
Neuroregenerative effects of lentiviral vector-mediated GDNF expression in reimplanted ventral roots
Upload
independentCategory
view
1download
0
ARCHIVAL REPORT
Ventral Tegmental Area Cannabinoid Type-1Receptors Control Voluntary Exercise Performance
Sarah Dubreucq, Audrey Durand, Isabelle Matias, Giovanni Benard, Elodie Richard,Edgar Soria-Gomez, Christelle Glangetas, Laurent Groc, Aya Wadleigh, Federico Massa,Dusan Bartsch, Giovanni Marsicano, Francois Georges, and Francis ChaouloffBackground: We have shown that the endogenous stimulation of cannabinoid type-1 (CB1) receptors is a prerequisite for voluntary running inmice, but the precise mechanisms through which the endocannabinoid system exerts a tonic control on running performance remain unknown.
Methods: We analyzed the respective impacts of constitutive/conditional CB1 receptor mutations and of CB1 receptor blockade onwheel-running performance. We then assessed the consequences of ventral tegmental area (VTA) CB1 receptor blockade on the wheel-running performances of wildtype (gamma-aminobutyric acid [GABA]-CB1
�/�) and mutant (GABA-CB1–/–) mice for CB1 receptors in brain
GABA neurons. Using in vivo electrophysiology, the consequences of wheel running on VTA dopamine (DA) neuronal activity wereexamined in GABA-CB1
�/� and GABA-CB1–/– mice.
Results: Conditional deletion of CB1 receptors from brain GABA neurons, but not from several other neuronal populations or fromastrocytes, decreased wheel-running performance in mice. The inhibitory consequences of either the systemic or the intra-VTAadministration of CB1 receptor antagonists on running behavior were abolished in GABA-CB1
–/– mice. The absence of CB1 receptors fromGABAergic neurons led to a depression of VTA DA neuronal activity after acute/repeated wheel running.
Conclusions: This study provides evidence that CB1 receptors on VTA GABAergic terminals exert a permissive control on rodentvoluntary running performance. Furthermore, it is shown that CB1 receptors located on GABAergic neurons impede negativeconsequences of voluntary exercise on VTA DA neuronal activity. These results position the endocannabinoid control of inhibitorytransmission as a prerequisite for wheel-running performance in mice.
Key Words: CB1 receptors, dopamine, GABA, physical exercise,ventral tegmental area, wheel running
Regular physical activity is beneficial to health, includingmental health (1–3). Although this finding is repeatedlypublicized, physical inactivity remains a major issue (4,5).
Physical inactivity is multifactorial, and the inability to experiencepleasure with exercise is considered one cause for the lack ofadherence to, or dropout from, exercise programs (6). Systemsgoverning reward-based motivation, such as the mesocortico-limbic dopamine (DA) system (7–9), might thus have an impacton voluntary exercise performance. Although voluntary wheelrunning in laboratory rodents models only certain aspects ofhuman adherence to exercise (10), it is worthy of mention thatwheel running has rewarding properties (10,11). This observationfits with the ability of wheel running to elicit temporary changesin DA neuronal activity in the ventral tegmental area (VTA) (12),the origin of the mesocorticolimbic DA system.
The endocannabinoid system (ECS), formed by cannabinoidtype-1 (CB1) and type-2 (CB2) receptors, their endogenous ligands
From the Institut National de la Sante et de la Recherche Medicale (INSERM)
U862 (SD, AD, IM, GB, ER, ES-G, AW, FM, GM, FC), NeuroCentre
Magendie, Physiopathologie de la Plasticite Neuronale; Centre National
de la Recherche Scientifique (CNRS) Unite Mixte de Recherche (UMR)
5297 (CG, LG, FG), Institut Interdisciplinaire de NeuroSciences; Universite
Bordeaux Segalen (SD, AD, IM, GB, ER, ES-G, CG, LG, AW, FM, GM, FG, FC),
Bordeaux, France; Department of Molecular Biology (DB), Central
Institute of Mental Health, Mannheim, Germany.
Authors GM, FG, and FC contributed equally to this work.
Address correspondence to Francis Chaouloff, PhD, Endocannabinoids and
NeuroAdaptation, INSERM U862, NeuroCentre Magendie, 146 rue Leo
Saignat, 33077 Bordeaux Cedex, France; E-mail: [email protected].
Received Jun 28, 2012; revised Oct 9, 2012; accepted Oct 26, 2012.
0006-3223/$36.00http://dx.doi.org/10.1016/j.biopsych.2012.10.025
(endocannabinoids), and the machinery for endocannabinoidsynthesis/degradation (13), controls wheel-running behavior.Thus, compared with wildtype littermates (CB1
�/�), mutant micelacking CB1 receptors (CB1
–/–) display a 30% to 50% decrease inwheel running (14,15), but not in locomotor (16), activity. Thequest for the mechanisms underlying this observation is complexgiven the plethora of ECS-regulated functions. CB1 receptors,which are mainly located on neuronal presynaptic terminals and onglial cells in numerous brain regions, are retrogradely stimulated byendocannabinoids released “on demand” from postsynaptic cells(13,17). This stimulation reduces the presynaptic release of severalneurotransmitters (e.g., gamma-aminobutyric acid [GABA], glutamate),providing to the ECS a key position in the regulation of centralnervous system functions (13,17–19). Several of these, including motorcontrol (20), metabolism (19,21), pain (22), mood (23), and rewardseeking (24,25), might be involved in running performance. As forreward seeking, the ECS, through CB1 receptors located on VTAGABAergic and glutamatergic terminals that synapse onto DA neurons(26,27), modulates mesocorticolimbic dopaminergic activity (27–31).Hence, the ECS controls the motivation to seek rewards (24,25,31), butwhether this rule includes voluntary running is unknown.
This study investigated the mechanisms underlying the involve-ment of CB1 receptors in wheel-running behavior. We show that CB1
receptors located on VTA GABAergic terminals exert a permissivecontrol on running performance. Moreover, the absence of CB1
receptors from GABAergic neurons is associated with inhibitoryconsequences of running on VTA DA neuronal activity, suggestingthat the ECS might play a pivotal role in voluntary exercise.
Methods and Materials
AnimalsAll experiments complied with the French and European rules
on animal experimentation. This study involved 2-month-old
BIOL PSYCHIATRY 2013;73:895–903& 2013 Society of Biological Psychiatry

896 BIOL PSYCHIATRY 2013;73:895–903 S. Dubreucq et al.
male C57BL/6N mice and 2- to 3-month-old male CB1 receptormutant and wildtype animals, including constitutive CB1 receptormutant mice (CB1
–/–), conditional mutant mice lacking CB1
receptors from principal neurons (CaMK-CB1–/–), brain GABAergic
neurons (GABA-CB1–/–), cortical glutamatergic neurons (Glu-CB1
–/–),serotonergic neurons (TPH2-CB1
–/–), dopamine D1 receptor-expressing neurons (D1-CB1
–/–), glial fibrillary acidic protein-expressing astrocytes (GFAP-CB1
–/–), and their respective wildtypelittermates (32–37) (see Methods in Supplement 1 for additionaldetails). The influence of Cre expression in the wheel-runningperformance of GABA-CB1
–/– mice was assessed by using Dlx5/6-Cre-positive and Dlx5/6-Cre-negative (wildtype) mice (33,35).Although Dlx5/6-Cre is thought to confer a forebrain-specificdeletion of CB1 receptors from GABAergic neurons (38) (Figure S1in Supplement 1), the possibility that CB1 receptors are deleted inother GABAergic neuronal populations cannot be excluded (seeDiscussion).
Wheel-Running ProtocolsOn arrival, all mice were housed with 25-cm diameter running
wheels (IntelliBio, Nomeny, France) (14) set free for limited/unlimited periods or blocked permanently (controls). The wheelswere connected to a computer that recorded all runningvariables (distance covered, running duration and maximal speed;ActiviWheel software, IntelliBio). These wheels were unlocked 3hours per day, except in one series of experiments (unlimitedaccess to the wheels). The 3-hour daily running episodes alwaysstarted at the onset of the dark phase (7:00 PM) of the 12-hourlight–dark cycle, excepted from several experiments in whichwheels were set free from 12:00 AM to 3:00 AM or from 4:00 AM to7:00 AM (see below and Methods in Supplement 1).
Injection and Infusion ProtocolsSystemic injections and surgery for bilateral cannulae implan-
tation always occurred when mice reached their plateau level ofperformance (sixth–seventh day). Intracerebral infusions wereperformed at least 1 week after surgery when mice displayedtheir presurgery body weight and wheel-running performances.Mice were injected/infused with the drug (or the vehicle) and 2 to3 days later with the vehicle (or the drug). In all cases, the effectsof drugs and vehicles on wheel-running performance were goneby the following day.
Drug Infusion in the VTAMice were anaesthetized by the intraperitoneal injection of a
mixture of ketamine/xylazine, and placed into a stereotaxicapparatus (David Kopf Instruments, Tujunga, California). Micewere bilaterally implanted with 2.7-mm stainless cannulae target-ing the VTA with the following coordinates: anteroposterior –3.0;lateral � .5; dorsoventral –4.7 (39). The cannulae were securedwith dental cement, and the mice were allowed to recover for atleast another week. Drugs were bilaterally infused over 2 min andthe injectors left in place for 1 min to allow further diffusion (seeMethods in Supplement 1).
Locomotor ActivityLocomotor activity was assessed in soundproof cubicles
hosting individual cages equipped with food, water, and infraredsensors allowing to detect horizontal crossings (Imetronic, Pessac,France) (35). All animals were housed therein 5 to 7 days beforetesting (see Methods in Supplement 1).
www.sobp.org/journal
In Vivo ElectrophysiologyMice were housed 1 day (acute running) or 1 to 3 weeks
(repeated running) with wheels that were either permanentlylocked (controls) or set free 3 hours per day (4:00–7:00 AM;runners). Electrophysiologic experiments, which started 3 to 4hours or 24 hours after the end of the running session, wereperformed as described previously (40,41) (see also Methods inSupplement 1).
DrugsRimonabant and JZL195 were from Caiman Chemical (Inter-
chim, Montluc-on, France). AM251, O-2050, JWH133, and AM630were from Tocris Bioscience (Bristol, United Kingdom). D9-tetra-hydrocannabinol (THC) was from Sigma-Aldrich (Saint QuentinFallavier, France). All drugs and their respective vehicles wereprepared and administered according to standard methods (seeMethods in Supplement 1).
StatisticsAll analyses were performed with GB-Stat software (version 10;
Dynamic Microsystems, Silver Spring, Maryland). Comparisonswere achieved by means of Student t tests (two-group compar-isons) and analyses of variance with/without repeated factors(multiple-group comparisons). In the latter case, post hoc groupcomparisons (Tukey’s test) were performed only if interactionsbetween main variables were found significant. When necessary,data were log-transformed to reach homogeneity of the variances.
Results
CB1 Receptors on Brain GABAergic Neurons ControlVoluntary Running
Mutant mice lacking CB1 receptors from the whole body(CB1
–/–) and their wildtype littermates (CB1�/�), bred as in
Dubreucq et al. (14), were offered 3-hour daily access to runningwheels for 2 weeks. This paradigm allowed observation ofplateau levels of performance in both genotypes (Figure 1A).CB1
–/– mice displayed low running activity, compared with CB1�/�
mice [F(1,35) ¼ 8.22, p ¼ .007], the amplitude of this differenceincreasing with the number of running sessions [F(13,455) ¼ 3.23,p ¼ .0001 for the genotype � time interaction] (Figure 1A).Repeated intraperitoneal injections (30 min before the onset ofwheel-running sessions on Days 1–8) with 3 mg/kg of the CB1
receptor antagonist rimonabant (SR141716) to C57BL/6N micemimicked the negative impact of the CB1 receptor mutation[F(13,130) ¼ 27.49, p � .0001; Figure 1B]. The inhibitory effect ofrimonabant was rapidly reversible and was not accounted for byan interaction with wheel-running acquisition. Thus, rimonabantalso decreased wheel running when acutely administered to micethat had reached their maximal daily performance (Figure 1B).Conversely, rimonabant administration to mice housed in cagesequipped with activity detectors did not affect locomotor activity(71.8 � 13.7 beam brakes/3 hours; n ¼ 5), compared with vehicletreatment (69.2 � 12.2 beam brakes/3 hours; n ¼ 5). To ensurethat CB1 receptors mediated rimonabant effects on running,rimonabant was administered to CB1
�/� and CB1–/– mice. Besides
genotype differences [F(1,12) ¼ 15.17, p ¼ .0021], this experimentrevealed that acute rimonabant reduced running performances inCB1
�/� mice but not in CB1–/– mice [F(1,12) ¼ 12.47, p ¼ .0041 for
the genotype � rimonabant interaction; Figure 1C].These results indicated that CB1 receptors exert a tonic
stimulatory influence on voluntary running. To identify the

Figure 1. CB1 receptors on gamma-aminobutyric acid (GABA)ergic neurons and wheel-running performance. (A) Distance ran by CB1�/� and CB1
–/– miceoffered daily 3-hour access to running wheels over 2 weeks (n ¼ 17 to 20 mice/genotype). (B) Respective effects of vehicle (V) and of the selective CB1
receptor antagonist rimonabant (R) on the daily distance covered by C57BL/6N mice (n ¼ 6 mice/group). (C) Time-dependent effects of acute vehicle andrimonabant administration 30 min before wheel access in CB1
�/� and CB1–/– mice (n ¼ 7 mice/genotype injected during the second week of wheel
running). (D) The deletion of CB1 receptors from GABA neurons (GABA-CB1–/–) reduced wheel-running performance (n ¼ 8 mice/genotype). (E) GABA-CB1
–/–
mice display decreased wheel-running performance whatever the period during which they are allowed to run for 3 hours during the dark phase of thelight–dark cycle (n ¼ 8–9 mice/genotype, except for the 4:00–07:00 AM period, which includes three mice/genotype. (F) Time-dependent effects of acutevehicle and rimonabant administration 30 min before wheel access in GABA-CB1
�/� and in GABA-CB1–/– mice (n ¼ 7–9 mice/genotype injected during the
second week of wheel running). p values refer to overall genotype influences. All data are expressed as mean � SEM. *p � .05 and **p � .01 forgenotypic differences, and �p � .05 and ��p � .01 for the differences between rimonabant and vehicle.
S. Dubreucq et al. BIOL PSYCHIATRY 2013;73:895–903 897
neuronal/glial subpopulations harboring these receptors, we nextused conditional mutant mice bearing a deletion of the CB1 genein distinct cellular populations. The deletion of the CB1 gene fromprincipal forebrain neurons (CaMK-CB1
–/– mice), from corticalglutamatergic neurons (Glu-CB1
–/– mice), from serotonergic neu-rons (TPH2- CB1
–/– mice), from DA type-1 (D1) receptor-expressingneurons (D1-CB1
–/– mice), or from glial fibrillary acidic protein-expressing astrocytes (GFAP-CB1
–/– mice) did not influence wheel-running performance (Figure S2A–S2E in Supplement 1). Incontrast, the deletion of the CB1 receptor gene from brainGABAergic neurons (GABA-CB1
–/– mice), reduced running distanceby 25% to 30% [F(1,14) ¼ 8.19, p ¼ .0126; Figure 1D]. Such adifference extended to running duration [F(1,14) ¼ 11.41, p ¼.0045; Figure S3A in Supplement 1], but not to maximal runningspeed (Figure S3B in Supplement 1), compared to GABA-CB1
�/�
littermates. The phenotype of GABA-CB1–/– mice was not influ-
enced by the time period during which mice ran (Figure 1E),whereas the single presence of the recombinase (Dlx5/6-Cre)used to obtain GABA-CB1
–/– mice proved ineffective on runningbehavior (Figure S2F in Supplement 1). To ensure that CB1
receptors on brain GABAergic neurons were involved in theinhibitory impact of rimonabant on running, the effects ofrimonabant on wheel-running activity were measured in GABA-CB1
�/� and GABA-CB1–/– mice. The temporal analysis of these
effects revealed genotype-dependent influences [F(1,14) ¼ 13.42,p ¼ .0026], the antagonist bearing inhibitory impacts in GABA-CB1
�/� mice, but not in GABA-CB1–/– mice (Figure 1F). An analysis
over the 3-hour running period confirmed this observation[F(1,14) ¼ 6.73, p ¼ .021]; rimonabant triggered a 28% decreasein running distance in GABA-CB1
�/� mice (2.31 � .12 and1.65 � .2 km in vehicle- and rimonabant-injected mice, respec-tively; n ¼ 9 and p � .05) but was ineffective in GABA-CB1
–/– mice(1.33 � .18 and 1.31 � .20 km in vehicle- and rimonabant-injected mice, respectively; n ¼ 7). These results indicate thatrimonabant-induced reduction in wheel-running performance(Figure 1B,C) was fully accounted for by the blockade of CB1
receptors on GABAergic neurons. In keeping with our initialobservation that CB1
–/– mice differ from CB1�/� mice when offered
unlimited access to the running wheels (14), we asked whetherGABA-CB1
�/� mice would differ from GABA-CB1–/– mice under this
setting. GABA-CB1–/– mice ran less than GABA-CB1
�/� mice whenprovided free access to the wheels [F(1,24) ¼ 16.63, p ¼ .0004;Figure 2A], a difference fully accounted for by a reduction inrunning during the active period of the light–dark cycle [F(1,24)¼ 19.85, p ¼ .0002; Figure 2B]. This result led us to examinewhether the two genotypes differ in their nocturnal locomotoractivities. Daily locomotor patterns were actually identical inGABA-CB1
�/� and GABA-CB1–/– mice housed in cages equipped
with activity detectors (Figure 2C). Lastly, assays of soleus musclemitochondrial respiration (Methods in Supplement 1) indicatedthat muscular mitochondrial oxygen consumption under stimu-lated conditions did not differ between sedentary GABA-CB1
�/�
and GABA-CB1–/– mice (7.26 � .83 ng atom/min/mg fibers and
7.37 � .61 ng atom/min/mg fibers, respectively; n ¼ 4).
www.sobp.org/journal
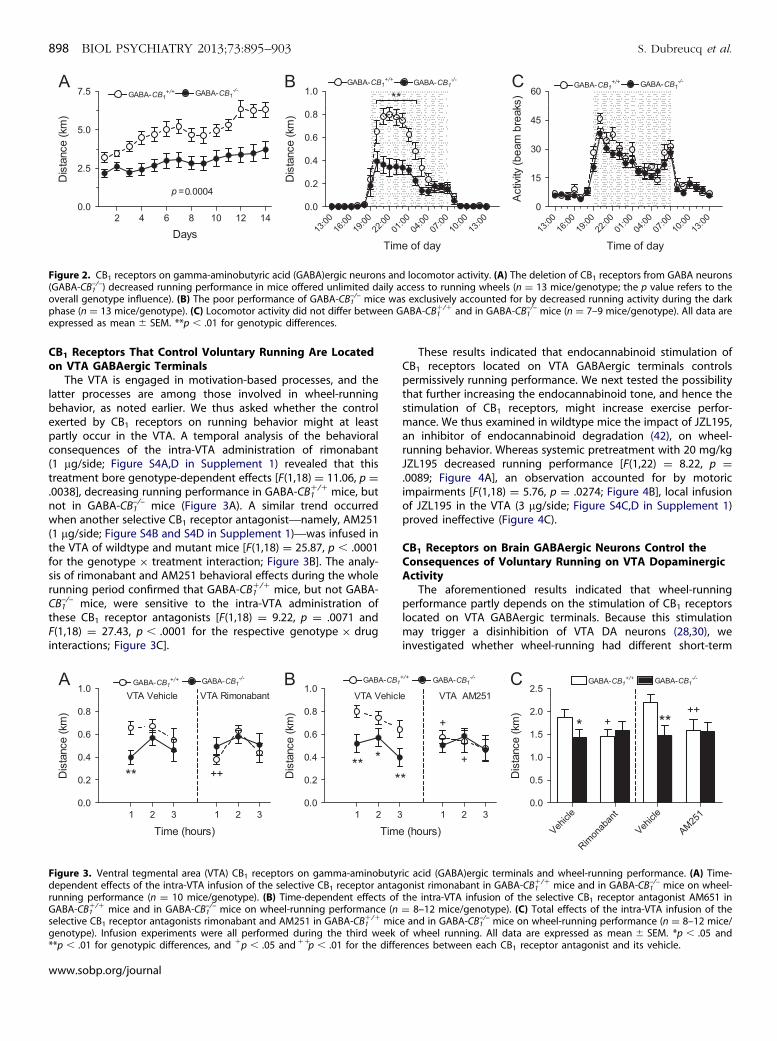
Figure 2. CB1 receptors on gamma-aminobutyric acid (GABA)ergic neurons and locomotor activity. (A) The deletion of CB1 receptors from GABA neurons(GABA-CB1
–/–) decreased running performance in mice offered unlimited daily access to running wheels (n ¼ 13 mice/genotype; the p value refers to theoverall genotype influence). (B) The poor performance of GABA-CB1
–/– mice was exclusively accounted for by decreased running activity during the darkphase (n ¼ 13 mice/genotype). (C) Locomotor activity did not differ between GABA-CB1
�/� and in GABA-CB1–/– mice (n ¼ 7–9 mice/genotype). All data are
expressed as mean � SEM. **p � .01 for genotypic differences.
898 BIOL PSYCHIATRY 2013;73:895–903 S. Dubreucq et al.
CB1 Receptors That Control Voluntary Running Are Locatedon VTA GABAergic Terminals
The VTA is engaged in motivation-based processes, and thelatter processes are among those involved in wheel-runningbehavior, as noted earlier. We thus asked whether the controlexerted by CB1 receptors on running behavior might at leastpartly occur in the VTA. A temporal analysis of the behavioralconsequences of the intra-VTA administration of rimonabant(1 mg/side; Figure S4A,D in Supplement 1) revealed that thistreatment bore genotype-dependent effects [F(1,18) ¼ 11.06, p ¼.0038], decreasing running performance in GABA-CB1
�/� mice, butnot in GABA-CB1
–/– mice (Figure 3A). A similar trend occurredwhen another selective CB1 receptor antagonist—namely, AM251(1 mg/side; Figure S4B and S4D in Supplement 1)—was infused inthe VTA of wildtype and mutant mice [F(1,18) ¼ 25.87, p � .0001for the genotype � treatment interaction; Figure 3B]. The analy-sis of rimonabant and AM251 behavioral effects during the wholerunning period confirmed that GABA-CB1
�/� mice, but not GABA-CB1
–/– mice, were sensitive to the intra-VTA administration ofthese CB1 receptor antagonists [F(1,18) ¼ 9.22, p ¼ .0071 andF(1,18) ¼ 27.43, p � .0001 for the respective genotype � druginteractions; Figure 3C].
Figure 3. Ventral tegmental area (VTA) CB1 receptors on gamma-aminobutyrdependent effects of the intra-VTA infusion of the selective CB1 receptor antagrunning performance (n ¼ 10 mice/genotype). (B) Time-dependent effects ofGABA-CB1
�/� mice and in GABA-CB1–/– mice on wheel-running performance (n
selective CB1 receptor antagonists rimonabant and AM251 in GABA-CB1�/� mic
genotype). Infusion experiments were all performed during the third week**p � .01 for genotypic differences, and �p � .05 and ��p � .01 for the diffe
www.sobp.org/journal
These results indicated that endocannabinoid stimulation ofCB1 receptors located on VTA GABAergic terminals controlspermissively running performance. We next tested the possibilitythat further increasing the endocannabinoid tone, and hence thestimulation of CB1 receptors, might increase exercise perfor-mance. We thus examined in wildtype mice the impact of JZL195,an inhibitor of endocannabinoid degradation (42), on wheel-running behavior. Whereas systemic pretreatment with 20 mg/kgJZL195 decreased running performance [F(1,22) ¼ 8.22, p ¼.0089; Figure 4A], an observation accounted for by motoricimpairments [F(1,18) ¼ 5.76, p ¼ .0274; Figure 4B], local infusionof JZL195 in the VTA (3 mg/side; Figure S4C,D in Supplement 1)proved ineffective (Figure 4C).
CB1 Receptors on Brain GABAergic Neurons Control theConsequences of Voluntary Running on VTA DopaminergicActivity
The aforementioned results indicated that wheel-runningperformance partly depends on the stimulation of CB1 receptorslocated on VTA GABAergic terminals. Because this stimulationmay trigger a disinhibition of VTA DA neurons (28,30), weinvestigated whether wheel-running had different short-term
ic acid (GABA)ergic terminals and wheel-running performance. (A) Time-onist rimonabant in GABA-CB1
�/� mice and in GABA-CB1–/– mice on wheel-
the intra-VTA infusion of the selective CB1 receptor antagonist AM651 in¼ 8–12 mice/genotype). (C) Total effects of the intra-VTA infusion of thee and in GABA-CB1
–/– mice on wheel-running performance (n ¼ 8–12 mice/of wheel running. All data are expressed as mean � SEM. *p � .05 andrences between each CB1 receptor antagonist and its vehicle.
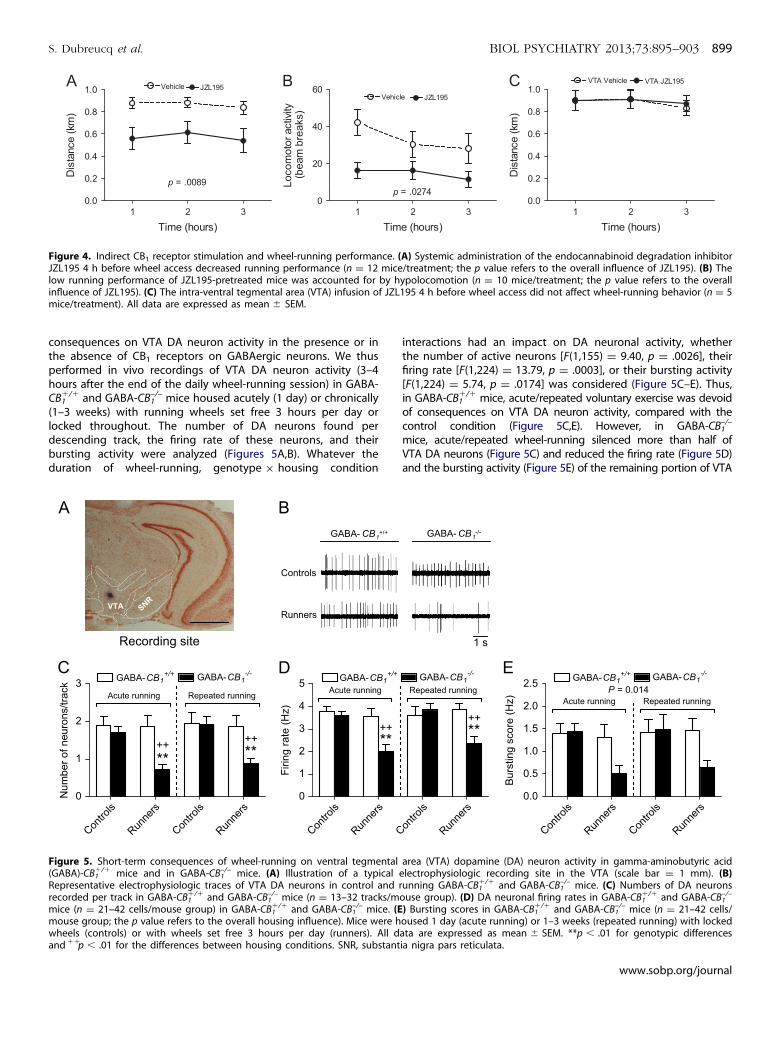
Figure 4. Indirect CB1 receptor stimulation and wheel-running performance. (A) Systemic administration of the endocannabinoid degradation inhibitorJZL195 4 h before wheel access decreased running performance (n ¼ 12 mice/treatment; the p value refers to the overall influence of JZL195). (B) Thelow running performance of JZL195-pretreated mice was accounted for by hypolocomotion (n ¼ 10 mice/treatment; the p value refers to the overallinfluence of JZL195). (C) The intra-ventral tegmental area (VTA) infusion of JZL195 4 h before wheel access did not affect wheel-running behavior (n ¼ 5mice/treatment). All data are expressed as mean � SEM.
S. Dubreucq et al. BIOL PSYCHIATRY 2013;73:895–903 899
consequences on VTA DA neuron activity in the presence or inthe absence of CB1 receptors on GABAergic neurons. We thusperformed in vivo recordings of VTA DA neuron activity (3–4hours after the end of the daily wheel-running session) in GABA-CB1
�/� and GABA-CB1–/– mice housed acutely (1 day) or chronically
(1–3 weeks) with running wheels set free 3 hours per day orlocked throughout. The number of DA neurons found perdescending track, the firing rate of these neurons, and theirbursting activity were analyzed (Figures 5A,B). Whatever theduration of wheel-running, genotype � housing condition
Recording site
SNRVTA
0
1
2
3
4
5Acute running
GABA-CB
Firin
g ra
te (H
z)
GABA-CB
Controls
Runners
Contro
ls
Contro
ls
Runne
rs
Runne
rs
Contro
ls
Runne
rs0
1
2
3Acute running Repeated running
GABA-CB GABA-CB
Num
ber o
f neu
rons
/trac
k
Figure 5. Short-term consequences of wheel-running on ventral tegmental(GABA)-CB1
�/� mice and in GABA-CB1–/– mice. (A) Illustration of a typical
Representative electrophysiologic traces of VTA DA neurons in control and rrecorded per track in GABA-CB1
�/� and GABA-CB1–/– mice (n ¼ 13–32 tracks/m
mice (n ¼ 21–42 cells/mouse group) in GABA-CB1�/� and GABA-CB1
–/– mice. (Emouse group; the p value refers to the overall housing influence). Mice were hwheels (controls) or with wheels set free 3 hours per day (runners). All dand ��p � .01 for the differences between housing conditions. SNR, substant
interactions had an impact on DA neuronal activity, whetherthe number of active neurons [F(1,155) ¼ 9.40, p ¼ .0026], theirfiring rate [F(1,224) ¼ 13.79, p ¼ .0003], or their bursting activity[F(1,224) ¼ 5.74, p ¼ .0174] was considered (Figure 5C–E). Thus,in GABA-CB1
�/� mice, acute/repeated voluntary exercise was devoidof consequences on VTA DA neuron activity, compared with thecontrol condition (Figure 5C,E). However, in GABA-CB1
–/–
mice, acute/repeated wheel-running silenced more than half ofVTA DA neurons (Figure 5C) and reduced the firing rate (Figure 5D)and the bursting activity (Figure 5E) of the remaining portion of VTA
Repeated runningGABA-CB
GABA-CB
1 s
Contro
ls
Runne
rs0.0
0.5
1.0
1.5
2.0
2.5 GABA-CB GABA-CB
Acute running Repeated runningP = 0.014
Bur
stin
g sc
ore
(Hz)
Contro
ls
Contro
ls
Runne
rs
Runne
rs
area (VTA) dopamine (DA) neuron activity in gamma-aminobutyric acidelectrophysiologic recording site in the VTA (scale bar ¼ 1 mm). (B)unning GABA-CB1
�/� and GABA-CB1–/– mice. (C) Numbers of DA neurons
ouse group). (D) DA neuronal firing rates in GABA-CB1�/� and GABA-CB1
–/–
) Bursting scores in GABA-CB1�/� and GABA-CB1
–/– mice (n ¼ 21–42 cells/oused 1 day (acute running) or 1–3 weeks (repeated running) with lockedata are expressed as mean � SEM. **p � .01 for genotypic differencesia nigra pars reticulata.
www.sobp.org/journal

900 BIOL PSYCHIATRY 2013;73:895–903 S. Dubreucq et al.
DA neurons. A 24-hour withdrawal from running was sufficient toreduce the proportion of silent DA neurons (38%; Figure S5A inSupplement 1) and to abolish running-induced reductions in VTADA neuronal firing rate and bursting activity (Figure S5B,C inSupplement 1) in 1-week running GABA-CB1
–/– mice.
Discussion
Mutant mice lacking CB1 receptors have been reported tospend less time running and to run shorter distances than theirwildtype littermates when housed with running wheels (14,15). Inthese studies, however, the animals had unlimited access to thewheels, thus questioning the role of CB1 receptors during shorterrunning activities. To solve this issue, we used a paradigm inwhich mice had daily 3-hour access to exercise. This exerciseduration was selected because it is sufficient to observe hippo-campal neurogenesis (43), a landmark of wheel running (44). Ourresults indicate that the inhibitory impact of the CB1 mutation onunrestricted voluntary running (14,15) extends to this restrictedexercise paradigm. Because mice needed 6 to 7 wheel-runningsessions to reach their maximal running performance, we con-sidered whether CB1 receptors might be involved in the acquisi-tion/learning phase of the wheel-running procedure. To addressthis point, C57BL/6N mice were pretreated with the CB1 receptorantagonist rimonabant either during the learning phase or there-after. Whatever the treatment day, rimonabant administration hada negative and reversible impact on wheel-running performance,an effect that was independent from motor-impairing effects.Rimonabant was ineffective in CB1 receptor mutant mice, indicat-ing that CB1 receptor stimulation is a prerequisite for wheel-running performance at each step of the running program (seealso Keeney et al. [45]). Moreover, the reductions in wheel runningafter systemic administration of rimonabant were also observed inmice of the CD1 strain (Figure S6 in Supplement 1), indicating thatour findings are not restricted to the C57BL/6N mouse strain. In thiscontext, it is noteworthy that most CB1 receptor antagonists(including rimonabant and AM251) bear CB1 inverse agonist proper-ties (46) that might have an impact on running behavior. This isunlikely, however, because the acute systemic administration ofO-2050, a neutral CB1 receptor antagonist with CB2 receptor ligandproperties (47,48) (but see Wiley et al. [49]), but not that witheffective concentrations (50) of either the CB2 receptor agonistJWH133 (46) or the CB2 receptor antagonist AM630 (46), decreasedrunning performance (Figure S7 in Supplement 1).
CB1 receptors are expressed in different neuronal (and glial)cell populations throughout the CNS (51,52), thus renderingcomplex the identification of the population(s) of CB1 receptorsinvolved in the tonic control of voluntary running. One means tocircumvent this task is the use of conditional mutagenesis inspecific cell populations. This approach has led us to identify thecell populations through which CB1 receptors regulate severalneuronal and behavioral functions (32–37). Herein, conditionalmutagenesis allowed us to exclude the hypothesis that the CB1
receptor population controlling wheel-running behavior waslocated on principal brain neurons, on cortical glutamatergicneurons, on serotonergic neurons, on D1 receptor-expressingneurons, or on GFAP-expressing astrocytes. In contrast, weobserved that wheel-running performance was reduced in micelacking CB1 receptors in brain GABAergic neurons (GABA-CB1
–/–
mice) (33,34). This observation was not accounted for by 1) thepresence of Dlx5/6-Cre, that is, the recombinase used to generateGABA-CB1
–/– mice (33,34), 2) an unexpected deletion of the CB1
www.sobp.org/journal
gene from peripheral tissues (Figure S1 in Supplement 1), or 3)alterations in basal locomotor activity. Indeed, the inability of theCB1 receptor antagonist rimonabant to decrease wheel-runningperformance in GABA-CB1
–/– mice indicated that the controlexerted by the ECS on wheel-running behavior is fully mediatedby CB1 receptors on GABAergic neurons. It is noteworthy that thecontrol of running performance exerted by CB1 receptors onGABAergic neurons extended to mice provided unrestrictedaccess to the wheels. These series of experiments also revealedthat GABA-CB1
�/� and GABA-CB1–/– mice had similar circadian
running and activity profiles. This may be surprising in view ofthe role exerted by CB1 receptors located on suprachiasmaticnucleus GABAergic terminals on circadian rhythms (53). However,these data were obtained by means of acute pharmacologicexperiments using CB1 receptor agonists (53), whereas theconsequences of CB1 receptor blockade on circadian rhythmsare still under debate (53,54). Nevertheless, we cannot excludethe possibility that our mice developed compensatory mechan-isms that masked the impact, if there was any, of the CB1
receptor mutation in GABAergic neurons on circadian rhythms.Central CB1 receptors are involved in the regulation of
numerous processes that could impact on running performance(discussed earlier). Because 1) wheel running is rewarding (55–58), 2) the ECS regulates reward-directed processes (24,25), and 3)rimonabant reduces the reinforcing properties of wheel runningin rats (58), we privileged first the hypothesis that CB1 receptorscontrol wheel-running performance by acting on reward-directedprocesses. The regulation of these processes by CB1 receptorsinvolves mainly the retrograde modulation of the excitatory andinhibitory inputs controlling mesocorticolimbic dopaminergicactivity (24,25), including in the VTA (30). Thus, VTA DA neurons,which are the targets of drugs of abuse and natural rewards(7–9), likely do not express CB1 receptors but do host the machineryallowing the synthesis/release of endocannabinoids to occur afterneuronal depolarization (26). Following their evoked release from DAcells, endocannabinoids stimulate in a retrograde manner CB1
receptors located on VTA GABAergic terminals and/or glutamatergicterminals (27–31). The resulting effect of endocannabinoid stimula-tion on the excitability of mesocorticolimbic DA neurons (disinhibi-tion or inhibition) depends on the levels of activation of therespective afferent pathways and thus on the nature of theenvironmental situation triggering endocannabinoid release (30).As an illustration, conditioned motivation to seek for food ispositively regulated by endocannabinoids in the VTA, likely througha disinhibition of VTA DA neurons (31). The report that wheelrunning targets the mesocorticolimbic DA system (12,57,59)prompted us to investigate the role of VTA CB1 receptors inwheel-running performance. Indeed, the intra-VTA perfusion ofrimonabant or AM251 reduced wheel-running performance inGABA-CB1
�/� mice, but not in GABA-CB1–/– mice, indicating that CB1
receptors on VTA GABAergic terminals control permissively wheel-running performance.
The pharmacologic and genetic tools used in this studysuggest that such a permissive impact of CB1 receptors accountsfor 25% to 30% of wheel-running performance. This result raisesseveral issues, among which is the identification of the source ofthese endocannabinoid-responsive VTA GABAergic terminals. Asindicated earlier, there is extensive evidence for the presence ofCB1 receptors on VTA GABAergic terminals, but the identity ofthese GABAergic neurons has not been defined yet. The VTAreceives major extrinsic inhibitory inputs from D1 receptor-expressing ventrostriatal medium spiny neurons and from ven-tropallidal neurons (60,61). Neither CaMK-CB1
–/– mice nor D1-CB1–/–

S. Dubreucq et al. BIOL PSYCHIATRY 2013;73:895–903 901
mice displayed alterations in running performances, suggestingthat the CB1 receptor-dependent control of voluntary runningdoes not involve these neuronal populations; however, wecannot exclude that the early deletion of CB1 receptors led tocompensatory mechanisms in these two mutant lines. In additionto the aforementioned extrinsic inhibitory inputs, 20% of VTAneurons in mice are local GABAergic interneurons (62), whichcould be endocannabinoid-responsive. In the embryonic brain,Dlx5/6 genes are expressed in forebrain GABAergic neuronswhere they control their migration and differentiation (38).Accordingly, our Dlx5/6-Cre mice are expected to bear anexclusive recombination of lox P sites in this forebrain neuronalpopulation (33,35), thus excluding VTA GABAergic neurons.However, the possibility remains that discrete neuronal subpo-pulations express Dlx5/6 genes out of the forebrain, including inthe midbrain VTA. This possibility is reinforced by a recenttransgenic study indicating that Dlx5/6 may be expressed in cellbodies of mouse midbrain (63) (Fu and Ess; written communica-tion; September 20, 2012). Moreover, transgenic expression ofthe Dlx1 gene in the cortex of mouse embryos leads to ectopicDlx5/6 expression in the ventral tegmentum (64), opening thepossibility that recombination events might have led to a partialloss of CB1 receptors in the VTA of our Dlx5/6-Cre mice. Futurein situ hybridization studies might help to examine this hypoth-esis. The second issue relates to the processes through which CB1
receptors exert a partial control on running performance.Although rimonabant has been reported to decrease the motiva-tion to perform wheel running in food-deprived rats (58), it isunknown whether such a process occurs under our experimentalsettings. One means to solve this issue requires the use ofoperant paradigms that assess the reinforcing properties ofwheel running (10,55–58), examining thereby whether GABA-CB1
�/� mice differ from GABA-CB1–/– mice in their motivation-
based processes. The third issue that merits consideration islinked to the extent to which the ECS has an impact on wheel-running performance. Systemic or intra-VTA administration of aCB1 receptor antagonist reduce conditioned food intake, indicat-ing that endocannabinoid stimulation of CB1 receptors on VTAGABAergic terminals is permissive to that behavior (31). Theamplification of such a stimulation by means of an inhibitor ofthe degradation of one principal endocannabinoid (namely, 2-arachidonoylglycerol) increases conditioned feeding (31). Theseresults indicate that the control exerted by the ECS in the VTA onthat behavior is both permissive and facilitatory. Herein, theamplification of the endocannabinoidergic tone by JZL195 (aninhibitor of the degradation pathways of the principal endocan-nabinoids 2-arachidonoylglycerol and anandamide) (42) failed toincrease running performance whether JZL195 was administeredin the VTA or systemically. The additional observation that thesystemic administration of the CB1/CB2 receptor agonists WIN55212 (65) and THC (Figure S8 in Supplement 1) failed to increasewheel-running performance indicates that the ECS exerts only apermissive control on voluntary running performance. Whetherthis observation is accounted for by the fact that the animalshave already reached their maximal running capacities is onepossibility. Finally, we believe that our knowledge of exerciseneurobiology, at least in rodents, would be improved by theidentification of the GABA receptor subtype (GABAA, GABAB)located on VTA DA neurons that is involved in the weak runningperformances of rimonabant/AM251-treated mice and GABA-CB1
–/– mice.It is likely that the CB1 receptor-dependent modulation of
voluntary exercise performance actually involves a disinhibition
of VTA DA neurons (30). However, our in vivo electrophysiologicdata bring evidence that acute/repeated wheel-running has noconsequences on VTA DA activity in control (i.e., wildtype) micewhen assessed 3 to 4 hours after the last running episode. Thisnegative finding reinforces the observation that VTA DA neuronalactivity is only temporarily increased by running activity, being soat the onset and at the offset of running activity (12). However, ifCB1 receptors are removed from GABA neurons, wheel-runningbears a negative impact on VTA DA activity, as revealed by thesilencing of half of VTA DA neurons and the reduction of theactivity of the second half of this neuronal population. Theseresults indicate that the tonic control exerted by CB1 receptors oninhibitory transmission is a prerequisite for the maintenance ofVTA DA neuron activity after acute and repeated voluntaryrunning. Thus, in the absence of these CB1 receptors, inhibitoryinputs on VTA DA neurons prevail over excitatory ones afterrunning. The possibility that these CB1 receptors are located onVTA GABAergic terminals cannot be addressed at presentbecause of methodologic limits, including the inability to performlocal drug infusion and electrophysiologic recordings in the VTAof the same animal.
In conclusion, this study reveals the key role exerted by theECS in the control of wheel-running performance. However, theextent to which our findings apply to human exercise remains tobe explored. As noted earlier, the motivational grounds forwheel-running are diverse (10), indicating the need for cautionbefore assuming that wheel-running models human exercise and,hence, that CB1 receptors control human exercise performance.
This work was supported by Institut National de la Sante et de laRecherche Medicale (INSERM) (to GM), EU-FP7 (REPROBESITY, GrantNo. HEALTH-F2-2008-223713 to GM), European Research Council(ENDOFOOD, Grant No. ERC-2010-StG-260515 to GM), NationalAlliance for Research on Schizophrenia and Depression (2008Independent Investigator Award to GM), Fondation pour laRecherche sur le Cerveau (GM), l’Agence Nationale pour laRecherche (FG, LG), the Aquitaine Region (GM, LG), the FyssenFoundation (ES-G), la Delegation Generale pour l’Armement duMinistere de la Defense (SD), BundesMinisterium fur Bildung undForschung (BMBF) Bernstein Center for Computational NeuroscienceHeidelberg/Mannheim (Grant No. 01GQ1003A, DB), and BMBFNGFN-Plus Grant (Grant No. 01GS0851 to DB).
We thank all the personnel of the Animal Facility and of theMouse Genotyping Platform of the NeuroCentre Magendie for theirhelp throughout the study. The members of GM’s laboratory arethanked for helpful discussions.
The authors report no biomedical financial interests or potentialconflicts of interest.
Supplementary material cited in this article is available online.
1. Martinsen EW, Morgan WP (1997): Antidepressant effects of physicalactivity. In: Morgan WP, editor. Physical Activity & Mental Health.Washington, DC: Taylor & Francis, 93–106.
2. Raglin JS (1997): Anxiolytic effects of physical activity. In: Morgan WP,editor. Physical Activity & Mental Health. Washington, DC: Taylor &Francis, 107–126.
3. Salmon P (2001): Effects of physical exercise on anxiety, depression,and sensitivity to stress: A unifying theory. Clin Psychol Rev 21:33–61.
4. World Health Organization (2003): Annual Global Move for HealthInitiative: A Concept Paper. Geneva: World Health Organization; 1–13.
5. Institut National de la Sante et de la Recherche Medicale (2008):Activite physique—contextes et effets sur la sante. Paris: INSERM.
www.sobp.org/journal

902 BIOL PSYCHIATRY 2013;73:895–903 S. Dubreucq et al.
6. Dishman RK, Buckworth J (1997): Adherence to physical activity. In:Morgan WP, editor. Physical Activity & Mental Health. Washington, DC:Taylor & Francis, 63–80.
7. Wise RA (2004): Dopamine, learning and motivation. Nat Rev Neurosci5:1–12.
8. Nestler EJ (2005): Is there a common molecular pathway for addic-tion? Nat Neurosci 8:1445–1449.
9. Luscher C, Malenka RC (2011): Drug-evoked synaptic plasticity inaddiction: From molecular changes to circuit remodeling. Neuron 69:650–663.
10. Sherwin CM (1998): Voluntary wheel running: A review and novelinterpretation. Anim Behav 56:11–27.
11. Novak CM, Burghardt PR, Levine JA (2012): The use of a runningwheel to measure activity in rodents: Relationship to energy balance,general activity, and reward. Neurosci Biobehav Rev 36:1001–1014.
12. Wang DV, Tsien JZ (2011): Conjunctive processing of locomotorsignals by the ventral tegmental area neuronal population. Plos One 6:e16528.
13. Piomelli D (2003): The molecular logic of endocannabinoid signalling.Nat Rev Neurosci 4:873–884.
14. Dubreucq S, Koehl M, Abrous DN, Marsicano G, Chaouloff F (2010):CB1 receptor deficiency decreases wheel-running activity: Conse-quences on emotional behaviours and hippocampal neurogenesis.Exp Neurol 224:106–113.
15. Miller LL, Picker MJ, Schmidt KT, Dykstra LA (2011): Effects ofmorphine on pain-elicited and pain-suppressed behavior in CB1kncokout and wildtype mice. Psychopharmacology 215:455–465.
16. Chaouloff F, Dubreucq S, Bellocchio L, Marsicano G (2011): Endocan-nabinoids and motor behavior: CB1 receptors also control runningactivity. Physiology 26:76–77.
17. Ohno-Shosaku T, Tanimura A, Hashimotodani Y, Kano M (2012):Endocannabinoids and retrograde modulation of synaptic transmis-sion. Neuroscientist 18:119–132.
18. Marsicano G, Lutz B (2006): Neuromodulatory functions of theendocannabinoid system. J Endocrinol Invest 29:27–46.
19. Pacher P, Batkai S, Kunos G (2006): The endocannabinoid system asan emerging target of pharmacotherapy. Pharmacol Rev 58:389–462.
20. El Manira A, Kyriakatos A (2010): The role of endocannbinoid signalingin motor control. Physiology 25:230–238.
21. Pagotto U, Marsicano G, Cota D, Lutz B, Pasquali R (2006): Theemerging role of the endocannabinoid system in endocrine regula-tion and energy balance. Endocr Rev 27:73–100.
22. Pertwee RG (2001): Cannabinoid receptors and pain. Prog Neurobiol63:569–611.
23. Hillard CJ, Weinlander KM, Stuhr KL (2012): Contributions of endo-cannabinoid signaling to psychiatric disorders in humans: Geneticand biochemical evidence. Neuroscience 204:207–229.
24. Gardner EL, Vorel SR (1998): Cannabinoid transmission and reward-related events. Neurobiol Dis 5:502–533.
25. Maldonado R, Valverde O, Berrendero F (2006): Involvement of theendocannabinoid system in drug addiction. Trends Neurosci 29:225–232.
26. Matyas F, Urban GM, Watanabe M, Mackie K, Zimmer A, Freund TF,Katona I (2008): Identification of the sites of 2-arachidonoylglycerolsynthesis and action imply retrograde endocannabinoid signaling atboth GABAergic and glutamatergic synapses in the ventral tegmentalarea. Neuropharmacology 54:95–107.
27. Kortleven C, Fasano C, Thibault D, Lacaille JC, Trudeau LE (2011): Theendocannabinoid 2-arachidonoylglycerol inhibits long-term potentia-tion of glutamatergic synapses onto ventral tegmental area dopa-mine neurons in mice. Eur J Neurosci 33:1751–1760.
28. Szabo B, Siemes S, Wallmichrath I (2002): Inhibition of GABAergicneurotransmission in the ventral tegmental area by cannabinoids. EurJ Neurosci 15:2057–2061.
29. Melis M, Pistis M, Perra S, Muntoni AL, Pillolla G, Gessa GL (2004):Endocannabinoids mediate presynaptic inhibition of glutamatergictransmission in rat ventral tegmental area dopamine neurons throughactivation of CB1 receptors. J Neurosci 24:53–62.
30. Lupica CR, Riegel AC (2005): Endocannabinoid release from midbraindopamine neurons: A potential substrate for cannabinoid receptorantagonist treatment of addiction. Neuropharmacology 48:1105–1116.
31. Oleson EB, Beckert MV, Morra JT, Lansink CS, Cachope R, Abdullah RA,et al. (2012): Endocannabinoids shape accumbal encoding of cue-
www.sobp.org/journal
motivated behavior via CB1 receptor activation in the ventraltegmentum. Neuron 73:360–373.
32. Marsicano G, Goodenough S, Monory K, Hermann H, Eder M, CannichA, et al. (2003): CB1 cannabinoid receptors and on-demand defenseagainst excitotoxicity. Science 302:84–88.
33. Monory K, Massa F, Egertova M, Eder M, Blaudzun H, Westenbroek R,et al. (2006): The endocannabinoid system controls key epileptogeniccircuits in the hippocampus. Neuron 51:455–466.
34. Monory K, Blaudzun H, Massa F, Kaiser N, Lemberger T, Schutz G, et al.(2007): Genetic dissection of behavioural and autonomic effects ofdelta9-tetrahydrocannabinol in mice. PLoS-Biol 5:e269.
35. Bellocchio L, Lafenetre P, Cannich A, Cota D, Puente N, Grandes P,et al. (2010): Bimodal control of stimulated food intake by theendocannabinoid system. Nat Neurosci 13:281–283.
36. Dubreucq S, Matias I, Cardinal P, Haring M, Lutz B, Marsicano M,Chaouloff F (2012): Genetic dissection of the role of cannabinoid type-1 receptors in the emotional consequences of repeated social stressin mice. Neuropsychopharmacology 37:1885–1900.
37. Han J, Kesner P, Metna-Laurent M, Duan T, Xu L, Georges F, Koehl M, et al.(2012): Acute cannabinoids impair working memory through astroglial CB1
receptor modulation of hippocampal LTD. Cell 148:1039–1050.38. Stuhmer T, Puelles L, Ekker M, Rubenstein JLR (2002): Expression of a
Dlx gene enhancer marks adult mouse cortical GABAergic neurons.Cereb Cortex 12:75–85.
39. Franklin KBJ, Paxinos G (2008): The Mouse Brain in StereotaxicCoordinates, 3rd edition. New York: Academic Press.
40. Georges F, Aston-Jones G (2002): Activation of ventral tegmental areacells by the bed nucleus of the stria terminalis: A novel excitatory aminoacid input to midbrain dopamine neurons. J Neurosci 22:5173–5187.
41. Jalabert M, Bourdy R, Courtin J, Veinante P, Manzoni OJ, Barrot M,Georges F (2011): Neuronal circuits underlying acute morphine actionon dopamine neurons. Proc Natl Acad Sci U S A 108:16446–16450.
42. Long JZ, Nomura DK, Vann RE, Walentiny DM, Booker L, Jin X, et al.(2009): Dual blockade of FAAH and MAGL identifies behavioralprocesses regulated by endocannabinoid cross-talk in vivo. Proc NatlAcad Sci USA 106:20270–20275.
43. Holmes MM, Galea LAM, Mistlberger RE, Kempermann G (2004): Adulthippocampal neurogenesis and voluntary wheel running activity:Circadian and dose-dependent effects. J Neurosci Res 76:216–222.
44. van Praag H (2009): Exercise and the brain: Something to chew on.Trends Neurosci 32:283–290.
45. Keeney BK, Raichlen DA, Meek TH, Wijeratne RS, Middleton KM,Gerdeman GL, Garland T Jr (2008): Differential response to a selectivecannabinoid receptor antagonist (SR141716: rimonabant) in femalemice from lines selectively bred for high voluntary wheel-runningbehaviour. Behav Pharmacol 19:812–820.
46. Pertwee RG, Howlett AC, Abood ME, Alexander SPH, Di Marzo V,Elphick MR, et al. (2010): International Union of Basic and ClinicalPharmacology. LXXIX. Cannabinoid receptors and their ligands:Beyond CB1 and CB2. Pharmacol Rev 62:588–631.
47. Gardner A, Mallet PE (2006): Suppression of feeding, drinking, andlocomotion by a putative cannabinoid receptor “silent antagonist.”Eur J Pharmacol 530:103–106.
48. Canals M, Milligan G (2008): Constitutive activity of the cannabinoidCB1 receptor regulates the function of co-expressed Mu opioidreceptors. J Biol Chem 283:11424–11434.
49. Wiley JL, Breivogel CS, Mahadevan A, Pertwee RG, Cascio MG,Bolognini D, et al. (2011): Structural and pharmacological analysis ofO-2050, a putative neutral cannabinoid CB1 receptor antagonist. Eur JPharmacol 651:96–105.
50. Xi ZX, Peng XQ, Li X, Song R, Zhang HY, Liu QR, et al. (2011): Braincannabinoid CB2 receptors modulate cocaine’s actions in mice. NatNeurosci 14:1160–1166.
51. Marsicano G, Lutz B (1999): Expression of the cannabinoid receptorCB1 in distinct neuronal subpopulations in the adult mouse forebrain.Eur J Neurosci 11:4213–4225.
52. Navarrete M, Araque A (2008): Endocannabinoids mediate neuron-astrocyte communication. Neuron 57:883–893.
53. Acuna-Goycolea C, Obrietan K, van den Pol AN (2010): Cannabinoidsexcite circadian clock neurons. J Neurosci 30:10061–10066.
54. Sanford AE, Castillo E, Gannon RL (2008): Cannabinoids and hamstercircadian activity rhythms. Brain Res 1222:141–148.

S. Dubreucq et al. BIOL PSYCHIATRY 2013;73:895–903 903
55. Lett BT, Grant VL, Koh MT (2001): Naloxone attenuates the condi-tioned place preference induced by wheel running in rats. PhysiolBehav 72:355–358.
56. Belke TW, Wagner JP (2005): The reinforcing property and therewarding aftereffect of wheel running in rats: a combination oftwo paradigms. Behav Processes 68:165–172.
57. Greenwood BN, Foley TE, Le TV, Strong PV, Loughridge AB, Day HE,Fleshner M (2011): Long-term voluntary wheel running is rewardingand produces plasticity in the mesolimbic reward pathway. BehavBrain Res 217:354–362.
58. Rasmussen EB, Hillman C (2011): Naloxone and rimonabant reducethe reinforcing properties of exercise in rats. Exp Clin Psychopharma-col 19:389–400.
59. Werme M, Messer C, Olson L, Gilden L, Thoren P, Nestler EJ, Brene S(2002): DFosB regulates wheel running. J Neurosci 22:8133–8138.
60. Geisler S, Zahm DS (2005): Afferents of the ventral tegmental area inthe rat—anatomical substratum for integrative functions. J CompNeurol 490:270–294.
61. Watabe-Uchida M, Zhu L, Ogawa SK, Vamanrao A, Uchida N (2012):Whole-brain mapping of direct inputs to midbrain dopamineneurons. Neuron 74:858–873.
62. Chieng B, Azriel Y, Mohammadi S, Christie MJ (2011): Distinctcellular properties of identified dopaminergic and GABAergicneurons in the mouse ventral tegmental area. J Physiol 589:3775–3787.
63. Fu C, Cawthon B, Clinkscales W, Bruce A, Winzenburger P, Ess KC(2012): GABAergic interneuron development and function is modu-lated by the Tsc1 gene. Cereb Cortex 22:2111–2119.
64. Stuhmer T, Anderson SA, Ekker M, Rubenstein JLR (2002): Ectopicexpression of the Dlx genes induces glutamic acid decarboxylase andDlx expression. Development 129:245–252.
65. Keeney BK, Meek TH, Middleton KM, Holness LF, Garland T Jr (2012):Sex differences in cannabinoid receptor-1 (CB1) pharmacology inmice selectively bred for high voluntary wheel-running behavior.Pharmacol Biochem Behav 101:528–537.
www.sobp.org/journal
Copyright © 2022 FDOKUMEN




















