
GFR?-1 mRNA in dopaminergic and nondopaminergic neurons in the substantia nigra and ventral...
12
GFRa-1 mRNA in Dopaminergic and Nondopaminergic Neurons in the Substantia Nigra and Ventral Tegmental Area AREZOU SARABI, 1 BARRY J. HOFFER, 1 LARS OLSON, 2 AND MARISELA MORALES 1 * 1 Cellular Neurophysiology, National Institute on Drug Abuse, Baltimore, Maryland 21224 2 Department of Neuroscience, Karolinska Institute, S-17177 Stockholm, Sweden ABSTRACT Glial cell line-derived neurotrophic factor (GDNF) is a survival factor for several types of neurons, including dopaminergic (DAergic) neurons. GDNF binds with high affinity to the GDNF family receptor a-1 (GFRa-1), which is highly expressed in the midbrain. Using anatomical and lesion techniques, we demonstrated that GFRa-1 was expressed in DAergic and non-DAergic neurons in the rat midbrain. Immunohistochemical characterization of GFRa-1-expressing neurons indicated that most of the neurons that were immunopositive for the DAergic marker tyrosine hydroxylase (TH) expressed GFRa-1 in the substantia nigra pars compacta (SNC). In contrast, fewer TH-containing neurons expressed GFRa-1 in the substantia nigra pars reticulata (SNR) and the ventral tegmental area (VTA). Depletion of GFRa-1/TH neurons was observed in the SNC following treatment with the neurotoxin 6-hydroxydopamine (6-OHDA); however, GFRa-1 expression remained in some neurons located in the SNR. The g-aminobutyric acid (GABA)ergic nature of GFRa-1-expressing neurons located in the SNR, which were resistant to (6-hydroxydopamine) 6-OHDA, was established by their expression of glutamic acid decarboxylase (GAD; the synthesizing en- zyme for GABA). Further analysis indicated that coexpression of GFRa-1 and GAD varied in a rostrocaudal gradient in the SNR, substantia nigra pars lateralis (SNL), and VTA. Mid- brain DAergic and GABAergic neurons have been previously classified according to their Ca 21 binding protein (CaBP) content; thus, we also sought to investigate the proportion of midbrain GFRa-1-expressing neurons containing parvalbumin (PV), calbindin (CB), and calretinin (CR) in the midbrain. Although GFRa-1 expression was found mainly in CB- and CR-immunoreactive neurons, it was rarely observed in PV-immunolabeled neurons. Analysis of the proportion of GFRa-1-expressing neurons for each CaBP subpopulation indicated the coexistence of GFRa-1 with CR in the VTA and all subdivisions of the SN; double-labeled GFRa-1/CR neurons were distributed in the SNC, SNR, SNL, and VTA. GFRa-1/CB neurons were also detected in the SNC, SNL, and VTA. Expression of GFRa-1 in DAergic and non-DAergic neurons in the rat SN and VTA suggests that GDNF, via GFRa-1, might modulate DAergic and GABAergic functions in the nigrostriatal, mesolimbic, and nigrotha- lamic circuits of the adult rat. J. Comp. Neurol. 441:106 –117, 2001. Published 2001 Wiley-Liss, Inc. Indexing terms: GDNF; interneurons; calcium-binding proteins; g-aminobutyric acid; glutamic acid decarboxylase; Parkinson’s disease Parkinson’s disease (PD) is a neurodegenerative disor- der characterized by progressive degeneration of dopa- mine (DA)-containing neurons (Bernheimer et al., 1973; Hornykiewicz and Kish, 1986). Glial cell line-derived neu- rotrophic factor (GDNF), originally cloned from a glial cell line (Lin et al., 1993), has become a candidate for the treatment of PD due to its neuroprotective and neurore- generative effects observed on DA neurons in animal mod- els of PD (Stromberg et al., 1993; Hoffer et al., 1994; Beck Grant sponsor: the Intramural Research Program, NIDA, NIH *Correspondence to: Marisela Morales, Cellular Neurophysiology, Na- tional Institute on Drug Abuse, 5500 Nathan Shock Drive, Baltimore, MD 21224. E-mail: [email protected] Received 26 December 2000; Revised 15 June 2001; Accepted 17 Sep- tember 2001 THE JOURNAL OF COMPARATIVE NEUROLOGY 441:106 –117 (2001) PUBLISHED 2001 WILEY-LISS, INC. ² This article is a US Government work and, as such, is in the public domain in the United States of America.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of GFR?-1 mRNA in dopaminergic and nondopaminergic neurons in the substantia nigra and ventral...

GFRa-1 mRNA in Dopaminergic andNondopaminergic Neurons in the Substantia
Nigra and Ventral Tegmental Area
AREZOU SARABI,1 BARRY J. HOFFER,1 LARS OLSON,2AND MARISELA MORALES1*
1Cellular Neurophysiology, National Institute on Drug Abuse, Baltimore, Maryland 212242Department of Neuroscience, Karolinska Institute, S-17177 Stockholm, Sweden
ABSTRACTGlial cell line-derived neurotrophic factor (GDNF) is a survival factor for several types of
neurons, including dopaminergic (DAergic) neurons. GDNF binds with high affinity to theGDNF family receptor a-1 (GFRa-1), which is highly expressed in the midbrain. Usinganatomical and lesion techniques, we demonstrated that GFRa-1 was expressed in DAergicand non-DAergic neurons in the rat midbrain. Immunohistochemical characterization ofGFRa-1-expressing neurons indicated that most of the neurons that were immunopositive forthe DAergic marker tyrosine hydroxylase (TH) expressed GFRa-1 in the substantia nigrapars compacta (SNC). In contrast, fewer TH-containing neurons expressed GFRa-1 in thesubstantia nigra pars reticulata (SNR) and the ventral tegmental area (VTA). Depletion ofGFRa-1/TH neurons was observed in the SNC following treatment with the neurotoxin6-hydroxydopamine (6-OHDA); however, GFRa-1 expression remained in some neuronslocated in the SNR. The g-aminobutyric acid (GABA)ergic nature of GFRa-1-expressingneurons located in the SNR, which were resistant to (6-hydroxydopamine) 6-OHDA, wasestablished by their expression of glutamic acid decarboxylase (GAD; the synthesizing en-zyme for GABA). Further analysis indicated that coexpression of GFRa-1 and GAD varied ina rostrocaudal gradient in the SNR, substantia nigra pars lateralis (SNL), and VTA. Mid-brain DAergic and GABAergic neurons have been previously classified according to theirCa21 binding protein (CaBP) content; thus, we also sought to investigate the proportion ofmidbrain GFRa-1-expressing neurons containing parvalbumin (PV), calbindin (CB), andcalretinin (CR) in the midbrain. Although GFRa-1 expression was found mainly in CB- andCR-immunoreactive neurons, it was rarely observed in PV-immunolabeled neurons. Analysisof the proportion of GFRa-1-expressing neurons for each CaBP subpopulation indicated thecoexistence of GFRa-1 with CR in the VTA and all subdivisions of the SN; double-labeledGFRa-1/CR neurons were distributed in the SNC, SNR, SNL, and VTA. GFRa-1/CB neuronswere also detected in the SNC, SNL, and VTA. Expression of GFRa-1 in DAergic andnon-DAergic neurons in the rat SN and VTA suggests that GDNF, via GFRa-1, mightmodulate DAergic and GABAergic functions in the nigrostriatal, mesolimbic, and nigrotha-lamic circuits of the adult rat. J. Comp. Neurol. 441:106–117, 2001. Published 2001 Wiley-Liss, Inc.†
Indexing terms: GDNF; interneurons; calcium-binding proteins; g-aminobutyric acid; glutamic
acid decarboxylase; Parkinson’s disease
Parkinson’s disease (PD) is a neurodegenerative disor-der characterized by progressive degeneration of dopa-mine (DA)-containing neurons (Bernheimer et al., 1973;Hornykiewicz and Kish, 1986). Glial cell line-derived neu-rotrophic factor (GDNF), originally cloned from a glial cellline (Lin et al., 1993), has become a candidate for thetreatment of PD due to its neuroprotective and neurore-generative effects observed on DA neurons in animal mod-els of PD (Stromberg et al., 1993; Hoffer et al., 1994; Beck
Grant sponsor: the Intramural Research Program, NIDA, NIH*Correspondence to: Marisela Morales, Cellular Neurophysiology, Na-
tional Institute on Drug Abuse, 5500 Nathan Shock Drive, Baltimore, MD21224. E-mail: [email protected]
Received 26 December 2000; Revised 15 June 2001; Accepted 17 Sep-tember 2001
THE JOURNAL OF COMPARATIVE NEUROLOGY 441:106–117 (2001)
PUBLISHED 2001 WILEY-LISS, INC. †This article is a USGovernment work and, as such, is in the public domain in theUnited States of America.
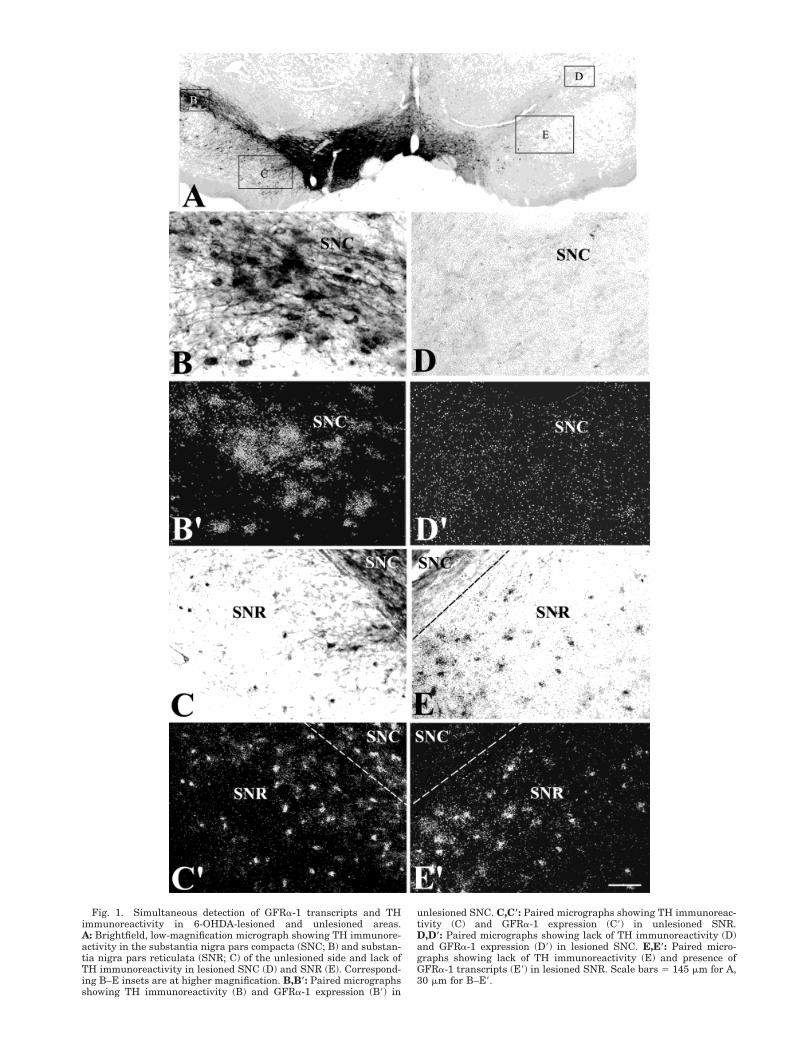
Fig. 1. Simultaneous detection of GFRa-1 transcripts and THimmunoreactivity in 6-OHDA-lesioned and unlesioned areas.A: Brightfield, low-magnification micrograph showing TH immunore-activity in the substantia nigra pars compacta (SNC; B) and substan-tia nigra pars reticulata (SNR; C) of the unlesioned side and lack ofTH immunoreactivity in lesioned SNC (D) and SNR (E). Correspond-ing B–E insets are at higher magnification. B,B*: Paired micrographsshowing TH immunoreactivity (B) and GFRa-1 expression (B9) in
unlesioned SNC. C,C*: Paired micrographs showing TH immunoreac-tivity (C) and GFRa-1 expression (C9) in unlesioned SNR.D,D*: Paired micrographs showing lack of TH immunoreactivity (D)and GFRa-1 expression (D9) in lesioned SNC. E,E*: Paired micro-graphs showing lack of TH immunoreactivity (E) and presence ofGFRa-1 transcripts (E9) in lesioned SNR. Scale bars 5 145 mm for A,30 mm for B–E9.

Figure 2
108 A. SARABI ET AL

et al., 1995; Gash et al., 1995; Bowenkamp et al., 1995;Winkler et al., 1996; Choi-Lundberg et al., 1997). In arecent study, it was also demonstrated that delivery ofGDNF cDNA into the nigrostriatal systems, using lenti-viral vectors, was able to prevent nigrostriatal degenera-tion and induce regeneration in primate models of PD(Kordower et al., 2000). In addition to DAergic neurons,GDNF promotes survival and differentiation of severalother populations of central and peripheral neurons (Hen-derson et al., 1994; Arenas et al., 1995; Buj-Bello et al.,1995; Li et al., 1995; Mount et al., 1995).
Initial studies demonstrated that responses to GDNFwere mediated through a receptor complex consisting ofthe ligand binding glycosyl-phosphatidyl inositol-linkedprotein, GFRa-1 (Jing et al., 1996; Treanor et al., 1996)and a transmembrane tyrosine kinase termed Ret (Durbecet al., 1996; Trupp et al., 1996). In this model of activation,GDNF binding to GFRa-1 induces tyrosine phosphoryla-tion (Durbec et al., 1996; Treanor et al., 1996; Trupp et al.,1996), which in turn triggers various intracellular signal-ing pathways (Creedon et al., 1997; Van Weering et al.,1997; Pong et al., 1998; Trupp et al., 1999), includingphosphatidylinositol 3-kinase (Creedon et al., 1997; VanWeering et al., 1997; Pong et al., 1998; Trupp et al., 1999)and Ras/ERK/MAPK pathways (Trupp et al., 1999). How-ever, recent evidence indicates that GDNF binding toGFRa-1 might also trigger signaling pathways indepen-dent of Ret, involving activation of Src kinase (Poteryaevet al., 1999; Trupp et al., 1999).
Following the cloning of GFRa-1 (Jing et al., 1996;Treanor et al., 1996), in situ hybridization studies wereperformed to determine the pattern of expression ofGFRa-1 as well as its anatomical relationship with itssignaling component, Ret (Nosrat et al., 1997; Trupp et al.,1997). Cellular comparison of expression of GFRa-1 andRet demonstrated the coexistence of both transcripts inseveral neurons, among them DAergic cells of the ventraltegmental area (VTA), substantia nigra (SN), anda-motoneurons (Nosrat et al., 1997; Trupp et al., 1997).This supports the idea that, in these cells, GDNF mediates
its responses through Ret receptor tyrosine kinase (Dur-bec et al., 1996; Jing et al., 1996; Treanor et al., 1996;Trupp et al., 1996). However, GFRa-1 is also expressed incells that are responsive to GDNF but lack Ret (Nosrat etal., 1997; Trupp et al., 1997). These cells might utilize aRet-independent GDNF-signaling pathway that involvesactivation of Src kinases (Poteryaev et al., 1999; Trupp etal., 1999). In agreement with functional studies demon-strating a GDNF role in the survival and differentiation ofDAergic neurons, GFRa-1expression is particularly highin the SN and VTA (Treanor et al., 1996; Nosrat et al.,1997; Trupp et al., 1997).
DAergic and g-aminobutyric acid (GABA)ergic neuronsare the two major populations of neurons present in themidbrain (Beckstead et al., 1979; Chevalier et al., 1981;Hardman et al., 1996). The substantia nigra pars com-pacta (SNC) consists of densely packed DAergic neurons,whereas both DAergic and GABAergic neurons arepresent in the substantia nigra pars reticulata (SNR),substantia nigra pars lateralis (SNL), and VTA (Beck-stead et al., 1979; Chevalier et al., 1981; Hardman et al.,1996). It is therefore possible that previously describedexpression of GFRa-1 within the midbrain may corre-spond to transcripts present in DAergic as well asGABAergic neurons. To address this question, we useddouble-labeling techniques alone or in combination with6-hydroxydopamine (6-OHDA) lesions and found thatDAergic and GABAergic neurons express GFRa-1 mRNAin the rat midbrain. Moreover, whereas GFRa-1/DAergicneurons were no longer detected in the SNC following6-OHDA lesions, GFRa-1/GABAergic neurons in the SNRwere clearly seen.
Subpopulations of midbrain neurons and projecting tar-gets can be identified according to their Ca21 bindingprotein (CaBP) content (Rogers and Resibois, 1992; Lianget al., 1996; McRitchie et al., 1996; Hontanilla et al., 1997;Nemoto et al., 1999). Accordingly, we used double-labelingtechniques to determine the degree of colocalization ofparvalbumin (PV), calbindin (CB), and calretinin (CR)with midbrain GFRa-1-expressing neurons. GFRa-1 tran-scripts were found in all three populations of CaBP-immunoreactive neurons but mainly in CB- and CR-labeled cells and rarely in PV-immunoreactive neurons.
Expression of GFRa-1 in DAergic and non-DAergic neu-rons in the rat midbrain suggests that GDNF could pro-mote survival and differentiation of both types of neurons.These findings also raise the possibility of the participa-tion of GDNF in modulating DAergic and GABAergic func-tions within the midbrain and projecting areas. Further-more, detection of coexistence of GFRa-1 with CB and CRsuggests that these molecules might provide neurons withspecific neuroprotective properties.
MATERIAL AND METHODS
Unilateral 6-OHDA lesion
Sixteen male Sprague-Dawley rats (250–300 g bodyweight) were anesthetized with chloral hydrate (35 mg/100 g) and placed in a stereotaxic apparatus. The skinover the skull was reflected. A small burr hole was made,and a 26-gauge Hamilton syringe was lowered into theskull opening. A 6-OHDA solution (8 mg/4 ml in normalsaline with 0.2 ml/ml ascorbic acid) was then infused over4 minutes into the medial forebrain bundle (MFB). Ten
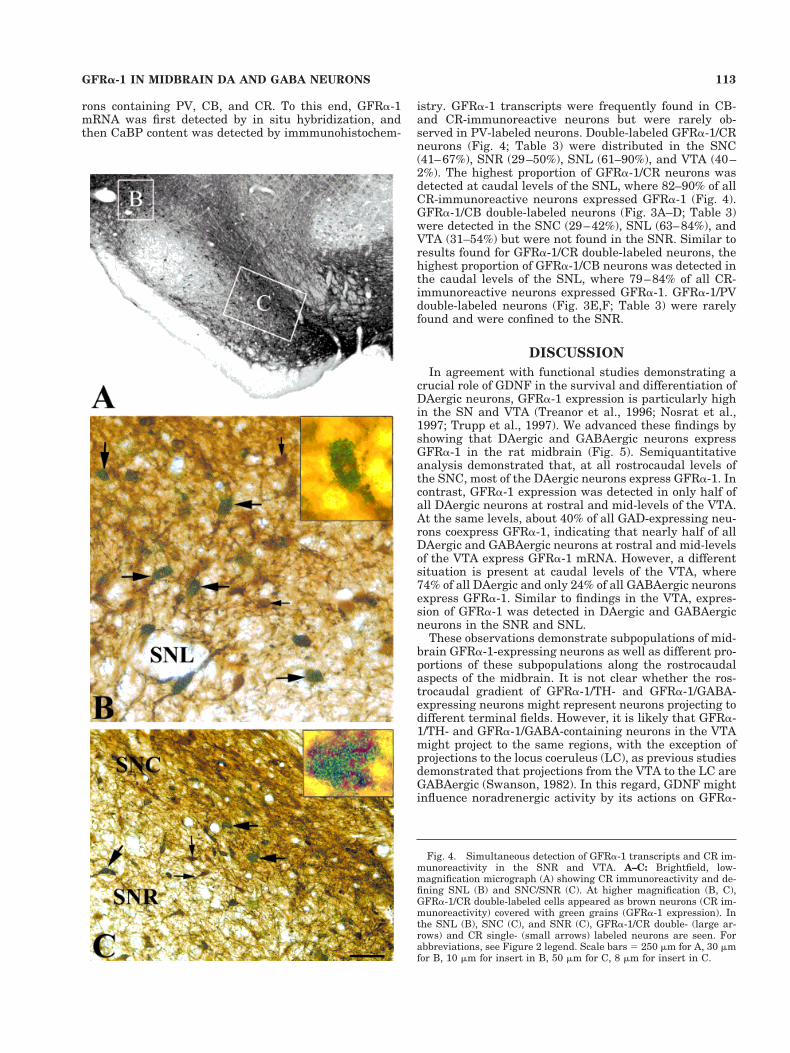
Fig. 2. Simultaneous detection of GFRa-1 transcripts and THimmunoreactivity in the substantia nigra pars reticulata (SNR) andthe ventral tegmental area (VTA). A: Brightfield, low-magnificationmicrograph showing TH immunoreactivity. B: In the VTA, GFRa-1/TH double-labeled cells appear as brown neurons (TH immunore-activity) covered with green grains (GFRa-1 expression). Examples ofdouble- (large arrows) and single- (small arrow) labeled cells areindicated. C: In the substantia nigra (SN), GFRa-1/TH double-labeledcells (large arrows) are clearly seen in the dorsal (SNCd) and ventral(SNCv) tiers of the substanatia nigra pars compacta (SNC). FewTH-immunoreactive neurons lacking GFRa-1 transcripts are seen(small arrow). Note that many GFRa-1-expressing neurons lack THsignal (arrowheads) in the SNR. D–H: Simultaneous detection ofGFRa-1 and GAD transcripts in the SN and VTA. Low magnificationof GAD-expressing neurons at rostral (D, F, G) and caudal (E, H)levels of the SN and VTA. At higher magnification, GFRa-1/GADdouble-labeled cells appear as dark neurons (GAD expression) coveredwith green grains (GFRa-1 expression). In the substantia nigra parslateralis (SNL; F), VTA (G), and SNR (H), GFRa-1/GAD double- (largearrows) and GAD single- (small arrows) labeled neurons are seen. InF, note many GFRa-1-expressing neurons lacking GAD transcripts inthe SNC (green aggregates). Scale bars 5 250 mm for A, 53 mm for B,9 mm for insert in B, 45 mm for C, 10 mm insert in C, 230 mm for D, 270mm for E, 45 mm for F, 11 mm for insert in F, 55 mm for G, 8 mm forinsert in G, 30 mm for H, 10 mm for insert in H.
109GFRa-1 IN MIDBRAIN DA AND GABA NEURONS

rats received 6-OHDA solution, and 6 control rats receivedvehicle (normal saline with 0.2 ml/ml ascorbic acid). TwoMFB lesion sites (unilateral) were targeted by using thefollowing coordinates; first target 4.4 mm posterior, 1.2mm lateral, and 7.8 mm below the dural surface; secondtarget 4.0 mm posterior, 0.8 mm lateral relative tobregma, and 8.0 mm below the dural surface (Paxinos andWatson, 1998). The surgical area was then closed withskin clips. Control animals were treated under the sameconditions, but 6-OHDA was omitted. Rotational testswere carried out in a rotometer system 3 weeks following6-OHDA lesion. Rats were placed in the rotometer bowls,and amphetamine (5 mg/kg, s.c.) was injected. The rota-tional data were recorded for 60 minutes. Lesioned ratsrotating more than 300 times during 60 minutes wereselected for perfusion. Control and 6-OHDA lesioned ratswere perfused 6 weeks after surgery.
Tissue preparation
Ten untreated adult Sprague-Dawley male rats (200–250 g body weight), four saline control, and six 6-OHDA-lesioned rats were anesthetized with chloral hydrate (35mg/100 g) and perfused transcardially with a solution of4% paraformaldehyde in 0.1 M phosphate buffer (PB), pH7.3. Brains were postfixed for 2 hours at 4°C, rinsed withPB, and sequentially transferred to 12, 14, and 18% su-crose solutions. Brains were frozen on dry ice and sec-tioned on a cryostat to obtain 30-mm coronal sections. Allprocedures were approved by the NIDA Animal Care andUse Committee.
In situ hybridization andimmunocytochemistry
In situ hybridization combined with immunolabelingwas performed as previously described (Morales et al.,1997). After prehybridization, sections were hybridizedwith [35S]-labeled sense or antisense [35S]-labeled GFRa-1probes (corresponding to nucleotides 273–1741 of the ratGFRa-1 cDNA; original GFRa-1 plasmid was kindly pro-vided by Dr. Andreas Tomac, NIDA) at 55°C for 16 hours.Sections were treated with 4 mg/ml RNase A at 37°C for 1hour and then washed in 1 3 standard saline citrate(SSC), 50% formamide at 55°C for 2 hours, and in 0.1 3SSC at 68°C for 1 hour. Material was rinsed with PB priorto processing for immunocytochemistry, incubated in 4%bovine serum albumin (BSA) supplemented with 0.3%
Triton X-100 in PB for 1 hour, followed by incubation withthe corresponding primary antibody for 24 hours at 4°C.
Primary antibody dilutions were done in PB containing1% BSA and 0.3% Triton X-100. The following antibodieswere used: anti-PV (1:2,000, Incstar, Stillwater, MN),anti-CB mouse monoclonal and anti-CR rabbit polyclonalantibodies (1:2,000, Swant, Bellinzona, Switzerland),anti-TH mouse monoclonal (1:250, Boehringer Mannheim,Indianapolis, IN). After rinsing 3 3 10 minutes in PB,sections were processed with an ABC kit (Vector, Burlin-game, CA). Material was incubated in a 1:200 dilution ofthe biotinylated secondary antibody. After rinsing withPB, sections were incubated with avidin-biotinylatedhorseradish peroxidase for 2 hours. Samples were rinsed,and the peroxidase reaction was developed with 0.05% 3,3-diaminobenzidine-4 HCl (DAB) and 0.003% hydrogenperoxide (H22O2). Sections were mounted on coated slides,air dried, dipped in nuclear track emulsion, and exposedfor several weeks prior to development. All CaBP antibod-ies used resulted in specific patterns of labeling as previ-ously described for PV (Baimbridge et al., 1992; McRitchieet al., 1996; Hontanilla et al., 1997), CR (Baimbridge et al.,1992; Rogers and Resibois, 1992; Nemoto et al., 1999) andCB (Baimbridge et al., 1992; Rogers and Resibois, 1992;Nemoto et al., 1999).
Double in situ hybridization
For double in situ hybridization, material was processedas described above by using a [35S]-labeled antisenseGFRa-1 probe and digoxigenin-labeled antisense probesfor the two isoforms of rat glutamic acid decarboxylase(GAD65 and GAD67; GAD plasmids were generously pro-vided by Dr. Allan Tobin, University of California LosAngeles, Los Angeles, CA). After this last SSC wash, sec-tions were rinsed with TBS buffer (20 mM Tris-HCl, 0.5 MNaCl), pH 8.2. Sections were incubated with an alkalinephosphatase-conjugated antibody against digoxigenin(DIG; Boehringer Mannheim) overnight at 4°C; the alka-line phosphatase reaction was developed with nitrobluetetrazolium and 5-bromo-4-chloro-3-indolyl phosphate(Life Technologies, Gaithersburg, MD). Sections weredipped in nuclear track emulsion and exposed for severalweeks prior to development.
Data analysis
Sections processed for in situ hybridization and immu-nocytochemistry were analyzed and photographed by us-ing brightfield or polarized microscopy. Photographs weretaken with a Leica DMR microscope (Leica, Deerfield, IL)
TABLE 1. Percentage of Neurons Expressing GFRa-1 and THImmunoreactivity in the Midbrain1
Region
% Double-labeled neurons (mean 6 SEM)
Level I Level II Level III
SNC 87 6 2.0 91 6 0.4 92 6 1.2(n 5 60) (n 5 53) (n 5 85)
SNR ND 44 6 0.1 66 6 0.8(n 5 40) (n 5 53)
SNL 46 6 4.0 60 6 2.0 67 6 3.0(n 5 20) (n 5 32) (n 5 25)
VTA 50 6 0.5 54 6 1.0 74 6 1.4(n 5 30) (n 5 35) (n 5 32)
1Total number of TH-immunoreactive cells (n) was counted in four sections from fourrats, and percentage of neurons expressing GFRa-1 and TH was calculated from thetotal population of TH-immunoreactive neurons. ND, double-labeled cells not detected.Three different levels of the substantia nigra (I, II, and III; see Materials and Methods)are shown. SNC, substantia nigra pars compacta; SNR, substantia nigra pars reticu-late; SNL, substantia nigra pars lateralis; VTA, ventral tegmental area; TH, tyrosinehydroxylase; GFRa-1, glial cell line-derived neurotrophic factor family receptor a-1.
TABLE 2. Percentage of Coexpression of GFRa-1 and GAD mRNAin the Midbrain1
Region
% Double-labeled neurons (mean 6 SEM)
Level I Level II Level III
SNR ND 33 6 3.0 48 6 4.6(n 5 44) (n 5 58)
SNL 52 6 1.4 37 6 1.4 34 6 0.6(n 5 24) (n 5 36) (n 5 38)
VTA 45 6 1.0 39 6 1.3 24 6 4.0(n 5 22) (n 5 30) (n 5 31)
1Total number of GAD-expressing cells (n) was counted in four sections from four rats,and percentage of neurons expressing GFRa-1 and GAD was calculated from the totalpopulation of GAD-positive cells. ND, double-labeled cells not detected. Three differentlevels of the substantia nigra (I, II, and III; see Materials and Methods) are shown.GAD, glutamic acid decarboxylase. For other abbreviations, see Table 1 footnote.
110 A. SARABI ET AL

by using Ektachrome 64T color reversal film from Kodak(EPY 135-36). The color slides were then scanned by usinga Nikon Coolscan LS2000 (Nikon, Melville, NY). Images
were opened and processed with a Photoshop 5.5 program(Adobe, Seattle, WA). Final micrographs were printed byusing a FUJIX pictrography 3000 color printer (FUJI-
Fig. 3. A–D: Simultaneous detection of GFRa-1 transcripts andCB immunoreactivity in the SNC, SNR, SNL, and VTA. Brightfield,low-magnification micrograph (A) showing CB immunoreactivity anddefining SNC/SNR (B), SNL (C), and VTA (D). At higher magnifica-tion (B–D), GFRa-1/CB double-labeled cells appeared as brown neu-rons (CB immunoreactivity) covered with blue grains (GFRa-1 expres-sion). In the SNC (B), SNL (C), and VTA (D), GFRa-1/CB double-(large arrows) and CB single- (small arrows) labeled neurons are seen.Note many GFRa-1 expressing neurons lacking CB immunoreactivity(arrowheads) in the SNR (B) and SNL (C). E,F: Simultaneous local-ization of GFRa-1 transcripts and PV-immunoreactive cells in SNR.
Brightfield, low-magnification micrograph (E) showing PV immuno-reactivity and defining SNC/SNR (F). At higher magnification (F),GFRa-1/PV double-labeled cells appeared as brown neurons (PV im-munoreactivity) covered with green grains (GFRa-1 expression).GFRa-1/PV double- (large arrows) and many PV single- (small ar-rows) labeled neurons are seen in the SNR. Note many GFRa-1-expressing neurons lacking PV immunoreactivity (green aggregates)in the SNC. For abbreviations, see Figure 2 legend. Scale bars 5 350mm for A, 45 mm for B–C, 11 mm for insert in C, 50 mm for D, 6 mm forinsert in D, 250 mm for E, 40 mm for F.
111GFRa-1 IN MIDBRAIN DA AND GABA NEURONS

FILM, Stamford, CT). The GFRa-1-expressing cells aswell as neurons containing GAD mRNA, TH, or CaBPimmunoreactivity were counted on coronal sections (Paxi-nos and Watson, 1998) through rostral (level I), mid (levelII), and caudal (level III) levels of the midbrain. The per-centage of cell profiles containing GFRa-1 mRNA andimmunoreactivity in the total population of immunola-beled neurons was calculated for each region (SNC, SNR,SNL, and VTA). A neuron was considered to be immuno-labeled when its soma was clearly seen as brown. Animmunopositive neuron was included in the calculation oftotal population of immunolabeled cells when the neuro-nal profile was at least 10 mm in diameter and was seen inthe same focal plane as the silver grains. A neuron wasconsidered double label when its soma was brown andcontained an aggregation of silver particles clearly abovethe immediately surrounding background. Similarly, thepercentage of cell profiles containing GFRa-1 mRNA andGAD mRNA in the total population of GAD mRNA-expressing neurons was calculated for each region (SNC,SNL, and VTA). A neuron was considered to express GADmRNA when its soma was clearly seen as purple. A GAD-expressing neuron was included in the calculation of totalpopulation of GAD-expressing cells when the neuronalprofile was at least 10 mm in diameter and was seen in thesame focal plane as the silver grains. A neuron was con-sidered double label when its soma was purple and con-tained an aggregation of silver particles clearly above theimmediately surrounding background. Background wasevaluated from slides hybridized with sense probes.Double-labeled cells were counted in 4–6 sections fromfour rats. Material was analyzed and photographed underbrightfield or epiluminescence microscopy by using a Ni-kon Eclipse E 800 microscope with 20 and 403 objectivelenses.
RESULTS
In agreement with previous observations, GFRa-1 tran-scripts were detected in the rat midbrain (Nosrat et al.,1997; Trupp et al., 1997). High levels of GFRa-1 tran-scripts were found throughout all levels of the SNC andVTA (Figs. 1, 2A–C). Within the SNC, GFRa-1 expressionwas equally strong in the dorsal and ventral tiers (Figs. 1,2A–C). In the SNR and SNL, GFRa-1 expression levelswere low at the rostral level and higher at the caudal level.
GFRa-1 mRNA in TH immunoreactiveneurons in the rat midbrain
Unilateral injection of 6-OHDA into the medial fore-brain bundle resulted in ipsilateral loss of TH immunore-activity in the SNC (Fig. 1A). As expected for colocaliza-tion of TH and GFRa-1 expression within DAergicneurons, loss of TH immunoreactivity was accompaniedby loss of GFRa-1 expression in the SNC (Fig. 1E,E9).However, GFRa-1 expression was maintained in the SNR(Fig. 1E9), suggesting that GFRa-1 transcripts were alsopresent in GABAergic neurons. To analyze the degree ofcolocalization of GFRa-1 expression within DAergic neu-rons in the midbrain of nonlesioned rats, GFRa-1 mRNAwas first detected by in situ hybridization and then THwas detected by immmunohistochemistry (Fig. 2A–C).SNC displayed the highest proportion of GFRa-1/TH-labeled neurons in the midbrain, where 87–92% of all
TH-immunopositive neurons expressed GFRa-1 mRNA atrostral to caudal levels, respectively (Table 1, Fig. 2A–C).Rostral levels of the SNR contained very low amounts ofGFRa-1 transcripts, whereas at more caudal levels 44–66% of all TH-containing neurons expressed GFRa-1 (Ta-ble 1). Within the SNL and VTA, there was a rostrocaudalincrease in the proportion of GFRa-1/TH-labeled neurons(Table 1); 46–67% of all TH-immunoreactive neurons ex-pressed GFRa-1 in the SNL, and 50–74% of all TH-immunoreactive neurons expressed GFRa-1 in the VTA.
GFRa-1 mRNA in GABAergic neurons inthe rat midbrain
Results from 6-OHDA-lesioned rats suggested thatGFRa-1 mRNA was expressed in GABAergic neurons inthe SNR. To examine GFRa-1 mRNA expression inGABAergic neurons in the SNL and VTA, we used doublein situ hybridization histochemistry in which GFRa-1 andGAD transcripts were simultaneously detected (Fig. 2D–H). GFRa-1/GAD coexpressing neurons were clearly seenin the SNR (Fig. 2H), SNL (Fig. 2F), and VTA (Fig. 2G) butnot in the SNC (Fig. 2F). Semiquantitative analysisshowed a rostrocaudal gradient of GFRa-1/GAD coex-pressing in the SNR and SNL (Table 2), with lowest coex-pression at the rostral level. Similarly, the subpopulationof GFRa-1/GAD coexpressing neurons varied in the VTA(Table 2): 45% of all GAD-expressing neurons coexpressedGFRa-1 at the rostral level, whereas this population wasonly 24% at the caudal level (Table 2).
GFRa-1 mRNA in different CaBP neuronsin the rat midbrain
Based on their CaBP content, it has been previouslyshown that midbrain DAergic and GABAergic neurons areheterogeneous populations. Thus, we sought to investi-gate the proportion of midbrain GFRa-1-expressing neu-
TABLE 3. Percentage of Neurons Expressing GFRa-1 and Calretinin,Calbindin, and Parvalbumin Immunoreactivity in the Midbrain1
Region
% Double-labeled neurons (mean 6 SEM)
Level I Level II Level III
Calretinin2
SNC 67 6 2.5 51 6 4.6 41 6 1.0(n 5 35) (n 5 55) (n 5 50)
SNR 35 6 3.2 42 6 1.4 50 6 5.5(n 5 30) (n 5 29) (n 5 35)
SNL 61 6 1.3 82 6 1.4 90 6 1.4(n 5 31) (n 5 33) (n 5 30)
VTA 62 6 1.2 59 6 1.4 40 6 2.8(n 5 29) (n 5 31) (n 5 25)
Calbindin3
SNC ND 29 6 0.5 42 6 3.5(n 5 10) (n 5 15)
SNL 63 6 3.0 79 6 4.0 84 6 4.0(n 5 22) (n 5 29) (n 5 31)
VTA 31 6 1.4 35 6 2.3 54 6 8.5(n 5 20) (n 5 18) (n 5 23)
Parvalbumin4
SNC ND ND NDSNR 3 6 0.7 7 6 2.8 16 6 1.0
(n 5 70) (n 5 97) (n 5 71)
1Total number of CaBP-immunoreactive neurons (n) was counted in four sections fromfour rats, and percentage of neurons expressing GFRa-1 was calculated from the totalpopulation of immunolabeled neurons. ND, double-labeled cells not detected. Threedifferent levels of the substantia nigra (I, II, and III; see Materials and Methods) areshown. CaBP, Ca21 binding protein. For other abbreviations, see Table 1 footnote.2Percentage of calretinin-immunoreactive cells that express GFRa-1.3Percentage of calbindin-immunoreactive cells that express GFRa-1.4Percentage of parvalbumin-immunoreactive cells that express GFRa-1.
112 A. SARABI ET AL
rons containing PV, CB, and CR. To this end, GFRa-1mRNA was first detected by in situ hybridization, andthen CaBP content was detected by immmunohistochem-
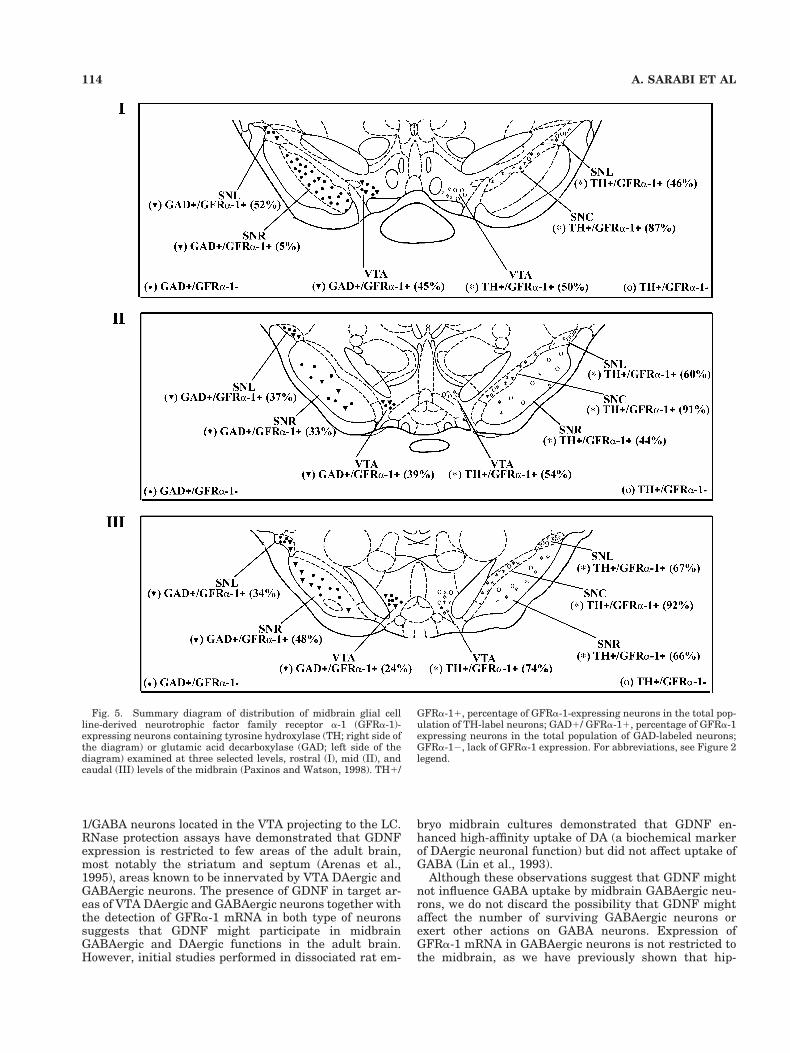
istry. GFRa-1 transcripts were frequently found in CB-and CR-immunoreactive neurons but were rarely ob-served in PV-labeled neurons. Double-labeled GFRa-1/CRneurons (Fig. 4; Table 3) were distributed in the SNC(41–67%), SNR (29–50%), SNL (61–90%), and VTA (40–2%). The highest proportion of GFRa-1/CR neurons wasdetected at caudal levels of the SNL, where 82–90% of allCR-immunoreactive neurons expressed GFRa-1 (Fig. 4).GFRa-1/CB double-labeled neurons (Fig. 3A–D; Table 3)were detected in the SNC (29–42%), SNL (63–84%), andVTA (31–54%) but were not found in the SNR. Similar toresults found for GFRa-1/CR double-labeled neurons, thehighest proportion of GFRa-1/CB neurons was detected inthe caudal levels of the SNL, where 79–84% of all CR-immunoreactive neurons expressed GFRa-1. GFRa-1/PVdouble-labeled neurons (Fig. 3E,F; Table 3) were rarelyfound and were confined to the SNR.
DISCUSSION
In agreement with functional studies demonstrating acrucial role of GDNF in the survival and differentiation ofDAergic neurons, GFRa-1 expression is particularly highin the SN and VTA (Treanor et al., 1996; Nosrat et al.,1997; Trupp et al., 1997). We advanced these findings byshowing that DAergic and GABAergic neurons expressGFRa-1 in the rat midbrain (Fig. 5). Semiquantitativeanalysis demonstrated that, at all rostrocaudal levels ofthe SNC, most of the DAergic neurons express GFRa-1. Incontrast, GFRa-1 expression was detected in only half ofall DAergic neurons at rostral and mid-levels of the VTA.At the same levels, about 40% of all GAD-expressing neu-rons coexpress GFRa-1, indicating that nearly half of allDAergic and GABAergic neurons at rostral and mid-levelsof the VTA express GFRa-1 mRNA. However, a differentsituation is present at caudal levels of the VTA, where74% of all DAergic and only 24% of all GABAergic neuronsexpress GFRa-1. Similar to findings in the VTA, expres-sion of GFRa-1 was detected in DAergic and GABAergicneurons in the SNR and SNL.
These observations demonstrate subpopulations of mid-brain GFRa-1-expressing neurons as well as different pro-portions of these subpopulations along the rostrocaudalaspects of the midbrain. It is not clear whether the ros-trocaudal gradient of GFRa-1/TH- and GFRa-1/GABA-expressing neurons might represent neurons projecting todifferent terminal fields. However, it is likely that GFRa-1/TH- and GFRa-1/GABA-containing neurons in the VTAmight project to the same regions, with the exception ofprojections to the locus coeruleus (LC), as previous studiesdemonstrated that projections from the VTA to the LC areGABAergic (Swanson, 1982). In this regard, GDNF mightinfluence noradrenergic activity by its actions on GFRa-
Fig. 4. Simultaneous detection of GFRa-1 transcripts and CR im-munoreactivity in the SNR and VTA. A–C: Brightfield, low-magnification micrograph (A) showing CR immunoreactivity and de-fining SNL (B) and SNC/SNR (C). At higher magnification (B, C),GFRa-1/CR double-labeled cells appeared as brown neurons (CR im-munoreactivity) covered with green grains (GFRa-1 expression). Inthe SNL (B), SNC (C), and SNR (C), GFRa-1/CR double- (large ar-rows) and CR single- (small arrows) labeled neurons are seen. Forabbreviations, see Figure 2 legend. Scale bars 5 250 mm for A, 30 mmfor B, 10 mm for insert in B, 50 mm for C, 8 mm for insert in C.
113GFRa-1 IN MIDBRAIN DA AND GABA NEURONS
1/GABA neurons located in the VTA projecting to the LC.RNase protection assays have demonstrated that GDNFexpression is restricted to few areas of the adult brain,most notably the striatum and septum (Arenas et al.,1995), areas known to be innervated by VTA DAergic andGABAergic neurons. The presence of GDNF in target ar-eas of VTA DAergic and GABAergic neurons together withthe detection of GFRa-1 mRNA in both type of neuronssuggests that GDNF might participate in midbrainGABAergic and DAergic functions in the adult brain.However, initial studies performed in dissociated rat em-
bryo midbrain cultures demonstrated that GDNF en-hanced high-affinity uptake of DA (a biochemical markerof DAergic neuronal function) but did not affect uptake ofGABA (Lin et al., 1993).
Although these observations suggest that GDNF mightnot influence GABA uptake by midbrain GABAergic neu-rons, we do not discard the possibility that GDNF mightaffect the number of surviving GABAergic neurons orexert other actions on GABA neurons. Expression ofGFRa-1 mRNA in GABAergic neurons is not restricted tothe midbrain, as we have previously shown that hip-
Fig. 5. Summary diagram of distribution of midbrain glial cellline-derived neurotrophic factor family receptor a-1 (GFRa-1)-expressing neurons containing tyrosine hydroxylase (TH; right side ofthe diagram) or glutamic acid decarboxylase (GAD; left side of thediagram) examined at three selected levels, rostral (I), mid (II), andcaudal (III) levels of the midbrain (Paxinos and Watson, 1998). TH1/
GFRa-11, percentage of GFRa-1-expressing neurons in the total pop-ulation of TH-label neurons; GAD1/ GFRa-11, percentage of GFRa-1expressing neurons in the total population of GAD-labeled neurons;GFRa-12, lack of GFRa-1 expression. For abbreviations, see Figure 2legend.
114 A. SARABI ET AL

pocampal GABAergic neurons express high concentra-tions of GFRa-1 transcripts (Sarabi et al., 2000). More-over, septal GABAergic neurons express high levels ofGFRa-1 mRNA (unpublished observations), and trans-plants of these GABAergic neurons manifest survival-promoting effects of GDNF (Price et al., 1996); fetal septalforebrain transplants in oculo showed a significant in-crease in the number of GABAergic neurons when theywere exposed to GDNF, compared with vehicle-treatedgrafts (Price et al., 1996).
Subpopulations of midbrain neurons and their targetscan be identified according to their CaBP contents (Rogersand Resibois et al., 1992; Liang et al., 1996; McRitchie et
al., 1996; Hontanilla et al., 1997; Nemoto et al., 1999).Considering the importance of midbrain DAergic neuronsin neurological disorders and the potential role of CaBPsand GDNF in neuroprotection, we used double-labelingtechniques to determine the degree of colocalization of PV,CB, and CR with GFRa-1 mRNA in midbrain neurons.Previous studies have shown that CB and CR proteins arepresent in DAergic neurons in the SNC as well as DAergicand GABAergic neurons in the SNR, SNL, and VTA, andthat PV is restricted to GABAergic neurons in the SNRand SNL (Rogers and Resibois et al., 1992; Liang et al.,1996; McRitchie et al., 1996; Hontanilla et al., 1997;Nemoto et al., 1999).
Fig. 6. Summary diagram of distribution of midbrain GFRa-1-expressing neurons containing CR (right side of the diagram) or CB(left side of the diagram) examined at three selected levels, rostral (I),mid (II) and caudal (III) levels of the midbrain (Paxinos & Watson,1998). CR1/ GFRa-11, percentage of GFRa-1-expressing neurons in
the total population of TH-labeled neurons; CB1/ GFRa-11, percent-age of GFRa-1-expressing neurons in the total population of GAD-labeled neurons; GFRa-12, lack of GFRa-1 expression. For abbrevi-ations, see legends to Figures 2 and 5.
115GFRa-1 IN MIDBRAIN DA AND GABA NEURONS

In agreement with the distribution of CB and CR inDAergic and GABAergic neurons, expression of GFRa-1was found mainly in CB- and CR-labeled cells in the SNand VTA. Nevertheless, GFRa-1 expression was rarelyfound in PV-immunoreactive neurons. The infrequent co-existence of PV and GFRa-1 in midbrain GABAergic neu-rons is in contrast with observations in the hippocampus,where about 70% of all PV-labeled neurons containGFRa-1 mRNA in the CA1 subfield (Sarabi et al., 2000).These results indicate that GFRa-1 expression inGABAergic/PV-containing neurons is brain region-specific. Our semiquantitative results (Fig. 6) agree withthe finding that about half of the DAergic neurons do notcontain either CR or CB in the SNC, and that in the SNLmore than half of the TH-immunoreactive cells have bothCR and CB (Liang et al., 1996; Nemoto et al., 1999). Aportion of DAergic neurons in the SN and VTA and somenon-DAergic nigral neurons project to the striatal matrix,whereas other DAergic SN neurons innervate the striatalpatches (Wright and Arbuthnott 1981; Herkenham et al.,1984; Gerfen et al., 1987). Based on CaBP contents, it wasestablished that CB-containing cells of VTA and SNproject to the striatal matrix and CR-containing neuronsto the striatal patches (Gerfen et al., 1985; Parent et al.,1996). The presence of GFRa-1 in projecting DAergic/CBand DAergic/CR neurons suggests that these neurons in-nervate both striatal matrix and striatal patches andtherefore might affect the entire striatum.
Although the functions of CaBPs are not clear, it hasbeen suggested that the buffering properties of these pro-teins might confer neuroprotective properties to neurons.Observations from human brains with PD and animalmodels of PD have shown that CB- and CR-immunoreactive neurons are less prone to degeneration inthe SNC, compared with SNC neurons lacking these pro-teins (Yamada et al., 1990; Lavoie and Parent; 1991; Ger-man et al., 1992; Iacopino et al., 1992; Hirsch, 1994;Mouatt-Prigent et al., 1994). These findings are in sharpcontrast with results in cases of Huntington’s disease(Kiyama et al., 1990) and Down’s syndrome (Kobayashi etal., 1990) where CB-containing neurons have proved to beparticularly vulnerable, indicating that the sole presenceof CB is insufficient to confer neuroprotection. The coex-istence of CB and GFRa-1 in DAergic neurons suggeststhat these two proteins might be part of a common neu-roprotective mechanism.
Interestingly, a previous study demonstrated thatGDNF treatment significantly increased the density ofCB- but not of CR-positive neurons in both rat and humanventral mesencephalic cultures (Meyer et al., 1999). Theseobservations suggest that midbrain neurons containingCB, which are responsive to GDNF, are likely to containGFRa-1, which is in agreement with our results showingthe coexistence of GFRa-1/CB in midbrain neurons. Inaddition to the presence or absence of CaBPs and GFRa-1,many other factors are likely to determine cell vulnerabil-ity. For instance, it has been found that mRNA for thedopamine transporter (DAT) is low in those DAergic cellsthat are spared from 1-methyl-4-phenyl-1,2,3,6-tetra-hydropyridine (MPTP)-induced degeneration in themouse. These findings indicate that the activity of DAT,which transports the MPTP toxin into the DA neurons,might play a role in determining which DA neurons arevulnerable to MPTP toxicity (Sanghera et., 1994).
Several studies have provided evidence for the neuro-protective and neuroregenerative effects of GDNF on DAneurons in animal models of PD (Stromberg et al., 1993;Hoffer et al., 1994; Beck et al., 1995; Gash et al., 1995;Bowenkamp et al., 1995; Winkler et al., 1996; Choi-Lundberg et al., 1997). Furthermore, in these animal mod-els GDNF reverses motor deficits by increasing DAergicneurotransmission (Hoffer et al., 1994). It is not clearwhether behavioral improvement induced by GDNFmight be also mediated by changes in GABAergic func-tions; however, the demonstration of GFRa-1 in midbrainGABAergic neurons suggests that GDNF might affect theGABAergic system in addition to the DAergic system.
ACKNOWLEDGMENTS
The authors thank Drs. Allan Tobin and Andreas Tomacfor providing original plasmids.
LITERATURE CITED
Arenas E, Trupp M, Akerud P, Ibanez CF. 1995. GDNF prevents degen-eration and promotes the phenotype of brain noradrenergic neurons invivo. Neuron 15:1465–1473.
Baimbridge KG, Celio MR, Rogers JH. 1992. Calcium-binding proteins inthe nervous system. TINS 15:303–308.
Beck KD, Valverde J, Alexi T, Poulsen K, Moffat B, Vandlen RA, RosenthalA, Hefti F. 1995. Mesencephalic dopaminergic neurons protected byGDNF from axotomy-induced degeneration in the adult brain. Nature373:339–341.
Beckstead RM, Domesick VB, Nauta WJ. 1979. Efferent connections of thesubstantia nigra and ventral tegmental area in the rat. Brain Res175:191–217.
Bernheimer H, Birkmayer W, Hornykiewicz O, Jellinger K, Seitleberger F.1973. Brain dopamine and the syndromes of Parkinson and Hunting-ton. Clinical, morphological and neurochemical correlations. J NeurolSci 20: 415–455.
Bowenkamp KE, Hoffman AF, Gerhardt GA, Henry MA, Biddle PT, HofferBJ, Granholm AC. 1995. Glial cell line-derived neurotrophic factorsupports survival of injured midbrain dopaminergic neurons. J CompNeurol 355: 479–489.
Buj-Bello A, Buchman VL, Horton A, Rosenthal A, Davies AM. 1995.GDNF is an age-specific survival factor for sensory and autonomicneurons. Neuron 15:821–828.
Chevalier G, Thierry AM, Shibazaki T, Feger J. 1981. Evidence for aGABAergic inhibitory nigrotectal pathway in the rat. Neurosci Lett21:67–70.
Choi-Lundberg DL, Lin Q, Chang YN, Chiang YL, Hay CM, Mohajeri H,Davidson BL, Bohn MC. 1997. Dopaminergic neurons protected fromdegeneration by GDNF gene therapy. Science 7:838–841.
Creedon DJ, Tansey MG, Baloh RH, Osborne PA, Lampe PA, Fahrner TJ,Heuckeroth RO, Milbrandt J, Johnson EM Jr. 1997. Neurturin sharesreceptors and signal transduction pathways with glial cell line-derivedneurotrophic factor in sympathetic neurons. Proc Natl Acad Sci USA94:7018–7023.
Durbec P, Marcos-Gutierrez CV, Kilkenny C, Grigorious M, WartiowaaraK, Suvanto P, Smith D, Ponder B, Costantini F, Saarma M, Sariola H,Pachnis V. 1996. GDNF signaling through the Ret receptor tyrosinekinase. Nature 381:789–792.
Gash DM, Zhang Z, Cass WA, Ovadia A, Simmerman L, Martin D, RussellD, Collins F, Hoffer BJ, Gerhardt GA. 1995. Morphological and func-tional effects of intranigrally administered GDNF in normal rhesusmonkeys. J Comp Neurol 363:345–358.
Gerfen CR, Baimbridge KG, Miller JJ. 1985. The neostriatal mosaic: com-partmental distribution of calcium-binding proteins and parvalbuminin the basal ganglia of the rat and monkey. Proc Natl Acad Sci USA82:8780–8784.
Gerfen CR, Herkenham M, Thibault J. 1987. The neostriatal mosaic: II.Patch- and matrix-directed mesostriatal dopaminergic and non-dopaminergic systems. J Neurosci 7:3915–3934.
116 A. SARABI ET AL

German DC, Manaye KF, Sonsalla PK, Brooks BA. 1992. Midbrain dopa-minergic cell loss in Parkinson’s disease and MPTP-induced parkin-sonism: sparing of calbindin-D28k-containing cells. Ann NY Acad Sci648:42–62.
Hardman CD, McRitchie DA, Halliday GM, Cartwright HR, Morris JGL.1996. Substantia nigra pars reticulata neurons in Parkinson’s disease.Neurodegeneration 5:49–55.
Henderson CE, Phillips HS, Pollock RA, Davies AM, Lemeulle C, ArmaniniM, Simmons L, Moffet B, Vandlen RA, Koliatsos VE, Rosenthal A.1994. GDNF: a potent survival factor for motoneurons present in pe-ripheral nerve and muscle. Science 266:1062–1064.
Herkenham M, Edley SM, Stuart J. 1984. Cell clusters in the nucleusaccumbens of the rat, and the mosaic relationship of opiate receptors,acetylcholinesterase and subcortical afferent termination. Neuro-science 11:561–593.
Hirsch EC. 1994. Biochemistry of Parkinson’s disease with special refer-ence to the dopaminergic systems. Mol Neurobiol 9:134–142.
Hoffer BJ, Hoffman A, Bowenkamp K, Huettl P, Hudson J, Martin D, LinL-FH, Gerhardt GA. Glial cell line-derived neurotrophic factor reversestoxin-induced injury to midbrain dopaminergic neurons in vivo. 1994.Neurosci Lett 182:107–111.
Hontanilla B, Parent A, Gimenez-Amaya JM. 1997. Parvalbumin andcalbindin D-28k in the entopeduncular nucleus, subthalamic nucleus,and substantia nigra of the rat as revealed by double-immunohistochemical methods. Synapse 25:359–367.
Hornykiewicz O, Kish SJ. 1986. Biochemical pathophysiology of Parkin-son’s disease. Adv Neurol 45:19–34.
Iacopino A, Chritakos S, German D, Sonsalla PK, Altar CA. 1992.Calbindin-D28k-containing neurons in animal models of neurodegen-eration: possible protection from excitotoxicty. Mol Brain Res 13:251–261.
Jing S, Wen D, Yu Y, Holst PL, Luo Y, Fang M, Tamir R, Antonio L, Hu Z,Cupples R, Louis J-C, Hu S, Altrock BW, Fox GM. 1996. GDNF-inducedactivation of the rat protein tyrosine kinase is mediated by GDNFR-aa novel receptor for GDNF. Cell 85:1113–1124.
Kiyama H, Emson PC, Ruth J, Morgan C. 1990. Sensitive non-radioisotopicin situ hybridization histochemistry: demonstration of tyrosine hydrox-ylase gene expression in the rat brain and adrenal. Mol Brain Res7:213–219.
Kobayashi K, Emson PC, Mountjoy CQ, Thornton SN, Lawson DE, MannDM. 1990. Cerebral cortical calbindin D28k and parvalbumin neuronsin Down’s syndrome. Neurosci Lett 18:17–22.
Kordower JH, Emborg ME, Bloch J, Ma SY, Chu Y, Leventhal L, McBrideJ, Chen EY, Palfi S, Roitberg BZ, Brown WD, Holden JE, Pyzalski R,Taylor MD, Carvey P, Ling Z, Trono D, Hantraye P, Deglon N, Aebi-scher P. 2000. Neurodegeneration prevented by lentiviral vector deliv-ery of GDNF in primate models of Parkinson’s disease. Science 290:767–773.
Lavoie B, Parent A. 1991. Dopaminergic neurons expressing calbindin innormal and parkinsonian monkeys. Neuroreport 2:601–604.
Li L, Wu W, Lin LF, Lei M, Oppenheim RW, Houenou LJ. 1995. Rescue ofadult mouse motoneurons from injury-induced cell death by glial cellline-derived neurotrophic factor. Proc Natl Acad Sci USA 92:9771–9775.
Liang CL, Sinton CM, German DC. 1996. Midbrain dopaminergic neuronsin the mouse: co-localization with calbindin-D28k and calretinin. Neu-roscience 75:523–533.
Lin LF, Doherty DH, Lile JD, Bektesh S, Collins F. 1993. GDNF: a glial cellline-derived neurotrophic factor for midbrain dopaminergic neurons.Science 260:1130–1132.
McRitchie DA, Hardman CD, Halliday GM. 1996. Cytoarchitectural dis-tribution of calcium binding proteins in midbrain dopaminergic regionsof rats and humans. J Comp Neurol 368:121–150.
Meyer M, Zimmer J, Seiler RW, Widmer HR. 1999. GDNF increases thedensity of cells containing calbindin but not of cells containing calreti-nin in culture rat and human fetal nigral tissue. Cell Transplant8:25–36.
Morales M, Bloom FE. 1997. The 5-HT3 receptor is present in differentsubpopulation of GABAergic neurons in the rat telencephalon. J Neu-rosci 17:3157–3167.
Mouatt-Prigent A, Agid Y, Hirsch EC. 1994. Does the calcium bindingprotein calretinin protect dopaminergic neurons against degenerationin Parkinson’s disease?. Brain Res 668:62–70.
Mount HTJ, Dean DO, Alberch J, Dreyfus CF, Black IB. 1995. Glial cellline-derived neurotrophic factor promotes the survival and morphologicdifferentiation of purkinje cells. Proc Natl Acad Sci USA 92:9092–9096.
Nemoto C, Hida T, Arai R. 1999. Calretinin and calbindin-D28k in dopa-minergic neurons of the rat midbrain:a triple-labeling immunohisto-chemical study. Brain Res 846:129–136.
Nosrat CA, Tomac A, Hoffer BJ, Olson L. 1997. Cellular and patterns ofexpression of Ret and glial cell line-derived neurotrophic factor recep-tor alpha mRNAs. Exp Brain Res 115:410–422.
Parent A, Fortin M, Cote P-Y, Cicchatti F. 1996. Calcium-binding proteinsin primate basal ganglia. Neurosci Res 25:309–334.
Paxinos G, Watson C. 1998. The rat brain in stereotaxic coordinates.Orlando: Academic Press.
Pong K, Xu RY, Baron WF, Louis JC, Beck KD. 1998. Inhibition of phos-phatidylinostitol 3-kinase activity blocks cellular differentiation medi-ated by glial cell line-derived neurotrophic in dopaminergic neurons.J Neurochem 71:1912–1919.
Poteryaev D, Titievsky A, Sun YF, Thomas-Crusells J, Lindahl, M, BillaudM, Arumae U, Saarma M. 1999. GDNF triggers a novel ret-independent src kinase family-coupled signaling via a GPI-linkedGDNF receptor alpha 1. FEBS Lett 463:63–66.
Price ML, Hoffer BJ, Granholm AC. 1996. Effect of GDNF on fetal septalforebrain transplant in ocula. Exp Neurol 141:181–189.
Rogers JH, Resibois A. 1992. Calretinin and calbindin-D28k in rat brain:patterns of partial co-localization. Neuroscience 51:843–865.
Sanghera MK, Manaye KF, Liang CL, Lacopino AM, Bannon MJ, GermanDC. 1994. Low dopamine transporter mRNA levels in midbrain regionscontaining calbindin. Neuroreport 5:1641–1644.
Sarabi A, Hoffer BJ, Olson L, Morales M. 2000. GFRa-1 is expressed inparvalbumin GABAergic neurons in the hippocampus. Brain Res 877:262–270.
Stromberg I, Bjorklund L, Johansson M, Tomac A, Collins F, Olson L,Hoffer BJ, Humpel C. 1993. Glial cell line-derived neurotrophic factoris expressed in the developing but not adult striatum and stimulatesdeveloping dopamine neurons in vivo. Exp Neurol 124:401–412.
Swanson LW. 1982. The projections of the ventral tegmental area andadjacent regions: a combined fluorescent retrograde tracer and immu-nofluorescence study in the rat. Brain Res Bull 9:321–353.
Treanor JJ, Goodman L, De Sauvage F, Stone DM, Poulsen KT, Beck CD,Gray C, Armanini MP, Pollock RA, Hefti F, Phillips HS, Goddard A,Moore MW, Buj-Bello A, Davies AM, Asai N, Takahashi M, Vandlen R,Henderson CE, Rosenthal A. 1996. Characterization of a multicompo-nent receptor for GDNF. Nature 382:80–83.
Trupp M, Arenas E, Fainzilber M, Nilsson AS, Sieber BA, Grigoriou M,Kilkenny C, Salazar-Grueso E, Pachnis V, Arumae U. 1996. Functionalreceptor for GDNF encoded by the c-ret proto-oncogene. Nature 381:785–789.
Trupp M, Belluardo N, Funakoshi H, Ibanez CF. 1997. Complementaryand overlapping expression of glial cell line-derived neurotrophic factor(GDNF), c-ret proto-oncogene, and GDNF receptor-a indicates multiplemechanisms of trophic action in the adult rat CNS. J Neurosci 17:3554–3567.
Trupp M, Scott R, Whittemore SR, Ibanez CF. 1999. Ret-dependent andindependent mechanisms of glial cell line-derived neurotrophic factorsignaling in neuronal cells. J Biol Chem 274:20885–20894.
Van Weering DH, Bos JL. 1997. Glial cell line-derived neurotrophic factorinduces Ret-mediated lamellipodia formation. J Biol Chem 272:249–254.
Winkler C, Sauer H, Lee CS, Bjorklund A. 1996. Short-term GDNF treat-ment provides long-term rescue of lesioned nigral dopaminergic neu-rons in a rat model of Parkinson’s disease. J Neurosci 16:7206–7215.
Wright AK, Arbuthnott GW. 1981. The pattern of innervation of the corpusstriatum by the substantia nigra. Neuroscience 6:2063–2067.
Yamada T, McGeer PL, Baimbridge KG, McGeer EG. 1990. Relative spar-ing in Parkinson’s disease of substantia nigra dopamine neurons con-taining calbindin-D28k. Brain Res 526:303–307.
117GFRa-1 IN MIDBRAIN DA AND GABA NEURONS




















