
Lesion of the ventral tegmental area amplifies stimulation-induced Fos expression in the rat brain
11
Research Report Lesion of the ventral tegmental area amplifies stimulation-induced Fos expression in the rat brain Irena Majkutewicz, Tomasz Cecot, Grażyna Jerzemowska, Dorota Myślińska, Karolina Plucińska, Weronika Trojniar, Danuta Wrona ⁎ Department of Animal Physiology, University of Gdańsk, 24 Kładki St., 80-822 Gdańsk, Poland ARTICLE INFO ABSTRACT Article history: Accepted 5 January 2010 Available online 14 January 2010 Unilateral lesions of the ventral tegmental area (VTA), the key structure of the mesolimbic system, facilitate behavioral responses induced by electrical stimulation of the VTA in the contralateral hemisphere. In search of the neuronal mechanism behind this phenomenon, Fos expression was used to measure neuronal activation of the target mesolimbic structures in rats subjected to unilateral electrocoagulation and simultaneously to contralateral electrical stimulation of the VTA (L/S group). These were compared to the level of mesolimbic activation after unilateral electrocoagulation of the VTA (L group), unilateral electrical stimulation of the VTA (S group) and bilateral electrode implantation into the VTA in the sham (Sh) group. We found that unilateral stimulation of the VTA alone increased the density of Fos containing neurons in the ipsilateral mesolimbic target structures: nucleus accumbens, lateral septum and amygdala in comparison with the sham group. However, unilateral lesion of the VTA was devoid of effect in non-stimulated (L) rats and it significantly amplified the stimulation-induced Fos-immunoreactivity (L/S vs S group). Stimulation of the VTA performed after contralateral lesion (L/S) evoked strong bilateral induction of Fos expression in the mesolimbic structures involved in motivation and reward (nucleus accumbens and lateral septum) and the processing of the reinforcing properties of olfactory stimuli (anterior cortical amygdaloid nucleus) in parallel with facilitation of behavioral function measured as shortened latency of eating or exploration. Our data suggest that VTA lesion sensitizes mesolimbic system to stimuli by suppressing an inhibitory influence of brain areas afferenting the VTA. © 2010 Elsevier B.V. All rights reserved. Keywords: Ventral tegmental area (VTA) Fos Lesion Contralateral stimulation Rat BRAIN RESEARCH 1320 (2010) 95 – 105 ⁎ Corresponding author. Fax: +48 58 523 64 22. E-mail address: [email protected] (D. Wrona). Abbreviations: Acb, accumbens nucleus; AcbC, accumbens nucleus, part core; AcbS, accumbens nucleus, part shell; ACo, anterior cortical amygdaloid nucleus; BLA, basolateral amygdaloid nucleus, anterior part; BST, bed nucleus of the stria terminalis; Ce, central amygdaloid nucleus; EC, entorhinal cortex; GABA, gamma-aminobutyric acid; L, unilateral lesion of the ventral tegmental area; LS, lateral septal nucleus; L/S, unilateral lesion and contralateral stimulation of the ventral tegmental area; Me, medial amygdaloid nucleus; MFB, medial forebrain bundle; MS, medial septal nucleus; NMDA, N-methyl-D-aspartate; PBS, phosphate buffered saline; S, unilateral stimulation of the ventral tegmental area; Sh, sham; VP, ventral pallidum; VTA, ventral tegmental area 0006-8993/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2010.01.009 available at www.sciencedirect.com www.elsevier.com/locate/brainres
Transcript of Lesion of the ventral tegmental area amplifies stimulation-induced Fos expression in the rat brain

B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
Lesion of the ventral tegmental area amplifiesstimulation-induced Fos expression in the rat brain
Irena Majkutewicz, Tomasz Cecot, Grażyna Jerzemowska, Dorota Myślińska,Karolina Plucińska, Weronika Trojniar, Danuta Wrona⁎
Department of Animal Physiology, University of Gdańsk, 24 Kładki St., 80-822 Gdańsk, Poland
A R T I C L E I N F O
⁎ Corresponding author. Fax: +48 58 523 64 22E-mail address: [email protected]: Acb, accumbens nucleus;
cortical amygdaloid nucleus; BLA, basolateramygdaloid nucleus; EC, entorhinal cortex; Gseptal nucleus; L/S, unilateral lesion and conmedial forebrain bundle; MS, medial septstimulation of the ventral tegmental area; Sh
0006-8993/$ – see front matter © 2010 Elsevidoi:10.1016/j.brainres.2010.01.009
A B S T R A C T
Article history:Accepted 5 January 2010Available online 14 January 2010
Unilateral lesions of the ventral tegmental area (VTA), the key structure of the mesolimbicsystem, facilitate behavioral responses induced by electrical stimulation of the VTA in thecontralateral hemisphere. In search of the neuronal mechanism behind this phenomenon,Fos expressionwas used tomeasure neuronal activation of the target mesolimbic structuresin rats subjected to unilateral electrocoagulation and simultaneously to contralateralelectrical stimulation of the VTA (L/S group). These were compared to the level ofmesolimbic activation after unilateral electrocoagulation of the VTA (L group), unilateralelectrical stimulation of the VTA (S group) and bilateral electrode implantation into the VTAin the sham (Sh) group. We found that unilateral stimulation of the VTA alone increased thedensity of Fos containing neurons in the ipsilateral mesolimbic target structures: nucleusaccumbens, lateral septum and amygdala in comparison with the sham group. However,unilateral lesion of the VTA was devoid of effect in non-stimulated (L) rats and itsignificantly amplified the stimulation-induced Fos-immunoreactivity (L/S vs S group).Stimulation of the VTA performed after contralateral lesion (L/S) evoked strong bilateralinduction of Fos expression in themesolimbic structures involved inmotivation and reward(nucleus accumbens and lateral septum) and the processing of the reinforcing properties ofolfactory stimuli (anterior cortical amygdaloid nucleus) in parallel with facilitation ofbehavioral function measured as shortened latency of eating or exploration. Our datasuggest that VTA lesion sensitizes mesolimbic system to stimuli by suppressing aninhibitory influence of brain areas afferenting the VTA.
© 2010 Elsevier B.V. All rights reserved.
Keywords:Ventral tegmental area (VTA)FosLesionContralateral stimulationRat
..pl (D. Wrona).AcbC, accumbens nucleus, part core; AcbS, accumbens nucleus, part shell; ACo, anterioral amygdaloid nucleus, anterior part; BST, bed nucleus of the stria terminalis; Ce, centralABA, gamma-aminobutyric acid; L, unilateral lesion of the ventral tegmental area; LS, lateraltralateral stimulation of the ventral tegmental area; Me, medial amygdaloid nucleus; MFB,al nucleus; NMDA, N-methyl-D-aspartate; PBS, phosphate buffered saline; S, unilateral, sham; VP, ventral pallidum; VTA, ventral tegmental area
er B.V. All rights reserved.
96 B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
1. Introduction
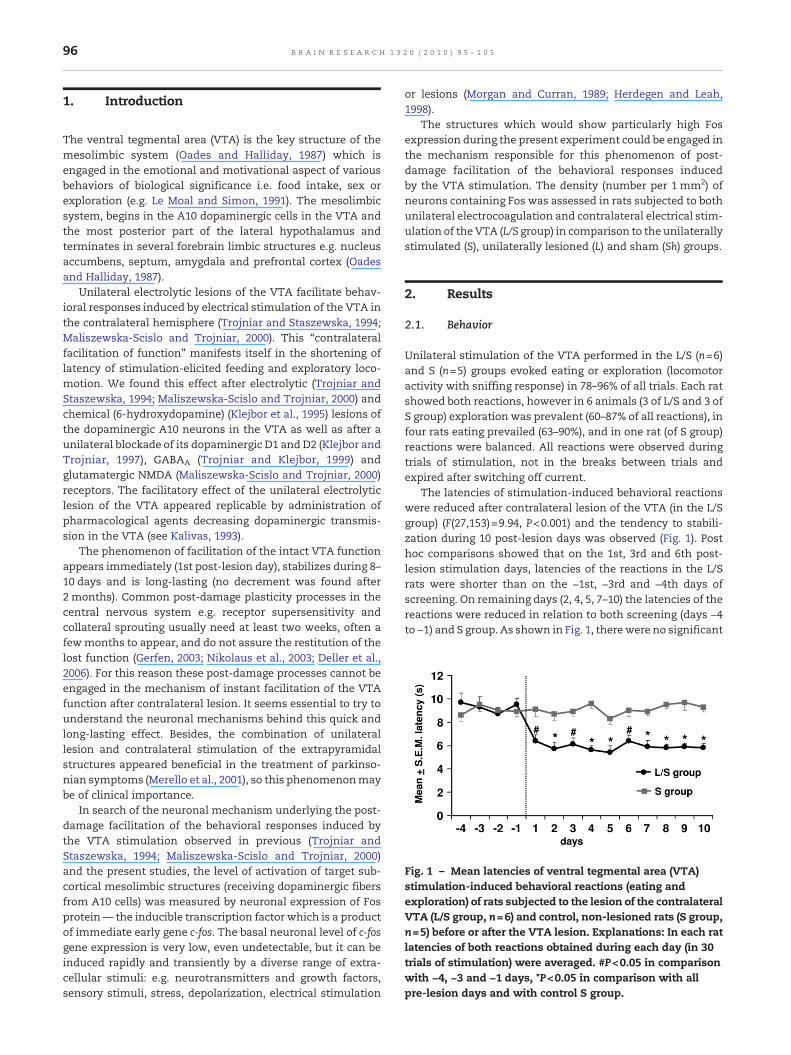
Fig. 1 – Mean latencies of ventral tegmental area (VTA)stimulation-induced behavioral reactions (eating andexploration) of rats subjected to the lesion of the contralateralVTA (L/S group, n=6) and control, non-lesioned rats (S group,n=5) before or after the VTA lesion. Explanations: In each ratlatencies of both reactions obtained during each day (in 30trials of stimulation) were averaged. #P<0.05 in comparisonwith −4, −3 and −1 days, *P<0.05 in comparison with allpre-lesion days and with control S group.
The ventral tegmental area (VTA) is the key structure of themesolimbic system (Oades and Halliday, 1987) which isengaged in the emotional and motivational aspect of variousbehaviors of biological significance i.e. food intake, sex orexploration (e.g. Le Moal and Simon, 1991). The mesolimbicsystem, begins in the A10 dopaminergic cells in the VTA andthe most posterior part of the lateral hypothalamus andterminates in several forebrain limbic structures e.g. nucleusaccumbens, septum, amygdala and prefrontal cortex (Oadesand Halliday, 1987).
Unilateral electrolytic lesions of the VTA facilitate behav-ioral responses induced by electrical stimulation of the VTA inthe contralateral hemisphere (Trojniar and Staszewska, 1994;Maliszewska-Scislo and Trojniar, 2000). This “contralateralfacilitation of function” manifests itself in the shortening oflatency of stimulation-elicited feeding and exploratory loco-motion. We found this effect after electrolytic (Trojniar andStaszewska, 1994; Maliszewska-Scislo and Trojniar, 2000) andchemical (6-hydroxydopamine) (Klejbor et al., 1995) lesions ofthe dopaminergic A10 neurons in the VTA as well as after aunilateral blockade of its dopaminergic D1 and D2 (Klejbor andTrojniar, 1997), GABAA (Trojniar and Klejbor, 1999) andglutamatergic NMDA (Maliszewska-Scislo and Trojniar, 2000)receptors. The facilitatory effect of the unilateral electrolyticlesion of the VTA appeared replicable by administration ofpharmacological agents decreasing dopaminergic transmis-sion in the VTA (see Kalivas, 1993).
The phenomenon of facilitation of the intact VTA functionappears immediately (1st post-lesion day), stabilizes during 8–10 days and is long-lasting (no decrement was found after2 months). Common post-damage plasticity processes in thecentral nervous system e.g. receptor supersensitivity andcollateral sprouting usually need at least two weeks, often afewmonths to appear, and do not assure the restitution of thelost function (Gerfen, 2003; Nikolaus et al., 2003; Deller et al.,2006). For this reason these post-damage processes cannot beengaged in the mechanism of instant facilitation of the VTAfunction after contralateral lesion. It seems essential to try tounderstand the neuronal mechanisms behind this quick andlong-lasting effect. Besides, the combination of unilaterallesion and contralateral stimulation of the extrapyramidalstructures appeared beneficial in the treatment of parkinso-nian symptoms (Merello et al., 2001), so this phenomenonmaybe of clinical importance.
In search of the neuronal mechanism underlying the post-damage facilitation of the behavioral responses induced bythe VTA stimulation observed in previous (Trojniar andStaszewska, 1994; Maliszewska-Scislo and Trojniar, 2000)and the present studies, the level of activation of target sub-cortical mesolimbic structures (receiving dopaminergic fibersfrom A10 cells) was measured by neuronal expression of Fosprotein— the inducible transcription factor which is a productof immediate early gene c-fos. The basal neuronal level of c-fosgene expression is very low, even undetectable, but it can beinduced rapidly and transiently by a diverse range of extra-cellular stimuli: e.g. neurotransmitters and growth factors,sensory stimuli, stress, depolarization, electrical stimulation
or lesions (Morgan and Curran, 1989; Herdegen and Leah,1998).
The structures which would show particularly high Fosexpression during the present experiment could be engaged inthe mechanism responsible for this phenomenon of post-damage facilitation of the behavioral responses inducedby the VTA stimulation. The density (number per 1 mm2) ofneurons containing Fos was assessed in rats subjected to bothunilateral electrocoagulation and contralateral electrical stim-ulation of the VTA (L/S group) in comparison to the unilaterallystimulated (S), unilaterally lesioned (L) and sham (Sh) groups.
2. Results
2.1. Behavior
Unilateral stimulation of the VTA performed in the L/S (n=6)and S (n=5) groups evoked eating or exploration (locomotoractivity with sniffing response) in 78–96% of all trials. Each ratshowed both reactions, however in 6 animals (3 of L/S and 3 ofS group) exploration was prevalent (60–87% of all reactions), infour rats eating prevailed (63–90%), and in one rat (of S group)reactions were balanced. All reactions were observed duringtrials of stimulation, not in the breaks between trials andexpired after switching off current.
The latencies of stimulation-induced behavioral reactionswere reduced after contralateral lesion of the VTA (in the L/Sgroup) (F(27,153)=9.94, P<0.001) and the tendency to stabili-zation during 10 post-lesion days was observed (Fig. 1). Posthoc comparisons showed that on the 1st, 3rd and 6th post-lesion stimulation days, latencies of the reactions in the L/Srats were shorter than on the −1st, −3rd and −4th days ofscreening. On remaining days (2, 4, 5, 7–10) the latencies of thereactions were reduced in relation to both screening (days −4to −1) and S group. As shown in Fig. 1, therewere no significant

97B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
changes in the latencies of stimulation-induced behaviorreactions in the S group during 10 post-lesion days.
Unilateral lesion of the VTA (L group) or electrode im-plantation (Sh group) did not influence spontaneous behavior.Neither aphagia nor adipsia was found after unilateral lesionof the VTA.
2.2. Histological verification
All lesions within the L/S (n=6) and L (n=5) groups were in theafflicted regions where, according to Tohyama and Takatsuji(1998), the dopaminergic A10 neurons are localized: themost posterior part of the lateral hypothalamus (−4.52 and−4.80 mmposterior to bregma— 6 rats), and the central part ofthe VTA (−5.20 mm posterior to bregma — 5 rats) (Fig. 2).
The tips of the stimulating electrodes of the L/S and S (n=5)rats were also found in the areas of the A10 neurons: thecaudal part of the lateral hypothalamus (6 rats), the centralpart of the VTA (2 rats), and in the posterior VTA (3 rats) (Fig. 2).
2.3. Fos expression
A two-way ANOVA (treatment×brain region) showed thatthe experimental treatment (lesion, stimulation or electrode im-plantation) significantly influenced the Fos expression F(2,484)=24.99,P<0.001, and thedensityof Fos+ cells differed in thevariousbrain regions F(10,484)=7.23, P<0.01. In addition, there was asignificant interaction between these factors F(32,484)=15.01,P<0.001.
A one-way ANOVA showed that various treatment signif-icantly influenced Fos-immunoreactivity in each studiedmesolimbic structure with an exception of the medial septum(AcbS: F(6,37)=5.93, P<0.001; AcbC: F(6,37)=4.68, P<0.01; ACo:F(6,37)=5.67, P<0.001; Me: F(6,37)=3.73, P<0.01; Ce: F(6,37)=2.45, P<0.05; BLA: F(6,37)=2.75, P<0.05; BST: F(6,37)=3.06,
Fig. 2 – Maximal extension of VTA lesions in the L/S (n=6) andstimulating electrodes in the L/S and S (n=5) (black dots) rats. PlaThe distance from the bregma of each plate is shown in the bottstimulation of the VTA; L — unilateral lesion of the VTA; S — un
P<0.05; LS: F(6,37)=5.57, P<0.001; MS: F(6,37)=1.62, P>0.05).Post hoc comparisons showed that unilateral stimulation ofthe VTA alone (S group) evoked a marked ipsilateral increasein Fos expression in the target mesolimbic structures.Interhemispheric differences (stimulated vs intact hemi-sphere) were found in the nucleus accumbens, part “shell”AcbS and “core” AcbC (Fig. 3) and all the tested regions of theamygdala (ACo, Me, Ce, BLA) and the BST (Fig. 4) and also inthe lateral septum (LS) (Fig. 5). The increase in the activity ofthe stimulated hemisphere of the S group was also significantas compared to the Sh group in all these structures. Themedial septum (MS) (Fig. 5) was unaffected.
In such structures as the nucleus accumbens (AcbS andAcbC) (Figs. 3, 6A), ACo (Figs. 4, 6B) and LS (Figs. 5, 6C),unilateral stimulation of the VTA that was made aftercontralateral lesion (in L/S group), evoked stronger activationmeasured by higher Fos expression than the unilateralstimulation of the VTA alone (S group). The increase in theFos-immunoreactivity in the L/S rats was also significant incomparison to L and S groups. The lesioned and stimulatedhemispheres of L/S rats were activated to a similar degreewithno interhemispheric differences being found.
Unilateral lesion of the VTA alone (L group) had no influenceon the neuronal activity of the tested mesolimbic structures.No significant interhemispheric difference (lesioned vs intacthemisphere) or difference with the sham (Sh) animals wasfound.
3. Discussion
In the present study, unilateral electrical stimulation of theVTA (in the S group) induced Fos expression in the ipsilateraltarget mesolimbic brain areas such as nucleus accumbens(AcbS and AcbC), lateral septum (LS), amygdala (Aco, Me, Ce,
L (n=5) rats (shading pattern areas) and localization oftes were taken from the atlas by Paxinos and Watson (1998).om right corner. Explanations: L/S — lesion and contralateralilateral stimulation of the VTA.
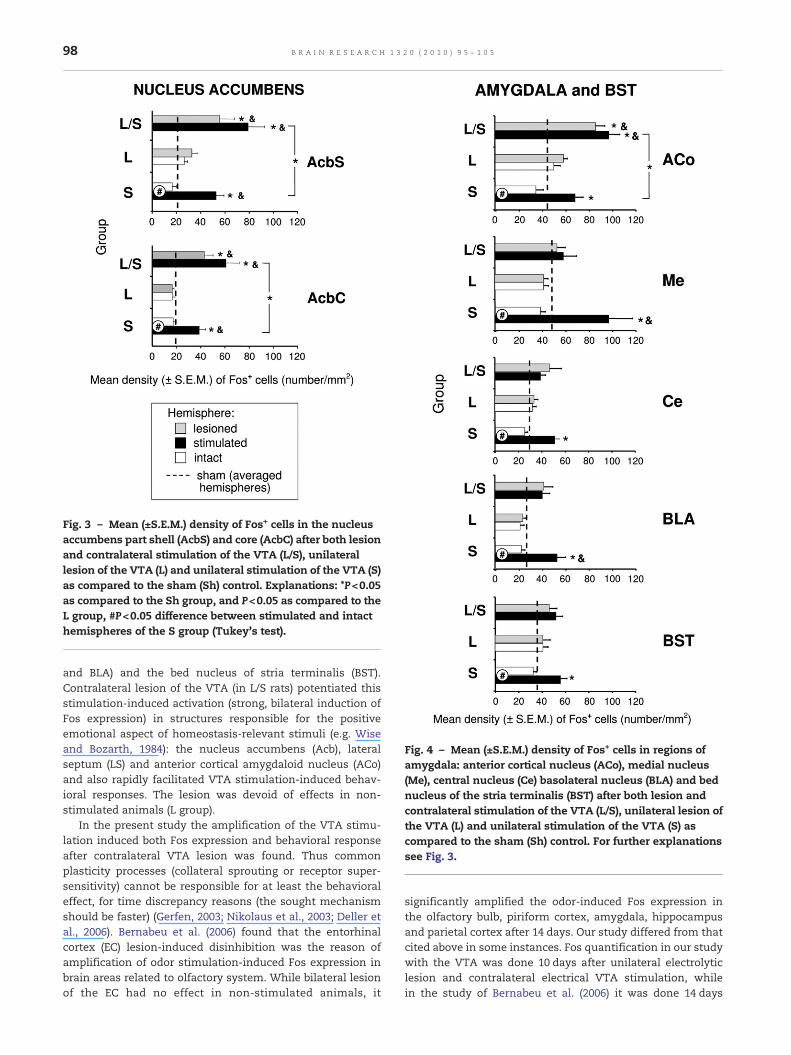
Fig. 3 – Mean (±S.E.M.) density of Fos+ cells in the nucleusaccumbens part shell (AcbS) and core (AcbC) after both lesionand contralateral stimulation of the VTA (L/S), unilaterallesion of the VTA (L) and unilateral stimulation of the VTA (S)as compared to the sham (Sh) control. Explanations: *P<0.05as compared to the Sh group, and P<0.05 as compared to theL group, #P<0.05 difference between stimulated and intacthemispheres of the S group (Tukey's test).
Fig. 4 – Mean (±S.E.M.) density of Fos+ cells in regions ofamygdala: anterior cortical nucleus (ACo), medial nucleus(Me), central nucleus (Ce) basolateral nucleus (BLA) and bednucleus of the stria terminalis (BST) after both lesion andcontralateral stimulation of the VTA (L/S), unilateral lesion ofthe VTA (L) and unilateral stimulation of the VTA (S) ascompared to the sham (Sh) control. For further explanationssee Fig. 3.
98 B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
and BLA) and the bed nucleus of stria terminalis (BST).Contralateral lesion of the VTA (in L/S rats) potentiated thisstimulation-induced activation (strong, bilateral induction ofFos expression) in structures responsible for the positiveemotional aspect of homeostasis-relevant stimuli (e.g. Wiseand Bozarth, 1984): the nucleus accumbens (Acb), lateralseptum (LS) and anterior cortical amygdaloid nucleus (ACo)and also rapidly facilitated VTA stimulation-induced behav-ioral responses. The lesion was devoid of effects in non-stimulated animals (L group).
In the present study the amplification of the VTA stimu-lation induced both Fos expression and behavioral responseafter contralateral VTA lesion was found. Thus commonplasticity processes (collateral sprouting or receptor super-sensitivity) cannot be responsible for at least the behavioraleffect, for time discrepancy reasons (the sought mechanismshould be faster) (Gerfen, 2003; Nikolaus et al., 2003; Deller etal., 2006). Bernabeu et al. (2006) found that the entorhinalcortex (EC) lesion-induced disinhibition was the reason ofamplification of odor stimulation-induced Fos expression inbrain areas related to olfactory system. While bilateral lesionof the EC had no effect in non-stimulated animals, it
significantly amplified the odor-induced Fos expression inthe olfactory bulb, piriform cortex, amygdala, hippocampusand parietal cortex after 14 days. Our study differed from thatcited above in some instances. Fos quantification in our studywith the VTA was done 10 days after unilateral electrolyticlesion and contralateral electrical VTA stimulation, whilein the study of Bernabeu et al. (2006) it was done 14 days
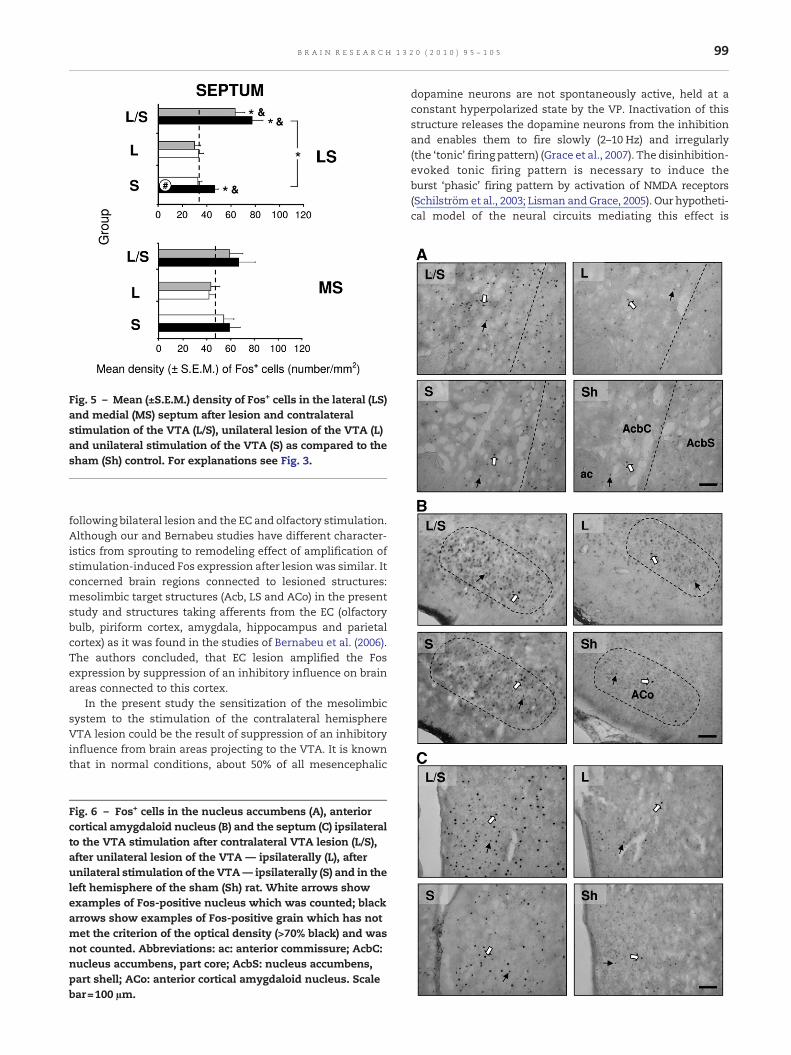
Fig. 5 – Mean (±S.E.M.) density of Fos+ cells in the lateral (LS)and medial (MS) septum after lesion and contralateralstimulation of the VTA (L/S), unilateral lesion of the VTA (L)and unilateral stimulation of the VTA (S) as compared to thesham (Sh) control. For explanations see Fig. 3.
99B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
following bilateral lesion and the EC and olfactory stimulation.Although our and Bernabeu studies have different character-istics from sprouting to remodeling effect of amplification ofstimulation-induced Fos expression after lesionwas similar. Itconcerned brain regions connected to lesioned structures:mesolimbic target structures (Acb, LS and ACo) in the presentstudy and structures taking afferents from the EC (olfactorybulb, piriform cortex, amygdala, hippocampus and parietalcortex) as it was found in the studies of Bernabeu et al. (2006).The authors concluded, that EC lesion amplified the Fosexpression by suppression of an inhibitory influence on brainareas connected to this cortex.
In the present study the sensitization of the mesolimbicsystem to the stimulation of the contralateral hemisphereVTA lesion could be the result of suppression of an inhibitoryinfluence from brain areas projecting to the VTA. It is knownthat in normal conditions, about 50% of all mesencephalic
Fig. 6 – Fos+ cells in the nucleus accumbens (A), anteriorcortical amygdaloid nucleus (B) and the septum (C) ipsilateralto the VTA stimulation after contralateral VTA lesion (L/S),after unilateral lesion of the VTA — ipsilaterally (L), afterunilateral stimulation of the VTA— ipsilaterally (S) and in theleft hemisphere of the sham (Sh) rat. White arrows showexamples of Fos-positive nucleus which was counted; blackarrows show examples of Fos-positive grain which has notmet the criterion of the optical density (>70% black) and wasnot counted. Abbreviations: ac: anterior commissure; AcbC:nucleus accumbens, part core; AcbS: nucleus accumbens,part shell; ACo: anterior cortical amygdaloid nucleus. Scalebar=100 µm.
dopamine neurons are not spontaneously active, held at aconstant hyperpolarized state by the VP. Inactivation of thisstructure releases the dopamine neurons from the inhibitionand enables them to fire slowly (2–10 Hz) and irregularly(the ‘tonic’ firing pattern) (Grace et al., 2007). The disinhibition-evoked tonic firing pattern is necessary to induce theburst ‘phasic’ firing pattern by activation of NMDA receptors(Schilström et al., 2003; Lisman andGrace, 2005). Our hypotheti-cal model of the neural circuits mediating this effect is

100 B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
presented in Fig. 7. Although disinhibition of dopamine cells oftheVTA is essential for their “burst” firingmode and conductiveto Fos expression, our hypothetical model was simplified toclearly show the lesion-induced “contralateral facilitation” ofthe function of the intact VTA. The leading role in thishypothetical mechanism plays the nucleus accumbens (Acb),which receives the greatest part of hyperpolarizing dopaminer-gic and GABA-ergic projections originating in the VTA (e.g.Thierry et al., 1980; Oades and Halliday, 1987; Steffensen et al.,1998; Tzschentke and Schmidt, 2000). The Acb sends inhibitiveGABA-ergic impulses predominantly to the ventral pallidum(VP), which strongly inhibits (also with GABA) the VTA (Geislerand Zahm, 2005; Grace et al., 2007). Numerous axons of the VPreach the VTA in the contralateral hemisphere (Geisler andZahm, 2005). Unilateral lesion of the VTA, causes disinhibitionof the ipsilateral Acb. Hyperactivation of the Acb during thestudied phenomenon causes inhibition of the VP, which stops
Fig. 7 – Hypothetical, simplified model of the neural circuitsmediating the facilitation of the Fos expression andbehavioral responses evoked by VTA stimulation aftercontralateral VTA lesion. The model shows only thesemesolimbic target structures, in which lesion of the VTAamplified stimulation-induced Fos expression: the nucleusaccumbens (Acb), lateral septum (LS), anterior corticalamygdaloid nucleus (ACo) and structures which project tothe contralateral VTA: ventral pallidum (VP) and lateralhypothalamus (LH). The VTA receives reciprocal afferentsfrom all structures to which projects (for clarity ipsilateralreciprocal projections are not shown). In basal conditions(A) VTA is balanced: it is under strong hyperpolarizinginfluence of (among others) the ventral pallidum (VP) andunder activating influences from pontomesencephaliccholinergic (ACh) and glutamatergic (Glu) neurons of pedunculopontine tegmental nucleus (PPTg), medial prefrontalcortex (mPFC), amygdala (Amg) and hippocampus (Hip). TheVTA in both brain hemispheres provides hyperpolarizingdopaminergic or GABA-ergic projections to ipsilateral targetmesolimbic structures, most prominent to the nucleusaccumbens (Acb). The Acb is restrained and weaklyinfluences the VP, which strongly inhibits the VTA in bothbrain hemispheres (only contralateral projections areshown). After unilateral lesion of the VTA (B) ipsilateral targetstructures (Acb, LS, and ACo) are disinhibited. GABA-ergicneurons in the Acb, which projects mainly to VP, becameoveractive. The effect is the inhibition of the VP anddisinhibition of contralateral VTA, which supplied with theexcitative cholinergic and glutamatergic PPTg, mPFC, Amgand Hip impulses also becomes hyperactive. As a result, thedopaminergic mesolimbic transmission increases and itcould lead to the facilitation of behavioral function andincreased Fos expression. The ACo in the lesionedhemisphere potentiates this effect by excitatory influence onthe Acb. Lateral septum (LS) after VTA lesion strongly inhibitsthe LH, and minimizes its influence on contralateral VTA.Due to the diversity of the LH projection (both excitatory andinhibitory) the effect of the LS disinhibition on thenon-lesioned VTA is difficult to predict. The thickness of thearrows reflects the intensity of impulsation.
the inhibition of the contralateral (non-lesioned) VTA. Disin-hibited VTA, which is constantly supplied with excitativecholinergic and glutamatergic pontomesencephalic impulsa-tion (Oakman et al., 1995; Parent et al., 1999) also becomeshyperactive, which can be the reason for the improvement ofbehavioral function and increased Fos expression in L/S rats.Moreover, for future studies, lesion of the corpus callosum anddirect stimulation of the ipsilateral VP after VTA lesion couldgive more precise information about the participation of the VPin this process.
As shown in our hypothetical model (Fig. 7), the VTA sendsinhibitory influence to the ipsilateral Acb, thus stimulating theVTA should decrease the number of Fos+ cells detected in thatAcb. However, in the present study, the long term (10 post-lesion days) induction of Fos protein expression in the Acb,higher following stimulation of the VTA (S and L/S groups)

101B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
than after VTA lesion (disinhibition of the Acb from the VTAinfluence) was observed. These discrepancies can be attribut-ed to the existence of additional important connectionsbetween the Acb, cortical and hippocampal structures. TheAcb receives afferent inputs from several frontal cortical(mPFC) and limbic cortical regions (Amg and Hip), withexcitatory glutamatergic afferents from these regions over-lapping with a VTA dopamine input onto single neurons inthis structure (Floresco et al., 2001). Therefore, the Acb ispositioned to integrate substantial amounts of informationunder the modulatory influence of the dopamine system forsubsequent processing by output regions, such as the ventralpallidum (VP).We think that the VTA stimulation-inducedAcbhyperactivation which is reflected in higher Fos proteinexpression is possibly in response to an activation of D1receptors by the high-amplitude, phasic dopamine signal thatpotentiates the drive of Acb neurons by glutamatergic inputsfrom the hippocampus (Goto and Grace, 2005; Grace et al.,2007).
In addition, the ACo and LS, which also were particularlyactive during the phenomenon, play supporting role in thismechanism. The ACo sends excitatory glutamatergic/asparta-tergic projections to the Acb (e.g. Ray et al., 1992; Ubeda-Bañonet al., 2007) and it additionally drives the Acb–VP–VTA circuit.The neurons in the LS are predominantly GABA-ergic (e.g.Sheehan et al., 2004) and their axons reach among others thelateral hypothalamus (LH), which is in turn the source ofdiverse (excitatory: glutamatergic, orexinergic or inhibitory:GABA-ergic, dopaminergic) transmission to the VTA in theipsi- and contralateral hemisphere (Gao and van den Pol, 2001;Geisler and Zahm, 2005). Lesion of the VTA releases the LSfrom the hyperpolarizing influence of dopamine. The LSstronger inhibits the LH, which is additionally restrained bythe nucleus accumbens. However, the effect of LH inhibitionon the activity of contralateral VTA neurons is difficult topredict, LS–LH–VTA is an alternative circuit, which can in-fluence this non-lesioned VTA.
We found previously that amplification of VTA stimula-tion-induced behavioral responses occurs not only after lost ofinhibitory (GABA-ergic or dopaminergic) transmission in theVTA but also after blockade of glutamatergic NMDA receptors(Maliszewska-Scislo and Trojniar, 2000) which are present onprojecting neurons (dopaminergic and GABA-ergic) in thisstructure (Seutin et al., 1990). Moreover, MK-801 which isNMDA receptors' antagonist, administered unilaterally intothe VTA lowered dopamine level in the ipsilateral Acb(Karreman et al., 1996; Westerink et al., 1996). Decreaseddopaminergic (and to a lesser extend GABA-ergic) transmis-sion to the Acb and other mesolimbic structures could beresponsible for the effects observed after unilateral lesion ofthe VTA in our study. Dopamine and GABA from the VTA havebeen shown to inhibit spiny projecting neurons in the nucleusaccumbens (Meredith et al., 1993; Tzschentke and Schmidt,2000), so lesion of the VTA or blockade of NMDA receptorsdisinhibits the nucleus accumbens and also triggers themechanism described above.
The increase in Fos expression in the mesolimbic targetstructures involved in this mechanism (e.g. Acb, LS and ACo)may be connected with both lesion and stimulation of theVTA. Strong bilateral induction of Fos expression, indistin-
guishable in both hemispheres was found in L/S rats. In thelesioned hemisphere target mesolimbic structures are disin-hibited (released from dopaminergic/GABA-ergic influences).They became susceptible for each, even weak stimulatoryinfluences. It is known that electrical stimulation of the VTAor the medial forebrain bundle carrying its axons, induces theFos expression mainly in the ipsilateral target structures(Arvanitogiannis et al., 1996; Chergui et al., 1996; Hunt andMcGregor, 1998). However, sensitized contralateral structurescould be activated even by the sparse contralateral VTAprojection (Oades and Halliday, 1987) or by stimuli revealedduring behavioral response to this stimulation (e.g. an odorand taste during eating response or environmental conditionsduring exploration).
There are no previous studies on Fos expression afterelectrolytic VTA lesion, so the present study is the pioneerone. We found no change in Fos expression in rats subjectedsolely to the unilateral VTA lesion. This result should also bediscussed with our model in Fig. 7. Although ipsilateral targetstructures lost their dopaminergic or GABA-ergic innervationsand became disinhibited (contralateral target structures areoverinhibited via the Acb–VP circuit-dependent disinhibitionof unlesioned VTA), there was no stimulus, which wasnecessary to induce Fos expression (e.g. Morgan and Curran,1989). Rats of our L group were not stimulated electrically orenvironmentally; they stayed in their home cages for 10 daysafter the lesion. Similarly, Seillier et al. (2003) and Bernabeuet al. (2006) did not observe increased Fos expression in thedenervated structures 14 and 15 days after lesion of theentorhinal cortex. Moreover, it has been shown that aunilateral electrolytic lesion of the entorhinal cortex (Haaset al., 1999) or the anteromedial cortex (Buytaert et al., 2001)leads to a rapid andwidespread induction of c-Fos protein andthis upregulation lasted until two days post-lesion, only. Inaddition Lanuza et al. (2008) have shown that unilateralelectrolytic lesions of the posterior intralaminar thalamicnucleus reduced footshock-induced Fos protein activation inthe lateral nucleus of the amygdala ipsilateral to the lesion inrats. Apparently the disinhibition of ipsilateral target struc-tures is relevant for the induction of Fos only when theneuronal circuit is externally excited (e.g. Herdegen and Leah,1998) which could be the reason for the lack of interhemi-spheric differences in our L rats. Not only electrical stimula-tion can be the stimulus triggering Fos expression, variousstimuli were also shown to increase the transcription of c-fosgene and production of Fos protein (for review see Herdegenand Leah, 1998). In the present study both groups in whichinduction of Fos expression were observed (L/S and S), werestimulated not only electrically but also behaviorally. Stimu-lation elicited feeding or exploratory behavior with 78–96%efficacy. It is impossible to separate the influences of electricaland behavioral stimulation on Fos expression in our study.The previous studies have shown that stimulation of theMFB inurethane anesthetized rats induced Fos-immunoreactivity inthe ipsilateral nucleus accumbens and lateral septum (Cherguiet al., 1996). This effect coexisted with increased DA levels inthese structures andwas dependent onD1 receptors. It emergesthat an increase in Fos expression in the Acb and LS couldbe independent on behavioral and environmental stimuli. How-ever, we found also increased Fos-immunoreactivity in the

102 B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
amygdala in electrical/behavioral stimulation conditions (in theS group). Our results are consistent with previous studies(Arvanitogiannis et al., 1996; Hunt and McGregor, 1998) wherebehaviorally relevant electrical stimulation or self-stimulationof the medial forebrain bundle (MFB), carrying afferents andefferents of the VTA, induced Fos expression in the ipsilateralamygdala, nucleus accumbens and lateral septum. There ismuch evidence that environmental, behavioral or emotionalstimuli can increase Fos expression not only in the amygdala(Nikolaev et al., 2002; Hale et al., 2008) but also in the Acb and LS(Cullinanet al., 1995;Duncanet al., 1996). Fromtheabovestudiesit emerges, that influences of electrical and behavioral/environ-mental stimuli on Fos expression in our L/S and S rats areintermingled and indistinguishable. Moreover, previous studies(Struthers et al., 2005; Weinberg et al., 2009) have shown thehabituation of Fos expression when the environmental orbehavioral stimuli were repeated, as in our L/S and S groups.Lesion-induced disinhibition in L/S rats could only amplify theeffect of stimulation, therefore the increase in Fos expression inL/S ratswasbilateralwhile innon-lesionedS ratsunilateral only.
We conclude that the lesion of the VTA leads some of itstarget structures (Acb, LS and ACo) to higher sensitivity tocontralateral stimulation, which manifests itself in strongbilateral induction of the Fos expression coexisting with thefacilitation of the behavioral function of the mesolimbicsystem. The mechanism of the facilitation of behavioralresponse induced by stimulation can be related to thedisinhibition of the VTA via the neuronal circuit composedof these hyperreactive target structures.
4. Experimental procedures
4.1. Animals
All animal experiments were carried out in accordance to theEuropean Communities Council Directive of 24 November1986 (86/609/EEC). The experiment was conducted under theauthority of the Local Ethical Committee for the Care and Useof Laboratory Animals of the Medical University of Gdansk,Poland. The number of animals used was the minimumjudged necessary to obtain significant results, and all effortswere made to minimize the animals' discomfort.
Studies were performed using male Wistar rats (TheInstitute of Work Medicine, Łódź, Poland, n=22) weighingapproximately 250–300 g at the time of arrival at the labora-tory. They were housed in individual cages with free access tofood and water under a 12 h light/12 h dark illumination cyclewith lights on at 6:00 AM. The animals were handled andadapted to the presence of an experimenter for 7 days. Therats were assigned to four groups: L/S (n=6) subjected tounilateral lesion and contralateral stimulation of the VTAconcomitantly, L (n=5) subjected to unilateral VTA lesion, S(n=5) subjected to unilateral stimulation of the VTA alone, andSh (n=6) bilaterally electrode implanted.
4.2. Surgery
All ratswere implanted, under pentobarbital anesthesia (50 mg/kg, i.p.) with bilateral VTAelectrodes (monopolar, stainless steel
electrodes; 0.3 mm diameter, insulated for their entire lengthexcept for the flat-cut tip). Coordinates for the ventral tegmentalarea were established according to Paxinos and Watson (1998):4.5 mm posterior to the bregma, 0.95mm lateral to the midlineand 8 mm ventral to the skull surface. The electrodes wereanchored to four stainless steel skull screwswith dental acrylic;stainless steel wire soldered to one screw served as the anodefor electrical stimulation.
After the surgery the rats of the L group stayed in theirhome cages for 11 days (until they were lesioned). The rats ofSh group stayed in their home cages for 21 days and thenweresacrificed.
4.3. Stimulation screening
After one-week recovery from the surgery the L/S and S groupswere screened for VTA stimulation-induced behaviors accord-ing to the method we described previously (Trojniar andStaszewska, 1994; Maliszewska-Scislo and Trojniar, 2000). Theaim of the screening was to determine individual (for eachelectrode) intensity of stimulating current which would, at astimulation frequency of 50 Hz, induce behavioral responsewith a mean latency of 8–10 s. The testing was carried out in asound attenuating chamber in a 250×350×440mm box withfood pellets covering the floor. The rats were taken from theirhome cages and placed in the testing box, where they had freeaccess to food and were allowed to explore the box for 30 minbefore testing, to allow for habituation to the experimentalconditions complete satiation. Trains of cathodal square-wave constant 0.1 ms current pulses were conducted fromthe stimulator to the electrode by flexible wire leads. Pulseduration, pulse frequency and stimulation intensity weremonitored with an oscilloscope. Screening was carried out for4 days using a fixed stimulation frequency of 50 Hz; currentintensity was raised incrementally in 30-s trials (30 trails/day,20 s rest between the trials) until exploration (forward loco-motion with sniffing) or eating were observed with 8–10 slatency. The latency was measured with the use of astopwatch which was switched on concurrently to startingstimulation and switched off when behavioral reactionstarted. The criterion for the beginning of the explorationwas making a few steps with all limbs following sniffing andfor feeding reaction starting to biting a food pellet. In each triallatencies of both reactions were measured and averaged toobtain a mean latency for each screening day. If a behavioralresponse was observed during one trail the current wasswitched off 5 s after its beginning to obtain only one reactionper trial of stimulation. The range of current intensity relevantto evoke these reactions was 120–240μA. Once determined thestimulation intensity was used for all the subsequent tests.Stimulation through each VTA electrode was tested in aseparate block of trials (15/each electrode); the one throughwhich stimulation induced a more reliable behavioral re-sponse (feeding or exploration) was chosen as a stimulatingelectrode for further experimentation.
4.4. Lesion
After completion of the screening procedure under lightketamine anesthesia (85 mg/kg, i.p.), rats of the L/S and L

103B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
groups were subjected to unilateral electrolytic lesion of theVTA. A cathodal current of 10 s duration and intensity of1.0 mA was applied through the electrode contralateral to thestimulation electrode chosen during the screening. Half of theanimals of the L/S received lesions in the right hemisphereand the other half in the left hemisphere. Three rats of the Lgroup were lesioned in the right, two in the left hemisphereand they had been staying in their home cages for 10 daysuntil they were sacrificed.
4.5. Stimulation
The rats of the L/S and S groups were subjected to daily VTAstimulation for 10 days after the lesion (L/S group) or aftercompletion of the screening procedure (S group). Chronic, 10-day stimulation (30 trials/day) was done to let the latency ofstimulation-induced behavioral reaction to stabilize. Stimula-tion conditions and procedure were identical as during thescreening except for the current intensity of the stimulationelectrode, which was constant (fixed in screening). Thelatencies of the reactions (both feeding and exploration —locomotion with sniffing) were measured in each trial andaveraged to obtain a mean latency in each day for each rat.One hour after the stimulation the rats were sacrificed andtheir brains were subjected to histological procedures.
4.6. Immunohistochemistry for Fos
The rats were overdosed with pentobarbital (120 mg/kg b.w.)and transcardially perfused (via left ventricle) with 200 ml of0.9% saline, followed by 400 ml of phosphate buffered 4%paraformaldehyde. The brains were removed from the skulland post-fixed in the same fixative for 24 h, and then storedfor 48 h in PBS containing 30% sucrose for cryoprotection. Thefrozen brains were cut with a cryostat at a thickness of 30μm.Prior to all the immunohistochemical stages, sections wererinsed several times in PBS, then incubated in 0.3% hydrogenperoxide in PBS for 10 min at room temperature, and rinsedagain in PBS. The sections were incubated for 48 h at 4°C withrabbit polyclonal antibodies to Fos protein (Santa Cruz Bio-technology, Inc.) (diluted in PBS containing 3% normal goatserum (Sigma) and 0.02% sodium azide) at a dilution of 1:500,and rinsed three times in PBS containing 0.02% sodium azideand 0.25% Triton X-100 (PBS+Triton). After a 2-h incubationwith biotinylated goat anti-rabbit IgG (a dilution of 1:200;Vector, USA) in PBS containing 0.02% sodium azide and 0.25%Triton X-100 at room temperature, the sections were rinsedwith PBS+Triton and incubated with avidin–biotin peroxidasecomplex (ABC) (a dilution of 1:100 in PBS; Vector Elite Kit, USA)for an additional hour at room temperature, then rinsed againand treated with 0.05% diaminobenzidine tetrahydrochloride(DAB) (Sigma), 0.2% glucosum (Cefarm PZF, Gdańsk, Poland),0.04% ammonium chloride (Sigma) and 0.1% ammoniumnickel (II) sulfate (Sigma) dissolved in PBS. After a fewminutesthe sections were finally incubated with 0.7 mg/100 ml ofglucose oxidase (Sigma) in distilled water, and allowed to reactfor 15–20 min. The Fos-immunoreactive cell nuclei appeareddark brown or black. The tissue sections were placed on slides,air dried and after dehydration with ethanol, mounted withDPX (Fluka).
Nissl staining was carried out both in the slices whereaccording to Tohyama and Takatsuji (1998), the dopaminergicA10 neurons are localized (ventral tegmental area and theposterior part of the lateral hypothalamus at sections 4.52 to5.80 mm posterior to Bregma) to determine the lesion andstimulating electrode placement. The sections were mountedon gelatin-coated slides, stained with Cresyl violet, dehy-drated and finally mounted with DPX (Fluka).
4.7. Counting of labeled cells
Sectionswere viewed at 4×10magnification by an investigatorblinded to the assignment of treatment groups under a lightmicroscope Nikon p600 connected to a Leica CCD-IRIS/RGBvideo camera. The image was exported to a computer with theLeica QWin software (serial no 4358). Fos-positive (Fos+) cellswere counted in both hemispheres in the whole area of eachtested mesolimbic structure: nucleus accumbens (AP 1.2 mm),shell (AcbS) and core (AcbC); septum (AP 0.2 mm): lateralnucleus (LS) andmedial nucleus (MS); amygdala (AP −2.3 mm):anterior cortical nucleus (ACo), medial nucleus (Me), centralnucleus (Ce) basolateral nucleus (BLA) and bed nucleus of thestria terminalis (BST) (AP −0.26 mm) considered to be a part ofthe “extended amygdala” (Alheid and Heimer, 1988). Theborders of the brain structures were determined on the basisof the Paxinos and Watson (1998) atlas, outlined by a counterand an area of the region was computed by the software. Theimagewas adjusted through a threshold procedure such that aneighboring area with only nonspecific background stainingwould contain no positive signals. The software has an opticaldensity and size filters which were calibrated to count darkgrains (>70% black) exceeding sizes of fifteen pixels (15 µm2),thus excluding any remaining particles inconsistent with thesize of a cell nucleus at that magnification. The density of Fos-positive cells was computed (number per 1 mm2). Whenpossible the value of three separate counts including bothbrain hemispheres of an individual brain region for eachanimal (3 sections) at a particular anterior–posterior level (mmto Bregma) according to the Paxinos and Watson (1998) atlaswas averaged and used as a single data point.
Sections at a level of 4.52 to 5.80 mm posterior to Bregma(Nissl stained) were viewed at a 2.5 magnification to deter-mine the lesion and stimulating electrode placement on thebasis of the Paxinos and Watson (1998) atlas.
4.8. Data analysis
Data are presented as a mean±standard error of the mean(S.E.M.).
The latency of behavioral response elicited by VTAstimulation was analyzed with a two-way analysis of variance(ANOVA) with the day of stimulation (14 levels) as the within-subject factor and group (lesioned L/S, non-lesioned S) as thebetween-subject factor. The differences in the means werefurther analyzed with Tukey's post hoc test.
Statistical evaluation of mean density of Fos+ cells wasperformed with a two-way ANOVA with treatment (lesion,stimulation,andelectrode implantation) as thebetween-subjectfactor and brain regions (9 levels) as the within-subject factor.For comparison of Sh animals, whose both brain hemispheres

104 B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
were subjected to identical treatment, one-way ANOVA withbrain hemisphere (left or right) as a factor was performed. Sincethere was no significant interhemispheric difference (F(1,107)=0.15,P>0.7), this datawas combined to giveonenumber for eachtested brain region. The mean number of Fos+ cells in the fourexperimental groups (L/S, L, S, andSh)was compared separatelyin each tested brain region with one-way ANOVA, the differ-ences in themeanswere further analyzedwithTukey's post hoctest, and P<0.05 was considered significant.
R E F E R E N C E S
Alheid, G.F., Heimer, L., 1988. New perspectives in basal forebrainorganization of special relevance for neuropsychiatricdisorders: the striatopallidal, amygdaloid, and corticopetalcomponents of substantia innominata. Neuroscience 27, 1–39.
Arvanitogiannis, A., Flores, C., Pfaus, J.G., Shizgal, P., 1996.Increased ipsilateral expression of Fos following lateralhypothalamic self-stimulation. Brain Res. 720, 148–154.
Bernabeu, R., Thiriet, N., Zwiller, J., Di Scala, G., 2006. Lesion of thelateral entorhinal cortex amplifies odor-induced expression ofc-fos, junB, and zif 268 mRNA in rat brain. Synapse 59, 135–143.
Buytaert, K.A., Kline, A.E., Montanez, S., Likler, E., Millar, C.J.,Hernandez, T.D., 2001. The temporal patterns of c-Fos andbasic fibroblast growth factor expression following a unilateralanteromedial cortex lesion. Brain Res. 894, 121–130.
Chergui, K., Nomikos, G.G., Mathe, J.M., Gonon, F., 1996. Burststimulation of the medial forebrain bundle selectivelyincreases Fos-like immunoreactivity in the limbic forebrain ofthe rat. Neuroscience 72, 141–156.
Cullinan, W.E., Herman, J.P., Battaglia, D.F., Akil, H., Watson, S.J.,1995. Pattern and time course of immediate early geneexpression in rat brain following acute stress. Neuroscience 64,477–505.
Deller, T., Haas, C.A., Freiman, T.M., Phinney, A., Jucker, M.,Frotscher, M., 2006. Lesion-induced axonal sprouting in thecentral nervous system. Adv. Exp. Med. Biol. 557, 101–121.
Duncan, G.E., Knapp, D.J., Breese, G.R., 1996. Neuroanatomicalcharacterization of Fos induction in rat behavioral models ofanxiety. Brain Res. 713, 79–91.
Floresco, S.B., Todd, C.L., Grace, A.A., 2001. Glutamatergic afferentsfrom the hippocampus to the nucleus accumbens regulateactivity of ventral tegmental area dopamine neurons.J. Neurosci. 21, 4915–4922.
Gao, X.B., van den Pol, A.N., 2001. Melanin concentrating hormonedepresses synaptic activity of glutamate and GABA neuronsfrom rat lateral hypothalamus. J. Physiol. 533, 237–252.
Geisler, S., Zahm, D.S., 2005. Afferents of the ventral tegmentalarea in the rat-anatomical substratum for integrativefunctions. J. Comp. Neurol. 490, 270–294.
Gerfen, C.R., 2003. D1 dopamine receptor supersensitivity in thedopamine-depleted striatum animal model of Parkinson'sdisease. Neuroscientist 9, 455–462.
Goto, Y., Grace, A.A., 2005. Dopamine-dependent interactionsbetween limbic and prefrontal cortical plasticity in the nucleusaccumbens: disruption by cocaine sensitization. Neuron 47,255–266.
Grace, A.A., Floresco, S.B., Goto, Y., Lodge, D.J., 2007. Regulation offiring of dopaminergic neurons and control of goal-directedbehaviors. Trends Neurosci. 30, 220–227.
Haas, C.A., Frotscher, M., Deller, T., 1999. Differential induction ofc-Fos, c-Jun and Jun B in the rat central nervous systemfollowing unilateral entorhinal cortex lesion. Neuroscience 90,41–51.
Hale, M.W., Hay-Schmidt, A., Mikkelsen, J.D., Poulsen, B., Shekhar,A., Lowry, C.A., 2008. Exposure to an open-field arena increases
c-Fos expression in a distributed anxiety-related systemprojecting to the basolateral amygdaloid complex.Neuroscience 155, 659–672.
Herdegen, T., Leah, J.D., 1998. Inducible and constitutivetranscription factors in the mammalian nervous system:control of gene expression by Jun, Fos and Krox, and CREB/ATFproteins. Brain Res. Rev. 28, 370–490.
Hunt, G.E., McGregor, I.S., 1998. Rewarding brain stimulationinduces only sparse Fos-like immunoreactivity indopaminergic neurons. Neuroscience 83, 501–515.
Kalivas, P.W., 1993. Neurotransmitter regulation of dopamineneurons in the ventral tegmental area. Brain Res. Rev. 18,75–113.
Karreman, M., Westerink, B.H., Moghaddam, B., 1996. Excitatoryamino acid receptors in the ventral tegmental area regulatedopamine release in the ventral striatum. J. Neurochem. 67,601–607.
Klejbor, I., Trojniar, W., 1997. Effect of unilateral blockade of VTAdopamine transmission on behavior elicited from contralateralVTA. Acta Neurobiol. Exp. 57 (supp), 21.
Klejbor, I., Trojniar, W., Tokarski, J., 1995. Unilateral VTA lesionsensitises locomotor response to contralateral VTAstimulation. Eur. J. Neurosci. 8 (supp), 134.
Lanuza, E., Moncho-Bogani, J., LeDoux, J.E., 2008. Unconditionedstimulus pathways to the amygdala: effects of lesions of theposterior intralaminar thalamus on foot-shock-induced c-Fosexpression in the subdivisions of the lateral amygdala.Neuroscience 2008 (155), 959–968.
Le Moal, M., Simon, H., 1991. Mesocorticolimbic dopaminergicnetwork: functional and regulatory roles. Physiol. Rev. 71,155–234.
Lisman, J.E., Grace, A.A., 2005. The hippocampal-VTA loop:controlling the entry of information into long-term memory.Neuron 46, 703–713.
Maliszewska-Scislo, M., Trojniar, W., 2000. Effect of unilateralinjection of MK-801 into the area of A10 cells on feedingevoked by stimulation of homologous area in thecontralateral hemisphere. Acta Neurobiol. Exp. 60,489–494.
Meredith, G.E., Pennartz, C.M., Groenewegen, H.J., 1993. Thecellular framework for chemical signalling in the nucleusaccumbens. Prog. Brain Res. 99, 3–24.
Merello, M., Starkstein, S., Nouzeilles, M.I., Kuzis, G., Leiguarda, R.,2001. Bilateral pallidotomy for treatment of Parkinson's diseaseinduced corticobulbar syndrome and psychic akinesiaavoidable by globus pallidus lesion combined withcontralateral stimulation. J. Neurol. Neurosurg. Psychiatry 71,611–614.
Morgan, J.I., Curran, T., 1989. Stimulus-transcription coupling inneurons: role of cellular immediate-early genes. TrendsNeurosci. 12, 459–462.
Nikolaev, E., Kaczmarek, L., Zhu, S.W., Winblad, B., Mohammed,A.H., 2002. Environmental manipulation differentially altersc-Fos expression in amygdaloid nuclei following aversiveconditioning. Brain Res. 957, 91–98.
Nikolaus, S., Larisch, R., Beu, M., Forutan, F., Vosberg, H.,Muller-Gartner, H.W., 2003. Bilateral increase in striataldopamine D2 receptor density in the6-hydroxydopamine-lesioned rat: a serial in vivo investigationwith small animal PET. Eur. J. Nucl. Med. Mol. Imaging 30,390–395.
Oades, R.D., Halliday, G.M., 1987. Ventral tegmental (A10) system:neurobiology: 1. Anatomy and connectivity. Brain Res. Rev. 12,117–165.
Oakman, S.A., Faris, P.L., Kerr, P.E., Cozzari, C., Hartman, B.K., 1995.Distribution of pontomesencephalic cholinergic neuronsprojecting to substantia nigra differs significantly fromthose projecting to ventral tegmental area. J. Neurosci. 15,5859–5869.

105B R A I N R E S E A R C H 1 3 2 0 ( 2 0 1 0 ) 9 5 – 1 0 5
Parent, A., Parent, M., Charara, A., 1999. Glutamatergic inputs tomidbrain dopaminergic neurons in primates. ParkinsonismRelat. Disord. 5, 193–201.
Paxinos, G., Watson, C., 1998. The Rat Brain in StereotaxicCoordinates. Academic Press, San Diego.
Ray, J.P., Russchen, F.T., Fuller, T.A., Price, J.L., 1992. Sources ofpresumptive glutamatergic/aspartatergic afferents to themediodorsal nucleus of the thalamus in the rat. J. Comp.Neurol. 320, 435–456.
Schilström, B., Rawal, N., Mameli-Engvall, M., Nomikos, G.G.,Svensson, T.H., 2003. Dual effects of nicotine on dopamineneurons mediated by different nicotinic receptor subtypes. Int.J. Neuropsychopharmacol. 6 (1), 1–11.
Seillier, A., Coutureau, E., Thiriet, N., Herbeaux, K., Zwiller, J., DiScala, G., Will, B., Majchrzak, M., 2003. Bilateral lesions of theentorhinal cortex differentially modify haloperidol- andolanzapine-induced c-fos mRNA expression in the ratforebrain. Neuropharmacology 45, 190–200.
Seutin, V., Verbanck, P., Massotte, L., Dresse, A., 1990. Evidence forthe presence of N-methyl-D-aspartate receptors in the ventraltegmental area of the rat: an electrophysiological in vitro study.Brain Res. 514, 147–150.
Sheehan, T.P., Chambers, R.A., Russel, D.S., 2004. Regulation ofaffect by the lateral septum: implications for neuropsychiatry.Brain Res. Rev. 46, 71–117.
Steffensen, S.C., Svingos, A.L., Pickel, V.M., Henriksen, S.J., 1998.Electrophysiological characterization of GABAergicneurons in the ventral tegmental area. J. Neurosci. 18,8003–8015.
Struthers,W.M., DuPriest, A., Runyan, J., 2005. Habituation reducesnovelty-induced FOS expression in the striatum and cingulatecortex. Exp. Brain Res. 167, 136–140.
Thierry, A.M., Deniau, J.M., Herve, D., Chevalier, G., 1980.Electrophysiological evidence for non-dopaminergicmesocortical andmesolimbic neurons in the rat. Brain Res. 201,210–214.
Tohyama, M., Takatsuji, K., 1998. Atlas of Neuroactive Substancesand Their Receptors in the Rat. University Press, Oxford.
Trojniar, W., Klejbor, I., 1999. Facilitatory effect of unilateral lesionof the ventral tegmental area on locomotor response tostimulation of the contralateral ventral tegmental area:involvement of GABAergic transmission. Brain Res. 641,333–430.
Trojniar, W., Staszewska, M., 1994. Unilateral damage to theventral tegmental area facilitates feeding induced bystimulation of the contralateral ventral tegmental area. BrainRes. 641, 333–340.
Tzschentke, T.M., Schmidt, W.J., 2000. Functional relationshipamong medial prefrontal cortex, nucleus accumbens, andventral tegmental area in locomotion and reward. Crit. Rev.Neurobiol. 14, 131–142.
Ubeda-Bañon, I., Novejarque, A., Mohedano-Moriano, A.,Pro-Sistiaga, P., de la Rosa-Prieto, C., Insausti, R.,Martinez-Garcia, F., Lanuza, E., Martinez-Marcos, A., 2007.Projections from the posterolateral olfactory amygdala to theventral striatum: neural basis for reinforcing properties ofchemical stimuli. BMC Neurosci. 8, 103.
Weinberg, M.S., Bhatt, A.P., Girotti, M., Masini, C.V., Day, H.E.,Campeau, S., Spencer, R.L., 2009. Repeated ferret odorexposure induces different temporal patterns ofsame-stressor habituation and novel-stressorsensitization in both hypothalamic–pituitary–adrenal axisactivity and forebrain c-fos expression in the rat.Endocrinology 150, 749–761.
Westerink, B.H., Kwint, H.F., deVries, J.B., 1996. Thepharmacology of mesolimbic dopamine neurons:a dual-probe microdialysis study in the ventral tegmental areaand nucleus accumbens of the rat brain. J. Neurosci. 16,2605–2611.
Wise, R.A., Bozarth, M.A., 1984. Brain reward circuitry: four circuitelements “wired” in apparent series. Brain Res. Bull. 12,203–208.




















