
return on investment: a multisite case study of cost-effective, high
Upload
independentCategory
view
1download
0
1042 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 6, JUNE 2007
Performance of Multisite Silicon MicroprobesImplanted Chronically in the Ventral
Cochlear Nucleus of the CatDouglas McCreery*, Albert Lossinsky, and Victor Pikov
Abstract—A central auditory prosthesis based on microstimula-tion within the ventral cochlear nucleus (VCN) offers a means ofrestoring hearing to persons whose auditory nerve has been de-stroyed bilaterally and cannot benefit from cochlear implants.
Arrays of silicon probes with 16 stimulating sites were implantedinto the VCN of adult cats, for up to 314 days. Compound neu-ronal responses evoked from the sites in the VCN were recordedperiodically in the central nucleus of the contralateral inferior col-liculus (ICC). The threshold and growth of most of the responseswere stable for at least 250 days after implantation of the arrays.The responses evoked from the deepest and shallowest electrodesites did exhibit some changes over time but none of the thresh-olds exceeded 10 A. The thresholds and growth of the compoundresponses from most of the stimulating sites were very stable overtime, and comparable to those of chronically implanted single-siteiridium microelectrodes. Multiunit neuronal activity evoked fromthe stimulating sites in the VCN was recorded along the dorsolat-eral-ventromedial (DLVM) axis of the ICC. The distribution, spanand degree of overlap of the multiunit activity demonstrated theutility of the multisite, multishank array configuration as a meansof accessing the neuronal populations in the VCN that encode var-ious acoustic frequencies. These findings are encouraging for theprospects of developing an auditory prosthesis employing multi-site silicon microprobes.
Index Terms—Auditory prostheses, cats, cochlear nucleus, mi-crostimulation, neural prostheses, silicon microprobes.
I. INTRODUCTION
COCHLEAR implants have restored hearing to tens ofthousands of persons with severe hearing loss, but pa-
tients who lack a functional auditory nerve cannot benefit fromthese devices. However, a prosthesis utilizing an electrodearray implanted in, or adjacent to, the cochlear nucleus (CN)can restore some hearing to these patients. Worldwide, severalhundred patients afflicted with Type II Neurofibromatosis(NF2) have received these auditory brainstem Implants. NF2occurs in about one in 40 000 live births, and these personstypically develop life-threatening vestibular schwannomas onthe 8th nerve, with high probability that the tumors will occurbilaterally [1], [2] The usual outcome is total destruction of the
Manuscript received May 11, 2006; revised November 15, 2006. This workwas supported in part by the National Institutes of Health (NIDCD) under Con-tract NO1-DC-4-0005. Asterisk indicates corresponding author.
*D. McCreery is with the Huntington Medical Research Institutes, NeuralEngineering Program, 734 Fairmount Ave., Pasadena, CA 91105 USA (e-mail:[email protected]).
A. Lossinsky and V. Pikov are with the Huntington Medical Research Insti-tutes, Neural Engineering Program, Pasadena, CA 91105 USA
Digital Object Identifier 10.1109/TBME.2007.891167
auditory and vestibular components of both 8th nerves. Afterthe tumor has been removed surgically, the electrode array isplaced within the lateral recess of the 4th ventricle.
Such an array of “macroelectrodes,” residing on the surfaceof the brainstem, can activate neurons throughout much of theunderlying CN. However, the stimulus current from these rela-tively large electrodes must spread quite broadly, providing onlylimited access to the spatial organization of the underlying nu-cleus, including its tonotopic organization [3]. In contrast, pen-etrating microelectrodes can deliver localized stimulation andin principle should allow for more precise and orderly accessto the tonotopic organization of the CN. Thus, a neural pros-thesis that employs microelectrodes, or a mixture of micro- andmacro-electrodes, might provide these patients with improvedhearing.
The feasibility of microstimulation within the CN has beeninvestigated in animal models in several laboratories [4]–[9],and a clinical trial of a microstimulating array employing 8 or 10single-site iridium microelectrodes is now in progress [10]. Animplant utilizing multiple silicon-substrate shanks, each withmultiple independent stimulating sites, should further improvethe devices’ functionality by allowing highly localized stimu-lation of many separate neuronal populations within the VCN.One attribute of such localized stimulation is the ability to ac-cess the tonotopic organization within each of the subdivisionsof the CN. Evans et al. [4] showed that microstimulation de-livered through adjacent sites on silicon probes implanted in theCN of guinea pigs produced systematic differences in the middlelatency evoked response, suggesting that activation of subpopu-lations of auditory brainstem neurons with multichannel siliconpenetrating microelectrodes is possible.
In all species in which it has been investigated, including thedomestic cat, a cochleotopic (tonotopic) sequence of termina-tions of auditory nerve axons in the ventral cochlear nucleus(VCN) has been demonstrated [11]–[16]. The terminals of theauditory nerve axons from a particular location along the basilarmembrane of the cochlea ramify into thin, parallel laminae thatspan the length and breadth of the anteroventral and postven-tral CN (PVCN); the “frequency-band” or “isofrequency” lam-inae. In an earlier study [7] we applied microstimulation at var-ious locations along the dorsal-ventral axis of the feline PVCNwhile recording multiunit activity (MUA) and local field poten-tials at 20 or more locations along the tonotopic axis of the cen-tral nucleus of the contralateral inferior colliculus (ICC). Thestudy demonstrated that microstimulation within the PVCN canaccess the tonotopic organization of the nucleus in an orderlymanner. This can be attributed not only to the highly localizednature of the microstimulation but also to the trajectory of the
0018-9391/$25.00 © 2007 IEEE

MCCREERY et al.: PERFORMANCE OF MULTISITE SILICON MICROPROBES IMPLANTED CHRONICALLY IN THE VCN OF THE CAT 1043
axons of the efferent projections from the feline CN. In the cat,the efferent axons traverse medially and slightly ventrally acrossthe nucleus, approximately parallel to the isofrequency laminae,as defined by the terminations of the auditory nerve axons, andthe same arrangement appears to be true in humans [17], [18].Were this not the case, each microelectrode would tend to ac-tivate axons from multiple isofrequency laminae, rendering itmore difficult to access the tonotopic organization of the VCNin an orderly manner.
Stimulating sites on the silicon probes are distributed alongthe probe shanks rather than at their tips as is the case for single-site iridium microelectrodes. Thus, while the stimulating siteson the single-site microelectrode are positioned above undis-turbed neural tissue, the sites on the silicon probes are adja-cent to tissue that has been disrupted during probe insertion.This insertion damage and the subsequent tissue response to thepresence of the shank induces neuronal loss [19], and promotesformation of a sheath composed of reactive astrocytes and mi-croglia [20], [21]. In the rat, the tissue response continues toevolve for up to 6–12 weeks after implantation [21]. The chroniccomponent of this tissue response has been implicated in theprogressive degradation of the quality of single-unit recordingsby such probes in the cerebral cortex, and it is unclear how thistissue response would affect the ability of a central auditoryprosthesis to activate neurons at low threshold and its ability tointroduce multiple channels of spectral information (tonotopicspecificity). In the present study, we investigated these issueswith arrays of multisite silicon substrate microprobes implantedchronically (195–314 days) in the feline VCN.
II. METHODS
A. Fabrication of the Microstimulating Array
Fig. 1(A) shows a multisite silicon substrate probe with 2shanks and 8 stimulating sites sputter-coated with iridium oxide.The probes were fabricated at the University of Michigan underthe direction of Design Engineer Jamille Hetke, and provided tous through the Center for Neural Communications Technology.The 4 electrode sites on each shank are 0.8–1.7 mm below theprobe’s transverse spine. After bonding of the Parylene-coatedgold lead wires, the transverse spines of two probes were en-capsulated in EpoTek 301 epoxy to form the button superstruc-ture, 2.5 mm in diameter. The 8 or 16 lead wires were woundinto a helical cable and encapsulated in silicone elastomer. Acomplete array of 2 probes (4 shanks and 16 electrode sites) ex-tending from the epoxy superstructure is shown in Fig. 1(B).
B. Array Implantation
Four young adult cats were used. The cat’s head was fixed ina stereotaxic frame and a bipolar recording electrode was intro-duced through a small craniectomy into the rostral pole of theICC. The recording electrode was a pair of stainless steel wires,100 in diameter, insulated with Parylene- C. Approximately1 mm of the Parylene insulation was removed from the tip re-gions. The recording member of the bipolar pair was insertedapproximately 5 mm ventral of the dorsal surface of the ICC,and the reference member was 10 mm dorsal of the recordingelectrode. A craniectomy was made over the contralateral cere-bellum and part of the lateral cerebellum was aspirated. Thearray of stimulating microelectrodes was secured against the
Fig. 1. (A) A 2-shank silicon substrate probe with 8 iridium stimulating sites.(B) A CN microstimulating array containing 2 of the silicon probes extendingfrom an epoxy button superstructure.
orifice of a vacuum wand, and advanced into the CN at approx-imately 45 from the vertical; approximately the angle at whicha microelectrode array would be inserted into the human CN,using the translabyrinthine approach to brainstem. The arraycable was fixed to the bone at the margin of the craniectomyand the cavity was filled with gelfoam.
C. Recording of Compound Evoked Responses
Periodically, the responses evoked from each of the micro-electrode sites in the CN were recorded via the electrode in therostral pole of the contralateral ICC. The cat was lightly anes-thetized with Propofol and the recordings were conducted insidea double-wall sound isolation booth (Audiometrics 120A-SP).The stimulus was cathodic-first, charge-balanced pulse pairs,each phase 150 in duration, ranging from 0 to 35 . 512to 2048 successive responses were averaged to obtain each aver-aged evoked compound response (AECR); an example is shownat the bottom of Fig. 3. The response growth functions (RGFs),which represent the recruitment of the neurons surrounding eachmicroelectrode in the VCN, were generated by plotting the am-plitude of the first component of each of the AECRs against theamplitude of the stimulus. The latency to the onset of the firstcomponent of the AECR is approximately 1 ms and, thus, wecan assume that it represents the activity of neurons excited bythe microstimulation in the VCN and projecting directly to thecontralateral ICC.
D. Multisite Recording in the ICC
This experiment was conduced just before the cats were sac-rificed. They were anesthetized with Isoflurane and oxygen, andtheir heads fixed in a stereotaxic frame. To facilitate delivery ofacoustic stimuli, a hollow earbar was used contralateral to theICC from which recordings were made. The ipsilateral earbarwas solid, in order to attenuate acoustic stimuli. A wide craniec-tomy was made over one cerebral hemisphere, and the occip-ital pole of the cerebrum was removed to expose the dorsal sur-face of the ICC. Throughout the experiment, respiration rate andend-tidal were monitored continuously. Core body temper-ature was maintained at 37–39 . All sound-generating equip-ment was outside of the sound isolation room.
The tonotopic organization of the anteroventral and PVCNis preserved in the projection from the CN to the ICC, wherethe iso-frequency laminae are oriented approximately perpen-dicular to its dorsolateral-ventromedial (DLVM) axis, with lowacoustic frequencies represented dorsally and laterally and high

1044 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 6, JUNE 2007
frequencies represented ventrally and medially [22]–[25]. Mul-tiunit neuronal activity was recorded at 16 locations separatedby 200 and spanning 3 mm along the DLVM axis of ICC,using a multisite probe fabricated by NeuroNexus, Inc. For eachinsertion of the recording probe, tone bursts ranging from 0.5to 25 kHz, and 100 ms in duration with a rise time of 10 ms,were delivered through a broad-band loudspeaker ( 5 db,200 Hz to 35 kHz). When a large, well-isolated extracellularaction potential was recorded at one of the sites in the ICC,its best frequency near threshold was determined by successivesmall adjustments of the acoustic frequency and reductions inthe stimulus intensity.
After the acoustic mapping, 1500 controlled-current, biphasicstimulus pulses (150 in duration at 50 pps) were appliedto each of the microelectrode sites in the CN. The stimuluscurrent was 10, 20, or 30 (1.5, 3, or 4.5 nC/phase). In thehuman patients who have been implanted with arrays of single-site iridium microstimulating electrodes in their cochlear nuclei,the thresholds for auditory percepts have been 1 to 2 nC/phasefor most of the penetrating microelectrodes [10].
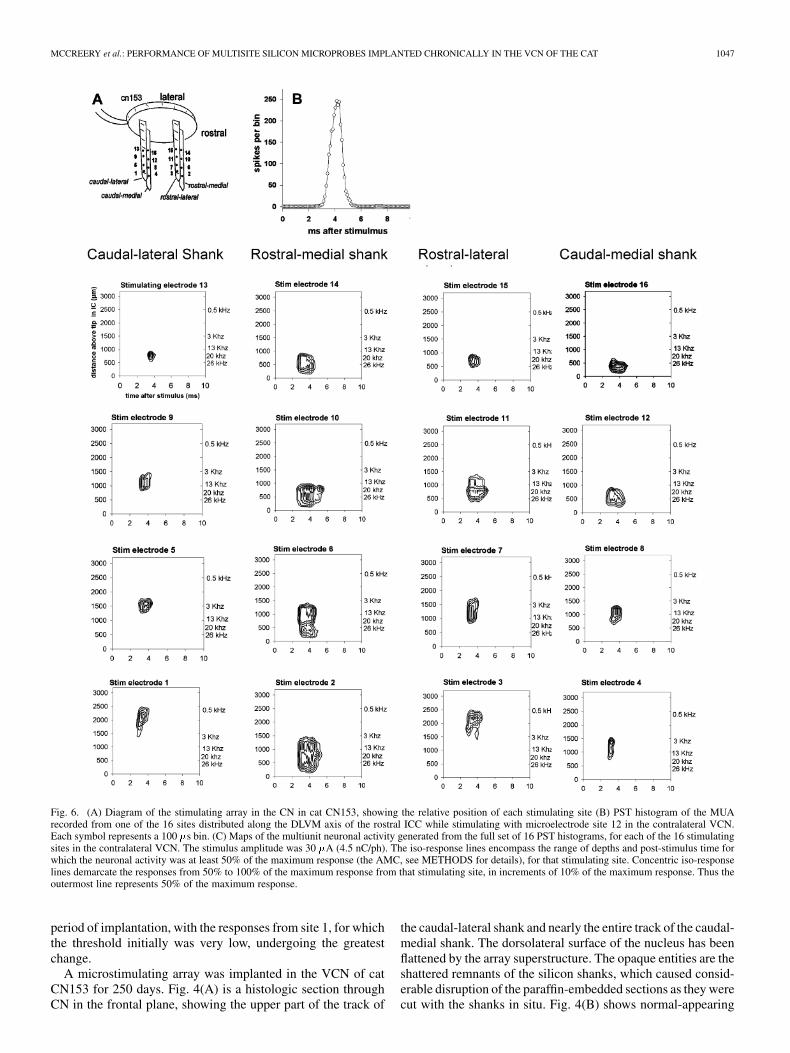
Extracellular action potentials (MUA) evoked by the stimu-lation in the VCN were separated from the large compound re-sponse using a common background suppression technique andbandpass filtering between 500 Hz and 8 kHz. We did not per-form formal point-process cross correlations of the neuronal ac-tivity recorded from adjacent sites in the ICC, but by inspec-tion of the raw traces, the risk of redundantly counting the samespike from adjacent recording sites was greatly reduced by usingevery other recording site, separated by 200 . Thus, the spanof the neural activity recorded along the DLVM axis of the ICCmore accurately reflects the actual neural activity along thataxis, rather than the spatial resolution of the recording array.Spikes were sorted into bins 100 in width and summed for the1500 presentation of the stimulus, yielding 16 poststimulus time(PST) histograms, one for each recording site in the ICC. The16 histograms represent, in PST and depth in the ICC, the MUAevoked in response to the stimulation in the VCN. A samplehistogram is shown in Fig. 6(A). Response maps of the MUAFig. 6(B) were generated from the 16 PST histograms, and thecentroids of the maps were computed as the means, in depth inthe ICC and PST, of the counts of the MUA from the bins com-prising the maps. Computation of the maps’ centroids can beseriously skewed by MUA that is far from the maximum neu-ronal response to the electrical stimulus in the VCN. This back-ground activity is present in most of the bins of the PST his-tograms, but it contributes only a few counts to each histogrambin [see Fig. 6(A)]. Thus, the maps were generated only fromhistograms bins in which the spike count was at least 50% ofthe “averaged maximum count” (AMC). As shown in the PSThistogram in Fig. 6(A), the histogram’s maxima is well definedby the average of the counts in the 4 bins (the 400 ms window)around the peak, so we computed this 4-bin average for each ofthe 16 histograms and selected the largest of these as the AMC.By computing the centroids only from the bins that are fairlyclose to the 4 bins contributing to the AMC, we also reduce theartifact whereby centroids near the upper or lower extremes ofthe recording array are displaced towards the center of the array.
In Fig. 6(B), the 5 iso-response (contour) lines correspond tothe range of bin counts between 50% and 100% of the AMC,in increments of 10% of the AMC. The extent of the 50%-of-
Fig. 2. A histologic section in the frontal plane, through the VCN of cat CN149.Portions of the tracks of the RL and RM silicon shanks are indicated. Cresylviolet (Nissl) stain.
AMC contour line around each centroid was used as the measureof dispersion (span) of the stimulation-induced MUA inducedalong the DLVM axis of the ICC, from that site in the VCN.Since each map is normalized on its own AMC, the computationof the activity span is not affected by the absolute magnitude ofthe neuronal activity evoked from each site in the VCN.
E. Histology
At the conclusion of the study, the cat was transitioned to deeppentobarbital anesthesia and perfused through the aorta withapproximately 100 ml of phosphate-buffered saline followedby 3 L of 4% buffered parformaldehyde. The microstimulatingarray was removed from the CN. During implantation monthsearlier, part of the cerebellum had been removed and the void hadfilled with loose connective tissue. While dissecting the arraycable from this tissue, most of the silicon shanks fractured fromthe epoxy superstructure and remained in the fixed tissue. TheCN was embedded into paraffin and sectioned in the frontal planeat 7 and the section stained with Cresyl violet (Nissl stain).The presence of the probe shanks did cause some disruption ofthe tissue sections, but the anatomical locations of the 4 probesand the condition of the neurons and neuropil adjacent to theprobe tracks still could be determined. The anatomical locationof the electrode tracks in the CN was determined from the dis-tance of the tracks from the caudal and lateral extremes of the CNcomplex and from the cell types surrounding the tracks, usingmaps of the cytoarchitecture of the feline CN complex [25].
III. RESULTS
A microstimulating array was implanted in the CN of catCN149 for 250 days. The array included 8 active electrode siteson the rostral-medial (RM) and rostral-lateral (RL) shanks. Fig. 2shows a histologic section in the frontal plane, close to the tip siteof the RM shank and approximately midway up the RL shank.The array’s epoxy superstructure had somewhat compressed thedorsolateral surface of the CN. The RM shank was in the PVCNand the RM shank was in the anterioventral CN.

MCCREERY et al.: PERFORMANCE OF MULTISITE SILICON MICROPROBES IMPLANTED CHRONICALLY IN THE VCN OF THE CAT 1045
Fig. 3. RGFs of the first component of the AECR for each of the 8 microelec-trode sites on the two rostral shanks of the microstimulating array in the ventralCN of cat CN149. A sample AECR is shown at bottom left, denoting the firstcomponent. The diagram at bottom right illustrates the relative position of themicrostimulating sites in the central CN. The responses were recorded near therostromedial pole of the contralateral ICC at 14, 43 and 237 days after implanta-tion of the stimulating array in the contralateral CN.
Fig. 3 shows the RGFs of the first component of the AECRsevoked from each of the 8 microelectrode sites in the CN, asrecorded near the rostromedial pole of the contralateral ICC.The responses were recorded 14, 43 and 237 days after arrayimplantation. The relative positions of the stimulating sites areshown in the inset at the bottom of the figure. Sites 13 and14 were deepest in the VCN, close to the axons of the trape-zoid body. The thresholds of the RGFs ranged from approx-
Fig. 4. Histologic sections in the frontal plan through the CN of cat CN153.(A) Tracks of the caudal silicon shanks. (B) Neurons near the caudal-lateralshank. (C) Tracks of the rostral shanks (D) Neurons near the RM shank. Cresylviolet stain.
imately 5–10 , and the thresholds and slopes of the RGFsfrom most of the stimulating sites were quite constant over the

1046 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 6, JUNE 2007
Fig. 5. RGFs evoked from each of the 16 electrode sites in the CN of cat CN153, at 22 and 250 days after implantation of the microstimulating array. The inset atthe bottom shows the relative position of each electrode site. Each frame is a plot of the amplitude of the earliest component of the compound response evoked fromthe particular site.
MCCREERY et al.: PERFORMANCE OF MULTISITE SILICON MICROPROBES IMPLANTED CHRONICALLY IN THE VCN OF THE CAT 1047
Fig. 6. (A) Diagram of the stimulating array in the CN in cat CN153, showing the relative position of each stimulating site (B) PST histogram of the MUArecorded from one of the 16 sites distributed along the DLVM axis of the rostral ICC while stimulating with microelectrode site 12 in the contralateral VCN.Each symbol represents a 100 �s bin. (C) Maps of the multiunit neuronal activity generated from the full set of 16 PST histograms, for each of the 16 stimulatingsites in the contralateral VCN. The stimulus amplitude was 30 �A (4.5 nC/ph). The iso-response lines encompass the range of depths and post-stimulus time forwhich the neuronal activity was at least 50% of the maximum response (the AMC, see METHODS for details), for that stimulating site. Concentric iso-responselines demarcate the responses from 50% to 100% of the maximum response from that stimulating site, in increments of 10% of the maximum response. Thus theoutermost line represents 50% of the maximum response.
period of implantation, with the responses from site 1, for whichthe threshold initially was very low, undergoing the greatestchange.
A microstimulating array was implanted in the VCN of catCN153 for 250 days. Fig. 4(A) is a histologic section throughCN in the frontal plane, showing the upper part of the track of
the caudal-lateral shank and nearly the entire track of the caudal-medial shank. The dorsolateral surface of the nucleus has beenflattened by the array superstructure. The opaque entities are theshattered remnants of the silicon shanks, which caused consid-erable disruption of the paraffin-embedded sections as they werecut with the shanks in situ. Fig. 4(B) shows normal-appearing

1048 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 6, JUNE 2007
Fig. 7. (A) Locations of the centroids of the multiunit neuronal activity induced from microstimulating sites on each silicon shank in the CN of cat CN153, asplotted in Fig. 6. The ordinate is distance along the DLVM axis of the ICC, above the deepest recording site in the ICC. The bar about each centroid represents thespan of the 50% iso-response lines along the DLVM axis. (B) The centroids ranked according to their depth in the ICC, and with the corresponding span of theMUA, showing generally good coverage of the tonotopic gradient, but with a gap in the low-frequency region.
neurons and neuropil within 100 of the caudal-lateral shank.Fig. 4(C) shows the tracks of the RM and RL shanks, 1.5 mmrostral of the caudal shanks, entering the ventral CN, and caudalof the entry zone of the auditory nerve. The RL shank was in therostral part of the posteroventral nucleus, while the RM shankwas in the caudal part of the anteroventral CN. Fig. 4(D) showsnormal-appearing neurons within 100 of the track of the RLshank.
Fig. 5 shows the RGFs recorded from the same cat, CN153,at 22 and 250 days after implantation of the microstimulatingarray. Between the 22nd and 250th day, the thresholds andslopes of the RGFs were very constant when evoked frommicrostimulating sites midway up each shank (Sites 5–12),while those from the shallowest and deepest sites showedmore change. The thresholds of the RGFs evoked from thedeepest sites decreased, and their slopes increased slightly.The changes in the slope and threshold of the RGFs evokedfrom the shallowest and deepest sites and the stability of theresponses from the sites midway up the shanks suggest someongoing movement of the array through the tissue. At autopsy,the array’s superstructure was seen to have depressed the dorso-lateral surface of the CN by about 0.5 mm [Fig. 4(A) and (C)].This may have been caused by small but continuous downwardforce exerted on the array superstructure by the cable, which inthe cat was routed dorsally.
In cats CN149 and CN153, the thresholds of the RGFs werecalculated by extrapolating the RGFs to the axis. For 24 elec-trode sites in cats CN149 and CN153, the thresholds range from3–15 (mean , range 3–15 ). This is verysimilar to the thresholds of the RGF from 30 single-site iridiummicroelectrodes chronically implanted in the VCN of 6 cats
[8], [26], [27].Fig. 6 shows the response maps of the multiunit neuronal ac-
tivity (spikes per 100 bin) recorded along the DLVM axisin the rostral part of the ICC of cat CN153 while stimulatingwith each of the 16 electrode sites in the contralateral VCN.The stimulus amplitude was 30 (4.5 nC/ph). In these maps,the ordinate is the distance above the deepest recording site inthe ICC (at the tip of the probe), and the abscissa is PST. The
depths of the near-threshold responses to the acoustic tone burstsof different frequencies are indicated near the right axis. Theiso-response lines in each map encompass the range of depthsin the ICC and PST for which the neuronal activity was at least50% of the maximum (AMC) for that stimulating site.
Fig. 7 shows the location in the ICC of the centroids ofmultiunit neuronal activity depicted in Fig. 6. The bars abouteach centroid represent the span of the 50%-of-maximumiso-response line about the centroid. Collectively, the 16 stim-ulating sites in the CN spanned acoustic frequencies fromapproximately 1 kHz to 25 kHz. Fig. 7(B) shows the centroidsranked according to their relative depth in the ICC. There wasgenerally good coverage of the full range of acoustic frequen-cies, but with less complete coverage in the low-frequencyregion. With the exception of the RM shank, which was in thecaudal part of the anteroventral CN, the centroids showed anorderly progression of high-to-low frequencies for stimulatingsites progressively deeper and more lateral in the CN, which isconsistent with the topology of the isofrequency laminae of thefeline VCN [15].
Fig. 8 shows the depths and the spans of the activity foci in-duced by stimulating in the CN at three different amplitudes (10,20, and 30 ), for the caudal-medial, caudal-lateral, and RLshanks in the VCN of cat CN153 (Not all stimulating sites inthe CN produced well-defined foci of activity in the ICC whenstimulating at 10 ). The depths of the centroids were es-sentially invariant with stimulus amplitude, but the span of theMUA along the DLVM axis of the ICC increased slightly, froma mean of 416 to 720 .
Silicon substrate microstimulating arrays were implantedchronically into two additional cats (CN152 for 195 days, andCN155 for 314 days). In both animals, some of the stimulatingsites became open-circuit and failed over time. However, asufficient number of electrode sites survived (6 sites in CN152,11 in CN155) to allow verification of the essential findings fromcat CN153, in which all 16 electrode sites were functional after250 days. Fig. 9 shows the locations of the centroids in the ICCand the span of the MUA from these animals. As in CN153,microstimulation in the ventral CN induced foci of neuronal

MCCREERY et al.: PERFORMANCE OF MULTISITE SILICON MICROPROBES IMPLANTED CHRONICALLY IN THE VCN OF THE CAT 1049
Fig. 8. Locations of the centroids and span of the MUA induced by stimulating at the various sites in the VCN of cat CN153, at three different stimulus amplitudes(10, 20 and 30 �A).
activity in the ICC spanning a wide range of acoustic frequen-cies. With a stimulus amplitude of 30 (4.5 nC/phase), theactivity foci evoked from adjacent sites on a particular shankdid not overlap completely.
IV. DISCUSSION
Since the data reported here were obtained from only 4 cats,and the most complete data set is from a single animal (CN153),it is unclear how some of the quantitative aspects of this studywould generalize to a larger sample, with various placementsof the stimulating array in the CN and of the recording arrayin the ICC. However, the findings do demonstrate a proof ofconcept for the utility of the 3-D microstimulating array basedon multisite silicon probes, as a central auditory prosthesis.
The findings from this study are encouraging for the prospectsof developing a CN auditory prosthesis based on multisite sil-icon microprobes. If the implantation and/or the long-term res-idence of penetrating microelectrodes were to cause a loss ofexcitable neural elements near the electrode sites, the ability toproduce localized excitation of neurons over a large dynamicrange would be compromised. In this study, the thresholds andslopes of most of the RGFs evoked by the intranuclear micros-timulation were stable for at least 250 days after implantation ofthe arrays. The RGFs evoked from the deepest and shallowestelectrode sites did exhibit some changes over time but none ofthe thresholds exceeded 10 . This implies a usable dynamic
range of at least 20 (4.5 nC/phase) between threshold and30 , at which amplitude there remained some separation ofthe responses induced along the tonotopic gradient of the ICC(Figs. 7–9). The changes in the slope and threshold of the RGFsfor the deepest and shallowest electrode sites in the CN mayhave been due to depression of the dorsal surface of the cat’sCN by the underside of the array superstructure, accompaniedby slow movement of the electrode sites through the VCN. Themay have been caused by downward pressure on the array fromthe dorsally routed array cable. As such, it probably would notbe indicative of what would occur in a human patient, in whichthe CN is covered by a thick glial limitans and in which thecable traverses laterally over a portion of the brainstem and thecerebellum [10].
In the mammalian nervous system, the relation between theamplitude, I, of a stimulus pulse that is just sufficient to evoke anaction potential and the distance, d, between the neuron and themicroelectrode is given by , where K ranges from ap-proximately 100–3000 for different types of neurons(assuming a pulse duration of 150–200 [28]. The K-valuesof the neurons in the feline VCN are unknown, but if we assumethat they fall within the range stated above, neurons within ap-proximately 50–200 of the electrode sites would be acti-vated by 6.3 currents pulses, the average threshold for the24 sites from cats CN149 and CN153. This is consistent withthe histologic results which show intact neurons within 100of the silicon shanks (Fig. 2).

1050 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 6, JUNE 2007
Fig. 9. Locations of the centroids of the multiunit neuronal activity along theDLVM axis of the ICC, and the span of the MUA, for cats CN152 and CN155,ranked according to their depth in the ICC. The plots show generally good cov-erage of the tonotopic gradient in the CN, but with less complete coverage of thelow-frequency region. The bars designate the corresponding spans of the MUA.
The chronically implanted multisite microstimulating arrayswere able to activate neurons across essentially the entire tono-topic axis of the feline ventral CN, with good coverage alongthe gradient (Figs. 6, 7, and 9). Also, stimulation at the highestamplitude (30 , 4.5 nC/phase) through sites separated by300–400 on a particular silicon shank activated neuronalpopulations that did not completely overlap. For comparison,patients who have been implanted with arrays of single-siteiridium microelectrodes in the CN report auditory percepts atthresholds of 1–2 nC, and maximum comfortable loudness atabout 4–5 nC/phase [10].
As illustrated in Fig. 8, the depth of the centroid in the ICCvaried little as a function of stimulus amplitude. In cat CN153,the spans of the activity in the ICC was rather weakly dependentupon stimulus amplitude and tended to plateau as stimulus am-plitude increased; the mean of the spans was 460 at 10 ,664 at 20 , and 766 at 30 . The explanation for
this behavior may lie in the current-distance relation for neu-ronal excitation . By this model, the spread of theeffective stimulus current along the tonotopic axis of the VCNis , which predicts the data from cat CN153 quite well( for the first-order regression of the plot of span inthe ICC vs ).
Unlike the domestic cat, the human CN is not a conspicuouspendulous structure on the surface of the brainstem. An advan-tage of a microstimulating array with multiple stimulating siteson each of several shanks is its ability to place many stimulatingsites along the tonotopic axis of the VCN, even when only a fewshanks actually enter the CN, and to do so without increasing thenumber of penetrating shanks and, thus, the risk of damage tothe nucleus and surrounding structures. Figs. 7 and 9 suggestanother advantage; the ability for two or more electrode siteson different shanks to activate neurons at essentially the samepoint on the CN’s tonotopic gradient but at different locationson a particular isofrequency lamina. Thus, interleaved stimu-lation applied at 2 or more such sites may afford a means ofdelivering high-rate stimulation to the same place on the tono-topic gradient, but via different neurons and, thus, without in-creasing the risk of neuronal injury or stimulation-induced de-pression of neuronal excitability, both of which increase whena microelectrode site is pulsed at a high rate [6], [8], [9]. Also,Leake and Snyder [16] showed that spiral ganglia neurons thatlie close to the scala vestibuli project to the medial portion ofeach isofrequency lamina in the CN while the spiral gangliacells closer to the scala tympani project to the lateral portionof the same lamina. It is quite likely that different portions ofeach isofrequency lamina subserve different functions that maybe exploited by a central auditory prosthesis with numerous in-dependent microstimulating sites.
The study also revealed some issues that must be resolvedbefore arrays of silicon probes can be used in a clinical pros-thesis. Figs. 7–9 demonstrate the value of the 3-D coveragewithin the CN that is afforded by the array design depicted inFig. 1(B). However, the relatively bulky epoxy superstructuresupporting the 3-D structure, and also the helical cable undoubt-edly placed stress on the thin (15 ) silicon shanks and inseveral instances, one or more shanks fractured from the su-perstructure during or soon after implantation. Similarly, thearrays must tolerate handling by the surgeon prior to implan-tation. Several of the microstimulating sites became open-cir-cuited during the months in vivo (e.g., cats CN152 and CN155).The likely culprit is the bonded junction between the gold leadwire and the bonding pads on the probe spines. These problemscertainly are surmountable, but they do demonstrate the need forfurther development of the implants and perhaps the need to em-ploy alternative fabrication technologies, such as Bosch-processdeep reactive ion etching, to produce probes that are thicker andstronger.
There was significant overlap of the neuronal activity evokedin the ICC from different microstimulating sites (Figs. 7–9),even at stimulus amplitudes only slightly above threshold(Fig. 8). As a measure of the spread of the induced MUA alongthe DLVM axis of the ICC, we used the span of the multiunitneuronal activity that was of the “averaged max-imum” for that stimulating site in the CN and for that stimulusamplitude. This measure is somewhat arbitrary, but it is largelyimmune to background neural activity that is not modulated by

MCCREERY et al.: PERFORMANCE OF MULTISITE SILICON MICROPROBES IMPLANTED CHRONICALLY IN THE VCN OF THE CAT 1051
the electrical stimulation, and it does convey an appreciation ofthe degree to which the neuronal activity induced from differentsites in the CN overlap. While Fig. 8 indicates that activationof partly separate neuronal populations is possible over a widerange of stimulus amplitudes, the degree of overlap implies alimit to the number of distinct “channels” of information thatcan be introduced into the CN, even with highly localized in-tranuclear microstimulation. However, Figs. 7 and 9 do suggestthat at least 16 channels are possible, perhaps with the additionof additional sites on each shank, or with a slightly differentdistribution of the inter-site spacings to provide more completecoverage of the region of the ventral CN where low acousticfrequencies are represented. The spans of the MUA induced inthe ICC by the chronically implanted multisite silicon probeswere slightly less than the activity spans induced by stimulatingin the VCN in acute experiments, using a single-site iridiummicroelectrode [7], (average span of approximately 850at a stimulus amplitude of 24 with the single-site iridiummicroelectrodes versus 660 or 760 for the silicon probes ata stimulus of 20 and 30 , respectively). The results from thetwo studies may not be directly comparable, since the earlierstudy used a single iridium microelectrode inserted acutelyinto the CN while the present study used chronically implantedsilicon probes. However, this does suggest that the frequencyresolution with the silicon substrate probes is comparable tothat obtainable with single-site iridium microelectrodes.
ACKNOWLEDGMENT
The authors would like to thank C. Graham for preparationof the histologic material, N. Kuleviciute and A. Kowalewskifor fabricating the microelectrode arrays, and L. Bullara forassistance with the design of the electrode arrays. Edna Smithmanaged the animal surgeries. They would like to thankE. Smith and the animal care staff for excellent care of theanimals. The animal studies were approved by the Animal Care& Use Committee of HMRI, and were performed under theguidelines set forth in the Guide to Care and Use of LaboratoryAnimals (1996 edition). The multisite silicon probes werefabricated at the University of Michigan under the direction ofDesign Engineer J. Hetke, and were donated through the Centerfor Neural Communications Technology. Most of the data anddescriptions of the methodologies upon which this paper isbased have appeared in the contract’s quarterly reports
REFERENCES
[1] D. G. Evans, S. M. Huson, D. Donnai, W. Neary, V. Blair, D. Teare, V.Newton, T. Strachan, R. Ramsden, and R. Harris, “A genetic study oftype 2 neurofibromatosis in the United Kingdom. I. Prevalence, muta-tion rate, fitness, and confirmation of maternal transmission effect onseverity,” J. Med. Genet., vol. 29, pp. 841–846, 1992.
[2] D. G. Evans, R. Lye, W. Neary, G. Black, T. Strachan, A. Wallace, andR. T. Ramsden, “Probability of bilateral disease in people presentingwith a unilateral vestibular schwannoma,” J. Neurol. Neurosurg., vol.66, pp. 764–767, 1999.
[3] J. Kuchta, S. R. Otto, R. V. Shannon, W. E. Hitselberger, and D. E.Brackmann, “The multichannel auditory brainstem implant: How manyelectrodes make sense?,” J. Neurosurg., vol. 100, pp. 16–23, 2004.
[4] D. E. Evans, J. K. Niparko, J. M. Miller, R. W. Jyung, and D. J. An-derson, “Multiple-channel stimulation of the cochlear nucleus,” Oto-laryngol. Head Neck Surg., vol. 101, pp. 651–657, 1989.
[5] J. K. Niparko, R. A. Altschuler, D. A. Evans, X. L. Xue, J. Farraye,and D. J. Anderson, “Auditory brainstem prosthesis: Biocompatibilityof stimulation,” Otolaryngol. Head Neck Surg., vol. 101, pp. 344–352,1989.
[6] D. B. McCreery, T. G. Yuen, W. F. Agnew, and L. A. Bullara, “A char-acterization of the effects on neuronal excitability due to prolongedmicrostimulation with chronically implanted microelectrodes,” IEEETrans. Biomed. Eng., vol. 44, no. 10, pp. 931–939, Oct. 1997.
[7] D. B. McCreery, R. V. Shannon, J. K. Moore, and M. Chatterjee, “Ac-cessing the tonotopic organization of the ventral cochlear nucleus byintranuclear microstimulation,” IEEE Trans. Rehabil. Eng., vol. 6, no.4, pp. 391–399, Dec. 1998.
[8] D. B. McCreery, T. G. Yuen, and L. A. Bullara, “Chronic microstimu-lation in the feline ventral cochlear nucleus: Physiologic and histologiceffects,” Hear. Res., vol. 149, pp. 223–238, 2000.
[9] D. B. McCreery, T. G. Yuen, W. F. Agnew, and L. A. Bullara, “Stim-ulus parameters affecting tissue injury during microstimulation in thecochlear nucleus of the cat,” Hear. Res., vol. 77, pp. 105–115, 1994.
[10] D. B. McCreery, A. S. Lossinsky, R. V. Shannon, and O. R. , Sr.,A cochlear Nucleus Auditory Prosthesis Based on Microstimulation(Contract # N01-DC-4-0005 NIDCD, From the National Institutes ofHealth, Bethesda, MD, Quart. Progress Reps. # 3,5,6,7, 2005 and 2006.
[11] T. R. Bourk, J. P. Mielcarz, and B. E. Norris, “Tonotopic organizationof the anteroventral cochlear nucleus of the cat,” Hear Res, vol. 4, pp.215–241, 1981.
[12] D. M. Fekete, E. M. Rouiller, M. C. Liberman, and D. K. Ryugo, “Thecentral projections of intracellularly labeled auditory nerve fibers incats,” J. Comp. Neurol., vol. 229, pp. 432–450, 1984.
[13] T. B. S. Powell and W. M. Cowen, “An experimental study of the pro-jection of the Cochlear.,” J. Anat., vol. 96, pp. 269–284, 1962.
[14] I. Sando, “The anatomical relationships of the cochlear nerve fibers,”Acta Oto-Laryngol., vol. 59, pp. 417–435, 1965.
[15] R. L. Snyder, P. A. Leake, and G. T. Hradek, “Quantitative analysisof spiral ganglion projections to the cat cochlear nucleus,” J. Comp.Neurol., vol. 379, pp. 133–149, 1997.
[16] P. A. Leake and R. L. Snyder, “Topographic organization of the centralprojections of the spiral ganglion in cats,” J. Comp. Neurol., vol. 281,pp. 612–629, 1989.
[17] J. K. Moore and K. K. Osen, “The cochlear nuclei in man,” Am. J. Anat.,vol. 154, pp. 393–418, 1979.
[18] J. K. Moore, “The human auditory brain stem: A comparative view,”Hear. Res., vol. 29, pp. 1–32, 1987.
[19] R. Biran, D. C. Martin, and P. A. Tresco, “Neuronal cell loss accompa-nies the brain tissue response to chronically implanted silicon micro-electrode arrays,” Exp. Neurol., vol. 195, pp. 115–126, 2005.
[20] D. H. Szarowski, M. D. Andersen, S. Retterer, A. J. Spence, M.Isaacson, H. G. Craighead, J. N. Turner, and W. Shain, “Brain re-sponses to micro-machined silicon devices,” Brain Res., vol. 983, pp.23–35, 2003.
[21] J. N. Turner, W. Shain, D. H. Szarowski, M. Andersen, S. Martins, M.Isaacson, and H. Craighead, “Cerebral astrocyte response to microma-chined silicon implants,” Exp. Neurol., vol. 156, pp. 33–49, 1999.
[22] M. N. Semple and L. M. Aitkin, “Representation of sound frequencyand laterality by units in central nucleus of cat inferior colliculus,” J.Neurophysiol., vol. 42, pp. 1626–1639, 1979.
[23] M. M. Merzenich and M. D. Reid, “Representation of the cochleawithin the inferior colliculus of the cat,” Brain Res., vol. 77, pp.397–415, 1974.
[24] M. Brown, W. R. Webster, and R. L. Martin, “The three-dimensionalfrequency organization of the inferior colliculus of the cat: A 2-de-oxyglucose study,” Hear. Res., vol. 104, pp. 57–72, 1997.
[25] K. K. Osen, “Cytoarchitecture of the cochlear nucleus in the cat,” J.Comp. Neurol., vol. 136, pp. 453–483, 1969.
[26] D. B. McCreery, T. G. Yuen, W. F. Agnew, and L. A. Bullara, “Stim-ulation with chronically implanted microelectrodes in the cochlear nu-cleus of the cat: Histologic and physiologic effects,” Hear. Res., vol.62, pp. 42–56, 1992.
[27] D. B. McCreery, T. G. H. Yuen, and L. A. Bullara, “Physiologic andHistologic effects of prolonged microstimulation in the feline ventralcochlear nucleus,” in Proc. Conf. Implantable Auditory Prostheses,Asilomar, CA, 2001, (Abstr.).
[28] E. J. Tehovnik, “Electrical stimulation of neural tissue to evoke behav-ioral responses,” J. Neurosci. Meth., vol. 65, pp. 1–17, 1996.

1052 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 6, JUNE 2007
Douglas McCreery received the B.Sc. and M.Sc.degrees in electrical engineering and the Ph.D. de-gree in biomedical engineering from the Universityof Connecticut, Storrs, in 1966, 1970, and 1975,respectively.
He is currently Director of the Neural Engineeringprogram at Huntington Medical Research Institutes.His research interests include the development ofneuroprostheses for the central nervous system, andthe physiologic and histologic effects of electricalstimulation of the central and peripheral nervous
system.Dr. McCreery is a member of the Society for Neuroscience.
Albert Lossinsky received the B.A. degree inbiology and chemistry from Kansas WesleyanUniversity, Salina, in 1969, the M.S. degree in mi-crobiology and electron microscopy from EmporiaState University, Emporia, KS, in 1974, and thePh.D. degree in Neurobiology from the MedicalResearch Center of the Polish Academy of Sciences,Warsaw, Poland, in 1994.
He is currently the Head of the Immunohistochem-istry and Scanning Electron Microscopy Laboratorieswithin the Neural Engineering Program at the Hunt-
ington Medical Research Institutes, Pasadena, CA. His research interests in-clude ultrastructural pathology, neurodegenerative changes after electrical stim-ulation of the brain and spinal cord, and transport across the blood-brain barrier.
Victor Pikov received the B.A. degree in bio-psychology from Vassar College, Poughkeepsie, NY,in 1995. In 2000, he received the Ph.D. degree in cellbiology from Georgetown University, Washington,DC.
He completed a Boswell postdoctoral fellowshipat the California Institute of Technology, Pasadena,in 2002. He is a Research Scientist in the NeuralEngineering Program at the Huntington MedicalResearch Institutes, Pasadena, CA. His researchinterest is in development of prostheses for the
central nervous system.
Copyright © 2022 FDOKUMEN




















