the one suggested by the reciprocal connection of the ventral pallidum with the subthalamic nucleus. Ahbre viations: see legend to Figure 1.
gently than the same or other combinations are required for optimal somatic-motor function. The possibility of such a more general functional role of the corpus striatum sug- gests itself from a remarkable parallel between the “lim- bic” and “nonlimbic” trans-striatal conduction lines: both prominently include channels over which part of their im- pulse flow can be led back to the domain of its origin in the cerebral hemisphere. To be more explicit: the familiar “ex- trapyramidal motor circuit,” motor cortex + striatum -+
pallidum + thalamic VA-VL complex -+ premotor cortex -+
motor cortex (Fig. 8A) appears to have limbic counterparts
in the sequences: limbic system -+ striatum -+ ventral pallidum -+ amygdala, and: limbic system -+ striatum -+
ventral pallidum -+ thalamic mediodorsal nucleus -, fron- tal and anterior limbic cortex (Fig. 8B). If it is considered likely on the basis of long-known anatomical connections that the function of the striatum expresses itself in part through a major circuit affecting the mechanisms of the motor cortex, the more recent anatomical data here re- viewed make it appear no less likely that the striatum, through an analogous though less massive circuit, can af- fect the mechanisms of the prefrontal cortex, and thus, a

334 S.N. HABER ET AL.
class of functions more likely to be cognitive than skeletomuscular.
ACKNOWLEDGMENTS This study was made possible by PHS grant 5:R23NS
20467 to S.N.H., NSF grant BNS-8306284 and PHS grants 1 R 0 1 NS 19945 and PO1 MH 31154 to WJ.H.N., and a grant from the Dutch Organization for the Advancement of Basic Research (ZWO) to H.J.G. E.A.G. was supported by a stipend from training grant NRSA 5 T32 GM 07484. It is a pleasure to acknowledge the superb assistance received from Diane Major, Rigmor Clark, and Peter Paskevich.
LITElRATURE CITED Beckstead, R.N. (1983) A pallidostriatal projection in the cat and monkey.
Brain Res. Bull. 11:629-632. Butcher, L.L. (1983) Acetylcholinesterase histochemistry. In A. Bdorklund
and T. Hokfelt (eds): Handbook of Chemical Neuroanatomy. Vol. I: Meth- ods in Chemical Neuroanatomy. Oxford: Elsevier Science Publishers B.V., pp. 1-49.
Dahlstrom, A,, and K. Fuxe ‘(1964) Evidence for the existence of morioamine- containing neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brainstem neurons. Acta Physiol. Scand. 62 (Suppl. 232):l--55.
Divac, I. (1975) Magnocellular nuclei of the basal forebrain project to neo- cortex, brain stem, and (olfactory bulb. Review of some functional corre- lates. Brain Res. 93:385-398.
Fallon, J.H., and C.E. Ribtik (1980) Multiple neurotransmitter szudies in the islands of Calleja complex of the basal forebraln. 111. Connections, correlations and reservakions. Sac. Neurosci. Abstr. 6:114.
Foix, C., and J. Nicolesco (1925) Anatomie Cerehrale. Les Noy.~ux Gris Centraux et la Region Mesencephalo-Sous-Optique. Paris: Masson.
Fox, C.A., H.N. Andrade, I. LuQui, and J.A. Rafols (1974) The primate globus pallidus. A Golgi and electron microscope study. J. Hirnforsch. 15:75-93.
Gerfen, C.R., and P.E. Sawrchenko (1984) An anterograde neuroariatomical tracing method that shows the detailed morphology of neurons, their axons and terminals: Immunohistochemical localization of an axonally transported plant lectin, Phaseolus vulgaris leucoagglutinin (PHA-L). Brain Res. 290219-238.
Goldschmidt, R.B., and L. Heimer (1980) The rat olfactory tubercle: Its connections and relation to the strio-pallidal system. Soc. Neuroscl. Abstr. 6271.
Graybiel, A.M., and C.W. Ragsdale (1983) Biochemical anatomy of the striatum. In P.C. Emson (ed): Chemical Neuroanatomy. New York: Raven Press, pp. 427-504.
Grillner, S., and M.T. Shik (1973) On the descending control of lurnbosacral spinal cord from the “mesencephalic locomotor region”. Acta Physiol. Scand. 87320-333.
Groenewegen, H.J., and C.A. van Dijk (1984) Efferent projecticns of the ventral pallidum in the rat as studied with the anterograde trmsport of Phaseolus vulgaris leucoagglutinin (PHA-L). Eur Soc. Neurosci. (in the press).
Groenewegen, H.J., and F.T. Russchen (1984) Organization of the efferent projections of the nuc‘leus accumbens to pallidal, hypothalamic, and mesencephalic structures: A tracing and immunohistochemical study in the cat. J. Comp. Neurirl. 223:347-367.
Grove, E.A., S.N. Haber, V.B. Domesick, and W.J.H Nauta (19831 Differen- tial projections from fiChE-positive and AChE-negative ventral-palli- dum cells in the rat. Soc. Neurosci. Abstr. 9:16.
Grove, E.A., and W.J.H. Nauta (1984) Light microscopic evidence for striatal and amygdaloid input to cholinergic cell group Ch 4 in the: rat. Soc. Neurosci. Abstr. 10:7.
Haber, S.N. (1984) Striatal Input to a basal forebrain AChE-positive cell group sets it apart from the rest of the forebrain cholinergic neurons. Soc. Neurosci. Abstr. 10:7.
Haber, S.N., and W.J.H. Nauta (1981) Substance P, but not enkephalin, immunoreactivity distiinguishes ventral from dorsal pallidum. Soc. Neu- rosci. Abstr. 7916.
Haber, S.N., H.J. Groeneviegen, and W.J.H. Nauta (1982) Efferent connec- tions of the ventral palllidurn in the rat. Soc. Neurosci. Abstr. 8:169.
Haber, S.IV., and W.J.H. Nauta (1983) Ramifications of the globus pallidus in the rat as indicated by patterns of immunohistochemistry Neurosci- ence 9245-260.
Heimer, I,. (1978) The olfactory cortex and the ventral striatum. In K.E. Livingston and 0. Hornykiewicz (eds): Limbic Mechanisms, the Contin- uing Evolution of the Limbic System Concept. New York: Plenum Press, pp. 95-187.
Heimer, I,., and R.D. Wilson (1975) The subcortical projections of the allo- cortex: Similarities in the neural associations of the hippocampus, the piriform cortex, and the neocortex. In M. Santini (ed): Golgi Centennial Symposium. New York: Raven Press, pp. 177-193.
Hill, J.M., and R.C. Switzer (1984) The regional distribution and cellular localization of iron in the rat brain. Neuroscience 11:595-603.
Jacobowibz, D.M. and M. Palkovits (1974) Topographic atlas of catechol- aminri and acetylcholinesterase-containing neurons in the rat brain. I. Forebrain (telencephalon, diencephalon). J. Comp. Neurol. 15713-28.
Jayaraman, A. (1983) Topographic organization and morphology of peripal- lidal and pallidal cells projecting to the striatum in cats. Brain Res. 275279-286.
Kelley, A.E., V.B. Domesick, and W.J.H. Nauta (1982) The amygdalostriatal projection in the rat-an anatomical study by anterograde and retro- grade tracing methods. Neuroscience 7:615-630.
Levey, A.I., B.H. Wainer, E.J. Mufson, and M.-M. Mesulam (19831 Co- localization of acetylcholinesterase and choline acetyltransferase in the rat cerebrum. Neuroscience 9:9-22.
McKinney, M., J.T. Coyle, and J.C. Hedreen (1983) Topographic analysis of the iinnervation of the rat neocortex and hippocampus by the basal forebrain cholinergic system. J. Comp. Neurol. 217103-121.
Mesulam, M-M. (1978) Tetramethylbenzidine for horseradish peroxidase neurochemistry: A non-carcinogenic blue reaction product with superior sensitivity for visualizing neural afferents and efferents. J. Histochem. Cytochem. 26: 106-117.
Mesulam, M-M. (1982) Simultaneous demonstration of AChE and HRP. In M-M. Mesulam (ed): Tracing Neural Connections with Horseradish Per- oxidase. Chichester, John Wiley & Sons, pp. 132-134.
Mesulam, M-M., E.J. Mufson, B.H. Wainer, and A.I. Levey (1983) Central cholinergic pathways in the rat: An overview based on an alternative nomenclature (Ch 1-Ch 6). Neuroscience IO:ll85-1201.
Mogenson, G.J., L.W. Swanson, and M. Wu (1983) Neural projections from nucleus accumbens to globus pallidus, substantia innominata and lat- eral preoptlc-lateral hypothalamic area: An anatomical and electrophys- iological investigation in the rat. J. Neurosci. 3:189-202.
Nagai, T., H. Kimura, T. Maeda, P.L. McGeer, F. Peng, and E.G. McGeer (198?!) Cholinergic projections from the basal forebrain of rat to the amygdala. J. Neurosci. 2513-520.
Nauta, 14.J.W. (1979) Projections of the pallidal complex: An autoradi- ographic study in the cat. Neuroscience 4:1853--1873.
Nauta, W.J.H., G.P. Smith, R.L.M. Faull, and V.B. Domesick (1978) Efferent connections and nigral afferents of the nucleus accumbens septi in the rat. Neuroscience 3385-401.
Otterson, O.P. (1980) Afferent connections to the amygdaloid complex of the rat and cat: 11. Afferents from the hypothalamus and the basal telence- phalon. J. Comp. Neurol. 194267-289,
Ribak, C.E., and J.H. Fallon (1983) The islands of Calleja complex of rat basal forebrain. I . Light and electron microscopic observations. J. Comp. Neu1-ol.205:207-218.
Ricardo, J. (1980) Efferent connections of the subthalamic region in the rat. I. The subthaiamic nucleus of Luys. Brain Res. 202257-271.
Saper, C.B. (1984) Organization of cerebral cortical afferent systems in the rat. [I. Magnocellular basal nucleus. J. Comp. Neurol. 222:313-342.
Staines. W.A., S. Atmadja, and H.C. Fibiger (1981) Demonstration of a pallido-striatal pathway by retrograde transport of HRP-labelled lectin. Brain Res. 206446-450.
Swanson, L.W. (1976) An autoradiographic study of the efferent connections of the preoptic region in the rat. J. Comp. Neurol. 167227-256.
Swanson, L.W., and W.M. Cowan (1975) A note on the connections and development of the nucleus accumbens. Brain Res. 29:324-330.
Swanson, L.W., G.J. Mogenson, C.R. Gerfen, and P. Robinson (1984) Evi- dence for a projection from the lateral preoptic area and substantia innominata to the ‘mesencephalic locomotor region’ in the rat. Brain Res. 295:161-178.
Switzer, R.C., and J. Hill (1979) Globus pallidus component in the olfactory tubercle: Evidence based on iron distribution. Soc. Neurosci. Abstr. 579.

EFFERENTS OF VENTRAL PALLIDUM 335
Switzer, R.C., J. Hill, and L. Heimer (1982) The globus pallidus and its rostroventral extension into the olfactory tubercle of the rat: A cyto- and chemoarchitectural study. Neuroscience 71891-1904.
Velayos, J.L., A. Llamas, and F. Reinoso-Suarez (1980) Prosencephalic con- nections of the mediodorsal nucleus of the thalamus. Xlth Int. Congr. Anat., Mexico City, p. 148.
Williams, D.J., A.R. Crossman, and P. Slater 11977) The efferent projections of the nucleus accumbens in the rat. Brain Res. 130217-227.
Wilson, R.D. (1972) Efferent Connections of the Nucleus Accurnbens in the Rat. Master’s Thesis, Massachusetts Institute of Technology.
Woolf, N.J., and L.L. Butcher 11982) Cholinergic projections to the basola- teral amygdala: A combined Evans Blue and acetylcholinesterase anal- ysis. Brain Res. Bull. 8351-763.
Young, W.S., G.F. Alheid, and L. Heimer (1984) The ventral pallidal projec- tion to the mediodorsal thalamus: A study with fluorescent retropade tracers and immunohistofluorescence. J. Neurosci. 4: 1626-1638.
Copyright © 2022 FDOKUMEN




















